Motor System Interactions in the Beta Band Decrease
during Loss of Consciousness
Nicole C. Swann1, Coralie de Hemptinne1, Ryan B. Maher2, Catherine A. Stapleton3,
Lingzhong Meng1, Adrian W. Gelb1, and Philip A. Starr1
Astratto
■ Communication between brain areas and how they are influ-
enced by changes in consciousness are not fully understood.
One hypothesis is that brain areas communicate via oscillatory
processes, utilizing network-specific frequency bands, that can
be measured with metrics that reflect between-region inter-
actions, such as coherence and phase amplitude coupling
(PAC). To evaluate this hypothesis and understand how these
interactions are modulated by state changes, we analyzed electro-
physiological recordings in humans at different nodes of one
well-studied brain network: the basal ganglia–thalamocortical
loops of the motor system during loss of consciousness induced
by anesthesia. We recorded simultaneous electrocorticography
over primary motor cortex (M1) with local field potentials from
subcortical motor regions (either basal ganglia or thalamus) In
15 movement disorder patients during anesthesia (propofol) induc-
tion as a part of their surgery for deep brain stimulation. Noi
observed reduced coherence and PAC between M1 and the sub-
cortical nuclei, which was specific to the beta band (∼18–24 Hz).
The fact that this pattern occurs selectively in beta underscores
the importance of this frequency band in the motor system and sup-
ports the idea that oscillatory interactions at specific frequencies are
related to the capacity for normal brain function and behavior. ■
INTRODUCTION
How do functionally related brain areas communicate
over long distances? One hypothesis is based on oscilla-
tory synchronization in specific frequency bands, usually
lower than ∼50 Hz (Siegel, Donner, & Engel, 2012).
Coherence is a simple measure of oscillatory synchroni-
zation that may support long-distance neural interactions.
It reflects interactions between brain areas related to the
consistency of the phases and amplitudes of their neu-
ronal signals (Fries, 2005). D'altra parte, higher-
frequency, broadband activity (“broadband gamma”,
70–250 Hz) seems to reflect local neural activity (Equipaggio,
Jacobs, Fried, & Kahana, 2009; Miller et al., 2007). Canolty
and colleagues demonstrated that, in some instances,
broadband gamma amplitude is modulated by the phase
of lower-frequency oscillatory rhythms (Canolty et al.,
2006). This phase amplitude coupling (PAC) provides a
mechanism that could explain how low-frequency oscil-
latory changes, which are well suited to coordinate activity
over long distances, can influence local neuronal activity
(Canolty & Knight, 2010; Fries, 2005). When PAC is calcu-
lated from the phase and amplitude of signals recorded
from two different brain areas, it may also reflect inter-
actions between brain regions.
1University of California San Francisco, 2Fidere Anesthesia
Consultants, Mountain View, CA, 3Alta Bates Summit Medical
Center, Berkeley, CA
© 2015 Istituto di Tecnologia del Massachussetts
Transitions in consciousness afford an opportunity to
evaluate how these basic measurements of brain inter-
actions change in the context of profound behavior/state
i cambiamenti. Although the underlying causal mechanisms of
consciousness remain unclear, there is converging evi-
dence that transitions of consciousness are characterized
by a change of network dynamics wherein local brain net-
works become more isolated from one another, disrupt-
ing the brain’s ability to integrate information (Lewis
et al., 2012; Alkire, Hudetz, & Tononi, 2008). Because
all but the most basic forms of behavior are only possible
in the conscious state, we hypothesized that the most
functionally important network interactions are only pres-
ent in the conscious state. To better understand how
those interactions might mediate behavior and, perhaps,
relate to communication in brain networks, we sought to
study how simple metrics of brain interactions (coherence
and PAC) in the motor system change during anesthesia-
induced transitions in consciousness.
To address this goal, we combined electrocortico-
graphy (ECoG) and subcortical local field potential
(LFP) recordings in movement disorder patients under-
going surgical implantation of deep brain stimulation
(DBS) leads (de Hemptinne et al., 2013, 2015). Questo
approach provides a unique opportunity to acquire field
potential data from functionally related cortical and sub-
cortical structures (primary motor cortex [M1] and motor
territories of the basal ganglia and thalamus). Because
DBS targets vary across individuals/diagnoses, we have
Journal of Cognitive Neuroscience 28:1, pag. 84–95
doi:10.1162/jocn_a_00884
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
e
j
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
8
8
/
4
1
1
/
9
8
5
4
0
/
2
1
6
7
1
8
o
4
C
5
N
3
_
0
UN
/
_
j
0
o
0
C
8
N
8
_
4
UN
P
_
D
0
0
B
8
sì
8
G
4
tu
.
e
P
S
D
T
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
T
/
.
F
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
the opportunity to sample from a number of different sub-
cortical nodes in the basal ganglia–thalamocortical (BGTC)
loop (although this does increase variability in our sample).
The BGTC circuit is particularly well suited to address ques-
tions concerning network interactions, because structural
connectivity in the motor loop is relatively well defined,
with connections from cortex to basal ganglia to thalamus
and then back to cortex (Alexander, DeLong, & Strick,
1986). Additionally, the electrophysiological signatures in
the motor system have been studied extensively. The beta
frequency range (13–30 Hz) dominates and is dynamically
modulated during movement (Brovelli et al., 2004; Kuhn
et al., 2004; Cassidy et al., 2002; Crone, Miglioretti, Gordon,
Sieracki, et al., 1998; Sanes & Donoghue, 1993; Murthy &
Fetz, 1992; Pfurtscheller, 1981). We hypothesized that loss
of consciousness would be characterized by reduced inter-
actions between cortical and subcortical regions, measured
with coherence and inter-region PAC, and that this would
be particularly prominent in the beta frequency band.
METHODS
Patients
Fifteen patients (4 women/11 men, average age = 62 years)
were recruited from the Surgical Movement Disorders
Clinic at the University of California, San Francisco. Patients
were scheduled to undergo surgery to implant a perma-
nent subcortical DBS lead to treat their movement dis-
orders. All patients were simultaneously participating in
a study of the contribution of cortical oscillatory activity
to movement disorders pathophysiology, using ECoG
from a subdural strip array inserted through the standard
frontal burr hole used for DBS insertion, and temporarily
placed over the primary motor cortex (de Hemptinne
et al., 2013; Shimamoto et al., 2013; Crowell et al.,
2012). Our standard surgical procedure requires that
patients be awake for microelectrode mapping and DBS
lead insertion, followed by anesthesia induction (for
wound closure). This procedure provides the opportunity
for brain recording during anesthesia-induced loss of con-
sciousness, avoiding the need to administer anesthetics
solely for research purposes.
The goal of this study was to examine interactions be-
tween nodes of the motor system during loss of con-
sciousness, rather than focusing on a specific movement
disorder. Therefore, we included patients with varying
diagnoses and varying subcortical targets within the
motor network to enable the identification of network
motifs common to changes in consciousness independent
of disease state. Twelve of the patients had Parkinson
disease (PD), two had essential tremor (ET), and one
had primary dystonia. Four PD patients had DBS leads
placed into the globus pallidus interna (GPi), whereas
the other eight had subthalamic nucleus (STN) leads,
with target choice dictated by clinical criteria (Follett
et al., 2010). The DBS target for ET patients was the ventro-
lateral thalamus. The target for the dystonia patient was
GPi. The study methodologies were approved by the insti-
tutional ethics committee and are in agreement with the
Declaration of Helsinki. All patients provided written
informed consent to participate in the study, and the use
of a temporary cortical ECoG array placed for research
purposes was an explicit part of the consent discussion.
