METHODS
Structure supports function: Informing directed
and dynamic functional connectivity with
anatomical priors
David Pascucci1,2
, Maria Rubega3, Joan Rué-Queralt2,4, Sebastien Tourbier4,
Patric Hagmann4, and Gijs Plomp2
1Brain Mind Institute, École Polytechnique Fédérale de Lausanne (EPFL), Lausanne, Svizzera
2Perceptual Networks Group, University of Fribourg, Fribourg, Svizzera
3Department of Neurosciences, University of Padova, Padova, Italy
4Connectomics Lab, Department of Radiology, University Hospital of Lausanne and University of Lausanne (CHUV-SUNIL),
Lausanne, Svizzera
a n o p e n a c c e s s
j o u r n a l
Keywords: Functional connectivity, Structural connectivity, Dynamic connectivity, Multimodal
imaging, Brain networks, Directed connectivity
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
N
e
N
_
UN
_
0
0
2
1
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
ABSTRACT
The dynamic repertoire of functional brain networks is constrained by the underlying topology
of structural connections. Despite this intrinsic relationship between structural connectivity
(SC) and functional connectivity (FC), integrative and multimodal approaches to combine the
two remain limited. Here, we propose a new adaptive filter for estimating dynamic and
directed FC using structural connectivity information as priors. We tested the filter in rat
epicranial recordings and human event-related EEG data, using SC priors from a meta-analysis
of tracer studies and diffusion tensor imaging metrics, rispettivamente. We show that, particularly
under conditions of low signal-to-noise ratio, SC priors can help to refine estimates of directed
FC, promoting sparse functional networks that combine information from structure and
function. Inoltre, the proposed filter provides intrinsic protection against SC-related false
negatives, as well as robustness against false positives, representing a valuable new tool for
multimodal imaging in the context of dynamic and directed FC analysis.
INTRODUCTION
The white matter architecture of the human brain constitutes the structural backbone for neu-
ronal communication. A fixed network of axonal pathways wires an extremely rich repertoire
of brain functions, from short-range interactions to large-scale dynamics that support percep-
zione, cognition, and action (Petersen & Sporns, 2015). As in all biological systems, properties of
the structure constrain the possible functions. Organizational principles of structural brain net-
works, such as small-world and modular architectures, determine the topological space for
functional interactions at the meso- and macroscale (Hagmann et al., 2008; Sporns, 2010).
At the microscale, the absence of a synaptic connection between two neurons makes a direct
functional coupling biologically impossible. Despite the inherent link between structural con-
nectivity (SC) and functional connectivity (FC), only a few existing studies have tried to
Citation: Pascucci, D., Rubega, M.,
Rué-Queralt, J., Tourbier, S., Hagmann,
P., & Plomp, G. (2022). Structure
supports function: Informing directed
and dynamic functional connectivity
with anatomical priors. Network
Neuroscience, 6(2), 401–419. https://doi
.org/10.1162/netn_a_00218
DOI:
https://doi.org/10.1162/netn_a_00218
Supporting Information:
https://doi.org/10.1162/netn_a_00218
https://github.com/ PscDavid/dynet
_toolbox
https://github.com/joanrue/pydynet
Received: 31 agosto 2021
Accepted: 23 novembre 2021
Corresponding Author:
David Pascucci
david.pascucci@epfl.ch
Handling Editor:
Olaf Sporns
Copyright: © 2021
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
Structurally informed STOK
Dynamic functional connectivity:
A loose term to define functional
connectivity measures that are
functions of time, accounting for
temporal changes and
reconfigurations of functional
network patterns.
Direct connectivity:
A metric that distinguishes between
direct and indirect flows in a
rete, quantifying the influence
that one signal produces directly on
another, without including indirect
paths.
Directed connectivity:
A directional measure of statistical
dependence among multiple signals,
quantifying the influence directed
from one signal to another in the
Granger causality framework.
combine the two modalities, particularly in the domain of directed and dynamic functional
connectivity (Lei et al., 2015; Suárez et al., 2020).
The relationship between structural wiring and functional coupling is at the core of several
statistical and biophysical models of brain networks (Honey et al., 2010). These models advo-
cate a substantial overlap between SC and FC, at both the mesoscopic and the macroscopic
scales. Empirical and modeling studies on resting-state brain networks provide converging
support, showing that the weights of structural and functional networks, as well as their topo-
logical features, tend to be correlated, and the strength of between-regions SC is typically a
good predictor of their FC (Deco et al., 2013; Honey et al., 2009; Mišić et al., 2016; Skudlarski
et al., 2008). Given the established overlap between structure and function, measures of FC
may be meaningfully improved by taking SC into account, as suggested by a few proposed
metodi. In the framework of Bayesian modeling, for instance, structural graphs have been
incorporated as priors for generative models of FC (Sokolov et al., 2019). A structural graph
is typically obtained from in vivo diffusion-weighted imaging (DWI) data that quantify the
anisotropy in the diffusion of water molecules along white matter tracts (Hagmann et al.,
2008). The connectivity graph is either a binary or a weighted undirected adjacency matrix
that provides information about the presence and strength of a physical link between distinct
brain regions. It has been shown how adding SC graphs as priors for effective connectivity
substantially improves model evidence (Sokolov et al., 2019). Allo stesso modo, constraining FC for
only present SC links and anatomically determined time lags may reduce false positives and
improve the spatial resolution of electroencephalography (EEG) source imaging (Filatova et al.,
2018; Takeda et al., 2019).
Whereas previous work has focused mainly on combining SC and FC for the analysis of
functional magnetic resonance imaging data (fMRI), similar integrative approaches are missing
for the emerging field of time-varying directed FC analysis (Eichenbaum et al., 2021). Time-
varying FC has been used to characterize the dynamics of directed neuronal interactions that
evolve at the millisecond scale, exploiting high-temporal-resolution recordings, such as local
field potentials and EEG source imaging data (Milde et al., 2010; Pascucci et al., 2018; Plomp
et al., 2014).
Recentemente, we introduced a variant of the classic Kalman filter, the self-tuning optimized
Kalman (STOK) filter (Pascucci et al., 2020), for modeling rapid changes in large-scale func-
tional networks during evoked brain activity. Here, we present an extension of this algo-
rithm that incorporates prior information on the structural connectivity: the structurally
informed STOK (si-STOK). The algorithm provides a straightforward tool to combine SC
(per esempio., diffusion tensor imaging [DTI]-derived metrics) with dynamic FC. In the proposed
approach, SC priors act as regularization terms in the estimation of “effects,” within the
Granger causality framework (Granger, 1969). Hence, the method is different from genera-
tive and predictive models that use SC as priors on the generation of neuronal dynamics
and interregional coupling (Stephan et al., 2009). The underlying assumption of our method
is that statistical influences among brain regions must follow the direct and directed nature
of brain interactions, which occur between nodes of a network that are structurally linked.
Hence, SC priors provide information about the fixed structure of directed links in which
dynamic FC is likely to occur, without forcing FC, at any time point, to be necessarily iden-
tical to SC (see Methods).
We tested the algorithm in benchmark data and evaluated the effect of different SC matrices
on the estimated FC. We demonstrated the advantages of combining SC with FC in terms of
noise resilience and consistency of the estimates. We then compared the two algorithms in
Network Neuroscience
402
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
N
e
N
_
UN
_
0
0
2
1
8
P
D
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Structurally informed STOK
Time-varying multivariate
autoregressive model:
A model that estimates lagged linear
effects within a multivariate system of
variables, providing recursive
estimates that are a function of time
and are collected for each time point.
event-related large-scale functional brain networks during face processing. Our results showed
that incorporating SC in dynamic FC promotes sparser and physiologically plausible topolo-
gies of functional networks, aiding the identification of the main network drivers and dynam-
ics. MATLAB and Python code for si-STOK are available on GitHub (Pascucci & Rubega,
2019, https://github.com/ PscDavid/dynet_toolbox; Rué-Queralt, 2019, https://github.com
/joanrue/pydynet).
RESULTS
Somatosensory Evoked Potentials in Rats
To incorporate structural priors in dynamic functional connectivity, we developed a variant of
the self-tuning optimized Kalman (STOK) filter (Pascucci et al., 2020). The STOK is an adap-
tive filter that derives time-varying multivariate autoregressive (tv-MVAR) coefficients using a
simple least squares regression of present on past signals. We exploited this core feature of the
filter to incorporate SC as shrinking priors of a least squares solution (see Methods, Equazione 7).
SC matrices were used to calibrate the variance of a prior expectation of zero FC between
each pair of nodes (Sokolov et al., 2019; Figura 1). Strong SC values correspond to large prior
variance, allowing FC to deviate from zero, whereas weak SC values reduce the prior variance
and shrink FC toward zero. Hence, the filter’s estimates combine the strength of SC with the
FC supported by the data (see Methods and Figure 1).
We tested the proposed algorithm, termed structurally informed STOK (si-STOK), on a
benchmark dataset of epicranial EEG recordings in rats, from a whisker stimulation protocol
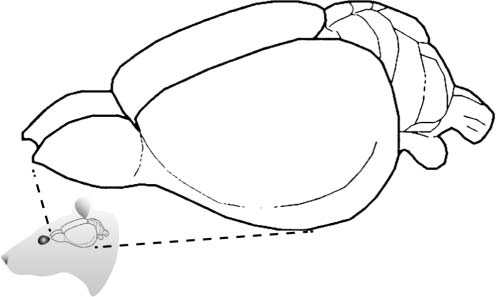
(Plomp et al., 2014; Quairiaux et al., 2011). After whisker stimulation, action potentials orig-
inate and propagate rapidly (per esempio., within 10–25 ms) from the contralateral primary sensory
cortex, following the underlying structural connectivity (Plomp et al., 2014) (see Figure 2A).
