Patterns of Modulation in the Activity and Connectivity
of Motor Cortex during the Repeated Generation
of Movement Sequences
Ella Gabitov1, David Manor1,2, and Avi Karni1,2
Astratto
■ It is not clear how the engagement of motor mnemonic
processes is expressed in online brain activity. We scanned par-
ticipants, using fMRI, during the paced performance of a finger-
to-thumb opposition sequence (FOS), intensively trained a day
earlier (T-FOS), and a similarly constructed, but novel, untrained
FOS (U-FOS). Both movement sequences were performed in
pairs of blocks separated by a brief rest interval (30 sec). We have
recently shown that in the primary motor cortex (M1) motor
memory was not expressed in the average signal intensity but
rather in the across-block signal modulations, questo è, when com-
paring the first to the second performance block across the
brief rest interval. Here, using an M1 seed, we show that for
the T-FOS, the M1–striatum functional connectivity decreased
across blocks; Tuttavia, for the U-FOS, connectivity within the
M1 and between M1 and striatum increased. Inoltre, In
M1, the pattern of within-block signal change, but not signal
variability per se, reliably differentiated the two sequences.
Only for the U-FOS and only within the first blocks in each pair,
the signal significantly decreased. No such modulation was
found within the second corresponding blocks following the
brief rest interval in either FOS. We propose that a network
including M1 and striatum underlies online motor working
memory. This network may promote a transient integrated
representation of a new movement sequence and readily re-
trieves a previously established movement sequence representa-
zione. Averaging over single events or blocks may not capture
the dynamics of motor representations that occur over multiple
timescales. ■
INTRODUCTION
The primary motor cortex (M1) not only controls specific
movements (Georgopoulos, Kalaska, Caminiti, & Massey,
1982) but also coordinates among them to generate
meaningful sequences (Ben-Shaul et al., 2004; Tanji,
2001; Carpenter, Georgopoulos, & Pellizzer, 1999; Karni
et al., 1998; Nudo, Milliken, Jenkins, & Merzenich, 1996).
There is evidence suggesting that lower-level motor areas,
including M1, not only generate the pattern of muscle
activity necessary to implement action plans but may also
play an active role in both the acquisition and retention
of complex motor skills in mammalian brains (Peters,
Chen, & Komiyama, 2014; Yang et al., 2014; Xu et al.,
2009; Yang, Pan, & Gan, 2009; Matsuzaka, Picard, & Strick,
2007; Ben-Shaul et al., 2004; Kleim et al., 2004; Carpenter
et al., 1999; Nudo et al., 1996). Animal studies indicate
that practice on a motor task may lead to rapid, Ma
long-lasting, synaptic reorganization in M1 ( Yang et al.,
2009, 2014; Xu et al., 2009). These experience-driven
synaptic modifications did not occur with motor activity
alone (Yang et al., 2014; Xu et al., 2009) and were cor-
related with delayed behavioral improvement (Xu et al.,
2009; Yang et al., 2009). Human studies using repetitive
1University of Haifa, Haifa, Israel, 2C. Sheba Medical Center, Ramat
Gan, Israel
© 2015 Istituto di Tecnologia del Massachussetts
TMS and anodal transcranial direct current stimulation
suggest that increasing the excitability of M1 during prac-
tice can improve motor sequence learning (Saucedo
Marquez, Zhang, Swinnen, Meesen, & Wenderoth, 2013;
Kantak, Mummidisetty, & Stinear, 2012; Stagg, Jayaram,
et al., 2011; Vines, Nair, & Schlaug, 2006; Kim, Park, Ko,
Jang, & Lee, 2004; Nitsche et al., 2003). These beneficial
effects on motor performance were specific to the trained
movement sequence (Stagg, Jayaram, et al., 2011; Nitsche
et al., 2003) and could not be induced by elevating excit-
ability in premotor and prefrontal cortices (Kantak et al.,
2012; Nitsche et al., 2003). Inoltre, fMRI resting
state studies have shown an increase in the amplitude of
signal fluctuation within the contralateral M1 as well as
changes in its functional connectivity following a single
session of motor training, suggesting early experience-
dependent changes in the representation of the move-
ments within the primary motor cortex (Tung et al., 2013;
Vahdat, Darainy, Milner, & Ostry, 2011).
Imaging studies in which multiple training sessions
were afforded suggested, in line with animal studies
(Matsuzaka et al., 2007; Kleim et al., 2004; Nudo et al.,
1996), that a learning-related relative enhancement in the
extent of the average M1 signal for a trained versus an
untrained movement sequence may become apparent
only after multisession training (Steele & Penhune, 2010;
Journal of Cognitive Neuroscience 27:4, pag. 736–751
doi:10.1162/jocn_a_00751
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
C
7
N
4
_
9
UN
/
_
j
0
o
0
C
7
N
5
1
_
UN
P
_
D
0
0
B
7
sì
5
G
1
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
F
T
/
.
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
Floyer-Lea & Matthews, 2005; Penhune & Doyon, 2002;
Karni et al., 1995, 1998). Tuttavia, early phases of prac-
tice on motor sequences were reported to be reflected
in decreased (Floyer-Lea & Matthews, 2005), increased
(Orban et al., 2011; Penhune & Doyon, 2005), or un-
changed (Steele & Penhune, 2010; Toni, Krams, Turner,
& Passingham, 1998; Karni et al., 1995) magnitude of the
mean BOLD-fMRI signal in the M1 contralateral to the
performing hand. Decreasing activation putatively reflects
reduced recruitment of unspecific neuronal resources,
whereas increasing activation is thought to reflect the re-
cruitment and evolution of additional neuronal substrates
with practice; the former presumably relate to the setting
of more efficient task representation with repeated ex-
perience (Poldrack, 2000) whereas the latter relate to the
establishment and development of task-specific represen-
tations with continued practice (Karni et al., 1998). How-
ever, the setting up of more efficient representation of
the skill may occur concurrently with increased neural re-
cruitment, making the interpretation of learning-related
changes in the averaged activity difficult. An alternative
approach was proposed (Gabitov, Manor, & Karni, 2014;
Karni et al., 1998) to assess the short-term dynamics, Piuttosto
than the averaged activity, in a given area or circuit. Corto-
term dynamic changes in M1 have been shown to be
induced by both the excitation of premotor cortical effer-
ents (Davare, Montague, Olivier, Rothwell, & Lemon, 2009;
Bestmann et al., 2008; Davare, Lemon, & Olivier, 2008) E
by the repetition of experience upon the repeated genera-
tion of a movement sequence after a brief interval of rest
(Gabitov et al., 2014; Karni et al., 1995). A short-term but
reproducible reduction in activity as a function of task
repetition (repetition suppression, RS) was shown to occur
in M1 in a variety of motor tasks (Chouinard & Goodale,
2009; Hamilton & Grafton, 2009; Dinstein, Hasson, Rubin,
& Heeger, 2007; Grafton & Hamilton, 2007; Karni et al.,
1995). Così, the pattern and magnitude of short-term
brain activity modulations upon task repetition, RS or
repetition enhancement (RE), rather than the averaged
evoked signal per se, may constitute a signature for the
level of experience with specific movement sequence.
We have recently shown that in M1 the previous expe-
rience with a motor sequence was not expressed in the
average signal intensity but rather in reproducible signal
modulations when comparing activity in performance
blocks before and after a brief rest interval (Gabitov
et al., 2014). Here, we explored changes in connectivity
in performance blocks before and after a brief rest in-
terval, between an M1 seed and other brain regions. Noi
tested whether novelty or experience is reflected in mod-
ulations of connectivity across blocks by comparing an
untrained to a trained movement sequence. Inoltre,
we tested the hypothesis that short-term signal modula-
zioni, within performance blocks as well as across blocks,
follow a consistent pattern and may provide a signature
for the engagement of motor mnemonic processes during
early motor sequence learning in M1. Finalmente, we tested
whether changes in the temporal variability of neural ac-
tivity reflect the accumulation of experience with the task
(Lui, 2013; Garrett, Kovacevic, McIntosh, & Grady, 2010,
2011; Stein, Gossen, & Jones, 2005). In the scanner, par-
ticipants performed two movement sequences, a novel,
untrained and a previously trained sequence (U-FOS and
T-FOS, rispettivamente). Both sequences were composed of
the same component movements and were performed
at an identical, paced rate, using the left, nondominant
hand.
METHODS
The data from the study reported by Gabitov et al. (2014)
were used in the current study.
Participants
Thirty-two healthy young adults participated in the current
study for payment: 17 participants (19–35 years, 25.7 ±
4.4, mean ± SD, five women) in the fMRI group and 15 par-
ticipants (n = 15, 20–35 years, 25.47 ± 2.73, mean ± SD,
eight women) in the control group. Both groups were
trained and behaviorally tested in an identical protocol,
but only participants of the fMRI group underwent an
additional imaging session. Così, the control group was
tested to evaluate the possible effects on subsequent per-
formance of the additional experience afforded during
the fMRI session. Two participants from the fMRI group
were not included in the analysis: One had difficulties with
executing the task in the scanner, and another withdrew
from the fMRI session for personal reasons. All participants
reported no prior history of neurological or psychiatric
illness or brain injury and no addiction to drugs, alcohol,
or cigarettes (nonsmokers or occasional smokers). Exclu-
sion criteria included current or chronic use of medica-
zione, any known learning disabilities and attention-deficit
disorder. Only individuals with little (less than 2 years)
or no formal music training participated in the current
study. Professional typists were excluded as well. All par-
ticipants affirmed that they had no sleep disorders and
reported at least 6 hr of proper night sleep during the
study period. Each participant was identified as strongly
right-handed using the Edinburg Handedness Inventory
(Oldfield, 1971). Before the study, all participants gave writ-
ten informed consent according to a protocol approved
by the C. Sheba Medical Centerʼs Ethics Committee.
Design and Procedures
Participants were trained to accurately perform a given
five-element finger-to-thumb opposition sequence (FOS),
either sequence A or sequence B, with their nondominant
left hand (Figure 1A). Both sequences consisted of iden-
tical component movements and were mirror-reversed
in relation to each other. Così, the two sequences were
matched for the number of movements per digit and
Gabitov, Manor, and Karni
737
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
C
7
N
4
_
9
UN
/
_
j
0
o
0
C
7
N
5
1
_
UN
P
_
D
0
0
B
7
sì
5
G
1
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
.
/
T
F
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
The performance test for each condition included four
consecutive blocks of 30 sec of duration separated by
30 sec of rest intervals. Before each test block, partici-
pants were asked to perform the movement sequence,
and the block was initiated only after the FOS was ac-
curately reproduced three times. Each test block was
initiated and terminated by an auditory “READY” and
“STOP” signal, rispettivamente. Participants were instructed
to perform the sequence continuously “as fast and as
accurately as possible.” Participants were instructed that
in case of an error being noted “not to correct errors
but rather to continue from the initial movement of the
assigned sequence as smoothly as possible.” No feedback
on performance was provided. The participantsʼ perfor-
mance during the test blocks was recorded by a video
camera and scored offline. For each test block, two mea-
sures of performance were determined from these record-
ing: (1) the number of correctly completed sequences
as a measure of speed and (2) the number of incorrect
sequences (errors) as a measure of accuracy.
Before the overnight performance tests, participants of
the fMRI group took part in a scanning session, wherein
they were asked to perform either the novel sequence
(U-FOS) or the sequence trained the day before (T-FOS),
using their trained (left) hand. The untrained (right)
hand was subsequently tested as well; the results are to
be reported elsewhere. The imaging session consisted of
three consecutive runs for each sequence (Figure 1C). In
Da questa parte, potential effects of proactive interference and
contextual retrieval that could be caused by switching
between the two sequences were minimized (Kiesel
et al., 2010; Cothros, Köhler, Dickie, Mirsattari, & Gribble,
2006). The order of sequences was counterbalanced
across participants. Experimental runs (each 144 sec long)
were separated by a 1.5- to 2-min break, which included
a verbal interaction with the participant. The component
movements of the sequences were paced by an auditory
signal at a fixed rate of 1.66 Hz to control rate-related
changes in the BOLD signal (Rao et al., 1996). The paced
performance enabled the assessment of signal differences
as a function of the order of the component movements
minimizing potential differences between the U-FOS and
T-FOS that were expected to result from training on one
but not on the other sequence (Korman et al., 2003; Karni
et al., 1995) as well as minimizing differences in perfor-
mance rates between individuals. Each imaging run was
initiated only after the explicitly designated FOS was ac-
curately reproduced three times. The run consisted of
two performance blocks (Block1 and Block2) separated
by a rest interval of 30 sec. Each block was initiated by an
auditory and visual “READY” cue (2 sec), after which par-
ticipants performed the required FOS continuously in a
paced manner for a total of eight repetitions of the FOS
(24 sec). The end of the performance block was marked
by an auditory and visual “STOP” cue (1 sec).
