Neural Systems underlying Lexical Competition:
An Eye Tracking and fMRI Study
Giulia Righi, Sheila E. Blumstein, John Mertus, and Michael S. Worden
Astratto
■ The present study investigated the neural bases of phonologi-
cal onset competition using an eye tracking paradigm coupled
with fMRI. Eighteen subjects were presented with an auditory
target (per esempio., beaker) and a visual display containing a pictorial
representation of the target (per esempio., beaker), an onset competitor
(per esempio., beetle), and two phonologically and semantically unrelated
objects (per esempio., shoe, hammer). Behavioral results replicated ear-
lier research showing increased looks to the onset competitor
compared to the unrelated items. fMRI results showed that lexi-
cal competition induced by shared phonological onsets recruits
both frontal structures and posterior structures. Specifically,
comparison between competitor and no-competitor trials elic-
ited activation in two nonoverlapping clusters in the left IFG,
one located primarily within BA 44 and the other primarily lo-
cated within BA 45, and one cluster in the left supramarginal
gyrus (SMG) extending into the posterior superior temporal
gyrus. These results indicate that the left IFG is sensitive to com-
petition driven by phonological similarity and not only to com-
petition among semantic/conceptual factors. Inoltre, Essi
indicate that the SMG is not only recruited in tasks requiring
access to lexical form but is also recruited in tasks that require
access to the conceptual representation of a word. ■
INTRODUCTION
In retrieving information in the service of a specific goal,
there are many instances in which more than one stimu-
lus representation may be accessed (Mugnaio & Cohen,
2001; Duncan, 1998; Desimone & Duncan, 1995). Così,
competition is created among these multiple representa-
zioni. Tuttavia, typically, only one of the active represen-
tations is appropriate as a response. As a consequence,
competition needs to be resolved in order to achieve a
specific goal or carry out a discrete response. This sce-
nario is not specific to any one cognitive domain, Ma
rather the maintenance of multiple representations, IL
resolution of competition, and ultimately, response selec-
tion are functions associated with the domain-general
mechanisms of cognitive control. Converging evidence
from neuropsychology and neurophysiology suggests
that prefrontal cortex is involved in cognitive control
(cf. Mugnaio & Cohen, 2001).
The resolution of competition is critical for the process-
ing of language. In order to recognize a word, Per esempio,
the listener must select the appropriate word candidate
from the thousands of words in the lexicon, many of which
share sound shape properties. Most models of word rec-
ognition suggest that as the auditory input unfolds over
time, this information is used on-line to continuously pare
down the set of potential competitors. Questo è, at the begin-
ning of a word, all of those words which share onsets are
Brown University, Providence, RI
initially activated; as the auditory input unfolds, those word
candidates that share sound properties remain partially ac-
tivated until sufficient auditory information is conveyed
such that the word is uniquely identified (Luce & Pisoni,
1998; Norris, 1994; McClelland & Elman, 1986; Marslen-
Wilson & Welsh, 1978). Not only is the sound shape of
lexical competitors partially activated but so are their mean-
ing. Per esempio, using a cross-modal priming paradigm,
subjects show semantic priming for words that share on-
sets (Marslen-Wilson, 1987). Presentation of a word such
as general, which has an onset competitor word generous,
primes gift when the prime word is presented before the
input string is disambiguated (cioè., gener…).
Recentemente, effects of lexical competition have been
found using an eye tracking or visual world paradigm
(per esempio., Allopenna, Magnuson, & Tanenhaus, 1998; Spivey-
Knowlton, 1996; Tanenhaus, Spivey-Knowlton, Eberhard,
& Sedivy, 1995). In this paradigm, subjects are presented
with a visual array of objects and are asked to find specific
targets as their eye movements are monitored. One of
the advantages of this technique over priming and lexical
decision judgments is that it provides information about
the time course of lexical access. Tanenhaus et al. (1995)
have shown that there is a systematic correspondence
between the time course of processing a referent name
and the eye movements to a referent object, indicating
that monitoring eye movements over time provides a
continuous measure of how word recognition unfolds
(cf. Yee & Sedivy, 2006). Inoltre, the eye tracking para-
digm allows for the investigation of lexical processing
© 2009 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 22:2, pag. 213–224
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
2
l
2
e
–
2
P
1
D
3
F
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
C
3
N
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
2
0
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
F
.
T
.
/
.
o
N
1
8
M
UN
sì
2
0
2
1
in a more “ecologically valid” manner than most other
paradigms used to investigate word recognition, as sub-
jects are not required to make a metalinguistic judgment
about the lexical status, the sound shape, or meaning of a
stimulus (cf. Yee & Sedivy, 2006).
Previous studies have shown that when subjects are
presented with an auditory target (per esempio., beaker) and a
visual display containing the target object (per esempio., beaker),
an onset competitor (per esempio., beetle), and two phonologically
and semantically unrelated objects (per esempio., shoe, hammer),
they show increased looks to the onset competitor com-
pared to the unrelated items before looking consistently
at the correct target (Dahan, Magnuson, & Tanenhaus,
2001; Dahan, Magnuson, Tanenhaus, & Hogan, 2001;
Allopenna et al., 1998; Tanenhaus et al., 1995). These
findings suggest that as the auditory input unfolds, both
the target stimulus and its onset competitor are partially
activated, and that the subject must resolve this com-
petition among this set of potential lexical candidates in
order to select the correct item in the visual display.
Despite the richness of the literature investigating lexi-
cal competition effects with behavioral methods, less
is known about the neural systems underlying the resolu-
tion of lexical competition. The present study set out to
investigate the neural basis of phonological onset compe-
tition using fMRI coupled with the eye tracking paradigm.
The paradigm used required subjects to find a target
object named auditorily within a visual display containing
four objects while their eye movements were being
tracked. In some trials, two of the object names shared
phonological onsets, whereas in other trials, no competi-
tor was present in the visual display.
A recent study by Yee, Blumstein, and Sedivy (2008)
provides some suggestions about the brain areas that
might be recruited under conditions of onset competi-
zione. Using the same eye tracking paradigm with Brocaʼs
and Wernickeʼs aphasics, they showed that both groups
demonstrated pathological patterns. Specifically, Essi
found that Brocaʼs aphasics with frontal lesions includ-
ing the inferior frontal gyrus (IFG) showed a very weak
onset competitor effect. Questo è, they made few fixations
toward the onset competitor in the visual display. On the
other hand, Wernickeʼs aphasics with temporo-parietal
lesions including the posterior portions of the superior
temporal gyrus (STG) showed a larger competitor effect
than age-matched controls. These results suggest that
both anterior and posterior language processing areas
are influenced by phonological onset competition. How-
ever, the differences between these two subject groups
also suggest that these regions might have different func-
tional roles.
Posterior temporo-parietal brain structures are involved
in the initial processing of a lexical entry both at the pho-
nological and the semantic level. Hickok and Poeppel
(2000) have proposed that mapping from sound to mean-
ing recruits a network of regions including the posterior
STG, the supramarginal gyrus (SMG), and the angular
gyrus (AG). The left temporo-parietal region, including
the SMG and the AG, is also involved in mapping of sound
structure to a phonological representation and in storing
this representation (Paulesu, Frith, & Frackowiak, 1993).