This pattern was accurately recovered by the STOK filter, which detected an overall larger
magnitude of outgoing directed influences (MDI; see Methods) from the contralateral sensory
cortex (e4), at early poststimulus latencies (13 ms; see Figure 2A and Pagnotta et al., 2018;
Pagnotta & Plomp, 2018; Plomp et al., 2014). We used a directed, weighted matrix of SC that
we derived from a meta-analysis of reported structural connections (Bota et al., 2015; Swanson
et al., 2017) (see Methods). Compared with the regular STOK filter, the inclusion of SC priors
provided qualitatively similar results, with clearer dynamics and visible but minor changes
(Figure 2A). The similarity between the results of the two algorithms was a consequence of
the use of a dense SC matrix, with connection weights that did not deviate drastically from
the estimated FC. When evaluating the magnitude of outgoing influences from e4 to all the
other nodes, Infatti, the results were highly comparable (Figure 2B). Tuttavia, incorporating
a sparser SC matrix, with only 25% of the strongest SC connections, led to evident changes.
When one of the expected FC connections (from e4 to e2) was absent in SC, the resulting
estimate decreased considerably (Figure 2B). Nevertheless, FC from e4 to e2 was still larger
compared with two other SC-absent connections (from e4 to e1 and from e4 to e8) for which
weak or no FC was supported by the data. Conversely, for connections with weak FC, strong
SC did not drastically increase FC. This demonstrated that in the present modeling framework
the inclusion of structural priors has a low risk of producing false negatives (the downscaling
of FC for absent SC depends also on the strength of FC), while it is also robust against the risk
of introducing false positives driven by strong SC in the absence of FC.
To better appreciate the advantages of combining SC and FC, we compared the perfor-
mance of the two algorithms under noise perturbations. We evaluated two criteria (Vedere
Methods): (UN) the consistency of the estimated network at the e4 peak latency; E (B) IL
Network Neuroscience
403
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
N
e
N
_
UN
_
0
0
2
1
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Structurally informed STOK
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
N
e
N
_
UN
_
0
0
2
1
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
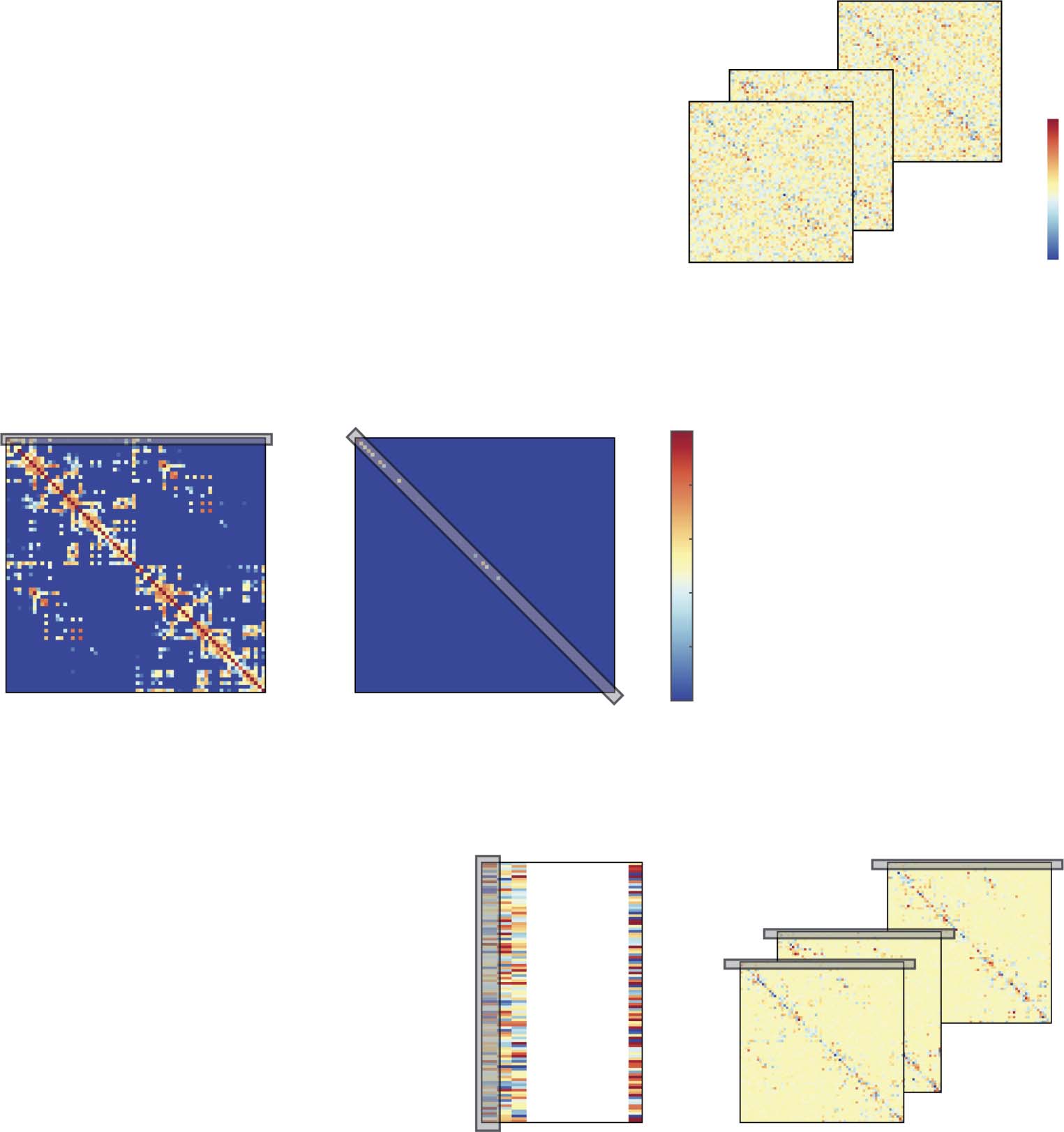
Figura 1. Estimation of structurally informed dynamic FC using the si-STOK filter. (UN) A least squares solution is used to estimate the coef-
ficient matrix A of a time-varying multivariate autoregressive model by regressing past H on present (Z) values of the multivariate time series
(see Equation 4; the symbol + stands for the matrix pseudoinverse). (B) SC priors (per esempio., DTI metrics) are incorporated in the filter as the prior
variance on the expected zero FC from all the sender nodes to each receiver node. Weak SC corresponds to small prior variance, shrinking the
coefficient estimates toward zero; strong SC corresponds to large prior variance, allowing the estimated coefficients to deviate more from zero
when supported by the data. (C) The regularizing matrix Q informs the least squares solution with priors on the variance of autoregressive
coefficients based on SC, resulting in MVAR models that combine FC and SC.
ability to detect e4 as the main driver compared with the average network activity. The two
criteria were tested by varying the ratio of noise to signal. As evident in Figure 2C, the si-STOK
outperformed the regular STOK for both criteria at all the noise levels. This highlighted an
additional important feature of the new filter: when FC is informed by SC, the estimated net-
works become more resilient to noise and present a consistent topology and nodal strength
under perturbations.
Network Neuroscience
404
Structurally informed STOK
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
N
e
N
_
UN
_
0
0
2
1
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
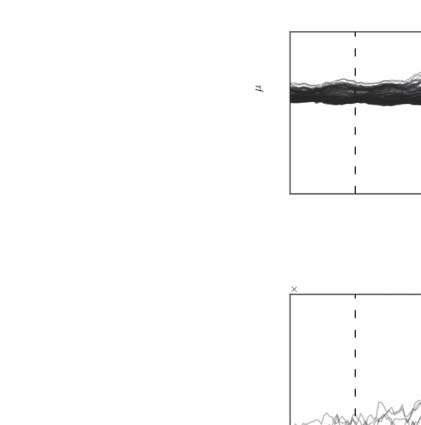
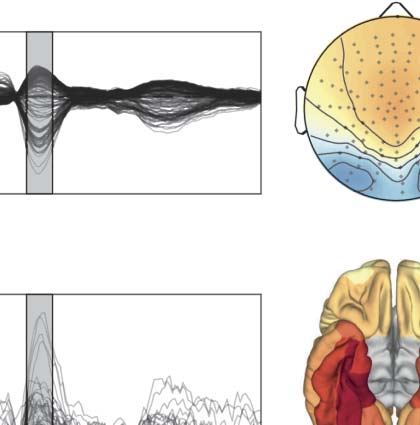
Figura 2. Validation in benchmark rat EEG data. (UN) Contralateral evoked potentials after whisker stimulation, showing the peak in the pri-
mary sensory cortex (e4). Activity propagates rapidly after stimulation from the primary sensory cortex to nearby parietal and frontal regions (e2
and e6). The role of the primary sensory cortex as the main driver of evoked activity is evident from the summed magnitude of outgoing
directed influences (MDI) estimated with both the STOK and the si-STOK filters. (B) Effect of incorporating SC priors in the estimated directed
influences from e4 to the rest of the network at the peak latency of the evoked activity (13 ms). Boxplots summarize the results of the STOK
filter with (empty boxplots) and without (filled boxplots) SC priors, across ten animals. By using a dense SC matrix with almost uniform priors,
the estimated FC are highly similar with and without SC priors. Retaining only 25% of the strongest SC connections demonstrates the relative
shrinkage of SC-absent FC and the resistance of the algorithm against SC-related false positives: Under an SC-absent prior, the expected FC
from e4 to e2 was still larger compared with other connections, whereas strong SC priors did not inflate FC when weak FC was supported by
the data (per esempio., from e4 to e5). (C) Performance evaluation under noise perturbations, after varying the proportion of signal to noise (additive) In
the original data. Compared with the regular STOK, the si-STOK FC estimates showed overall larger consistency with the FC estimated in the
absence of additional noise, as the proportion of noise perturbing the data increased (left panel). Allo stesso modo, the si-STOK showed a higher ability
to detect the contralateral primary sensory cortex as the main driver of network activity at peak evoked latencies (right panel; the black line
indicates the estimated e4 outflow, subtracted from the average network outflow at 13 ms, with the si-STOK in the absence of noise pertur-
bations; the dashed line indicates the estimate obtained with the regular STOK). SC priors lead to an increased ability to detect e4 as the main
network driver at all noise levels tested. Shaded lines in panel A are 95% CI of the mean. The lack of a noise-absent reference value in the
Consistency plot of panel C is due to the fact that Consistency values for noise-free estimates are always 1.