ECoG Strip Placement
ECoG was recorded from one hemisphere during the
surgery. For patients receiving unilateral DBS, the ECoG
was placed ipsilateral to the side of the DBS. For patients
receiving bilateral surgery, the side of the ECoG was
determined based on the clearest anatomic demarcation
of the central sulcus on preoperative MRI. Eleven patients
had left side recordings, whereas four had right.
The data from the dystonia patient and five PD patients
were collected with a six-electrode ECoG strip (1 cm spac-
ing between electrodes; see Figure 1A), and the other
seven PD patients and both ET patients were recorded
with a 28-electrode ECoG strip (4 mm spacing between
electrodes; see Figure 1B). The type of ECoG strip used
depended on the parent study of movement disorders
physiology, because during this study period, we transi-
tioned from the lower spatial resolution recording to a
higher-resolution technique. Each strip was placed with
at least one electrode covering M1. The intended target
location for the center of the ECoG strip was the arm area
of M1, 3 cm from the midline and slightly medial to the hand
knob ( Yousry et al., 1997). Adequate localization of the
ECoG strip was confirmed using either preoperative MRI
merged to an intraoperative CT (13 participants) or using
intraoperative lateral fluoroscopy with a radio-opaque
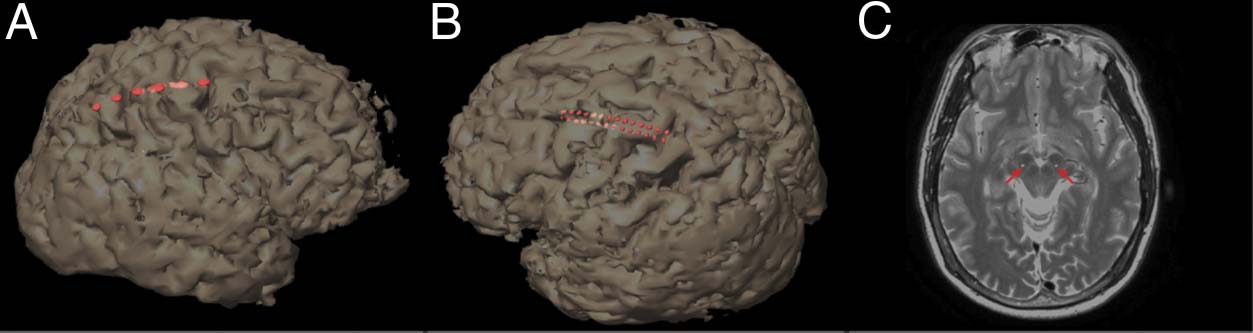
Figura 1. 3D brain
reconstructions of each
participant’s MRI with the
low-resolution (UN) E
high-resolution (B) ECoG strips
and the subcortical DBS electrode,
positioned in STN (indicated
with red arrows) (C). Electrode
locations (shown in red) are
visualized by merging an
intraoperative CT with a
preoperative MRI.
Swann et al.
85
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
e
j
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
8
8
/
4
1
1
/
9
8
5
4
0
/
2
1
6
7
1
8
o
4
C
5
N
3
_
0
UN
/
_
j
0
o
0
C
8
N
8
_
4
UN
P
_
D
0
0
B
8
sì
8
G
4
tu
.
e
P
S
D
T
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
F
T
.
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
marker placed on the skin indicating the intended target
in M1 that was visible relative to the electrode strip (2 par-
ticipants), as previously described (de Hemptinne et al.,
2013, 2015; Shimamoto et al., 2013; Crowell et al., 2012).
In all participants, functional localization was also exam-
ined using somatosensory evoked potentials (SSEPs, fre-
quency = 2 Hz, pulse width = 200 μs, pulse train length =
160, amplitude = 25–40 mAmp), as has been reported
(de Hemptinne et al., 2013; Shimamoto et al., 2013; Crowell
et al., 2012). Note that for the 28-electrode strip, there
were two rows of 14 electrodes (see Figure 1B), so it
was expected that two electrodes localized immediately
anterior to the central sulcus would show a phase reversal
relative to the postcentral sulcus electrodes.
Tavolo 1. MOAA/S Responsiveness Scale
State
Responsiveness
5
4
3
2
1
0
Responds readily to name spoken in normal tone
Lethargic response to name spoken in normal tone
Responds only after name is called loudly
and/or repeatedly
Responds only after mild prodding or shaking
Responds only after painful trapezius squeeze
No response after painful trapezius squeeze
Rating criteria for each state.
DBS Electrode Implantation
ECoG and LFP Recordings
Anatomic targeting of the desired subcortical structure
was performed as previously described (Starr et al.,
2002, 2006; Papavassiliou et al., 2004). For the GPi and
STN targets, the proper location was verified by elic-
iting movement-related single-cell discharge patterns
(Starr et al., 2002). For all regions, the correct placement
was verified by test stimulation, as well as intraoperative
fluoroscopy (all patients), intraoperative CT (13 patients;
Shahlaie, Larson, & Starr, 2011), and postoperative MRI
(all patients).
Consciousness Assessment and
Anesthesia Parameters
Continuous recordings were taken after DBS placement,
but prior to burr hole closure and DBS pulse generator
placement, while the patients were slowly anesthetized
with propofol. Data from PD patients were recorded
after at least 12 hr off antiparkinsonian medications.
Consciousness was assessed every 3 min using the Mod-
ified Observers Assessment of Alertness/Sedation Scale
(MOAA/S; Vedi la tabella 1; Chernik et al., 1990). Ratings were
recorded by an anesthesiologist (AG, LM). When the
clock started for the assessment session (time zero), UN
button was pressed, which generated a voltage deflection
in an auxiliary channel digitized with the electrophysiology
dati. This synchronized the times noted by the anesthe-
siologist with the electrophysiology data.
Anesthesia induction with propofol was performed
slowly with the goal of achieving an estimated plasma
level (Marsh Model) Di 4 mcg/ml at approximately 15 min
(Smith et al., 1994). Induction ended when all patients no
longer responded to painful stimulation according to
MOAA/S scale (Vedi la tabella 1). Propofol sedation had also
been used during the drilling of the burr holes prior to
intracranial recording but was stopped for subcortical
mapping and DBS lead testing, almeno 60 min prior to
initiating the recordings analyzed here. This is ample time
for the effects of propofol to wear off (Raz, Eimerl, Zaidel,
Bergman, & Israel, 2010; Fechner et al., 2004).
ECoG recordings were performed using either the Alpha
Omega Microguide Pro, Alpha Omega, Inc. (Alpharetta, GA;
for the six-electrode strip; Figure 1A) or the Tucker Davis
Technologies Recording System (for the 28 electrode
strip; Figure 1B). For the Alpha Omega system, dati
were sampled at 3000 Hz. Data from each of the five
more posterior electrodes were referenced to the most
anterior electrode. A needle electrode in the scalp served
as the ground. Data were band-pass filtered at 1–500 Hz.
For the Tucker Davis System, data were sampled at
3051 Hz. All electrodes were referenced to a scalp needle
electrode that also served as the ground. Data were low-
pass filtered at 1500 Hz. Because there was no high-pass
filter applied during data acquisition, the mean of each
electrode was subtracted during offline preprocessing
to detrend the data. See Figure 2A for an example of the
raw ECoG data.
