Psychophysiological Markers of Performance and
Learning during Simulated Marksmanship in
Immersive Virtual Reality
Sicong Liu1, Jillian M. Clements1, Elayna P. Kirsch1, Hrishikesh M. Rao2,
David J. Zielinski1, Yvonne Lu1, Boyla O. Mainsah1, Nicholas D. Potter1,3,
Marc A. Sommer1, Regis Kopper4, and Lawrence G. Appelbaum1
Astratto
■ The fusion of immersive virtual reality, kinematic movement
tracking, and EEG offers a powerful test bed for naturalistic neuro-
science research. Here, we combined these elements to investigate
the neuro-behavioral mechanisms underlying precision visual–
motor control as 20 participants completed a three-visit, visual–
motor, coincidence-anticipation task, modeled after Olympic
Trap Shooting and performed in immersive and interactive virtual
reality. Analyses of the kinematic metrics demonstrated learning of
more efficient movements with significantly faster hand RTs, earlier
trigger response times, and higher spatial precision, leading to an
average of 13% improvement in shot scores across the visits. As
revealed through spectral and time-locked analyses of the EEG
beta band (13–30 Hz), power measured prior to target launch
and visual-evoked potential amplitudes measured immediately
after the target launch correlated with subsequent reactive kine-
matic performance in the shooting task. Inoltre, both launch-
locked and shot/feedback-locked visual-evoked potentials became
earlier and more negative with practice, pointing to neural mecha-
nisms that may contribute to the development of visual–motor pro-
ficiency. Collectively, these findings illustrate EEG and kinematic
biomarkers of precision motor control and changes in the neuro-
physiological substrates that may underlie motor learning. ■
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
C
N
_
UN
_
0
1
7
1
3
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
INTRODUCTION
Humans are adept at interacting skillfully in dynamically
changing environments. We are able to maintain internal
and external representations of our bodies, extrapolate
motion of objects, and engage physically with these ob-
jects to achieve complex and precise results. These skills
are largely because of the remarkable visual–motor inte-
gration enabled by our brains, occhi, and bodies.
Studies addressing visual–motor integration have led to a
wealth of knowledge about the mechanisms that translate
goals and states into motor commands (Elliott, Hayes, &
Bennett, 2012; Wolpert, Ghahramani, & Jordan, 1995),
motor learning ( Wolpert & Flanagan, 2016; Schmidt &
Lee, 2011), and the neural characteristics of expertise in
highly practiced individuals (Yarrow, Brown, & Krakauer,
2009). Visually guided movements have been studied in a
range of contexts to characterize the influence of visual
information on movement control, demonstrating a bal-
ance between preprogrammed ballistic movements and
visual-feedback-mediated refinements (Urbin, Stodden,
Fischman, & Weimar, 2011; Elliott et al., 2010; Desmurget
& Grafton, 2000; Meyer, Keith-Smith, Kornblum, Abrams,
1Duke University School of Medicine, 2MIT Lincoln Laboratory,
Lexington, MA, 3US Olympic Committee, Colorado Springs,
CO, 4University of North Carolina at Greensboro
© 2021 Istituto di Tecnologia del Massachussetts
& Wright, 1990), as classically characterized by Fitts’ Law
(Fitts, 1954).
Scalp-recorded EEG has been particularly instrumental
in understanding the cortical activity involved in visual–
motor integration. This high-temporal resolution approach
has allowed for the identification of processes essential for
visual scene (Appelbaum, Ales, & Norcia, 2012; Anokhin
et al., 2006) and motion perception ( Jensen, Kaiser, &
Lachaux, 2007; Kuba, Kubová, Kremlácek, & Langrová,
2007), such as is characterized by contralateralized visual-
evoked potentials (VEPs) within the first ∼200 msec after
stimulus presentation. Inoltre, EEG has been widely
used to understand attention (Appelbaum & Norcia, 2009;
Jensen et al., 2007) and memory (Robitaille et al., 2010;
Klimesch, 1999) processes necessary for goal-oriented
behaviors, as well as error-related responses that have been
shown to reflect learning through reweighting of sensory
informazione (Cohen, Elger, & Ranganath, 2007; Hajcak,
Moser, Holroyd, & Simons, 2006). These studies have led
to a broad understanding of the chronometry of processes
within the visual–motor cascade, including evidence that
higher attentional states, indexed by less default-mode pro-
cessing and lower alpha power, are associated with better
processing of visual stimuli (per esempio., Macdonald, Mathan, &
Yeung, 2011). These findings and the ease-of-use of EEG
have led to a growing number of studies evaluating expertise
Journal of Cognitive Neuroscience 33:7, pag. 1253–1270
https://doi.org/10.1162/jocn_a_01713
in activities such as marksmanship (Berka, Behneman, Kintz,
Johnson, & Raphael, 2010; Janelle & Hatfield, 2008;
Hatfield, Haufler, Hung, & Spalding, 2004; Hillman,
Apparies, Janelle, & Hatfield, 2000), golf putting (Arns,
Kleinnijenhuis, Fallahpour, & Breteler, 2008; Babiloni et al.,
2008), table tennis (Hülsdünker, Ostermann, & Mierau,
2019), badminton (Hülsdünker, Strüder, & Mierau, 2017),
and archery (Seo et al., 2012; Landers, Han, Salazar, &
Petruzzello, 1994). Inoltre, because the EEG power spec-
tra and ERPs associated with sensory processing, decision
making, and error recognition in these tasks correlate with
trial-by-trial response speed (Hülsdünker et al., 2019) E
different levels of accomplishment (Hatfield et al., 2004),
they may provide potentially useful biomarkers in closed-
loop neurofeedback and neurostimulation approaches
(Gruzelier, 2014; Paulus et al., 2009).
The prospect of investigating brain dynamics of actively
behaving participants in complex 3-D environments has
been challenging. In most natural environments, conditions
cannot be controlled and are difficult to replicate. In
aggiunta, studies using noninvasive modalities for recording
brain activity, such as EEG, usually aim to minimize artifact-
producing muscle activity by limiting mobility. Despite this,
real-world tasks involve dynamic movements that must be
achieved to fully engage perception, planning, and motor
control for executing complex objectives. Studies that limit
mobility may not capture all the cognitive and neural pro-
cesses involved in performing natural, full-body movements,
thereby creating a need for more advanced platforms for
naturalistic brain and body monitoring.
Recent improvements in simulation technology, motion
tracking, and mobile EEG have contributed to the develop-
ment of mobile brain/body imaging (MoBI), an approach
that investigates the links between distributed brain dynam-
ics and natural behavior (Gramann et al., 2011; Makeig,
Gramann, Jung, Sejnowski, & Poizner, 2009). Integration
of simulation technologies, such as immersive and interac-
tive virtual reality (VR) with brain and behavioral measure-
menti, allow for complex tasks to be performed in
controlled, indoor 3-D environments with high ecological
validity. In particular, previous studies have used the
MoBI approach to investigate physical interactions with
dynamically moving objects ( Jungnickel & Gramann,
2016), cognitive control during locomotion ( Wagner,
Makeig, Gola, Neuper, & Müller-Putz, 2016), and physical
exertion during high-intensity cycling (Enders et al.,
2016). Inoltre, skill training in VR translates successfully
to many real-world tasks that require procedural (Ragan,
2010), motor (dos Santos Mendes et al., 2012), and athletic
(Gray, 2017; Miles, Pop, Watt, Lawrence, & John, 2012; Fink,
Foo, & Warren, 2009; Aggarwal et al., 2006) apprendimento, and can
be used to simulate critical elements of stress and pressure
providing an avenue for widespread application (Sanz,
Multon, & Lécuyer, 2015; Stinson & Bowman, 2014).
In the current study, we combined these elements to in-
vestigate the neural and behavioral mechanisms underlying
visual–motor skill learning during a three-visit simulated
marksmanship training protocol based on the rules,
physics, and timing of Olympic Trap Shooting. In this
coincidence-anticipation task, individuals shot a simulated
flying clay pigeon with a firearm game controller while
kinematic movement tracking and 13-channel EEG were
recorded within a six-sided immersive and interactive VR
ambiente. Building upon our previous findings demon-
strating kinematic changes in the ballistic and refinement
stages of movement on this task (Rao et al., 2018), the goals
of the current study were to characterize the cascade of
kinematic and electrophysiological activity that contribute
to the successful shooting of targets. In particular, the pri-
mary aim of this study was to test if EEG biomarkers early in
the neural processing cascade correlated with ensuing be-
havioral outcomes, including RTs and eventual shot preci-
sion. As such, this study sought to establish biomarkers of
performance that unfold over time by leveraging the high
temporal resolution of EEG and the characteristic sequence
of actions involved in the trap shooting event. Therefore,
based on the temporal dependence of this trap shooting
task, we sought to establish brain states early in a trial
(either preparatory prior to the launch of the pigeon or
early in the evoked cascade of sensory responses) Quello
are able to reliably capture variance in measured kinematics
and EEG occurring later in the trial. Secondary goals were to
characterize changes in the latency and amplitude of kine-
matic and electrophysiological responses that occur as par-
ticipants practice this task and to determine if error-related
responses at the end of a trial contribute to performance in
the next trial, as would be expected from reinforcement
learning theories (Walsh & Anderson, 2012).
Based on the timing and movement constructs of this
trap shooting simulation and the literature describing
neuro-kinematic processes involved in visual–motor or-
ienting, several hypotheses were formulated and tested.
A central expectation was that VEPs would be present
contralateral to the launch direction of the target and that
larger amplitudes for this potential would correlate with
better shooting performance. A second expectation in this
study was that EEG spectral power over para-Rolandic,
motor control areas during the preparatory interval prior
to the launch of each target would correlate with move-
ment outcomes of the trial. Finalmente, although highly ex-
ploratory, it was hypothesized that the occurrence of
misses, versus hits, on each trial would elicit characteris-
tic feedback-related negativity (FRN; Holroyd & Coles,
2002). Such findings would add to understanding of
motor system physiology and aid in the development of
a naturalistic neuroscience approaches that can be used
for evaluation and training in applied contexts.
METHODS
Participants
Twenty-four participants (mean age = 25 ± 4.28 years,
13 women) were recruited to take part in this study. Four
1254
Journal of Cognitive Neuroscience
Volume 33, Numero 7
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
C
N
_
UN
_
0
1
7
1
3
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
participants were excluded from subsequent analyses for
the following reasons: Two participants were left-handed,
creating discrepancies in the lateralized EEG analyses; one
participant had extensive marksmanship experience; E
one participant did not complete the entire protocol. IL
remaining 20 participants constituted a homogenous set of
right-handed novice marksmen, with normal or corrected-
to-normal vision, no history of neurological deficits or
family history of photosensitive epileptic seizures, and no
previous experience with the protocol presented in this
study. Participation was voluntary, and participants were
compensated $10/hr for their involvement. The experi-
mental protocol was approved by the Duke University
institutional review board.
Equipment
VR and Motion Tracking
The study was conducted in the Duke immersive Virtual
Environment (DiVE; Figure 1A), a six-sided cave automatic
virtual environment (CAVE) VR system (Cruz-Neira,
Sandin, & DeFanti, 1993), where participants stood in
the center of a room-size 3 × 3 × 3 m cube with projectors
directed at each of the cube’s six walls. Projectors were
run at 120 Hz with a total resolution of 1920 × 1920 pixels
per wall. Participants wore 3-D shutter glasses operating at
60 Hz to view stereoscopic graphics. A head tracking de-
vice was mounted on the glasses to control the system
viewpoint according to the participant’s head movements.
An Intersense IS-900 tracking system was used to record
the 3-D position and orientation of the controller and head
throughout the experiment. Data from both the controller
and head tracking sensors were sampled at 60 Hz.
An Xbox Top Shot Elite firearm game controller was used
for target shooting. The controller, furnished with a six
degrees-of-freedom tracking sensor, was held in the partic-
ipant’s right hand and stabilized with the opposite hand
placed along the barrel of the controller, as shown in
Figure 1B. Target acquisition was performed using a ray
casting technique, but rather than a visible ray, only a white
dot was shown at the target depth to mimic a laser sight.
EEG Recording
EEG signals were recorded using a BrainVision actiCAP sys-
tem with 13 active gel-based electrode channels, Quale
were placed on the scalp according to the 10–20 system
at locations F3, Fz, F4, C3, Cz, C4, T5, T6, P3, Pz, P4, O1,
and O2, with mastoid references. Two additional elec-
trodes were placed on the right and left outer canthi of
the eyes to record horizontal eye movements via HEOG.
