Learning Increases the Survival of Newborn
Neurons Provided That Learning Is Difficult
to Achieve and Successful
Daniel M. Curlik II and Tracey J. Shors
D
o
w
N
l
o
UN
D
e
D
Astratto
■ Learning increases neurogenesis by increasing the survival of
new cells generated in the adult hippocampal formation [Shors,
T. J. Saving new brain cells. Scientific American, 300, 46–52,
2009]. Tuttavia, only some types of learning are effective. Re-
cent studies demonstrate that animals that learn the conditioned
risposta (CR) but require more trials to do so retain more new
neurons than animals that quickly acquire the CR or that fail to
acquire the CR. In these studies, task parameters were altered to
modify the number of trials required to learn a CR. Here, we
asked whether pharmacological manipulations that prevent or
facilitate learning would decrease or increase, rispettivamente,
the number of cells that remain in the hippocampus after train-
ing. To answer this question, we first prevented learning with
the competitive N-methyl-D-aspartate (NMDA) receptor antago-
nist (RS)-3-(2-carboxypiperazin-4-yl) propyl-1-phosphonic acid.
As a consequence, training did not increase cell survival. Secondo,
we facilitated learning with the cognitive enhancer D-cycloserine,
which increases NMDA receptor activity via its actions at the
glycine binding site. Administration of D-cycloserine each day
before training increased the number of learned responses and
the number of cells that survived. All animals that learned the
CR retained more of the new cells, but those that learned very
quickly retained fewer than those that required more training
trials to learn. Together, these results demonstrate that NMDA
receptor activation modifies learning and as a consequence alters
the number of surviving cells in the adult hippocampus. ■
INTRODUCTION
The dentate gyrus of the hippocampal formation generates
new granule neurons throughout life (Kornack & Rakic,
1999; Eriksson et al., 1998; Kaplan & Hinds, 1977; Altman
& Das, 1965). It is estimated that the adult dentate gyrus
generates 5000–10,000 new cells per day (Cameron &
McKay, 2001), the vast majority of which differentiate into
neurons (Hastings & Gould, 1999; Markakis & Gage, 1999;
Cameron, Woolley, McEwen, & Gould, 1993). Tuttavia,
over half of these cells die within a few weeks of their birth
(Gould, Beylin, Tanapat, Reeves, & Shors, 1999). One week
after they are born, these neurons display a remarkable sen-
sitivity to various forms of learning that can prevent their
death, thereby increasing the number of new neurons that
survive in the adult brain (Shors, 2009; Waddell & Shors,
2008; Leuner et al., 2004; Dayer, Ford, Cleaver, Yassaee, &
Cameron, 2003; Ambrogini et al., 2000; Gould et al., 1999).
The first study to report that learning increases neuronal
survival indicated that tasks that depend on the hippocam-
pus are the most effective (Gould et al., 1999). Since then,
it has been determined that hippocampal dependence,
per se, is not necessary to increase the number of surviving
cells (reviewed in Shors, 2008). Per esempio, animals that
were trained with a hippocampal-independent form of
Rutgers University, Piscataway, NJ
trace conditioning, known as contiguous trace condi-
tioning, possessed more new neurons after learning than
naive controls (Dalla, Bangasser, Edgecomb, & Shors,
2007). That said, the most effective tasks tend to depend
on the hippocampus for learning.
Generalmente, tasks that are dependent on the hippocampus
tend to be more difficult to learn than their non-hippocampal-
dependent counterparts. Perhaps task difficulty is the crit-
ical factor. Infatti, this appears to be the case. Animals that
Imparare, but do so after more training trials, rescue more new-
born neurons than animals that quickly acquire the same
learned response ( Waddell & Shors, 2008; Dalla et al.,
2007). Inoltre, apprendimento, not merely training, is likewise
critical (Anderson, Sisti, Curlik, & Shors, 2010). Così, quelli
animals that are trained but do not learn do not retain any
more cells than naive animals. Overall, these results indicate
that animals that fail to acquire a learned response or do so
very rapidly will have fewer surviving newborn neurons
than animals that learn but require many trials of training
to do so. These conclusions were drawn from studies in
which task parameters were manipulated to alter acquisi-
tion and asymptotic performance. If these conclusions can
be of general significance, then they should apply to other
learning situations, including those in which learning is
altered pharmacologically. In the current set of studies
we pharmacologically manipulated acquisition of trace
eyeblink conditioning because learning this task reliably
© 2011 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 23:9, pag. 2159–2170
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
3
l
9
e
–
2
P
1
D
5
F
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
C
9
N
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
P
C
D
N
.
B
sì
2
0
G
1
tu
0
e
.
S
T
2
o
1
N
5
9
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
T
/
.
F
.
o
N
1
8
M
UN
sì
2
0
2
1
increases cell survival. In the first experiment, we pre-
vented learning with a competitive N-methyl-D-aspartate
(NMDA) receptor antagonist, (RS)-3-(2-carboxypiperazin-
4-yl) propyl-1-phosphonic acid (CPP). In the second ex-
periment, we enhanced learning with the partial glycine
receptor agonist D-cycloserine (DCS). The effects of these
agents on learning and the consequent effects on the
number of surviving cells were determined.
METHODS
General Methods for All Experiments
Subjects
Adult male Sprague–Dawley rats ranging between 60 E
90 days of age were individually housed and given access to
food and water ad libitum. The animals were maintained on
a 12-hr light–dark cycle, with the lights turning on at 7:00 a.m.
Rats were handled by the experimenter at least 1 week
before surgery. The experiments were designed to fully
comply with the rules and regulations set forth by the PHS
Policy on Humane Care and Use of Laboratory Animals and
the Guide for the Care and Use of Laboratory Animals.
Surgical Procedures
Animals were anesthetized with sodium pentobarbital
(15 mg /kg), which was supplemented with isoflurane
gas. They then underwent stereotaxic surgery for the im-
plantation of four periorbital electrodes, which were
placed in the muscle of the right eyelid. The electrodes
were composed of a stainless steel wire (0.005 In. diameter)
that was inserted through the eyelid, under the skin of the
scalp, to a headstage that was mounted on the skull. Dental
cement secured the headstage to four small screws that
were partially embedded within the skull (Servatius &
Shors, 1996). All animals were given a minimum recovery
period of 5 days before receiving one single intraperito-
neal injection (200 mg/kg) of bromodeoxyuridine (BrdU),
a thymidine analog that labels cells in the S-phase of the
cell cycle (Cameron & McKay, 2001; Mugnaio & Nowakowski,
1988).
Classical Eyeblink Conditioning
Six days after the BrdU injection, animals were acclimated
to the conditioning chambers for 1 hr while spontaneous
blinking activity was recorded. The next day (1 week after
the BrdU injection), groups of animals began training with
either paired trace or explicitly unpaired eyeblink condi-
tioning. Training continued each day for 4 days (for the ex-
perimental time line, see Figure 1A). During each day of
training, the animals were injected with either the drug
or the physiological saline before or after the training ses-
sion. All animals were trained with 200 trials per day. Each
trial of trace conditioning consisted of an 82-dB white noise
conditioned stimulus (CS), followed by a 500-msec trace in-
terval, which was followed by a 100-msec unconditioned
stimulus ( US) Di 0.65 mA of periorbital stimulation to
the right eyelid. The intertrial interval was 25 ± 5 sec.
Explicitly unpaired training consisted of the same num-
ber of CS and US presentations as trace conditioning; how-
ever, the CS and the US were presented in an explicitly
unpaired manner.
The occurrence of an eyeblink was determined from
EMG recording of the right eyelid muscle. During trace
conditioning a baseline recording was taken 250 msec be-
fore every trial. A conditioned response (CR) was counted
on that trial if a response occurred during the 500-msec
trace interval and if that response had an amplitude greater
than the maximum amplitude from the baseline recording
for that trial plus four times the baseline recordingʼs standard
deviation. A similar baseline recording was taken 250 msec
before the onset of the CS during unpaired training. CRs dur-
ing unpaired training were measured as responses that
occurred up to 500 msec after the offset of the CS. Trials
were divided into blocks of 100 consecutive trials. The first
100 trials were also examined in blocks of 20 trials. IL
number of trials required for an animal to reach a behav-
ioral criterion of 60% CRs during any block of 100 trials was
determined. Animals that reached this criterion were consid-
ered to have successfully learned the CR (Waddell & Shors,
2008; Dalla et al., 2007; Moyer, Thompson, & Disterhoft,
2000). Inoltre, we examined another common measure
of acquisition of the CR: the number of trials an animal
required to emit a CR on eight of nine consecutive trials
during any one block of 100 trials ( Dalla, Papachristos,
Whetstone, & Shors, 2009; Nokia, Penttonen, Korhonen,
& Wikgren, 2008).
