Predication Drives Verb Cortical Signatures
Mireia Hernández1,2, Scott L. Fairhall3, Alessandro Lenci4,
Marco Baroni3, and Alfonso Caramazza2,3
Astratto
■ Verbs and nouns are fundamental units of language, Ma
their neural instantiation remains poorly understood. Neuro-
psychological research has shown that nouns and verbs can be
damaged independently of each other, and neuroimaging re-
search has found that several brain regions respond differen-
tially to the two word classes. Tuttavia, the semantic–lexical
properties of verbs and nouns that drive these effects remain
unknown. Here we show that the most likely candidate is pre-
dication: a core lexical feature involved in binding constituent
arguments (boy, candies) into a unified syntactic–semantic
structure expressing a proposition (the boy likes the candies).
We used functional neuroimaging to test whether the intrinsic
“predication-building” function of verbs is what drives the
verb–noun distinction in the brain. We first identified verb-
preferring regions with a localizer experiment including verbs
and nouns. Then, we examined whether these regions are
sensitive to transitivity—an index measuring its tendency to
select for a direct object. Transitivity is a verb-specific prop-
erty lying at the core of its predication function. Neural activ-
ity in the left posterior middle temporal and inferior frontal
gyri correlates with transitivity, indicating sensitivity to predi-
catione. This represents the first evidence that grammatical
class preference in the brain is driven by a wordʼs function
to build predication structures. ■
INTRODUCTION
Verbs and nouns represent elemental building blocks for
the grammatical rules that constrain how words can be
combined into sentences (Greenberg, 1966; Robins,
1952). The grammatical distinction between nouns and
verbs is of fundamental importance, because a wordʼs
grammatical category determines the types of phrases
in which it appears and the morphological transforma-
tions it undergoes (The boy likes the candies, but not
*The likes boy the candies; The boy liked the candies,
but not *The boyed like the candies)—see Haspelmath
(2012) for an overview of the debate about the extent
to which the verb–noun distinction is universal across
languages. There is considerable evidence showing
neural differentiation between verbs and nouns. There
are reports of grammatical category-specific deficits fol-
lowing brain damage—selective damage to either verbs
or nouns (per esempio., Hernández et al., 2008; Shapiro, Shelton,
& Caramazza, 2000; Damasio & Tranel, 1993; Caramazza
& Hillis, 1991; McCarthy & Warrington, 1985; Miceli, Silveri,
Villa, & Caramazza, 1984), and functional neuroimaging
studies have found regions that are particularly sensitive
to a wordʼs grammatical category (per esempio., Tyler, Randall, &
Stamatakis, 2008; Shapiro, Moo, & Caramazza, 2006; Perani
et al., 1999). Tuttavia, the precise basis for the observed
neural distinction remains elusive, in part because it is
1Universitat Pompeu Fabra, 2Harvard University, 3University of
Trento, 4Università di Pisa
© 2014 Istituto di Tecnologia del Massachussetts
not obvious that the observed noun–verb effects reflect
grammatical as opposed to semantic distinctions be-
tween the two word categories. Questo è, the putative gram-
matical category effects could reflect the effect of different
semantic dimensions that are correlated with grammatical
category—for example, verbs prototypically refer to actions
whose meanings include more motion features relative to
concrete objects—rather than the effect of specifically
grammatical properties.
An additional difficulty is posed by the less-than-clear
picture that has emerged of the neural correlates of the
verb–noun distinction. No systematic relationship has
been found between type of grammatical category deficit
and lesion site (see Vigliocco, Vinson, Druks, Barber, &
Cappa, 2011, for a review). And although a number of func-
tional neuroimaging studies have found verb-preferring
regions, these vary across studies (see Crepaldi, Berlingeri,
Paulesu, & Luzzatti, 2011, for a critical review challenging
the existence of a grammatical class distinction in the brain
on the basis of across-study inconsistency). Ancora, a number
of recent fMRI studies have consistently implicated the
left lateral temporal cortex (LTC), including the posterior
middle temporal gyrus (pMTG), in processing verbs rela-
tive to nouns (per esempio., Peelen, Romagno, & Caramazza,
2012; Romagno, Rota, Ricciardi, & Pietrini, 2012; Willms
et al., 2011; Burton, Krebs-Noble, Gullapalli, & Berndt,
2009; Bedny, Caramazza, Grossman, Pascual-Leone, &
Saxe, 2008; Tyler et al., 2008; Shapiro et al., 2006; Yokoyama
et al., 2006; Kable, Kan, Wilson, Thompson-Schill, &
Chatterjee, 2005) E, less consistently, different left frontal
Journal of Cognitive Neuroscience 26:8, pag. 1829–1839
doi:10.1162/jocn_a_00598
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
C
5
N
8
_
9
UN
/
_
j
0
o
0
C
5
N
9
8
_
UN
P
_
D
0
0
B
5
sì
9
G
8
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
2
io
3
e
S
/
j
F
T
/
.
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
regions. For the latter regions, some studies have not found
any effects (per esempio., Peelen et al., 2012), whereas others have
implicated middle frontal (per esempio., Willms et al., 2011; Shapiro
et al., 2006) or inferior frontal (per esempio., Bedny et al., 2008)
regions. The reliable observation of verb preference in
LTC across recent studies suggests that this region might
represent core grammatical or semantic properties of verbs.
Tuttavia, as already noted, the precise lexical-grammatical
or semantic properties that drive the preference remain to
be determined.
The LTC has not only been implicated in processing
verbs relative to nouns but also in the conceptual repre-
sentation of actions, as shown by studies that considered
the relationship between lexical deficits and lesion sites
(Kemmerer, Rudrauf, Manzel, & Tranel, 2012; Kalénine,
Buxbaum, & Coslett, 2010; Tranel, Kemmerer, Adolphs,
Damasio, & Damasio, 2003) and studies that compared
the neural activity in response to action versus different
types of nonaction words such as colors (Martin, Haxby,
Lalonde, Wiggs, & Ungerleider, 1995) or abstract words
(Noppeney, Josephs, Kiebel, Friston, & Price, 2005). Questo
pattern of results encouraged the hypothesis that differ-
ences in the type of information associated with action
and object concepts were likely to be behind LTC verb-
preferring responses. Tuttavia, recent studies have under-
mined this hypothesis. Bedny et al. (Bedny, Caramazza,
Pascual-Leone, & Saxe, 2012; Bedny et al., 2008) observed
that LTC regions that preferentially respond to verbs
relative to nouns, in both sighted and congenitally blind
individuals, were not engaged in processing visual motion
information associated with action concepts—for example,
think and cut induced similar responses. These results
show that verb preference in LTC does not reflect the
representation of sensory motor properties of action con-
cepts. Peelen et al. (2012) tested whether LTC verb prefer-
ence reflects the processing of more abstract information
prototypically associated with actions (rather than objects)
such as the understanding of actions as dynamic events
unfolding over time (which may involve visual motion
informazione [per esempio., walk] or not [per esempio., watch]). Tuttavia,
neither of two independently defined verb-preferring LTC
regions—the left STS and pMTG—responded more to
verbs or nouns referring to dynamic events (per esempio., he
watches, the accident) relative to verbs or nouns referring
to states and conditions persistent in time (per esempio., he exists,
the aptitude). Inoltre, verb-preferring responses in left
STS and pMTG were not restricted to event verbs but
showed equally greater activity for event and state verbs
relative to event and state nouns. Crucially, Peelen et al.
found greater neural activity for action relative to state
verbs in a more posterior portion of the pMTG that was
not verb preferring, implying that verbs and action con-
cepts are represented in nearby but separate regions (Vedere
Watson, Cardillo, Ianni, & Chatterjee, 2013, for a meta-
analysis suggesting that the pMTG may be a “hub” region
for action concept processing). In short, extant results
suggest that (1) the verb effect in the LTC is not driven
by the representation of semantic properties associated
with action concepts and (2) LTC verb-preferring regions
are adjacent to more posterior LTC regions representing
action concepts.
