Landmark-dependent Navigation Strategy Declines across
the Human Life-Span: Evidence from Over
37,000 Participants
Greg L. West1*, Zita Eva Patai2,3*, Antoine Coutrot4, Michael Hornberger5,
Véronique D. Bohbot6**, and Hugo J. Spiers2**
Astratto
■ Humans show a remarkable capacity to navigate various
environments using different navigation strategies, and we
know that strategy changes across the life span. Tuttavia, Questo
observation has been based on studies of small sample sizes. A
this end, we used a mobile app–based video game (Sea Hero
Quest) to test virtual navigation strategies and memory perfor-
mance within a distinct radial arm maze level in over 37,000 par-
ticipants. Players were presented with six pathways (three open
and three closed) and were required to navigate to the three
open pathways to collect a target. Prossimo, all six pathways were
made available and the player was required to visit the pathways
that were previously unavailable. Both reference memory and
working memory errors were calculated. Crucially, at the end
of the level, the player was asked a multiple-choice question
about how they found the targets (cioè., a counting-dependent
strategy vs. a landmark-dependent strategy). As predicted from
previous laboratory studies, we found the use of landmarks
declined linearly with age. Those using landmark-based strate-
gies also performed better on reference memory than those
using a counting-based strategy. These results extend previous
observations in the laboratory showing a decreased use of
landmark-dependent strategies with age. ■
INTRODUCTION
The human brain changes across the life span—during
ageing people experience decreases in executive and
memory functions, which are associated with gray matter
loss in the frontal cortex and the hippocampus, respec-
tively (Thambisetty et al., 2010; Moffat, Elkins, & Resnick,
2006; Raz, Rodrigue, Head, Kennedy, & Acker, 2004;
Small, Tsai, DeLaPaz, Mayeux, & Stern, 2002). The integ-
rity of the hippocampus during ageing is of particular
interest to researchers as a decline in behavioral perfor-
mance supported by this structure, such as wayfinding
ability (Mapstone, Steffenella, & Duffy, 2003; Klein et al.,
1999; Passini, Rainville, Marchand, & Joanette, 1995) COME
well as gray matter loss, are an early sign of cognitive
decline and Alzheimer’s disease (AD; Jessen et al., 2010;
Apostolova et al., 2006; Jagust et al., 2006). Changes in
hippocampal gray matter during ageing also correlates
with changes in behaviors related to other aspects of
learning and memory such as navigational strategies used
1University of Montréal, Quebec, Canada, 2University College
London, United Kingdom, 3King’s College London, United
Kingdom, 4University of Lyon, France, 5University of East
Anglia, Norwich, United Kingdom, 6McGill University, Montréal,
Quebec, Canada
*Listed as co-first authors.
**Listed as co-senior authors.
© 2022 Istituto di Tecnologia del Massachussetts
when learning a new environment ( West et al., 2018;
Bohbot, Lerch, Thorndycraft, Iaria, & Zijdenbos, 2007;
Iaria, Petrides, Dagher, Pike, & Bohbot, 2003). The hip-
pocampus is required during navigation when relation-
ships between multiple landmarks in the environment
are learned to form a cognitive map, allowing for naviga-
tion irrespective of the starting position of the observer
and is commonly referred to as spatial learning (Ekstrom
& Ranganath, 2018; Bohbot et al., 2007; Bohbot, Iaria, &
Petrides, 2004; Iaria et al., 2003; Packard, Hirsh, & White,
1989; O’Keefe & Nadel, 1978). In contrasto, when a person
navigates by using a series of rigid turns from one specific
starting point in the environment, stimulus–response
associations are formed in the striatum, which includes
the caudate nucleus, and this is referred to as response
apprendimento (Bohbot et al., 2004, 2007; Packard et al., 1989;
O’Keefe & Nadel, 1978).
Numerous studies in rodents and humans have demon-
strated that spatial learning is supported by the hippocam-
pus while response learning recruits the caudate nucleus
(West et al., 2018; Konishi et al., 2013; Konishi & Bohbot,
2013; Lerch et al., 2011; Bohbot et al., 2007; Iaria et al.,
2003; Packard & McGaugh, 1992, 1996; McDonald & White,
1993; Packard et al., 1989). Per esempio, when rodents
were presented with a dual-solution task that could be
completed using spatial or response learning in a plus
maze, increased acetylcholine in the hippocampus was
Journal of Cognitive Neuroscience 35:3, pag. 452–467
https://doi.org/10.1162/jocn_a_01956
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
C
N
_
UN
_
0
1
9
5
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
measured in rodents that would spontaneously use a spa-
tial strategy, whereas increased acetylcholine was observed
in the caudate nucleus of rodents who would spontane-
ously use a response strategy (Chang & Gold, 2003).