All electrode impedances were kept below 10 kΩ to record
high-quality signals that were sampled at 1000 Hz. IL
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
C
N
_
UN
_
0
1
7
1
3
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
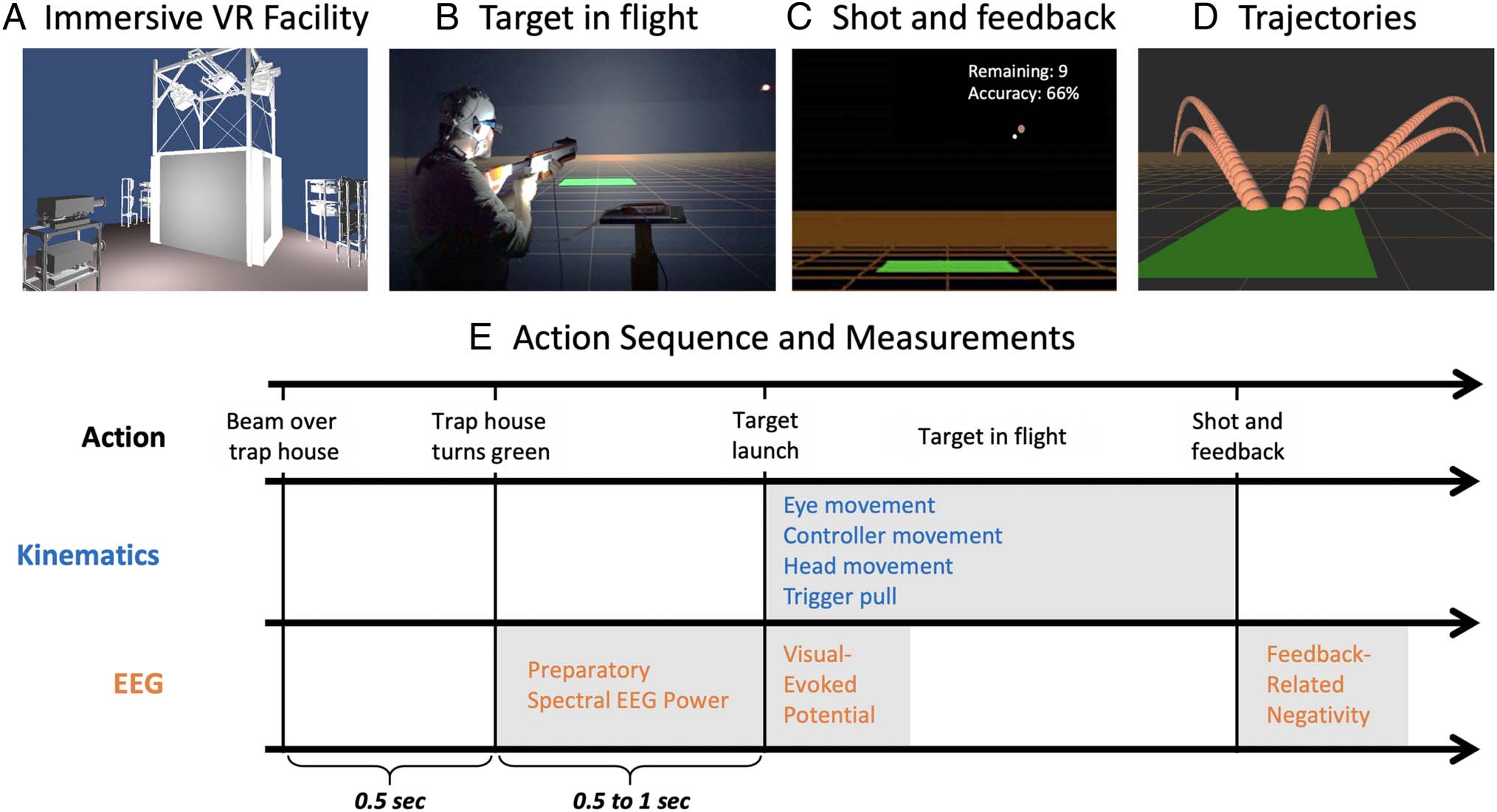
Figura 1. (UN) Rendering of the DiVE immersive and interactive VR facility where participants took part in the study. (B) Picture of an individual
participating in the marksmanship simulation wearing EEG and holding firearm controller, while target is in mid-flight. Each trial began when the
controller was aimed at the green trap house, after which the pigeon was launched and the participant was given one chance to shoot and hit the
moving target. (C) Screen immediately following a missed shot, showing the position of the target and controller, and feedback if it was a hit (green)
or miss (red), as well as the remaining trials and running accuracy in that task block. (D) Sequential screenshots illustrating paths of the six target
trajectories. (E) Schematic illustration of the time course in a given trial, including event actions, bodily kinematics, and derived EEG markers.
Liu et al.
1255
dominant environmental artifact at 60 Hz (power line
frequency) was attenuated using a 0.1- to 30-Hz bandpass
filter.
Experimental Design
Simulated Marksmanship Task
The task presented inside the DiVE was modeled after the
Olympic Trap Shooting event (International Shooting
Sport Federation, 2013), where the goal for the participant
was to shoot a digital clay pigeon that was launched from
behind a trap house. The design of the simulation abided
by the laws of physics for projectile motion (including
gravitational pull, air resistance, and lift force) to mimic
realistic projectile flight.
To begin the task, participants aimed the controller
toward a digital trap house, which was displayed as a rect-
angle above the ground, 54 ft in front of them in simulated
spazio. After an initial 500-msec waiting period, the trap
house changed color from red to green and a second
waiting period began that was variable between 0.5 E
1.0 sec. During the initial waiting period, if the participant
aimed the controller away from the trap house before the
color changed from red to green, the timer was reset and
did not begin until the participant aimed the controller
back toward the trap house. At the end of the second wait-
ing period, a digital target (displayed as an orange sphere
of radius 1 piedi) was launched in one of six directions with the
initial speed of 95.34 ft/sec.
Participants were given one chance to shoot the target.
The longer a participant took to pull the trigger, IL
further the target moved away from them, making it more
difficult to hit because of the change in depth and corre-
sponding reduction in visual angle. Once the trigger was
pulled, the screen immediately froze to display feedback
that included the participant’s shot location relative to
the location of the target, their cumulative shot success
(percent hits), and the number of trials remaining, COME
shown in Figure 1C. If the controller’s ray was in contact
with the target at the time of the shot, the target changed
color from orange to green, indicating a hit. If not, IL
color changed from orange to red to indicate a miss. IL
feedback remained on the screen until the controller was
returned to a position over the trap house, which started
the next trial sequence.
The six possible target trajectories, illustrated in
Figure 1D, consisted of three horizontal directions relative
to the center of the trap house (left = −45°, center = 0°,
right = 45°) and two elevations relative to the ground
plane (upper = 25.17°, lower = 12.95°). To increase eco-
logical validity (per esempio., fluctuations in outdoor environmental
conditions such as wind currents), random horizontal
noise ranging from −3° to 3° was added to the launch
angle. The maximum flight times for the target were
1.772 sec and 3.085 sec for the lower and upper trajectory
elevations, rispettivamente.
Training Protocol
Participants practiced the simulated marksmanship task
on three separate visits within 1 week. During each visit,
which lasted approximately 2 hr, the practice session
was split into eight blocks of 60 trials. Within each block,
all six target trajectories were presented 10 times in a ran-
dom order. During the first visit, participants were given
two practice rounds to become acquainted with the task
(approximately 7 min total). The first practice round con-
sisted of trials where the target was launched only in the
upper right direction, with the target at a larger size than in
the real simulation. The second practice round consisted
of six trials where a normal-sized target was launched in
each possible direction. The second practice round was
administered twice if the participants’ hit rate was below
50%. After completing the practice rounds, participants
began the eight blocks of training and had the option to
take a break between Blocks 2, 4, E 6 to prevent fatigue.
Psychophysiological Measures
The shooting task presented in this study is particularly
useful for investigating psychophysiological skill learning
because it produces discrete measures of performance
(participants either hit or miss the target) while still re-
quiring complex visual–motor coordination. In order to
determine the types of skill learning that occurred through
practice, dependent variables were extracted from the
kinematic motion tracking and EEG data to analyze
changes that occurred over time and to study the relation-
ships between motor proficiency and neural activity.
Figure 1E illustrates the sequence of event actions within
a single trial from which the EEG and kinematic variables
were extracted. Gray shaded regions indicate approximate
intervals and biomarkers during which kinematic (blue) E
EEG (orange) responses were calculated. These psycho-
physiological measures are discussed individually below.
Behavioral and Kinematic Measures
Shot success and shot error were used as indicators to de-
termine performance improvements through practice.
Shot success was calculated as the percentage of target
hits out of the total number of shots taken. Shot error
was measured as the Euclidean distance (in millimeters)
between the location of the shot and the center of the
target at the time of the trigger pull and therefore reflects
the precision of the shot attempt. High shot success rates
and lower shot error (defined by shorter distances to the
target) signify superior performance that is reproducible
and consistent.
The timings of key behavioral events, such as the onset of
movement, were extracted from the motion tracking and
HEOG data. RT—the elapsed time from the target launch
to the onset of movement—was calculated for the move-
ment of the eyes, controller, and head. Controller RT and
1256
Journal of Cognitive Neuroscience
Volume 33, Numero 7
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
C
N
_
UN
_
0
1
7
1
3
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
head RT were calculated using 10% of the peak acceleration
trace, as measured with the Intersense motion trackers (van
Donkelaar & Franks, 1991). Acceleration was computed as
the derivative of the speed trace, after the speed trace was
smoothed with a seventh order finite impulse response
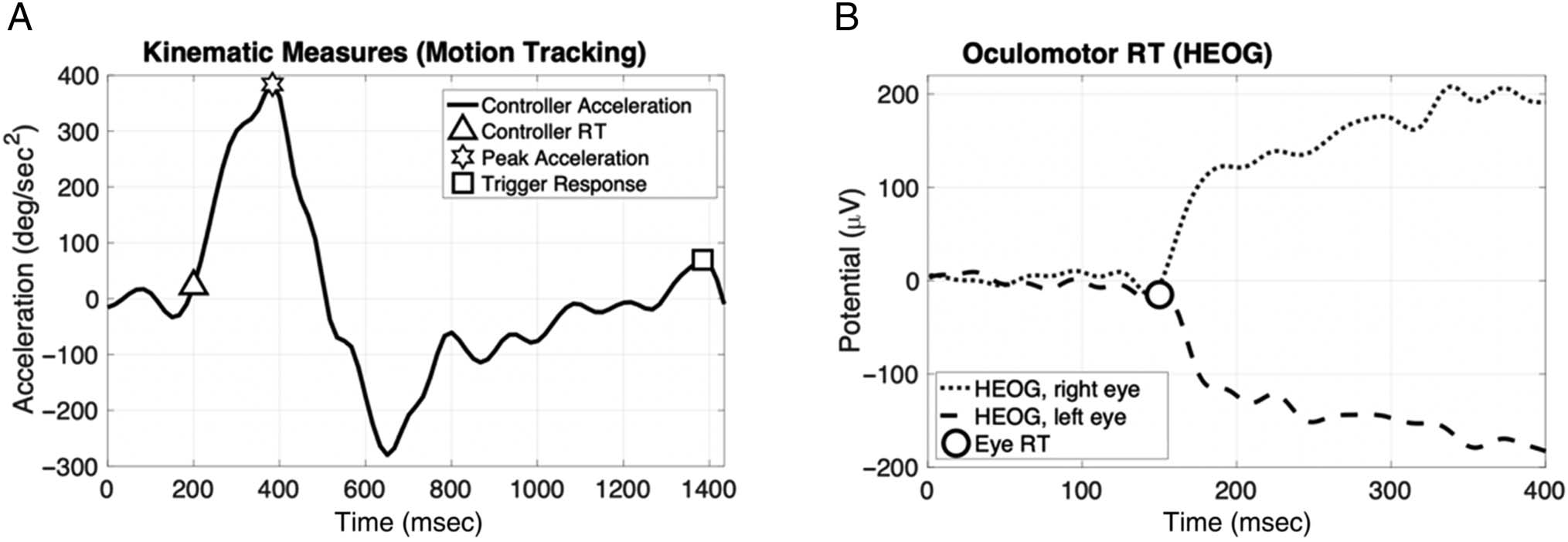
filter. An example acceleration trace collected for a single
trial is shown in Figure 2A. Oculomotor RT was calculated
using a rectified sum of the corneoretinal potential mea-
sured from the two HEOG channels for the left and right
horizontal target trajectories, as shown in Figure 2B. IL
corneoretinal potential occurred when the positive part of
the eye (the cornea) rotated closer to either the left or right
HEOG electrode, resulting in an upward voltage deflection
for the closer electrode and a downward deflection for the
opposite electrode. The final oculomotor RT was recorded
when a threshold of 3 SDs above a baseline mean (100 msec
postlaunch) was exceeded. Trigger response time was
calculated as the elapsed time between the target launch
and when the trigger was pulled to shoot. Trials were ex-
cluded from the analyses if the participant did not pull
the trigger to shoot (1.67%, 482 trials) or the movement
was initiated too quickly for a given trial, defined as a con-
troller RT of less than 16.667 msec, or one rendered video
frame (0.39%, 23 trials).