These previous studies did not provide a positive re-
sponse to the question of what dimension(S) drives verb
preference in left LTC but revealed a grammatical cate-
gory effect that is not reducible to a number of semantic
properties linked to action concepts. This led us to con-
sider the alternative hypothesis that verb preference in
the left LTC and/or the less-studied frontal regions is
driven by grammatical properties linked to the specific
role of verbs in sentence structure—that is, predication:
binding the different components of a sentence into a
unified structure expressing a meaningful proposition.
Così, Per esempio, it is the verb “like” that binds the
arguments “the boy” and “the candies” into “The boy
likes the candies,” a proposition stating a fact about the
current state of the world. It is worth stressing that we use
here the term predication to refer to this proposition-
constructing role of verbs, in line with the standard
semantic view of verbs as functions from entities to state-
ments about states of the world (per esempio., Heim & Kratzer,
1998). This function, incidentally, also distinguishes verbs
from adjectives, which cannot, by themselves, project
entities to statements. The term predication has also been
applied to other concepts in the linguistic literature (per esempio.,
by Croft, 1991, 2000, 2001)—concepts that we do not
want to imply here. The fundamental predication role of
verbs, distinguishing them from nouns, may be what
drives verb-preferring activity in LTC. If such were the
case, we should observe effects of predication in verb-
preferring regions when comparing verbs differing in
predicative function.
Ma, how does one measure the verbsʼ predicative
function? One way is through a verb-specific property—
transitivity—that is crucially related to their predication
role. Transitive verbs (per esempio., destroy) select for a direct object;
intransitive verbs do not select for a direct object—they
either do not select for any other argument besides the
subject (per esempio., die) or select for prepositional complements
(per esempio., wait for Bill ). It is worth emphasizing that the
transitive–intransitive opposition does not apply to other
grammatical categories: Transitivity is in fact related to
verb-specific morphosyntactic operations (per esempio., passiviza-
tion and accusative case assignment), which are in turn
inherently connected to the core and unique role of verbs
in forming predication. Because many verbs function as
intransitive or transitive depending on the semantic
context—to sink is intransitive in “The ship sank” but tran-
sitive in “The submarine sank the ship”—we computed a
continuous transitivity score as a probabilistic gradient
reflecting the ratio between the frequency of a verbʼs tran-
sitive uses and the frequency of the intransitive occur-
rences of the same verb.
Few previous studies have examined the neural cor-
relates of verbs differing in predicative function (cioè.,
1830
Journal of Cognitive Neuroscience
Volume 26, Numero 8
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
C
5
N
8
_
9
UN
/
_
j
0
o
0
C
5
N
9
8
_
UN
P
_
D
0
0
B
5
sì
9
G
8
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
2
io
3
e
S
/
j
T
.
/
F
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
transitive vs. intransitive). None of those studies, how-
ever, posed the question of whether predication would
be encoded in verb-specific regions, but simply aimed at
examining whether different types of verbs (per esempio., intran-
sitive vs. transitive) showed different neural correlates
anywhere in the brain. Unfortunately, extant results paint
a complicated picture about the possibility that verb-
specific LTC and frontal regions encode predication. In
two studies, Assadollahi et al. (Assadollahi, Meinzer,
Flaisch, Obleser, & Rockstroh, 2009; Assadollahi &
Rockstroh, 2008) report mixed results. In a study using
magnetoencephalography (Assadollahi & Rockstroh,
2008), they found larger effects for intransitive than
transitive verbs in left MTG and left inferior IFG (infIFG).
In a later study using fMRI (Assadollahi et al., 2009), Essi
found greater activation for intransitive than transitive
verbs in the left inferior temporal fusiform gyrus and in
the right infIFG. An equally complicated picture emerges
from the fMRI studies by Thompson and colleagues
(Thompson, Bonakdarpour, & Fix, 2010; den Ouden,
Fix, Parrish, & Thompson, 2009; Thompson et al.,
2007), who report predication effects in both directions
(intransitive > transitive and transitive > intransitive) In
a wide range of brain regions. In one study (Thompson
et al., 2007), they observed greater neural activity in the
left supramarginal and angular gyri for transitive relative
to intransitive verbs. In a follow-up study (Thompson
et al., 2010) using the same task and materials, Essi
observed similar results but this time restricted to the
angular gyrus. In a different study, Meltzer-Asscher,
Schuchard, den Ouden, and Thomphon (2013) observed
greater neural activity in the bilateral angular and supra-
marginal gyri, as well as in the right posterior superior
temporal gyrus (STG), the right MTG, and the right
middle and superior frontal gyri for verbs that can act as
transitive or intransitive versus verbs that must exclusively
act as intransitive. Tuttavia, in an fMRI study using an
overt action picture naming task (den Ouden et al.,
2009), they found a wide range of regions that respond
differentially to intransitive and transitive verbs, in both
directions: transitive > intransitive and intransitive >
transitive. Larger intransitive relative to transitive verb
effects were reported for the right hemisphere in the
thalamus, precentral gyrus, and STS (encompassing Heschlʼs
gyrus and insula) and for left hemisphere in STG extend-
ing into Heschlʼs gyrus; larger transitive relative to intran-
sitive verbs were reported in bilateral fusiform gyrus,
middle occipital gyrus (extending into inferior occipital
gyrus in the left hemisphere), pMTG and angular and
supramarginal gyri, and the precuneus, as well as left hemi-
sphere activation in the postcentral and precentral gyri,
middle frontal gyrus (MFG), and infIFG. E, finally, In
an fMRI study using written Hebrew sentences on which
participants performed a semantic judgment task, Shetreet,
Palti, Friedmann, and Hadar (2007, Experiment 1) exam-
ined whether the neural activity correlated with the num-
ber of components that each verb type could bind, Quale
included unergative (binding no components), transitive
(two components), and ditransitive (three components)
verbs. The results showed greater neural activity as a func-
tion of the number of components in brain regions typi-
cally not associated with language processing—the right
precuneus and right anterior cingulate. In contrasto, In
Experiment 2, they found that the neural activity in brain
regions more typically associated with language—left
STG and IFG—increased with the number of different
types of complements that a verb can take (per esempio., “discover”
can take a direct object [per esempio., John discovered
covered that
At least part of the inconsistency across these studies
in terms of brain regions involved in transitivity effects
may derive from undercontrolled linguistic confounds.
Per esempio, verb-specific properties related to the goal
of the actions (per esempio., telicity or subject agentivity) may
account for Thompson et al.ʼs (2007, 2010) risultati,
because the supramarginal and angular gyri have been
implicated in the neural representation of action goals
(per esempio., Oosterhof, Tipper, & Downing, 2012; Spunt,
Satpute, & Lieberman, 2011; Desmurget et al., 2009;
Hamilton & Grafton, 2006, 2008; Lestou, Pollick, &
Kourtzi, 2008; Majdandzić et al., 2007; Sirigu et al.,
2004). Allo stesso modo, the predication effects in a number of
the regions reported by den Ouden et al. (2009) may
be attributed to visual complexity confounds in their
naming task (per esempio., occipital cortex). In contrasto, the incon-
sistency across and within studies in the direction of the
transitivity effects may reflect the encoding of different
processes associated with the predication function. For
esempio, the greater neural activity for transitive relative
to intransitive verbs may reflect the processing of verb
syntactic frame complexity—that is, the set of slots rep-
resenting the different arguments that a particular verb
can bind in a sentence (per esempio., subject, direct object), con
transitive verbs being more complex than intransitive
verbs. It is unclear, Tuttavia, whether this greater com-
plexity could also be attributed to other dimensions (per esempio.,
telicity, subject agentivity) that have been undercontrolled
in previous studies. Alternatively, the opposite direction of
the effect—greater activity for intransitive relative to tran-
sitive verbs—could index processes reflecting the proto-
typicality of a verbʼs syntactic frame, with transitive verbs
being more prototypical than intransitive verbs, and there-
fore easier to process. Note that these are tentative inter-
pretations and that it is still unknown why transitivity
effects have been observed in both directions. Never-
theless, what it is relevant for the purpose of our study is
Quello (In)transitivity effects, regardless of their direction, are
related to the verbsʼ binding role of arguments. Therefore,
in this study, we measure (In)transitivity as an index of
sensitivity to predication without regard to the direction
of the effects.