Rodents trained on the Morris Water Maze by learning
the relationship between visual landmarks (spatial learn-
ing) displayed larger hippocampal volume after 5 days of
training whereas a separate group trained to solve the
maze using a single beacon stimulus while landmarks were
hidden with a curtain (response learning) displayed a
larger volume in the striatum, which includes the caudate
nucleus (Lerch et al., 2011). In humans, hippocampal vol-
ume positively correlated with peoples’ ability to use a
learned cognitive map of a university campus to locate
relative building locations (Schinazi, Nardi, Newcombe,
Shipley, & Epstein, 2013). Inoltre, when people
navigated in a dual-solution virtual maze that allowed for
the use of both spatial or response navigation learning,
people who navigated using spatial strategies showed
greater activity in the hippocampus (Iaria et al., 2003)
and also greater hippocampal gray matter (Bohbot et al.,
2007). In contrasto, people who used response strategies
showed increased activity (Iaria et al., 2003) and gray
matter (Bohbot et al., 2007) in the caudate nucleus. These
observations were replicated in older adults where the use
of spatial strategies was associated with greater activity
(Konishi et al., 2013) and gray matter (Konishi & Bohbot,
2013) in the hippocampus compared with those using
response strategies who had more gray matter in the
caudate nucleus (Sodums & Bohbot, 2020).
Infatti, many studies have demonstrated a significant
relationship between navigational ability and both healthy
ageing and neuropathology. Per esempio, older adults
display lower route learning and spatial learning during
navigation that does not improve to the same degree as
younger adults with training (Nemmi, Boccia, & Guariglia,
2017). This is thought to occur in ageing in part because of
increased path integration errors that accumulate with
travel distance (Stangl, Kanitscheider, Riemer, Fiete, &
Wolbers, 2020). When considering early neuropathology
associated with AD, asymptomatic preclinical AD patients
(biomarkers of beta amyloid and tau protein) show
reduced hippocampus-dependent wayfinding ability
compared with healthy controls (Allison, Fagan, Morris,
& Head, 2016), while these groups have shown no
difference when using conventional cognitive tests (Laczo
et al., 2022).
Given that atrophy in the hippocampus is associated
with age-related cognitive decline (Moffat et al., 2006;
Raz et al., 2004; Small et al., 2002; Lupien et al., 1998), Esso
is hypothesized that the use of hippocampus-dependent
spatial strategies, previously shown to be associated with
higher hippocampal volume, also decline with age. For
esempio, numerous studies have shown that the integrity
of the hippocampus is associated with healthy cognition in
ageing. Higher volume in the hippocampus is associated
with better learning and memory performance in ageing
as measured by virtual versions of the Morris Water Maze
and Transverse Patterning Discrimination tasks (Driscoll
et al., 2003) and higher estimates of global cognition as
measured by the Montreal Cognitive Assessment, a test
that is sensitive to dementia (Ritter, Hawley, Banks, &
Mugnaio, 2017; O’Shea, Cohen, Porges, Nissim, & Woods,
2016). Inoltre, lower gray matter in the hippocam-
pus and the functionally connected entorhinal cortex
(EC) are predictors of future diagnosis of AD (Apostolova
et al., 2006). Related to this, older adult participants who
use hippocampus-dependent spatial strategies had higher
scores on the Montreal Cognitive Assessment. These par-
ticipants also displayed better wayfinding performance
and increased hippocampal volume (Konishi, McKenzie,
Etchamendy, Roy, & Bohbot, 2017). Inoltre, spatial
strategies are associated with increased gray matter in
the hippocampus, despite the presence of an APOE4
allele. The APOE4 allele, present in approximately 10–
15% of people, increases the risk for AD, lowers the age
of onset, and is also associated with less gray matter in
the hippocampus (O’Dwyer et al., 2012; Pievani et al.,
2011) and the EC (Donix et al., 2010). È interessante notare, people
who are APOE4 carriers who use nonspatial strategies dis-
play decreased gray matter in the hippocampus and the
EC, whereas in contrast, APOE4 carriers who use spatial
strategies have gray matter levels comparable to non-
APOE4 carriers (Konishi et al., 2016). Così, continued
experience using spatial strategies is possibly associated
with increased gray matter in the hippocampus and EC,
which in turn could potentially reduce biological risk fac-
tors for developing dementia during ageing. Longitudinal
studies examining behavioral and imaging data are needed
to confirm the causal relationship between APOE4 status
and navigational strategies. Relatedly, it was found that
wayfinding performance, which is supported in part by
the hippocampus (per esempio., Konishi et al., 2017; Guderian
et al., 2015; Spiers, Burgess, Hartley, Vargha-Khadem, &
O’Keefe, 2001), within the mobile game Sea Hero Quest
(SHQ) was lower for APOE4 carriers compared with non-
APOE4 carriers (Coughlan et al., 2019).