EEG Measures
EEG data were divided into three separate segments of the
task for analysis: (1) the prelaunch preparatory EEG
phase, which is thought to represent the participant’s
mental state prior to target launch; (2) the postlaunch
VEP that reflects visual-cortical processing of the target
prior to kinematic movement; E (3) the postshot FRN
that reflects the participants’ response to hits versus mis-
ses on each trial.
Preparatory EEG signals were extracted from a
500-msec window in the EEG data after the trap house
turned green. Baseline drifts were corrected by subtract-
ing the mean value of the signal in the preceding 50-msec
finestra. The preparatory phase was characterized by
spectral power. The baseline-corrected EEG signals were
transformed to compute the power spectral density esti-
mates using Welch’s method (window size = 250 msec,
overlap = 50%) and divided into the standard EEG fre-
quency bands for analysis: delta (0.1–4 Hz), theta (4–8 Hz),
alpha (8–13 Hz), and beta (13–30 Hz). Trials containing
excess muscle artifact were rejected by removing voltages
that exceeded a threshold of 200 μV (0.97%, 279 out of
28,800 trials).
VEPs were extracted from the 200-msec window imme-
diately following a target launch in the left or right direc-
tions to assess the contralateral/ipsilateral visual response
in the brain. Baseline correction was performed by sub-
tracting the average voltage in the 50-msec prelaunch
finestra. Electrodes P3 and P4 were selected for analysis
because of their posterior locations over the left and right
hemispheres of the visual cortex, rispettivamente, E
because of their higher signal quality as compared to
Channels O1 and O2, which tended to include more noise
because of cap fitting issues and neck muscle artifacts
produced during movement. Single-trial VEP amplitudes
were calculated using the average over a 20-msec window
centered on the participant’s peak VEP latency for that
visit. Trials were excluded from the averages if they ex-
ceeded a threshold of ± 35 μV (3.02%, 580 out of 19,200
trials) or contained data outside of 5 SDs of the joint prob-
ability distribution observed at each time point (0.35%, 67
out of 19,200 trials). There were no systematic differences
in rejection rates across the different trajectory directions.
The use of EEG artifact removal techniques (as opposed to
rejection) would have been challenging because of the
lack of clear biological templates (per esempio., ocular artifact cor-
rection) to base exclusion on. Inoltre, based on the low
prevalence of rejected trials (3.37% total), it was evident
Figura 2. (UN) Example of an acceleration trace measured from the firearm controller during a single trial. Kinematic dependent variables are
indicated by the triangle, star, and square markers. (B) Example of eye movement signals for a target launched rightward, captured via HEOG and
time-locked to the target launch at 0 msec that are used to calculate oculomotor RT, indicated here by the circle.
Liu et al.
1257
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
C
N
_
UN
_
0
1
7
1
3
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
that the signal under consideration offered an unimpeded
view of the neural activity that was meant to be scrutinized
in the planned hypothesis tests.
The FRN was calculated by extracting data epochs span-
ning from 200 msec before the trigger pull to 800 msec
after the trigger pull at channel Cz. These “shot-locked
potentials” were baseline corrected by subtracting the
average of a 100-msec preshot window. Individual trial
data were separately averaged for all successful shots (hits)
and for all unsuccessful shots (misses). Preliminary analy-
ses determined nonsignificant differences over the shot
elevations (high vs. low) and shot directions (left vs. center
vs. right). Therefore, the FRN was calculated using the
average over all trajectories, separately on each visit of
the protocol for all 20 participants. Single-trial FRN ampli-
tudes were calculated using the average of the 40 msec
surrounding the peak latency for that visit, whereas single
trial onset latencies were calculated as the latency at 1/3
peak amplitude.
Statistical Analysis
The purpose of this study was twofold: (1) to test if EEG
biomarkers early in the cascade of processes induced
during the coincidence anticipation task correlated with
ensuing behavioral outcomes, including shooting move-
ment RTs and eventual shot precision, E (2) to analyze
skill learning as a function of practice by exploring changes
in both EEG and behavior over time to determine if error-
related responses at the end of a trial contributed to per-
formance in the next trial.
All the preprocessing of EEG recordings was performed
in MATLAB (The MathWorks) using the EEGLab toolbox
(Delorme & Makeig, 2004). To investigate relationships
between EEG variables and motor behavior variables in a
data set showing substantial clustering structure (cioè., In-
traclass coefficient estimates are in [0.36, 0.92]), linear
mixed-effects models (LMEMs) were fit and tested using
the lme4 and lmerTest package in R Version 3.5.3
(R Core Team, 2019). LMEMs were fit to EEG and behav-
ioral data that included trial-level observations (task block
and visit [as a statistical nuisance factors to offset variable
discontinuities], EEG measures [preparatory delta/theta/
alpha/beta band power and VEP amplitude], launch direc-
zione), nested within individual participants. In different
models, controller RT or shot error served as outcome var-
iables while allowing for the intercept parameter to vary
among participants. The LMEMs involving controller RT
or shot error also included an exponential function term
of trial number (cioè., e-Trial Number), whose slope parameter
was treated as a random effect and allowed to vary among
participants. All the LMEMs reached normal convergence
using restricted maximum likelihood estimator. Nel
LMEMs, the statistical significance of the parameter esti-
mates was tested using t distribution with degree of freedom
adjusted using Satterthwaite’s method (see Kuznetsova,
Brockhoff, & Christensen, 2017).
Repeated-measures analysis of variance (rANOVA) era
performed in SPSS v25 to test skill learning across time
on both EEG and behavioral variables. A given rANOVA in-
volved either the main effect of Visit (cioè., Visit 1–3) or the
main effects of Block (cioè., Block 1–8) and interactions
from Visit and Block, depending on the specific question
being evaluated. Mauchly’s method was used to examine
the assumption of sphericity in the data. In case(S) Di
assumption violation, model outcomes based on the
Greenhouse–Geisser correction were adopted.
A final analysis was performed on the FRN to test its role
in reinforcement learning, where epochs were resegmen-
ted to create the sequential categories: hits following hits,
hits following misses, misses following hits, and misses
following misses. Single-trial mean amplitudes within the
40 msec surrounding the peak FRN for each of the four
segment classes were submitted to rANOVA to evaluate
main effects and interactions between the Shot Success
(cioè., hit or miss) of successive trial pairs. Effects were
considered statistically significant if multiple-comparison
adjusted p values were less than .05.
RESULTS
The data set in this study consists of metrics characterizing
task performance, movement kinematics, and electro-
physiology that change over the course of learning and il-
lustrate relationships between the brain and body. Nel
following sections, results from these three classes of met-
rics are presented to infer the detailed chronometry and
influence of psychophysiological activity associated with
performance on the marksmanship task.
Task Performance
Shot success rate—the percentage of hits out of total shots
taken—was calculated for each block and visit for all 20
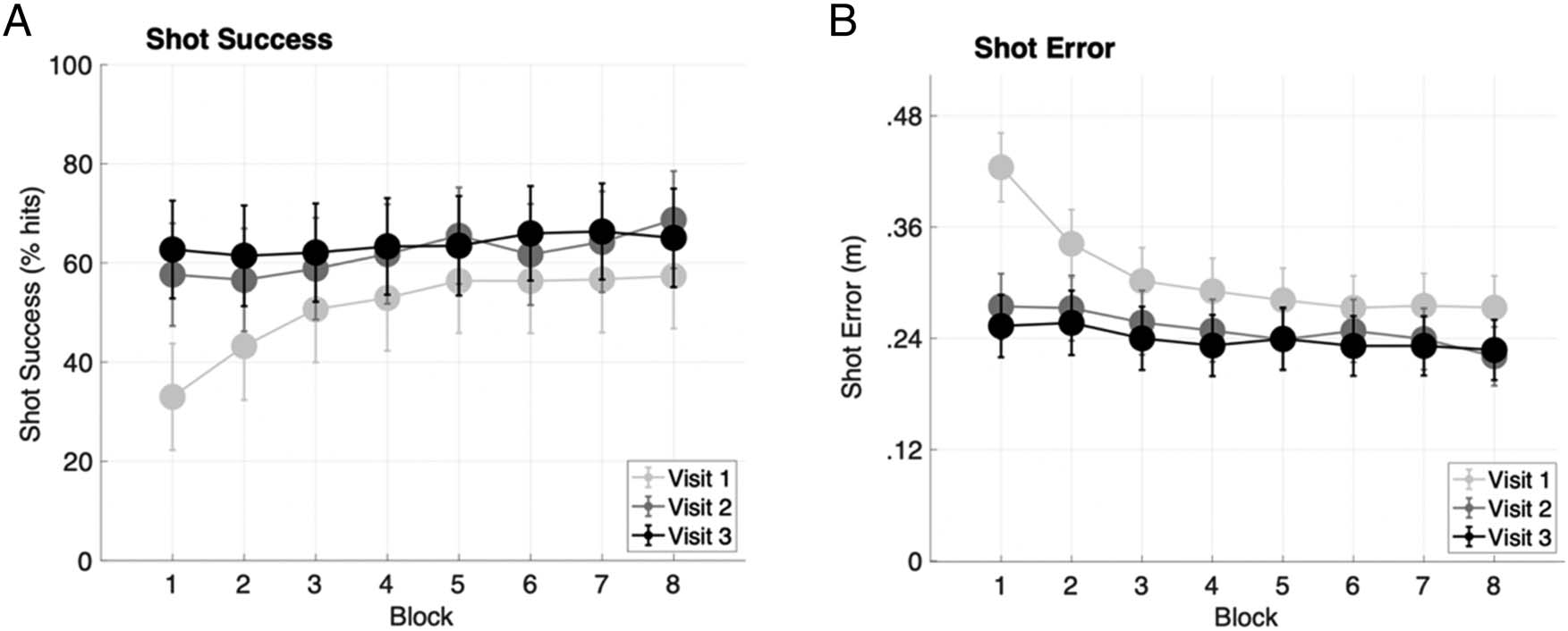
participants. The average results are shown in Figure 3A.
As expected, participants significantly improved their suc-
cess rate through practice, with rANOVA demonstrating
main effects of Visit, F(2, 38) = 68.50, P < .001, η2 =
.78, and Block on the first visit, F(7, 133) = 22.78, p <
.001, η2 = .55, and the second visit, F(7, 133) = 10.04,
p < .001, η2 = .346. However, there was not a main effect
of Block on the third visit, F(7, 133) = 1.54, p = .16, η2 =
.08, suggesting that performance may have begun to pla-
teau as participants became more familiar with the task.
Shot error—the Euclidean distance between the shot
location and the center of the target—significantly de-
creased across visits, F(2, 38) = 50.65, p < .001, η2 =
.73, as shown in Figure 3B. A significant decrease across
block was also observed on the first visit, F(7, 133) = 26.91,
p < .001, η2 = .59; the second visit, F(7, 133) = 11.27, p <
.001, η2 = .37; and the third visit, F(7, 133) = 3.27, p < .001,
η2 = .15. Here, lower values represent shots that were
closer to the target. Therefore, decreases in this distance
1258
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
c
n
_
a
_
0
1
7
1
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
c
n
_
a
_
0
1
7
1
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. (A) Increases in shot success rate, measured as the proportion of hits versus misses on each block, and (B) decreases in shot error,
measured as the Euclidian distance error from the target (lower is better), both illustrated that performance improved significantly over the course of
practice. Error bars denote the within-subject standard deviation for each condition.
show that participants continuously enhanced their per-
formance as they practiced.
Kinematic Behavior
In addition to the shot success and shot error outcome
measurements described above, kinematic tracking of
the participants allowed for calculating onset movement
times of eye, hand, and head. As illustrated in Table 1,
these movement times proceeded first with oculomotor
RTs indexing the start of horizontal eye movements re-
corded through the HEOG channels at approximately
194 msec after target launch. This was followed within
the first 300 msec by controller RTs and then head RTs,
which marked the onset of hand and head movement,
respectively. Finally, trigger response time occurred at
approximately 1.5 sec, marking the average elapsed time
between the target launch and the execution of the shot.
Whereas oculomotor RTs were unchanged throughout
the study and did not show a main effect of block or visit,
other measures sped up throughout task practice.
Namely, a significant decrease in controller RT was
observed across visits, F(2, 38) = 27.53, p < .001, η2 =
.59, and blocks on the first visit, F(7, 133) = 13.27, p <
.001, η2 = .41; the second visit, F(7, 133) = 7.13,
p < .001, η2 = .27; and the third visit, F(7, 133) = 5.75,
p < .001, η2 = .23. These reductions indicate that hand
movements became faster as participants practiced the
task throughout the entirety of the training protocol. A
significant decrease in head RT was also observed across
visits, F(2, 38) = 3.92, p < .05, η2 = .17, but was not
observed across blocks on any of the visits. Finally, trigger
response time significantly decreased across visits, F(2,
38) = 4.70, p < .05, η2 = .20, as participants became faster
overall at performing the task.