To test the hypothesis that predication drives neural
verb preference, we first performed a localizer experiment
Hernández et al.
1831
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
C
5
N
8
_
9
UN
/
_
j
0
o
0
C
5
N
9
8
_
UN
P
_
D
0
0
B
5
sì
9
G
8
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
2
io
3
e
S
/
j
T
F
.
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
that included separate verb and noun conditions to loca-
lize verb-specific regions reported in earlier studies.
Then, we sought transitivity effects in those regions by
means of an experiment that included verbs varying in
the transitivity index. If verb-preferring LTC and/or fron-
tal regions respond to predication, we would expect
activity in those regions to correlate (positively or nega-
tively) with the probabilistic gradient of transitivity after
controlling for relevant intrinsic properties of words
(length, frequency, imageability, number of senses)
and difficulty of word processing (through RT). We also
controlled for other verb-specific dimensions that might
be correlated with transitivity and that could contribute
to determining the syntactic–semantic role of a particu-
lar verb. Those dimensions were telicity, dynamicity, E
subject agentivity. Telicity refers to the extent to which
an action entails a natural endpoint: telic verbs such as
arrive (someone is now at a location where she was
not before) versus atelic verbs such as walk (it describes
a way of walking without specifying any natural point at
which there is a transition from walking to not walking);
dynamicity refers to the expression of change or process:
dynamic verbs such as run or kill versus static verbs such
as exist or sleep; agentivity refers to an action intention-
ally performed by the subject: agentive verbs such as kill
(the subject decides to perform the action) versus non-
agentive verbs such as like (the subject involuntarily
experiences the action). Controlling for these dimen-
sions is of particular relevance because (UN) transitive
verbs tend to be telic (Levin & Rappaport Hovav, 2005;
Levin, 1999; Dowty, 1991); (B) subject agentivity may
also correlate with transitivity, because the subject of
static actions (per esempio., think, exist) tend to be nonintentional
and intransitive; (C) Peelen et al. (2012) observed that the
verb-specific region in the LTC responded more to states
than dynamic actions. Note that, among all these dimen-
sions, we hypothesize that predication is the property driv-
ing verb preference in the brain because of its role in
binding the different arguments of a sentence—an exclu-
sive role of verbs. Consider, Per esempio, deverbal nouns
(cioè., nouns derived from verbs) such as destruction.
Despite its belonging to the grammatical category of
nouns, this word carries semantic properties related to
the verb it was derived from, namely, “destroy,” which is
telic, dynamic, and subject agentive. Così, destruction
expresses an event entailing a natural endpoint (something
has been destroyed) that unfolded over time and was
probably performed intentionally by a subject. Tuttavia,
in its grammatical quality of noun, destruction lacks the
function of building a predication nexus like the one
expressed by a verb in a sentence: Both “Alexanderʼs
destruction of the city” and “Alexander destroyed the
city” refer to the same event, but only the latter expresses
a full-fledged meaningful proposition. Nevertheless,
because all verb-specific dimensions are interrelated, we
also examine whether any verb-preferring region that
responds to transitivity also represents some of these
other syntactic–semantic dimensions related to verb
processing.
METHODS
Participants
Fourteen right-handed healthy volunteers (10 women;
mean age = 25 years, range = 18–38 years) took part in
the lexical decision and the localizer experiments. All par-
ticipants gave written informed consent for their par-
ticipation. The experimental procedures were approved
by the ethical committee for experiments involving hu-
mans at the University of Trento.
Functional Localizer
To identify cortical regions more sensitive to verb relative
to noun processing, we used two runs (∼8 min each) Di
a version of the paradigm used in Peelen et al. (2012).
Stimuli consisted of 21 Italian verbs and 21 Italian nouns
presented in two different syntactic contexts—as single
parole (per esempio., create, book) or short phrases (per esempio., she
creates, the book). The two syntactic contexts were not
analyzed separately in this study. Verbs were different
from those included in the main experiment and referred
to actions and events. Nouns referred to objects of dif-
ferent semantic categories with the exception of tools
because they elicit action concepts, which activate por-
tions of the LTC (per esempio., Kemmerer et al., 2012; Kable
et al., 2005). Verbs and nouns were matched for length
in letters (verbs: 7.25, nouns: 7.38; T(249) < 1). Verbs
had slightly higher logarithmic (log) lemma frequency
values than nouns (verbs: 4.49, nouns: 4.32; t(249) =
2.11, p < .036). Note, however, that any potential verb-
specific effect cannot be driven by difficulty confounds
related to this difference in log frequency, because verbs
were more frequent than nouns. Because the localizer
was meant to replicate the typical verb-preferring find-
ings reported in the literature—that is, regardless of tran-
sitivity effects—we did not control for verb transitivity.
Participants indicated whether a memory probe was iden-
tical to one of the three previous stimuli by pressing the
right/left buttons on a response pad. In an event-related
design, each run consisted of 42 blocks. Each block con-
sisted of four stimuli of one of the two conditions (nouns,
verbs), with each condition occurring 21 times within a
run in a random order. The fourth stimulus was the
memory probe, which was presented in green instead
of white. The first three stimuli were visually displayed
for 1.5 sec, each followed by a 0.5-sec interval. After an
extra 1.5-sec interval, the memory probe was presented
for 1 sec, followed by an intertrial interval of 2.5 sec. For
“identical” trials, the memory probe stimulus was ran-
domly selected from one of the three preceding stimuli;
for nonidentical trials, the memory probe stimulus was
randomly selected from the remaining set of 18 stimuli.
1832
Journal of Cognitive Neuroscience
Volume 26, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
c
5
n
8
_
9
a
/
_
j
0
o
0
c
5
n
9
8
_
a
p
_
d
0
0
b
5
y
9
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
One of the runs was administered first, and the other last
in the experimental session, with the main experiment in
between.
to the number of times the same verb occurs with a direct
object. The transitivity score ranged from 0 to 1, with higher
values for strongly intransitive verbs and lower values for
strongly transitive ones.
Main Experiment
We used a lexical decision task (four runs of ∼6 min). The
experimental stimuli consisted of 200 Italian verbs, pre-
sented in the gerund or indicative mood with two sepa-
rate runs for each grammatical mood—none of the
verbal forms used could serve as both nouns and verbs.
The two verb moods were not analyzed separately in this
study. In addition, 100 pseudoverbs were created by
replacing one letter in the stem of half of the experimen-
tal verbs. Participants indicated whether the letter string
presented in each trial was an Italian verb or not by
pressing the right/left buttons on a response pad. In an
event-related design, letter strings were visually dis-
played for 1 sec followed by 1 sec of fixation cross (+).
Thirty-eight null events consisting of a fixation cross for
2 sec were randomly interspersed with the letter strings.
One hundred verbs and 50 pseudoverbs were randomly
presented within each run. That is, the order in which
participants were presented with verbs, pseudoverbs,
and null events was left random. Each verb and pseudo-
verb was presented once within a run but twice across
the whole experiment (once in gerund and once in
one of six alternative forms of the indicative mood).