These regions have also been shown to be sensitive to pho-
nological competition. In particular, using a lexical decision
paradigm, Prabhakaran, Blumstein, Myers, Hutchinson, E
Britton (2006) showed increased activation in the left SMG
when subjects performed a lexical decision task on words
that were from high-density neighborhoods (cioè., had a lot
of phonological competitors) compared to words from low-
density neighborhoods (cioè., had few phonological competi-
tori) (cf. Luce & Pisoni, 1998 for discussion of behavioral
effects of neighborhood density on word recognition).
Although posterior regions appear to be involved in
processing the sound structure and meaning of words,
as discussed earlier, frontal regions appear to be involved
in “executive control functions” (per esempio., Duncan, 2001; Mugnaio
& Cohen, 2001; Duncan & Owen, 2000; Smith & Jonides,
1999). Within prefrontal cortex there is evidence that the
left IFG is recruited in the resolution of competition and
in the ultimate selection of the appropriate response.
Thompson-Schill and colleagues (Thompson-Schill,
DʼEsposito, & Kan, 1999; Thompson-Schill et al., 1998;
Thompson-Schill, DʼEsposito, Aguirre, & Farah, 1997) Avere
suggested that the left IFG is involved in making a selec-
tion among competing conceptual representations. In a re-
cent study, Snyder, Feignson, and Thompson-Schill (2007)
showed increased activation in the left IFG as a function of
semantic conflict during both semantic and phonological
compiti. Allo stesso modo, Gold and Buckner (2002) found increased
activation in the same portions of left inferior prefrontal cor-
tex during controlled retrieval of both semantic and phono-
logical information.
Although the IFG is clearly involved in executive con-
trol, several recent proposals have suggested that there
is a functional division within the IFG. Some have pro-
posed that the left IFG is partitioned as a function of
linguistic domain with a phonology-specific processing
mechanism in the posterior portion of the IFG and
a semantic-specific processing mechanism in the ante-
rior portion of the IFG (Burton, 2001; Poldrack et al.,
1999; Fiez, 1997; Buckner, Raichle, & Petersen, 1995).
Others (Badre & Wagner, 2007) have proposed that the
left IFG can be divided on the basis of different functional
processes. They propose that the most anterior portion is
responsible for the maintenance of multiple conceptual
representations that have been activated either by top–
down or bottom–up processes, and the more posterior
portion of the IFG is responsible for selecting the ap-
propriate task-relevant representation (Badre & Wagner,
2007).
The goal of this study is to investigate the neural bases
of phonological onset competition. Based on the findings
reviewed above, it is hypothesized that both posterior
regions, specifically the left SMG, and anterior regions,
specifically the left IFG, will be recruited under condi-
214
Journal of Cognitive Neuroscience
Volume 22, Numero 2
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
2
l
2
e
–
2
P
1
D
3
F
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
C
3
N
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
2
0
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
/
F
T
.
.
.
o
N
1
8
M
UN
sì
2
0
2
1
tions of phonological onset competition. Although the
role of the SMG is uncontroversially related to phono-
logical analysis and access of lexical form, the role of
the IFG in resolving phonological competition is less
clear. Prabhakaran et al. (2006) failed to show increased
IFG activation as a function of phonological competition,
suggesting that this area may be recruited only in re-
solving competition among conceptual representations.
Activation should occur in the IFG in the current study
because the appropriate conceptual representation
needs to be selected from the competing conceptual
representations in order to look at the correct object in
the visual array. Tuttavia, IFG activation could also be
modulated by phonological competition, independent
of semantic/conceptual competition. In that case, IL
IFG would appear to play a domain general role in re-
solving competition not only as a function of competing
conceptual representations but also as a function of com-
peting phonological representations. Given that in our
paradigm, competition is based upon similarities in the
phonological form among words, and that no specific
semantic or phonological judgment is required, we hope
to provide evidence to further specify the role of the left
IFG in the resolution of lexical competition.
METHODS
Norming Experiments
Two preliminary norming experiments were conducted
in order to select easily identifiable visual exemplars for
each of the object names to be used as stimuli in the eye
tracking experiment.
Subjects
Forty college-aged subjects were tested. All subjects were
recruited from the Brown University community and
were paid for their participation. All subjects were native
speakers of American English and had either normal or
corrected-to-normal vision and no hearing or neurological
deficit.
Materials
Sixty names of highly pictureable common objects served
as target words. Each of the target words was paired with a
highly pictureable noun that shared either the entire first
syllable or the onset and the vowel of the first syllable with
Esso (per esempio., lamb–lamp) to create a set of 60 onset competi-
tori. The target–onset competitor pairs were chosen from
those used in previous experiments and unpublished data
that have investigated the behavioral effects of phono-
logical onset competition (McMurray, personal communi-
catione; Yee et al., 2008; Yee & Sedivy, 2006). An additional
120 highly pictureable nouns were selected to serve as
filler items. The filler items were chosen from the MRC
psycholinguistic database (The University of Western
Australia), such that there were no significant differences,
as assessed by t tests, in mean word frequency, number of
syllables, concreteness, and imageability scores between
the sets of targets, onset competitors, and fillers. The vi-
sual stimuli consisted of two distinct color photographs of
each target, onset competitor, and filler object. The color
photographs were taken from the Hemera Photo Object
database (Hemera Technologies, Toronto, Canada) E
Google Images.
Procedure
The first preliminary experiment was a naming task de-
signed to make sure that subjects consistently named
each of the pictures chosen, and that the name given
to each object corresponded to the name intended to
be associated with each picture.
Twenty participants divided into two groups partici-
pated in this experiment. Each subject group saw only
one exemplar per object, for a total of 240 images. In
each trial, subjects saw a picture of an object on a white
background and were asked to type the name of the ob-
ject they thought it represented. The picture remained
on the screen for 1 sec, and subjects had unlimited time
to type the name. If more than one third of the subjects
misidentified a picture, a new image was selected to be
paired with the object in question.
The second preliminary experiment was conducted to
validate the new set of images that met the identification
criterion from the previous experiment plus the new ex-
emplars selected to replace those that did not meet the
above specified criterion. In this second experiment, sub-
jects were asked to make a yes/no judgment about the
word–picture pairs to provide a measure of identification
accuracy, and a measure of ease of identification by re-
cording RT latencies.
Two groups of 10 new subjects participated in this ex-
periment. Each subject group saw only one visual exemplar
per object name, for a total of 240 images. Each picture
appeared on the screen for 1 sec and was followed by a
visually presented word that was either the correct name
for the object presented or a different object name. Sub-
jects were asked to decide whether the word they read
was the correct name for the object presented or not. Ac-
curacy and RTs were measured. Items were eliminated
from the stimulus set on the basis of three criteria: (1) if
more than one third of the subjects gave incorrect answers
in the word–picture matching trials; (2) if more than one
third of the subjects had RT latencies longer than two
standard deviations over their individual mean RT laten-
cies for correct responses; E (3) if the across-subjects
mean RT for the correct responses to a specific item was
more than two standard deviations above the across-
subjects mean for the set of correct responses for the match-
ing trials.
Righi et al.
215
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
2
l
2
e
–
2
P
1
D
3
F
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
C
3
N
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
2
0
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
/
.