The participantsʼ performance during the fMRI ses-
sion was recorded by a video camera focused on the
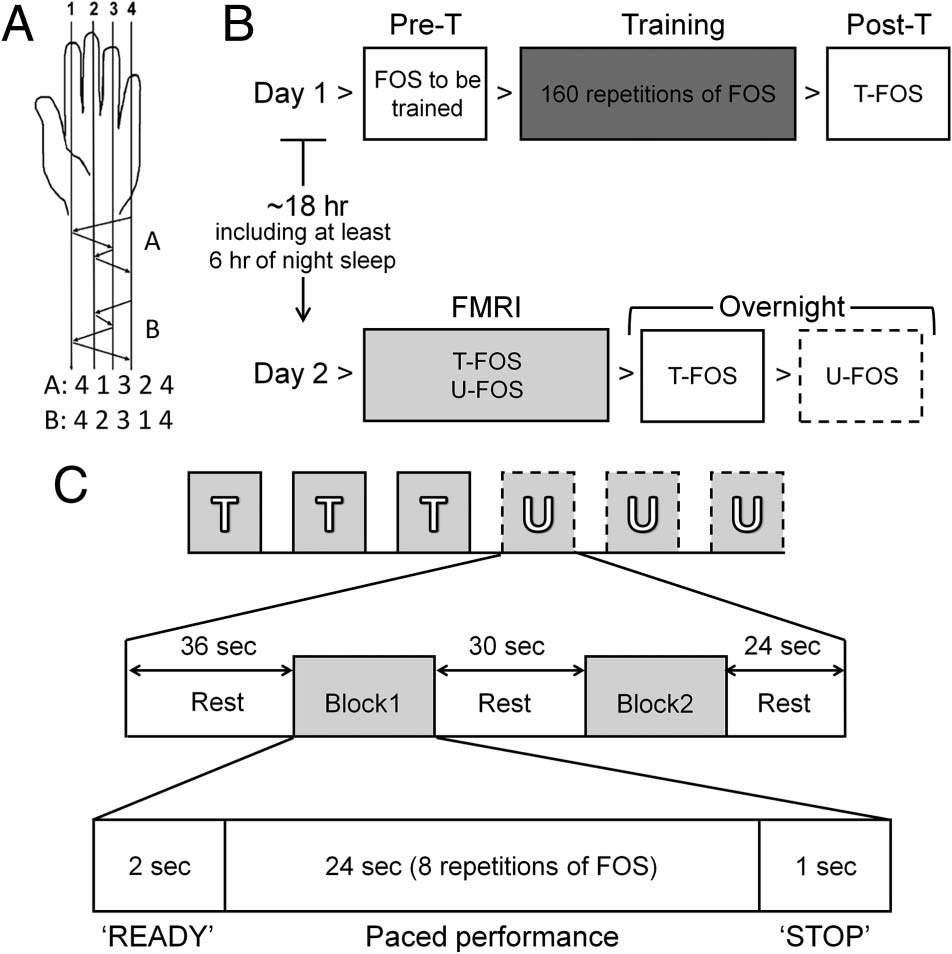
Figura 1. Study design. (UN) FOS. The two sequences were matched
for a number of movements per digit and mirror-reversed in relation
to each other (in terms of order). (B) The overall study design. Day 1:
a pretraining performance test (Pre-T), a structured training session
(Training: 10 blocks, 16 repetitions of a given sequence per block),
and an immediate posttraining performance test (Post-T). Day 2:
performance tests of the trained sequence and the untrained sequence
(Overnight: T-FOS and U-FOS, rispettivamente). Only participants of the
fMRI group took part in the scanning session (fMRI), immediately
preceding overnight performance tests. The control (non-fMRI) group
was tested to evaluate the effect of the additional experience afforded
during the fMRI session on subsequent performances. (C) The fMRI
session design. T = T-FOS; U = U-FOS; Block1, Block2 = two blocks
of FOS performance. Note that both sequences were performed at
an identical auditory-paced rate of 1.66 Hz per movement.
differed only in their order. If the sequence assigned for
training was A (T-FOS), then sequence B was used as
the novel untrained sequence (U-FOS) and vice versa.
The movement sequence was randomly assigned and
explicitly instructed. In all sessions and tests, the partici-
pants performed the instructed movement sequence lying
supine. The executing hand was positioned beside the
trunk in direct view (palm-up) of a video camera to allow
the recording of all digit movements. Visual feedback was
not afforded at any time.
Each participant took part in two study sessions con-
ducted on two consecutive days, separated by 18 hr of in-
terval that included nocturnal sleep (Figure 1B). On Day 1,
all participants were trained and tested according to a
standard FOS training protocol (Korman et al., 2007;
Korman, Raz, Flash, & Karni, 2003). For details, see our
previous report (Gabitov et al., 2014). On the second
day, all participants were retested on the performance of
the trained sequence and then the untrained sequence
(Overnight: T-FOS and U-FOS, rispettivamente) using the
trained (left) as well as the untrained (right) hand. The re-
sults for the untrained hand will be reported elsewhere.
738
Journal of Cognitive Neuroscience
Volume 27, Numero 4
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
C
7
N
4
_
9
UN
/
_
j
0
o
0
C
7
N
5
1
_
UN
P
_
D
0
0
B
7
sì
5
G
1
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
F
.
T
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
performing hand and evaluated by at least one trained
observer, online and offline. Performance was evaluated
for accuracy, timing (cioè., initiation and termination of
FOS performance) and performance rate to ensure an
appropriate task execution. Errors occurred very rarely
and when noted by the experimenters or the partici-
pants, the run was repeated. No additional errors were
observed during evaluation of performance offline. Only
runs with errorless performance were included in the
analyses. This experiment was realized using Cogent
2000 developed by the Cogent 2000 team at the FIL
and the ICN and Cogent Graphics developed by John
Romaya at the LON, Wellcome Department of Imag-
ing Neuroscience and implemented in MATLAB (IL
Mathworks, Inc., Natick, MA).
fMRI Data Acquisition and Analyses
Acquisition Parameters
fMRI scanning was carried out at the C. Sheba Medical
Center, Tel-Hashomer, using a 3-T whole-body high-
definition system (GE Excite 3 HD, Fairfield, CT) equipped
with an eight-channel head coil. A high-resolution full-
brain 3-D structural images were acquired in the axial
orientation using a T1*-weighted echo-planar sequence
(repetition time = 7.3 msec, echo time = 3 msec, flip
angle = 20°, field of view = 256 × 256 mm2, matrix size =
256 × 256 voxels, voxel size = 1 × 1 × 1 mm3). BOLD-
sensitive functional images were obtained using a gradient-
echo planar T2*-sequence (repetition time = 3000 msec,
echo time = 35 msec, flip angle = 90°, field of view = 220 ×
220 mm2, matrix size = 64 × 64 voxels, voxel size = 3.4 ×
3.4 × 3.4 mm3, no gap, ascending) con 40 axial oblique
slices, covering the whole brain.
Preprocessing
The structural and functional images were converted to
Neuroimaging Informatics Technology Initiative (NIfTI)
format using MRIcron (University of South Carolina). Pre-
processing and statistical analysis of the data were carried
out with Statistical Parametric Mapping (SPM8; Wellcome
Department of Cognitive Neurology, London, UK) oper-
ating under Matlab R2012a. For each run, the four initial
scans were discarded to allow for magnetic saturation
and equilibration effects. All images were reorientated
to stereotactic space. Functional data were realigned
and unwarped, adjusting for interactions between move-
ment and local field inhomogeneity (Hutton, Andersson,
Deichmann, & Weiskopf, 2013; Andersson, Hutton,
Ashburner, Turner, & Friston, 2001). Normalization to
the Montreal Neurological Institute (MNI) space was
performed using parameters obtained from the seg-
mentation procedure of the structural data, following co-
registration. The normalized functional images were
resampled to voxel dimensions of 3 mm3. Finalmente, func-
tional images were spatially smoothed with a Gaussian
kernel of 8 mm FWHM. Before statistical analyses, head
motion artifact detection was applied to the preprocessed
data using the Artifact Detection Tools (Mazaika, Hoeft,
Glover, & Reiss, 2009). No significant head motion artifacts
were detected (normalized z-threshold = 2, movement
threshold = 2 mm, rotation threshold = 0.05 rad).
Whole-brain Analyses
Statistical analyses of BOLD signal changes were per-
formed using a general linear model (GLM; Friston et al.,
1995). Individual models were specified separately for
each sequence (U-FOS, T-FOS) using a multisession de-
sign while each session included data from a single run
(three runs). Regressors of interest for each performance
block (Block1, Block2) were modeled as a boxcar func-
tion with a length of 24 sec convolved with the canonical
hemodynamic response function. A high pass filter of
128 sec was used to remove low-frequency noise. For
the block design, inclusion of motion covariates has a del-
eterious impact on GLM sensitivity when even moderate
correlation existed between motion and the experimental
progetto (Johnstone et al., 2006). Therefore, movement
parameters derived from realignment of the functional
volumes were not included as covariates. Following the
model parameters estimation, a linear contrast for perfor-
mance-related changes in BOLD-fMRI signal was defined
for each sequence versus rest (U-FOS > Rest, T-FOS >
Rest). The individual contrast images were introduced to
a second-level random effects analysis, separately for each
sequence, using a one-sample t test.
ROI Definition
Because of intersubject anatomical variability (Nieto-
Castañón & Fedorenko, 2012; Fedorenko, Hsieh, Nieto-
Castañón, Whitfield-Gabrieli, & Kanwisher, 2010; Woods,
1996) and the variability in the representation of hand
movements within the motor cortex of a given individual
(Nudo et al., 1996; Schlaug, Knorr, & Seitz, 1994; Nudo,
Jenkins, Prejean, & Grenda, 1992), the ROI within M1
was defined in each individual brain using a combined
anatomical and functional approach. The central sulcus
and the hand knob (Yousry et al., 1997), contralateral to
the performing hand, were used for anatomical identifica-
tion of the primary motor hand area of each individual.
The functional voxels relevant to the task performance
were identified on an individual level from activation maps
of a whole-brain analysis for each sequence (U-FOS >
Rest, T-FOS > Rest) using family-wise error rate (FWE)
correction at p < .05. The MNI coordinates of the most
active voxel (local maxima) within the right M1 hand
area, corresponding to the performing hand, were then
extracted for each individual and each sequence. Indi-
vidual ROIs were defined as a sphere centered at mean
Gabitov, Manor, and Karni
739
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
MNI coordinates across the two sequences with a radius
of 6 mm.
Functional Connectivity Analyses
A seed-driven approach was applied to explore changes
in functional connectivity during FOS performance. Indi-
vidual ROIs within M1 were used as a seed. Connectivity
analyses on preprocessed functional images were run
using the Functional Connectivity Toolbox (Conn) for
SPM (Whitfield-Gabrieli & Nieto-Castanon, 2012). This
toolbox allows condition-dependent functional connectiv-
ity analysis (e.g., for resting state network analyses and
block design studies). Before connectivity analyses, the
data underwent additional temporal preprocessing. Six
parameters obtained by rigid body head motion correc-
tion (three rotation and three translation parameters) plus
six additional parameters representing the corresponding
first-derivative terms were used as temporal covariates to
reduce the impact of motion within performance blocks.
Main effect of block may affect within-block connectivity
estimates in the presence of possible voxel-specific differ-
ences in hemodynamic delay. Therefore, main effect of
each block (Block1, Block2; each block 24 sec long con-
volved with the canonical hemodynamic response func-
tion) and the corresponding first-derivative terms were
included as additional temporal confounding factors.
Temporal covariates were removed from the BOLD func-
tional data using linear regression. The resulting residual
BOLD time series were band-pass filtered (0.008 Hz <
f < 0.1 Hz).
The preprocessed BOLD time series were divided into
scans associated with each block (Block1, Block2) sepa-
rately for each sequence (U-FOS, T-FOS). To take into
account the hemodynamic delay, block regressors were
convolved with a canonical hemodynamic response
function and rectified. Temporal connectivity maps were
generated for each block (Block1, Block2) separately for
each sequence (U-FOS, T-FOS) by estimating Pearsonʼs
correlation coefficients between the BOLD signal from
the seed region (i.e., ROI within the M1) and that at every
other brain voxel. All seed-to-voxel correlation coefficients
were converted to normally distributed scores using Fisherʼs
transformation to allow for second level GLM analyses. The
whole-brain connectivity patterns with the M1 seed were
tested in second-level analyses for the main effect of
sequence (U-FOS > T-FOS, U-FOS < T-FOS), the main
effect of block (Block1 > Block2, Block1 < Block2), and
directional sequence by block interactions, that is, testing
for a greater increase in connectivity (with the M1 seed)
across blocks, during the U-FOS performance com-
pared with the T-FOS performance ([U-FOS: Block1 <
Block2] × [T-FOS: Block1 > Block2]) and vice versa
([U-FOS: Block1 > Block2] × [T-FOS: Block1 < Block2]).