Spatial strategies are also associated with lower volume
in the caudate nucleus, a structure involved in automatiza-
tion of behavior and reward ( West et al., 2018; Bohbot
et al., 2007; Iaria et al., 2003). Related to this, young adults
who use hippocampus-dependent spatial strategies dis-
play lower risk-taking behaviors in the Iowa Gambling
Task (Aumont, Blanchette, Bohbot, & West, 2019) E
report consuming fewer illicit substances (Bohbot, Del
Balso, Conrad, Konishi, & Leyton, 2013). Therefore,
hippocampal-dependent spatial strategy use is associated
with fewer reward-seeking behaviors.
Crucially, spatial learning, which depends on the use of
landmarks, declines throughout the life span. Relative to
young adult rodents, senescent rats shift from the use of
spatial memory strategies to the use of response strategies
to learn the location of a target arm in a plus maze (Barnes,
Nadel, & Honig, 1980). È interessante notare, when the striatum of
West et al.
453
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
C
N
_
UN
_
0
1
9
5
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
older rats was inactivated with lidocaine, they instead
favored the use of hippocampus-dependent spatial strate-
gies when tested on a similar task (Gardner, Gold, & Korol,
2020). These results suggest that the down-regulation of
striatal processing during ageing can help reinforce the
use of hippocampus-dependent strategies. When older
mice are given a choice between solving a maze using
response or spatial learning, they choose to use response
strategies although they demonstrated being capable of
using spatial strategies (Nicolle, Prescott, & Bizon,
2003). In humans, older adults tested on a virtual analog
of a rodent radial maze displayed decreased spatial strat-
egy use when completing the task (Etchamendy, Konishi,
Pike, Marighetto, & Bohbot, 2012). Another study testing
younger and older adults in a wayfinding task that allowed
participants to use either a flexible spatial strategy involv-
ing the use of efficient shortcuts or an inflexible response
strategy involving route-following found that older adults
consistently used the response strategy. This was in con-
trast to the younger adults who flexibly switched to the
more efficient spatial strategy during the wayfinding task
(Harris & Wolbers, 2014). Older adults display impair-
ment when required to revisit locations memorized rela-
tive to external landmarks, likely reflecting impairment of
relative positional encoding of environmental stimuli
(Bates & Wolbers, 2014). Aging has also been associated
with a decline in other landmark-dependent navigation
processes including impairment in linking directional
knowledge to the location of environmental landmarks
and decreased memory for the sequence in which land-
marks are encountered (Lester, Moffat, Wiener, Barnes,
& Wolbers, 2017; Zhong & Moffat, 2016; Head & Isom,
2010). Related to this, a cross sectional study in humans
examining the relationship between age and the use of
spatial strategies in a radial arm maze found that the
proportion of 8-year-old children who were identified as
spatial learners was approximately 85%, whereas this
proportion decreased to 50% in young adulthood and
then continued to decline to only 35% among people
over the age of 60 years in favor of the use of more rigid
stimulus response strategies (Bohbot et al., 2012). Simi-
larly, among 107 older adults between 55 E 80 years of
age, wayfinding performance linearly decreased with age
and correlated with a concurrent decrease of gray matter
within the hippocampus (Konishi et al., 2017).
The shift away from spatial strategies toward reliance on
habit-based response strategies might not simply be
because of ageing, but because of experience during
young adulthood. Per esempio, action video games (per esempio.,
Call of Duty) require players to perform many rapid
stimulus–response actions and navigate by way of the
external global positioning system (GPS) cues ( West
et al., 2018). When nonplayers were trained on 90 hr of
action video games (per esempio., Call of Duty), it was found that
the use of environmental landmarks to guide wayfinding
decreased and this correlated with gray matter loss in
the hippocampus ( West et al., 2018). Therefore, this study
shows that one’s everyday experiences can shape the brain
and could even affect how they perceive the world (Cia-
lone, Tenbrink, & Spiers, 2018). Further highlighting this
observation, a recent study found that people who rely
more on the in-car GPS systems to navigate, over time, dis-
played a steeper decline in hippocampal-dependent spa-
tial memory performance (Dahmani & Bohbot, 2020). In
a longitudinal segment of this study, GPS was found to
cause a decrease in the ability to use spatial landmarks
years later, independently of participants’ awareness of
their navigation ability. These results suggest that when
people are exposed to tasks or environments that pro-
mote response learning, it can reduce the ability to use
landmarks, reduce ability to learn the relationships
between landmarks, and is associated with a decreased
gray matter in the hippocampus, at any age, even in young
adults when brain tissue is healthy.