Human EEG Data
After validating the algorithm in benchmark data recorded in rats, we employed the si-STOK
filter to model FC in event-related human EEG data. We modeled FC in a large-scale network
Di 68 brain regions in response to faces and scrambled stimuli (Desikan et al., 2006) (Vedere
Methods and Figure 3A). On the electrode level, the comparison of evoked responses between
Network Neuroscience
405
Structurally informed STOK
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
N
e
N
_
UN
_
0
0
2
1
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
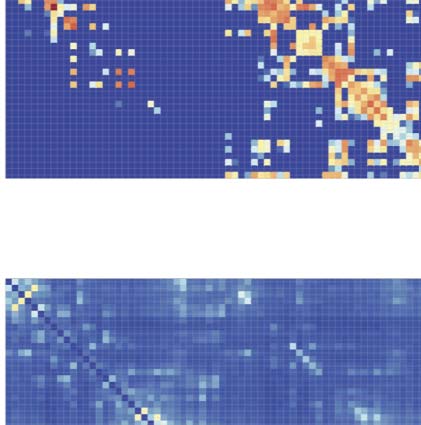
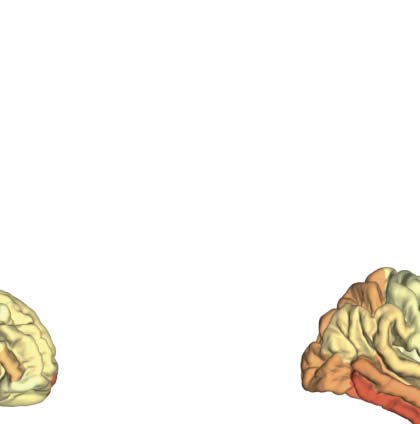
SC priors for large-scale FC analysis of human EEG data. (UN) Example of the sequence of events in one trial of the face detection
Figura 3.
task. Face and scrambled stimuli were randomly interleaved across trials. EEG data were time-locked to the onset of the stimuli. (B) Scalp-
evoked responses showing the typical topography and time course of the face-related N170 component (faces minus scrambled, peak at
160 ms, upper panels). Source reconstruction localized the source of face-selective responses in the right fusiform and nearby occipitotem-
poral areas (lower panels). (C) The group SC prior matrix (upper panel) and the FC (MDI index) estimated with the regular STOK (central
panel) and the si-STOK (lower panel) in response to face stimuli, averaged in a time window around the face-selective N170 response (from
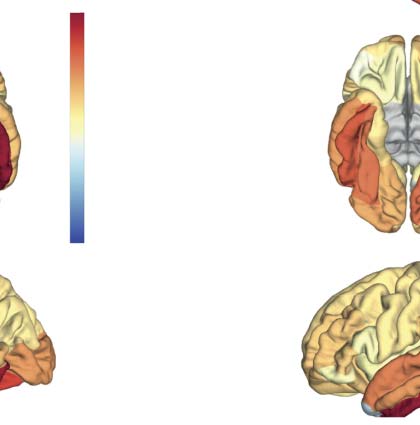
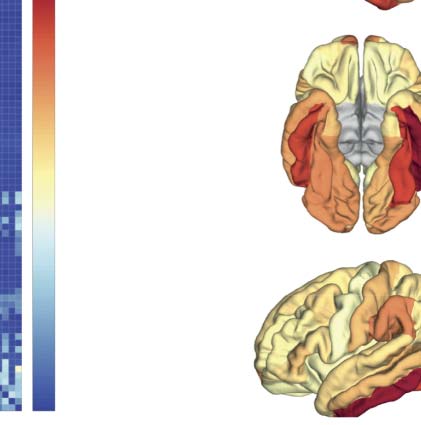
140 A 180 ms poststimulus). (D) Ranked summed outflow in response to faces (faces minus scrambled) from all the 68 areas, obtained
using the regular STOK (bar plot on the left) and the si-STOK (bar plot on the right). See Supplementary File 2 for abbreviations. IL
inclusion of SC priors resulted in a less scattered topology of face-selective outflows, with the largest network drivers localized in primary,
secondary visual areas and regions of the fusiform and inferior-temporal cortex.
Network Neuroscience
406
Structurally informed STOK
faces and scrambled images revealed the typical topography and time course of the face-
related N170 component (faces minus scrambled, peak at 160 ms), with the largest difference
between conditions localized in the right fusiform and nearby occipitotemporal areas (Vedere
Figure 3B).
Figure 3C shows the effect of incorporating SC priors on the estimated FC at a time window of
interest around the N170 response. FC matrices obtained through the si-STOK filter showed the
clear shrinkage of functional connections for weak and absent SC, leading to FC matrices that
partly inherit the structure of SC but preserved intrinsic patterns of FC coupling (see Figure 3C), In
line with the rat EEG benchmark results. For the same time window of evoked activity, we com-
pared the summed outflow from each area between the two conditions (faces minus scrambled,
summed MDI) as a measure of changes in nodal strength during face processing. In the ranked
outflow, the two filters agreed in identifying the bilateral inferior temporal gyrus and the left fusi-
form gyrus as the three areas with the largest increase in outflow in response to faces (Vedere
Figure 3D). Without structural priors, Tuttavia, frontal regions were also ranked among the larg-
est drivers at short poststimulus latencies (per esempio., rFPol, lFPol; see Supplementary File 2 for abbre-
viations), and the outflow increase in response to faces was more or less pronounced throughout
the entire network. With the inclusion of structural priors, IL 10 largest drivers of face-related
activity were all located in the primary and secondary visual cortex, including the bilateral fusi-
form, lateral occipital cortex, lingual gyrus (per esempio., V1), and regions in the temporal cortex (Vedere
Figure 3D), while the summed outflow from the rest of the network decreased progressively.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
N
e
N
_
UN
_
0
0
2
1
8
P
D
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Effect of SC priors on the evoked dynamics of directed interactions in a subset of brain areas of the core face network. Shaded lines
Figura 4.
are standard errors of the mean. Red and blue horizontal bars highlight statistically significant differences (faces vs. scrambled, permutation
test, P(unc.) < 0.01).
Network Neuroscience
407
Structurally informed STOK
These comparisons suggest that the inclusion of SC priors refines the topology of FC net-
works and portrays the contribution of each node in a more physiologically plausible way.
This feature can aid the identification of hubs and critical modules in large-scale FC analysis.
A further important question is whether SC priors also affect the temporal dynamics of FC. We
evaluated this aspect in a final analysis where we compared the estimated changes in directed
influences in response to face and scrambled stimuli, time-locked to the stimulus onset (from
−100 to 500 ms). For this analysis, we considered a subset of regions in the core face network:
the right inferior temporal gyrus (rIT), the right fusiform (rFu), and the right superior temporal
sulcus (rB). The functional role of these brain areas in face processing is well documented (Fox
et al., 2009; Haxby et al., 2000), and their SC is a predictive feature of face-specific activity
(Saygin et al., 2011), representing a functionally specialized module with known structure-
function relationships. Figure 4 shows the estimated time-varying MDI with and without SC
priors. This comparison revealed clear differences that extended well beyond the basic out-
flow summary described above, specifically the following: (a) SC priors led to quantitative
increases in unidirectional interactions at specific latencies, consistent with face-related
evoked dynamics (e.g., from rFu to rIT, from rIT to rB); (b) SC priors more clearly show the
sequence of unidirectional interactions between nodes (e.g., from rIT to rFu, followed by
rFu to rIT); (c) SC priors shrunk and underestimated weak FC for SC-absent connections,
reducing the risk of false positives (e.g., the connection from rIT to rB at very short poststimulus
latencies).
DISCUSSION
We introduced and validated a new adaptive filter for combining structural and dynamic func-
tional connectivity, the si-STOK. The algorithm builds on a recent variant of the general linear
Kalman filter (Kalman, 1960; Milde et al., 2010; Pascucci et al., 2020) and allows incorporat-
ing structural priors in a multivariate autoregressive modeling framework with high temporal
resolution. Structural priors are used as starting assumptions on the structure of a regularizing
matrix applied to the estimation of time-varying multivariate autoregressive models, in the
framework of the generalized Tikhonov regularization method (Plato & Vainikko, 1990).
We tested the filter in rat epicranial recordings and human EEG data, using SC priors from a
meta-analysis of tracer studies and DTI metrics, respectively. Compared with FC without SC
priors, we found the following advantages. First, FC estimates were more consistent and more
resilient to noise. Second, SC priors promoted sparser FC networks and favored a more accu-
rate identification of the main network drivers at expected poststimulus latencies. Third, using
SC as prior variance provided solutions with intrinsic protection against SC-related false neg-
atives (for discordant SC-FC, the filter relied more on the data and less on the prior), and
robustness against false positives (strong SC did not inflate FC unless supported by the data).