LFP recordings were from the subcortical target ipsi-
lateral to ECoG placement were recorded in a bipolar
configuration from the middle two cylindrical contacts
(1.5 mm height, 1.2 mm diameter) of a quadripolar lead
(Medtronic model 3389, 0.5 mm between contacts, eight
patients or Medtronic model 3387, 1.5 mm between con-
tacts, seven patients), with Electrode 1 as the active and
Electrode 2 as the reference. The guide tube for the DBS
lead served as ground. For the six patients for whom the
six-electrode cortical strip was used, the LFP recordings
were done with the Alpha Omega system using the same
Alpha Omega parameters described above for ECoG. For
all the PD patients who were tested with the 28-electrode
strip, LFP data were still collected using the hardware
from the Alpha Omega system (which was necessarily
present as it is FDA-approved for clinical microelectrode
recordings for mapping); Tuttavia, the analog signal was
streamed to the Tucker Davis system and digitized there
at 25,000 Hz sampling rate and then subsequently down-
sampled. For the sessions with ET patients, the Alpha
Omega system was not present in the room, as micro-
electrode recordings were not used for ET. Così, LFPs were
recorded in the same manner as the ECoG electrodes (dati
86
Journal of Cognitive Neuroscience
Volume 28, Numero 1
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
e
j
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
8
8
/
4
1
1
/
9
8
5
4
0
/
2
1
6
7
1
8
o
4
C
5
N
3
_
0
UN
/
_
j
0
o
0
C
8
N
8
_
4
UN
P
_
D
0
0
B
8
sì
8
G
4
tu
.
e
P
S
D
T
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
/
T
.
F
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
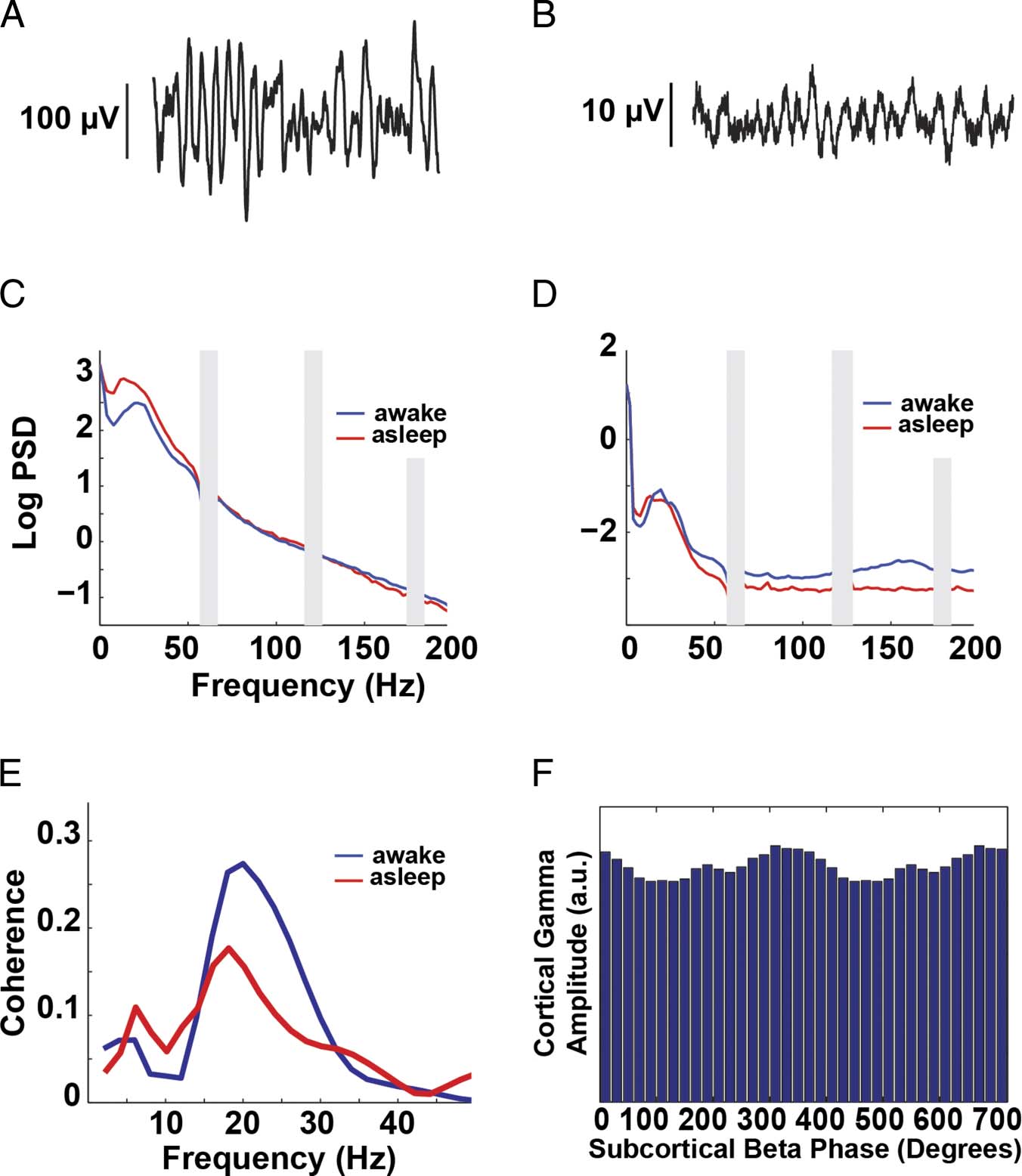
Figura 2. Example data
from individual participant 6.
One second of raw M1 ECoG
(UN) data and raw LFP data
(from STN) (B). Log PSD
from M1 (C) and STN (D).
The gray bars indicate 60-Hz
noise and the harmonics.
(E) Coherence between
M1 and STN. (F) Histogram
of mean M1 broadband
gamma amplitude (70–150 Hz)
binned by subcortical beta
phase (18–20 Hz). Here,
modulation is apparent.
A flat distribution would
suggest no PAC.
from Electrodes 1 E 2 of the DBS electrode were both re-
corded with the needle electrode as the reference and
ground) and then subsequently referenced in a bipolar con-
figuration offline. See Figure 2B for an example of the raw
LFP data.
Electrophysiological Preprocessing
All data were initially down-sampled to 1000 Hz, and line
frequency noise (at 60 Hz, 120 Hz, E 180 Hz) was re-
moved using a third-order Butterworth notch filter,
which spanned a 4-Hz range centered at each frequency
(cioè., 58–62 Hz for 60 Hz). Data were then re-referenced.
For the six-electrode strip, a bipolar montage was used
such that each electrode was referenced to the anterior
electrode (C1–C2, C2–C3, eccetera.). For the 28-electrode
strip a common average reference was used including
all ECoG electrodes (excluding any with obvious, con-
tinuous noise). Each reference strategy is best suited
for the given electrode configuration: An average refer-
ence montage is not appropriate when there are only
five active electrodes to contribute to the reference, E
a bipolar reference is less optimal for the 28 electrode
strip with small contacts and very close spacing. To deter-
mine if the different referencing schemes influenced our
risultati, we re-ran our analyses using an average reference
scheme for all participants, and the results were similar;
così, this difference in analysis is unlikely to have strongly
influenced our findings. Subcortical LFP electrodes were
processed in the same way as above (down-sampled and
filtered). All raw data were manually inspected and
periods of artifact (per esempio., electrical artifact caused by medical
equipment or movement) were noted and excluded from
analysis.
During data recording, the anesthesiologist performed
ratings every 3 min to assess the patient’s level of con-
sciousness (Vedi la tabella 1). Data were assigned a conscious-
ness rating according to the assessment at the end of
each 3-min time segment. For instance, if after 3 min
the anesthesiologist rated the patient’s alertness as a
5 and at 6 min rated it as a 4, the segment of the electro-
physiology data corresponding to 0–3 min would be
marked as “State 5” and times 3–6 min would be marked
as “State 4.” Not all patients were observed at each state.
Swann et al.
87
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
e
j
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
8
8
/
4
1
1
/
9
8
5
4
0
/
2
1
6
7
1
8
o
4
C
5
N
3
_
0
UN
/
_
j
0
o
0
C
8
N
8
_
4
UN
P
_
D
0
0
B
8
sì
8
G
4
tu
.
e
P
S
D
T
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
/
.