In the current study, we employed a mobile video game
designed to measure human spatial navigation ability through
gameplay—SHQ—to test if the use of landmark-dependent
navigation strategies changes across the life span. SHQ
is a virtual reality navigation task for mobile and tablet
devices that was funded by T-Mobile owned by Deutsche
Telekom and was designed as a possible early diagnostic
tool for AD (Spiers, Coutrot, & Hornberger, 2021;
Coughlan et al., 2018, 2019, 2020; Coutrot et al., 2018).
SHQ was recently further validated using real-world navi-
gation tasks demonstrating that performance in SHQ pre-
dicts performance in the real-world (Coutrot et al., 2019).
Whereas previous reports using SHQ focused on wayfind-
ing and path integration levels, here we analyze data from
the radial maze elements of the game, thereby providing
the first summary of this data set. One of the unique
elements of the radial maze is that it explores the use of
different strategies via direct behavior in the task, così
extending findings on navigation ability beyond those
based solely on self-assessment questionnaires not linked
to recent experience. The current study aimed to validate
and extend findings from radial arm maze tasks found in
smaller studies conducted in the laboratory using a larger,
more representative worldwide sample. Because land-
mark use is highly correlated with spatial navigation strat-
egies (Andersen, Dahmani, Konishi, & Bohbot, 2012) E
spatial learners display increased visual attention to tar-
gets in the environment (Drisdelle et al., 2017), we used
this measure as an approximation of the use of spatial
strategies within the SHQ Virtual Radial Maze. Using a
sample of over 37,000 participants, we examined whether
landmark use during the completion of the SHQ Virtual
Radial Maze varied across an age range between 19 E
69 years and whether there was an interaction with sex.
Spatial memory performance within the SHQ Virtual
Radial Maze was also recorded. We predicted that, as in
previous studies conducted in the laboratory suggest,
navigational strategies that depend on the use of land-
marks will decline with age in favor of more rigid
stimulus–response strategies. We also predicted that, COME
454
Journal of Cognitive Neuroscience
Volume 35, Numero 3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
C
N
_
UN
_
0
1
9
5
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
demonstrated by previous studies, landmark-dependent
navigation will be associated with better cognitive per-
formance within the SHQ Virtual Radial Maze.
METHODS
Game Design
As detailed in Coutrot et al. (2018) and Spiers et al. (2021),
SHQ was developed in collaboration with the indepen-
dent video games design company Glitchers Ltd and used
Unity 3D (Unity Technologies) for smart phones and
tablets (apple and android devices). SHQ (https://www
.alzheimersresearchuk.org/research/for-researchers
/resources-and-information) was released on May 4, 2016,
on the App Store for iOS and on Google Play for Android
and was available in 17 languages. Through the game, par-
ticipants followed a sea captain as he tries to recover his
father’s lost memories. The player manipulates the game
through four controls, (cioè., tap left to turn left, tap right to
turn right, swipe up to speed up, and swipe down to halt).
Players were asked a set of optional questions, Quale
included their age, sex and nationality.
SHQ was designed to reproduce classic navigation tasks
used in the literature. Players were tasked to complete
game levels that tested performance in three different
types of tasks found in the literature. The data from the
first two task types, wayfinding and path integration, Avere
been described in Coutrot et al. (2018). The third task
type, performance in a virtual radial maze (SHQ Virtual
Radial Maze), has previously not been examined and is
the focus of the current research report. The experimental
tasks in SHQ were accessed by unlocking levels sequen-
tially. These levels were grouped into five themed areas,
each containing 15 levels that contained the wayfinding
and path integration tasks. At the end of each themed area,
players were asked to complete a bonus level (five in
total), which consisted of the SHQ Virtual Radial Maze,
which we focus on in this article. The completion of the
bonus SHQ Virtual Radial Maze levels was optional, E
players had the option to skip them to progress to the next
themed area.