.
F
T
.
o
N
1
8
M
UN
sì
2
0
2
1
Based on the results of the two norming experiments,
54 target word–object pairs, 54 objects depicting phono-
logical onset competitors, E 108 filler items were se-
lected as stimuli for the eye tracking experiment. IL
stimuli used in the experiment are listed in the Appendix.
fMRI and Eye Tracking Experiment
Participants
Eighteen participants (14 women) ranging in age from 18
A 32 years (mean age = 23 years) took part in one scan-
ning session. Participants were all right-handed as assessed
by the Oldfield Handiness Inventory (Oldfield, 1971). Par-
ticipants reported normal hearing and normal or corrected-
to-normal vision, and no neurological impairment. Tutto
participants gave written informed consent according to
the guidelines of the Human Subjects Committee of
Brown University. All participants were screened for MR
safety prior to the scanning session. Data from one partici-
pant were excluded from both the behavioral and fMRI
analyses because of technical difficulties.
fMRI Acquisition
Scanning was performed on a 3-T Siemens TIM Trio scan-
ner at Brown University, using a standard eight-channel
head coil, outfitted with a mirror for back projection, an
infrared illuminator, and an infrared mirror for eye track-
ing. High-resolution anatomical images were collected
using a 3-D T1-weighted magnetization prepared rapid ac-
quisition gradient-echo sequence (TR = 2.25 sec, TE =
2.98 msec, 1 mm3 isotropic voxel size). Functional images
were acquired using a multislice, ascending, interleaved
EPI sequence (TR = 2.7 sec, TE = 28 msec, FOV = 192,
45 slices, 3 mm × 3 mm in-plane resolution, 3 mm slice
thickness). A total of 164 volumes were acquired during
each run. Inoltre, two “dummy” volumes were ac-
quired at the start of each run to allow the MR signal to reach
steady state; these volumes were discarded by the scanner.
Materials
The stimulus set consisted of 54 target auditory stimuli
(mean duration = 480 msec) E 108 visual displays.
The auditory target stimuli were recorded by a male
speaker in a sound-attenuated room. Each visual display
consisted of a 3 × 3 grid on a white background contain-
ing four pictures, one picture in each of the corners of
the grid. The visual grid subtended 16° × 16° visual angle,
with each cell being approximately 5° × 5° visual angle.
The competitor trials consisted of 54 visual displays con-
taining four objects corresponding to the target auditory
stimulus (per esempio., beaker), the onset competitor (per esempio., beetle),
and two fillers (per esempio., train, hammer) (Guarda la figura 1). IL
remaining 54 displays served as the no-competitor trials
and contained an object corresponding to the target audi-
Figura 1. Example of a competitor trial display. The target object
(beaker) shares the onset with one of the objects in the display
(beetle), whereas the other two objects are unrelated semantically
and phonologically.
tory stimulus (per esempio., beaker) and three filler objects that
were phonologically, semantically, and visually unrelated
to the target (per esempio., ship, marble, comb). Each auditory
target token was paired with two displays, one containing
the target, the onset competitor, and two fillers (competi-
tor trial), and one containing the target and three fillers
(no-competitor trial). Although each auditory token was
repeated across the two conditions, none of the object
pictures was seen more than once. Questo è, subjects saw
two different exemplars of each object, one in the com-
petitor trials and one in the no-competitor trials. The posi-
tion of targets and competitors within the grid was
counterbalanced, such that targets and competitors ap-
peared an equal number of times in all four positions.
Inoltre, all repeated types (cioè., there were two visual
exemplars for each object) never appeared in the same
location across runs.
Procedure
An SMI iView × MRI (SensoMotoric Instruments, Needham,
MA) eye tracker was used. An infrared camera located at the
edge of the MRI bed was used to monitor participantsʼ eye
movements. The camera recorded the participantʼs eye
movements at 60 Hz, with accuracy of greater than 1° visual
angle. Stimuli were presented with Bliss software (Mertus,
2002) on a Dell laptop, connected to an LCD projector,
and back-projected to the head coil mirror, and through
sound-attenuating pneumatic headphones.
Subjects participated in two experimental runs of an
event-related design, each consisting of 54 stimulus pre-
216
Journal of Cognitive Neuroscience
Volume 22, Numero 2
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
2
l
2
e
–
2
P
1
D
3
F
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
C
3
N
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
2
0
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
.
T
.
.
/
F
o
N
1
8
M
UN
sì
2
0
2
1
sentations. In each run, they saw an equal number of
target–competitor and target–filler displays in random
order and were exposed once to all of the auditory targets;
one-half of the targets were paired with target–competitor
displays and the other half were paired with target–filler
displays. Hence, if an auditory target was paired with a
target–competitor display in the first run, the same audi-
tory target would be paired with a target–filler display in
the second run. The occurrence of target–competitor and
no-competitor displays was counterbalanced across con-
ditions. Overall, subjects were presented with 54 target–
competitor trials and 54 target–filler trials.
Each trial began with the presentation of the four-
picture array in order to provide the subject with a
chance to briefly scan the display. One second later, UN
red fixation cross appeared in the center of the screen,
and the objects disappeared. Participants were instructed
to fixate on the cross when it appeared. The fixation cross
remained on the screen for 1 sec followed by the re-
appearance of the four objects with the simultaneous
auditory presentation of the name of one of the objects
present in the display (target). The four objects remained
on the screen for 1.5 sec. Subjects were instructed to
find and look at the object that had been named until it
disappeared. Presentation of stimuli was jittered according
to a uniform distribution of nine trial onset asynchronies
(TOA values ranging from 4.5 A 11.7 sec in 0.9-sec steps).
During the TOA intervals, the screen was left blank. Each
TOA bin was used an equal number of times in each run,
and an equal number of competitor and noncompetitor
trials were assigned to each TOA bin in each run.
Data Analysis Methods
fMRI Data Preprocessing
fMRI data were analyzed using AFNI (Cox & Hyde, 1997;
Cox, 1996). Preprocessing steps included slice acquisi-
tion time correction for each run separately. The runs
were then concatenated for each subject to carry out head
motion correction by aligning all volumes to the fourth
collected volume using a six-parameter rigid-body trans-
form. These data were then warped to Talairach and
Tournoux (1988) space and were resampled to 3-mm iso-
tropic voxels. Lastly, the data were spatially smoothed
using a 6-mm full-width half-maximum Gaussian kernel.
fMRI Statistical Analysis
Each subjectʼs preprocessed EPI data were regressed to
estimate the hemodynamic response function for the two
experimental conditions (competitor condition vs. non-
competitor condition). Response functions were esti-
mated by convolving vectors containing the onset times
of each auditory stimulus with a stereotypic gamma-variate
hemodynamic response function provided by AFNI (Cox
& Hyde, 1997; Cox, 1996). The calculation of the raw
fit coefficients for each voxel was carried out using the
AFNI 3dDeconconvolve program. The raw coefficients
were then converted to percent signal change by dividing
each voxel coefficient by the experiment-wise mean acti-
vation for that voxel. Because of the repetition of audi-
tory targets across the two runs, a group analysis was
carried out by entering the percent signal change data
into a mixed factor two-way ANOVA, with subject as a
random factor, and condition (competitor vs. no com-
petitor) and run (first run vs. second run) as fixed factors.