Values for different variables were collected for the 200
experimental verbs. Except for length and number of
senses, multiple decimal places were used. Imageability
values were collected through 19 native Italian speakers
rating whether each verb was easy to imagine or not and
averaging their choices to obtain an imageability score
ranging between 0 and 1, with higher values for highly
imageable verbs—it is of note that, although a binary
scale was used for this rating, variations in participantsʼ
judgments of the imageability of specific verbs yield an
approximately uniform, continuous distribution in the
imageability values used in this study. Log lemma fre-
quency was estimated on the 1.5 billion-word itWac
corpus (Baroni, Bernardini, Ferraresi, & Zanchetta, 2009).
The number of senses of each verb was derived from the
Italian WordNet (Pianta, Bentivogli, & Girardi, 2002).
Values for telicity, dynamicity, and subject agentivity were
collected through five linguistsʼ ratings. Telicity values
were obtained through a two-choice decision and ranged
from 0 to 1, with higher values for telic verbs. Subject
agentivity and dynamicity values were obtained through a
three-choice decision, allowing the rating of few ambig-
uous cases and ranged from 1 to 3, with higher values
for strongly nonintentional subject and strongly dynamic
actions, respectively. The transitivity index was computed
over the 325 million-word La Repubblica Corpus, using
LexIt (Lenci, Lapesa, & Bonansinga, 2012) as A/A + B,
where A refers to the number of times a given verb occurs
with no other arguments besides the subject and B refers
Data Acquisition
Functional and structural data were collected with a Bruker
BioSpin MedSpec 4T scanner (Bruker BioSpin GmbH,
Rheinstetten, Germany) at the Center for Mind/Brain
Sciences, University of Trento, Italy. Functional images
were acquired using echo-planar T2*-weighted scans. We
used 43 axial slices acquired in ascending interleaved order
(repetition time = 2000 msec, echo time = 33 msec, flip
angle = 73°, field of view = 192 mm, matrix = 64 × 64,
voxel size = 3 × 3 × 3 mm), covering the whole cerebral
cortex. Structural images were acquired with a T1-weighted
3-D MP-RAGE sequence (sagittal slice orientation, voxel
resolution = 1 × 1 × 1 mm, field of view = 256 × 224, GRAPPA
acquisition with acceleration factor of 2, repetition time =
2700 msec, inversion time = 1020 msec, flip angle = 7°).
Data Analysis
Data were analyzed using SPM8 software (www.fil.ion.ucl.
ac.uk/spm/). After the first four volumes were discarded to
avoid T1 saturation, functional data were slice time-
corrected, realigned to the first volume, motion-corrected,
normalized into standard stereotactic space using the
Montreal Neurological Institute (MNI) template, and
smoothed with a Gaussian kernel (6 mm FWHM). The time
series were high-filtered to eliminate low-frequency com-
ponents (filter 128 sec). Then the data were modeled
based on a conventional general linear model (GLM) for
each participant, convolving events with a canonical hemo-
dynamic response function and including the motion
parameters as regressors of no interest.
Functional Localizer
Events were 2-sec long and consisted of each word and
short phrase including memory probes. In the first-level
analysis, each participantʼs GLMs were created to model
single verbs, verb phrases, single nouns, and noun
phrases separately. After estimating the model and extract-
ing β-weights, contrasts against baseline were extracted
for these four conditions. Then, a subject factor and the
four conditions were included in a group-level flexible
factorial design. We performed a whole-brain F-contrast
to look for verb-specific clusters that allowed us to define
the ROIs—the Verb > Noun contrast, collapsing single
words and short phrases.
Verb-preferring ROIs
To define the ROIs, we first considered the group-level
clusters significantly activated for the Verbs > Nouns
Hernández et al.
1833
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
C
5
N
8
_
9
UN
/
_
j
0
o
0
C
5
N
9
8
_
UN
P
_
D
0
0
B
5
sì
9
G
8
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
2
io
3
e
S
/
j
F
.
T
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
Tavolo 1. Verb-preferring Regions
Brain Region
L MTG
L superior frontal/bilateral cingulate gyrus
L IFG
MFG
BA
22
21
32
32
6
6
44
44
47
44
6
6
6
Cluster Size
Cluster, pFWE
Voxel Z
265
246
.0001
.0001
160
.001
5.4
4.87
5.03
4.35
4.23
3.37
4.53
3.8
3.79
3.64
4.3
4.3
4.19
MNI Coordinates
X
−48
−54
−6
9
−6
−6
−54
−51
−48
−54
−36
−39
−45
sì
−43
−28
17
20
8
11
17
14
32
17
−1
2
5
z
4
−2
40
40
55
64
19
4
−2
1
34
37
40
Regions more active for verbs than nouns in a whole-brain analysis of the localizer experiment.
BA = Brodmannʼs area; Cluster, pFWE = cluster-level, P < .05 FWE-corrected. Subregions denote local maxima; Voxel Z = peak voxel Z score.
contrast of the localizer experiment using a voxelwise
threshold of p < .001 (uncorrected) and a clusterwise
threshold of p < .05 (family-wise error [FWE]-corrected).
There were four clusters showing a greater response to
verbs than to nouns. Those clusters included the left
MTG (BA 21 and BA 22), the left superior frontal gyrus
extending into the bilateral cingulate gyri (BA 6 and
BA 32), the left IFG (BA 44 and 47), and the left MFG
(BA 6; see Table 1). Because the greater activation of
left superior frontal regions does not seem to reflect
grammatical class-specific processes (Willms et al.,
2011) and the cingulate gyrus has not been reported as
playing a role in the verb versus noun distinction, we did
not consider those areas for the ROIs definition. This
criterion reduced the probability of including ROIs where
the higher activity for verbs relative to nouns is because
of any potential confound unrelated to verb processing.
Then, we defined six spheres of 7-mm radius around peak
coordinates so that most of the cluster was included
while avoiding ROIs overlapping. We only considered
Table 2. ROI Selection
MNI Coordinates
Location of the Spheres
x
y
z
Verb > Baseline Contrast Value
T
SE
P
L MTG
pMTG
midMTG
L IFG
supIFG
infIFG(UN)
infIFG(B)
L MFG
−48
−54
−54
−48
−54
−45
−43
−28
17
32
17
5
4
−2
19
−2
1
40
1.45
1.03
1.46
0.15
0.83
2.84
3.78
3.07
3.36
0.48
2.32
6.69
0.384
0.336
0.435
0.313
0.358
0.425
.00024
.0026
.001
.63
.022
.000001
We defined 7-mm spheres in three clusters showing greater response to verbs than to nouns in the localizer experiment. Spheres were defined
around peak coordinates of those clusters avoiding overlap. Any sphere not showing greater response to the contrast verbs versus baseline in the
main experiment ( P < .05 uncorrected for multiple comparisons) was excluded as ROI (i.e., infIFG(a)).
1834
Journal of Cognitive Neuroscience
Volume 26, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
c
5
n
8
_
9
a
/
_
j
0
o
0
c
5
n
9
8
_
a
p
_
d
0
0
b
5
y
9
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Table 3. Lexicality Effect
Brain Region
L angular gyrus/STG
L superior frontal gyrus/MFG
BA
39
39
8
10
9
Cluster Size
Cluster, pFWE
Voxel Z
120
325
.01
.0001
6.16
5.10
5.29
4.65
4.64
MNI Coordinates
x
−48
−42
−15
−6
−6
y
−70
−58
38
62
56
z
34
31
46
7
25
Regions more active for verbs than pseudoverbs in a whole-brain analysis of the main experiment.
BA = Brodmannʼs area; Cluster, pFWE = cluster-level, p < .05 FWE-corrected. Subregions denote local maxima; Voxel Z = peak voxel Z score.
as ROIs those spheres showing greater response to verbs
versus baseline ( p < .05, uncorrected for multiple com-
parisons) in the ROI analysis of the main experiment,
using the same flexible factorial design by which we
estimated linear increases in activation during verb pro-
cessing as a function of the parametric modulators (see
below; see Table 2). This criterion allowed us to include
all ROIs that were at least minimally sensitive to verb
processing during the lexical decision task, resulting in
five final ROIs (see Results section).