were used during navigation. At the beginning of each
SHQ Virtual Radial Maze level, the player’s boat in which
they control begins in the center of a platform surrounded
by six radial arms (Guarda la figura 1). The environment that
surrounds the radial arms contains both proximal and dis-
tal landmarks (per esempio., icebergs, animals, volcanoes, moun-
tains). The player is instructed to collect as many stars
as possible. In Part 1, three of the six radial arms are
blocked and the player’s goal is to visit the three available
arms. When the player enters an arm, a star appears and is
collected. When all three stars are collected, the player
moves on to the second part of the level. Players were
unable to see the stars from the center; Perciò, Essi
had to remember which arms contained stars and which
did not to perform the second part of the task without
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
C
N
_
UN
_
0
1
9
5
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
SHQ Virtual Radial Maze Task
The focus of the current article centers around perfor-
mance in the SHQ Virtual Radial Maze levels based on
radial mazes used in the rodent and human literatures
(Bohbot et al., 2007; Iaria et al., 2003; Packard et al.,
1989; Olton & Papas, 1979). Specifically, the SHQ Virtual
Radial Maze task was modeled after the 4 SU 8 virtual maze
(4/8VM), a computerized behavioral task used in the lab-
oratory to assess learning and memory performance and
navigation strategy (Bohbot et al., 2007; Iaria et al., 2003).
The player is presented with a novel environment with
distinct landmarks in each level. The SHQ Virtual Radial
Maze was used to assess the degree to which landmarks
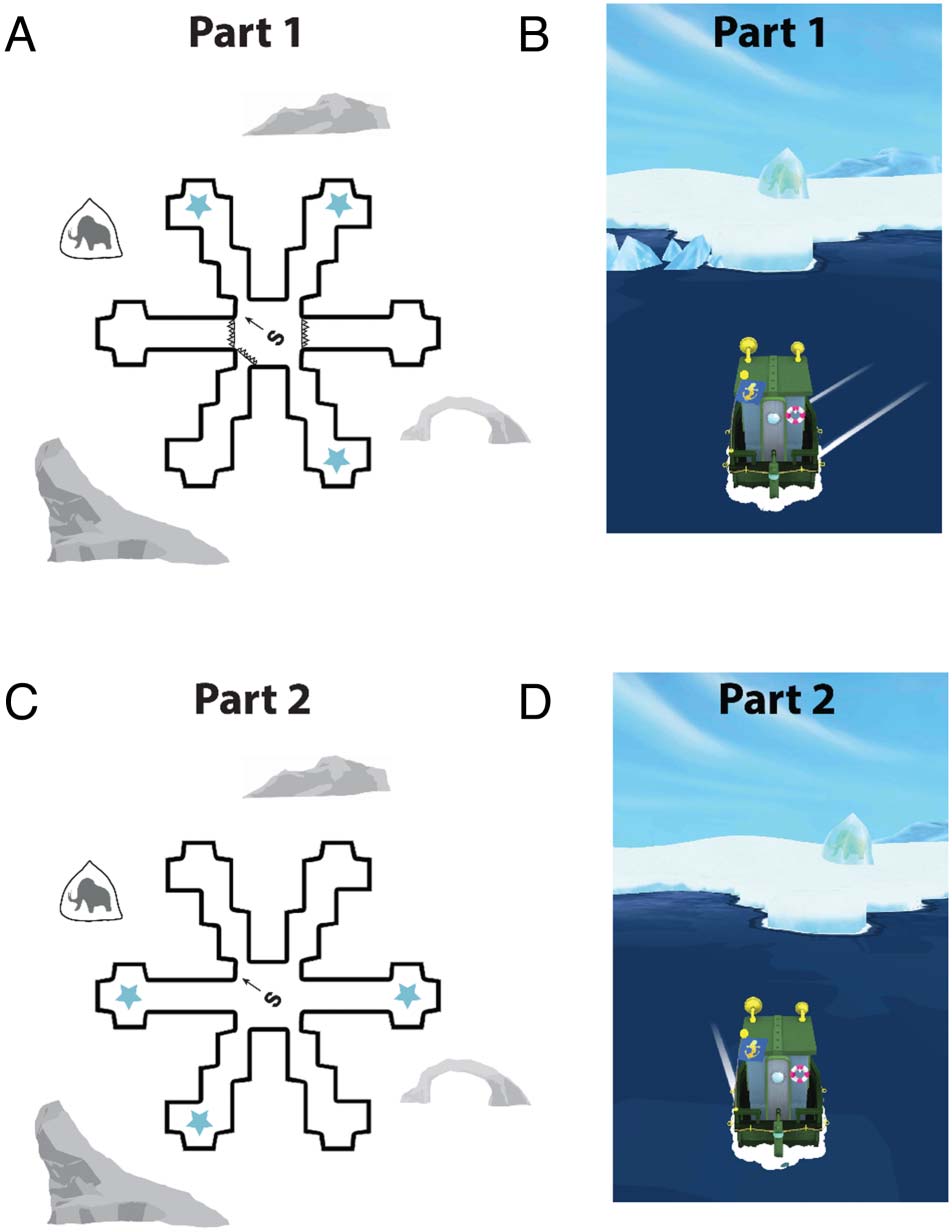
Figura 1. An example of a SHQ Virtual Radial Maze level. The radial
maze levels contain six pathways surrounded by proximal and distal
landmarks (per esempio., ice bergs, mountains) and consist of two parts. Part 1 È
shown from both an overview perspective (UN) and a third-person
perspective. (B) Three of the six pathways are blocked, and the player is
required to navigate the boat to visit the three open pathways to collect
a star that pops out of the water. Part 2 is shown from both an overview
perspective (UN) and a third-person perspective. (B) All six pathways are
made available, and the player is required to visit the pathways that
were unavailable during Part 1 to collect the remaining three stars.