T
F
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
The longest artifact-free period of each state was used
for analysis. On average, this was 218.1 sec and was
always greater than 31 sec. Because both coherence and
PAC can vary over time even without an overt change in
behavioral state, we included the longest data segments
possible for each participant and each level of conscious-
ness. Tuttavia, because coherence and PAC values can
also be influenced by differences in the length of data
analyzed, we conducted a control analysis to verify that
differences in data length were not systematically driving
our observed differences. In this analysis, for each par-
ticipant, data of the same length from each state were
used for comparison.
For each patient, the cortical electrode(S) that most
closely corresponded to M1 were selected based on
both the anatomical localization and SSEP waveforms.
This was done prior to examining the data from the
anesthesia induction file. For the 28-electrode strip, there
were two rows of 14 electrodes, so we selected one
M1 electrode from each row that met these same criteria.
In questo caso, power, coherence, and PAC (described be-
low) were calculated separately for each of these elec-
trodes and then averaged for each electrode pair. For
the six-electrode strip, the electrode selection procedure
described above typically led to the selection of one elec-
trode, which clearly lay over M1 (at the border of the cen-
tral sulcus and the precentral gyrus), that also showed a
clear SSEP phase reversal. For the 28-electrode strip,
where the electrodes were smaller and spaced closer to-
gether, definitive selection of the two optimal M1 elec-
trodes was less clear (regardless of reference scheme
used). To test whether ambiguous selection of the M1
electrode may have influenced our results, we conducted
a test wherein all electrodes on each strip were included in
the analysis. (In questo caso, signal processing calculations
described below were calculated for each electrode
separately, and then the final results were averaged,
such that each participant contributed only one data point
to the group statistics.) Although this method clearly
provides less spatial specificity, results were similar even
when all electrodes were included. Così, ambiguity in
the selection of the optimal M1 electrode did not distort
our results. The fact that results were similar for the M1
electrode, and the entire strip, most likely reflects the fact
that the strip was placed to span precentral and postcentral
gyri, which generate similar oscillatory signatures (Crone,
Miglioretti, Gordon, Sieracki, et al., 1998).
Electrophysiological Signal Processing
All analyses used a combination of custom Matlab scripts
and EEGLAB functions (Delorme & Makeig, 2004). Noi
implemented two main types of analyses. Primo, we ana-
lyzed the activity of each area (M1 and the subcortical
region) separately. We calculated power spectral density
(PSD) and PAC (Tort et al., 2008; Canolty et al., 2006)
within each region. Analyses for each patient, state, E
region (M1 and subcortical) were calculated separately
in Matlab.
PSD analysis used the Welch method ( pwelch function
in Matlab with a 512-msec window, 256 msec of overlap;
see Figure 2C and D for an example of PSD in one par-
ticipant). Statistics were then calculated based on the log
PSD values. Tuttavia, results were similar if non-log-
transformed values were used.
PAC was calculated using the Kullback–Leibler-based
modulation index method, which has been previously
described (de Hemptinne et al., 2013; Tort et al., 2008).
In brief, the M1 ECoG and subcortical LFP signal were
filtered separately using a two-way FIR1 filter (eegfilt with
‘fir1’ parameters). Low-frequency signals were filtered
individually at frequencies ranging from 2 A 50 Hz, con
a 2-Hz bandwidth, and the phase was extracted from this
signal using a Hilbert transform. Allo stesso modo, the amplitude
of the high-frequency broadband gamma signal was
extracted by taking the Hilbert transform of the band-
pass filtered data (70–150 Hz). Then the distribution of
the instantaneous amplitude envelope was computed
for every 20° interval of the instantaneous phase (Vedere
Figure 2F). The coupling (modulation index) between
the phase of each low-frequency rhythm and the high-
frequency amplitude was then determined by computing
the entropy values of this distribution and normalizing
by the maximum entropy value (Tort et al., 2008).
Secondo, we analyzed subcortical–cortical interactions by
calculating coherence and cross-structure PAC. Analyses
were performed for each participant and state separately,
using signals from both the M1 ECoG and the subcortical
LFP. We calculated coherence for frequencies ranging
from 2 A 50 Hz, with a 2-Hz bandwidth, by filtering both
the M1 ECoG and subcortical LFP using the same two-way
FIR1 filter used to calculate PAC. Complex signals were
then obtained for each filtered signal by taking the Hilbert
transform. Coherence between the ECoG electrode(S)
and the subcortical LFP was calculated using the cor-
responding autospectra (Wxx and Wyy) and cross-spectra
(Wxy) of the filtered complex signals.
Coh fð Þ ¼
(cid:1)
(cid:1)
(cid:1)
(cid:1)
Wxy fð Þ
√Wxx fð Þ√Wyy fð Þ
Here x and y refer to data from the two regions (cioè., cor-
tical and subcortical). Wxy was calculated by taking the
sum over time of the complex signal of x multiplied by
the conjugate of the complex signal of y. Wxx and Wyy
were calculated as the sum over time of the amplitude
of each (x and y) complex signal. See Figure 2E for an
example of coherence at all frequencies in one patient.
Between-region PAC was calculated in the same way as
within-region PAC described above, except that the low-
frequency phase component and the high-frequency
amplitude component were derived from signals from dif-
ferent brain regions (M1 ECoG and subcortical LFP). Noi
examined PAC using both the phase of the subcortical
88
Journal of Cognitive Neuroscience
Volume 28, Numero 1
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
e
j
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
8
8
/
4
1
1
/
9
8
5
4
0
/
2
1
6
7
1
8
o
4
C
5
N
3
_
0
UN
/
_
j
0
o
0
C
8
N
8
_
4
UN
P
_
D
0
0
B
8
sì
8
G
4
tu
.
e
P
S
D
T
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
/
T
F
.
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
LFP and amplitude of M1 ECoG and the opposite configu-
ration. See Figure 2F for an example of PAC in one patient.
Statistical Analysis of Data Grouped across Patients
For statistical comparisons of the change across patients
during anesthesia induction, a nonparametric paired
sign-rank test was computed comparing, for each patient,
the metric of interest (power, within region PAC, coher-
ence, or between region PAC) for data corresponding to
the state during which the patient was most awake (State
5, or closest to 5) to the data corresponding to the state
during which the patient was most asleep (State 0, O
closest to 0). This was done separately for each fre-
quency. Results were then corrected for multiple compar-
isons (for all frequencies examined) using a false discovery
rate (FDR) correction.
RESULTS
Propofol and Monitoring Results
Each patient reached an unconscious state (cioè., MOAA/S
value = 0 O 1). This was achieved on average in 22.5 min
from the start of the file recording (SD = 8 min). For
each patient, a minimum of three anesthetic states were
observed.