Connectivity maps generated from the second level
GLM analyses were thresholded at p ≤ .001 and overlaid
on the mean structural image of all participants or the
surface rendered from the participantsʼ mean structural
segmented images using SPM8 and Functional Imaging
Visualization Environment (FIVE; nmr.mgh.harvard.edu/
harvardagingbrain/ People/AaronSchultz/OrthoView.
html). Statistical inferences were performed on the peak-
level using p values FWE-corrected for multiple compar-
isons over a small VOI. Areas of interest for small volume
corrections were defined, for structures within the motor-
related (Hardwick, Rottschy, Miall, & Eickhoff, 2013;
Halsband & Lange, 2006) network, as follows: (1) the right
sensorimotor cortex, defined as a union between the right
primary sensory and motor cortices using human motor
area template (Mayka, Corcos, Leurgans, & Vaillancourt,
2006) as well as (2) the right and (3) the left putamen
using automated anatomical labeling (AAL; Tzourio-
Mazoyer et al., 2002). Clusters that survived p < .05
(uncorrected) on the cluster level were reported as well.
Finally, Fisher-transformed correlation coefficients were
calculated for clusters, wherein connectivity with the M1
seed showed significant sequence by block interaction.
These correlation coefficients were entered to Statistical
Package for the Social Sciences (SPSS Statistics for
Windows, Version 19.0; IBM Corp., Armonk, NY ) for
post hoc analyses applying paired samples t tests. Statistical
inferences were performed at 0.05 level corrected for
multiple tests (i.e., the number of clusters) using Bonferroni
adjustments.
Time-course Analyses
Raw ROI time courses were extracted from preprocessed
functional images for each run using the MarsBar tool-
box for SPM (Brett, Anton, Valabregue, & Poline, 2002).
These raw BOLD signals were converted to percent signal
change. To reduce the low-frequency noise because of
the scanner drift, the BOLD signal at the performance
blockʼs onset (“READY” cue) was used as the blockʼs
baseline. To explore within-block dynamics each block
was divided into two equal phases (Phase1, Phase2), each
consisting of four successive time points (signal measure-
ments), with Phase1 beginning 6 sec after the “READY”
cue and Phase2 including the “STOP” cue; exclusion of
time points corresponding to the first 6 sec following
the “READY” cue minimized the effects of hemodynamic
delay.
Analyses of temporal BOLD signal variability were per-
formed on BOLD signals converted to percent signal
change relative to the mean evoked signal across all time
points for each run. To assess the magnitude of temporal
BOLD signal variability, we used the mean squared suc-
cessive difference (SSD) measure ( Von Neumann, Kent,
Bellinson, & Hart, 1941). Mean SSD was suggested as a
more reliable estimator of the true underlying temporal
variability of the BOLD signal, compared with SD, with no
need for particular assumption about a functional form of
the expected signal drift or variation of the mean signal
across conditions (Mohr & Nagel, 2010). Individual mean
740
Journal of Cognitive Neuroscience
Volume 27, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
SSD of percent signal change were calculated separately
for each phase within each block (i.e., Phase1 and Phase2
separately for Block1 and Block2) as well as for each rest
interval (before Block1, ending in the first “READY” cue;
between Block1 and Block2, starting 9 sec after the first
“STOP” cue and ending in the second “READY” cue; and
after Block2, starting 9 sec after the second “STOP” cue
and including the last scan; exclusion of time-points cor-
responding to the first 9 sec following the “STOP” cue
allowed for the hemodynamic response to return to
baseline). In addition, BOLD signal variability was analyzed
in terms of SD for each period of interest (Garrett et al.,
2010, 2011). The analyses were designed as within-subject
comparisons. Repeated-measures ANOVAs or paired
samples t tests were run using SPSS. The results were cor-
rected for nonsphericity violation using the Greenhouse–
Geisser adjustment.
Behavioral Data Analyses
For each test block two performance measures for each
individual were determined: the number of correctly
completed sequences as a measure of speed and the
number of sequences with ordering errors as a measure
of accuracy. For statistical analyses, these measures were
averaged across the four test blocks for each perfor-
mance test. In addition, the slope for speed as a mea-
sure of within-test improvement and the SD for speed
as a measure of within-test variability were determined.
The slope was calculated as a gradient of linear regres-
sion line through 4 data points; each point represented
speed achieved during one test block. The SD for speed
within each performance test was converted to per-
centages relative to individual mean speed achieved in
that specific test. Unless otherwise stated, the analyses
were designed as within-subject comparisons. Separate
repeated-measures ANOVAs for each performance mea-
sure with sequence (U-FOS, T-FOS) as within-subject fac-
tor were run using SPSS. The results were corrected for
nonsphericity violation using the Greenhouse–Geisser
adjustment.
RESULTS
fMRI Results
The whole-brain analyses did not show significant dif-
ferences in the performance-driven changes in neural
activity between the two sequences (Gabitov et al., 2014).
Group effects of performance-related increases in neural
activity for each sequence (U-FOS > Rest, T-FOS > Rest)
are shown in Figure 2A.
ROI Definition
The MNI coordinates of the most active voxel within the
right hemisphere hand knob (Yousry et al., 1997), questo è,
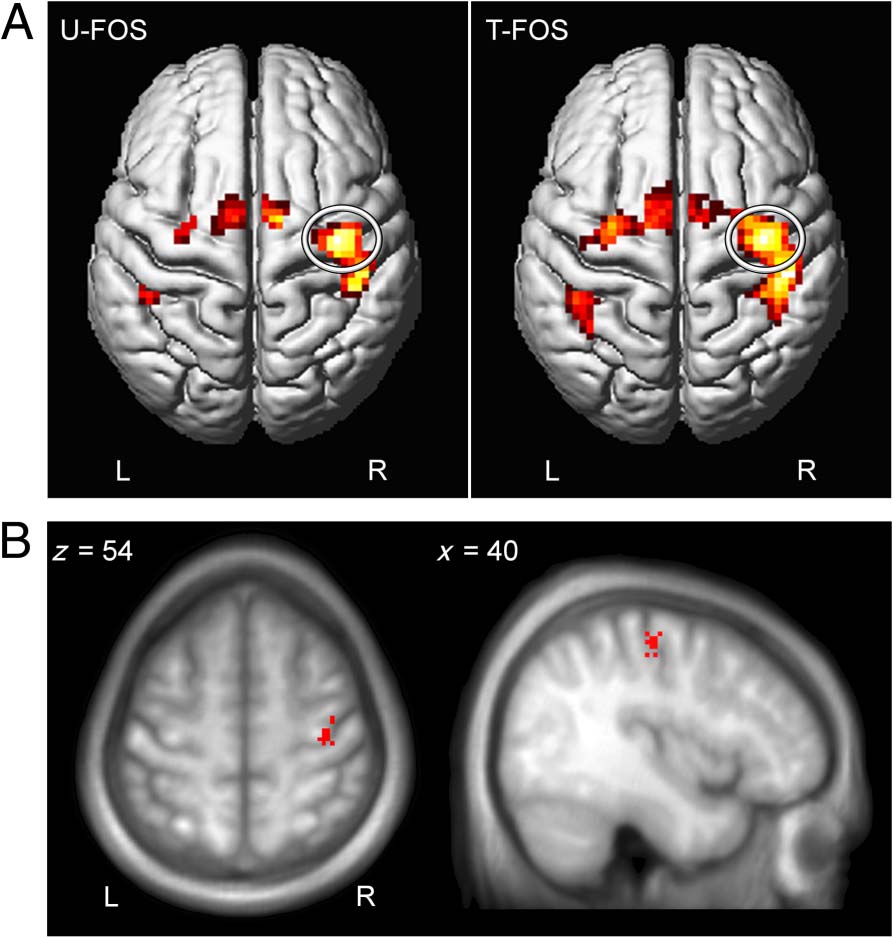
Figura 2. Task-related activity and ROI. (UN) Activation maps of
group effects showing areas activated during performance of the U-FOS
(left) and the T-FOS (right) overlaid on the surface rendered from the
mean structural segmented images of all participants. The maps were
thresholded using FWE correction at p < .05. Ellipse = motor hand
area corresponding to the performing hand. (B) Individual locations of
the most active voxels averaged across sequences within the M1 hand
area were used as a center for sphered ROI with a radius of 6 mm.
the primary motor hand area contralateral to the per-
forming hand, did not differ between the two sequences.
A repeated-measures multivariate ANOVA with Sequence
(U-FOS, T-FOS) as a within-subject factor with three
levels, that is, coordinates (x, y, and z), showed no sig-
nificant effect of Sequence (F(3, 12) = 1.06, p = .40).
Therefore, individual MNI coordinates were averaged
across the two sequences for each participant, and the
ROI was defined as a sphere (r = 6 mm) centered on that
mean location (40 ± 0.41, −19 ± 0.82, 54 ± 0.95, mean ±
SEM for x, y, and z, respectively; Figure 2B).
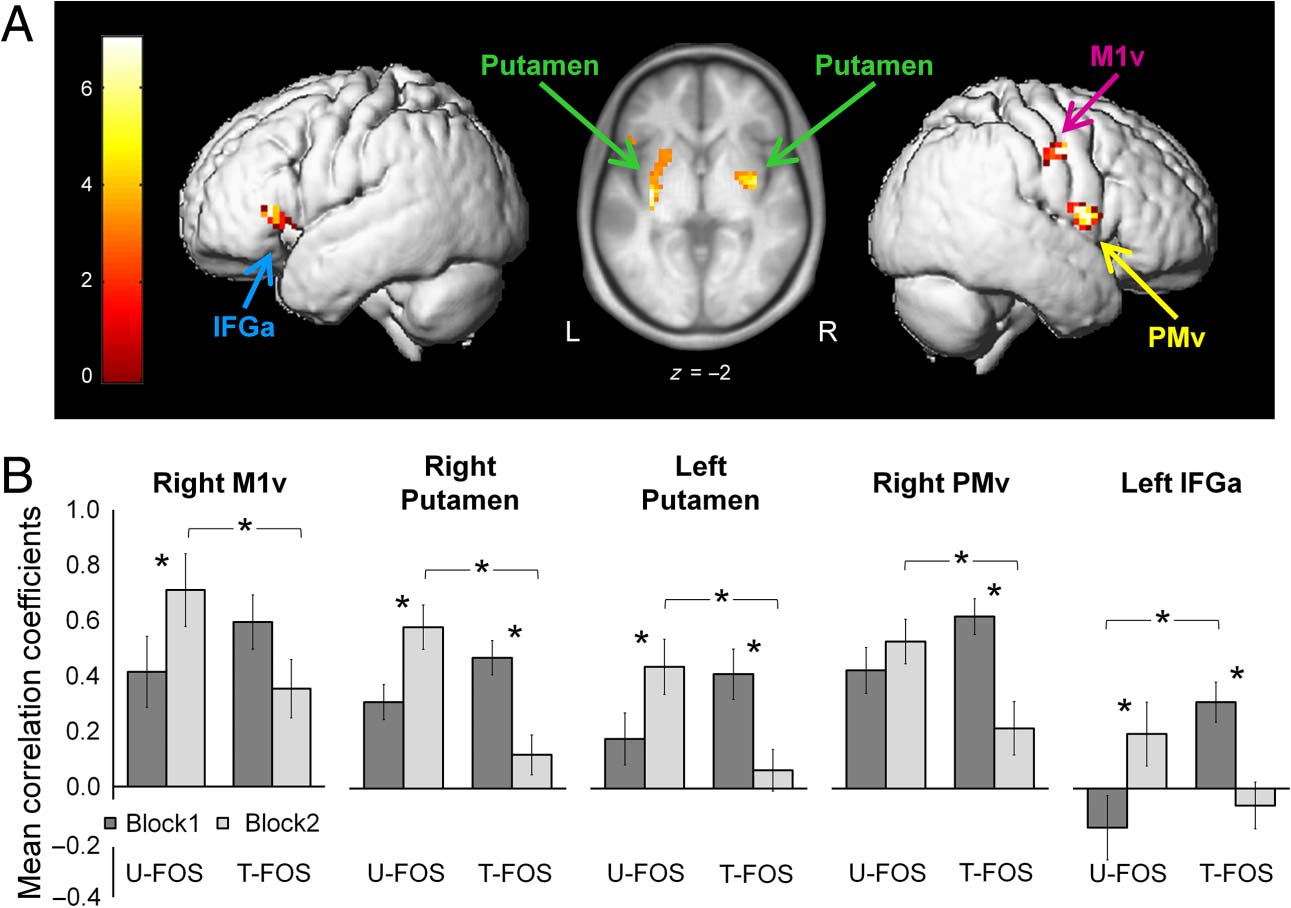
Connectivity Analyses
Areas wherein functional connectivity with the M1 seed
showed significant effects are listed in Table 1. Compari-
son of the connectivity maps with the M1 hand area as
the seed, generated for the performance blocks during
execution of the U-FOS and the T-FOS, showed no signif-
icant main effect of Sequence (U-FOS vs. T-FOS). Analyses
of the main effect of Block revealed a significant decrease
(i.e., Block1 > Block2) in connectivity between the M1
seed and the lateral anterior part of the left inferior frontal
gyrus (Tavolo 1, labeled as Frontal_Inf_Orb (AAL)), a cluster
conjoining Brodmannʼs areas (BA) 45, 46, E 47.