The resulting statistical maps were corrected for multi-
ple comparisons using Monte Carlo simulations. A voxel-
level threshold of p < .025 and a cluster-level threshold
of p < .05 (48 contiguous voxels) were used for all com-
parisons. The atlases used to locate the anatomical struc-
tures were the Anatomy Toolbox atlases (Eickhoff et al.,
2007; Eickhoff, Heim, Zilles, & Amunts, 2006; Eickhoff
et al., 2005).
RESULTS
Eye Tracking Results
Eye tracking data were initially processed by counting the
number of fixations that each subject made on target,
competitor, and filler objects in each trial, sampled every
17 msec. Only fixations initiated after the onset of the
auditory target and lasting longer than 100 msec were in-
cluded in the analysis. We defined four regions that con-
tained target, competitor, and filler object pictures. Each
region was a 5° × 5° of visual angle square in each of
the four corners of the display. A fixation to a specific ob-
ject was defined as consisting of the time that a saccade
moved the eye into the specific region until a saccade
moved the eye out of that region. Thus, saccades in
which the eye did not move out of the region were in-
cluded as part of the fixation time for that region.
If the subject did not fixate on the target by the end of the
trial, that trial was excluded from the analysis. On average,
4% of the trials were excluded for 15 out of the 17 subjects.
The remaining two subjects proved to be difficult for the
eye tracker to calibrate. Thus, approximately one third of
their trials were excluded from the analysis. For each sub-
ject, the proportion of fixations to target, competitor, and
filler objects was computed by averaging the number of
fixations to each object in seven 200-msec windows, and
dividing it by the total number of fixations across all objects
in the same time window. This analysis was performed on
the data of both runs averaged together, and on the data
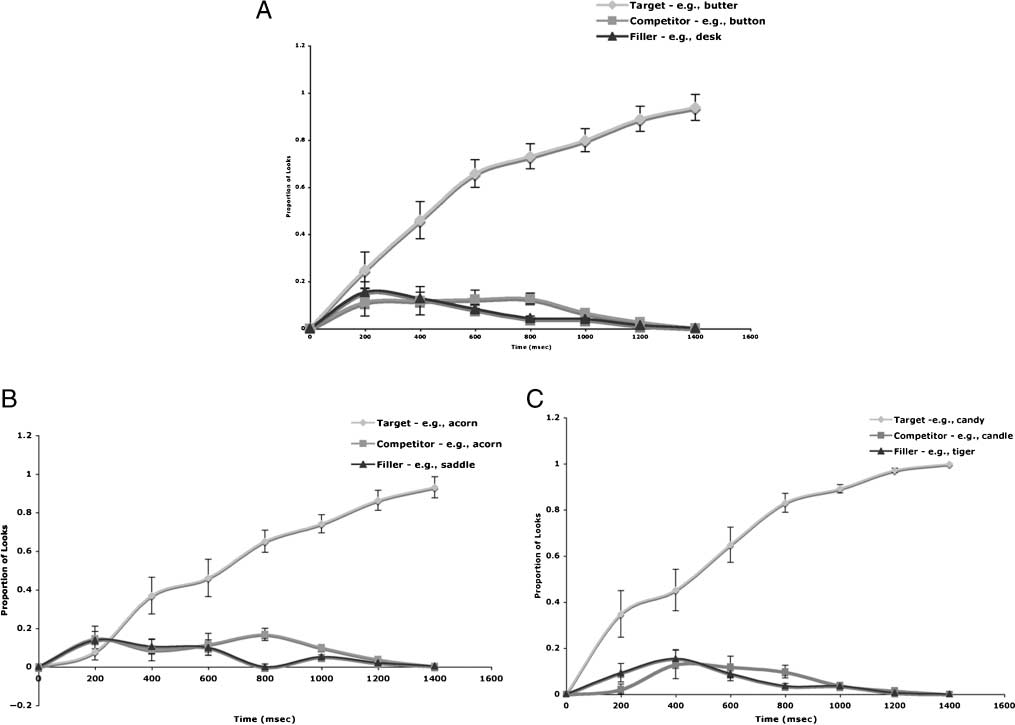
from each run alone. Figure 2A shows the mean propor-
tion of looks to the target, onset competitor, and the aver-
age of the two unrelated pictures in the competitor trials
across both runs, Figure 2B shows the same analysis done
on the data from the first run, and Figure 2C shows the
results of the same analysis from the second run.
The data were statistically analyzed to determine
whether a significant competitor effect emerged and to
ascertain the time course of this effect. For the purpose
of the analysis, a trial was defined as starting 200 msec after
Righi et al.
217
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
2
e
-
2
p
1
d
3
f
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
c
3
n
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
2
0
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
/
.
t
.
.
f
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
2
e
-
2
p
1
d
3
f
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
c
3
n
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
2
0
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
t
/
.
f
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Proportion of fixations over time to the target, the onset competitor, and the average of the two unrelated items in competitor trials.
Zero corresponds to the onset of the auditory target. Standard error bars are shown for every data point. (A) Data averaged from both runs.
(B) Data from the first run. (C) Data from the second run.
auditory stimulus onset, because it takes about 200 msec to
launch a saccade (Altman & Kamide, 2004; Hallet, 1986),
and ending when the display disappeared from the screen.
A two-way repeated measures ANOVA of the proportion of
looks with object (competitor, filler) and time bin (seven
200-msec bins) as factors revealed a marginally significant
main effect of condition [F(1, 16) = 4.04, p < .07], show-
ing that overall competitors were fixated on more often
than filler items. There was no significant main effect for
time bin [F(6, 16) = 1.7, p < .13]. Moreover, no significant
interaction was found [F(6, 96) = 1.6, p < .16].
In order to assess the presence of competitor effects
across the two runs, separate two-way (Condition × Time
bin) repeated measures ANOVAs were also conducted.
Results for the first run revealed a significant main effect
for condition [F(1, 16) = 5.8, p < .05], showing that com-
petitors were fixated on significantly more than filler ob-
jects. The analysis also revealed a marginally significant
main effect for time bin [F(6, 16) = 1.9, p < .08], but no
significant interaction [F(6, 96) = 1.5, p < .2]. Results from
the second run revealed no significant main effect for condi-
tion [F(1, 16) = 0.4, p < .6] and no significant interaction
[F(6, 96) = 1.7, p < .12]. However, there was a marginally
significant main effect for time bin [F(6, 16) = 2.1 p < .06].
fMRI Results
Table 1 provides a summary of the results of the two-way
(Condition × Run) ANOVA. Table 1A lists those clusters
that showed a main effect of condition and Table 1B lists
those clusters that showed a main effect of run ( p < .05,
corrected). No clusters showed a significant Condition ×
Run interaction. The focus of discussion will be on those
clusters falling within regions previously identified to be
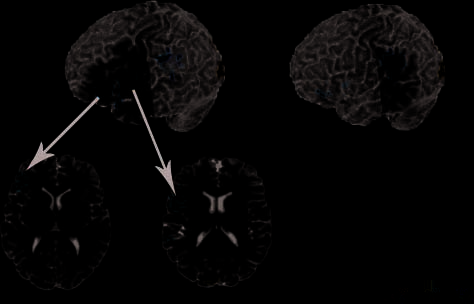
involved in language processing (see Figure 3).