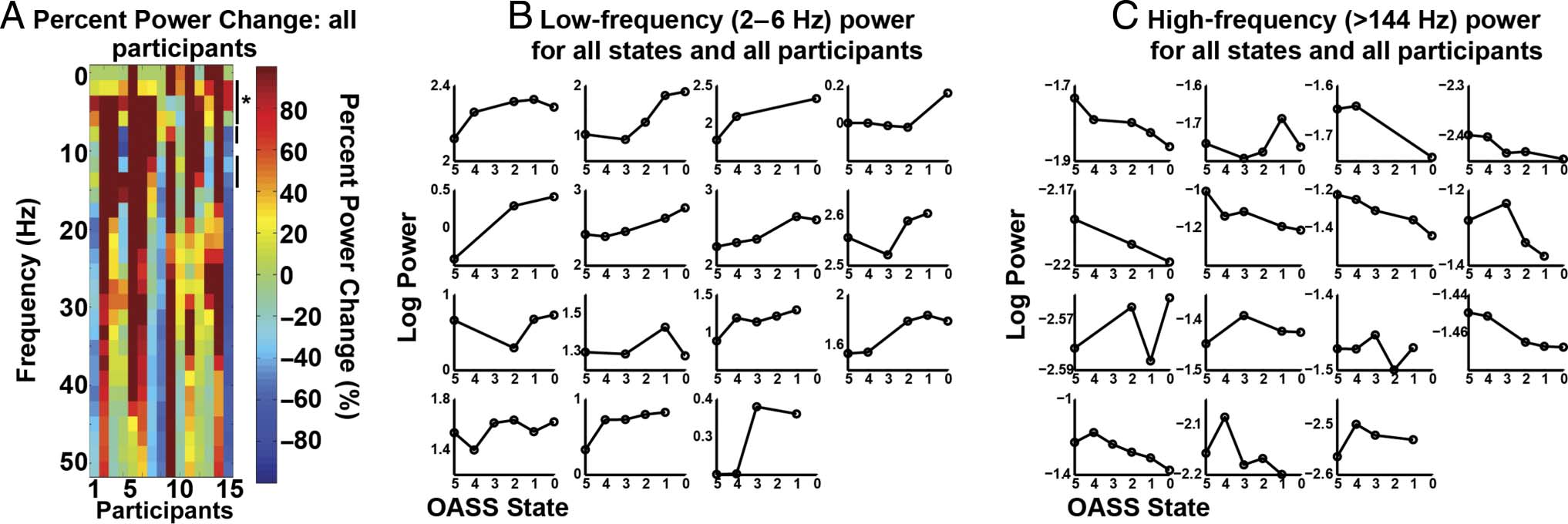
56 μV for the most awake state and 64 μV for the most
asleep state ( P < .04, with a paired, sign-rank test). The
subcortical LFP root mean square values were on average
8 μV (not significantly different for awake vs. asleep, p >
.15; Figura 2).
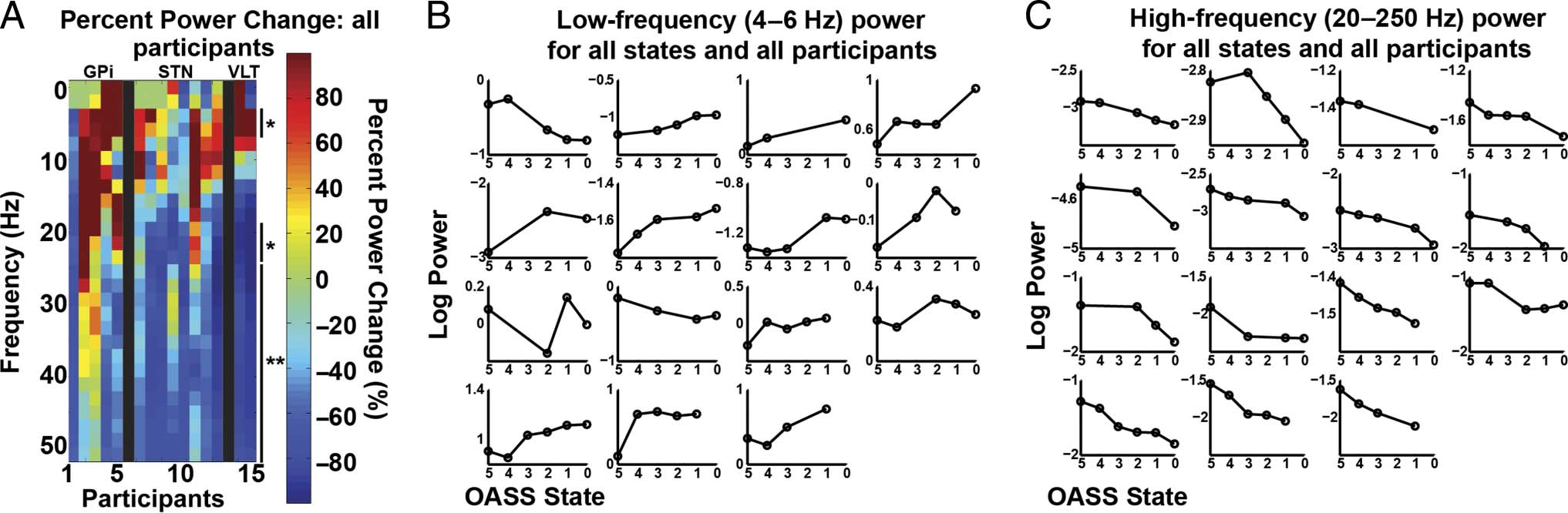
During induction, the subcortical LFP was charac-
terized by an increase in low-frequency power (4–6 Hz,
P < .05, FDR-corrected) and a decrease in higher-frequency
(broadband) activity (all frequencies 20–250 Hz, p <
.05, FDR-corrected; Figure 3). In the cortex, a similar
pattern was observed for the low-frequency power increase
(2–6 Hz, p < .05, FDR-corrected) and high-frequency
decreases (144–250 Hz, but not all frequencies in this range
were significant, p < .05). However, the decrease in high-
frequency power in the cortex was not significant when
correcting for multiple comparisons (Figure 4). Although
the cortical high-frequency effect was weak, previous
ECoG work has shown similar cortical changes in these
frequency ranges during propofol induction (Verdonck,
Reed, Hall, Gotman, & Plourde, 2014; Breshears et al.,
2010). Thus, although our result was not statistically
robust, it trends in the direction reported by others.
Of note, M1 beta power did not change with induction
of anesthesia. Although subcortical beta power did de-
crease during induction, this was a nonspecific effect,
because all frequencies above 20 Hz were reduced.
High-frequency Spectral Power Decreases and
Low-frequency Power Increases in the
Cortex and Subcortical Nuclei
The signal amplitudes were typical for ECoG and LFP
data, with an average root mean square value for M1 of
Coherence between Cortex and Subcortical
Nuclei Decreases during Induction
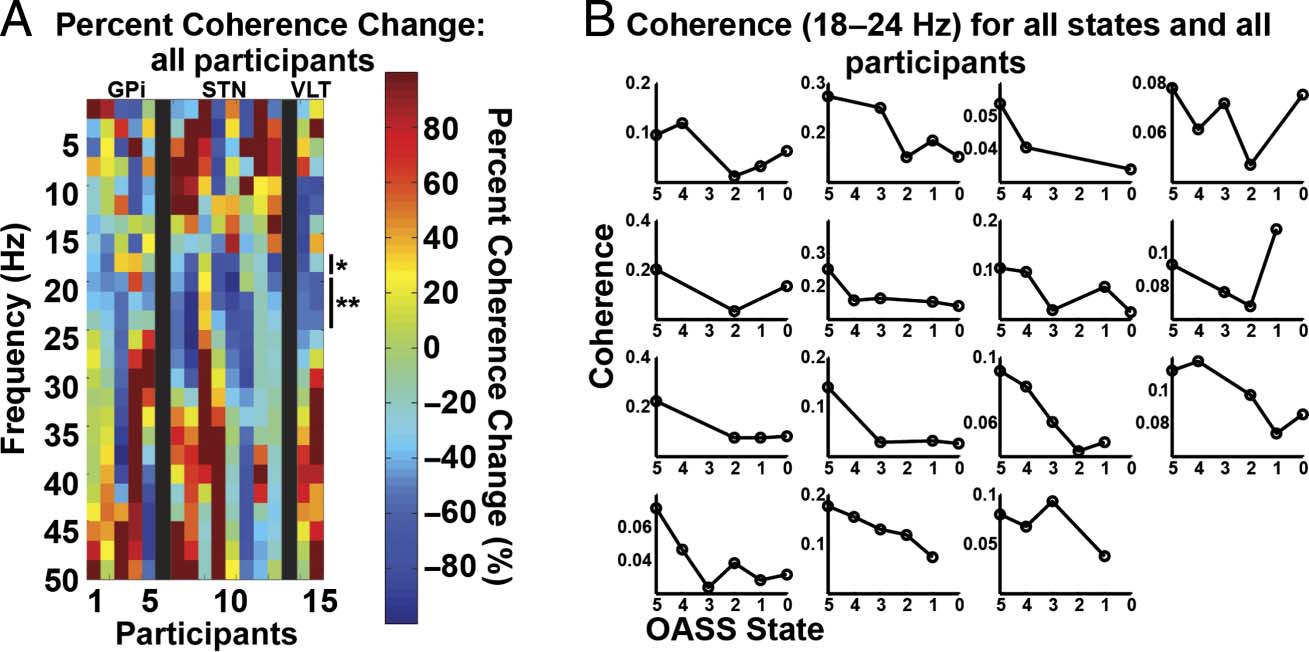
Across patients, there was a spectrally specific decrease in
coherence in the beta band (18–24 Hz) during propofol-
induced loss of consciousness ( p < .05, FDR-corrected;
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
8
8
/
4
1
1
/
9
8
5
4
0
/
2
1
6
7
1
8
o
4
c
5
n
3
_
0
a
/
_
j
0
o
0
c
8
n
8
_
4
a
p
_
d
0
0
b
8
y
8
g
4
u
.