The whole-brain connectivity patterns, with the M1
seed, were differentially modulated by the repeated per-
formance of the two sequences (Figura 3 and Table 1).
Gabitov, Manor, and Karni
741
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
C
7
N
4
_
9
UN
/
_
j
0
o
0
C
7
N
5
1
_
UN
P
_
D
0
0
B
7
sì
5
G
1
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
F
.
T
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
Tavolo 1. Functional Connectivity with the M1 Seed
MNI Coordinates
Label
U-FOS > T-FOS
No significant results
U-FOS < T-FOS No significant results Block1 < Block2 No significant results Block1 > Block2
Frontal_Inf_Orb
X
–
–
–
sì
–
–
–
l
−48
44
[U-FOS: Block1 < Block2] × [T-FOS: Block1 > Block2]
Precentral
Putamen
Putamen
Rolandic_Oper
Frontal_Inf_Tri
R
R
l
R
l
45
36
−33
60
−39
−7
−7
−16
8
26
[U-FOS: Block1 > Block2] × [T-FOS: Block1 < Block2]
No significant results
–
–
z
–
–
–
−8
43
−2
−2
10
7
–
Cluster Size ( Voxels)
z-Score
–
–
–
36
38
52
74
54
31
–
–
–
–
3.64
4.31
4.15
4.52
4.05
3.93
–
p
–
–
–
.03
.02*1
<.01*2
.02FWE
*3
<.01
.03
–
Labeling clusters obtained from connectivity maps thresholded at p < .001 using AAL (Tzourio-Mazoyer et al., 2002). pFWE = cluster-level
FWE-corrected over the entire brain volume; p = cluster-level uncorrected; *1–*3 = significant peak at .05 level FWE-corrected over a small VOI,
1–3 refers to an area of interest used for small volume correction: 1 the right sensorimotor cortex defined as a union between the right primary
sensory and motor cortexes using Human Motor Area Template (Mayka et al., 2006), 2 the right and 3 the left putamen using AAL.
Figure 3. Functional
connectivity analyses using
M1 hand area as a seed.
(A) Areas wherein functional
connectivity patterns were
differentially modulated by
the repeated performance of
the two sequences using M1
as a seed (interaction: [U-FOS:
Block1 < Block2] × [T-FOS:
Block1 > Block2]). Connectivity
map of group effects overlaid
on the mean structural image
of all participants or the
surface rendered from the
participantsʼ mean structural
segmented images. The map
was thresholded at p < .001.
The color bar represents
t values. M1v = primary motor
cortex ventral to the hand
area; PMv = ventral premotor
cortex; IFGa = anterior part
of the inferior frontal gyrus.
(B) Mean correlation coefficients between the M1 seed and each of the clusters, wherein connectivity with the M1 seed showed significant sequence
by block interaction. Columns = mean Fisher-transformed correlation coefficients; bars = SEMs. *Significant differences at 0.05 level corrected for
multiple tests (i.e., a number of clusters) using Bonferroni adjustments.
742
Journal of Cognitive Neuroscience
Volume 27, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Overall, there was a pattern of increased connectivity with
the M1 seed, across blocks (i.e., comparing the functional
connectivity in performance blocks before and after the
brief rest intervals), for the U-FOS but a reduction in con-
nectivity for the T-FOS. Analyses of sequence by block
interactions showed significant changes only in one direc-
tion, toward greater increase in connectivity with the M1
seed across blocks during the U-FOS performance com-
pared with the T-FOS performance ([U-FOS: Block1 <
Block2] × [T-FOS: Block1 > Block2]). This interaction
was significant for clusters within the right M1 ventral to
the hand area and the striatum, bilaterally (Figure 3A and
Tavolo 1, labeled as precentral and putamen (AAL), respec-
tively). The peak voxels within the striatum were located
in the ventral posterior parts of the putamen, bilaterally,
corresponding to the sensorimotor territories of the BG
(Lehéricy et al., 2004, 2005). Post hoc analyses showed that
across blocks during the U-FOS performance the connec-
tivity between the M1 seed and the right M1 ventral to the
hand area increased (T(14) = 4.11, P < .01; Figure 3B).
There was also an increase in the M1 connectivity with
the striatum (t(14) = 3.18, p < .05; t(14) = 2.92, p =
.05, correlation coefficients for the M1 seed with the right
and the left putamen, respectively). However, performance
of the T-FOS led to a relative decrease in the M1 connec-
tivity with the striatum across blocks (t(14) = −5.95, p <
.001; t(14) = −3.51, p < .05, correlation coefficients for
the M1 seed with the right and the left putamen, respec-
tively; Figure 3B). Additional clusters that showed a signif-
icantly greater increase in connectivity with the M1 seed,
across blocks, for the U-FOS compared with the T-FOS
were located within the right rolandic operculum, cor-
responding to the ventral premotor cortex (PMv), and
the anterior part of the left inferior frontal gyrus (IFGa,
BA 45 and 47; Figure 3A and Table 1, labeled as Rolandic_
Oper and Frontal_Inf_Tri (AAL), respectively). Post hoc
analyses showed a relative increase in the right-M1–
left-IFGa connectivity across blocks, during the U-FOS per-
formance (t(14) = 3.41, p < .05), but a relative decrease
in the right-M1–right-PMv as well as the right-M1–left-IFGa
connectivity during the performance of the T-FOS (t(14) =
−4.82, p < .01; t(14) = −4.70, p < .01, correlation co-
efficients for the M1 seed with the right PMv and the left
IFGa, respectively; Figure 3B).
Correlation coefficients with the M1 seed were also
compared between the two sequences (U-FOS, T-FOS)
separately, for each block (Block1, Block2). Connectivity
for the M1 seed with the right M1 area ventral to the hand
knob and the striatum was similar for both sequences dur-
ing the first blocks, but after the brief rest interval during
the second blocks in the pairs connectivity was stronger
for the U-FOS compared with the T-FOS (t(14) = 4.18,
p < .01; t(14) = 5.02, p < .001; t(14) = 3.90, p = .01,
correlation coefficients for the M1 seed with the ventral
part of the right M1, the right and the left putamen,
respectively; Figure 3B). A similar pattern was observed
for the right-M1–right-PMv connectivity values (t(14) =
3.16, p < .05). The right IFGa was the only cluster that
showed stronger connectivity with the M1 seed for the
T-FOS compared with the U-FOS during the first block
(t(14) = −3.47, p < .05; Figure 3B).
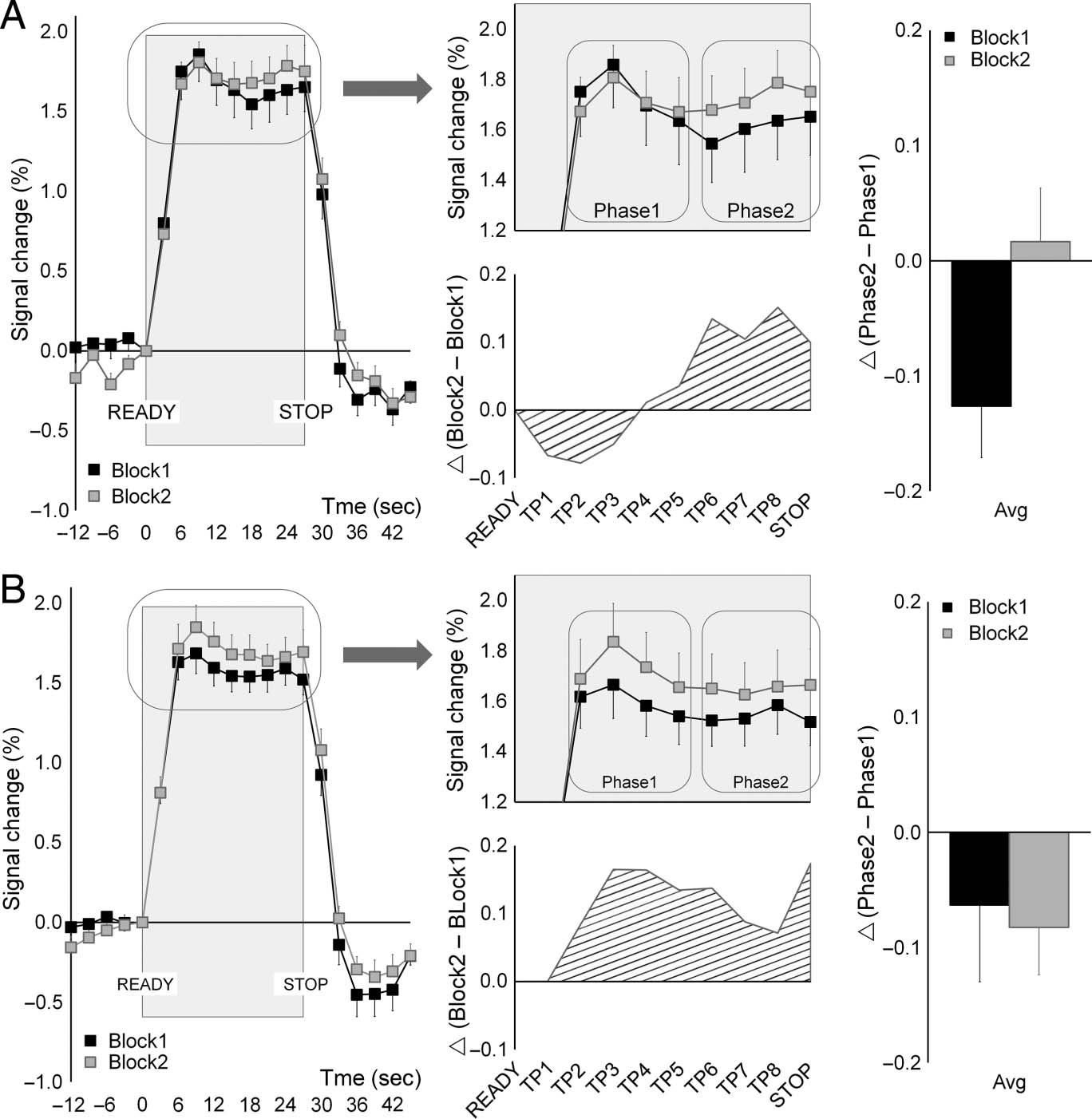
Time-course Analyses
The time-course series, for the two sequences, in terms
of percent signal change extracted from the M1 hand
area, are shown in Figure 4. A repeated-measures ANOVA
with Sequence (U-FOS, T-FOS), Run (1–3), Block (Block1,
Block2), and Phase (Phase1, Phase2) as within-subject fac-
tors showed no significant effect of Run (F(1.86, 26.09) =
0.43, p = .64). There were neither significant differences
in the magnitude of evoked BOLD signals between the
two sequences (F(1, 14) = 2.75, p = .12) nor a differential
modulation of the mean signal across blocks (F(1, 14) =
0.94, p = .35; F(1, 14) = 0.53, p = .48, Block effect and
Sequence × Block interaction, respectively). Thus, the
mean magnitude of the evoked BOLD signal within each
block did not differentiate between the two sequences.
However, there was a significant Sequence × Block ×
Phase interaction (F(1, 14) = 8.62, p = .01) indicating that
activity varied between Phase1 and Phase2 as a function
of the sequence and its repetition across the brief rest
interval.
Post hoc repeated-measures ANOVAs were performed
separately for each sequence (U-FOS, T-FOS). Repetition
effects across blocks (Block2–Block1) averaged across
runs are shown in Figure 4 (bottom middle). On average
there was a trend toward relative enhancement (RE) in the
BOLD signals across blocks when participants executed
the T-FOS; these differences, however, were not signifi-
cant (F(1, 14) = 1.28, p = .28). There was a significant
Phase × Block interaction only for the U-FOS (F(1, 14) =
11.18, p < .01; F(1, 14) = 0.13, p = .73, U-FOS and T-FOS,
respectively). Repeated-measures ANOVAs performed
separately for each phase (Phase1, Phase2) during the
U-FOS performance did not show any significant differ-
ences across blocks (F(1, 14) = 0.22, p = .65; F(1, 14) =
1.11, p = .31, Phase1 and Phase2, respectively). However,
when the phases where compared within blocks, there
was a significant reduction of BOLD signal from Phase1
to Phase2 in the first block, that is, within-block RS, (F(1,
14) = 8.67, p = .01), but no BOLD signal modulation
between the two phases in the second, repeated block
(F(1, 14) = 0.25, p = .63; Figure 4A, right plot). No signifi-
cant changes in the evoked BOLD signals were observed
within blocks during the T-FOS performance (Figure 4B,
right plot). The differential within-block modulations in
the BOLD signal were replicable across runs (Figure 5).