Several clusters showed a significant effect of condi-
tion with greater activation for competitor trials than for
no-competitor trials. As Figure 3 shows, the left inferior
frontal gyrus (LIFG) contains three clusters of activation
that responded more strongly in the competitor condition
compared to the no-competitor condition. The largest of
these clusters was located primarily within the pars oper-
cularis (BA 44, 71% of active voxels), but also extended
into the pars triangularis (BA 45, 20% of active voxels).
218
Journal of Cognitive Neuroscience
Volume 22, Number 2
Table 1. Significant Clusters Activated in the Condition × Run ANOVA
Cortical Region
Brodmannʼs Area
Cluster Size
x
y
z
Talairach Coordinates
(A) Main Effect of Condition
Competitor > No-Competitor
Left supramarginal gyrus
Left inferior frontal
Left cingulate gyrus
Left inferior frontal gyrus
Right inferior parietal lobule
Left insula
Competitor < No-Competitor Left cuneus Left lingual gyrus Right lingual gyrus (B) Main Effect of Run First Run > Second Run
Left declive
Right superior parietal lobule
Right middle temporal cortex
Left superior/inferior parietal
Left middle occipital gyrus
Left postcentral gyrus
First Run < Second Run
Posterior cingulate
Left superior parietal lobule
40/2/13
44/45
24/23
45/46/10
40
13/47
18/17
18/17
18/17
17/18
7/2/40
37/19
7
18/19/37
3/40/22
31/23/7
7/5/39
139
118
77
70
69
51
59
55
50
330
145
127
109
90
81
510
160
58.5
49.5
1.5
46.5
−49.5
31.5
4.5
19.5
−10.5
1.5
−34.5
−52.5
31.5
25.5
58.5
1.5
37.5
46.5
−4.5
4.5
−43.5
43.5
−13.5
91.5
97.5
82.5
73.5
46.5
61.5
55.5
88.5
25.5
49.5
64.5
35.5
17.5
38.5
11.5
50.5
−9.5
26.5
−6.5
−6.5
−12.5
62.5
−0.5
56.5
11.5
35.5
17.5
50.5
Clusters thresholded at a cluster-level threshold of p < .05 with a minimum of 48 contiguous voxels, and at a voxel-level threshold of p < .025, t =
2.459 (corrected). Coordinates indicate the maximum intensity voxel for that cluster in Talairach and Tournoux space. The first Brodmann’s area
listed corresponds to the location of the maximum intensity voxel. If the cluster extended into other BAs, those are also listed (see text for details).
The second cluster was located primarily within the pars
triangularis (BA 45, 95% of active voxels), and extended
marginally into the middle frontal gyrus (4% of active
voxels). A third frontal cluster was located primarily in
the left insula (56% of active voxels), but extended into
the pars orbitalis (BA 47, 28% of active voxels). It is im-
portant to note that, despite the proximity of these three
frontal clusters, they are anatomically distinct.
Beyond the frontal regions, there were also significant
clusters of activation in the left and right temporo-parietal
region showing increased activation in the competitor con-
dition compared to the no-competitor condition. The left
hemisphere cluster was located within the SMG (50% of ac-
tive voxels). This cluster extended into the inferior parietal
lobule (22% of active voxels) and the posterior portion of
the left STG (7% of active voxels). The right hemisphere
cluster was primarily located in the right SMG (71% of active
voxels) and extended into the inferior parietal lobule (27%
of active voxels). No clusters emerged in language areas in
which there was greater activation in the noncompetitor
condition than in the competitor condition.
The two-way ANOVA also revealed several clusters that
showed a significant main effect of run. The clusters that
showed a stronger response in the first run compared
to the second run were located in left middle occipital
cortex, the superior parietal lobules bilaterally, the left
postcentral gyrus extending into the posterior portion
of the STG, and right middle temporal cortex extend-
ing into extrastriate visual cortex. The clusters showing
a larger response in the second run compared to the first
run were located in the left superior parietal lobule and
posterior cingulate cortex.
Righi et al.
219
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
2
e
-
2
p
1
d
3
f
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
c
3
n
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
2
0
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
/
.
f
.
t
.
o
n
1
8
M
a
y
2
0
2
1
DISCUSSION
In the present study, the neural bases of phonological
onset competition were investigated. The results show
that lexical competition induced by shared phonological
onsets recruits both frontal structures (i.e., the LIFG) and
posterior structures (i.e., the SMG).
Behavioral Findings
The behavioral results replicate previous findings showing
that subjects look more at pictures of objects that share
onsets with an auditory target than to pictures of unrelated
objects (Dahan, Magnuson, & Tanenhaus, 2001; Dahan,
Magnuson, Tanenhaus, et al., 2001; Allopenna et al.,
1998; Tanenhaus et al., 1995). This effect was marginally
significant across both runs, and statistically significant in
the first run. The lack of significance in the second run
suggests that the repetition of auditory targets reduced
the strength of the competition effect. However, as shown
in Figure 2C, even in the second run, the proportion of
looks between competitors and filler items showed greater
looks to competitor trials than to no-competitor trials.
Thus, the presence of an onset competitor in the stimulus
array affects access to the auditorily presented lexical tar-
get. It is worth noting that the initial divergence between
the proportion of looks to the competitor and filler items
emerged about 200–400 msec later than that reported in
previous studies. Previous studies have shown that a di-
vergence between looks to the competitor and filler ob-
jects emerges as early as 200 msec after word onset (Yee
et al., 2008; Yee & Sedivy, 2006).
Figure 3. Maps thresholded at a voxel-level threshold of p < .025,
t = 2.459, and clusters shown correspond to a corrected significant
level of p < .05. (A) Clusters in the LIFG showing greater activation
for competitor trials compared to no-competitor trials. Sagittal slice
shown at x = 35 and coronal cut shown at y = 15. (B) Cluster in
the LIFG (BA 45). Axial slice shown at z = 11. (C) Cluster in the
LIFG (BA 44/45). Axial slice shown at z = 17. (D) Cluster in the left
temporo-parietal region showing greater activation for competitor
trials compared to no-competitor trials. Sagittal slice shown at
x = 50, coronal slice shown at y = 20.
In the current study, the delay of the onset competitor
effect may be due to a number of methodological differ-
ences. First, traditionally, a small number of competitor
trials are presented together with a larger number of no-
competitor trials to ensure that the subject remains unaware
of the experimental manipulation. In the present study, an
equal number of competitor and no-competitor trials were
used, because the addition of more no-competitor trials
would have lengthened significantly the amount of time
subjects had to spend in the scanner. It is possible that
the shift in the ratio of competitor to no-competitor trials
influenced the size and latency of the competitor effect ob-
served here. Second, earlier studies required subjects not
only to look at the target but also to overtly point to the tar-
get with their finger or with a mouse. In the current study,
subjects were only required to look at the target picture.