The first two aspects represent desired features considering the expected sparsity of FC net-
works and the sparse topology of the underlying structural links (Markov et al., 2012; Pagnotta
et al., 2019; Valdés-Sosa et al., 2005). Previous work has shown how sparse and regularized
approaches to FC analysis can decrease spurious connections, increase robustness to noise
(Pagnotta et al., 2019), and counteract issues due to limited data points (Antonacci et al.,
2019; Valdés-Sosa et al., 2005). Informing sparse solutions through the fixed topology of SC
links has the additional benefit of introducing a biologically grounded space for regularization.
This represents an advantage in conditions where regularization may have a strong impact on
the model structure, such as under multicollinearity and nonindependence among time-series
(e.g., under linear mixing of source EEG activity due to volume conduction; Anzolin et al.,
Network Neuroscience
408
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
2019; Haufe et al., 2013). The resulting FC partly inherits the sparsity and topological prop-
erties of the SC matrices, while preserving the strength and directedness of functional inter-
actions. This may ultimately facilitate graph analysis of functional networks, such as the
identification of FC hubs, modules, and nodal properties without additional sparsity-based
and consensus-based thresholding, which may lead to unstable and threshold-dependent
network estimates (Garrison et al., 2015).
The third important feature of the si-STOK is the protection against false positives due to
invalid SC priors. Previous studies, including large-scale validations of tractography pipe-
lines, have reported a high ratio of invalid connections and a substantial amount of false
positives (Maier-Hein et al., 2017). This caveat undermines the possibility to simply mask
or weight FC by SC, increasing the risk of inflating FC for invalid SC connections. The pro-
posed algorithm employs the generalized Tikhonov method (Plato & Vainikko, 1990), a pow-
erful and versatile regularization scheme under a Bayesian perspective where priors are
combined with the observed data. Strong SC, therefore, does not necessarily inflate FC, as
demonstrated in our test in benchmark data. Conversely, strong FC can still be detected in
the absence of SC. This was also evident from the results in rat EEG data, where we
observed FC between the primary somatosensory and the parietal cortex (e4 to e2; see
Figure 2) even under a strong prior of no SC. Although reduced, FC between these areas
was still larger compared with other connections for which SC was present but only weak
FC was expected from physiology (e.g., from e4 to e8). This accommodates the possible
divergence between FC and SC connections (Honey et al., 2009; Lim et al., 2019), which
may arise from indirect structural connections, false negatives in SC (Damoiseaux & Greicius,
2009), or because of the differential engagement of specific functional modules under different
task demands (Sokolov et al., 2019).
From our test in benchmark data it is also clear that, under certain circumstances, SC priors
have minor effects on FC. This may occur, for instance, when using dense SC matrices in con-
ditions where the signal-to-noise ratio is high, or when SC does not deviate drastically from FC.
The rat EEG data analyzed here are an example. The rat cortex is essentially flat (lissencepha-
lic) with few expected deep sources. The signal recorded at each electrode is therefore an
accurate representation of the activity flowing through the structural pathways in the cortex
underneath (Quairiaux et al., 2011). Using dense SC priors, generally in agreement with the
expected FC, provided no additional or diverging information (see Figure 2B). In many other
applications, however, signals are contaminated with multiple sources of noise and the record-
ings are not direct measurements of brain activity and connectivity. Under these common cir-
cumstances, SC priors may have more appreciable and beneficial effects on FC estimates. Our
manipulation of added noise confirmed this advantage (Figure 2C).
Recently, a method to combine structural and functional connectivity in the framework of
autoregressive modeling has been proposed and tested in fMRI data (Crimi et al., 2021). This
approach relies on stationary (e.g., time-invariant) MVAR models in which a set of constraints,
based on SC, force the model to estimate parameters only for existing, direct structural con-
nections. The estimation is performed using a gradient descent approach. While our algorithm
bears similarities to this approach, the key differences are that si-STOK is developed for time-
varying MVAR modeling of fast, nonstationary dynamics, and that it incorporates structural
information as regularizing priors, rather than strong constraints on model estimation. The
approach presented here, therefore, should provide solutions that are more robust against
SC false negatives, as SC provides information about the variance of FC, rather than imposing
the presence/absence of a connection.
Network Neuroscience
409
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
The proposed algorithm is the first to integrate SC in a directed and dynamic measure of FC.
We evaluated the effect of SC priors on large-scale dynamics of directed interactions in human
source EEG activity during a face detection task. We considered dynamic interactions in a
subset of regions corresponding to key nodes of the core face network (Haxby et al., 2000;
Saygin et al., 2011). Our results showed that SC priors can also shape the temporal dynamics
of FC. Particularly, incorporating SC priors revealed an initial face-selective increase of FC
from the right inferior temporal gyrus to the fusiform, followed by an increase of FC from
the fusiform to the inferior temporal gyrus and from the inferior temporal gyrus to the superior
temporal sulcus. Without SC priors, a precise sequence of FC changes was less distinguish-
able, and face-evoked increases in FC were also present at unreasonably short poststimulus
latencies (e.g., from the inferior temporal gyrus to the superior temporal sulcus; see Figure 4).
The observed pattern may well reflect the buildup of face-specific processing supported by a
hierarchy of recurrent interactions in the core face network: the inferior temporal cortex may
first relay information about global aspects (e.g., the shape and coarse structure of the face
stimuli used here) to the fusiform which then feed back information about finer details
(Goffaux et al., 2011; Sugase-Miyamoto et al., 2011; Tovée, 1995). Although the interpre-
tation of these results remains speculative and goes beyond the purpose of the current
work, the results of this analysis are a clear example of how SC priors can enhance or
downregulate time-varying dynamics in FC.
Our adaptive filter follows the aim of recent biological and mathematical modeling
approaches that use anatomical information for predictive and generative models of functional
interactions (Crimi et al., 2021; Jirsa et al., 2010). Particularly, structural graphs have been
used as priors in the Bayesian framework of dynamic causal modeling (Sokolov et al.,
2019). In this context, SC represents a prior on the generation of neuronal dynamics and inter-
regional coupling, with the target of identifying candidate mechanisms that support the bio-
physical transfer of neuronal activity. By contrast, the si-STOK comes from the framework of
Granger causality (Granger, 1969), in which causal influences are described through statistical
dependencies between the past and present of multivariate time series. Hence, the target prop-
erty is the effect and not the mechanism. The use of SC information has therefore a different
meaning than in a generative framework. SC priors act as constraints on the estimation of
“effects”—linear and directed statistical dependencies between signals recorded at multiple
brain areas. While similar priors could in principle be used with dynamic causal models
(Sokolov et al., 2019), the MVAR-based si-STOK formulation is computationally more advan-
tageous for large networks. Si-STOK thus provides an efficient way to exploit structural con-
straints in the estimation of large-scale, high-temporal-resolution FC, building on an algorithm
suited to track nonstationary signals with rapid and abrupt changes, such as those recorded
during evoked brain activity (Pascucci et al., 2020).
As a final note, it is worth mentioning that FC, in general, refers to a statistical covariation
between two signals and, therefore, FC can also exist without a structural link, as reported in
many studies (Reid et al., 2019). The measure proposed here incorporates the physiologically
grounded assumption that direct and directed statistical influences must occur between nodes
of a network that are structurally linked (e.g., not as the result of indirect and mediating
variables).
In sum, we provide a new method for dynamic FC that incorporates priors on structural
connectivity for the analysis of multivariate electrophysiological signals. The algorithm offers
a simple and powerful tool for multimodal imaging that can meaningfully contribute to inte-
grative approaches in network neuroscience (Crimi et al., 2016; Lei et al., 2015; Sokolov et al.,
2019; Stephan et al., 2009). It allows the incorporation of various types of SC, weighted or
Network Neuroscience
410
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
binary, symmetric or directed. Because of its simple form, the algorithm can flexibly incorpo-
rate different types of SC priors, from basic metrics, such as the number of white matter fibers
and the Euclidean distance, to graph-derived metrics, such as the path length and communi-
cability (Vázquez-Rodríguez et al., 2019), as well as FC priors from other modalities.
METHODS
Benchmark EEG Data
Rat EEG recordings. Benchmark data are publicly available EEG recordings from a grid of
16 stainless steel electrodes placed directly on the skull bone of 10 young Wistar rats (P21;
half males). Data were collected during unilateral whisker stimulations under light isoflurane
anesthesia (available from https://osf.io/fd5ru). Details about the recording can be found in
the original publication (Plomp et al., 2014; Quairiaux et al., 2011). Data were acquired at
2,000 Hz, bandpass filtered online, and downsampled to 1,000 Hz before connectivity anal-
ysis. All animal handling procedures were approved by the Office Vétérinaire Cantonal
(Geneva, Switzerland) following Swiss federal laws.
Rat structural connectome. Structural priors for the rat EEG benchmark were obtained from a
published meta-analysis of histologically defined axonal connections between cortical regions
in rats (Bota et al., 2015; Swanson et al., 2017). Particularly, we used one dataset containing
ranked connection weights based on reported association and commissural connections.
Details of the dataset can be found in the original publication (Dataset S3; from Swanson
et al., 2017). Ranked connection weights for pairs of regions corresponding to the electrodes
recording sites were manually selected by an expert biologist from primary visual areas,
somatosensory, primary, and secondary motor areas, and cingulate cortex (see Supplementary
File 1). Connection weights, ranging from 0 to 12 (from absent to very strong; see Supplemen-
tary File 1) were organized into a 16-by-16 structural connection matrix whose main diagonal
elements (e.g., self-connections) were set to the maximum value of 12. The structural matrix
was then normalized to the maximum value and used as a prior for the time-varying connec-
tivity analyses of the rat EEG data.