Signal variability within the M1 significantly decreased
during the performance of the both sequences compared
with rest (Figure 6 and Table 2). A repeated-measures
ANOVA on the mean SSD (see Methods) with Sequence
(U-FOS, T-FOS), Run (1–3), and Period of interest (Phase1
and Phase2 within each block as well as three periods of
Gabitov, Manor, and Karni
743
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 4. Time courses of
BOLD signal within the M1
hand area and repetition effects:
(A) U-FOS and (B) T-FOS.
Mean time courses in percent
signal change (vs. performance
onset, “READY” cue) across
all sequence-specific runs
are plotted separately for
each performance block
(Block1, Block2) versus time
(in seconds, counted from a
performance onset, that is,
“READY” cue = 0 sec; left
and top middle plots). Data
points = group mean percent
signal changes at a single
time-point; bars = SEMs.
Repetition effects across blocks
were measured as differences
(▵) between the two blocks
(i.e., Block2 − Block1; bottom
middle plots). Vertices = ▵
at corresponding time points
averaged across runs. Negative
values correspond to RS
effects across blocks; positive
values correspond to RE
effects across blocks. Repetition
effects within blocks were
measured as differences (▵)
between the two phases
(i.e., Phase2 − Phase1; right
plots). Columns = within-block
repetition effects averaged
across runs; bars = SEMs.
rest) as within-subject factors resulted in significant effect
of Period of interest (F(1.91, 26.76) = 6.78, p < .01) irre-
spective of the sequence (F(1, 14) = 0.10, p = .76; F(2.41,
33.80) = 0.42, p = .70, main effect of Sequence and
Sequence × Period-of-interest interaction, respectively)
or run (F(1.82, 25.46) = 0.66, p = .51). Post hoc pairwise
comparisons revealed that variability during all rest periods
was significantly higher than during the actual performance
blocks (Table 2A). A repeated-measures ANOVA with
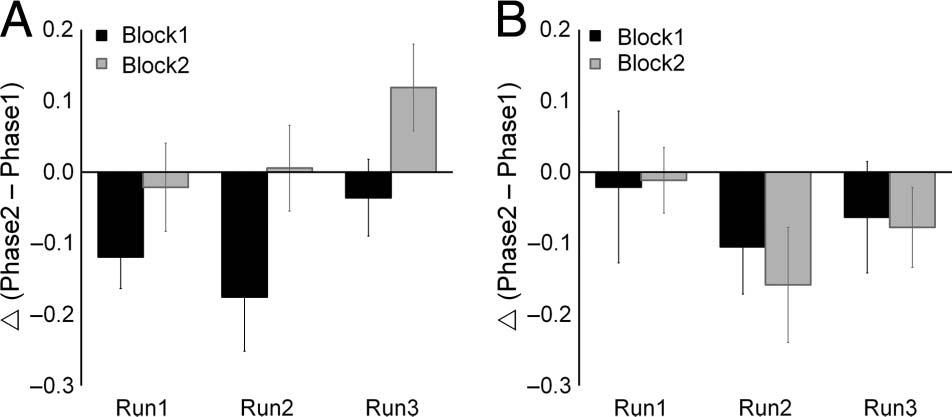
Figure 5. Within-block repetition effects within the M1 hand area
for each run: (A) U-FOS and (B) T-FOS. Repetition effects within
blocks, measured as differences (▵) between the two phases (i.e.,
Phase2 − Phase1), are plotted separately for each block and run.
Columns = within-block repetition effects; bars = SEMs.
Sequence (U-FOS, T-FOS), Run (1–3), Block (Block1,
Block2), and Phase (Phase1, Phase2) as within-subject
factors showed a significant decrease in variability from
Phase1 to Phase2 (F(1, 14) = 22.33, p < .001) irrespective
of a sequence, run, or block.
Analyses of SD for each period of interest showed
similar results (Figure 6B and Table 2B). Thus, while signal
variability within M1 significantly decreased during the
performance of the two sequences compared with rest,
as well as within blocks (i.e., across phases), the signal
variability, within blocks or across blocks, did not reflect
sequence specificity.
Behavioral Results
The results of the behavioral performance tests under-
taken after the imaging session, in comparison with the
nonscanned control group are shown in Figure 7. The
performance of the T-FOS was significantly faster, more
accurate, and less variable compared with the U-FOS in
both groups. A repeated-measures ANOVA with Group
(fMRI, control) as a between-subject factor and Sequence
(Overnight T-FOS and U-FOS) as a within-subject fac-
tor showed a significant effect of Sequence for both,
the number of correct sequences (i.e., speed) and the
744
Journal of Cognitive Neuroscience
Volume 27, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
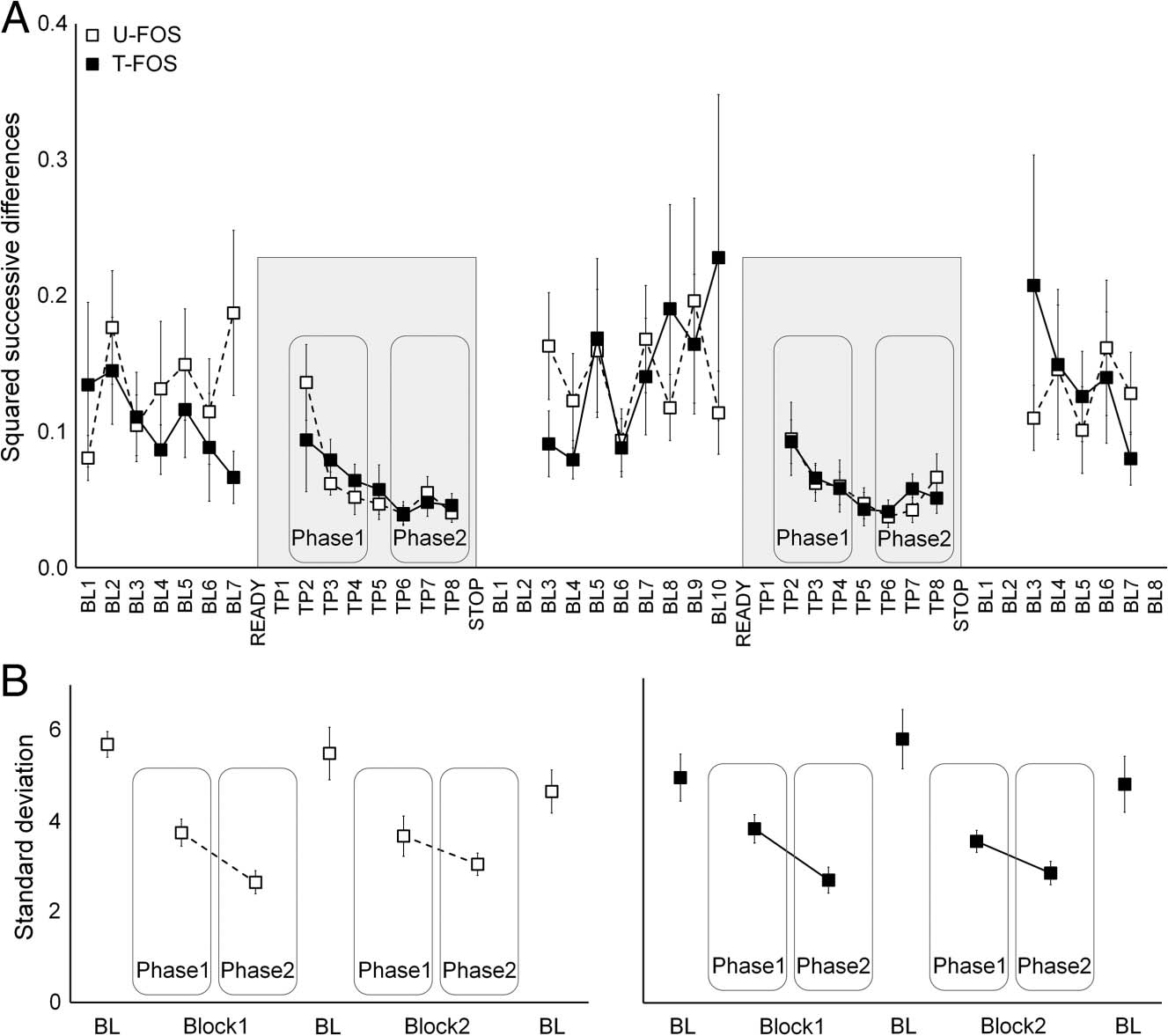
Figure 6. Temporal variability
of BOLD signal within the
M1 hand area. (A) SSDs
averaged across runs are plotted
separately for each sequence
(U-FOS = white squares;
T-FOS = black squares)
versus time points with
distinct indexing for each
period of interest: BL = rest;
TP = performance block.
Data points = group mean
SSD of signal change (%)
between the current and the
next time-point; bars = SEMs.
(B) SD for each period of
interest averaged across runs
is plotted separately for each
sequence (U-FOS = left plot,
T-FOS = right plot). BL = rest;
data points = group mean SD;
bars = SEMs.
number of errors (F(1, 28) = 93.10, p < .001; F(1, 28) =
36.98, p < .001, respectively). The effect of Sequence was
also significant for the within-test rate of improvement in
speed (slope) and the within-test speed variability, that
is, SD (F(1, 28) = 9.09, p < .01; F(1, 28) = 18.14, p <
.001, respectively). There was no significant Group effect
for any of the behavioral measures. However, the analy-
ses of errors and slope showed a significant Sequence ×
Group interaction (F(1, 28) = 36.98, p < .001; F(1, 28) =
5.01, p < .05, respectively). A post hoc t test performed
separately for each sequence showed significant dif-
ferences in performance between the two groups only
Table 2. Statistics of Temporal Variability for BOLD Signal within the M1 Hand Area
A. Mean SSDs
Phase
Rest1 (0.121 ± 0.015)
Rest2 (0.143 ± 0.030)
Rest3 (0.135 ± 0.032)
Phases Difference
Block1 Phase1 (0.081 ± 0.010)
Block1 Phase2 (0.045 ± 0.005)
Block2 Phase1 (0.072 ± 0.008)
0.001
0.001
0.003
Block2 Phase2 (0.050 ± 0.006)
<0.001
0.005
0.005
0.008
<0.001
0.015
0.006
0.013
0.000
0.001
0.019
B. SD
Phase
Rest1 (5.274 ± 0.340)
Rest2 (5.588 ± 0.571)
Rest3 (4.683 ± 0.474)
Phases Difference
Block1 Phase1 (3.749 ± 0.231)
Block1 Phase2 (2.648 ± 0.236)
Block2 Phase1 (3.576 ± 0.306)
Block2 Phase2 (2.923 ± 0.206)
<0.001
<0.001
0.001
<0.001
0.002
<0.001
0.004
<0.001
0.060
0.001
0.056
0.002
<0.001
0.041
Statistics ( p values) for mean differences in variability measures (mean SSD and SD) between periods of interest (Phase1 and Phase2 for each block as well
as three periods of rest) resulted from post hoc pairwise comparisons. Mean values of variability for each period of interest are shown in parentheses
(mean ± SEM ).
Gabitov, Manor, and Karni
745
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
terns of dynamic changes in motor cortex activity and
connectivity during the repeated performance of the
sequences can be considered as neural signatures of
novelty and experience with a motor task.
There were no significant differences between the two
sequences in the location of the most active voxel within
the primary motor hand area contralateral to the perform-
ing hand or in the mean magnitude of activity evoked
during the performance. The current results, therefore,
are in line with the results of animal studies showing that
the representation of motor sequences in M1 may not
be reliably assessed by averaging neuronal activity (Zelenin
et al., 2011; Ben-Shaul et al., 2004; Hatsopoulos, Paninski,
& Donoghue, 2003). There are data suggesting that dif-
ferent tasks can be reliably expressed in the modulation
patterns rather than in the population mean of activity
of motor cortex neurons (Zelenin et al., 2011), presumably
reflecting the fact that the same neuronal pool in the
motor cortex can be recruited in different tasks ( Yang
et al., 2014; Zelenin et al., 2011).
During the performance blocks signal variability in the
contralateral M1 significantly decreased, compared with
rest as well as within blocks (i.e., across phases), for both
sequences (Figure 6). These results are in agreement
with recent reports (He, 2013) of a reduction in variability
in the fMRI BOLD signals following stimulus onset, as
well as with animal studies that showed that intrinsically
generated spontaneous fluctuations in neuronal activity
undergo suppression during task-evoked activity through-
out the cortex in a broad range of conditions (White, Abbott,
& Fiser, 2012; Churchland et al., 2010; Churchland, Yu, Ryu,
Santhanam, & Shenoy, 2006). The decline in variability
implies that cortical circuits can be stabilized by an input
or a task and may support information encoding (He,
2013; White et al., 2012; Churchland et al., 2010). In the
current study, however, the large decreases in the M1 signal
variability with task performance were not differentially
modulated by the level of experience with the specific
movement sequence or by its repetition across the brief
rest interval.