Immunohistochemistry for BrdU
Twenty-one days after the BrdU injection, all animals
were deeply anesthetized with sodium pentobarbital
(100 mg/kg) and transcardially perfused with 4% parafor-
maldehyde. Brains were extracted and postfixed in 4%
paraformaldehyde for a minimum of 48 hr before being
sectioned. Coronal sections (40 μM) were cut through the
entire rostral-caudal extent of the dentate gyrus of one
hemisphere with an oscillating tissue slicer. UN 1:12 series
of slices was mounted onto glass slides for BrdU immuno-
histochemistry. To stain for the presence of BrdU, these
slices were heated in 0.1 M citric acid (pH 6.0), incubated
in trypsin followed by 2N HCl, and then incubated over-
night in primary antimouse BrdU (1:200; Becton Dickson,
Franklin Lakes, NJ) E 0.5% Tween 20. The next day, IL
tissue was incubated in biotinylated antimouse antibody
(1:200; Vector Laboratories, Burlingame, CA), followed
by avidin–biotin–horseradish peroxidase (1:100; Vector
Laboratories). The slices were then placed in diamino-
benzidine, counterstained with cresyl violet, and cover-
slipped. All slides were coded so that the experimenter
was blind to the experimental condition while they counted
2160
Journal of Cognitive Neuroscience
Volume 23, Numero 9
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
3
l
9
e
–
2
P
1
D
5
F
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
C
9
N
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
P
C
D
N
.
B
sì
2
0
G
1
tu
0
e
.
S
T
2
o
1
N
5
9
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
F
T
.
/
.
o
N
1
8
M
UN
sì
2
0
2
1
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
3
l
9
e
–
2
P
1
D
5
F
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
C
9
N
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
P
C
D
N
.
B
sì
2
0
G
1
tu
0
e
.
S
T
2
o
1
N
5
9
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
T
.
.
/
F
.
o
N
1
8
M
UN
sì
2
0
2
1
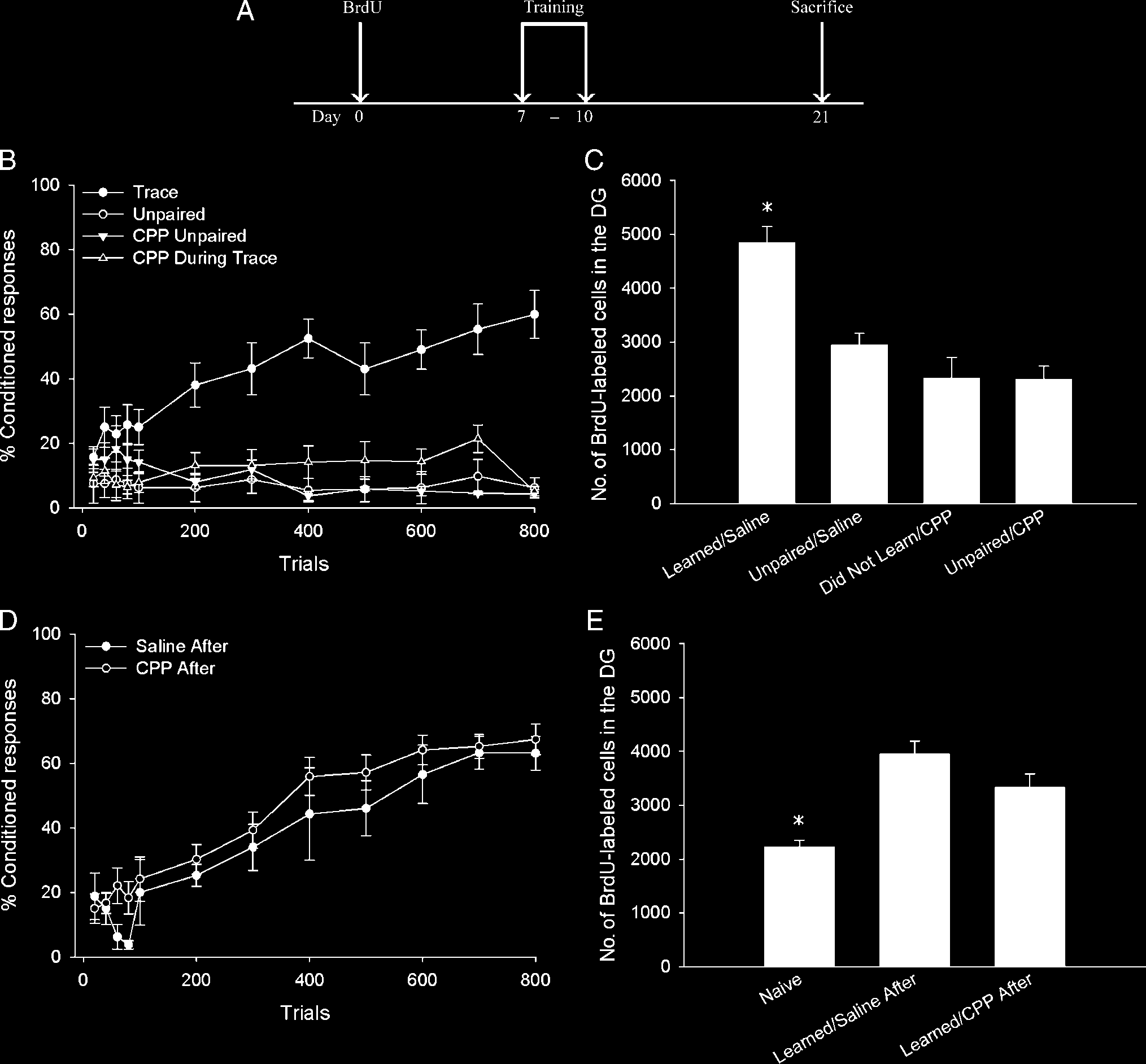
Figura 1. (UN) A diagram depicting the time line of both experiments. Animals underwent stereotaxic surgery at least 5 days before the BrdU
injection. CPP, DCS, or saline was administered during the 4 days of training (days 7–10 post-BrdU). (B) Administration of the NMDA receptor
antagonist CPP during trace eyeblink conditioning (CPP During Trace) prevented acquisition of the CR. CPP did not alter responding to unpaired
stimuli (CPP Unpaired). Additional animals received saline during training with trace eyeblink conditioning (Trace) or explicitly unpaired stimuli
(Unpaired). (C) Animals that received saline during training with trace conditioning that reached criterion ( Learned/Saline) retained more
BrdU-labeled cells than animals that received CPP during trace conditioning, none of which reached criterion ( Did Not Learn/CPP). The saline-
treated animals that learned retained more new cells than animals that received saline during unpaired training ( Unpaired/Saline) and CPP
during unpaired training (Unpaired/CPP). The number of BrdU-labeled cells in animals that received CPP during trace conditioning was similar
to the number of cells in animals that received saline or CPP during unpaired training. (D) CPP did not alter acquisition or asymptotic
performance of trace eyeblink conditioning when it was administered 6 hr after training. (E) CPP did not alter the number of BrdU-labeled cells
when administered 6 hr after training, as animals that received saline or CPP after training that learned retained more newborn cells than
experimentally naive animals.
the number of BrdU-labeled cells present in the entire
dentate gyrus of each slice. To accurately estimate the
number of BrdU-labeled cells throughout both hemi-
spheres of the entire dentate gyrus of one animal, IL
number of BrdU-labeled cells in the slices from that
animal was multiplied by 24.
Statistical Analysis
Performance during eyeblink conditioning was analyzed
with repeated measures ANOVA, with trial blocks as the
repeated measure and number of CRs as the dependent
measure. Post hoc comparisons were made between
Curlik and Shors
2161
groups using Tukeyʼs HSD test. The number of BrdU-labeled
cell was analyzed with one-way and two-way ANOVAs.
Independent samples t tests and Mann–Whitney U tests
were performed as needed.
Experiment 1 Methods: Does the NMDA
Receptor-dependent Blockade of Learning
Prevent the Increase in Cell Survival?
The competitive NMDA receptor antagonist CPP (Tocris
Bioscience) was dissolved in 0.9% saline and injected in-
traperitoneally (10 mg/ kg) each day, 1 hr before training.