Human EEG Data
Task and stimuli. EEG data were recorded while 20 participants (3 males, mean age = 23 ±
3.5) performed a face detection task (see Figure 3A) in a dimly lit and electrically shielded
room. Each trial lasted 1.2 s and started with a blank screen (500 ms). After the blank screen,
one image (either a face or a scrambled image of a face) was presented for 200 ms, and par-
ticipants had the remaining 1,000 ms to respond. The task was to report whether they saw a
face or not (yes/no task) by pressing two buttons in a response box with their right hand
(ResponsePixx, VPixx technologies). Faces and scrambled faces were randomly interleaved
across trials. After the response and a random interval (from 600 to 900 ms), a new trial
began. The experiment consisted of four blocks of 150 trials each, for a total of 600 trials,
that is, 300 with faces and 300 with scrambled faces. Face stimuli were female and male
faces (4 by 4 degrees of visual angle, dva) taken from online repositories and cropped with
a Gaussian kernel to smooth the borders. Scrambled images were obtained by fully random-
izing the phase spectra of the original images (Ales et al., 2012). Stimuli were generated using
PsychoPy (Peirce, 2008) and presented on a VIEWPixx/3D display system (1920 × 1080
pixels, refresh rate of 100 Hz). All participants provided written informed consent before
the experiment and had a normal or corrected-to-normal vision. The experiment was
approved by the local ethical committee.
Network Neuroscience
411
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
EEG acquisition and preprocessing. Data were recorded at 2,048 Hz with a 128-channel Bio-
semi Active Two EEG system (Biosemi, Amsterdam, the Netherlands). Signal quality was
ensured by monitoring and maintaining the offset between the active electrodes and the
common mode sense–driven right leg (CMS-DRL) feedback loop under a standard value
of ±20 mV. After each recording session, individual 3D electrode positions were digitized
using an ultrasound motion capture system (Zebris Medical GmbH). One participant was
excluded because of too many motion artifacts, leaving 19 datasets for analysis. Further
details on the recordings and preprocessing pipeline can be found in the original manuscript
of the VEPCON dataset (OpenNeuro Dataset ds003505; Pascucci et al., 2021).
EEG source imaging. EEG source imaging was performed using Cartool (Brunet et al., 2011)
and custom-made scripts in MATLAB R2020b (9.9.0.1524771 Update 2). Source reconstruc-
tion was based on individual MRI data and the LAURA algorithm implemented in Cartool (reg-
ularization 6; spherical model with anatomical constraints, LSMAC), limiting the solution
space to gray matter voxels. Source activity for freely oriented dipoles was extracted from
all the source points inside each of the 68 cortical areas and projected to a representative
single direction for each area, using the singular values decomposition approach (Rubega
et al., 2019; time window for estimating the main direction: 140–250 ms poststimulus). Before
functional connectivity analysis, a global z-score transformation was applied to the entire data-
set of each participant. Epochs of source activity corresponding to trials with behavioral errors
were then removed and the dataset was divided into two conditions, according to trials con-
taining faces or scrambled stimuli.
MRI acquisition and preprocessing. A detailed description of the MR acquisition and preprocess-
ing can be found in the original manuscript of the VEPCON dataset (OpenNeuro Dataset
ds003505; Pascucci et al., 2021).
Structural connectome. Structural connectivity matrices were estimated from the reconstructed
fiber orientation distribution (FOD) image using the SD_stream deterministic streamline trac-
tography algorithm implemented in MRtrix 3.0.0-RC1 (Tournier et al., 2019). Fiber streamline
reconstruction started from seeds in the white matter that were spatially random, and the
whole process was completed when 1M fiber streamlines were reconstructed. At each stream-
line step of 0.5 mm, the local FOD was sampled, and from the current streamline tangent
orientation, the orientation of the nearest FOD amplitude peak was estimated via a Newton
optimization on the sphere. Fibers were stopped if a change in direction was greater than
45 degrees. Fibers with a length not in the 5–200 mm range were discarded. The streamline
reconstruction process was complete when both ends of the fiber left the white matter mask.
Then, for each scale, the parcellation was projected to the native DTI space after symmetric
diffeomorphic coregistration between the T1w scan and the diffusion-free B0 using ANTs
2.2.0. Finally, the connectivity matrix was built according to the Desikan parcellation atlas,
using the log of the number of fibers as the connectivity measure. Only 68 areas from cortical
volumes were included in the structural matrix and used for functional connectivity analysis.
A consensus group-representative structural brain connectivity matrix was generated from the
connectomes of all participants’ connectomes using the method introduced in Betzel et al.
(2019). Each participant connectivity matrix was then thresholded by preserving the group-
representative connection density independently for intra- and interhemispheric connections.
This allows retaining more interhemispheric connections in comparison to simple connec-
tome thresholding. The resulting connection density is set to 30%. The median of the
obtained structural connectomes across participants was then normalized to its maximum
and used as a group structural prior for connectivity analysis.
Network Neuroscience
412
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
Linear adaptive filter:
A linear filter that self-adjusts its
transfer function and coefficients
over time, according to an
optimization rule. In the domain of
functional brain networks, adaptive
filters adapt to the time-varying
properties of neuronal signals.
Adaptive Filtering
Self-tuning optimized Kalman filter (STOK). To incorporate structural priors in dynamic func-
tional connectivity, we used a linear adaptive filter, the self-tuning optimized Kalman (STOK)
filter (Pascucci et al., 2020), as the base algorithm. STOK is a high-temporal-resolution and
noise-resilient filter for modeling time-varying multivariate autoregressive (tv-MVAR) processes
of the following form:
Yt ¼
X
p
k¼1 Ak;t Yt−k þ εt ; t 2 1; …; N
½
(cid:2);
(1)
where Y is a multitrial and multivariate set of d time series of dimension [trials × d] (e.g., activ-
ity signals from different brain regions), t refers to time samples (with N the total length of the
time segment considered), Ak,t are [d × d × p × N] matrices of autoregressive coefficients for
each lag k of a chosen model order p, and εt is zero-mean white noise with covariance matrix
(cid:2)ε (also called the innovation process). Equation 1 can be represented in the following state-
space form:
xt ¼ Φt−1xt−1 þ ωt−1;
zt ¼ Ht xt þ νt ;
(2)
(3)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
where Equation 2 represents the latent state xt (e.g., the MVAR process) as a random walk from
the previous state xt −1 [d × d × p], with transition matrix Φt −1 and uncorrelated zero-mean
noise ω. In Equation 3, the observed data zt are expressed as a linear combination of the latent
state xt and a projection matrix Ht, under white noise perturbation νt. The link with Equation 1
is established by recursively defining Ht from the past of the time series in Y (from t − 1 to the
model order p), and zt as the values of Y at present time t. This leads to the least squares esti-
mate:
^x
þð Þ
t
¼
^x
−ð Þ
t
þ c H~þ
1 þ c
t zt
;
(4)
þð Þ
t
is a weighted average of the previous state ^x
−ð Þ
in which the recursive update of ^x
t and a least
squares reconstruction from recent measurements H~þ
is the damped Moore-
Penrose pseudoinverse (+) of H, in which small singular values are attenuated to retain a pre-
specified portion of the variance (here we retained 0.99 of the variance for the rat EEG data
and 0.9 for the human EEG data). The variable c is a self-tuning adaptation constant that
automatically updates the speed of the filter depending on its residuals. The complete
derivation of the STOK can be found in Pascucci et al. (2020).
t zt. The matrix H~þ
Structural priors. The prior information is incorporated in the filter as a regularizing operator.
Recall that the update of ^x
in Equation 4—that is, the estimated matrix of tv-MVAR
coefficients—requires the ordinary least squares solution to H~þ
þð Þ
t
t zt,
bols
t
¼ H~þ
t zt ;
(5)
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
(dp × d ) representing the matrix of autoregressive coefficients that updates the pre-
with bols
t
vious state ^x
Tikhonov regularization, which admits the closed-form solution:
−ð Þ
t
. A principled way to incorporate priors in Equation 5 is the use of a generalized
(cid:2)
¼ x0;i←j þ H~T
b *
i←j;t
H~ þ Q−1
i←j
(cid:3)þ
(cid:4)
H~T
zi;t − H~ x0;i←j
(cid:5)
;
½
j 2 1; …; d
(cid:2);
(6)
Network Neuroscience
413
Structurally informed STOK
where x0 is the expected value of b, and Q−1 is the inverse covariance matrix, or precision
matrix, of x0. Q−1
i←j acts as a regularizing or Tikhonov matrix in Equation 6. The inclusion of
a regularizing matrix allows specifying penalties on the estimated coefficients. When the reg-
ularizing matrix is a multiple of the identity matrix, the solution corresponds to the classical L2
norm. When the main diagonal contains distinct elements, however, the solution penalizes the
coefficients differently, depending on the strength of the corresponding value in the regulariz-
ing matrix. This form of regularization offers a straightforward solution for incorporating struc-
tural priors in a tv-MVAR model. By solving Equation 6 for each channel separately (e.g., for
each signal in the multivariate time series), elements on the main diagonal of Q−1 can be used
to penalize the inflow from channel j to channel i (i ← j ) depending on structural priors; that is,
an a priori structure can be imposed on the contribution of all the channels to the activity
observed in each one.