Main Experiment
Events were 2-sec long and consisted of each verb pre-
sented during the experiment. Telicity, log lemma fre-
quency, number of senses, imageability, length, response
latencies (RTs), subject agentivity, event dynamicity, and
transitivity were included as parametric modulators.
Because parametric modulators are serially orthogonalized
with respect to those previously entered in the GLM, the
modulator of interest (i.e., transitivity) was entered last
while the order of the remaining eight was left random.
Pseudoverbs in the two grammatical moods and errors
were modeled separately. β-weights estimated from each
subjectʼs GLM were used to create statistical maps of
contrasts of interest. For verb processing-related activity,
we contrasted verbs and pseudoverbs against baseline.
The effect of lexicality was estimated in a group-level
analysis by including a subject factor and a condition
factor (verbs, pseudoverbs) in a flexible factorial design.
We performed whole-brain F-contrast to assess the verb
processing-related activity compared with that of pseudo-
verbs (i.e., the effect of lexicality) using a voxelwise thresh-
old of p < .001 (uncorrected) and a clusterwise threshold
of p < .05 (FWE-corrected). To estimate linear increases
in activation during verb processing as a function of the
parametric modulators, we contrasted each of them across
baseline—contrasts were performed considering only
verbs (i.e., verbs and pseudoverbs were never collapsed).
All contrasts were combined in a group-level model based
on a flexible factorial design that included a subject factor
along with the “verb” condition as well as the nine para-
metric modulators. We used MarsBaR toolbox (Brett,
Anton, Valabregue, & Poline, 2002) to obtain the mean
parameter estimates at the group level for the contrasts
of the nine parametric modulators within each of the
five predefined ROIs. Results were considered at an α of
0.001, based on Bonferroni correction for multiple com-
parisons, taking into account 45 comparisons (5 ROIs,
8 control modulators, 1 parametric modulator of interest
[transitivity], p = .05/45). Note that, because of the ortho-
gonalization process, only the potential effect of the para-
metric modulator entered last in the GLM (i.e., transitivity)
persists independently of any other modulator. In the case
of the control modulators, however, any potential effect
could be driven by the influence of any modulator entered
later in the GLM. For the sake of completeness, in case any
control modulator showed a significant linear correlation
with activity in any ROI, we redid the analyses placing that
modulator last.
RESULTS
Behavioral Results
Responses were faster and more accurate for verbs
(820 msec, 93.8% correct) than pseudoverbs (927 msec,
84.2% correct; both p < .0001). Neither RT (r = −.14, p <
.296) nor accuracy (r = −.007, p < .591) correlated with
the parametric modulator of interest (i.e., transitivity).
Whole-brain Analysis
We observed an expected effect of lexicality reflected by
greater neural activity for verbs relative to pseudoverbs in
the left angular gyrus extending into the STG and the left
superior frontal gyrus, including MFG areas (see Table 3).
But no modulation of the neural activity as a function of
transitivity was observed anywhere in the brain.
Verb-preferring ROIs
We were able to localize the most commonly reported
verb-preferring region—the left LTC—that we divided
Hernández et al.
1835
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
c
5
n
8
_
9
a
/
_
j
0
o
0
c
5
n
9
8
_
a
p
_
d
0
0
b
5
y
9
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
in two ROIs, the pMTG [−48 −43 4] and the middle MTG
(midMTG) [−54 −28 −2]. Additionally, we also identified
verb-preferring left frontal regions whose detectability has
been rather elusive in previous functional neuroimaging
studies. Those regions were the MFG [−45 5 40], infIFG
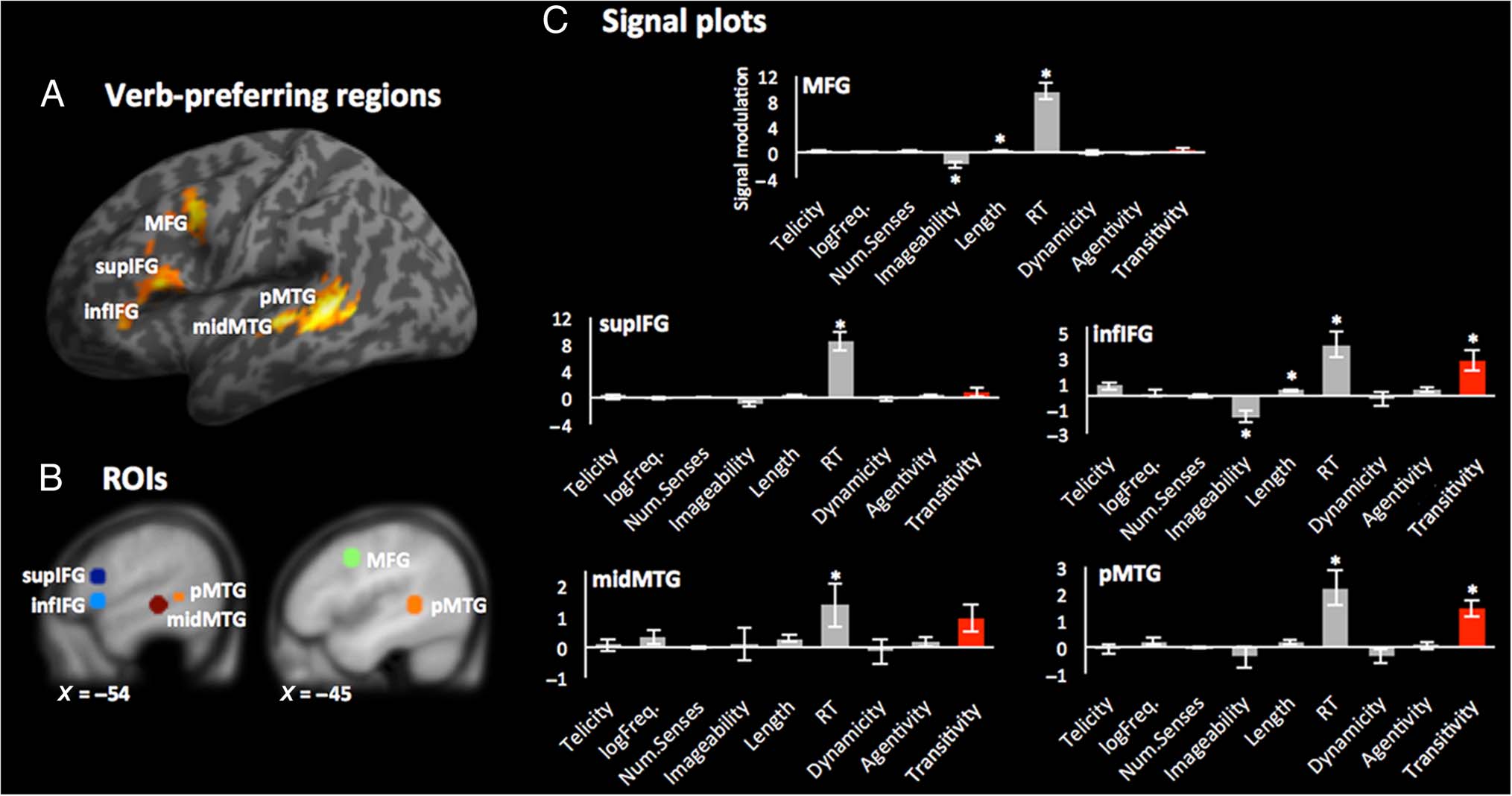
[−54 17 1], and superior IFG (supIFG) [−54 17 19] (Fig-
ure 1A, B).
Are Verb-preferring ROIs Sensitive to Transitivity?