Previous research has demonstrated that pretraining ad-
ministration of this dose of CPP prevents acquisition of trace
eyeblink conditioning (Leuner, Falduto, & Shors, 2003). IL
experiment consisted of seven groups. Group 1 (n = 7)
received injections of CPP each day, 1 hr before training
with trace eyeblink conditioning (7–10 days after the BrdU
injection). Group 2 (n = 7) received saline injections at the
same time before trace conditioning. Group 3 (n = 6)
received injections of CPP 1 hr before explicitly unpaired
training, whereas Group 4 (n = 4) received saline injec-
zioni 1 hr before unpaired training. Comparison of these
first four groups (Groups 1–4) allowed us to determine
whether pretraining administration of CPP prevented
trace eyeblink conditioning and the subsequent increase
in cell survival that is normally seen after learning. Com-
parison of the BrdU-labeled cell counts in the two groups
trained with unpaired stimuli (Groups 3 E 4) revealed
whether CPP administration itself, irrespective of its effects
on learning, increased the number of surviving cells.
Group 5 (n = 12) received injections of CPP every day,
6 hr after trace conditioning, when CPP administration did
not interfere with acquisition of the CR. Group 6 (n = 4)
received saline injections every day, 6 hr after trace condi-
tioning. The remaining group, Group 7 (n = 4), received
daily saline injections at the same time as Groups 5 E 6;
Tuttavia, this group was not trained with any form of eye-
blink conditioning. The cell counts from Groups 5 A 7 were
used to determine whether CPP decreased cell survival irre-
spective of its effects on learning.
Experiment 2 Methods: Does the NMDA Receptor
Modulator DCS Facilitate Learning and Thereby
Increase the Number of Cells That Survive?
DCS (Sigma-Aldrich, Atlanta, GA), a partial agonist of the
strychnine-insensitive glycine binding site, was used to
facilitate acquisition of the CR during trace eyeblink con-
ditioning. The experiment consisted of five groups. Group
1 (n = 7) received intraperitoneal injections of DCS dis-
solved in 0.9% saline (15 mg/ kg) 30 min before each
session of trace conditioning. This dose of DCS has pre-
viously been shown to facilitate learning during trace
eyeblink conditioning (Waddell, Mallimo, & Shors, 2010;
Thompson & Disterhoft, 1997; Thompson, Moskal, &
Disterhoft, 1992). Group 2 (n = 9) received daily injec-
tions of saline 30 min before trace conditioning. Compari-
sons between these two groups allowed us to determine
whether DCS facilitated acquisition of the trace eyeblink re-
sponse, and if so, if that facilitated learning resulted in an
increase in the number of surviving cells. Two additional
groups (Groups 3 E 4) received DCS (n = 7) or saline
(n = 4) 30 min before training with explicitly unpaired
stimuli. These two groups were examined to determine
whether administration of DCS itself might increase cell
survival, irrespective of its effects on learning. A fifth group
of animals (n = 5) received daily injections of DCS 6 hr
after trace conditioning, at a time point when administra-
tion of DCS did not alter acquisition of the CR. A compari-
son of the number of BrdU-labeled cells between this
group and Group 1, which received saline before trace
condizionata, allowed us to determine whether DCS de-
creased cell survival, irrespective of its effects on learning.
RESULTS
Experiment 1 Results: Administration of an NMDA
Receptor Antagonist before but not after Training
Prevents Learning and the Enhancement in Cell
Survival Seen after Learning
Administration of the NMDA receptor antagonist CPP be-
fore training each day completely prevented acquisition of
the CR. A repeated measures ANOVA using the drug con-
dizione (CPP vs. saline) and the training protocol (paired vs.
unpaired) as the independent measures, the trial blocks
(100 trial blocks, two blocks per day) as the repeated mea-
sure, and the percentage of CRs as the dependent measure
revealed a significant interaction between the drug condition
and the training protocol, F(1, 20) = 38.71, P < .01, indicating
that CPP only prevented conditioned responding when it was
administered before trace but not unpaired conditioning.
Furthermore, a significant interaction was found between
training blocks and drug condition, F(7, 140) = 2.40, p <
.05, revealing that CPP prevented any increase in conditioned
responding during trace conditioning (Figure 1B).
A two-way ANOVA was performed with drug condition
(CPP vs. saline) and training protocol (paired vs. unpaired)
as the independent measures and the percentage of CRs
emitted during the last session as the dependent mea-
sure. Results revealed a significant effect of drug condition,
F(1, 23) = 20.93, p < .01, and training protocol, F(1, 23) =
31.73, p < .01, with an interaction between drug condition
and training protocol, F(1, 23) = 14.89, p < .01. Therefore,
animals that received saline before trace conditioning
emitted a greater percentage of CRs during the last day
of training than either of the two groups that received un-
paired training or the group that received CPP before trace
conditioning. Furthermore, the percentage of CRs emitted
by the group that received CPP before trace conditioning
was not different than the percentages emitted by the
two unpaired groups. Therefore, administration of CPP
before trace eyeblink conditioning completely prevented
2162
Journal of Cognitive Neuroscience
Volume 23, Number 9
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
9
e
-
2
p
1
d
5
f
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
c
9
n
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
9
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
f
/
t
.
o
n
1
8
M
a
y
2
0
2
1
acquisition of the CR. In addition, CPP administration did
not alter responding to explicitly unpaired stimuli.
Nearly all (86%) of the animals that received saline dur-
ing trace conditioning reached the 60% response criterion
(Learned/Saline), whereas none of the animals that were
trained with unpaired stimuli (Unpaired/Saline; Unpaired/
CPP) or those that received CPP before trace conditioning
did (Did Not Learn/CPP). To examine the difference in cell
counts between animals that learned versus those that did
not, a two-way ANOVA was conducted, with drug condi-
tion and training protocol as the independent measures
and the number of BrdU-labeled cells as the dependent
measure. Only saline-treated animals that reached the
60% criterion were included in this analysis. This was done
to ensure that we compared the cell counts of one entire
group of animals that successfully learned the task to one
group that did not. Results revealed a significant interaction
between drug condition and training protocol, F(1, 22) =
8.24, p < .05, indicating that only the animals that re-
ceived saline before trace conditioning (and that reached
criterion) displayed an increase in the number of surviving
cells (Figure 1C).
To examine the possibility that CPP may decrease cell
survival irrespective of its effects on learning, we examined
the number of new cells in two additional groups that re-
ceived saline (Saline After) or CPP (CPP After) 6 hr after
each day of training, a time point when CPP administration
did not interfere with acquisition of the CR. A repeated
measures ANOVA was conducted, with drug condition as
the independent measure, trial blocks as the repeated
measure, and percentage of CRs as the dependent mea-
sure. The results revealed a significant main effect of trial
block, F(7, 98) = 23.39, p < .01, indicating that the percent-
age of CRs emitted by these two groups increased over the
course of training. There was no effect of drug condition,
F(1, 14) = 1.00, p > .05, and no interaction between drug
condition and trial block, F(7, 98) = .08, p > .05. There-
fore, administration of the NMDA receptor antagonist
6 hr after every day of trace conditioning did not prevent
acquisition of the CR (Figure 1D). Inoltre, adminis-
tration of CPP 6 hr after training did not alter responding
on the last session of training, T(14) = .38, p > .05 (Saline
After vs. CPP After), nor did it change the number of trials
required to reach the 60% criterion (U = 8.0, p > .05, Saline
After vs. CPP After) or the number of trials required to emit
eight of nine consecutive CRs (U = 19.0, p > .05).
An ANOVA was used to compare the number of BrdU-
labeled cells in animals that received saline or CPP after
training, which reached the 60% criterion, to the number
of BrdU-labeled cells in experimentally naive animals,
which received saline injections, but no training (naive).
There was a difference in cell number among these groups,
F(2, 17) = 7.12, P < .01. Post hoc Tukey comparisons indi-
cated that animals that reached the 60% criterion despite
being injected with saline or CPP each day after training
possessed more BrdU-labeled cells than the naive controls
( p values < .05; Figure 1E).
Thus, animals could still acquire the CR as long as the
NMDA receptor antagonist was given after training. In
fact, their performance was no different than that of ani-
mals injected with saline and trained with paired stimuli.
Because the cell counts from these animals that received
CPP after training were not different from those that were
injected with saline and trained with paired stimuli, we
can conclude that antagonism of NMDA receptors before
but not 6 hr after training blocks learning of the CR and
thereby prevents the increase in cell survival normally
seen after learning. In other words, the effect of CPP on
cell survival is via its effect on acquisition of the CR and
not an effect of the drug alone.