Although this approach is deterministic, it has a natural Bayesian interpretation: Equation 6
is equivalent to expressing a prior belief on the functional connections bi←j entering each
channel, under a multivariate normal distribution N(bi←j; x0, Qi←j). This regularization scheme
requires the nontrivial conversion of SC to FC priors. A commonly employed strategy is to set
the prior expectations on x0 to zero (e.g., no functional connectivity; Sokolov et al., 2019;
Stephan et al., 2009), and to define the prior variance Qi←j based on the strength of SC. Under
the mild assumption of a positive and monotonic relationship between structure and function,
strong SC can be translated into large FC prior variance Q−1
i←j, corresponding to small regular-
ization values in Q−1
i←j that favor nonzero FC when supported by the data. Conversely, weak SC
can be translated into small prior variance, increasing the effect of regularization, and shrink-
ing the tv-MVAR coefficients toward their expected value of zero. Mapping SC to FC priors
also requires the scaling of SC to a range of suitable values, given that regularization acts
on the magnitude and scale of autoregressive coefficients. Normalized SC values are in the
0–1 range. For the sake of the present work, we scaled SC from 10−4 to 0.1, a range that pro-
duced clear effects of regularization for both rat and human EEG data, without an excessive
shrinking of all the coefficients. This range was chosen ad hoc, by varying the maximum of the
scaling range and selecting a value for which (a) the correlation between SC and FC increased,
and (b) the correlation between STOK (noninformed by SC) and si-STOK estimates would not
drastically decrease. Note that the choice of smaller scaling maxima reduces the regulariza-
tion effects and provides estimates that approximate the STOK results. The choice of a larger
scaling maxima increases the effect of regularization, introducing an excessive shrinkage of
the coefficients. An alternative, data-driven method to select the scaling maxima is presented
as Supplementary File 3.
Hence, the Tikhonov matrix Q−1
i←j in Equation 6 is a diagonal matrix whose nonzero ele-
ments represent the inverse prior variance for the functional connectivity between j input
sources and a receiver node i. By setting x0 equal to zero, we obtain the following:
(cid:2)
¼ H~T
bi←j;t*
H~ þ Q−1
i←j
(cid:3)þ
H~T
zi;t :
(7)
i←j in Equation 7 is equal to diag
(cid:3)
In a Bayesian view, the precision matrix Q−1
, where SCi←j
stands for the magnitude of structural connection between node j and node i. Hence, Q−1
i←j
determines the extent to which the posterior estimate (e.g., the functional connectivity
between two nodes) can deviate from its expected value of zero. As a result, the estimated
matrices of tv-MVAR coefficients combine information from both functional and structural
connectivity: Weak SC decreases the prior variance and increases regularization (e.g.,
1
SCi←j
(cid:2)
Network Neuroscience
414
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
expressing a strong belief that a functional connection is likely to be absent, to reduce false-
positive connections). Strong SC increases the prior variance and decreases regularization,
allowing functional connections to deviate from zero when supported by the functional data.
The new coefficients bt* obtained from Equation 7 (posteriors) are then substituted in
Equation 4, leading to the recursive update equation with structural priors:
^x
þð Þ
t
¼
^x
−ð Þ
t
þ cbt*
1 þ c
;
(8)
which ultimately provides an estimate of tv-MVAR coefficients A informed by the properties
of SC. The structural connectivity matrix can be either symmetric or asymmetric (e.g., for
unidirectional priors), binary or weighted, with values that increase as a function of the
expected strength of a connection between two nodes.
Time-Varying Directed Functional Connectivity
In the analysis of FC, we used a measure of directed connec-
Directed connectivity measure.
tivity derived from the estimated matrices of tv-MVAR coefficients. This measure, which we
termed magnitude of directed influence (MDI), is a function of time t and corresponds to
the magnitude of autoregressive coefficients a over lags (k = [1, 2, …, p]), quantifying the
time-domain unidirectional influence from a sender j to a target node i:
MDIi←j;t ¼
:
(9)
v
u
u
t
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
Xp
a2
i←j;k;t
k¼1
A similar measure, called direct causality (DC), has previously been used to estimate the
amount of direct causal influences in multivariate systems (Kamiński et al., 2001; Porcaro
et al., 2009).
Rat EEG functional connectivity. We tested the new algorithm and compared the performance
against the regular STOK using benchmark EEG data from epicranial recordings in rats. These
data provide a good benchmark for time-varying connectivity analysis because of the known
structural and functional connectivity patterns. After whisker stimulation, the evoked FC is
expected to follow the underlying SC, with the activity that propagates from the contralateral
primary somatosensory cortex to nearby parietal and frontal regions, at short latencies (5–25 ms
poststimulation). This pattern was extensively validated in previous work (Pagnotta & Plomp,
2018; Pascucci et al., 2020; Plomp et al., 2014).
As a proof of concept, we first verified the effect of adding structural priors with different
levels of sparsity. Two SC matrices were used as priors for the si-STOK, one obtained from the
original SC (see Rat structural connectome in the Methods section and Supplementary File 1),
the other obtained by applying proportional thresholding to the same SC, retaining only 25%
of the strongest connections, setting the remaining to zero and the diagonal (self-self connec-
tions) to one. The MDI metric was derived from tv-MVAR models with a model order of 4
(Pagnotta & Plomp, 2018; Pascucci et al., 2020). In the comparison, we focused on the out-
flow from the contralateral somatosensory cortex to the rest of the network, at the peak latency.
The peak latency was estimated from the results of the regular STOK as 13 ms poststimulus, in
line with previous reports (Pascucci et al., 2020; Plomp et al., 2014).
In a second step, we compared the performance of the two filters under noise perturbation. We
mixed the original data with white noise signals of the same size as the data and with an amplitude
corresponding to the 95th percentile of the data. We varied the mixing ratio between the original
Network Neuroscience
415
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
data and the noise signals in five levels (from 0 to 0.5, in steps of 0.1; 0 = 100% of the data and
0% of the noise). The noise at each level was regenerated 30 times. For each iteration, we
estimated (a) the difference between the outflow from the contralateral somatosensory cortex
and the average outflow of the network, at peak latency; and (b) a binary, directed adjacency
matrix preserving 50% of the strongest connections at peak latency. From the first measure, we
estimated the difference between the outflow from the primary somatosensory cortex and the
network average outflow, across animals—that is, the ability of the two algorithms to
discriminate the somatosensory cortex as the main driver under noise perturbations. From the
second measure, we derived a metric of consistency of the estimated networks under increasing
noise. The consistency was obtained as the proportion of binary connections in the adjacency
matrix that, for each level of noise larger than 0, were identical to those estimated in the absence
of noise—that is, the consistency of the estimated strongest connections under increasing noise.
For both measures, the zero-noise level was used as a baseline.
In the analysis of human EEG data, we evaluated
Human EEG directed functional connectivity.
the results of the STOK and si-STOK filter in estimating large-scale FC during face processing
(see Figure 3A). For both filters, we used a model order of 10, in line with values used before
(Pascucci et al., 2018; Pascucci et al., 2020). We first compared evoked activity after face and
scrambled stimuli at the scalp and source level (see Figure 3B). We selected a time window
centered on the scalp and the source N170 peak component (140–180 ms) as the latency of
interest for FC results. In the N170 window, we evaluated the estimated difference in network
MDI across conditions. For this difference, we compared the summed outflow obtained with
the two filters, from each node of the 68-area network (see Figure 3D). In a subsequent anal-
ysis, we focused on dynamic directed interactions among a subset of areas known to be
involved in face processing (e.g., regions of the core face network; Haxby et al., 2000). We
considered three areas, all in the right hemisphere: the fusiform gyrus (rFu), the inferior tem-
poral gyrus (rIT), and the superior temporal sulcus (rB; see Figure 4). We then compared
directed influences among each pair of areas in response to face or scrambled stimuli across
time. Significance was assessed using group-permutation statistics, as the proportion of group-
averaged MDI differences that were larger or smaller than the observed ones, after shuffling the
sign of the difference across participants 100,000 times. An uncorrected p value (p < 0.01) was
used as the threshold, with the exploratory aim to qualitatively compare the results with and
without the inclusion of SC priors.
ACKNOWLEDGMENTS
The authors thank Guru Prasad Padmasola and Charles Quairiaux for providing guidelines and
useful material for the analysis of the rat benchmark data, and Larry W. Swanson and Joel
Hahn for providing the rat structural connectivity dataset.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00218.
AUTHOR CONTRIBUTIONS
David Pascucci: Conceptualization; Data curation; Formal analysis; Investigation; Methodol-
ogy; Software; Validation; Visualization; Writing – original draft. Maria Rubega: Conceptuali-
zation; Investigation; Methodology; Software; Validation; Writing – review & editing. Joan
Rué-Queralt: Data curation; Methodology; Resources; Software; Writing – review & editing.
Network Neuroscience
416
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
Sebastien Tourbier: Formal analysis; Methodology; Resources; Software. Patric Hagmann:
Conceptualization; Funding acquisition; Methodology; Project administration; Resources;
Supervision; Writing – review & editing. Gijs Plomp: Conceptualization; Funding acquisition;
Investigation; Project administration; Resources; Supervision; Writing – original draft; Writing –
review & editing.
FUNDING INFORMATION
Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (https://dx
.doi.org/10.13039/501100001711), Award ID: PP00P1_183714. Schweizerischer National-
fonds zur Förderung der Wissenschaftlichen Forschung (https://dx.doi.org/10.13039
/501100001711), Award ID: PP00P1_190065. Schweizerischer Nationalfonds zur Förderung
der Wissenschaftlichen Forschung (https://dx.doi.org/10.13039/501100001711), Award ID:
PZ00P1_179988. Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen
Forschung (https://dx.doi.org/10.13039/501100001711), Award ID: CRSII5-170873.