Once controlling for all other factors, we observed a clear
linear increase of activity as a function of transitivity in
one LTC ROI, the pMTG ( p < .000004). The correlation
between activity and transitivity also survived the thresh-
old imposed by the correction for multiple comparisons
( p < .001; see Methods) in one frontal ROI, the infIFG
( p < .0004). The correlation between activity and tran-
sitivity reflected greater activity for strongly intransitive
relative to strongly transitive verbs in these regions. No
other verb-preferring ROI was influenced by transitivity
(all ps > .03; Figure 1C).
Effect of Variables Other than Transitivity
Neural activity was not modulated by any other semantic–
syntactic property of verbs (cioè., telicity, dynamicity, E
subject agentivity) in any verb-preferring region. How-
ever, the activity in the two verb-preferring ROIs sensitive
to transitivity also showed influence of RT (pMTG and
infIFG), imageability (infIFG), and length (infIFG; Figure 1C).
When redoing the analyses placing each of these variables
as the last parametric modulator in the GLM (see Methods
for details), we still observed the effects of RT and length
in infIFG (and a marginal RT effect in pMTG), as well as
effects of RT in other verb-preferring ROIs not sensitive
to transitivity—the MFG and supIFG. In contrasto, the effects
of imageability in infIFG vanished and were not observed
in any other ROI (Vedi la tabella 4).
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
DISCUSSION
This study was designed to explore which verb property
is encoded (relative to nouns) in verb-preferring regions.
We hypothesized that the fundamental syntactic–semantic
distinction between verbs and nouns—predication—is
what drives the widely observed verb-preferring activity.
Because the predication-building function is limited to
verbs, this question can only be addressed by testing
whether verb-preferring regions distinguish between
verbs with different predicative properties. To test this
hypothesis, we first replicated previous studies showing
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
C
5
N
8
_
9
UN
/
_
j
0
o
0
C
5
N
9
8
_
UN
P
_
D
0
0
B
5
sì
9
G
8
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
2
io
3
e
S
/
j
.
F
T
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
Figura 1. (UN) Verb-preferring clusters resulting from contrasting verbs with nouns in the localizer experiment that were considered for ROI
definition. (B) Verb-preferring ROIs defined as 7-mm spheres around peak coordinates of the clusters shown in A. (C) Signal plots for the modulation
of the BOLD response as a function of transitivity (red bars) and all control modulators (gray bars). Note: Because of the serial orthogonalization
of the modulators in the GLM, they were entered in the order (da sinistra a destra) that is shown in the plots—leaving transitivity last to make sure its
effects persisted independently of any control modulator (see Methods; Vedi la tabella 4 for the results of the complementary analyses in which we
placed last in the GLM those control modulators showing here a significant linear correlation with activity). Error bars represent the SEM. *p value
fulfills correction for multiple comparisons ( P < .001; see Methods).
1836
Journal of Cognitive Neuroscience
Volume 26, Number 8
Table 4. Effect of Variables Other than Transitivity
ROI
MFG
Variable
Signal
T
SE
p
RT
Length
9.8
0.26
6.76
1.45
.000001a
2.61
0.1
.01
Imageability −0.97 −1.66
0.584
.099
supIFG
RT
Length
8.51
0.18
6.44
1.321
.000001a
1.85
0.097
Imageability −0.12 −0.21
0.571
.06
.83
infIFG
RT
Length
3.9
0.42
3.99
0.977
.0001a
3.6
0.117
.00045a
Imageability −1.16 −1.14
1.018
.25
midMFG RT
Length
Imageability
pMTG
RT
Length
Imageability
1.55
0.25
0.37
2.41
0.15
0.03
1.94
0.799
.054
2.6
0.096
0.47
0.787
.01
.64
3.29
0.733
.0013
1.97
0.076
0.05
0.6
.05
.96
We re-did the analysis three times, each placing last one of the three
parametric modulators of no interest that showed a significant linear
correlation with activity in any ROI in the first analysis (i.e., RT, length,
imageability). Because of the orthogonalization process (see Methods),
this action allowed us to examine the effects of these three variables
unaffected by any other modulator.
aRegion survives correction for multiple comparisons ( p < .001; see
Methods).
verb-preferring responsivity in the left LTC and in the
less-consistently observed frontal regions by contrasting
verbs with nouns in a localizer experiment. Then we
examined whether different ROIs within those verb-
preferring regions (LTC: pMTG, midMTG; frontal regions:
MFG, infIFG, supIFG) were sensitive to the property of
predication by seeking effects of verb transitivity (here
defined as a gradient contrast between transitive and
intransitive verbs) on neural activity. The results showed
that, after controlling for relevant intrinsic properties of
words and task difficulty, the activity in two verb-preferring
ROIs—pMTG and infIFG—showed a positive correlation
with transitivity. No other semantic–syntactic property of
verbs (i.e., telicity, dynamicity, and subject agentivity) inde-
pendently modulated activity in any ROI.
To the extent that (in)transitivity is a core aspect of
predication structures defined by verbs, the correlation
between transitivity and activity in pMTG and infIFG
indicates that the role of verbs as “predication builders”—a
role that nouns do not take—drives the grammatical class
distinction in those regions. This hypothesis is silent on the
specific direction of the effect: It does not speak to why
neural activity increased for intransitive relative to transitive
verbs (see also Assadollahi et al., 2009; den Ouden et al.,
2009; Assadollahi & Rockstroh, 2008). Nonetheless, as
argued in the Introduction, the correlation between tran-
sitivity and neural activity may be tentatively interpreted
in terms of verb prototypicality. The lower neural activity
in pMTG and infIFG for transitive versus intransitive verbs
would reflect the fact that verbs binding agents and objects
(transitive) are easier to process in those regions because
of their higher prototypicality compared with verbs that do
not take a direct object (intransitive).
A question that arises when interpreting the role of
pMTG in representing a wordʼs predication property is
how this hypothesis fits with previous data implicating this
region in processing semantic information of actions (e.g.,
Kemmerer et al., 2012; Kalénine et al., 2010; Noppeney
et al., 2005; Tranel et al., 2003; Martin et al., 1995). However,
as already noted, we have found in several studies (Bedny
et al., 2008, 2012; Peelen et al., 2012; Shapiro et al., 2006)
that the verb-preferring regions do not show greater pref-
erence for action verbs over other verb types. Indeed, if
anything, state verbs and abstract verbs, more generally,
produce greater activation in these regions (Peelen et al.,
2012). The latter results are consistent with a broader char-
acterization of the role of pMTG in the representation of
verbs. The predication function of verbs does not merely
consist in linking arguments within a sentence but also in
determining the roles and relations that arguments (typi-
cally nouns) take—verbs inform about what an agent does
(e.g., John runs), its properties (e.g., John stinks), or what it
does to or with something else (e.g., John rides the bike).
This makes predication a semantic–syntactic interface
where semantic information is crucial to determine that,
for instance, the syntactic structure “The boy likes” is not
felicitous because the verb “like” needs an object “to be
liked.” It is then plausible that the pMTG, as a region in-
volved in representing word meaning, provides the relevant
neural substrate for the representation of the semantic–
syntactic interface captured by a verbʼs predication role.
Transitivity is related to the number of arguments
required by a verb. However, verbs are not the only cate-
gory that can project an argument structure. An interest-
ing question that follows from our results is whether the
pMTG and infIFG would respond more to those nouns
that also express argument structure to some degree (e.g.,
Alexiadou & Grimshaw, 2008). Consider, for example,
father (e.g., Johnʼs father—where father expresses a rela-
tionship between two agents; in this context, it is interesting
to note that, in some languages, kinship relations are
expressed by verbs; Evans, 2000), author (e.g., John is
the author of a best seller—where author embeds the
concept of an agent writing something), or killer (e.g.,
The killer of the man—where killer is a deverbal noun
derived from the verb kill ). It is worth considering for
future research whether brain regions sensitive to the pre-
dicative role of verbs respond differentially to nouns with
or without argument structure. Alternatively, it is possible
that those regions only respond to predication in the
narrower sense that we suggested in the Introduction—
the possibility of creating propositional expressions of
Hernández et al.