In the past, our laboratory has reported a positive cor-
relation between the number of trials that an individual
animal requires to learn a task and the number of surviv-
ing cells in that animalʼs dentate gyrus (Waddell & Shors,
2008). When we examined the data from all animals in
Experiment 1, including those animals that did not reach
the 60% criterion, we observed a significant positive cor-
relation between the percentage of CRs an animal emitted
over all 800 trials and the number of surviving cells in that
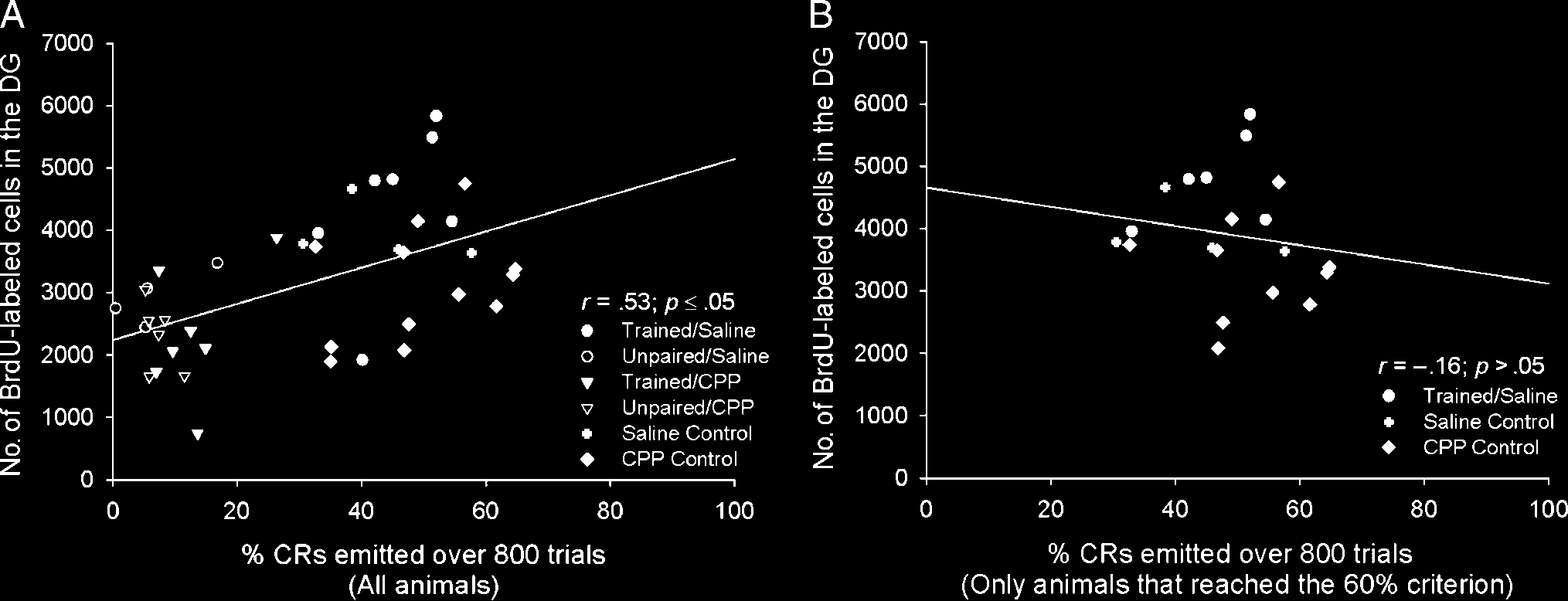
animalʼs dentate gyrus (r = .53, p < .05; Figure 2A). This
result confirms that animals that successfully learned the
CR retained more new cells than animals that failed to ac-
quire the CR. When we performed this analysis using only
the data from animals that reached the 60% criterion, we
observed no such correlation between the percentage of
CRs emitted during training and the number of surviving
cells (r = −.16, p > .05; Figure 2B).
Experiment 2 Results: DCS Facilitates Trace
Conditioning and Neurogenesis
Pretraining administration of DCS has previously been
demonstrated to facilitate learning of trace eyeblink con-
ditioning (Waddell et al., 2010; Thompson & Disterhoft,
1997; Thompson et al., 1992). The current experiment
was conducted to determine whether DCS-facilitated
learning would result in an increase in the number of sur-
viving newborn cells in the adult dentate gyrus. The experi-
ment consisted of five groups. Groups 1 E 2 received
DCS or saline before training with trace eyeblink condi-
tioning. Groups 3 E 4 received DCS or saline before
training with explicitly unpaired stimuli. Group 5 received
DCS 6 hr after training with trace eyeblink conditioning. A
determine whether DCS facilitated acquisition, we exam-
ined early acquisition, which was defined as the percent-
age of CRs emitted during the first day of training. An
independent samples t test comparing the percentage of
CRs emitted during the first day indicated that learning in
the presence of DCS significantly enhanced early acquisi-
tion of the CR, T(14) = 2.63, P < .05 (Trace vs. DCS during
Trace). A separate independent samples t test revealed
that DCS did not alter responding to unpaired stimuli dur-
ing the first day of training, t(9) = 0.85, p > .05 ( Unpaired
vs. DCS Unpaired).
Curlik and Shors
2163
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
3
l
9
e
–
2
P
1
D
5
F
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
C
9
N
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
P
C
D
N
.
B
sì
2
0
G
1
tu
0
e
.
S
T
2
o
1
N
5
9
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
F
.
T
.
.
/
o
N
1
8
M
UN
sì
2
0
2
1
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
3
l
9
e
–
2
P
1
D
5
F
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
C
9
N
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
P
C
D
N
.
B
sì
2
0
G
1
tu
0
e
.
S
T
2
o
1
N
5
9
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
F
.
.
T
.
/
o
N
1
8
M
UN
sì
2
0
2
1
Figura 2. (UN) When the data from all animals in the first experiment were examined, we observed a significant positive correlation between the
percentage of CRs an individual animal emitted over the entire course of training and the number of BrdU-labeled cells in that animalʼs dentate gyrus.
(B) Tuttavia, when we analyzed the data from only those animals that reached the 60% criterion during the first experiment, we did not observe
a significant correlation.
The total number of CRs emitted during the entire 4 days
of training differed between the five groups, F(4, 27) =
25.18, P < .05. Post hoc analysis indicated that pretraining
administration of DCS did not alter the total number of CRs
emitted during the 4 days of trace conditioning, ( p > .05
Trace vs. DCS during Trace; Figure 3A). Therefore, al-
though administration of DCS before trace conditioning
facilitated early acquisition of the CR, it had no effect on
overall responding during the remaining days of training.
Inoltre, DCS did not alter the total number of CRs
emitted during training when it was administered 6 hr after
training ( p > .05, Trace vs. DCS After, data not shown;
Ledgerwood, Richardson, & Cranney, 2003) or when it
was administered before training with explicitly unpaired
stimuli ( p > .05, Unpaired vs. DCS unpaired).
Groups that were trained with trace conditioning (Trace,
DCS during Trace, and DCS After) increased the percentage
of CRs that they emitted across training blocks, F(7, 126) =
20.144, P < .05, with no interaction between group and
training block, F(14, 126) = 1.25, p > .05, revealing that
all trace conditioned groups successfully acquired the CR
over the course of training. There was no increase in respond-
ing in animals exposed to unpaired stimuli, F(7, 63) = 1.18,
p > .05, and no difference in overall responding between
the two unpaired groups, F(1, 9) = 1.27, p > .05. Così, only
animals that were trained with trace conditioning success-
fully acquired the CR. DCS did not alter the number of re-
sponses emitted during explicitly unpaired training. These
results suggest that DCS does not increase conditioning
simply by increasing nonspecific responding to the CS.
There were no differences in responding during the last
day of training among those groups exposed to trace con-
ditioning, F(2, 18) = 0.82, p > .05. Together, these results
indicate that the effects of DCS on trace eyeblink condi-
tioning are limited to early acquisition of the CR. More-
Sopra, DCS only facilitated acquisition when it was admin-
istered 30 min before but not 6 hr after training.
Most (78%) animals given saline during trace condition-
ing reached our 60% response criterion (Learned/Saline),
whereas all animals (100%) given DCS during (Learned/
DCS) or after trace conditioning (DCS After) reached this
criterion. None of the animals that received unpaired training
reached the criterion. È interessante notare, all trace-conditioned and
no unpaired animals reached the eight of nine response cri-
ionizzato. Inoltre, there was a strong positive correlation
between these two criteria (Rho = 0.83, P < .01). However,
animals tended to emit eight of nine consecutive CRs before
they reached the 60% criterion.