EEG Biomarkers
Scalp-recorded EEG provides detailed chronometry of
the neural activity associated with performance on the
marksmanship task. The following sections characterize
the prelaunch preparatory-spectral, visual-evoked, and
shot-locked EEG responses that occurred before, during,
and after each trial of the task, respectively.
Prelaunch Preparatory EEG
To assess the relationship between prelaunch preparatory
EEG and postlaunch motor performance, spectral power
was calculated from channels C3, C4, and Cz using the sig-
nals extracted in the 500-msec window after the trap house
turned green (prior to the target launch). Time–frequency
Table 1. Average Value and Standard Deviation of Reaction and Response Times (in milliseconds) for Each Visit
Oculomotor RT (msec)
Controller RT (msec)
Head RT (msec)
Visit 1
194 ± 46
213 ± 46
297 ± 13
Visit 2
194 ± 44
204 ± 39
284 ± 12
Visit 3
193 ± 49
199 ± 39
288 ± 12
Trigger response time (msec)
1,571 ± 399
1,529 ± 364
1,514 ± 337
Liu et al.
1259
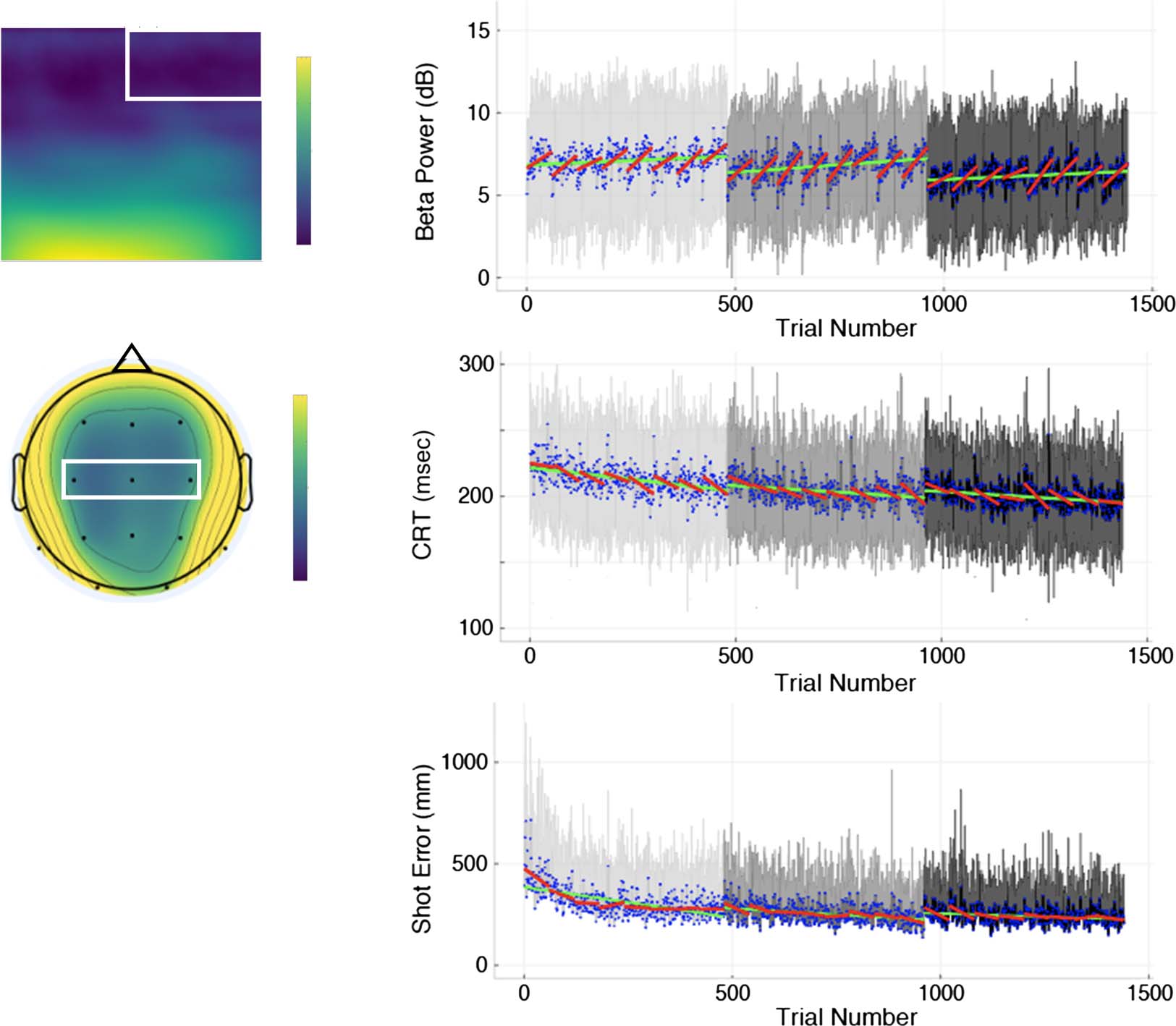
analysis of the spectral power time-locked to the moment
of trap house turning green produced spectral power
estimates that generally decreased at higher frequencies
and were broadly distributed over the center of the head, as
illustrated in the group average spectrogram in Figures 4A
and topography for beta band power in Figure 4B.
Two LMEMs were fit to explore changes in preparatory
alpha and beta power, over the course of practice with the
marksmanship task. Trial number, task block, and visit
were included in the LMEMs as fixed-effects terms with
the intercept treated as a random effect among partici-
pants. In both LMEMs, results showed that alpha and
beta power tonically increased with the growth factor of
trial number ( p < .001), controlling for other factors.
Moreover, an LMEM was fit to test the predictive power
of prelaunch preparatory EEG frequency band power, task
block, and visit on postlaunch motor behavior measured
by controller RT and shot error, respectively, given the ran-
dom effects of intercept and nonlinear term (i.e., expo-
nential function) among participants. The LMEM run on
controller RT showed reasonable fit to the data, account-
ing for 4%, 13%, and 52% of the variance associated with
the Level 1 residual term, Level 2 intercept term, and Level 2
exponential term, respectively, given that the intraclass
coefficient estimates suggested a high proportion of data
variance coming from Level 2 random variables (Cohen,
Cohen, West, & Aiken, 2013). Table 2 demonstrates pa-
rameter estimates for the fixed-effects part of the LMEM,
revealing that preparatory beta power was a significant
predictor of controller RT, t(27907.30) = −3.53, p <
.001. The regression coefficient can be interpreted such
that, for each dB increase in beta power, controller RT
was faster by 0.34 msec, when controlling for all other fac-
tors. Results of LMEM for shot error indicated that EEG
preparatory power did not correlate with shot error.
Overall, this evidence implicated a functional role of beta
band EEG in the marksmanship task toward predicting
controller RTs. Figure 4C–4E illustrates the tonic variability
pattern of beta power, controller RT, and shot error across
the entire sequence of experimental trials in the three-visit
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
c
n
_
a
_
0
1
7
1
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. (A) Time–frequency plot from para-Rolandic channels in a 1000-msec time window time-locked to the moment the trap house turning
green with the white rectangle highlighting the extracted beta power signal from the preparatory period. (B) Scalp topography map of beta power
during the preparatory period. White triangle highlighting the para-Rolandic channels. Group means and standard deviations across trials with block-
and visit-wide regression lines are shown for (C) preparatory period beta power, (D) controller RT (CRT), and (E) shot error. Note that only the
upper standard deviation bars were plotted for shot error to avoid possible misleading illustration that shot error can be negative-valued.
1260
Journal of Cognitive Neuroscience
Volume 33, Number 7
Table 2. LMEM Fixed-Effect Results for Predicting Controller RT
(Intercept)
Alpha power
Beta power
Delta power
Theta power
Task block
Visit 2
Visit 3
***p < .001.
Estimate (SE)
216.34 (4.52)
−0.14 (0.12)
−0.34 (0.10)
0.04 (0.04)
−0.02 (0.09)
−1.21 (0.08)
−9.27 (0.46)
−13.06 (0.46)
t [df]
47.87 [22.37]
−1.17 [27897.04]
−3.53 [27907.30]
1.00 [27900.42]
−0.24 [27900.18]
−14.92 [27852.89]
−20.34 [27885.91]
−28.39 [27884.37]
p Value
< .001***
.24
< .001***
.32
.81
< .001***
< .001***
< .001***
protocol. Block-wise (red) and visit-wise (green) regres-
sion lines are descriptively plotted along with group
means and standard deviations.
Visually Evoked Potential
Based on the lateralization of the visual system, an impor-
tant a priori hypothesis in this study was the expectation
that visual cortical responses would differ in the hemi-
sphere contralateral to left or right trajectory launches.
Moreover, based on the expectation that physiological
responses in the visual cortex would correlate with
behavior, it was expected that this contralateralized
potential would be earlier and larger on trials with better
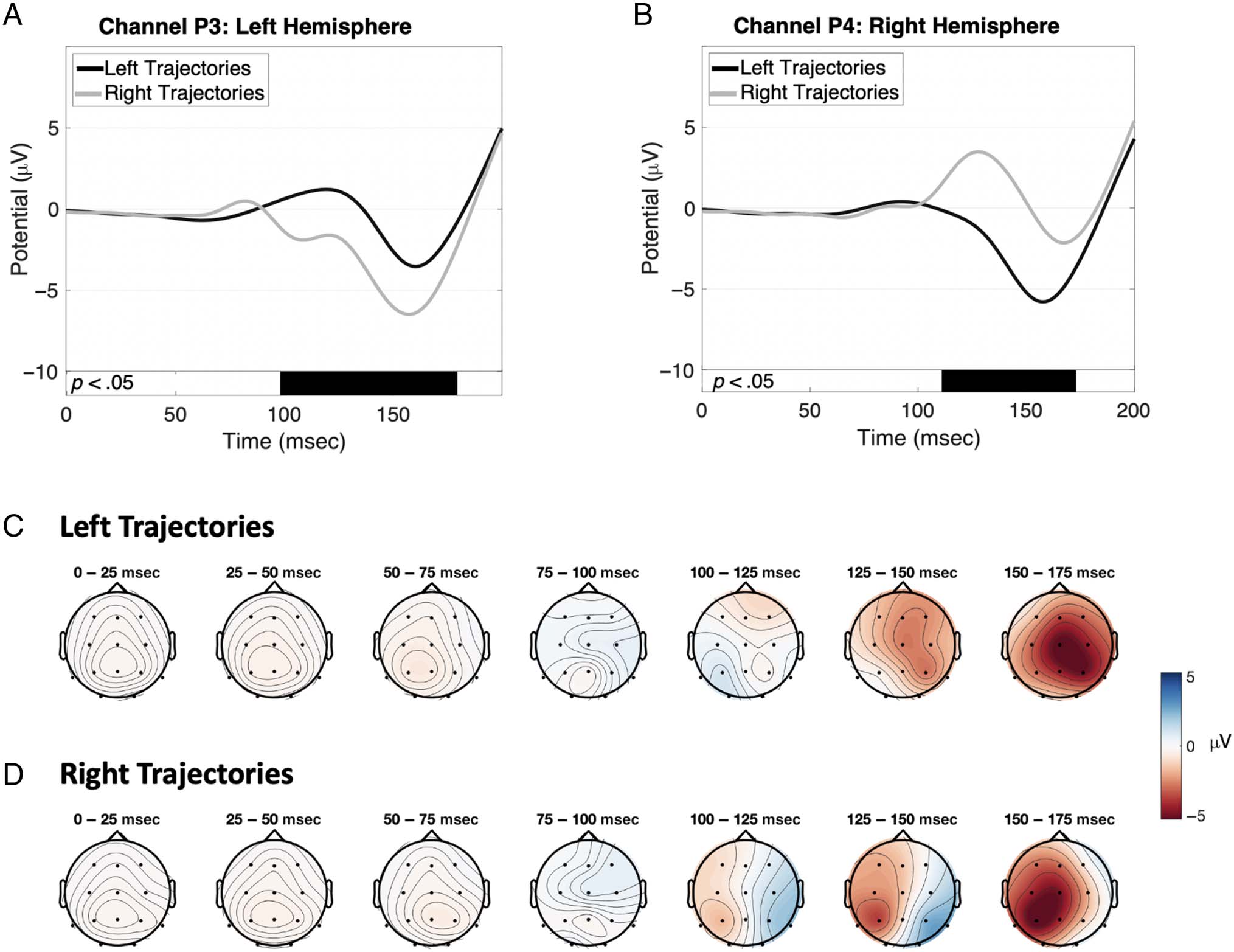
behavioral performance. As shown in Figure 5A and 5B,
an early ipsilateral positive VEP began at around 100 msec,
followed by the onset of a larger contralateral negative
VEP that began at 110–120 msec and ended before the
onset of oculomotor RT at around 194 msec. Statistical
tests performed between 0 and 200 msec, with Bonferroni
method to adjust for multiple comparisons, showed that
significant differences existed between the VEPs for
the left and right trajectory directions in the range of
98–179 msec and 111–173 msec, respectively, as indicated
by the black bars below the VEPs in each plot. Figure 5C
and 5D illustrate the evolution of the topographic dis-
tribution of the evoked responses for left and right tra-
jectories, respectively, demonstrating the presence of
contralateralized responses over the posterior cortex,
prior to the initiation of eye movements.