1837
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
c
5
n
8
_
9
a
/
_
j
0
o
0
c
5
n
9
8
_
a
p
_
d
0
0
b
5
y
9
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
some state of affairs that can be true or false (e.g., The man
was killed )—which is exclusively reserved to verbs. For
instance, the noun phrase The killer of the man contains
a predicative noun, but it does not express a state of affairs,
because it only refers to an entity. Instead, the sentence
The man was killed is a statement that can be true or false.
Moreover, it is worth stressing that the direct object of the
transitive verb kill is obligatory (The soldier killed the
man vs. *The soldier killed ), whereas the argument of
the corresponding deverbal noun is fully optional.
Both the pMTG (marginally) and infIFG were also sensi-
tive to RT, and the infIFG was also sensitive to length. How-
ever, because the effect of predication survived after
regressing out both RT and word length, we may confi-
dently conclude that the observed transitivity effect in
these regions is not because intransitive verbs are asso-
ciated with higher RTs or length. In addition, other verb-
preferring regions not sensitive to transitivity also showed
sensitivity to RT (see Table 4), indicating that the presence
or absence of the transitivity effect was independent of
whether the ROIs also responded to other variables. In
any case, the orthogonalization process used in this experi-
ment (see Methods for details) was designed precisely to
ensure that any surviving factor, such as transitivity, cannot
be explained as the influence of a control modulator.
Finally, the contrast verbs versus nouns in the localizer
experiment also gave significant verb-preferring activity
in three other ROIs (midMTG, MFG, and supIFG). No
effect of transitivity or any other verb-specific property
(i.e., telicity, dynamicity, and subject agentivity) was ob-
served in these regions. Therefore, at present, we do not
have enough information to assess what grammatical dis-
tinction, if any, these regions might capture, leaving the
question of what role these areas might play in determin-
ing their verb specificity to future research.
In short, we have observed that two verb-preferring
regions—the left pMTG and infIFG—are sensitive to tran-
sitivity as a measure of the verb-specific predication func-
tion. Following the rationale that verbsʼ fundamental role
is that of predication whereas nouns lack this property,
we can conclude that verb preference in these regions is
driven by this syntactic–semantic function. These results
represent the first positive evidence on the question of
what grammatical class distinctions are captured in the
brain. In addition, our results reveal that, at least in the
posterior LTC and inferior frontal cortex, these distinctions
reflect the representation of grammatical (verbs vs. nouns)
and not semantic properties associated with action con-
cepts (e.g., dynamicity). Further research is needed to
shed light into how other verb-preferring regions capture
the distinction between verbs and nouns.
Acknowledgments
This research was supported by the Provincia Autonoma di
Trento and the Fondazione Cassa di Risparmio di Trento e
Rovereto. This research was also supported by a Marie Curie
International Outgoing Fellowship within the 7th European
Community Framework Programme (FP7/2007-2013) under
grant agreement no. 271671 awarded to Mireia Hernández.
Reprint requests should be sent to Mireia Hernández, Cognitive
Neuropsychology Laboratory, Harvard University, William James
Hall, Room 918, 33 Kirkland Street, Cambridge, MA 02138, or
via e-mail: mireiahp@gmail.com.
REFERENCES
Alexiadou, A., & Grimshaw, J. (2008). In F. Schäfer (Ed.), Verbs,
nouns and affixation (pp. 1–16). Working Papers of the SFB
732. Incremental Specification in Context 01 (2008).
Assadollahi, R., Meinzer, M., Flaisch, T., Obleser, J., &
Rockstroh, B. S. (2009). The representation of the verbʼs
argument structure as disclosed by fMRI. BMC Neuroscience,
10, doi:10.1186/1471-2202-10-3.
Assadollahi, R., & Rockstroh, B. S. (2008). Representation of the
verbʼs argument-structure in the human brain. BMC
Neuroscience, 9, doi:10.1186/1471-2202-9-69.
Baroni, M., Bernardini, S., Ferraresi, A., & Zanchetta, E. (2009).
The WaCky wide web: A collection of very large linguistically
processed web-crawled corpora. Language Resources &
Evaluation, 43, 209–226.
Bedny, M., Caramazza, A., Grossman, E., Pascual-Leone, A., &
Saxe, R. (2008). Concepts are more than percepts: The case
of action verbs. Journal of Neuroscience, 28, 11347–11353.
Bedny, M., Caramazza, A., Pascual-Leone, A., & Saxe, R. (2012).
Typical neural representations of action verbs develop
without vision. Cerebral Cortex, 22, 286–293.
Brett, M., Anton, J. L., Valabregue, R., & Poline, J. B. (2002).
Region of interest analysis using an SPM toolbox [abstract].
Presented at the 8th International Conference on Functional
Mapping of the Human Brain, June 2–6, 2002, Sendai, Japan.
Neuroimage. 16(2): Elsevier.
Burton, M. W., Krebs-Noble, D., Gullapalli, R. P., & Berndt, R. S.
(2009). Functional neuroimaging of grammatical class:
Ambiguous and unambiguous nouns and verbs. Cognitive
Neuropsychology, 26, 148–171.
Caramazza, A., & Hillis, A. (1991). Lexical organisation of nouns
and verbs in the brain. Nature, 349, 788–790.
Crepaldi, D., Berlingeri, M., Paulesu, E., & Luzzatti, C. (2011).
A place for nouns and a place for verbs? A critical review of
neurocognitive data on grammatical-class effects. Brain and
Language, 116, 33–49.
Croft, W. (1991). Syntactic categories and grammatical
relations. Chicago: University of Chicago Press.
Croft, W. (2000). Parts of speech as typological universals and as
language particular categories. In P. Vogel & B. Comrie (Eds.),
Approaches to the typology of word classes (pp. 65–102).
Berlin: Mouton de Gruyter.
Croft, W. (2001). Radical construction grammar. Oxford, UK:
Oxford University Press.
Damasio, A. R., & Tranel, D. (1993). Nouns and verbs are retrieved
with differently distributed neural systems. Proceedings of the
National Academy of Sciences, U.S.A., 90, 4957–4960.
den Ouden, D. B., Fix, S., Parrish, T. B., & Thompson, C. K. (2009).
Argument structure effects in action verb naming in static and
dynamic conditions. Journal of Neurolinguistics, 22, 196–215.
Desmurget, M., Reilly, K. T., Richard, N., Szathmari, A.,
Mottolese, C., & Sirigu, A. (2009). Movement intention after
parietal cortex stimulation in humans. Science, 324, 811–813.
Dowty, D. (1991). Thematic proto-roles and argument
selection. Language, 67, 547–619.
Evans, N. (2000). Kinship verbs. In P. Vogel & B. Comrie (Eds.),
Approaches to the typology of word classes (pp. 103–173).
Berlin: Mouton de Gruyter.
1838
Journal of Cognitive Neuroscience
Volume 26, Number 8
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
c
5
n
8
_
9
a
/
_
j
0
o
0
c
5
n
9
8
_
a
p
_
d
0
0
b
5
y
9
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Greenberg, J. H. (1966). Some universals of grammar with
Peelen, M. V., Romagno, D., & Caramazza, A. (2012).
particular reference to the order of meaningful elements.
In J. H. Greenberg (Ed.), Universals of language (2nd ed.,
pp. 73–113). Cambridge, MA: MIT Press.
Hamilton, A. F., & Grafton, S. T. (2006). Goal representation in
human anterior intraparietal sulcus. Journal of Neuroscience,
26, 1133–1137.
Hamilton, A. F., & Grafton, S. T. (2008). Action outcomes are
represented in human inferior frontoparietal cortex. Cerebral
Cortex, 18, 1160–1168.