The number of BrdU-labeled cells in the entire dentate
gyrus was different between the trained groups, F(5, 27) =
13.27, p < .01. Post hoc comparisons revealed that saline-
treated animals that reached the 60% response criterion
retained more newborn cells than saline-treated animals
that were trained with trace conditioning that did not reach
criterion ( p < .05, Learned/Saline vs. Did Not Learn/Saline).
The saline-treated animals that reached this criterion also
retained more cells than animals that received saline dur-
ing unpaired training ( p < .01, Learned/Saline vs. Unpaired/
Saline) and DCS during unpaired training ( p < .01, Learned/
Saline vs. Unpaired/DCS).
Interestingly, although animals that received DCS dur-
ing training expressed more CRs at the beginning of train-
ing, they did not retain more new neurons than animals
that learned in the absence of the drug ( p > .05, Learned/
DCS vs. Learned/Saline; Figure 3B). Inoltre, administra-
tion of DCS by itself did not increase the survival of newborn
neurons ( p > .05, Unpaired/DCS vs. Unpaired/Saline).
Inoltre, the cell counts from animals that received
DCS 6 hr after training (which did not alter acquisition of
the CR) were no different from those of animals that learned
2164
Journal of Cognitive Neuroscience
Volume 23, Numero 9
in the absence of the drug ( p > .05, DCS After vs. Learned/
Saline, data not shown), indicating that administration of
DCS itself did not decrease the number of surviving cells.
Consistent with previous results (Waddell & Shors, 2008),
we observed a significant positive correlation between the
number of trials that an individual animal required to reach
IL 60% response criterion and the number of BrdU-labeled
cells in that animalʼs dentate gyrus (Rho = .59, P < .05;
Figure 3C). In addition, there was a positive correlation be-
tween the number of trials required to emit eight of nine con-
secutive CRs and the number of BrdU-labeled cells (Rho =
.47, p < .05; Figure 3D). As in the first experiment, when we
examined the data from all animals, including those that did
not reach the 60% criterion, we observed a positive correla-
tion between the percentage of CRs emitted over the entire
course of training and the number of surviving BrdU-labeled
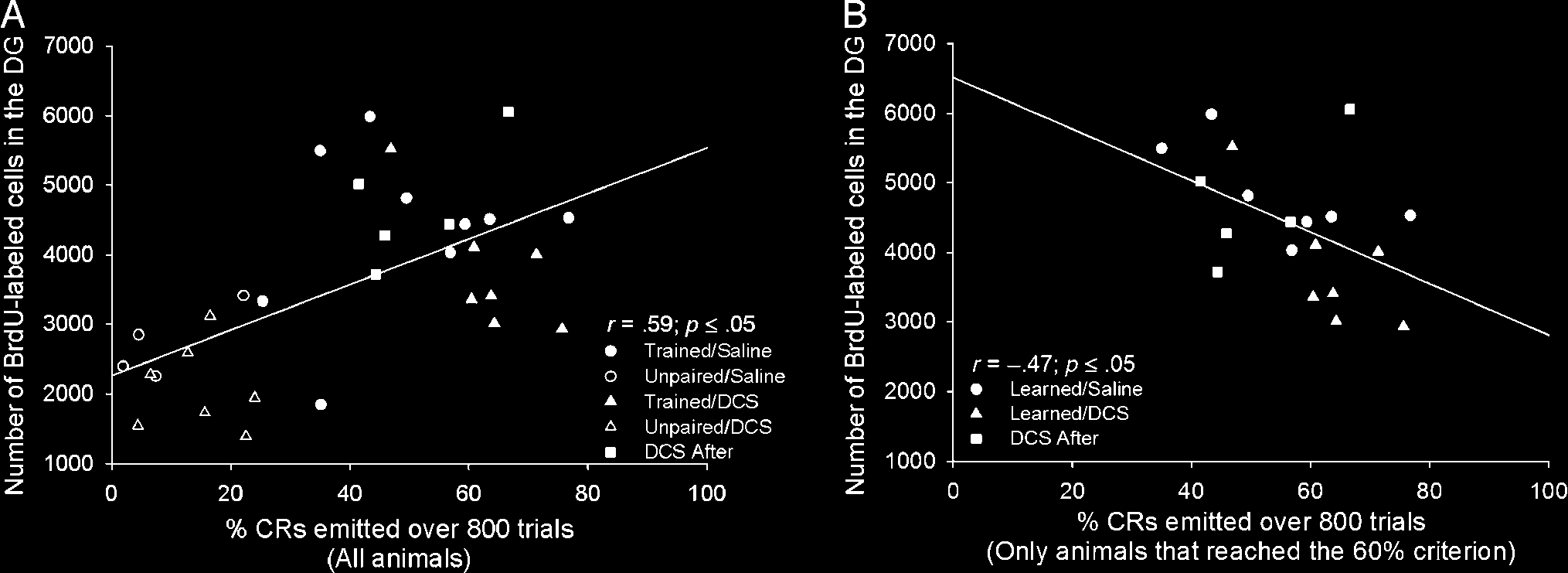
cells (r = .59; Figure 4A). Furthermore, when we examined
the data from animals that successfully learned the task (i.e.,
those that reached the 60% criterion), we found a strong
negative correlation between the percentage of CRs an indi-
vidual animal emitted during the 800 trials of trace condition-
ing and the number of surviving cells in the granule cell layer
of that animalʼs dentate gyrus (r = −.47, p < .05; Figure 4B).
Thus, animals that successfully acquired the CR and required
more trials to do so retained more of the newborn cells.
DISCUSSION
It is now well established that training with various learn-
ing tasks can increase the number of newly born cells that
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
9
e
-
2
p
1
d
5
f
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
c
9
n
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
9
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
/
f
.
t
o
n
1
8
M
a
y
2
0
2
1
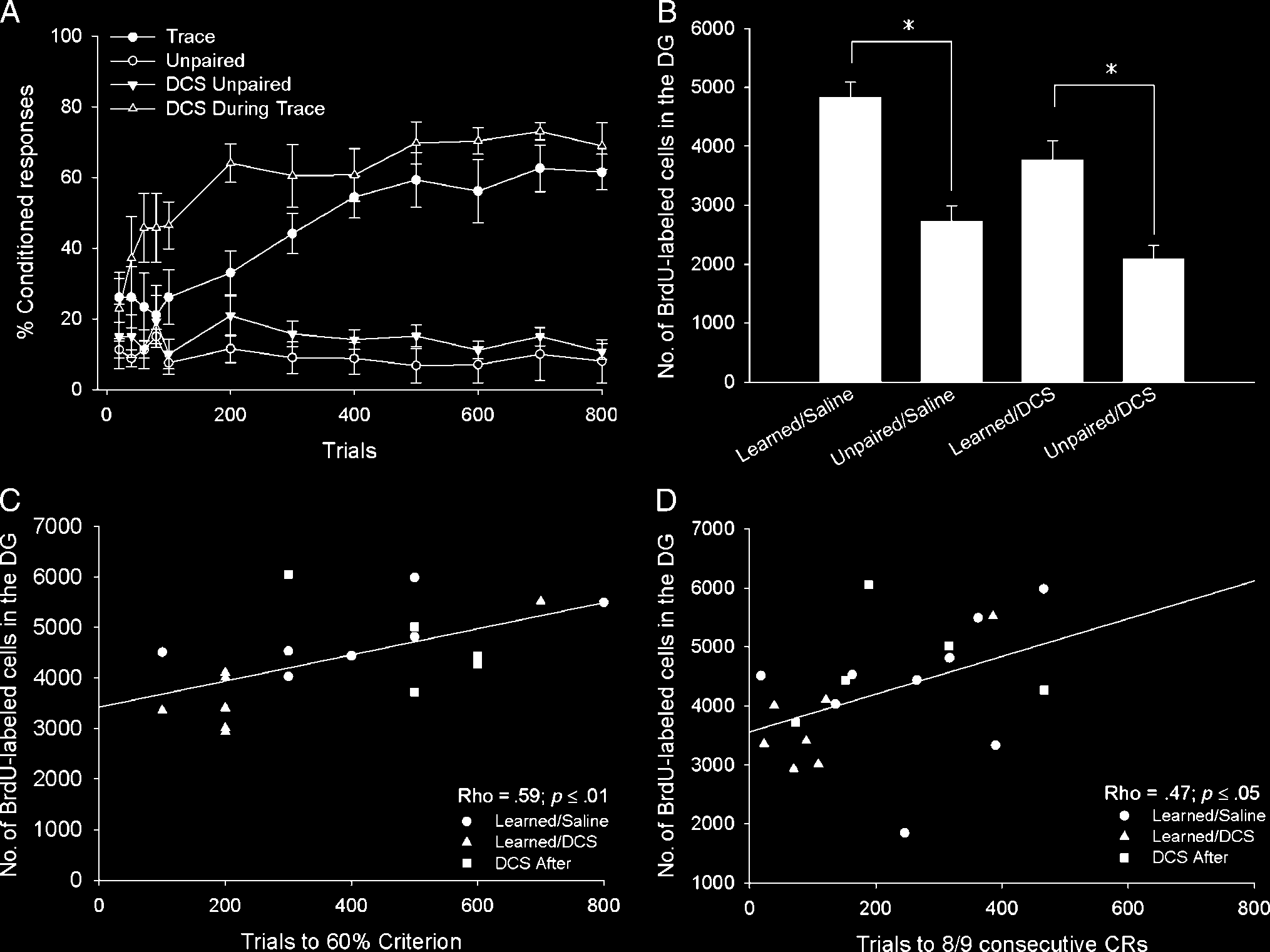
Figure 3. (A) Intraperitoneal administration of DCS before training with trace eyeblink conditioning (DCS during Trace) facilitated acquisition of the
CR, as evidenced by an increase in conditioned responding during the first day of training. DCS did not alter responding to explicitly unpaired stimuli
(DCS Unpaired). For comparison, animals received saline during training with trace conditioning (Trace) or explicitly unpaired stimuli (Unpaired).