In order to understand the role of the VEP in behavior,
LMEMs were fit to test the predictive power of the contra-
lateral VEP amplitude, launch direction, task block, and
visit on controller RT and shot error, given the random
effect of intercept and nonlinear exponential term among
participants. In addition, the LMEMs included a fixed-
effects factor, launch direction, to account for whether
the trial observation came from channel P3 or P4. The
LMEM was run with data excluding trials of central launch
trajectory (to be consistent with the requirement of calcu-
lating contralateral VEP amplitude).
Overall, the LMEM showed reasonable fit to the data,
explaining 6%, 15%, and 72% of the variance associated
with the Level 1 residual term, Level 2 intercept term,
and Level 2 exponential term, respectively. Results of the
fixed-effects part in the LMEM run on controller RT are
shown in Table 3. VEP amplitude was identified as a signif-
icant predictor of controller RT, regardless of channel
location, t(19076.33) = 11.10, p < .001.1 Namely, for each
μV increment in the negative-going VEP amplitude, the
controller RT became faster by 0.21 msec, controlling for
all other factors. In addition, the results suggested that
participants, on average, reacted to targets launched to
the right 5.94 msec faster than to those launched to the
left, t(19069.34) = −15.70, p < .001. Results of LMEM
run on shot error indicated VEP amplitude did not predict
shot error.
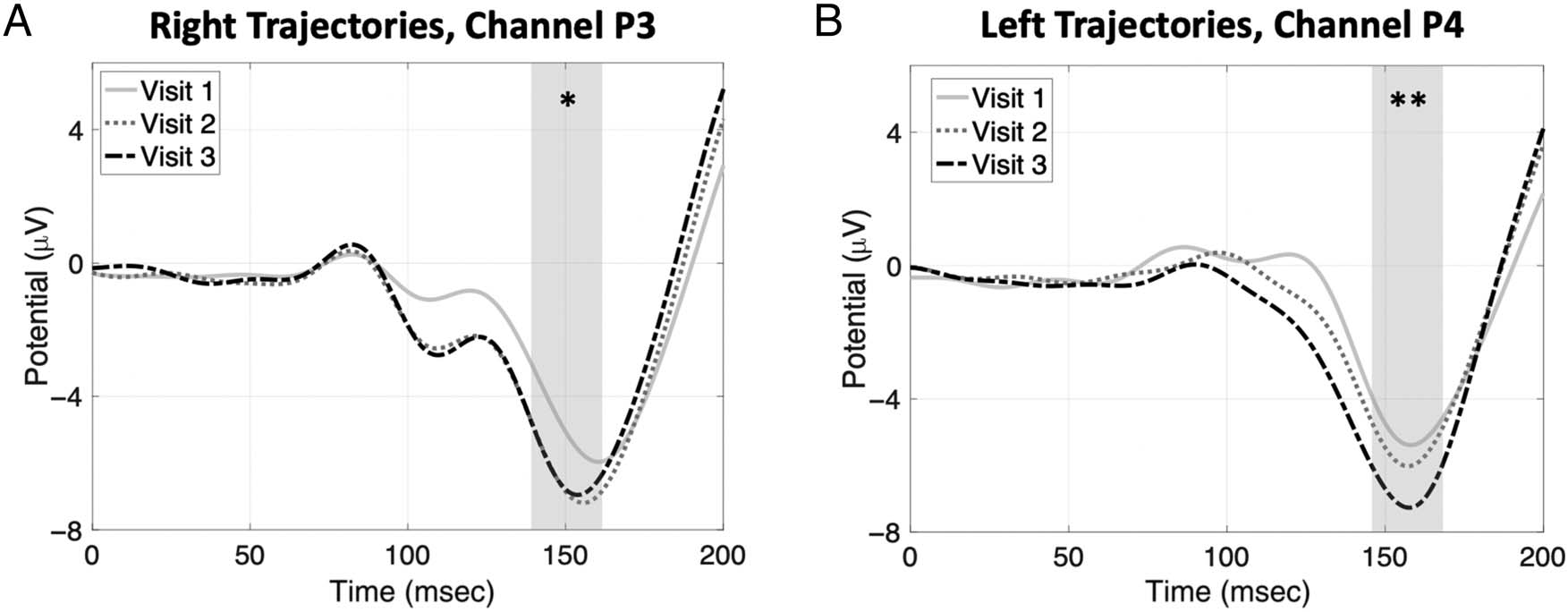
To further evaluate learning that occurred over visits,
additional rANOVAs were performed on the individual
daily average VEP amplitudes for channels P3 and P4.
The rANOVAs revealed significant decreases in amplitude
across visits in both channel P3, F(2, 38) = 4.59, p < .05,
η2 = .20, and channel P4, F(2, 38) = 9.18, p = .001, η2 =
.33. These findings are illustrated for the grand average
VEPs in Figure 6A and 6B, where successive decreases in
amplitudes can be seen across consecutive visits.
Combined Model Predicting
Marksmanship Performance
To obtain an overall model of the behavioral and EEG mea-
sures leading up to shot execution that predict marksman-
ship performance, an LMEM was run regressing controller
RT on the preshot EEG-based variables (EEG power bands
and VEP), task block, visit, and launch direction, while
allowing parameters associated with the intercept and
nonlinear exponential term to vary among participants.
The LMEM was run with identical trial observations used
Liu et al.
1261
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
c
n
_
a
_
0
1
7
1
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
c
n
_
a
_
0
1
7
1
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. (A–B) Grand average VEPs for the left (black) and right (gray) target trajectories in channels P3 and P4. The black bars below the VEPs
indicate latencies where the signals differ significantly between trajectory directions. (C–D) Grand average topographic distributions for left (top) and
right (bottom) target trajectories, both showing posterior contralateral distributions from approximately 100 to 150 msec.
in the previous section of testing VEP amplitude’s predic-
tion on controller RT. Overall, the LMEM showed reason-
able fit to data, accounting for 6%, 17%, and 66% of the
variance associated with the Level 1 residual term, Level 2
intercept term, and Level 2 exponential term, respectively.
Results from the fixed-effects part of the LMEM model are
shown in Table 4. In particular, the results included previ-
ous findings that both beta power, t(19073.51) = −3.14,
p = .002, and VEP amplitude, t(19072.35) = 11.05, p <
.001, significantly predicted controller RT. Moreover,
Table 3. LMEM Fixed-Effect Results for Predicting Controller RT Using VEP Amplitude, Trial Number, Task Block, Visit, and Launch
Direction
(Intercept)
VEP amplitude
Right launch direction
Task block
Visit 2
Visit 3
***p < .001.
Estimate (SE)
214.25 (3.69)
0.21 (0.02)
−5.94 (0.38)
−1.00 (0.08)
−8.22 (0.47)
−11.05 (0.46)
t [df]
58.00 [15.70]
11.10 [19076.33]
−15.70 [19069.34]
−12.10 [18992.47]
−17.60 [19043.30]
−23.80 [19042.31]
p Value
< .001***
< .001***
< .001***
< .001***
< .001***
< .001***
1262
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
c
n
_
a
_
0
1
7
1
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Grand average contralateralized VEPs for each visit, time-locked to the target launch. (A) VEPs in channel P3 for targets launched rightward
and (B) VEPs in channel P4 for targets launched leftward. Average VEP amplitudes became significantly more negative across consecutive visits,
suggesting that the brain’s visual response to the target launch increased with practice. Gray shading illustrates 20-msec window over which
amplitudes were averaged. * p < .05. ** p < .01.
participants, on average, reacted to targets launched to the
right 5.98 msec faster than those launched to the left,
t(19038.00) = −15.82, p < .001.
FRN and Sequential Trial Effects
A final set of exploratory analyses was performed to deter-
mine if a response similar to the FRN was present during
the time period immediately following the trigger pull. In
the present design, the pull of the trigger instantaneously
froze the display, turned the target green or red depend-
ing on shot success, and displayed the participant’s shot
location relative to the location of the target. Because
this happened in the context of ongoing movement to
intercept the target, a first objective was to assess for the
presence of an FRN-like response that is frontal-centrally
distributed with greater negative amplitudes for misses,
relative to hits. In the presence of such a response, a sec-
ond objective was to determine whether the amplitude
and/or latency of the FRN changed with practice and
whether or not such changes correlated with behavioral
learning. Finally, based on past demonstrations that the
amplitude of the FRN predicts whether participants will
learn to avoid an erroneous response on subsequent
attempts (van der Helden, Boksem, & Blom, 2010),
sequential-trial behavioral and brain effects were then
evaluated under the hypotheses that (1) behavioral mea-
sures would be faster for trials following misses than hits
Table 4. LMEM Fixed-Effect Results for Predicting Controller RT Using All Preparatory Power Bands, VEP Amplitude, Trial Number,
Visit, and Launch Direction
(Intercept)
Alpha power
Beta power
Delta power
Theta power
VEP amplitude
Task block
Right launch direction
Visit 2
Visit 3
**p < .01.
***p < .001.
Estimate (SE)
217.34 (3.74)
−0.14 (0.12)
−0.31 (0.10)
0.002 (0.04)
−0.02 (0.09)
0.21 (0.02)
−0.98 (0.08)
−5.98 (0.38)
−8.39 (0.47)
−11.47 (0.47)
t [df]
58.04 [18.62]
−1.13 [19066.90]
−3.14 [19073.51]
0.06 [19068.20]
0.24 [19072.72]
11.05 [19072.35]
−11.83 [18994.55]
−15.82 [19038.00]
−17.99 [19038.65]
−24.40 [19038.00]
p Value
< .001***
.26
.002**
.96
.81
< .001***
< .001***
< .001***
< .001***
< .001***
Liu et al.
1263
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
c
n
_
a
_
0
1
7
1
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
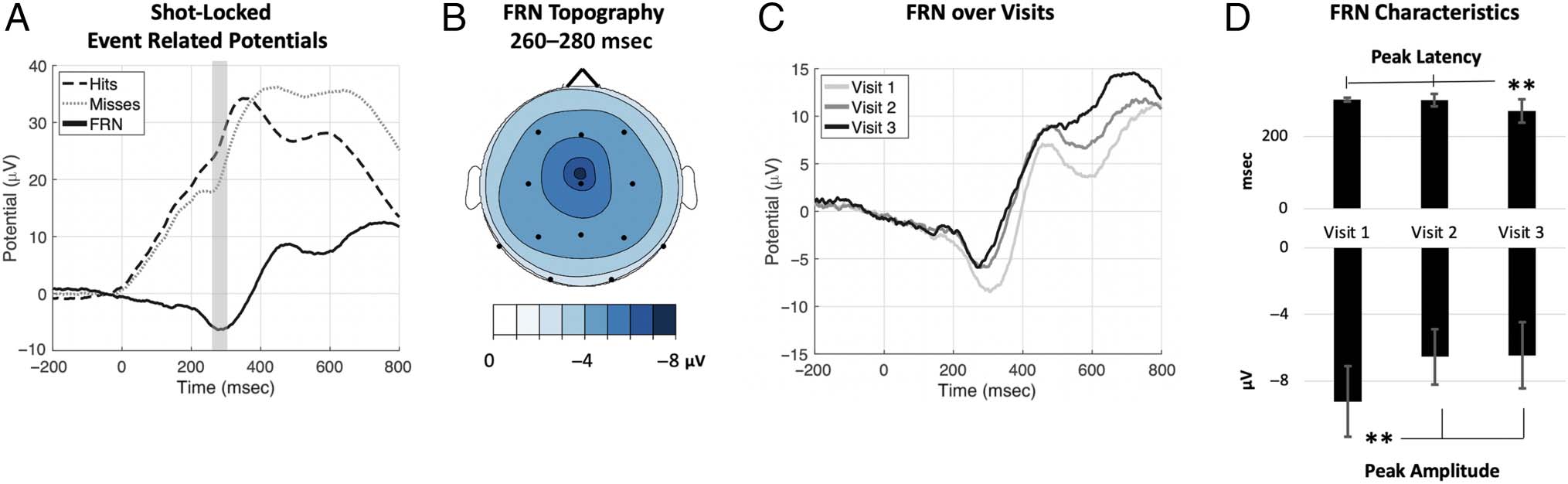
Figure 7. (A) Grand average ERPs for hits (dashed black line), misses (dotted gray line), and the miss-minus-hit differences (solid black line) at
channel Cz, illustrating the FRN-like response. Gray shading indicates ± 20-msec window centered on the peak for this condition. (B) Average
topography of FRN 260- to 280-msec posttrigger pull. (C) FRN over visits at channel Cz. (D) Means and SDs of FRN peak latency and amplitude over
visits. ** p < .01 pairwise comparison.
and (2) that greater FRN amplitudes would be followed
by more successful behavior on the subsequent trials.