REFERENCES
Ales, J. M., Farzin, F., Rossion, B., & Norcia, A. M. (2012). An
objective method for measuring face detection thresholds using
the sweep steady-state visual evoked response. Journal of Vision,
12(10). https://doi.org/10.1167/12.10.18, PubMed: 23024355
Antonacci, Y., Toppi, J., Mattia, D., Pietrabissa, A., & Astolfi, L.
(2019). Single-trial connectivity estimation through the least
absolute shrinkage and selection operator. 2019 41st Annual
International Conference of the IEEE Engineering in Medicine
and Biology Society (EMBC), 6422–6425. https://doi.org/10
.1109/EMBC.2019.8857909, PubMed: 31947312
Anzolin, A., Presti, P., Van De Steen, F., Astolfi, L., Haufe, S., &
Marinazzo, D. (2019). Quantifying the effect of demixing
approaches on directed connectivity estimated between recon-
structed EEG sources. Brain Topography, 32(4), 655–674.
https://doi.org/10.1007/s10548-019-00705-z, PubMed:
30972604
Betzel, R. F., Griffa, A., Hagmann, P., & Mišić, B.
(2019).
Distance-dependent consensus thresholds for generating group-
representative structural brain networks. Network Neuroscience,
3(2), 475–496. https://doi.org/10.1162/netn_a_00075, PubMed:
30984903
Bota, M., Sporns, O., & Swanson, L. W. (2015). Architecture of the
cerebral cortical association connectome underlying cognition.
Proceedings of the National Academy of Sciences, 112(16),
E2093–2101. https://doi.org/10.1073/pnas.1504394112,
PubMed: 25848037
Brunet, D., Murray, M. M., & Michel, C. M. (2011). Spatiotemporal
analysis of multichannel EEG: CARTOOL. Computational Intelli-
gence and Neuroscience, 2011, 2. https://doi.org/10.1155/2011
/813870, PubMed: 21253358
Crimi, A., Dodero, L., Murino, V., & Sona, D. (2016). Effective brain
connectivity through a constrained autoregressive model. Inter-
national Conference on Medical Image Computing and
Computer-Assisted Intervention, 140–147. https://doi.org/10
.1007/978-3-319-46720-7_17
Crimi, A., Dodero, L., Sambataro, F., Murino, V., & Sona, D. (2021).
Structurally constrained effective brain connectivity. Neuro-
Image, 239, 118288. https://doi.org/10.1016/j.neuroimage.2021
.118288, PubMed: 34147631
Damoiseaux, J. S., & Greicius, M. D. (2009). Greater than the sum
of its parts: A review of studies combining structural connectivity
and resting-state functional connectivity. Brain Structure and
Function, 213(6), 525–533. https://doi.org/10.1007/s00429-009
-0208-6, PubMed: 19565262
Deco, G., Ponce-Alvarez, A., Mantini, D., Romani, G. L., Hagmann,
P., & Corbetta, M. (2013). Resting-state functional connectivity
emerges from structurally and dynamically shaped slow linear
fluctuations. Journal of Neuroscience, 33(27), 11239–11252.
https://doi.org/10.1523/ JNEUROSCI.1091-13.2013, PubMed:
23825427
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson,
B. C., Blacker, D., Buckner, R. L., Dale, A. M., Maguire,
R. P., & Hyman, B. T. (2006). An automated labeling system
for subdividing the human cerebral cortex on MRI scans into
gyral based regions of interest. NeuroImage, 31(3), 968–980.
https://doi.org/10.1016/j.neuroimage.2006.01.021, PubMed:
16530430
Eichenbaum, A., Pappas, I., Lurie, D., Cohen, J. R., & D’Esposito,
M. (2021). Differential contributions of static and time-varying
functional connectivity to human behavior. Network Neurosci-
ence, 5(1), 145–165. https://doi.org/10.1162/netn_a_00172,
PubMed: 33688610
Filatova, O. G., Yang, Y., Dewald, J. P. A., Tian, R., Maceira-Elvira,
P., Takeda, Y., Kwakkel, G., Yamashita, O., & van der Helm,
F. C. T. (2018). Dynamic information flow based on EEG and
diffusion MRI in stroke: A proof-of-principle study. Frontiers in
Neural Circuits, 12, 79. https://doi.org/10.3389/fncir.2018
.00079, PubMed: 30327592
Fox, C. J., Iaria, G., & Barton, J. J. S. (2009). Defining the face pro-
cessing network: Optimization of the functional localizer in
Network Neuroscience
417
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
fMRI. Human Brain Mapping, 30(5), 1637–1651. https://doi.org
/10.1002/hbm.20630, PubMed: 18661501
Garrison, K. A., Scheinost, D., Finn, E. S., Shen, X., & Constable,
R. T. (2015). The (in)stability of functional brain network mea-
sures across thresholds. NeuroImage, 118, 651–661. https://doi
.org/10.1016/j.neuroimage.2015.05.046, PubMed: 26021218
Goffaux, V., Peters, J., Haubrechts, J., Schiltz, C., Jansma, B., &
Goebel, R. (2011). From coarse to fine? Spatial and temporal
dynamics of cortical face processing. Cerebral Cortex, 21(2),
467–476. https://doi.org/10.1093/cercor/ bhq112, PubMed:
20576927
Granger, C. W. (1969). Investigating causal relations by economet-
ric models and cross-spectral methods. Econometrica: Journal of
the Econometric Society, 424–438. https://doi.org/10.2307
/1912791
Hagmann, P., Cammoun, L., Gigandet, X., Meuli, R., Honey, C. J.,
Wedeen, V. J., & Sporns, O. (2008). Mapping the structural core
of human cerebral cortex. PLoS Biology, 6(7), e159. https://doi
.org/10.1371/journal.pbio.0060159, PubMed: 18597554
Haufe, S., Nikulin, V. V., Müller, K.-R., & Nolte, G. (2013). A critical
assessment of connectivity measures for EEG data: A simulation
study. NeuroImage, 64, 120–133. https://doi.org/10.1016/j
.neuroimage.2012.09.036, PubMed: 23006806
Haxby, J. V., Hoffman, E. A., & Gobbini, M. I. (2000). The distrib-
uted human neural system for face perception. Trends in Cogni-
tive Sciences, 4(6), 223–233. https://doi.org/10.1016/S1364
-6613(00)01482-0, PubMed: 10827445
Honey, C. J., Sporns, O., Cammoun, L., Gigandet, X., Thiran, J. P.,
Meuli, R., & Hagmann, P. (2009). Predicting human resting-state
functional connectivity from structural connectivity. Proceedings
of the National Academy of Sciences, 106(6), 2035–2040. https://
doi.org/10.1073/pnas.0811168106, PubMed: 19188601
Honey, C. J., Thivierge, J.-P., & Sporns, O. (2010). Can structure
predict
function in the human brain? NeuroImage, 52(3),
766–776. https://doi.org/10.1016/j.neuroimage.2010.01.071,
PubMed: 20116438
Jirsa, V., Sporns, O., Breakspear, M., Deco, G., & McIntosh, A. R.
(2010). Towards the virtual brain: Network modeling of the intact
and the damaged brain. Archives Italiennes de Biologie, 148(3),
189–205. https://doi.org/10.4449/aib.v148i3.1223, PubMed:
21175008
Kalman, R. E. (1960). A new approach to linear filtering and predic-
tion problems. Journal of Basic Engineering, 82(1), 35–45. https://
doi.org/10.1115/1.3662552
Kamiński, M., Ding, M., Truccolo, W. A., & Bressler, S. L. (2001).
Evaluating causal relations in neural systems: Granger causality,
directed transfer function and statistical assessment of signifi-
cance. Biological Cybernetics, 85(2), 145–157. https://doi.org
/10.1007/s004220000235, PubMed: 11508777
Lei, X., Wu, T., & Valdes-Sosa, P. (2015). Incorporating priors for
EEG source imaging and connectivity analysis. Frontiers in Neu-
roscience, 9, 284. https://doi.org/10.3389/fnins.2015.00284,
PubMed: 26347599
Lim, S., Radicchi, F., van den Heuvel, M. P., & Sporns, O. (2019).
Discordant attributes of structural and functional brain connec-
tivity in a two-layer multiplex network. Scientific Reports, 9(1),
2885. https://doi.org/10.1038/s41598-019-39243-w, PubMed:
30814615
Maier-Hein, K. H., Neher, P. F., Houde, J.-C., Côté, M.-A., Garyfallidis,
E., Zhong, J., Chamberland, M., Yeh, F.-C., Lin, Y.-C., Ji, Q., Reddick,
W. E., Glass, J. O., Chen, D. Q., Feng, Y., Gao, C., Wu, Y., Ma, J.,
Renjie, H., Li, Q., … Descoteaux, M. (2017). The challenge of
mapping the human connectome based on diffusion tractography.
Nature Communications, 8(1), 1349. https://doi.org/10.1038
/s41467-017-01285-x, PubMed: 29116093
Markov, N., Ercsey-Ravasz, M. M., Ribeiro Gomes, A. R., Lamy, C.,
Magrou, L., Vezoli, J., Misery, P., Falchier, A., Quilodran, R., &
Gariel, M. A. (2012). A weighted and directed interareal connec-
tivity matrix for macaque cerebral cortex. Cerebral Cortex, 24(1),
17–36. https://doi.org/10.1093/cercor/ bhs270, PubMed:
23010748
Milde, T., Leistritz, L., Astolfi, L., Miltner, W. H., Weiss, T., Babiloni,
F., & Witte, H. (2010). A new Kalman filter approach for the esti-
mation of high-dimensional time-variant multivariate AR models
and its application in analysis of laser-evoked brain potentials.