e
p
s
d
t
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 3. Subcortical power changes. (A) Percent power change for all frequencies and participants. Percent change was calculated as: ((asleep −
awake) / awake) * 100. Significant differences are indicated with one ( p < .05, FDR-corrected) or two asterisks ( p < .01, FDR-corrected). Participants
are grouped by region of LFP recordings, indicated with labels (i.e., GPi, STN, and ventrolateral thalamus [VLT]) and separated by black bars.
Participant 1 was the dystonia patient, Participants 2–5 and 6–13 were PD patients, and Participants 14–15 were ET patients. To be consistent
with subsequent figures, only frequencies below 50 Hz are shown, although higher frequencies were analyzed and described in C. (B) Individual
participant log power for each state averaged across the frequencies with a significant increase in power (4–6 Hz). Participant order is the same
as in A, with Participants 1–4 making up the first row. (C) Same as B, but showing average log power for all frequencies with a significant decrease
during induction (20–250 Hz).
Swann et al.
89
Figure 4. M1 power changes. (A) Same as Figure 3A, except M1 power changes. To be consistent with subsequent figures, only frequencies
up to 50 Hz are shown, although higher frequencies were analyzed and described in C. The asterisk indicates significant frequencies ( p < .05,
FDR-corrected), and the line without an asterisk indicates significance at p < .05, uncorrected. Participants 1, 2, 3, 6, 7, and 8 were recorded with the
low-resolution strip. The others were recorded with the high-resolution strip. (B and C) Same as Figure 3B and C except showing M1 power changes for
frequencies with a significant increase (B, 2–6 Hz, p < .05, FDR-corrected) and (C) significant decrease (>144 Hz, P < .05, uncorrected).
Figure 5). This occurred in the setting of no significant
change in cortical beta power and a nonspecific decrease
in power in all frequencies above 20 Hz in the subcortical
LFP. The control analysis for which the same recording
durations were used for both the “awake” and “asleep”
epochs produced similar results (significant decrease in
coherence for 20–24 Hz, p < .05 FDR-corrected). Thus,
variability in data length is not driving the observed effect.
We also examined the change in the phase of the beta
coherence, and although some patients showed a
change, it was not consistent across patients (data not
shown). Coherence patterns at other frequencies were
variable across patients.
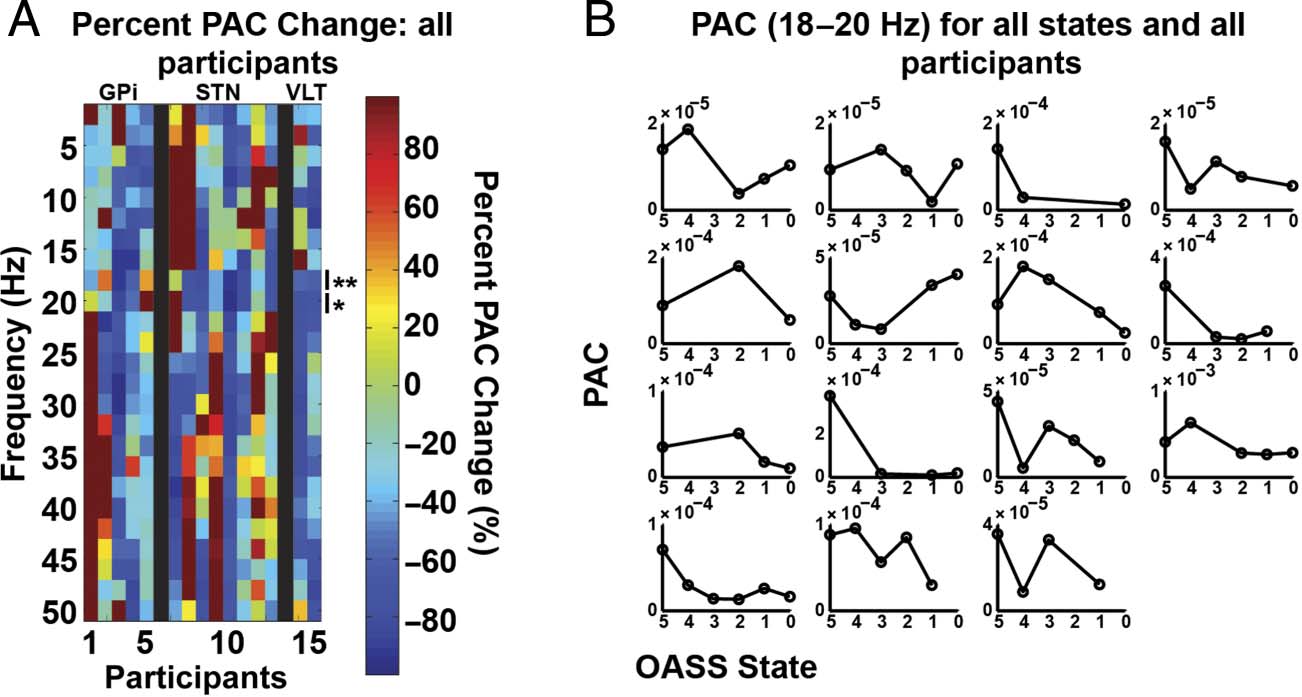
Phase Amplitude Coupling between Cortex and
Subcortical Nuclei Decreases during Induction
We calculated PAC in two ways. First, we tested the
within-region PAC for M1 and the subcortical target
separately and did not observe any significant effects
associated with anesthesia induction when correcting for
multiple comparisons.
We then calculated inter-region PAC taking the phase
from the subcortical target and the amplitude from the
M1 ECoG and vice versa. With phase from the subcorti-
cal target and amplitude from the M1 ECoG, there was a
significant decrease in PAC during induction, again in
the beta band (18–20 Hz, p < .05, FDR-corrected; see
Figure 6). The control analysis for which the same re-
cording durations were analyzed for both the “awake”
and “asleep” epochs produced similar, although slightly
weaker results (significant decrease in inter-region PAC
for 18–20 Hz, p < .05, uncorrected). Thus variability in
data length is unlikely to be driving the observed effect.
Results in other frequency ranges were variable across
participants. Calculating the PAC using the cortical phase
and subcortical amplitude revealed no significant results
associated with anesthesia induction.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
8
8
/
4
1
1
/
9
8
5
4
0
/
2
1
6
7
1
8
o
4
c
5
n
3
_
0
a
/
_
j
0
o
0
c
8
n
8
_
4
a
p
_
d
0
0
b
8
y
8
g
4
u
.
e
p
s
d
t
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
t
.