Relative changes in the magnitude of the mean M1
activity across blocks were observed for the T-FOS, with
a trend toward RE (Figure 4B). These RE effects have been
shown to be significant for participants that expressed
delayed “offline” performance gains overnight (i.e., in ad-
dition to the gains achieved immediately after the training;
Gabitov et al., 2014). RE effects in the M1 contralateral to
the performing hand were reported to occur for practiced
sequences (Karni et al., 1995, 1998). However, unlike the
previous results of Karni et al. (1995, 1998), no across-
blocks reductions in the magnitude of the mean M1 ac-
tivity upon repetition (RS) were observed for the U-FOS
in the current study. It has been proposed that RS effects
in the primary motor cortex may reflect an adaptation
phase which relate to repeated component movements
rather than to their specific order in a sequence in early
phases of experience (Karni et al., 1995, 1998). The
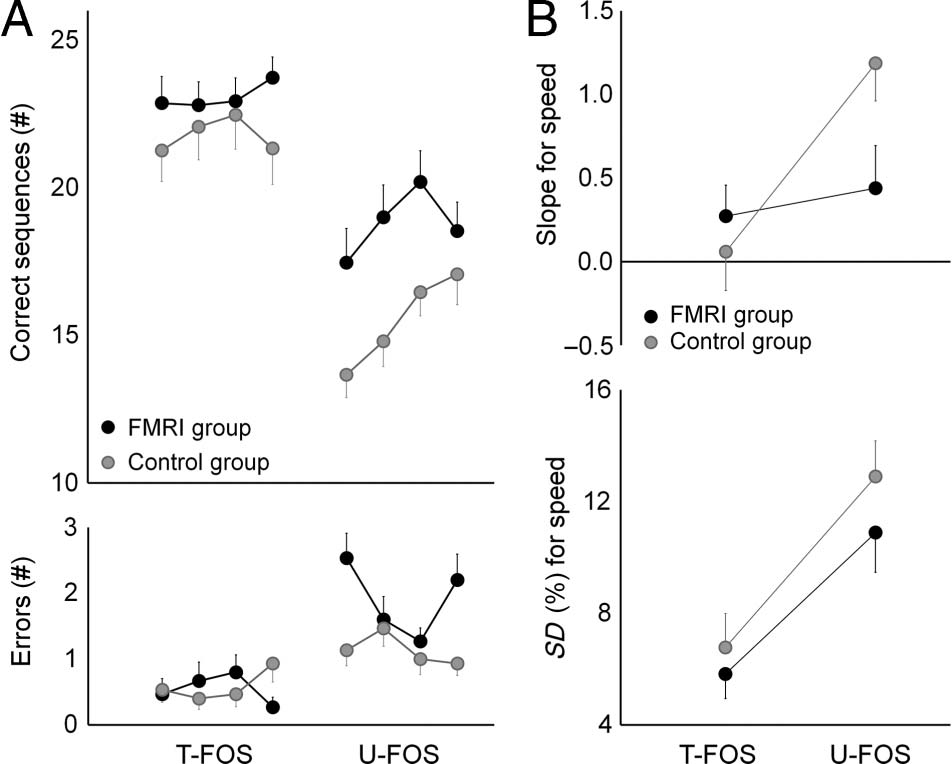
Figure 7. Behavioral results. Performances of fMRI and control
group overnight. (A) The number of correctly completed sequences
(i.e., speed, top) and the number of sequences with ordering errors
(bottom). Data points = group mean values for each of the four test
blocks; bars = SEMs. (B) Within-test change in speed (slope, top)
and within-test speed variability (SD, bottom). Data points = group
mean values; bars = SEMs.
for the U-FOS (Overnight U-FOS). As can be seen in
Figure 7, U-FOS performance was faster in the fMRI
group with a reduced within-test rate of improvement
(T(28) = 2.61, p = .01; T(28) = −2.20, p < .05, speed and
slope, respectively, fMRI vs. control group) indicating that
the experience with the novel sequence inside the scanner
contributed to the subsequent performance. However,
this experience was insufficient to reduce the within-test
variability for speed (SD) during the subsequent U-FOS
performance (T(28) = −1.05, p = .30, fMRI vs. control
group). The number of errors was significantly higher in
the fMRI group (T(28) = 2.64, p = .01, fMRI vs. control
group), although the size of this effect was very small
because of the overall small number of errors committed
(1.9 ± 0.23; 1.13 ± 0.18, mean ± SEM, fMRI and control
group, respectively). This small difference in accuracy
reflected in part the unintended switching to the T-FOS.
DISCUSSION
Our results suggest that short-term BOLD signal modula-
tions, within the performance blocks as well as across
blocks separated by the brief rest interval, reflect the level
of experience with the movement sequence and may,
thus, reliably reflect underlying mnemonic processes.
Both the patterns of neural activity and the functional con-
nectivity of the primary motor hand area (contralateral
to the performing hand) were differentially modulated
by the repeated performance of the untrained and trained
sequences. However, different levels of motor experience
were not reflected in the averaged signal intensities or in
signal variability per se. The transient but consistent pat-
746
Journal of Cognitive Neuroscience
Volume 27, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
absence of coherent RS effects across blocks for the
U-FOS in the current study may, therefore, reflect the
prior experience with the component movements that
underwent stabilization overnight. Nevertheless, there
were significant reductions in the M1 activity within the
initial blocks of the pairs, that is, from the first to the sec-
ond phase (Figures 4A and 5A), during the U-FOS perfor-
mance. Repetition-related reductions in neural activity
were also observed in perceptual systems (for a review,
see Grill-Spector, Henson, & Martin, 2006) and were
suggested to reflect tuning and task optimization pro-
cesses (Schacter & Buckner, 1998; Wiggs & Martin, 1998;
Desimone, 1996). From this point of view, the activity of
cells that poorly represent the stimulus-specific features
is reduced upon repetition, leaving cells carrying critical
information for task performance (Desimone, 1996). We
propose that the reduction of the M1 activity within the
initial blocks reflects the tuning (i.e., adaptation) of the
M1 representation for the novel sequence of movements.
This within-block adaptation-like effect saturated by the
second phase of the first block and did not recur across
the brief rest interval, that is, BOLD signal levels, tended
to stabilize during the second, repeated block. These
modulations, during the performance of the U-FOS but
not the T-FOS, presumably reflect the short-term accu-
mulation of experience with the novel order (sequence,
syntax) of movements as both sequences were com-
posed of the same component opposition movements.
It has been shown that the initial phases of motor task
acquisition are characterized by various activity patterns
of movement-related neurons selected and engaged
from a more extensive pool in motor cortex; these activ-
ity patterns stabilize and a more restricted population is
consistently engaged after extensive training (Peters
et al., 2014). The transient stabilization of neural activity
upon repeated performance blocks, in the current study,
may result from the selection of a particular subset of
excitatory neurons that were “tried out” during the cor-
responding initial blocks.
Our results suggest that the initial acquisition (encoding)
of procedural knowledge about a novel order (sequencing)
of component movements is characterized not only by
short-term stabilizations of activity but also by an increase
in functional connectivity between voxels within M1. An
increase in functional connectivity within the primary
motor cortex was previously observed during learning
new muscle synergies (McNamara et al., 2007) and, re-
cently, following noninvasive cortical stimulation using
transcranial direct current stimulation (Sehm, Kipping,
Schäfer, Villringer, & Ragert, 2013; Polanía, Paulus, &
Nitsche, 2012). The increase of coherence and the tran-
sient signal stabilization within the M1 may be the neural
signature of working memory processes (Fuster, 2001)
whereby relevant information about the sequence of
movements is maintained “in mind” for brief periods of
time (Langner et al., 2013). There is evidence that M1 is
capable of storing procedural information in STM (Classen,
Liepert, Wise, Hallett, & Cohen, 1998). The capacity of the
primary motor cortex to undergo short-term (transient)
plastic modifications by practiced movements has been
proposed to constitute the first step in skill acquisition
and may be crucial for the long-term formation of a new
motor skill representation (Classen et al., 1998; Karni
et al., 1995). Differential repetition-driven short-term
plasticity in M1 may be related to short-term modula-
tions of GABA concentration and excitatory–inhibitory
balance changes in relation to practice (Stagg, Bachtiar, &
Johansen-Berg, 2011; Floyer-Lea, Wylezinska, Kincses, &
Matthews, 2006; Bütefisch, Khurana, Kopylev, & Cohen,
2004). It has been suggested that rapid, regionally specific
short-term decreases in GABA concentration in M1 may
be associated primarily with encoding of the task during
the period of task performance, rather than its longer-
term consolidation (Floyer-Lea et al., 2006).
The fMRI-BOLD signal correlations between the right M1
hand area and the posterior ventral striatum, corresponding
to the sensorimotor territories of the BG (Lehéricy et al.,
2004, 2005), showed a differential pattern for the two
sequences. For the U-FOS, there were relative increases
in the right-M1–striatum connectivity after the brief rest
interval. Repetition of the T-FOS resulted in relative de-
creases in the right-M1–striatum connectivity across blocks.
The differential modulations of the M1–striatum connec-
tivity upon repeated performance of the U-FOS compared
with the T-FOS are in line with the notion that cortical and
striatal circuits exhibit remarkable but dissociable plasticity
as a function of the level of prior experience with a given
task (Costa, Cohen, & Nicolelis, 2004). It has been pro-
posed that high-level associative circuits with frontoparietal
regions and associative regions of the striatum and the
cerebellum are recruited during the early phase of motor
learning, whereas sensorimotor regions may take over
during later learning phases (Doyon et al., 2009; Doyon &
Benali, 2005; Hikosaka, Nakamura, Sakai, & Nakahara,
2002). However, a dynamic shift of activation from the asso-
ciative to the sensorimotor territories of the striatopallidal
complex may occur early during training (Lehéricy et al.,
2005). Lesions of the sensorimotor striatum in mice lead
to significant and selective deficit in the acquisition of serial
order in lever pressing (Yin, 2010).
Additional, albeit less robust, sequence-specific changes
were found in functional connectivity of the right M1
hand area with the IFGa (BA 45 and 47). This region cor-
responds to Brocaʼs area (Broca, 1861) and was implicated
in various motor functions such as planning, recognition
and imitation of actions (for reviews, see Fadiga, Craighero,
& DʼAusilio, 2009; Binkofski & Buccino, 2006). The relative
increase in functional connectivity between the right M1
hand area and the left IFGa during the U-FOS performance
may reflect a recruitment of syntactic processing routines
within the dominant hemisphere (Roy et al., 2013; Fazio
et al., 2009) in early stages of sequence practice. On the
other hand, the relative reduction in the coupling of the
right M1 with the IFGa and the PMv across performance
Gabitov, Manor, and Karni
747
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
blocks during the performance of the T-FOS may reflect a
reduction in allocation of cognitive resources presumably
involved in task aspects such as explicit sequence control
and attention (Barber, Caffo, Pekar, & Mostofsky, 2013;
Hikosaka et al., 2002; Honda et al., 1998).
The most common application of functional connec-
tivity is examining intrinsic correlations determined dur-
ing task-free intervals (rest). It has been proposed that
resting-state functional connectivity approaches can not
only reveal the underlying anatomical connectivity but also
contribute to our understanding of brain dynamics (Deco,
Jirsa, & McIntosh, 2011; Raichle, 2010). Although func-
tional connectivity during rest may be significantly altered
by motor learning (Vahdat et al., 2011; Albert, Robertson,
& Miall, 2009), it may also be modified by rote button
presses (Tung et al., 2013). Significant changes in func-
tional connectivity patterns were observed for resting-
state data, across imaging sessions (Honey et al., 2009),
between runs within the same imaging session (Shehzad
et al., 2009) as well as within the same run on a timescale
of seconds to minutes (Chang & Glover, 2011) with no
specific learning experience afforded. Thus, the assess-
ment of functional connectivity during performance inter-
vals, rather than during rest, may more directly reflect
task-specific mnemonic processes.
It is not known whether and how the length and nature
of the rest interval inserted between the two performance
blocks affect the modulation of neural activity and con-
nectivity upon task repetition. The number and rate of
task iterations (the block length) may also be important
factors in the modulation of BOLD signal to task repeti-
tion; however, the effects of changing these time param-
eters remain to be determined. Adaptation studies in
the visual system showed that the magnitude of repetition
effects was decreased with longer ISIs (Henson, Shallice, &
Dolan, 2000; Grill-Spector et al., 1999) and increased with
longer exposures to the stimuli (Grill-Spector et al., 1999).