Reference memory errors were defined as entering a pathway entered
during Part 1 that needed to be avoided during Part 2. Working memory
errors were defined as re-entering a pathway in Part 2 that was already
entered previously during Part 2. When the radial arm maze level is
completed, the player is asked a multiple-choice question about how
they found the stars (cioè., a counting-dependent strategy vs. a landmark-
dependent strategy).
West et al.
455
errors. In Part 2, all six radial arms are now open. IL
player’s goal in Part 2 is to visit the three radial arms that
were blocked during Part 1 and collect the remaining
three stars. Because 1) the SHQ Virtual Radial Maze levels
are very different compared with the standard levels and
only begin to be presented to the player after 15 standard
levels are completed and 2) no overt instructions are
given to the player about the goal of the level, SHQ Virtual
Radial Maze Level 1 was counted as a practice trial. It
should be noted that this makes the SHQ Virtual Radial
Maze different from the 4/8VM because the latter includes
detailed instructions to the participants.
Two types of memory errors are recorded during Part 2
of SHQ Virtual Radial Maze levels. Primo, a “reference mem-
ory error” is recorded when a participant visits a radial arm
that does not contain an object in Part 2, questo è, entering a
radial arm that they previously visited in Part 1 of that trial
and needed to avoid during Part 2. A “spatial working
memory error” is recorded in Part 2 when a participant
visits an arm that they had already visited before collecting
all three stars, questo è, entering a radial arm that they
already visited in the same Part 2 of that trial.
While data from these game levels did not provide specific
details about navigational strategies used (per esempio., did you use
the relationship between multiple landmarks), reports of
whether landmarks were used or not to complete the radial
maze were obtained. To assess the degree to which land-
marks were used, players were presented with a multiple-
choice question after each SHQ Virtual Radial Maze level that
asked players what they did to remember which radial arms
they needed to visit and which ones they needed to avoid:
“How did you navigate?" (1) Counted from the start; (2)
used multiple landmarks; (3) counted from a landmark. UN
player was identified as using landmarks or not using land-
marks when solving the maze based on their answer to a
question after the first experimental level was complete
(SHQ Virtual Radial Maze Level 2, which was done after
the practice trial in Level 1). If a player answered that they
used a landmark to navigate, they were categorized as using
a landmark-based strategy (cioè., Answer 2 O 3). In contrasto, if
the player indicated that they counted the radial arms from
the start and did not use any landmarks (cioè., Answer 1), Essi
were categorized as spontaneously using a counting strat-
egy. An option to skip this question was also provided; if this
was chosen, their strategy was classified as “none reported”
and was not included in the analysis.
There are several important differences between the
SHQ Virtual Radial Maze and the 4/8VM that is used in
the laboratory. Primo, a detailed verbal report from the
player about how they completed the task was not
obtained, as is the case with the 4/8VM, and we therefore
could not distinguish between people’s strategies that
used the relationship between multiple landmarks (spatial
strategy) and strategies that used a single landmark or mul-
tiple landmarks in conjunction with a rigid pattern or
counting (response strategy). Because of this, we com-
bined people into two groups: People reported using a
strategy that relied on landmarks and people who
reported using a strategy that relied on counting only.