(B) Animals treated with saline during trace conditioning that reached the 60% criterion (Learned/Saline) retained more BrdU-labeled cells than
animals that received saline during unpaired training (Unpaired/Saline), or animals that received DCS during unpaired training (Unpaired /DCS).
Those animals that received DCS during Trace conditioning (Learned/DCS) retained significantly more BrdU-labeled cells than animals that received
DCS before unpaired training (Unpaired/ DCS). (C) A significant positive correlation was observed between the number of trials an individual
animal required to reach criterion and the number of BrdU-labeled cells in that animalʼs dentate gyrus. Only animals that successfully reached
the 60% criterion were included in this analysis. (D) The number of BrdU-labeled cells also positively correlated with the number of trials required
to reach eight of nine consecutive CRs during any one block of trials.
Curlik and Shors
2165
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
9
e
-
2
p
1
d
5
f
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
c
9
n
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
9
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
f
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Figure 4. (A) A significant positive correlation was observed between the percentage of CRs an individual animal emitted over the entire course
of training (800 trials) and the number BrdU-labeled cells in that animalʼs dentate gyrus. (B) After analyzing only data from animals that learned
(i.e., reached the learning criterion), there was a significant negative correlation between the percentage of CRs an individual animal emitted during
training and the number of BrdU-labeled cells in the granule cell layer of that animalʼs dentate gyrus.
survive to become neurons in the adult hippocampus
(Shors, 2009). The goal of these experiments was to assess
the role of learning (per se) in this phenomenon. To do
this, we manipulated performance of a learned response
in two ways and directions. First, we completely prevented
learning by using an NMDA receptor antagonist—which
was injected each day before training with trace condition-
ing. Animals that received the antagonist before training
did not learn, and they did not retain any more cells in
their hippocampus than animals that were trained with
unpaired stimuli, which also did not learn the CR. These
effects were not due to adverse consequences of the drug
itself because animals that were injected with the antago-
nist after training were able to learn the CR and retained
as many new cells as the animals that were trained in the
presence of saline. As noted, animals that learned the CR
retained many more new cells than those trained with
unpaired stimuli. Thus, preventing learning with an NMDA
receptor antagonist prevents the increase in cell number
that typically occurs after training. These effects are clear,
albeit not necessarily surprising, because our previous
studies have suggested that the effects of training on cell
survival only occur in animals that actually learn (Dalla
et al., 2007; Sisti, Glass, & Shors, 2007). However, these
results extend our findings to demonstrate that pharma-
cological manipulations of learning determine how many
cells ultimately survive to become neurons in the adult
hippocampus. Because the cells that are rescued from
death by learning remain in the hippocampus for months,
at least (Leuner et al., 2004), these pharmacological effects
of CPP presumably have similarly persistent effects on the
circuitry of the adult hippocampal formation.
In the second experiment, we manipulated learning in
the opposite way, again using a drug that modulates neuro-
nal activity at the NMDA receptor. The drug, DCS, is a partial
agonist of the strychnine-insensitive glycine binding site,
which facilitates NMDA receptor-mediated excitatory trans-
mission (Rouaud & Billard, 2003; Henderson, Johnson,
& Ascher, 1990; Hood, Compton, & Monahan, 1989) and
learning (Waddell et al., 2010; Thompson & Disterhoft,
1997; Thompson et al., 1992). Similarly, we observed an
increase in performance during trace eyeblink condition-
ing in response to the drug. As a consequence, we report
that the number of cells that were retained in the group
that received DCS before trace conditioning was elevated
when compared with the number of cells in animals that
received unpaired training in the presence of the drug.
However, DCS did not increase cell numbers beyond those
observed in animals that learned in the presence of saline.
Furthermore, those animals that naturally failed to acquire
the CR did not retain any more of the new cells than those
trained with unpaired stimuli. These data suggest that the
overall increase in performance in the presence of the drug
was not sufficient to increase the number of surviving cells
beyond what learning without the drug would accomplish.
This result is consistent with that of a previous study,
where we found that learning, when it occurred, rescued
nearly all, if not all, of the cells that were available to be
rescued (Waddell & Shors, 2008). One might surmise that
there are simply no more new cells left to rescue—at least
not of this particular cohort. Despite a potential ceiling on
the number of cells that can be rescued, animals that were
trained in the presence of DCS tended to learn better and
more of them did so. These results are consistent with re-
cent findings, indicating that pretraining administration of
DCS facilitates declarative learning in humans (Onur et al.,
2010). Moreover, DCS is being used in conjunction with
cognitive behavioral therapy to treat patients with various
mental disorders (Davis, Ressler, Rothbaum, & Richardson,
2006; Hofmann et al., 2006). On the basis of the findings
reported here, one might propose that the learning that
occurs with DCS during cognitive behavioral therapy
2166
Journal of Cognitive Neuroscience
Volume 23, Number 9
increases the number of new cells that survive to become
mature neurons in these human subjects, a process that is
likely to benefit them in the future.
It is noted that exposure to DCS alone did not seem to
affect cell number. Animals that were injected with DCS
before unpaired training did not possess any more or fewer
cells than those injected with saline and exposed to un-
paired training. Moreover, animals injected with DCS after
paired training, which learned, retained as many cells as
those trained with trace conditioning after an injection of
saline. Because DCS is known to enhance excitability
through facilitating activation of NMDA receptors, these
data suggest that this activation is not sufficient to alter
the survival of 1- to 2-week-old hippocampal cells.
The most intriguing results related to individual differ-
ences in learning and the number of surviving cells. Across
both experiments, we observed a strong positive correla-
tion between the percentage of learned responses that an
individual animal emitted over the entire 800 trials of train-
ing and the number of surviving BrdU-labeled cells. Thus,
animals that tended to learn better (i.e., emitted more CRs
over the course of training) also tended to retain more of
the new cells in their hippocampus. Because the animals
were sacrificed 3 weeks after training, these cells would al-
ready be mature neurons. This relationship between learn-
ing and neurogenesis can be further parceled into those
animals that learned quickly versus those that did not learn
quickly but still did learn. In this case, we examined the
data from animals that reached a criterion of 60% CRs in
at least one block of training trials. This criterion is an es-
tablished one in eyeblink conditioning because it tends to
capture the vast majority of animals that will learn regard-
less of how many trials are given (Waddell & Shors, 2008;
Dalla et al., 2007). Using this criterion, we correlated the
number of trials an individual animal required to reach cri-
terion with the number of BrdU-labeled cells in that animalʼs
dentate gyrus. There was a strong and positive correlation
in the experiment with DCS (Rho = .59), irrespective of
whether the animals received the drug or not. In other
words, of the animals that learned those that required
more trials to reach the 60% learning criterion tended to
retain more of the new cells. These results were replicated
with a second commonly used criterion; the number of trials
required for an animal to emit a CR on eight of nine con-
secutive trials (Dalla et al., 2009; Nokia et al., 2008). Thus,
in the DCS study, animals that learned well retained more
new neurons than animals that did not learn. However,
those animals that required more trials to do so retained
more cells than those that learned with fewer training trials
(Figure 5). These results are consistent with previous re-
ports from our laboratory using task parameters to manipu-
late learning and rates of learning (Waddell & Shors, 2008).