NeuroImage, 50(3), 960–969. https://doi.org/10.1016/j
.neuroimage.2009.12.110, PubMed: 20060483
Mišić, B., Betzel, R. F., de Reus, M. A., van den Heuvel, M. P.,
Berman, M. G., McIntosh, A. R., & Sporns, O. (2016). Network-
level structure-function relationships in human neocortex. Cere-
bral Cortex, 26(7), 3285–3296. https://doi.org/10.1093/cercor
/bhw089, PubMed: 27102654
Pagnotta, M. F., Dhamala, M., & Plomp, G. (2018). Benchmarking
nonparametric Granger causality: Robustness against downsam-
pling and influence of spectral decomposition parameters. Neuro-
Image, 183, 478–494. https://doi.org/10.1016/j.neuroimage.2018
.07.046, PubMed: 30036586
Pagnotta, M. F., & Plomp, G. (2018). Time-varying MVAR algo-
rithms for directed connectivity analysis: Critical comparison in
simulations and benchmark EEG data. PLoS ONE, 13(6),
e0198846. https://doi.org/10.1371/journal.pone.0198846,
PubMed: 29889883
Pagnotta, M. F., Plomp, G., & Pascucci, D. (2019). A regularized
and smoothed General Linear Kalman Filter for more accurate
estimation of time-varying directed connectivity. 2019 41st
Annual International Conference of the IEEE Engineering in Med-
icine and Biology Society (EMBC), 611–615. https://doi.org/10
.1109/EMBC.2019.8857915, PubMed: 31945972
Pascucci, D., Hervais-Adelman, A., & Plomp, G. (2018). Gating by
induced α-γ asynchrony in selective attention. Human Brain
Mapping, 39(10), 3854–3870. https://doi.org/10.1002/ hbm
.24216, PubMed: 29797747
Pascucci, D., & Rubega, M. (2019). DyNet toolbox, GitHub, https://
github.com/PscDavid/dynet_toolbox
Pascucci, D., Rubega, M., & Plomp, G. (2020). Modeling
time-varying brain networks with a self-tuning optimized Kalman
filter. PLoS Computational Biology, 16(8), e1007566. https://doi
.org/10.1371/journal.pcbi.1007566, PubMed: 32804971
Pascucci, D., Tourbier, S., Rue-Queralt, J., Carboni, M., Hagmann,
P., & Plomp, G. (2021). Source imaging of high-density visual
evoked potentials with multi-scale brain parcellations and
connectomes. bioRxiv. https://doi.org/10.1101/2021.03.16
.435599
Peirce, J. W. (2008). Generating stimuli for neuroscience using Psy-
choPy. Frontiers in Neuroinformatics, 2. https://doi.org/10.3389
/neuro.11.010.2008, PubMed: 19198666
Network Neuroscience
418
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Structurally informed STOK
Petersen, S. E., & Sporns, O. (2015). Brain networks and cognitive
architectures. Neuron, 88(1), 207–219. https://doi.org/10.1016/j
.neuron.2015.09.027, PubMed: 26447582
Plato, R., & Vainikko, G. (1990). On the regularization of projection
methods for solving III-posed problems. Numerische Mathema-
tik, 57(1), 63–79. https://doi.org/10.1007/BF01386397
Plomp, G., Quairiaux, C., Michel, C. M., & Astolfi, L. (2014). The
physiological plausibility of time-varying Granger-causal model-
ing: Normalization and weighting by spectral power. Neuro-
Image, 97, 206–216. https://doi.org/10.1016/j.neuroimage.2014
.04.016, PubMed: 24736179
Porcaro, C., Zappasodi, F., Rossini, P. M., & Tecchio, F. (2009).
Choice of multivariate autoregressive model order affecting real
network functional connectivity estimate. Clinical Neurophysiol-
ogy, 120(2), 436–448. https://doi.org/10.1016/j.clinph.2008.11
.011, PubMed: 19109060
Quairiaux, C., Mégevand, P., Kiss, J. Z., & Michel, C. M. (2011).
Functional development of large-scale sensorimotor cortical net-
works in the brain. Journal of Neuroscience, 31(26), 9574–9584.
https://doi.org/10.1523/ JNEUROSCI.5995-10.2011, PubMed:
21715622
Reid, A. T., Headley, D. B., Mill, R. D., Sanchez-Romero, R.,
Uddin, L. Q., Marinazzo, D., Lurie, D. J., Valdés-Sosa, P. A.,
Hanson, S. J., & Biswal, B. B. (2019). Advancing functional con-
nectivity research from association to causation. Nature Neuro-
science, 22, 1751–1760. https://doi.org/10.1038/s41593-019
-0510-4, PubMed: 31611705
Rubega, M., Carboni, M., Seeber, M., Pascucci, D., Tourbier, S.,
Toscano, G., Van Mierlo, P., Hagmann, P., Plomp, G., Vulliemoz,
S., & Michel, C. M. (2019). Estimating EEG source dipole orien-
tation based on singular-value decomposition for connectivity
analysis. Brain Topography, 32(4), 704–719. https://doi.org/10
.1007/s10548-018-0691-2, PubMed: 30511174
Rué-Queralt, J. (2019). PyDyNet, GitHub, https://github.com
/joanrue/pydynet
Saygin, Z. M., Osher, D. E., Koldewyn, K., Reynolds, G., Gabrieli,
J. D. E, & Saxe, R. R. (2011). Anatomical connectivity patterns predict
face-selectivity in the fusiform gyrus. Nature Neuroscience, 15(2),
321–327. https://doi.org/10.1038/nn.3001, PubMed: 22197830
Skudlarski, P., Jagannathan, K., Calhoun, V. D., Hampson, M.,
Skudlarska, B. A., & Pearlson, G. (2008). Measuring brain connec-
tivity: Diffusion tensor imaging validates resting state temporal
correlations. NeuroImage, 43(3), 554–561. https://doi.org/10
.1016/j.neuroimage.2008.07.063, PubMed: 18771736
Sokolov, A. A., Zeidman, P., Erb, M., Ryvlin, P., Pavlova, M. A., &
Friston, K. J. (2019). Linking structural and effective brain connec-
tivity: Structurally informed Parametric Empirical Bayes (si-PEB).
Brain Structure and Function, 224(1), 205–217. https://doi.org/10
.1007/s00429-018-1760-8, PubMed: 30302538
Sporns, O. (2010). Networks of the brain. MIT Press. https://doi.org
/10.7551/mitpress/8476.001.0001
Stephan, K. E., Tittgemeyer, M., Knösche, T. R., Moran, R. J., &
Friston, K. J. (2009). Tractography-based priors for dynamic
causal models. NeuroImage, 47(4), 1628–1638. https://doi.org
/10.1016/j.neuroimage.2009.05.096, PubMed: 19523523
Suárez, L. E., Markello, R. D., Betzel, R. F., & Misic, B. (2020). Link-
ing structure and function in macroscale brain networks. Trends
in Cognitive Sciences, 24(4), 302–315. https://doi.org/10.1016/j
.tics.2020.01.008, PubMed: 32160567
Sugase-Miyamoto, Y., Matsumoto, N., & Kawano, K. (2011). Role of
temporal processing stages by inferior temporal neurons in facial
recognition. Frontiers in Psychology, 2. https://doi.org/10.3389
/fpsyg.2011.00141, PubMed: 21734904
Swanson, L. W., Hahn, J. D., & Sporns, O. (2017). Organizing prin-
ciples for the cerebral cortex network of commissural and asso-
ciation connections. Proceedings of the National Academy of
Sciences, 114(45), E9692–E9701. https://doi.org/10.1073/pnas
.1712928114, PubMed: 29078382
Takeda, Y., Suzuki, K., Kawato, M., & Yamashita, O. (2019). MEG
source imaging and group analysis using VBMEG. Frontiers in
Neuroscience, 13. https://doi.org/10.3389/fnins.2019.00241,
PubMed: 30967756
Tournier, J.-D., Smith, R., Raffelt, D., Tabbara, R., Dhollander, T.,
Pietsch, M., Christiaens, D., Jeurissen, B., Yeh, C.-H., & Connelly,
A. (2019). MRtrix3: A fast, flexible and open software framework
for medical image processing and visualisation. NeuroImage,
202, 116137. https://doi.org/10.1016/j.neuroimage.2019
.116137, PubMed: 31473352
Tovée, M. J. (1995). Face recognition: What are faces for? Current
Biology, 5(5), 480–482. https://doi.org/10.1016/S0960-9822(95)
00096-0
Valdés-Sosa, P. A., Sánchez-Bornot, J. M., Lage-Castellanos, A.,
Vega-Hernández, M., Bosch-Bayard, J., Melie-García, L., &
Canales-Rodríguez, E. (2005). Estimating brain functional con-
nectivity with sparse multivariate autoregression. Philosophical
Transactions of the Royal Society of London B: Biological Sci-
ences, 360(1457), 969–981. https://doi.org/10.1098/rstb.2005
.1654, PubMed: 16087441
Vázquez-Rodríguez, B., Suárez, L. E., Markello, R. D., Shafiei, G.,
Paquola, C., Hagmann, P., van den Heuvel, M. P., Bernhardt,
B. C., Spreng, R. N., & Misic, B. (2019). Gradients of structure–
function tethering across neocortex. Proceedings of the National
Academy of Sciences, 116(42), 21219–21227. https://doi.org/10
.1073/pnas.1903403116, PubMed: 31570622
Network Neuroscience
419
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
6
2
4
0
1
2
0
2
8
1
2
1
n
e
n
_
a
_
0
0
2
1
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3