Therefore, this task is able to distinguish between people
who were inclined to use external landmarks during the
resolution of the task and those who were not. Tuttavia,
the current methodology cannot distinguish between
more fine-grained strategy differences as done in the past
based on data collected in the laboratory (per esempio., Aumont,
Arguin, Bohbot, & West, 2019; West et al., 2018; Bohbot
et al., 2007, 2013; Konishi & Bohbot, 2013; Iaria et al.,
2003). Secondo, because of time constraints, there were
six arms in total instead of eight. Third, the SHQ Virtual
Radial Maze only presented players with each environ-
ment once per level (cioè., one trial), whereas the standard
4/8VM repeats a minimum of three trials in the same envi-
ronment and required people to learn the environment.
Because the SHQ Virtual Radial Maze does not repeat
the same environment across multiple trials, we only com-
pared memory errors within Parts 1 E 2 of a single trial.
Our operationalization of a reference memory error in the
SHQ Virtual Radial Maze therefore differs from previous
reports using the 4/8VM: A reference memory error in
the SHQ Virtual Radial Maze is recorded when a partici-
pant visits a radial arm that does not contain an object in
Part 2 of the same trial; in contrast, a reference memory
error in the 4/8VM is recorded when a participant visits a
radial arm that does not contain an object in Part 2 Di
multiple trials that repeat the same environment.
Data Collection
Within the opening screen menu, participants were made
aware of the purpose of the game. They were then asked
to give their consent to share their data. Inoltre, IL
opt out was always available in the game’s settings. Data
integrity and data privacy were adhered to according to
German data security law. For full details please see
Coutrot et al. (2018).
Data Analysis
We did not include data of people who indicated that they
were 18 years old as this was the minimum age required to
consent to data sharing and likely contained a large num-
ber of people who were under the age of 18 years. In other
parole, people under the age of 18 years who accessed the
game more than likely selected the lowest age possible,
adding a potential confound. Significantly lower sample
sizes were obtained for people older than 69 years, so
we chose this age as the higher end cutoff. To analyze
the factor of age, we organized the data in bins that ranged
10 years (19–29, 30–39, 40–49, 50–59, 60–69). As stated
above, participants completed a first practice level before
completing the experimental levels. We chose to focus our
analyses on SHQ Virtual Radial Maze Level 2 (cioè., the first
experimental level) because (UN) this level provided data for
the initial measures of spontaneous navigation strategy
456
Journal of Cognitive Neuroscience
Volume 35, Numero 3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
C
N
_
UN
_
0
1
9
5
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Tavolo 1. Participant Breakdown by Age, Sex, and Strategy
19–29
30–39
n = 3338
n = 4742
n = 1000
n = 1215
Age
40–49
n = 506
n = 590
n = 5439
n = 8937
n = 2775
n = 3705
n = 1399
n = 1713
Female
Counting
Landmark
Male
Counting
Landmark
50–59
60–69
n = 234
n = 324
n = 516
n = 602
n = 117
n = 128
n = 154
n = 151
used and spatial memory performance within the radial
maze and, (B) as noted in Coutrot et al. (2018), the sample
size falls dramatically the later into the game the sample is
taken from (Coutrot et al., 2018). Other demographics
included sex and home environment. Home environment
was analyzed because of the fact that people who grow up
in rural environments are hypothesized to have more
opportunity to use distal landmarks during navigation while
people who grow up in cities are more reliant on Cartesian
coordinates, thereby engaging spatial memory to a signifi-
cantly lower degree. For simplicity, home environment
was collapsed across noncity categories (rural, suburbs,
mixed) and was compared with those participants who
reported living in cities (Coutrot et al., 2018, 2019, 2022).
Data Analysis and Predictions
Linear and logistic regressions were performed using
MATLAB’s fitglm function (MATLAB and Statistics Toolbox
Release 2022b, The MathWorks, Inc.). Odd ratios were
calculated by taking the exponential of the model estimate
of the relevant variable (per esempio., esp(β1)). Follow-up effects
sizes (mes/mestab) and two-way ANOVAs (mes2way) were
calculated using Hentschke’s effect size toolbox as
described in Hentschke and Stuttgen (2011). Computa-
tion of measures of effect size for neuroscience data sets
(Hentschke & Stuttgen, 2011).