The coupling of a motor and visual action may have a fa-
cilitatory effect on subjectsʼ responses that is not observed
when only a visual response is required. Third, in contrast to
earlier studies that used a head-mounted eye tracker that
allowed subjects to move their heads as they were doing
the task, participants were unable to move their heads in
the scanner. This difference might have affected the ease
with which subjects moved their eyes, as it is more natural
to follow eye movements with head movements. Lastly, the
stress of doing the task while in the scanner might have also
affected the ease with which participants launched eye
movements. In fMRI studies, it is not uncommon to find
slower RTs in behavioral tasks while in the scanner com-
pared to the same tasks outside of the scanner. For exam-
ple, RT latencies in a lexical decision task were on the order
of 200–300 msec slower in the scanner than outside the
scanner (cf. Prabhakaran et al., 2006 and Luce & Pisoni,
1998 where the same stimuli were used). It is possible that
eye movements are subject to a similar phenomenon.
The Effects of Competition in
Temporo-parietal Structures
The comparison between competitor and no-competitor
trials showed increased activation in the left posterior
STG and SMG. Activation of the left STG and SMG has
been previously identified to be involved in phonological
processing (e.g., Gelfand & Bookheimer, 2003; Binder &
Price, 2001; Hickok & Poeppel, 2000). More recently, it
has been shown that the left SMG is recruited under con-
ditions of phonological competition (Prabhakaran et al.,
2006). Prabhakaran et al. (2006) observed increased activa-
tion in the SMG when subjects performed a lexical deci-
sion task on words with many phonological competitors
compared to words with few phonological competitors.
Of interest, the current study showed activation not only
in the left SMG but also in the right SMG. However, it is
worth noting that although not discussed, other studies
have shown that the right SMG is also recruited in phono-
logical tasks (Price, Moore, Humphreys, & Wise, 1997).
That the right SMG is activated in the current study sug-
220
Journal of Cognitive Neuroscience
Volume 22, Number 2
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
2
e
-
2
p
1
d
3
f
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
c
3
n
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
2
0
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
/
.
f
.
t
o
n
1
8
M
a
y
2
0
2
1
gests that right hemisphere mechanisms are also recruited
under conditions of phonological competition.
The present findings confirm the recruitment of the
SMG under conditions of competition driven by similari-
ties in phonological form. However, these results differ
from those of Prabhakaran et al. (2006) in two important
ways. First, the findings of the current study show that the
SMG is not only recruited in a task requiring access to
lexical form but it is also recruited in a task that requires
access to the conceptual representation of a word. In the
Prabhakaran et al. study, participants had to make a lexical
decision on a singly presented auditory target stimulus.
Thus, the subjects had only to overtly access the lexical
form of the word to make a decision. In contrast, in the
current study, subjects were required to look at a named
picture. In order to do so, they had to access the concep-
tual representation of the picture in order to match its
shape to the auditorily presented target. Second, the find-
ings of the current study show that the SMG is not only
recruited when phonological competition is implicit but
is also recruited in a task in which competition is explicitly
present in the stimulus array. That is, in the Prabhakaran
et al. study the phonological competitors were never pre-
sented, whereas in the current study the phonological
competitors were presented in the stimulus array. Thus,
findings of the present study indicate that the left SMG
is sensitive not only to phonological competition intrinsic
to a stimulus but also to phonological competition that is
reinforced by conceptual representations. The influence of
conceptual representations on activation in the SMG could
result from two-way connections with frontal areas as-
sumed to be involved in the manipulations of these repre-
sentations. This is not surprising given that a previous study
(Gold & Buckner, 2002) has shown that the left SMG co-
activates with domain-general frontal regions when subjects
are performing a controlled phonological judgment.
The Effects of Competition in Frontal Structures
There is a large body of evidence suggesting that ventro-
lateral portions of prefrontal cortex, which include the
IFG, are involved in guiding response selection under con-
ditions of conflict or competition (cf. Badre & Wagner,
2004; Miller & Cohen, 2001; Desimone & Duncan, 1995).
Selection is undoubtedly present in our task. Thus, it is
not surprising that increased left IFG activation was found.
However, there are a number of experimental factors that
could have contributed to the modulation of IFG activa-
tion. In the present task, the presence of an onset compe-
titor in the stimulus array results not only in the activation
of the phonological form of the target stimulus and its pho-
nological competitor but also in activating the conceptual
representations of these competing stimuli. Thus, competi-
tion needs to be resolved at both phonological and con-
ceptual levels of representation.
Thompson-Schill et al. (1997, 1998, 1999) have pro-
posed that the left IFG is a domain-general mechanism
that guides selection among competing conceptual repre-
sentations. In support of this hypothesis, a recent study by
Snyder et al. (2007) found increased activation in the left
IFG when multiple semantic representations were compet-
ing regardless of whether the subject performed a phono-
logical or semantic judgment task. Thus, in their study,
competition was driven by semantic factors. In the current
study, competition was driven by phonological factors,
and activation of the IFG was modulated by the presence
of phonological onset competition. Taken together, these
findings support the hypothesis that the IFG is domain-
general in that it is responsive to phonological as well as
semantic/conceptual competition and it is recruited even
when the response is not dependent on either semantic
or phonological judgments.
Nonetheless, competitor trials elicited more activation
than noncompetitor trials in the current study in three non-
overlapping clusters in the left IFG. The largest cluster was
located primarily within BA 44 (71% of active voxels) and
the second cluster was located within BA 45 (95% of active
voxels). A third cluster in the insula extended into BA 47.
The presence of these distinct clusters of activation sug-
gests that there is a functional division of the IFG. It is
to this issue that we now turn.
There are several neuroimaging studies showing a func-
tional distinction between the anterior and posterior IFG
on the basis of whether semantic or phonological informa-
tion is processed. These studies have shown that the ante-
rior portion of the left IFG, corresponding to BA 45/47, is
activated in tasks requiring semantic processing, whereas
the posterior portion of the left IFG, corresponding to
BA 44, is activated in tasks requiring phonological pro-
cessing (Burton, Small, & Blumstein, 2000; Poldrack et al.,
1999; Fiez, 1997; Buckner et al., 1995). This framework
would suggest that the clusters found in the present study
are responding separately to competition at the semantic/
conceptual level and at the phonological level. Thus, the
emergence of a cluster in BA 44 is consistent with the view
that it is responsive to phonological factors, whereas the
emergence of clusters in BA 45 and including BA 47 is con-
sistent with the view that they are responsive to semantic/
conceptual factors. Given that the presence of phonological
competition appeared to activate more than one concep-
tual representation, thus increasing conceptual competition
in the competitor condition, it is reasonable to suggest that
BA 45/47 and BA 44 are tightly coupled.
Other researchers have proposed that there is a func-
tional subdivision of the IFG based upon different processes
involved in cognitive control. Badre and Wagner (2007) sug-
gest that, within the IFG, two distinct subregions perform
two different functions: controlled retrieval and postretrieval
selection (Badre & Wagner, 2007). Controlled retrieval re-
fers to the top–down activation of semantic knowledge rel-
evant to the task at hand and it is suggested to recruit BA 47.
If more than one knowledge representation becomes ac-
tive, postretrieval selection is needed to resolve competi-
tion, regardless of the form of these representations (e.g.,
Righi et al.
221
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
2
e
-
2
p
1
d
3
f
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
c
3
n
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
2
0
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
t
.
f
.