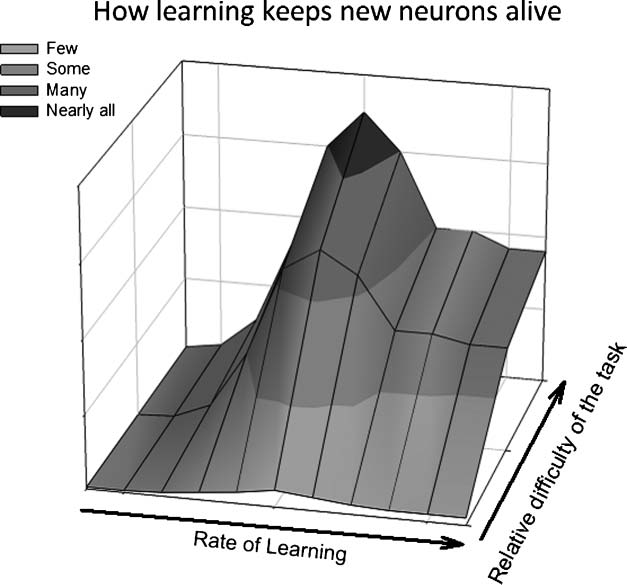
On the basis of these findings, we propose a model in which
the number of trials required to learn a task and the relative
difficulty of a task interact to determine the number of
new neurons that survive after a learning experience (Fig-
ure 6). In animals that are able to master the behavioral
response, those that require more trials to do so retain
more new neurons. This effect is especially robust when
animals are trained on tasks that are difficult to master.
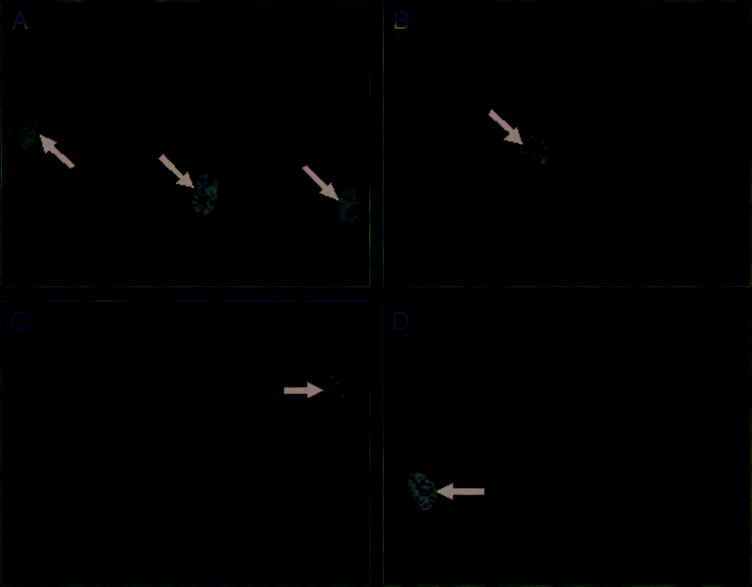
Figure 5. Representative
BrdU-labeled cells from animals
that learned or did not learn
in the presence of saline, DCS,
or NMDA receptor antagonist:
(A) Learned/Saline, (B) Unpaired/
Saline, (C) Learned/DCS, and
(D) Did Not Learn/CPP. Arrows
indicate BrdU-labeled cells.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
9
e
-
2
p
1
d
5
f
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
c
9
n
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
9
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
/
.
t
o
n
1
8
M
a
y
2
0
2
1
Curlik and Shors
2167
Figure 6. A model of how
learning influences the survival
of adult-born hippocampal
neurons. The relative difficulty
of the task being learned and
the rate at which an individual
animal learns the task
determine how many newborn
neurons survive in the adult
dentate gyrus. Successful
learning of a relatively easy task
rescues very few neurons,
whereas learning a difficult task
rescues many neurons. Of those
animals that successfully learn,
those that take many trials to do
so retain more newborn
neurons than those that rapidly
learn.
It is noted that there are other explanations for the ob-
served findings. For example, one injection of BrdU pro-
vides a “snapshot” of one population of cells—those that
were dividing at the time of or shortly after the BrdU in-
jection. It is possible that animals that rapidly acquire the
CR, which possess fewer BrdU-labeled cells, may be res-
cuing cells that are slightly younger or slightly older than
the population labeled with BrdU. Although this is a pos-
sibility, it does seem unlikely because we and others have
found that there is a critical period during which the new
cells can be rescued: Cells that are less than 3 days of age
or greater than 3 weeks were not preferentially retained
after learning (Anderson & Shors, submitted; Epp, Spritzer,
& Galea, 2007).
A correlation between rate of acquisition and cell num-
ber did not exist for the data from the second experiment,
in which an NMDA receptor antagonist was injected before
and after training. There was much less variability among
the individual animals in terms of learning, which likely ex-
plains the absence of a correlation. Alternatively, exposure
to the antagonist may have decreased cell survival on its
own. Indeed, we did observe a slight decrease in the num-
ber of surviving cells in animals that were given CPP after
training (although they learned). This effect was not signif-
icant. However, others have reported that antagonism of
NMDA receptors decreases cell survival (Tashiro, Zhao, &
Gage, 2006). Still others report that NMDA receptor activa-
tion after the learning experience is necessary to retain
some memories (Burgos-Robles, Vidal-Gonzalez, Santini,
& Quirk, 2007). Perhaps if we had infused the antagonist
persistently after training, more cells would have died as a
consequence.
During training of the classically conditioned eyeblink
response cells in area CA1 of the hippocampus become
more excitable. This effect is most prevalent in those an-
imals that successfully learn the CR (Moyer, Thompson, &
Disterhoft, 1996). During trace fear conditioning, granule
cells in the dentate also increase their responsiveness to
the CS (Gilmartin & McEchron, 2005). Both of these effects
do not appear to persist beyond the learning phase. Thus, it
is possible that an increase in cell excitability in either CA1
or dentate gyrus contributes to the increase in cell survival.
In other words, the new neurons in the DG of animals that
require more trials (and time) to learn the CR would be
exposed to a longer period of excitability when compared
with the amount in animals that quickly learn the task or
animals that fail to learn. How an increase in excitability
would modulate neuronal survival is currently unknown,
although one possibility may be through the activation of
NMDA receptors in the hippocampal network. Regardless,
the present findings indicate that learning will induce the
survival of new neurons when that learning is both suffi-
ciently difficult to achieve and successful (Figure 6).
Acknowledgments
This work was supported by the National Institutes of Health
(grant nos. MH-59970 and ARRA-3R01MH059970-10S1) and
the National Science Foundation (grant nos. IOB-0444364 and
IOS-0914386) to T. J. S.
2168
Journal of Cognitive Neuroscience
Volume 23, Number 9
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
9
e
-
2
p
1
d
5
f
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
c
9
n
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
9
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
/
f
.
o
n
1
8
M
a
y
2
0
2
1
Reprint requests should be sent to Tracey J. Shors, Department of
Psychology and Center for Collaborative Neuroscience, Rutgers
University, 152 Frelinghuysen Road, Piscataway, NJ 08854-8020,
or via e-mail: shors@rutgers.edu.
REFERENCES
Altman, J., & Das, G. D. (1965). Autoradiographic and
histological evidence of postnatal hippocampal
neurogenesis in rats. Journal of Comparative Neurology,
124, 319–335.
Ambrogini, P., Cuppini, R., Cuppini, C., Ciaroni, S., Cecchini, T.,
Ferri, P., et al. (2000). Spatial learning affects immature granule
cell survival in adult rat dentate gyrus. Neuroscience Letters,
286, 21–24.
Anderson, M., & Shors, T. (submitted). The effects of learning
on neurogenesis: Survival versus proliferation.
Anderson, M. L., Sisti, H. M., Curlik, D. M., & Shors, T. J. (2010).
Associative learning increases adult neurogenesis during a
critical period. European Journal of Neuroscience, 33, 175–181.
Burgos-Robles, A., Vidal-Gonzalez, I., Santini, E., & Quirk, G. J.