Haspelmath, M. (2012). How to compare major word-classes
across the worldʼs languages. In T. Graf, D. Paperno,
A. Szabolcsi, & J. Tellings (Eds.), Theories of everything: In
honor of Edward Keenan (pp. 109–130) (UCLA Working
Papers in Linguistics, 17). Los Angeles: UCLA.
Heim, I., & Kratzer, A. (1998). Semantics in generative grammar.
Oxford, UK: Blackwell.
Hernández, M., Caño, A., Costa, A., Sebastián-Gallés, N.,
Juncadella, M., & Gascón-Bayarri, J. (2008). Grammatical
category-specific deficits in bilingual aphasia. Brain and
Language, 107, 68–80.
Kable, J. W., Kan, I. P., Wilson, A., Thompson-Schill, S. L., &
Chatterjee, A. (2005). Conceptual representations of action in
the lateral temporal cortex. Journal of Cognitive Neuroscience,
17, 1855–1870.
Kalénine, S., Buxbaum, L. J., & Coslett, H. B. (2010). Critical
brain regions for action recognition: Lesion symptom
mapping in left hemisphere stroke. Brain, 133, 3269–3280.
Kemmerer, D., Rudrauf, D., Manzel, K., & Tranel, D. (2012).
Behavioral patterns and lesion sites associated with impaired
processing of lexical and conceptual knowledge of actions.
Cortex, 48, 826–848.
Lenci, A., Lapesa, G., & Bonansinga, G. (2012). LexIt: A
computational resource on Italian argument structure. In
Proceedings of LREC 2012. Istanbul, Turkey.
Lestou, V., Pollick, F. E., & Kourtzi, Z. (2008). Neural substrates
for action understanding at different description levels in the
human brain. Journal of Cognitive Neuroscience, 20, 324–341.
Levin, B. (1999). Objecthood: An event structure perspective.
CLS, 35, 223–247.
Levin, B., & Rappaport Hovav, M. (2005). Argument realization,
research surveys in linguistics series. Cambridge, UK:
Cambridge University Press.
Majdandzić, J., Grol, M. J., Van Schie, H. T., Verhagen, L., Toni, I.,
& Bekkering, H. (2007). The role of immediate and final goals
in action planning: An fMRI study. Neuroimage, 37, 589–598.
Independent representations of verbs and actions in the left
lateral temporal cortex. Journal of Cognitive Neuroscience,
24, 2096–2107.
Perani, D., Cappa, S. F., Schnur, T., Tettamanti, M., Collina, S.,
Rosa, M. M., et al. (1999). The neural correlates of verb and
noun processing. A PET study. Brain, 122, 2337–2344.
Pianta, E., Bentivogli, L., & Girardi, C. (2002). MultiWordNet:
Developing an aligned multilingual database. In Proceedings
of the First Global WordNet Conference. Mysore, India.
Robins, R. H. (1952). Noun and verb in universal grammar.
Language, 28, 289–298.
Romagno, D., Rota, G., Ricciardi, E., & Pietrini, P. (2012). Where
the brain appreciates the final state of an event: The neural
correlates of telicity. Brain and Language, 123, 68–74.
Shapiro, K. A., Moo, L. R., & Caramazza, A. (2006). Cortical
signatures of noun and verb production. Proceedings of the
National Academy of Sciences, U.S.A., 103, 1644–1649.
Shapiro, K., Shelton, J., & Caramazza, A. (2000). Grammatical
class in lexical production and morphological processing:
Evidence from a case of fluent aphasia. Cognitive
Neuropsychology, 17, 665–682.
Shetreet, E., Palti, D., Friedmann, N., & Hadar, U. (2007).
Cortical representation of verb processing in sentence
comprehension: Number of complements, subcategorization,
and thematic frames. Cerebral Cortex, 17, 1958–1969.
Sirigu, A., Daprati, E., Ciancia, S., Giraux, P., Nighoghossian, N.,
Posada, A., et al. (2004). Altered awareness of voluntary
action after damage to the parietal cortex. Nature
Neuroscience, 7, 80–84.
Spunt, R. P., Satpute, A. B., & Lieberman, M. D. (2011).
Dissociable neural systems support retrieval of how and
why action knowledge. Journal of Cognitive Neuroscience,
23, 63–74.
Thompson, C. K., Bonakdarpour, B., & Fix, S. F. (2010).
Neural mechanisms of verb argument structure processing
in agrammatic aphasic and healthy age-matched listeners.
Journal of Cognitive Neuroscience, 22, 1993–2011.
Thompson, C. K., Bonakdarpour, B., Fix, S. C., Blumenfeld,
H. K., Parrish, T. B., Gitelman, D. R., et al. (2007). Neural
correlates of verb argument structure processing. Journal of
Cognitive Neuroscience, 19, 1753–1767.
Tranel, D., Kemmerer, D., Adolphs, R., Damasio, H., &
Damasio, A. R. (2003). Neural correlates of conceptual
knowledge for actions. Cognitive Neuropsychology, 20,
409–432.
Martin, A., Haxby, J. V., Lalonde, F. M., Wiggs, C. L., &
Tyler, L. K., Randall, B., & Stamatakis, E. A. (2008). Cortical
Ungerleider, L. G. (1995). Discrete cortical regions associated
with knowledge of color and knowledge of action. Science,
270, 102–105.
McCarthy, R. A., & Warrington, E. K. (1985). Category specificity
in an agrammatic patient: The relative impairment of verb
retrieval and comprehension. Neuropsychologia, 23, 709–727.
Meltzer-Asscher, A., Schuchard, J., den Ouden, D. B., &
Thompson, C. K. (2013). The neural substrates of complex
argument structure representations: Processing “alternating
transitivity” verbs. Language and Cognitive Processes, 28,
1154–1168.
Miceli, G., Silveri, M. C., Villa, G., & Caramazza, A. (1984). On
the basis of agrammaticsʼ difficulty in producing main verbs.
Cortex, 20, 217–220.
Noppeney, U., Josephs, O., Kiebel, S., Friston, K. J., & Price, C. J.
(2005). Action selectivity in parietal and temporal cortex.
Brain Research, Cognitive Brain Research, 25, 641–649.
Oosterhof, N. N., Tipper, S. P., & Downing, P. E. (2012).
Viewpoint (in)dependence of action representations: An
MVPA study. Journal of Cognitive Neuroscience, 24, 975–989.
differentiation for nouns and verbs depends on
grammatical markers. Journal of Cognitive Neuroscience,
20, 1381–1389.
Vigliocco, G., Vinson, D. P., Druks, J., Barber, H., & Cappa,
S. F. (2011). Nouns and verbs in the brain: A review of
behavioural, electrophysiological, neuropsychological and
imaging studies. Neuroscience and Biobehavioral Reviews,
35, 407–425.
Watson, C. E., Cardillo, E. R., Ianni, G. R., & Chatterjee, A.
(2013). Action concepts in the brain: An activation likelihood
estimation meta-analysis. Journal of Cognitive Neuroscience,
25, 1191–1205.
Willms, J. L., Shapiro, K. A., Peelen, M. V., Pajtas, P. E., Costa, A.,
Moo, L. R., et al. (2011). Language-invariant verb processing
regions in Spanish–English bilinguals. Neuroimage, 57,
251–261.
Yokoyama, S., Miyamoto, T., Riera, J., Kim, J., Akitsuki, Y., Iwata,
K., et al. (2006). Cortical mechanisms involved in the
processing of verbs: An fMRI study. Journal of Cognitive
Neuroscience, 18, 1304–1313.
Hernández et al.
1839
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
6
/
8
2
6
1
/
8
8
2
/
9
1
1
8
9
2
4
9
7
/
8
1
4
7
7
8
o
1
c
5
n
8
_
9
a
/
_
j
0
o
0
c
5
n
9
8
_
a
p
_
d
0
0
b
5
y
9
g
8
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1