Analysis of the FRN revealed differences between hits
(black dashed line) and misses (gray dotted line), shown
graphically in Figure 7A. The miss-minus-hit difference
wave (black solid line) consists of a broadly distributed
negative potential that peaks at approximately 280 msec,
similar to the commonly reported FRN in the literature
(Cohen et al., 2007; Hajcak et al., 2006), and is relatively
unchanged in distribution over visits. The FRN scalp to-
pography is displayed in Figure 7B. As displayed in wave-
forms in Figures 7C and evidenced in rANOVA results in
Figure 7D, the peak latency of this potential occurred ear-
lier across visits, F(1.44, 27.24) = 17.158, p < .001, η2 =
.475, shifting from 303 msec, to 300 msec, to 270 msec
across the three visits. Similarly, rANOVA performed on
the peak amplitude extracted within ± 20 msec of this
peak also demonstrate significant differences across visits,
F(1.77, 33.68) = 57.08, p < .001, η2 = .75, with successive
reductions in the amplitude over visits.
To better understand sequential trial effects, all trials
were reclassified into four conditions: hits following misses,
hits following hits, misses following misses, and misses
following hits. Based on past research showing that indi-
viduals exert greater control following errors (Allain,
Burle, Hasbroucq, & Vidal, 2009; Rabbitt, 1966), it was ex-
pected that participants would be more likely to hit the
target following miss trials relative to hit trials. However,
contrary to these expectations, rANOVA comparing the
proportion of hits following hits to the number of hits
following misses showed that participants were more
likely to hit the target following a successful trial than an
unsuccessful trial, F(1, 19) = 4.85, p = .04, η2 = .20. Across
the three visits, participants were successful 67.5% of the
time following hits, but only 65.7% of the time following
misses. A 2 (Outcome: hit or miss) × 2 (Subsequent
Shot: hit or miss) × 3 (Visit) rANOVA performed on the
FRN amplitudes indicated that, although there was a main
effect of Shot Outcome on FRN amplitudes, F(1, 18) =
25.21, p < .001, η2 = .58, this did not interact with the
sequential shot result, F(1, 18) = 0.07, p = .79, indicating
that performance on a given trial could not be predicted
based on the amplitude of the shot-locked FRN on the
previous trial. As such, the current exploratory analyses
provide indication of an FRN response that changes with
practice and evidence of behavioral sequential trial effects
(akin to the “hot hand” where a person who experiences
a successful outcome has a greater chance of success in
further attempts), but do not provide support that the
amplitude or latency of the FRN indexes behavioral
changes across trials.
DISCUSSION
In this study, a simulated trap shooting task was used to
investigate the behavioral and brain processes underlying
visual–motor control and learning. Repeated natural
movement patterns were measured with kinematic track-
ing, whereas brain activity was simultaneously measured
with EEG as participants practiced and became, on aver-
age, 13.03% more accurate at shooting targets. Analysis
of the behavioral measures revealed that performance im-
provements were accompanied by faster controller RTs,
lower shot errors, and earlier trigger response times, indi-
cating that less time was needed for motor planning, exe-
cution, and error correction as training progressed.
Analysis of the EEG data reveals several markers that cor-
related with upcoming motor performance and changed
over time with practice. First in this sequence, beta band
power measured in the brief 500-msec preparatory period
before the target launch correlated with RTs of the upcom-
ing shot. Next, following the target launch, a contralateral
VEP was observed over posterior cortex, indicating that
1264
Journal of Cognitive Neuroscience
Volume 33, Number 7
cortical processing of the target occurred before the onset of
eye, hand, and head movements captured by the kinematics.
The amplitude of this VEP became more negative with prac-
tice and correlated with controller RT, pointing to a neural
mechanism that may contribute to the development of
visual–motor proficiency. Last, upon execution of the trig-
ger pull, there was a broadly distributed potential that was
more negative for misses than hits, suggestive of the com-
monly reported FRN (Holroyd & Coles, 2002). Although
this response became significantly smaller and earlier over
the course of practice, it did not differ for subsequent hits
versus misses as would be expected based on reinforce-
ment learning models that have associated this response
with reward prediction. Methodologically, it was shown
that an MoBI approach is feasible for recording and
analyzing EEG during a simulated trap shooting event
and, by extension, other tasks of similar motor demand
when conducted in an immersive and interactive CAVE-
like VR system.
Collectively, these findings build on our previous results
demonstrating kinematic motor learning (Rao et al., 2018;
Zielinski, Rao, Potter, Appelbaum, & Kopper, 2016) and
biomarker identification (Clements et al., 2018), test for
changes in brain activity and body movements that accom-
pany skill learning, and establish biomarkers that can be
used to infer behavioral performance. In the following
discussion, we first address the rapidly growing field of
MoBI then provide context for the observed learning
and brain–behavior correlation findings, before closing
with limitations and future directions that can help move
this field forward.
MoBI in an Immersive and Interactive
Virtual Environment
Traditional EEG experiments often record with motor
responses in the form of minimal movements, such as
button presses. In contrast, MoBI experiments simulta-
neously record multiple sources of information under
naturalistic contexts allowing a more complete representa-
tion of the full-body behavior (Rao et al., 2020; Makeig,
2009). The current study used such an MoBI approach
to provide important new insights into the brain dynamics
accompanying the interception of a moving target in a
naturalistic performance context with multiple levels of
kinematic and brain measurement. Kinematic tracking
performed inside a CAVE-like VR environment provided
the exact timing of key events in simultaneous EEG re-
cordings, which allowed for calculating brain responses
throughout the full cascade of preparatory and execution
steps of the target interception task. Importantly, although
it might seem counterintuitive, EEG data collection in this
immersive and interactive VR environment produced
notably clean recordings. This is likely because of the fact
that the participants stood in the center of the chamber
with image projection from outside the cave onto the
fabric walls, so the participants were actually quite far from
interfering electrical noise sources. The fact that expected
EEG metrics could be measured with quiet baseline inter-
vals (demonstrating characteristic 1/f + alpha power spec-
trum), low artifact rejection rates (< 3.5% using 35-μV
rejection threshold), and significant conditional
differences indicates further that the psychophysiological
approach used here is conducive to identifying EEG bio-
markers of behavior outcomes. Moreover, these findings
may offer important information for how to integrate
EEG into the emerging use of VR in motor learning appli-
cations, performance training (Ahir, Govani, Gajera, &
Shah, 2020; Appelbaum & Erickson, 2018), and rehabilita-
tion (Levac et al., 2016; Levin, Weiss, & Keshner, 2015).
EEG Biomarkers of Motor Behavior
One of the main objectives in this research was to identify
EEG markers of motor behavior during the simulated
marksmanship task. The term “biomarker” commonly re-
fers to a measurable indicator of a specific biological state,
which can be used to describe a psychophysiological pro-
cess underlying behavior. In recent years, there has been
an increasing interest in the identification of biomarkers to
assess skill in performance-related research (Lee et al.,
2017; Cheron et al., 2016; Paulus et al., 2009). Validated
biomarkers have the potential to aid coaches, athletes,
and researchers in the design of better training programs
that can increase their effectiveness. Despite interest in re-
search on motor skills during dynamic activities, these
studies typically involve minimally active laboratory tasks
where the same principles may not apply to motor skills
in more complex real-world situations.
In this study, the addition of EEG recording in a VR train-
ing environment added a new dimension into the investi-
gation of the psychophysiological biomarkers that reflects
complex motor behaviors. With this framework, candi-
dates of behavior biomarker were extracted from EEG data
and tested in three different sets of multilevel models
(which are capable of accounting for the natural clustering
structure in the data); one focused on the preparatory
spectral power, one testing contralateral VEP amplitudes,
and a combined model that each give different views of the
correlational structure of these variables at different start
(preparation, processing, and reaction) and end points
(RT, response time, and shot error).
As observed in the LMEMs, beta band power appeared as
a reliable correlate of upcoming performance. This may
mirror past findings demonstrating a role for beta in
visual–motor integration during preparatory motor pro-
cesses (Zaepffel, Trachel, Kilavik, & Brochier, 2013; Perez,
Lundbye-Jensen, & Nielsen, 2006). In this model, it is
thought that phase locking of neural populations within
the beta band reflects greater entrainment that facilitates
neural information transfer during motor planning and ex-
ecution (Rueda-Delgado et al., 2014; van Wijk, Beek, &
Daffertshofer, 2012). When interpreted in this context,
the finding may indicate the development of an internal
Liu et al.
1265
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
c
n
_
a
_
0
1
7
1
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
mental model that aids participants in shot execution
(Monfared, Tenenbaum, & Folstein, 2019). Interestingly,
studies comparing expert and novice marksmen have
shown that experts exhibit greater alpha band power in
the seconds leading up to the trigger pull (Berka et al.,
2010; Janelle & Hatfield, 2008; Hatfield et al., 2004;
Hillman et al., 2000). This has been interpreted as indicating
that expert shooters possess greater default mode network
activity and lower active cognition when executing shots
(i.e., more automaticity and less top–down control). As
such, contrasting experts and novices on alpha power
may reflect differences in brain responses at different levels
of learning. Nonetheless, because of the limited temporal
window under scrutiny (500 msec) in this shooting simula-
tion, these data are treated as tonic markers that may not
have the same association as phasic changes often described
in the literature. Future studies are thus encouraged to slow
the pace of shooting tasks to allow better access to these
important processes. Through subtle modifications to the
shooting task, future studies may therefore be able to
explore the connection between tonic and phasic variability
in EEG spectral power and how they map to motor control
and learning process.
A further finding of interest was the observation that the
amplitude of the contralateralized VEP time-locked to the
target launch correlated positively with the controller RT.
This link suggests that, in this time-limited target-
interception task, greater processing resources early in
the sensory cascade, indicated by more negative-going
VEP amplitude, may have led to faster motor actions to in-
tercept the target. This evoked potential, elicited to the vi-
sual detection of the target launches, is theorized to reflect
early visual and attentional processes, occurring in the
contralateral occipital–parietal cortex. This is thought to
be similar to the N2pc reported in other rapid attentional
orienting tasks (An et al., 2012), but motor planning cir-
cuits may also contribute to this evoked potential. We
therefore encourage forthcoming investigations to further
clarify these relevant ERP components in similar naturalis-
tic motor tasks.
Neural Signatures of Learning
An important question in this research concerned the
mechanisms of motor skill learning during the simulated
marksmanship task. In particular, this task offers an inter-
esting view to evaluate how visual-evoked responses and
error detection signals may have changed with practice
and contributed to skill acquisition. Although novel in its
implementation, this approach extends past skill learning
research with EEG, which helps exploit the contralaterali-
zation of the visual and motor cortex, and to derive later-
alized ERPs that changed through practice (van den Berg,
Appelbaum, Clark, Lorist, & Woldorff, 2016; Clark,
Appelbaum, van den Berg, Mitroff, & Woldorff, 2015).
Based on these and other studies (e.g., An et al., 2012),
it has been proposed that learning is accompanied by
reorganizations at multiple stages of the neural hierarchy
with dynamically interacting reorganizations at each
stage (Makino, Hwang, Hedrick, & Komiyama, 2016).
One observation from the current data was that the am-
plitude of the contralateralized VEP became significantly
more negative over the course of practice, implying that
changes in the brain processes might occur through train-
ing. This change in VEP amplitude is mirrored by observa-
tions in the LMEM results, which consistently demonstrated
significant effects of trial number in that participants were
improving over the course of the experiment, a phenome-
non descriptively illustrated in Figure 4D and 4E for con-
troller RT and shot error, respectively.
Finally, although highly exploratory in the context of this
study, the FRN, which is known to occur in EEG recordings
after a participant recognizes an error, was also found to oc-
cur at earlier latencies and with lower mean amplitudes on
successive days of practice. Such changes in sensory pro-
cessing have been observed with learning in other domains,
including perceptual learning (Appelbaum, Wade, Vildavski,
Pettet, & Norcia, 2006), visual search (Arns et al., 2008;
Aggarwal et al., 2006), and reward learning (dos Santos
Mendes et al., 2012). As observed in Figure 7, subsequent
differences in the later positive P3 component may also
have indexed changes on a trial-by-trial or visit-by-visit
basis. However, because these differences may arise from
uncontrolled processes (e.g., lingering on the feedback or
score), it is not possible to uniquely associate these with
error processing. Having demonstrated here that the FRN
can be captured, future studies may construct designs that
can better index the multiple processes underlying this
component.