(2007). Consolidation of fear extinction requires NMDA
receptor-dependent bursting in the ventromedial prefrontal
cortex. Neuron, 53, 871–880.
Cameron, H. A., & McKay, R. D. (2001). Adult neurogenesis
produces a large pool of new granule cells in the dentate
gyrus. Journal of Comparative Neurology, 435, 406–417.
Cameron, H. A., Woolley, C. S., McEwen, B. S., & Gould, E.
(1993). Differentiation of newly born neurons and glia in the
dentate gyrus of the adult rat. Neuroscience, 56, 337–344.
Dalla, C., Bangasser, D. A., Edgecomb, C., & Shors, T. J. (2007).
Neurogenesis and learning: Acquisition and asymptotic
performance predict how many new cells survive in the
hippocampus. Neurobiology of Learning and Memory,
88, 143–148.
Dalla, C., Papachristos, E. B., Whetstone, A. S., & Shors, T. J.
(2009). Female rats learn trace memories better than male
rats and consequently retain a greater proportion of new
neurons in their hippocampi. Proceedings of the National
Academy of Sciences, U.S.A., 106, 2927–2932.
Davis, M., Ressler, K., Rothbaum, B. O., & Richardson, R.
(2006). Effects of D-cycloserine on extinction: Translation
from preclinical to clinical work. Biological Psychiatry, 60,
369–375.
Dayer, A. G., Ford, A. A., Cleaver, K. M., Yassaee, M., & Cameron,
H. A. (2003). Short-term and long-term survival of new
neurons in the rat dentate gyrus. Journal of Comparative
Neurology, 460, 563–572.
Epp, J. R., Spritzer, M. D., & Galea, L. A. (2007). Hippocampus-
dependent learning promotes survival of new neurons in
the dentate gyrus at a specific time during cell maturation.
Neuroscience, 149, 273–285.
Eriksson, P. S., Perfilieva, E., Bjork-Eriksson, T., Alborn, A. M.,
Nordborg, C., Peterson, D. A., et al. (1998). Neurogenesis
in the adult human hippocampus. Nature Medicine, 4,
1313–1317.
Gilmartin, M. R., & McEchron, M. D. (2005). Single neurons
in the dentate gyrus and CA1 of the hippocampus exhibit
inverse patterns of encoding during trace fear conditioning.
Behavioral Neuroscience, 119, 164–179.
Gould, E., Beylin, A., Tanapat, P., Reeves, A., & Shors, T. J.
(1999). Learning enhances adult neurogenesis in the
hippocampal formation. Nature Neuroscience, 2,
260–265.
Hastings, N. B., & Gould, E. (1999). Rapid extension of axons
into the CA3 region by adult-generated granule cells. Journal
of Comparative Neurology, 413, 146–154.
Henderson, G., Johnson, J. W., & Ascher, P. (1990). Competitive
antagonists and partial agonists at the glycine modulatory site
of the mouse N-methyl-D-aspartate receptor. Journal of
Physiology, 430, 189–212.
Hofmann, S. G., Meuret, A. E., Smits, J. A., Simon, N. M.,
Pollack, M. H., Eisenmenger, K., et al. (2006). Augmentation
of exposure therapy with D-cycloserine for social anxiety
disorder. Archives of General Psychiatry, 63, 298–304.
Hood, W. F., Compton, R. P., & Monahan, J. B. (1989).
D-Cycloserine: A ligand for the N-methyl-D-aspartate
coupled glycine receptor has partial agonist characteristics.
Neuroscience Letters, 98, 91–95.
Kaplan, M. S., & Hinds, J. W. (1977). Neurogenesis in the adult
rat: Electron microscopic analysis of light radioautographs.
Science, 197, 1092–1094.
Kornack, D. R., & Rakic, P. (1999). Continuation of
neurogenesis in the hippocampus of the adult macaque
monkey. Proceedings of the National Academy of Sciences,
U.S.A., 96, 5768–5773.
Ledgerwood, L., Richardson, R., & Cranney, J. (2003). Effects
of D-cycloserine on extinction of conditioned freezing.
Behavioral Neuroscience, 117, 341–349.
Leuner, B., Falduto, J., & Shors, T. J. (2003). Associative memory
formation increases the observation of dendritic spines in
the hippocampus. Journal of Neuroscience, 23, 659–665.
Leuner, B., Mendolia-Loffredo, S., Kozorovitskiy, Y., Samburg,
D., Gould, E., & Shors, T. J. (2004). Learning enhances
the survival of new neurons beyond the time when the
hippocampus is required for memory. Journal of Neuroscience,
24, 7477–7481.
Markakis, E. A., & Gage, F. H. (1999). Adult-generated neurons
in the dentate gyrus send axonal projections to field CA3
and are surrounded by synaptic vesicles. Journal of
Comparative Neurology, 406, 449–460.
Miller, M. W., & Nowakowski, R. S. (1988). Use of
bromodeoxyuridine-immunohistochemistry to examine the
proliferation, migration and time of origin of cells in the
central nervous system. Brain Research, 457, 44–52.
Moyer, J. R., Jr., Thompson, L. T., & Disterhoft, J. F. (1996).
Trace eyeblink conditioning increases CA1 excitability
in a transient and learning-specific manner. Journal of
Neuroscience, 16, 5536–5546.
Moyer, J. R., Jr., Thompson, L. T., & Disterhoft, J. F. (2000).
Increased excitability of aged rabbit CA1 neurons after
trace eyeblink conditioning. Journal of Neuroscience, 20,
5476–5482.
Nokia, M. S., Penttonen, M., Korhonen, T., & Wikgren, J.
(2008). Hippocampal theta (3–8 Hz) activity during classical
eyeblink conditioning in rabbits. Neurobiology of Learning
and Memory, 90, 62–70.
Onur, O. A., Schlaepfer, T. E., Kukolja, J., Bauer, A., Jeung, H.,
Patin, A., et al. (2010). The N-methyl-D-aspartate receptor
co-agonist D-cycloserine facilitates declarative learning and
hippocampal activity in humans. Biological Psychiatry, 67,
1205–1211.
Rouaud, E., & Billard, J. M. (2003). D-Cycloserine facilitates
synaptic plasticity but impairs glutamatergic neurotransmission
in rat hippocampal slices. British Journal of Pharmacology,
140, 1051–1056.
Servatius, R. J., & Shors, T. J. (1996). Early acquisition, but not
retention, of the classically conditioned eyeblink response is
N-methyl-D-aspartate (NMDA) receptor dependent.
Behavioral Neuroscience, 110, 1040–1048.
Shors, T. J. (2008). From stem cells to grandmother cells: How
neurogenesis relates to learning and memory. Cell Stem Cell,
3, 253–258.
Shors, T. J. (2009). Saving new brain cells. Scientific American,
300, 46–52, 54.
Curlik and Shors
2169
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
9
e
-
2
p
1
d
5
f
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
c
9
n
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
9
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
/
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
Sisti, H. M., Glass, A. L., & Shors, T. J. (2007). Neurogenesis
Thompson, L. T., Moskal, J. R., & Disterhoft, J. F. (1992).
and the spacing effect: Learning over time enhances memory
and the survival of new neurons. Learning and Memory,
14, 368–375.
Tashiro, A., Zhao, C., & Gage, F. H. (2006). Retrovirus-mediated
single-cell gene knockout technique in adult newborn
neurons in vivo. Nature Protocols, 1, 3049–3055.
Thompson, L. T., & Disterhoft, J. F. (1997). Age- and dose-
dependent facilitation of associative eyeblink conditioning by
D-cycloserine in rabbits. Behavioral Neuroscience, 111,
1303–1312.
Hippocampus-dependent learning facilitated by a
monoclonal antibody or D-cycloserine. Nature, 359,
638–641.
Waddell, J., Mallimo, E., & Shors, T. (2010). D-Cycloserine
reverses the detrimental effects of stress on learning in
females and enhances retention in males. Neurobiology of
Learning and Memory, 93, 31–36.
Waddell, J., & Shors, T. J. (2008). Neurogenesis, learning and
associative strength. European Journal of Neuroscience, 27,
3020–3028.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
9
e
-
2
p
1
d
5
f
9
/
1
2
9
3
4
/
2
9
2
/
2
2
9
1
o
5
c
9
n
/
1
2
0
7
1
7
0
6
2
2
1
1
1
5
9
/
7
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
9
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
.
f
/
o
n
1
8
M
a
y
2
0
2
1
2170
Journal of Cognitive Neuroscience
Volume 23, Number 9