Robust repetition effects were observed in M1 and can be
reproduced (recovered) in successive runs when these
are separated by breaks (1.5–2 min) dedicated to verbal
interaction with participants (Gabitov et al., 2014; Karni
et al., 1995, 1998). One cannot rule out, however, that
the verbal interaction with the participants during the
breaks between runs may have an effect on the recovery
of the repetition effects.
In the current study, the performance of the untrained
sequence was significantly slower, less accurate, and more
variable than the performance of the trained sequence,
composed of identical opposition movements (Figure 7).
However, during the scanning session, the differences in
the rate of sequence execution and accuracy were mini-
mized, because participants performed the component
movements of both sequences at a comfortable, exter-
nally paced rate. Thus, the differences in patterns of
modulation in neural activity and connectivity were not
directly related to task execution speed or the number
and nature of the component movements, but more likely
reflected the differences in prior experience with the two
sequences.
The behavioral data acquired immediately after the
fMRI session suggest that the imaging session constituted
a learning experience; experience with the U-FOS inside
the scanner contributed to its faster performance and
led to saturation of within-test improvements compared
with the control group (Figure 7). The less accurate per-
formance of the U-FOS in the fMRI group compared with
the controls may be a consequence of proactive inter-
ference from the previously learned T-FOS (Cothros
et al., 2006) or task-switching costs (Kiesel et al., 2010).
The additional experience afforded during the fMRI ses-
sion with the T-FOS had no significant impact on its
subsequent performance.
Conclusions
Altogether, the current results support the idea that tem-
poral signal modulations can reflect nonrandom consis-
tent differential patterns of brain activity evolving as
a function of the statistics of accumulated experience.
Brain function may be underappreciated when using
mean-based brain measures (He, 2013; Garrett et al.,
2010, 2011; Stein et al., 2005), but measures of signal var-
iability per se may not reflect critical aspects of experience-
driven changes in brain activity. We propose that a network
including M1 and striatum underlies online motor work-
ing memory whereby motor representations of specific
movement sequences are retained across short periods of
time (Langner et al., 2013). This network may promote
a transient integrated representation of a new movement
sequence and readily retrieves a previously established
movement sequence representation.
Acknowledgments
E. G. was partially supported by a fellowship from the E. J.
Safra Brain Research Center for the Study of Learning Disabil-
ities. The research leading to these results has received fund-
ing from the European Union Seventh Framework Program
(FP7/2007-2013) under grant agreement no. 604102 (Human
Brain Project). We thank Tammi Kushnir, Ph.D., director of
research at the C. Sheba Medical Center, for her technical and
administrative support.
Reprint requests should be sent to Ella Gabitov, University of
Haifa, 199 Abba Hushi Blvd., Mount Carmel, Haifa 3498838,
Israel, or via e-mail: gabitovella@gmail.com.
REFERENCES
Albert, N. B., Robertson, E. M., & Miall, R. C. (2009). The
resting human brain and motor learning. Current Biology,
19, 1023–1027.
Andersson, J. L. R., Hutton, C., Ashburner, J., Turner, R., &
Friston, K. J. (2001). Modeling geometric deformations
in EPI time series. Neuroimage, 13, 903–919.
Barber, A. D., Caffo, B. S., Pekar, J. J., & Mostofsky, S. H.
(2013). Effects of working memory demand on neural
748
Journal of Cognitive Neuroscience
Volume 27, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
mechanisms of motor response selection and control.
Journal of Cognitive Neuroscience, 25, 1235–1248.
Ben-Shaul, Y., Drori, R., Asher, I., Stark, E., Nadasdy, Z., &
Abeles, M. (2004). Neuronal activity in motor cortical areas
reflects the sequential context of movement. Journal of
Neurophysiology, 91, 1748–1762.
Bestmann, S., Swayne, O., Blankenburg, F., Ruff, C. C.,
Haggard, P., Weiskopf, N., et al. (2008). Dorsal premotor
cortex exerts state-dependent causal influences on activity
in contralateral primary motor and dorsal premotor cortex.
Cerebral Cortex, 18, 1281–1291.
Binkofski, F., & Buccino, G. (2006). The role of ventral
premotor cortex in action execution and action
understanding. Journal of Physiology Paris, 99, 396–405.
Brett, M., Anton, J.-L., Valabregue, R., & Poline, J.-B. (2002).
Region of interest analysis using an SPM toolbox [abstract].
Presented at the 8th International Conference on Functional
Mapping of the Human Brain, June 2–6, 2002, Sendai,
Japan. Available on CD-ROM in Neuroimage, Vol 16.
Broca, P. P. (1861). Remarques sur le siège de la faculté du
langage articulé; suivies dʼune observation dʼaphémie,
1861. Bulletin de la Société Anatomique (Paris), 2,
330–357.
Bütefisch, C. M., Khurana, V., Kopylev, L., & Cohen, L. G.
(2004). Enhancing encoding of a motor memory in the
primary motor cortex by cortical stimulation. Journal of
Neurophysiology, 91, 2110–2116.
Carpenter, A. F., Georgopoulos, A. P., & Pellizzer, G. (1999).
Motor cortical encoding of serial order in a context-recall
task. Science, 283, 1752–1757.
Chang, C., & Glover, G. H. (2011). Time-frequency dynamics
of resting-state brain connectivity measured with fMRI.
Neuroimage, 50, 81–98.
Chouinard, P. A., & Goodale, M. A. (2009). fMRI adaptation
during performance of learned arbitrary visuomotor
conditional associations. Neuroimage, 48, 696–706.
Churchland, M. M., Yu, B. M., Cunningham, J. P., Sugrue,
L. P., Cohen, M. R., Corrado, G. S., et al. (2010). Stimulus
onset quenches neural variability: A widespread cortical
phenomenon. Nature Neuroscience, 13, 369–378.
Churchland, M. M., Yu, B. M., Ryu, S. I., Santhanam, G., &
Shenoy, K. V. (2006). Neural variability in premotor cortex
provides a signature of motor preparation. Journal of
Neuroscience, 26, 3697–3712.
Desimone, R. (1996). Neural mechanisms for visual memory
and their role in attention. Proceedings of the National
Academy of Sciences, U.S.A., 93, 13494–13499.
Dinstein, I., Hasson, U., Rubin, N., & Heeger, D. J. (2007).
Brain areas selective for both observed and executed
movements. Journal of Neurophysiology, 98, 1415–1427.
Doyon, J., Bellec, P., Amsel, R., Penhune, V. B., Monchi, O.,
Carrier, J., et al. (2009). Contributions of the basal ganglia
and functionally related brain structures to motor learning.
Behavioural Brain Research, 199, 61–75.
Doyon, J., & Benali, H. (2005). Reorganization and plasticity
in the adult brain during learning of motor skills. Current
Opinion in Neurobiology, 15, 161–167.
Fadiga, L., Craighero, L., & DʼAusilio, A. (2009). Brocaʼs area
in language, action, and music. Annals of the New York
Academy of Sciences, 1169, 448–458.
Fazio, P., Cantagallo, A., Craighero, L., DʼAusilio, A., Roy,
A. C., Pozzo, T., et al. (2009). Encoding of human action
in Brocaʼs area. Brain, 132, 1980–1988.
Fedorenko, E., Hsieh, P.-J., Nieto-Castañón, A., Whitfield-
Gabrieli, S., & Kanwisher, N. (2010). New method for fMRI
investigations of language: Defining ROIs functionally in
individual subjects. Journal of Neurophysiology, 104,
1177–1194.
Floyer-Lea, A., & Matthews, P. M. (2005). Distinguishable
brain activation networks for short- and long-term motor
skill learning. Journal of Neurophysiology, 94, 512–518.
Floyer-Lea, A., Wylezinska, M., Kincses, T., & Matthews, P. M.
(2006). Rapid modulation of GABA concentration in human
sensorimotor cortex during motor learning. Journal of
Neurophysiology, 95, 1639–1644.
Friston, K. J., Holmes, A. P., Worsley, K. J., Poline, J.-B., Frith,
C. D., & Frackowiak, R. S. J. (1995). Statistical parametric
maps in functional imaging: A general linear approach.
Human Brain Mapping, 2, 189–210.
Fuster, J. M. (2001). The prefrontal cortex—An update: Time
is of the essence. Neuron, 30, 319–333.
Gabitov, E., Manor, D., & Karni, A. (2014). Done that: Short-term
repetition related modulations of motor cortex activity as a
stable signature for overnight motor memory consolidation.
Journal of Cognitive Neuroscience, 26, 2716–2734.
Garrett, D. D., Kovacevic, N., McIntosh, A. R., & Grady, C. L.
(2010). Blood oxygen level-dependent signal variability is more
than just noise. Journal of Neuroscience, 30, 4914–4921.
Classen, J., Liepert, J., Wise, S. P., Hallett, M., & Cohen,
Garrett, D. D., Kovacevic, N., McIntosh, A. R., & Grady,
L. G. (1998). Rapid plasticity of human cortical movement
representation induced by practice. Journal of
Neurophysiology, 79, 1117–1123.
Costa, R. M., Cohen, D., & Nicolelis, M. A. L. (2004). Differential
corticostriatal plasticity during fast and slow motor skill
learning in mice. Current Biology, 14, 1124–1134.
Cothros, N., Köhler, S., Dickie, E. W., Mirsattari, S. M., &
Gribble, P. L. (2006). Proactive interference as a result
of persisting neural representations of previously learned
motor skills in primary motor cortex. Journal of Cognitive
Neuroscience, 18, 2167–2176.
Davare, M., Lemon, R. N., & Olivier, E. (2008). Selective
modulation of interactions between ventral premotor
cortex and primary motor cortex during precision grasping
in humans. Journal of Physiology, 586, 2735–2742.
Davare, M., Montague, K., Olivier, E., Rothwell, J. C., &
Lemon, R. N. (2009). Ventral premotor to primary motor
cortical interactions during object-driven grasp in humans.
Cortex, 45, 1050–1057.
Deco, G., Jirsa, V. K., & McIntosh, A. R. (2011). Emerging
concepts for the dynamical organization of resting-state
activity in the brain. Nature Reviews Neuroscience, 12,
43–56.
C. L. (2011). The importance of being variable. Journal
of Neuroscience, 31, 4496–4503.
Georgopoulos, A. P., Kalaska, J. F., Caminiti, R., & Massey,
J. T. (1982). On the relations between the direction of
two-dimensional arm movements and cell discharge in
primate motor cortex. Journal of Neuroscience, 2,
1527–1537.
Grafton, S. T., & Hamilton, A. F. (2007). Evidence for a
distributed hierarchy of action representation in the
brain. Human Movement Science, 26, 590–616.
Grill-Spector, K., Henson, R. N., & Martin, A. (2006). Repetition
and the brain: Neural models of stimulus-specific effects.
Trends in Cognitive Sciences, 10, 14–23.
Grill-Spector, K., Kushnir, T., Edelman, S., Avidan, G.,
Itzchak, Y., & Malach, R. (1999). Differential processing
of objects under various viewing conditions in the human
lateral occipital complex. Neuron, 24, 187–203.
Halsband, U., & Lange, R. K. (2006). Motor learning in man:
A review of functional and clinical studies. Journal of
Physiology-Paris, 99, 414–424.
Hamilton, A. F., & Grafton, S. T. (2009). Repetition suppression
for performed hand gestures revealed by fMRI. Human
Brain Mapping, 30, 2898–2906.
Gabitov, Manor, and Karni
749
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
t
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Hardwick, R. M., Rottschy, C., Miall, R. C., & Eickhoff, S. B.
(2013). A quantitative meta-analysis and review of motor
learning in the human brain. Neuroimage, 67, 283–297.
Hatsopoulos, N. G., Paninski, L., & Donoghue, J. P. (2003).
Sequential movement representations based on correlated
neuronal activity. Experimental Brain Research, 149,
478–486.
He, B. J. (2013). Spontaneous and task-evoked brain activity
negatively interact. Journal of Neuroscience, 33, 4672–4682.
Henson, R. N., Shallice, T., & Dolan, R. (2000). Neuroimaging
evidence for dissociable forms of repetition priming.
Science, 287, 1269–1272.
Hikosaka, O., Nakamura, K., Sakai, K., & Nakahara, H.
(2002). Central mechanisms of motor skill learning.
Current Opinion in Neurobiology, 12, 217–222.
Honda, M., Deiber, M.-P., Ibáñez, V., Pascual-Leone, A., Zhuang,
P., & Hallett, M. (1998). Dynamic cortical involvement
in implicit and explicit motor sequence learning. A PET
study. Brain, 121, 2159–2173.
Honey, C. J., Sporns, O., Cammoun, L., Gigandet, X.,
Thiran, J.-P., Meuli, R. A., et al. (2009). Predicting human
resting-state functional connectivity from structural
connectivity. Proceedings of the National Academy of
Sciences, U.S.A., 106, 2035–2040.
Hutton, C., Andersson, J., Deichmann, R., & Weiskopf, N.
(2013). Phase informed model for motion and susceptibility.