Conclusions
The present results highlight a multiscale approach to link
kinematic measurements of eye, hand, and head move-
ments with EEG recordings during natural dynamic actions
in a fully immersive and interactive VR environment. The
full-body orienting task required participants to actively in-
teract with their environments using fast, accurate move-
ments, with evidence for both kinematic and neural
learning over the course of task practice. Moreover, both
beta band activity in the short preparatory period prior
to target launch and contralateral VEP amplitudes in the
period immediately after the target launch correlated with
faster hand RTs, signaling important aspects of the neural
upstream that contributes to motor performance. Taken
together, the present protocol demonstrates the ability to
quantify neurophysiological substrates of motor control
and learning, while providing an empirical platform for
the continued development of MoBI for applied uses.
Future work should continue to incorporate ecologically
valid measures in testing expert marksmen with the even-
tual goal of deriving reliable biomarkers in real time to guide
motor performance and learning.
1266
Journal of Cognitive Neuroscience
Volume 33, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
c
n
_
a
_
0
1
7
1
3
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Acknowledgments
The authors would like to thank all of the participants for their
time and effort during this study.
Reprint requests should be sent to Lawrence G. Appelbaum,
Department of Psychiatry, Duke University, 400 Trent Dr.,
Durham NC 27710, or via e-mail: greg@duke.edu.
Funding Information
This research was funded by a grant support to L. G. A.
through the United States Army Research Office [W911NF-
15-1-0390].
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 to 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by estimated
gender identification of first author/last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .408, W(oman)/M = .335,
M/ W = .108, and W/ W = .149, the comparable proportions
for the articles that these authorship teams cited were
M/M = .579, W/M = .243, M/ W = .102, and W/ W = .076
(Fulvio et al., JoCN, 33:1, pp. 3–7). Consequently, JoCN
encourages all authors to consider gender balance explicitly
when selecting which articles to cite and gives them the
opportunity to report their article’s gender citation balance.
Note
1. A linear mixed effects model
involving the interaction
between VEP amplitude and launch direction was tested on con-
troller RT, and the interaction was not statistically significant
( p > .14).
REFERENCES
Aggarwal, R., Grantcharov, T. P., Eriksen, J. R., Blirup, D.,
Kristiansen, V. B., Funch-Jensen, P., et al. (2006). An
evidence-based virtual reality training program for novice
laparoscopic surgeons. Annals of Surgery, 244, 310–314.
https://doi.org/10.1097/01.sla.0000218094.92650.44, PubMed:
16858196
Ahir, K., Govani, K., Gajera, R., & Shah, M. (2020). Application
on virtual reality for enhanced education learning, military
training and sports. Augmented Human Research, 5, 7.
https://doi.org/10.1007/s41133-019-0025-2
Allain, S., Burle, B., Hasbroucq, T., & Vidal, F. (2009). Sequential
adjustments before and after partial errors. Psychonomic
Bulletin & Review, 16, 356–362. https://doi.org/10.3758/PBR
.16.2.356, PubMed: 19293107
An, A., Sun, M., Wang, Y., Wang, F., Ding, Y., & Song, Y. (2012).
The N2pc is increased by perceptual learning but is
unnecessary for the transfer of learning. PLoS One, 7, e34826.
https://doi.org/10.1371/journal.pone.0034826, PubMed:
22485189
Anokhin, UN. P., Golosheykin, S., Sirevaag, E., Kristjansson, S.,
Rohrbaugh, J. W., & Heath, UN. C. (2006). Rapid discrimination
of visual scene content in the human brain. Brain Research,
1093, 167–177. https://doi.org/10.1016/j.brainres.2006.03.108,
PubMed: 16712815
Appelbaum, l. G., Ales, J. M., & Norcia, UN. M. (2012). The time
course of segmentation and cue-selectivity in the human
visual cortex. PLoS One, 7, e34205. https://doi.org/10.1371
/journal.pone.0034205, PubMed: 22479566
Appelbaum, l. G., & Erickson, G. (2018). Sports vision training:
A review of the state-of-the-art in digital training techniques.
International Review of Sport and Exercise Psychology,
11, 160–189. https://doi.org/10.1080/1750984X.2016.1266376
Appelbaum, l. G., & Norcia, UN. M. (2009). Attentive and pre-
attentive aspects of figural processing. Journal of Vision, 9,
1–12. https://doi.org/10.1167/9.11.18, PubMed: 20053081
Appelbaum, l. G., Wade, UN. R., Vildavski, V. Y., Pettet, M. W., &
Norcia, UN. M. (2006). Cue-invariant networks for figure and
background processing in human visual cortex. Journal of
Neuroscience, 26, 11695–11708. https://doi.org/10.1523
/JNEUROSCI.2741-06.2006, PubMed: 17093091
Arns, M., Kleinnijenhuis, M., Fallahpour, K., & Breteler, R. (2008).
Golf performance enhancement and real-life neurofeedback
training using personalized event-locked EEG profiles.
Journal of Neurotherapy, 11, 11–18. https://doi.org/10.1080
/10874200802149656
Babiloni, C., Del Percio, C., Iacoboni, M., Infarinato, F., Lizio, R.,
Marzano, N., et al. (2008). Golf putt outcomes are predicted by
sensorimotor cerebral EEG rhythms. Journal of Physiology,
586, 131–139. https://doi.org/10.1113/jphysiol.2007.141630,
PubMed: 17947315
Berka, C., Behneman, A., Kintz, N., Johnson, R., & Raphael, G.
(2010). Accelerating training using interactive neuro-
educational technologies: Applications to archery, golf and
rifle marksmanship. International Journal of Sport and
Società, 1, 187–104. https://doi.org/10.18848/2152-7857/CGP
/v01i04/54040
Cheron, G., Petit, G., Cheron, J., Leroy, A., Cebolla, A., Cevallos,
C., et al. (2016). Brain oscillations in sport: Toward EEG
biomarkers of performance. Frontiers in Psychology, 7, 246.
https://doi.org/10.3389/fpsyg.2016.00246
Clark, K., Appelbaum, l. G., van den Berg, B., Mitroff, S. R., &
Woldorff, M. G. (2015). Improvement in visual search with
practice: Mapping learning-related changes in neurocognitive
stages of processing. Journal of Neuroscience, 35,
5351–5359. https://doi.org/10.1523/JNEUROSCI.1152-14
.2015, PubMed: 25834059
Clements, J. M., Kopper, R., Zielinski, D. J., Rao, H. M., Sommer,
M. A., Kirsch, E., et al. (2018). Neurophysiology of visual–motor
learning during a simulated marksmanship task in immersive
virtual reality. In Proceedings of the 25th IEEE Conference on
Virtual Reality and 3D User Interfaces. Reutlingen, Germany.
https://doi.org/10.1109/ VR.2018.8446068
Cohen, J., Cohen, P., West, S. G., & Aiken, l. S. (2013). Applied
multiple regression/correlation analysis for the behavioral
sciences (3rd ed.). Mahwah, NJ: Erlbaum. https://doi.org/10
.4324/9780203774441
Cohen, M. X., Elger, C. E., & Ranganath, C. (2007). Reward
expectation modulates feedback-related negativity and EEG
spectra. Neuroimage, 35, 968–978. https://doi.org/10.1016/j
.neuroimage.2006.11.056, PubMed: 17257860
Cruz-Neira, C., Sandin, D. J., & DeFanti, T. UN. (1993). Surround-
screen projection-based virtual reality: The design and
implementation of the CAVE. Proceedings of the 20th Annual
Conference on Computer Graphics and Interactive
Techniques, 135–142). https://portal.acm.org/citation.cfm
?doid=166117.166134. https://doi.org/10.1145/166117
.166134
Liu et al.
1267
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
C
N
_
UN
_
0
1
7
1
3
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21. https://doi.org/10.1016/j.jneumeth.2003
.10.009, PubMed: 15102499
Desmurget, M., & Grafton, S. (2000). Forward modeling allows
feedback control for fast reaching movements. Trends in
Cognitive Sciences, 4, 423–431. https://doi.org/10.1016/S1364
-6613(00)01537-0, PubMed: 11058820
error-related negativity. Psychological Review, 109, 679–709.
https://doi.org/10.1037/0033-295X.109.4.679, PubMed:
12374324
Hülsdünker, T., Ostermann, M., & Mierau, UN. (2019). The speed
of neural visual motion perception and processing
determines the visuomotor reaction time of young elite table
tennis athletes. Frontiers in Behavioral Neuroscience, 13,
165. https://doi.org/10.3389/fnbeh.2019.00165, PubMed:
31379535
dos Santos Mendes, F. A., Pompeu, J. E., Modenesi Lobo, A.,
Hülsdünker, T., Strüder, H. K., & Mierau, UN. (2017). Visual
Guedes da Silva, K., Oliveira, T., Peterson Zomignani, A., et al.
(2012). Motor learning, retention and transfer after virtual-
reality-based training in Parkinson’s disease—Effect of motor
and cognitive demands of games: A longitudinal, controlled
clinical study. Physiotherapy, 98, 217–223. https://doi.org/10
.1016/j.physio.2012.06.001
Elliott, D., Hansen, S., Grierson, l. E., Lyons, J., Bennett, S. J., &
Hayes, S. J. (2010). Goal-directed aiming: two components
but multiple processes. Psychological Bulletin, 136,
1023–1044. https://doi.org/10.1037/a0020958, PubMed:
20822209
Elliott, D., Hayes, S. J., & Bennett, S. J. (2012). 125 years
of perceptual-motor skill research. American Journal
of Psychology, 125, 9–23. https://doi.org/10.5406/amerjpsyc
.125.1.0009, PubMed: 22428422
Enders, H., Cortese, F., Maurer, C., Baltich, J., Protzner, UN. B., &
Nigg, B. M. (2016). Changes in cortical activity measured with
EEG during a high-intensity cycling exercise. Journal of
Neurophysiology, 115, 379–388. https://doi.org/10.1152/jn
.00497.2015, PubMed: 26538604
Fink, P. W., Foo, P. S., & Warren, W. H. (2009). Catching fly balls
in virtual reality: A critical test of the outfielder problem.
Journal of Vision, 9, 1–8. https://doi.org/10.1167/9.13.14,
PubMed: 20055547
Fitts, P. M. (1954). The information capacity of the human
motor system in controlling the amplitude of movement.
Journal of Experimental Psychology, 47, 381–391. https://doi
.org/10.1037/h0055392, PubMed: 13174710
Gramann, K., Gwin, J. T., Ferris, D. P., Oie, K., Jung, T. P., Lin, C. T.,
et al. (2011). Cognition in action: Imaging brain/body
dynamics in mobile humans. Reviews in the Neurosciences,
22, 593–608. https://doi.org/10.1515/RNS.2011.047, PubMed:
22070621
Gray, R. (2017). Transfer of training from virtual to real baseball
batting. Frontiers in Psychology, 8, 2183. https://doi.org/10
.3389/fpsyg.2017.02183, PubMed: 29326627
Gruzelier, J. H. (2014). EEG-neurofeedback for optimising
performance. IO: A review of cognitive and affective outcome
in healthy participants. Neuroscience & Biobehavioral
Reviews, 44, 124–141. https://doi.org/10.1016/j.neubiorev
.2013.09.015, PubMed: 24125857
Hajcak, G., Moser, J. S., Holroyd, C. B., & Simons, R. F. (2006).
The feedback-related negativity reflects the binary evaluation
of good versus bad outcomes. Biological Psychology, 71,
148–154. https://doi.org/10.1016/j.biopsycho.2005.04.001,
PubMed: 16005561
Hatfield, B. D., Haufler, UN. J., Hung, T. M., & Spalding, T. W. (2004).
Electroencephalographic studies of skilled psychomotor
performance. Journal of Clinical Neurophysiology, 21,
144–156. https://doi.org/10.1097/00004691-200405000-00003,
PubMed: 15375345
Hillman, C. H., Apparies, R. J., Janelle, C. M., & Hatfield, B. D.
(2000). An electrocortical comparison of executed and
rejected shots in skilled marksmen. Biological Psychology,
52, 71–83. https://doi.org/10.1016/S0301-0511(99)00021-6,
PubMed: 10686373
Holroyd, C. B., & Coles, M. (2002). The neural basis of human
error processing: Reinforcement learning, dopamine, and the
motion processing subserves faster visuomotor reaction in
badminton players. Medicine and Science in Sports and
Exercise, 49, 1097–1110. https://doi.org/10.1249/MSS
.0000000000001198, PubMed: 28072633
International Shooting Sport Federation. (2013). Shotgun
rules for trap, double trap, and skeet. In ISSF Official
statutes, rules, and regulations (pag. 401–470). Munich,
Germany: ISSF.