Figure 5. M1 and subcortical
coherence changes. (A) Same
as Figure 3A, except M1 and
subcortical coherence changes.
Significant differences are
indicated with one ( p < .05,
FDR-corrected) or two asterisks
( p < .01, FDR-corrected).
(B) Individual participant
coherence shown for each state
for 18–24 Hz. Participant order
is the same as in Figure 3B.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
90
Journal of Cognitive Neuroscience
Volume 28, Number 1
Figure 6. M1 and subcortical
PAC changes. PAC is calculated
using subcortical phase and
broadband M1 amplitude
(70–150 Hz). (A) Same as
Figure 3A, except PAC changes.
Significant differences are
indicated with one ( p < .05,
FDR-corrected) or two asterisks
( p < .01, FDR-corrected).
(B) Individual participant PAC
averaged between 18 and 20 Hz.
Participant order is the same
as in Figure 3B.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
e
j
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
1
2
8
8
/
4
1
1
/
9
8
5
4
0
/
2
1
6
7
1
8
o
4
c
5
n
3
_
0
a
/
_
j
0
o
0
c
8
n
8
_
4
a
p
_
d
0
0
b
8
y
8
g
4
u
.
e
p
s
d
t
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
DISCUSSION
To evaluate changes in interactions between connected
brain regions of the motor network associated with loss
of consciousness, we collected multisite field potential
recordings from structures in the motor network in 15 par-
ticipants undergoing DBS surgery and examined changes
in oscillatory activity. We found a decrease in interactions,
measured with coherence and inter-region PAC, between
brain regions in BGTC loops during loss of consciousness,
specific to the beta band. Importantly, the beta frequency
band was the only band to have a consistent change in
between-structure coherence and PAC across participants,
and this was not the case for cortical or subcortical spectral
power. We also observed a decrease in broadband gamma
activity in the subcortical nuclei and a similar, although
weaker, decrease in cortex. This was accompanied by an
increase in low-frequency power in both cortex and sub-
cortical nuclei.
Throughout the brain, cortical structures communicate
with functionally homologous subcortical modulators,
and this communication is thought to be mediated in
part by synchronization in oscillatory activity. The motor
system represents one such pair, with well-characterized
anatomic connections from cortical motor areas to basal
ganglia and thalamus (Alexander et al., 1986), and robust
oscillatory activity that is predominant in the alpha or
beta range (van Wijk, Beek, & Daffertshofer, 2012; Crone,
Miglioretti, Gordon, Sieracki, et al., 1998; Pfurtscheller,
1981). Our focus on the motor system was motivated
by the fact that it can be ethically studied using invasive
electrophysiological methods during DBS surgery. We
expect that the motor system is a reasonable model for
other brain networks and that some of the observed
patterns may be generally applicable, with network-
dependent variation in the particular frequency that is
most strongly influenced. Changes in coherence and
PAC, reflecting interactions between brain regions, have
been shown to vary with changes in behavior in a number
of different brain networks (Siegel et al., 2012; Canolty &
Knight, 2010). Furthermore, some of the cortical patterns
we observe in the motor system are similar to those ob-
served in another ECoG study, which examined more
widespread regions of cortex (Breshears et al., 2010).
However, because we focused specifically on the motor
system, we cannot be sure of the generalizability of our
results.
Support for the Importance of Synchronized
Beta Activity in the Motor System
Converging evidence suggests that one consequence of
loss of consciousness is functional disconnection be-
tween distributed brain regions (Sarasso et al., 2014; Lewis
et al., 2012; Alkire et al., 2008; Laureys, 2005; Engel &
Singer, 2001). For cortical and subcortical interactions, in
particular, this hypothesis is supported by functional imag-
ing studies of the recovery of awareness following trau-
matic unconsciousness (Laureys & Schiff, 2012; Goldfine
& Schiff, 2011) and by the finding that cortical–subcortical
structural connectivity correlates with levels of awareness
in patients with severe head trauma (Zheng, Reggente,
Lutkenhoff, Owen, & Monti, 2014; Fernandez-Espejo et al.,
2011). Our results support this hypothesis because two
measures that reflect interactions between brain regions,
coherence and cross-structure PAC, both decrease during
loss of consciousness. These measures are not fully inde-
pendent, because changes in coherence are likely to affect
cross-structure PAC.
Communication in different brain networks may occur
through synchronization at particular frequencies, with
the specific frequency varying, depending on the brain
network (Siegel et al., 2012; Fries, 2005). The motor net-
work is dominated by activity in the alpha–beta range
(Miller et al., 2012; Yanagisawa et al., 2012; Brovelli et al.,
2004; Kuhn et al., 2004; Cassidy et al., 2002; Crone, Miglioretti,
Gordon, Sieracki, et al., 1998; Sanes & Donoghue, 1993;
Murthy & Fetz, 1992; Pfurtscheller, 1981). Thus, the re-
duction in beta coherence seen in this study would be
Swann et al.
91
expected to impair communication between the motor
cortex and its subcortical modulators, consistent with the
“communication through coherence” hypothesis (Fries,
2005). PAC provides a mechanism to explain how co-
herence might influence local neural activity (Canolty &
Knight, 2010; Canolty et al., 2006). Thus, our observed
coherence and PAC decreases are consistent with a
general reduction in motor network connectivity and,
perhaps, communication, which can be detected with both
metrics. We predict that a similar phenomenon might be
observed in a different frequency range if a different brain
network (with a different predominant frequency range)
were studied.
Opposing Patterns of Functional Connectivity
in Consciousness
Some studies, similar to our own, show a decrease in
synchrony between distributed areas during loss of
consciousness (Chennu et al., 2014; Sarasso et al., 2014;
Bonhomme, Boveroux, Brichant, Laureys, & Boly, 2012;
Massimini, Ferrarelli, Sarasso, & Tononi, 2012; Laureys
et al., 1999), whereas others showed an increase (Mukamel
et al., 2014; Breshears et al., 2010; Ching, Cimenser,
Purdon, Brown, & Kopell, 2010; Arthuis et al., 2009;
Feshchenko, Veselis, & Reinsel, 2004). One explanation
for these seemingly contradictory findings may lie in the
distinction between “healthy” synchrony, necessary to
coordinate communication between separated brain areas
(Siegel et al., 2012; Fries, 2005), and abnormal oscillatory
patterns, which may break up normally synchronized activ-
ity or constrain neural activity in an inflexible pattern. An-
esthesia may cause some populations of neurons to fire in
an oscillatory, inflexible, way, which then precludes pat-
terns of synchronization necessary for conscious behavior.
Indeed, there have been studies that observe both patterns
during loss of consciousness, that is, both an increase in
oscillatory activity in certain frequency ranges or brain net-
works, coincident with a decrease in oscillatory activity in
others (Purdon et al., 2013; Lewis et al., 2012).
Alternative Interpretations of the
Coherence and PAC Results
We have observed a decrease in coherence and PAC dur-
ing anesthesia induction that is specific to the beta band
and not accompanied by a change in beta power in
cortex (and only a general decrease in all frequencies
>20 Hz in subcortical areas). We interpret this as a reflec-
tion of a reduction in communication throughout the
motor system, which occurs specifically in beta. How-
ever, alternative interpretations are possible. For in-
stance, the power decrease in the subcortical regions
may make the subcortical phase estimate of beta less
reliable, reducing coherence. We cannot rule out this
interpretation, Tuttavia, if this were the only factor driv-
ing the effect, we might expect a more broadband coher-
ence change, because all frequencies above 20 Hz
decreased in the subcortical areas. Inoltre, this inter-
pretation is not necessarily mutually exclusive with the
hypothesis that communication at specific frequencies is
decreased, because a decrease in communication could
also be driven by a decrease in power at the dominant
frequency for a particular structure within a brain network.