Human Brain Mapping, 34, 3086–3100.
Johnstone, T., Ores Walsh, K. S., Greischar, L. L., Alexander, A. L.,
Fox, A. S., Davidson, R. J., et al. (2006). Motion correction
and the use of motion covariates in multiple-subject fMRI
analysis. Human Brain Mapping, 27, 779–788.
Kantak, S. S., Mummidisetty, C. K., & Stinear, J. W. (2012).
Primary motor and premotor cortex in implicit sequence
learning-evidence for competition between implicit and
explicit human motor memory systems. European Journal
of Neuroscience, 36, 2710–2715.
Karni, A., Meyer, G., Jezzard, P., Adams, M. M., Turner, R.,
& Ungerleider, L. G. (1995). Functional MRI evidence for
adult motor cortex plasticity during motor skill learning.
Nature, 377, 155–158.
Karni, A., Meyer, G., Rey-Hipolito, C., Jezzard, P., Adams, M. M.,
Turner, R., et al. (1998). The acquisition of skilled motor
performance: Fast and slow experience-driven changes
in primary motor cortex. Proceedings of the National
Academy of Sciences, U.S.A., 95, 861–868.
Kiesel, A., Steinhauser, M., Wendt, M., Falkenstein, M., Jost, K.,
Philipp, A. M., et al. (2010). Control and interference in task
switching—A review. Psychological Bull, 136, 849–874.
Kim, Y.-H., Park, J.-W., Ko, M.-H., Jang, S. H., & Lee, P. K. W.
(2004). Facilitative effect of high frequency subthreshold
repetitive transcranial magnetic stimulation on complex
sequential motor learning in humans. Neuroscience Letters,
367, 181–185.
Kleim, J. A., Hogg, T. M., VandenBerg, P. M., Cooper, N. R.,
Bruneau, R., & Remple, M. (2004). Cortical synaptogenesis
and motor map reorganization occur during late, but not
early, phase of motor skill learning. Journal of Neuroscience,
24, 628–633.
Korman, M., Doyon, J., Doljansky, J., Carrier, J., Dagan, Y., &
Karni, A. (2007). Daytime sleep condenses the time course
of motor memory consolidation. Nature Neuroscience,
10, 1206–1213.
Korman, M., Raz, N., Flash, T., & Karni, A. (2003). Multiple
shifts in the representation of a motor sequence during
the acquisition of skilled performance. Proceedings of the
National Academy of Sciences, U.S.A., 100, 12492–12497.
Langner, R., Sternkopf, M. A., Kellermann, T. S., Grefkes, C.,
Kurth, F., Schneider, F., et al. (2013). Translating working
memory into action: Behavioral and neural evidence for
using motor representations in encoding visuo-spatial
sequences. Human Brain Mapping, 35, 3465–3484.
Lehéricy, S., Benali, H., Van de Moortele, P.-F., Pélégrini-Issac,
M., Waechter, T., Ugurbil, K., et al. (2005). Distinct basal
ganglia territories are engaged in early and advanced motor
sequence learning. Proceedings of the National Academy
of Sciences, U.S.A., 102, 12566–12571.
Lehéricy, S., Ducros, M., Krainik, A., Francois, C., Van de Moortele,
P.-F., Ugurbil, K., et al. (2004). 3-D diffusion tensor axonal
tracking shows distinct SMA and pre-SMA projections to the
human striatum. Cerebral Cortex, 14, 1302–1309.
Matsuzaka, Y., Picard, N., & Strick, P. L. (2007). Skill
representation in the primary motor cortex after long-term
practice. Journal of Neurophysiology, 97, 1819–1832.
Mayka, M. A., Corcos, D. M., Leurgans, S. E., & Vaillancourt,
D. E. (2006). Three-dimensional locations and boundaries
of motor and premotor cortices as defined by functional
brain imaging: A meta-analysis. Neuroimage, 31,
1453–1474.
Mazaika, P. K., Hoeft, F., Glover, G. H., & Reiss, A. L. (2009).
Methods and software for fMRI analysis of clinical subjects.
Annual Meeting of the Organization for Human Brain
Mapping. doi:10.1016/S1053-8119(09)70238-1.
McNamara, A., Tegenthoff, M., Dinse, H., Büchel, C., Binkofski,
F., & Ragert, P. (2007). Increased functional connectivity
is crucial for learning novel muscle synergies. Neuroimage,
35, 1211–1218.
Mohr, P. N. C., & Nagel, I. E. (2010). Variability in brain
activity as an individual difference measure in neuroscience?
Journal of Neuroscience, 30, 7755–7757.
Nieto-Castañón, A., & Fedorenko, E. (2012). Subject-specific
functional localizers increase sensitivity and functional
resolution of multi-subject analyses. Neuroimage, 63,
1646–1669.
Nitsche, M. A., Schauenburg, A., Lang, N., Liebetanz, D.,
Exner, C., Paulus, W., et al. (2003). Facilitation of implicit
motor learning by weak transcranial direct current
stimulation of the primary motor cortex in the human.
Journal of Cognitive Neuroscience, 15, 619–626.
Nudo, R. J., Jenkins, W. M., Prejean, T., & Grenda, R. (1992).
Neurophysiological correlates of hand preference in
primary motor cortex of adult squirrel monkeys. Journal
of Neuroscience, 12, 2918–2947.
Nudo, R. J., Milliken, G. W., Jenkins, W. M., & Merzenich,
M. M. (1996). Use-dependent alterations of movement
representations in primary motor cortex of adult squirrel
monkeys. Journal of Neuroscience, 16, 785–807.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsychologia,
9, 97–113.
Orban, P., Peigneux, P., Lungu, O., Debas, K., Barakat, M.,
Bellec, P., et al. (2011). Functional neuroanatomy
associated with the expression of distinct movement
kinematics in motor sequence learning. Neuroscience,
179, 94–103.
Penhune, V. B., & Doyon, J. (2002). Dynamic cortical and
subcortical networks in learning and delayed recall of
timed motor sequences. Journal of Neuroscience, 22,
1397–1406.
Penhune, V. B., & Doyon, J. (2005). Cerebellum and M1
interaction during early learning of timed motor sequences.
Neuroimage, 26, 801–812.
Peters, A. J., Chen, S. X., & Komiyama, T. (2014). Emergence of
reproducible spatiotemporal activity during motor learning.
Nature, 510, 263–267.
Polanía, R., Paulus, W., & Nitsche, M. A. (2012). Reorganizing
the intrinsic functional architecture of the human primary
750
Journal of Cognitive Neuroscience
Volume 27, Number 4
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
motor cortex during rest with non-invasive cortical stimulation.
PloS One, 7, e30971.
Poldrack, R. A. (2000). Imaging brain plasticity: Conceptual and
methodological issues—A theoretical review. Neuroimage,
12, 1–13.
Raichle, M. E. (2010). Two views of brain function. Trends
in Cognitive Sciences, 14, 180–190.
Rao, S. M., Bandettini, P. A., Binder, J. R., Bobholz, J. A., Hammeke,
T. A., Stein, E. A., et al. (1996). Relationship between finger
movement rate and functional magnetic resonance signal
change in human primary motor cortex. Journal of Cerebral
Blood Flow & Metabolism, 16, 1250–1254.
Tung, K.-C., Uh, J., Mao, D., Xu, F., Xiao, G., & Lu, H. (2013).
Alterations in resting functional connectivity because of
recent motor task. Neuroimage, 78, 316–324.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D.,
Crivello, F., Etard, O., Delcroix, N., et al. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Neuroimage, 15, 273–289.
Vahdat, S., Darainy, M., Milner, T. E., & Ostry, D. J. (2011).
Functionally specific changes in resting-state sensorimotor
networks after motor learning. Journal of Neuroscience,
31, 16907–16915.
Roy, A. C., Curie, A., Nazir, T., Paulignan, Y., des Portes, V.,
Vines, B. W., Nair, D. G., & Schlaug, G. (2006). Contralateral
Fourneret, P., et al. (2013). Syntax at hand: Common syntactic
structures for actions and language. PloS One, 8, e72677.
Saucedo Marquez, C. M., Zhang, X., Swinnen, S. P., Meesen, R.,
& Wenderoth, N. (2013). Task-specific effect of transcranial
direct current stimulation on motor learning. Frontiers in
Human Neuroscience, 7. doi:10.3389/fnhum.2013.00333.
Schacter, D. L., & Buckner, R. L. (1998). Priming and the
brain review. Neuron, 20, 185–195.
Schlaug, G., Knorr, U., & Seitz, R. J. (1994). Inter-subject
variability of cerebral activations in acquiring a motor skill:
A study with positron emission tomography. Experimental
Brain Research, 98, 523–534.
Sehm, B., Kipping, J., Schäfer, A., Villringer, A., & Ragert, P.
(2013). A comparison between uni- and bilateral tDCS
effects on functional connectivity of the human motor
cortex. Frontiers in Human Neuroscience, 7.
doi:10.3389/fnhum.2013.00183.
Shehzad, Z., Kelly, A. M. C., Reiss, P. T., Gee, D. G., Gotimer, K.,
Uddin, L. Q., et al. (2009). The resting brain: Unconstrained
yet reliable. Cerebral Cortex, 19, 2209–2229.
Stagg, C. J., Bachtiar, V., & Johansen-Berg, H. (2011). The role of
GABA in human motor learning. Current Biology, 21, 480–484.
Stagg, C. J., Jayaram, G., Pastor, D., Kincses, Z. T., Matthews,
P. M., & Johansen-Berg, H. (2011). Polarity and timing-
dependent effects of transcranial direct current stimulation
in explicit motor learning. Neuropsychologia, 49, 800–804.
Steele, C. J., & Penhune, V. B. (2010). Specific increases within
global decreases: A functional magnetic resonance imaging
investigation of five days of motor sequence learning.
Journal of Neuroscience, 30, 8332–8341.
Stein, R. B., Gossen, E. R., & Jones, K. E. (2005). Neuronal
variability: Noise or part of the signal? Nature Reviews
Neuroscience, 6, 389–397.
Tanji, J. (2001). Sequential organization of multiple movements:
Involvement of cortical motor areas. Annual Review of
Neuroscience, 24, 631–651.
Toni, I., Krams, M., Turner, R., & Passingham, R. E. (1998).
The time course of changes during motor sequence learning:
A whole-brain fMRI study. Neuroimage, 8, 50–61.
and ipsilateral motor effects after transcranial direct current
stimulation. NeuroReport, 17, 671–674.
Von Neumann, J., Kent, R. H., Bellinson, H. R., & Hart, B. I.
(1941). The mean square successive difference. Annals
of Mathematical Statistics, 12, 153–162.
White, B., Abbott, L. F., & Fiser, J. (2012). Suppression of
cortical neural variability is stimulus- and state-dependent.
Journal of Neurophysiology, 108, 2383–2392.
Whitfield-Gabrieli, S., & Nieto-Castanon, A. (2012). Conn:
A functional connectivity toolbox for correlated and
anticorrelated brain networks. Brain Connectivity, 2,
125–141.
Wiggs, C. L., & Martin, A. (1998). Properties and mechanisms
of perceptual priming. Current Opinion in Neurobiology,
8, 227–233.
Woods, R. P. (1996). Modeling for intergroup comparisons
of imaging data. Neuroimage, 4, S84–S94.
Xu, T., Yu, X., Perlik, A. J., Tobin, W. F., Zweig, J. A., Tennant,
K. A., et al. (2009). Rapid formation and selective stabilization
of synapses for enduring motor memories. Nature, 462,
915–919.
Yang, G., Lai, C. S. W., Cichon, J., Ma, L., Li, W., &
Gan, W.-B. (2014). Sleep promotes branch-specific
formation of dendritic spines after learning. Science,
344, 1173–1178.
Yang, G., Pan, F., & Gan, W.-B. (2009). Stably maintained
dendritic spines are associated with lifelong memories.
Nature, 462, 920–924.
Yin, H. H. (2010). The sensorimotor striatum is necessary
for serial order learning. Journal of Neuroscience, 30,
14719–14723.
Yousry, T. A., Schmid, U. D., Alkadhi, H., Schmidt, D.,
Peraud, A., Buettner, A., et al. (1997). Localization of
the motor hand area to a knob on the precentral gyrus.
A new landmark. Brain, 120, 141–157.
Zelenin, P. V., Deliagina, T. G., Orlovsky, G. N., Karayannidou,
A., Stout, E. E., Sirota, M. G., et al. (2011). Activity of motor
cortex neurons during backward locomotion. Journal of
Neurophysiology, 105, 2698–2714.
Gabitov, Manor, and Karni
751
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
4
2
7
7
/
3
4
6
/
1
7
9
3
4
6
8
/
9
1
6
7
3
8
o
2
c
7
n
4
_
9
a
/
_
j
0
o
0
c
7
n
5
1
_
a
p
_
d
0
0
b
7
y
5
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1