/
.
o
n
1
8
M
a
y
2
0
2
1
semantic, phonological, and perceptual). In this framework,
postretrieval selection is implemented in BA 45.
In the current study, BA 47 (which was part of a larger
cluster including the insula) could be recruited by the
activation of task-relevant semantic knowledge. The in-
creased activation found in BA 47 for competitor trials
could reflect the higher number of semantic represen-
tations that are activated in the competitor condition.
Namely, not only are the semantic representations asso-
ciated with the target stimulus activated but, in addition,
the semantic representations of the phonological com-
petitor are as well. The cluster found in BA 45 could reflect
the domain-general postretrieval selection mechanism. As
to the role of BA 44, Badre and Wagner (2007) agree with
its involvement in phonological processing and further
suggest that its proximity to speech production regions
might implicate it as a selection mechanism tied with a
specific overt response (Badre & Wagner, 2007). Thus,
the activation found in BA 44 could be interpreted as in-
volved in carrying out response-related selection.
To summarize, the present study showed that the left
IFG is sensitive to competition driven by phonological
similarity, and not only to competition among semantic/
conceptual factors. Moreover, the activation found in the
left IFG is consistent with a functional segregation of this
region in anterior and posterior portions both on the basis
of linguistic domain and on the basis of different pro-
cesses involved in cognitive control. Further studies will
be necessary to determine whether these interpretations
are mutually exclusive, or whether the IFG can be func-
tionally divided according to both models.
APPENDIX (continued )
(continued )
Target
Competitor
Filler 1
Filler 2
CANE
CANOE
CARROT
CASKET
CAT
CAKE
STATUE
LION
CASSETTE
OLIVE
FEATHER
CARRIAGE
SPEAKER
FOOTBALL
CASTLE
TOASTER
MARBLE
CAB
NET
WATCH
CHOCOLATE
CHOPSTICKS
LOBSTER
HELMET
CLOWN
COAT
CRAYON
DOLLAR
HEN
HOLE
HORN
CLOUD
COMB
CRADLE
DOLPHIN
HEAD
HOME
HORSE
BOX
TRUCK
RACKET
MIRROR
SHIP
BEER
CHAIR
SOAP
GLASS
IRON
HONEY
DRILL
DRESS
MAP
HORNET
HORSHOE
ONION
DOCTOR
LAMB
LETTER
MONEY
LAMP
ROCK
GLOVE
LETTUCE
TRUMPET
SCISSORS
MONKEY
PEPPER
BRACELET
MUSTARD
MUSHROOM
BADGE
PADDLE
PADLOCK
LIGHTER
PANCAKE
PENNY
PIER
PANDA
PENCIL
PEACH
DANCER
BANANA
KNIFE
WHEEL
BULLET
ORANGE
ANCHOR
COW
PITCHER
PITCHFORK
HAMSTER
BALLOON
GUITAR
RIBBON
RING
WALNUT
CUP
APPLE
CHERRY
SCALE
MOUSE
TOILET
PIANO
CIGAR
DRUM
BRAIN
DESK
MATCH
BRUSH
GARLIC
BELT
ARROW
BOTTLE
TREE
DAISY
NEST
PUMPKIN
BARREL
QUARTER
SOLDIER
SANDAL
SANDWICH
RABBIT
CHESTNUT
SEAHORSE
SEESAW
RULER
ROPE
SNAKE
SOFA
TIGER
TOWEL
TULIP
TURTLE
SNAIL
SODA
TIRE
TOWER
TUBA
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
2
e
-
2
p
1
d
3
f
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
c
3
n
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
2
0
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
f
/
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Competitor
Filler 1
Filler 2
APRON
BABY
NEEDLE
PIPE
SADDLE
SHOVEL
BATHTUB
NECKLACE
PENGUIN
PIZZA
PLATE
PUPPY
ROAD
PEANUT
PLANE
PUPPET
ROLL
BANJO
BEETLE
PACKAGE
JACKET
ROCKER
ROCKET
HAMMER
TRAIN
ROOSTER
BEEHIVE
LADDER
ARMCHAIR
ROSE
APPENDIX
Target
ACORN
BAGEL
BASKET
BATTER
BEAKER
BEAVER
BEE
BELL
BOWL
BEACH
BED
BONE
CHAIN
HAT
KEY
POT
VAN
SHIRT
LEMON
BUCKLE
BUCKET
ENGINE
BUG
BUS
TELEPHONE
TENT
BUTTER
BUTTON
SQUIRREL
RIFLE
CABBAGE
CABIN
CAMEL
CANDLE
CAMERA
CANDY
MEDAL
MUFFIN
PILLOW
BLANKET
TURKEY
HANGER
SHUTTER
WHISTLE
WINDOW
CANNON
BACON
222
Journal of Cognitive Neuroscience
Volume 22, Number 2
Acknowledgments
This research was supported in part by NIH Grant RO1 DC006220
from the National Institute on Deafness and Other Communi-
cation Disorders. The content is solely the responsibility of the
authors and does not necessarily represent the official views of
the National Institute on Deafness and Other Communication
Disorders or the National Institutes of Health. We thank Emily
Myers for help with the analysis of the data, Kathleen Kurowski
for help in the preparation of the auditory stimuli, and Eiling Yee
and Bob McMurray for providing competitor pair candidates.
Reprint requests should be sent to Sheila E. Blumstein, Depart-
ment of Cognitive and Linguistic Sciences, Brown University, Pro-
vidence, RI 02912, or via e-mail: Sheila_Blumstein@brown.edu.
REFERENCES
Allopenna, P. D., Magnuson, J. S., & Tanenhaus, M. K. (1998).
Tracking the time course of spoken word recognition using
eye movements: Evidence for continuous mapping models.
Journal of Memory and Language, 38, 419–439.
Altman, G. T. M., & Kamide, Y. (2004). Now you see it, now
you donʼt: Meditating the mapping between language and
the visual world. In J. Henderson & F. Ferreira (Eds.), The
interface of language, vision, and action (pp. 347–386).
New York: Psychology Press.
Badre, D., & Wagner, A. W. (2004). Selection, integration,
and conflict monitoring: Assessing the nature and generality
of prefrontal cognitive control mechanisms. Neuron, 41,
473–487.
Badre, D., & Wagner, A. W. (2007). Left ventrolateral
prefrontal cortex and the cognitive control of memory.
Neuropsychologia, 45, 2883–2901.
Binder, J. R., & Price, C. J. (2001). Functional neuroimaging
of language. In R. Cabeza & A. Kingstone (Eds.), Handbook
of functional neuroimaging (pp. 187–251). Cambridge,
MA: MIT Press.
Buckner, R. L., Raichle, M. E., & Petersen, S. E. (1995).
Dissociation of human prefrontal cortical areas across
different speech production tasks and gender groups.
Journal of Neurophysiology, 74, 2163–2173.
Burton, M. W. (2001). The role of inferior frontal cortex in
phonological processing. Cognitive Science, 25, 695–709.
Burton, M. W., Small, S. L., & Blumstein, S. E. (2000). The
role of segmentation in phonological processing: An fMRI
investigation. Journal of Cognitive Neuroscience, 12,
679–690.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers
and Biomedical Research, 29, 162–173.