Another interpretation is that movement could have
influenced the recordings, because beta is modulated
by movement (Crone, Miglioretti, Gordon, Sieracki,
et al., 1998; Pfurtscheller, 1981). Although all our patients
were at rest during the recordings, perhaps subtle, unde-
tectable movements were present in the early portions of
the recording that decreased over time with anesthesia
induction. Tuttavia, this interpretation would predict
changes opposite to those observed here (cioè., an in-
crease in coherence with anesthesia induction), because
movement has been associated with a decrease in coher-
ence in the alpha/beta range between cortical and sub-
cortical motor structures (Alegre et al., 2010; Lalo et al.,
2008; Cassidy et al., 2002).
Changes in Power in Specific Brain Regions May
Reflect Reduced Neuronal Activity or Relate
to Disconnection between Brain Regions
The propofol-induced reduction in cortical broadband
activity that we observed was weak but is in general agree-
ment with previous human studies examining ECoG dur-
ing anesthesia induction ( Verdonck et al., 2014; Breshears
et al., 2010). The subcortical broadband gamma reduc-
tion confirms similar findings in rodent studies (Reed,
Plourde, Tobin, & Chapman, 2013) and in one small series
of thalamic recordings in humans ( Verdonck et al., 2014).
Broadband gamma power in cortex is thought to be an
index of local neural activity and a surrogate for neural
spiking (Manning et al., 2009; Miller et al., 2007). IL
interpretation of broadband gamma in subcortical struc-
tures is less clear but may similarly represent the sum of
local neuronal spiking. Infatti, event-related broadband
gamma changes similar to those reported in cortex have
been observed in subcortical structures (Hamame, Alario,
Llorens, Liegeois-Chauvel, & Trebuchon-Da Fonseca,
2014; Ray et al., 2012).
Gamma power decreases are often associated with low-
frequency power increases (Crone, Miglioretti, Gordon, &
Lesser, 1998; Crone, Miglioretti, Gordon, Sieracki, et al.,
1998). The low-frequency power increases (2–6 Hz) Quello
we observe in both cortex and subcortex may relate to
this same process. Therefore, one interpretation is that
the changes in power observed during loss of conscious-
ness relate to decreased neural activity in regions that play
key roles in generating behavior.
An alternative explanation is that the low-frequency (2–
6 Hz) power changes in both cortex and subcortex have
a different etiology than the high-frequency power changes.
Several studies have reported low-frequency power
92
Journal of Cognitive Neuroscience
Volume 28, Numero 1
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
e
j
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
1
2
8
8
/
4
1
1
/
9
8
5
4
0
/
2
1
6
7
1
8
o
4
C
5
N
3
_
0
UN
/
_
j
0
o
0
C
8
N
8
_
4
UN
P
_
D
0
0
B
8
sì
8
G
4
tu
.
e
P
S
D
T
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
.
T
F
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
increases in cortex during anesthesia induction (Verdonck
et al., 2014; Purdon et al., 2013; Lewis et al., 2012; Breshears
et al., 2010). In some cases, this emerging low-frequency
activity is asynchronous across cortex and has been inter-
preted as a mechanism whereby different brain networks
become disconnected from one another (Purdon et al.,
2013; Lewis et al., 2012). In many of these studies, IL
low-frequency activity is in a slightly lower-frequency
range (<2 Hz) than is observed here; nevertheless, other
studies have shown these changes over a broader
frequency range ( Verdonck et al., 2014), and so it is
possible that similar mechanism involved.
Limitations
There are several limitations to our study. Although our
recording methods high temporal resolution, the
precision with which the state of consciousness is
assessed, using standard anesthesia scales, less tem-
porally precise. This may limited we
could detect. All participants were movement dis-
order patients who necessarily abnormal motor
networks, subcortical recording sites limited
to clinically indicated targets for ethical reasons. We
sought minimize this problem by including patients
with different diagnoses, presumably differ-
ent pathophysiologies, focusing on patterns that
were consistent across patients. Nevertheless, an alter-
native interpretation activity associated
with induction reflect cessation symptoms, which
occurs anesthesia. Additionally, inclusion dif-
ferent diagnoses further increased heterogeneity of
our sample, decreased sensitivity
for finding effects makes more com-
plex. Likewise, although examination multiple sub-
cortical allowed us focus motifs common
to nodes BGTC motor loop, also in-
creased potential signal variability. Thus, we
may be insensitive present most
strongly in only certain regions. Finally, the
use ECoG strip arrays can inserted via burr
hole spatial sampling. Our results not
generalize entire cortex (although there simi-
larities between data recorded greater
spatial sampling; see Breshears 2010). Investigation
of other, distributed brain networks was precluded
by constraints working human patients
in operating room.
Conclusion
Using method simultaneously collect
high resolution from human
cortex nuclei, we narrow-
band reduction system interactions beta
band during loss consciousness.
These support communication
throughout involves synchronous
oscillatory frequency-specific breaks
down when organized recruitment network
for behavior impossible (i.e., unconsciousness).
These shed light possible general mechanisms
of neural communication brain.
Acknowledgments
We would like thank Dr. Jill Ostrem help patient
recruitment, Oana Maties assess-
ments, Bradley Voytek helpful comments the
manuscript. We all who
participated work supported the
National Institutes Health (grant R01NS069779 P. A. S.).
Reprint requests should sent Nicole C. Swann, Health
Sciences East, Rm #823, 513 Parnassus Avenue, San Francisco,
CA 94143, or e-mail: Nicole.Swann@ucsf.edu.
REFERENCES
Alegre, M., Rodriguez-Oroz, M. C., Valencia, Perez-Alcazar,
M., Guridi, J., Iriarte, al. (2010). Changes subthalamic
activity observation Parkinson’s disease:
Is mirror mirrored basal ganglia? Clinical
Neurophysiology, 121, 414–425.
Alexander, G. E., DeLong, R., & Strick, L. (1986). Parallel
organization functionally segregated circuits linking basal
ganglia cortex. Annual Review Neuroscience, 9,
357–381.
Alkire, T., Hudetz, G., Tononi, (2008). Consciousness
and Science, 322, 876–880.
Arthuis, Valton, L., Regis, Chauvel, P., Wendling, F.,
Naccache, (2009). Impaired during
temporal lobe seizures related long-distance
cortical-subcortical synchronization. Brain, 132, 2091–2101.
Bonhomme, V., Boveroux, Brichant, J. F., Laureys, S., Boly,
M. (2012). Neural correlates general
anesthesia functional magnetic resonance imaging
(fMRI). Archives Italiennes de Biologie, 150, 155–163.
Breshears, D., Roland, Sharma, Gaona, M.,
Freudenburg, Z. Tempelhoff, Stable and
dynamic cortical electrophysiology and
emergence propofol Proceedings Academy Sciences, U.S.A., 107, 21170–21175.
Brovelli, A., Ding, Ledberg, Chen, Y., Nakamura, &
Bressler, S. (2004). Beta oscillations large-scale
sensorimotor network: Directional influences
revealed Granger causality. National
Academy 101, 9849–9854.
Canolty, R. Edwards, Dalal, Soltani, Nagarajan,
S. Kirsch, H. (2006). High gamma power is
phase-locked theta neocortex.
Science, 313, 1626–1628.
Canolty, Knight, T. The role of
cross-frequency coupling. Trends Cognitive Sciences,
14, 506–515.
Cassidy, Mazzone, Oliviero, Insola, Tonali, P.,
Di Lazzaro, (2002). Movement-related in
synchronization ganglia. 125,
1235–1246.
Chennu, Finoia, Kamau, Allanson, Williams, B.,
Monti, (2014). Spectral signatures reorganised
brain disorders consciousness. PLoS
Computational Biology, 10, e1003887.
Swann al.
93
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
>