Janelle, C. M., & Hatfield, B. D. (2008). Visual attention and
brain processes that underlie expert performance:
Implications for sport and military psychology. Military
Psychology, 20(Suppl. 1), S39–S69.
Jensen, O., Kaiser, J., & Lachaux, J. P. (2007). Human gamma-
frequency oscillations associated with attention and memory.
Trends in Neurosciences, 30, 317–324. https://doi.org/10
.1016/j.tins.2007.05.001, PubMed: 17499860
Jungnickel, E., & Gramann, K. (2016). Mobile brain/body
imaging (MoBI) of physical interaction with dynamically
moving objects. Frontiers in Human Neuroscience, 10,
306. https://doi.org/10.3389/fnhum.2016.00306, PubMed:
27445747
Klimesch, W. (1999). EEG alpha and theta oscillations reflect
cognitive and memory performance: A review and analysis.
Brain Research Reviews, 29, 169–195. https://doi.org/10.1016
/S0165-0173(98)00056-3, PubMed: 10209231
Kuba, M., Kubová, Z., Kremlácek, J., & Langrová, J. (2007).
Motion-onset VEPs: Characteristics, metodi, and diagnostic
use. Vision Research, 47, 189–202. https://doi.org/10.1016/j
.visres.2006.09.020, PubMed: 17129593
Kuznetsova, A., Brockhoff, P. B., & Christensen, R. H. B. (2017).
lmerTest package: Tests in linear mixed effects models.
Journal of Statistical Software, 82, 1–26. https://doi.org/10
.18637/jss.v082.i13
Landers, D. M., Han, M., Salazar, W., & Petruzzello, S. J. (1994).
Effects of learning on electroencephalographic and
electrocardiographic patterns in novice archers.
International Journal of Sport Psychology, 25, 313–330.
Lee, E. C., Fragala, M. S., Kavouras, S. A., Queen, R. M., Pryor, J. L.,
& Casa, D. J. (2017). Biomarkers in sports and exercise:
Tracking health, performance, and recovery in athletes.
Journal of Strength and Conditioning Research, 31,
2920–2937. https://doi.org/10.1519/JSC.0000000000002122,
PubMed: 28737585
Levac, D. E., Glegg, S. M., Sveistrup, H., Colquhoun, H., Mugnaio,
P., Finestone, H., et al. (2016). Promoting therapists’ use of
motor learning strategies within virtual reality-based stroke
rehabilitation. PLoS One, 11, e0168311. https://doi.org/10
.1371/journal.pone.0168311, PubMed: 27992492
Levin, M. F., Weiss, P. L., & Keshner, E. UN. (2015). Emergence
of virtual reality as a tool for upper limb rehabilitation:
Incorporation of motor control and motor learning principles.
Physical Therapy, 95, 415–425. https://doi.org/10.2522/ptj
.20130579, PubMed: 25212522
Macdonald, J. S., Mathan, S., & Yeung, N. (2011). Trial-by-trial
variations in subjective attentional state are reflected in
ongoing prestimulus EEG alpha oscillations. Frontiers in
Psychology, 2, 82. https://doi.org/10.3389/fpsyg.2011.00082,
PubMed: 21687452
1268
Journal of Cognitive Neuroscience
Volume 33, Numero 7
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
C
N
_
UN
_
0
1
7
1
3
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Makeig, S. (2009). Mind monitoring via mobile brain–body
imaging. In International Conference on Foundations of
Augmented Cognition. Berlin, Heidelberg: Springer.
Makeig, S., Gramann, K., Jung, T. P., Sejnowski, T. J., & Poizner,
H. (2009). Linking brain, mind and behavior. Internazionale
Journal of Psychophysiology, 73, 95–100. https://doi.org/10
.1016/j.ijpsycho.2008.11.008, PubMed: 19414039
Makino, H., Hwang, E. J., Hedrick, N. G., & Komiyama, T.
(2016). Circuit mechanisms of sensorimotor learning.
Neuron, 92, 705–721. https://doi.org/10.1016/j.neuron.2016
.10.029, PubMed: 27883902
Meyer, D. E., Keith-Smith, J. E., Kornblum, S., Abrams, R. A., &
Wright, C. E. (1990). Speed-accuracy tradeoffs in aimed
movements: Towards a theory of rapid voluntary action.
In M. Jeannerod (Ed.), Attention and performance
(pag. 173–226). Hillsdale, NJ: Erlbaum. https://doi.org/10.4324
/9780203772010-6
Miles, H. C., Pop, S. R., Watt, S. J., Lawrence, G. P., & John, N. W.
(2012). A review of virtual environments for training in ball
sports. Computers & Graphics, 36, 714–726. https://doi.org/10
.1016/j.cag.2012.04.007
Monfared, S. S., Tenenbaum, G., & Folstein, J. R. (2019).
Anticipation in sharp shooting: Cognitive structures in
detecting performance errors. Psychology of Sport and
Exercise, 45, 101555. https://doi.org/10.1016/j.psychsport
.2019.101555
Paulus, M. P., Potterat, E. G., Taylor, M. K., Van Orden, K. F.,
Bauman, J., Momen, N., et al. (2009). A neuroscience
approach to optimizing brain resources for human
performance in extreme environments. Neuroscience and
Biobehavioral Reviews, 33, 1080–1088. https://doi.org/10
.1016/j.neubiorev.2009.05.003, PubMed: 19447132
Perez, M. A., Lundbye-Jensen, J., & Nielsen, J. B. (2006).
Changes in corticospinal drive to spinal motoneurones
following visuo-motor skill learning in humans. Journal of
Physiology, 573, 843–855. https://doi.org/10.1113/jphysiol
.2006.105361, PubMed: 16581867
R Core Team. (2019). R: A language and environment for
statistical computing. Vienna, Austria: R Foundation for
Statistical Computing. https://www.r-project.org/.
Rabbitt, P. M. (1966). Errors and error correction in choice-
response tasks. Journal of Experimental Psychology, 71,
264–272. https://doi.org/10.1037/h0022853, PubMed:
5948188
Ragan, E. D. (2010). The effects of higher levels of immersion
on procedure memorization performance and implications
for educational virtual environments. In Presence (Vol. 19,
pag. 527–543). Cambridge, MA: CON Premere. https://doi.org/10
.1162/pres_a_00016
Rao, H. M., Khanna, R., Zielinski, D. J., Lu, Y., Clements, J. M.,
Potter, N. D., et al. (2018). Sensorimotor learning during a
marksmanship task in immersive virtual reality. Frontiers in
Psychology, 9, 58. https://doi.org/10.3389/fpsyg.2018.00058,
PubMed: 29467693
Rao, H. M., Smalt, C. J., Rodriguez, A., Wright, H. M., Mehta, D. D.,
Brattain, l. J., et al. (2020). Predicting cognitive load and
operational performance in a simulated marksmanship task.
Frontiers in Human Neuroscience, 14, 222. https://doi.org/10
.3389/fnhum.2020.00222, PubMed: 32719593
Robitaille, N., Marois, R., Todd, J., Grimault, S., Cheyne, D.,
& Jolicoeur, P. (2010). Distinguishing between lateralized
and nonlateralized brain activity associated with visual
short-term memory: fMRI, MEG, and EEG evidence from
the same observers. Neuroimage, 53, 1334–1345. https://doi
.org/10.1016/j.neuroimage.2010.07.027, PubMed: 20643214
Rueda-Delgado, l. M., Solesio-Jofre, E., Serrien, D. J.,
Mantini, D., Daffertshofer, A., & Swinnen, S. P. (2014).
Understanding bimanual coordination across small time
scales from an electrophysiological perspective. Neuroscience
& Biobehavioral Reviews, 47, 614–635. https://doi.org/10
.1016/j.neubiorev.2014.10.003, PubMed: 25445184
Sanz, F. A., Multon, F., & Lécuyer, UN. (2015). A methodology for
introducing competitive anxiety and pressure in VR sports
training. Frontiers in Robotics and AI, 2, 10. https://doi.org
/10.3389/frobt.2015.00010
Schmidt, R. A., & Lee, T. D. (2011). Motor control and learning
(5th ed.). Champaign, IL: Human Kinestics.
Seo, J., Kim, Y. T., Song, H. J., Lee, H. J., Lee, J., Jung, T. D., et al.
(2012). Stronger activation and deactivation in archery
experts for differential cognitive strategy in visuospatial
working memory processing. Behavioural Brain Research,
229, 185–193. https://doi.org/10.1016/j.bbr.2012.01.019,
PubMed: 22266924
Stinson, C., & Bowman, D. UN. (2014). Feasibility of training
athletes for high-pressure situations using virtual reality. IEEE
Transactions on Visualization and Computer Graphics, 20,
606–615. https://doi.org/10.1109/TVCG.2014.23, PubMed:
24650988
Urbin, M. A., Stodden, D. F., Fischman, M. G., & Weimar, W. H.
(2011). Impulse-variability theory: Implications for ballistic,
multijoint motor skill performance. Journal of Motor
Behavior, 43, 275–283. https://doi.org/10.1080/00222895
.2011.574172, PubMed: 21598159
van den Berg, B., Appelbaum, l. G., Clark, K., Lorist, M. M.,
& Woldorff, M. G. (2016). Visual search performance is
predicted by both prestimulus and poststimulus electrical
brain activity. Scientific Reports, 6, 37718. https://doi.org/10
.1038/srep37718, PubMed: 27901053
van der Helden, J., Boksem, M. A., & Blom, J. H. (2010). IL
importance of failure: Feedback-related negativity predicts
motor learning efficiency. Cerebral Cortex, 20, 1596–1603.
https://doi.org/10.1093/cercor/bhp224, PubMed: 19840974
van Donkelaar, P., & Franks, IO. M. (1991). Preprogramming vs.
on-line control in simple movement sequences. Acta
Psychologica, 77, 1–19. https://doi.org/10.1016/0001-6918
(91)90061-4, PubMed: 1950633
van Wijk, B. C., Beek, P. J., & Daffertshofer, UN. (2012).
Differential modulations of ipsilateral and contralateral beta
(Di)synchronization during unimanual force production.
European Journal of Neuroscience, 36, 2088–2097. https://
doi.org/10.1111/j.1460-9568.2012.08122.x, PubMed:
22583034
Wagner, J., Makeig, S., Gola, M., Neuper, C., & Müller-Putz, G.
(2016). Distinct β band oscillatory networks subserving
motor and cognitive control during gait adaptation. Journal
of Neuroscience, 36, 2212–2226. https://doi.org/10.1523
/JNEUROSCI.3543-15.2016, PubMed: 26888931
Walsh, M. M., & Anderson, J. R. (2012). Learning from
experience: Event-related potential correlates of reward
processing, neural adaptation, and behavioral choice.
Neuroscience & Biobehavioral Reviews, 36, 1870–1884.
https://doi.org/10.1016/j.neubiorev.2012.05.008, PubMed:
22683741
Wolpert, D. M., & Flanagan, J. R. (2016). Computations
underlying sensorimotor learning. Current Opinion in
Neurobiology, 37, 7–11. https://doi.org/10.1016/j.conb.2015.12
.12.003, PubMed: 26719992
Wolpert, D. M., Ghahramani, Z., & Jordan, M. IO. (1995). An
internal model for sensorimotor integration. Scienza, 269,
1880–1882. https://doi.org/10.1126/science.7569931,
PubMed: 7569931
Yarrow, K., Brown, P., & Krakauer, J. W. (2009). Inside the
brain of an elite athlete: The neural processes that support
high achievement in sports. Nature Reviews Neuroscience,
10, 585–596. https://doi.org/10.1038/nrn2672, PubMed:
19571792
Liu et al.
1269
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
C
N
_
UN
_
0
1
7
1
3
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Zaepffel, M., Trachel, R., Kilavik, B. E., & Brochier, T. (2013).
Zielinski, D. J., Rao, H., Potter, N., Appelbaum, l. G., & Kopper, R.
Modulations of EEG beta power during planning and
execution of grasping movements. PLoS One, 8, e60060.
https://doi.org/10.1371/journal.pone.0060060, PubMed:
23555884
(2016). Evaluating the effects of image persistence on
dynamic target acquisition in low frame rate virtual
environments. In 3D user interfaces (3DUI), 2016 IEEE
Symposium (pag. 319–320). IEEE.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
3
7
1
2
5
3
1
9
2
1
2
8
8
/
/
j
o
C
N
_
UN
_
0
1
7
1
3
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
1270
Journal of Cognitive Neuroscience
Volume 33, Numero 7