Cox, R. W., & Hyde, J. S. (1997). Software tools for analysis and
visualization of fMRI data. NMR in Biomedicine, 10, 171–178.
Dahan, D., Magnuson, J. S., & Tanenhaus, M. K. (2001). Time
course of frequency effects in spoken-word recognition:
Evidence from eye movements. Cognitive Psychology, 42,
317–367.
Dahan, D., Magnuson, J. S., Tanenhaus, M. K., & Hogan, E. M.
(2001). Subcategorical mismatches and the time course of
lexical access: Evidence for lexical competition. Language
and Cognitive Processes, 16, 507–534.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of
selective visual attention. Annual Review of Neuroscience,
18, 193–222.
Duncan, J. (1998). Converging levels of analysis in the
cognitive neuroscience of visual attention. Philosophical
Transcripts of the Royal Society of London, Series B, 353,
1307–1317.
Duncan, J. (2001). An adaptive model of neural function in
prefrontal cortex. Nature Reviews Neuroscience, 2, 820–829.
Duncan, J., & Owen, A. M. (2000). Common regions of the
human frontal lobe recruited by diverse cognitive demands.
Trends in Neurosciences, 23, 475–483.
Eickhoff, S. B., Heim, S., Zilles, K., & Amunts, K. (2006). Testing
anatomically specified hypotheses in functional imaging
using cytoarchitectonic maps. Neuroimage, 32, 570–582.
Eickhoff, S. B., Paus, T., Caspers, S., Grosbras, M. H., Evans,
A. C., Zilles, K., et al. (2007). Assignment of functional
activations to probabilistic cytoarchitectonic areas revisited.
Neuroimage, 26, 511–521.
Eickhoff, S. B., Stephan, K. E., Mohlberg, H., Grefkes, C.,
Fink, G. R., Amunts, K., et al. (2005). A new SPM toolbox
for combining probabilistic cytoarchitectonic maps and
functional imaging data. Neuroimage, 25, 1325–1335.
Fiez, J. A. (1997). Phonology, semantics, and the role of the
left inferior prefrontal cortex. Human Brain Mapping, 5,
79–83.
Gelfand, J. R., & Bookheimer, S. Y. (2003). Dissociating
neural mechanisms of temporal sequencing and processing
phonemes. Neuron, 38, 831–842.
Gold, B. T., & Buckner, R. L. (2002). Common prefrontal
regions coactivate with dissociable posterior regions
during controlled semantic and phonological tasks.
Neuron, 35, 803–812.
Hallet, P. E. (1986). Eye movements. In K. Boff, L. Kaufman,
& J. Thomas (Eds.), Handbook of perception and human
performance (pp. 10-1–10-112). New York: Wiley.
Hickok, G., & Poeppel, D. (2000). Towards a functional
neuroanatomy of speech perception. Trends in Cognitive
Sciences, 4, 131–138.
Luce, P. A., & Pisoni, D. B. (1998). Recognizing spoken words:
The neighborhood activation model. Ear and Hearing,
19, 1–36.
Marslen-Wilson, W. (1987). Functional parallelism in spoken
word recognition. Cognition, 25, 71–102.
Marslen-Wilson, W., & Welsh, A. (1978). Processing interactions
during word-recognition in continuous speech. Cognitive
Psychology, 10, 29–63.
McClelland, J. L., & Elman, J. L. (1986). The TRACE model
of speech perception. Cognitive Psychology, 18, 1–86.
Mertus, J. A. (2002). BLISS: The Brown Lab interactive
speech system. Providence, RI: Brown University.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory of
prefrontal cortex function. Annual Review of Neuroscience,
24, 167–202.
Norris, D. (1994). Shortlist: A connectionist model of
continuous speech recognition. Cognition, 52, 189–234.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh Inventory. Neuropsychologia,
9, 97–113.
Paulesu, E., Frith, C. D., & Frackowiak, R. S. J. (1993). The
neural correlates of the verbal component of working
memory. Nature, 362, 342–345.
Poldrack, R. A., Wagner, A. D., Prull, M. W., Desmond, J. E.,
Glover, G. H., & Gabrielli, J. D. (1999). Functional
specialization for semantic and phonological processing in
the left inferior prefrontal cortex. Neuroimage, 10, 15–35.
Prabhakaran, R., Blumstein, S. E., Myers, E. B., Hutchinson, E.,
& Britton, B. (2006). An event-related investigation of
phonological–lexical competition. Neuropsychologia, 44,
2209–2221.
Price, C., Moore, C. J., Humphreys, G. W., & Wise, R. J. S. (1997).
Segregating semantic from phonological processes during
reading. Journal of Cognitive Neuroscience, 9, 727–733.
Smith, E. E., & Jonides, J. (1999). Storage and executive
processes in the frontal lobes. Science, 283, 1657–1661.
Righi et al.
223
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
2
e
-
2
p
1
d
3
f
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
c
3
n
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
2
0
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
/
f
t
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Snyder, H. R., Feignson, K., & Thompson-Schill, S. L. (2007).
Prefrontal cortical response to conflict during semantic
and phonological tasks. Journal of Cognitive Neuroscience,
19, 761–775.
Spivey-Knowlton, M. K. (1996). Integration of visual
and linguistic information: Human data and model
simulations. Unpublished Ph.D. thesis, University of
Rochester, Rochester, NY.
Tanenhaus, M. K., Spivey-Knowlton, M. J., Eberhard, K. M.,
& Sedivy, J. C. (1995). Integration of visual and linguistic
information in spoken language comprehension. Science,
268, 1632–1634.
Thompson-Schill, S. L., DʼEsposito, M., Aguirre, G. K., &
Farah, M. J. (1997). Role of the inferior prefrontal cortex
in retrieval of semantic knowledge: A reevaluation.
Proceedings of the National Academy of Sciences, U.S.A.,
94, 14792–14797.
Thompson-Schill, S. L., DʼEsposito, M., & Kan, I. P. (1999).
Effects of repetition and competition on activity in left
prefrontal cortex during word generation. Neuron, 23,
513–522.
Thompson-Schill, S. L., Swick, D., Farah, M. J., DʼEsposito, M.,
Kan, I. P., & Knight, R. T. (1998). Verb generation in
patients with focal frontal lesions: A neuropsychological
test of neuroimaging findings. Proceedings of the National
Academy of Sciences, U.S.A., 95, 15855–15860.
Talairach, J., & Tournoux, P. (1988). A co-planar stereotaxic
atlas of a human brain. Stuttgart, Germany: Thieme.
Yee, E., Blumstein, S., & Sedivy, J. C. (2008). Lexical–semantic
activation in Brocaʼs and Wernickeʼs aphasia: Evidence
from eye movements. Journal of Cognitive Neuroscience,
20, 1–21.
Yee, E., & Sedivy, J. (2006). Eye movements reveal transient
semantic activation during spoken word recognition.
Journal of Experimental Psychology: Learning, Memory,
and Cognition, 32, 1–14.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
2
l
2
e
-
2
p
1
d
3
f
1
/
9
2
3
2
8
/
5
2
3
/
2
2
o
1
c
3
n
/
1
2
0
7
0
6
9
9
1
2
5
1
2
2
0
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
2
0
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
t
/
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
224
Journal of Cognitive Neuroscience
Volume 22, Number 2