RESULTS
Participants
Participants were included in analyses if they (UN) provided
their age and sex, (B) completed a minimum of the first
two SHQ Virtual Radial Maze levels (cioè., the practice level
[SHQ Virtual Radial Maze Level 1] and the first experi-
mental level [SHQ Virtual Radial Maze Level 2]), (C)
reported a navigation strategy, E (D) were between
the ages of 19 E 69 years. Six thousand thirty-seven
did not report a navigation strategy and therefore were
not included in the analyses. This resulted in a total n =
37,585 being included for analysis (n = 12,194 women
(mean age [SEM] = 29.3 [0.09]); n = 25,391 men (mean
age [SEM] = 30.4 [0.06])). See Table 1 for a total break-
down by age range, sex, and navigational strategy.
Planned a priori comparisons were made to examine the
factors of age and sex when analyzing the dependent
measures of navigational strategy, reference memory
errors, and spatial working memory errors.
Navigational Strategy
We first analyzed the spontaneous navigational strategies
people used to complete SHQ Virtual Radial Maze Level 2.
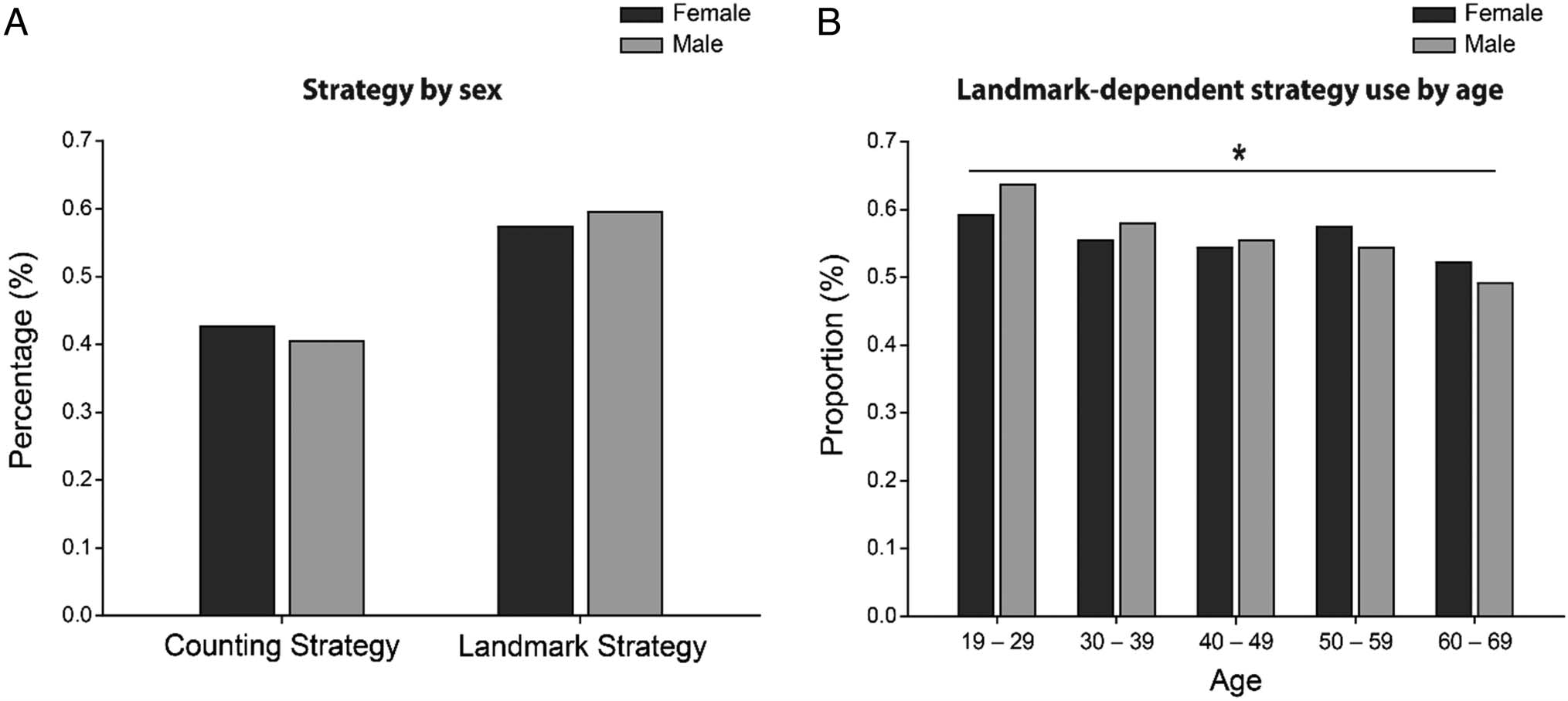
Overall, 42% of participants reported using a counting-
based strategy whereas 58% of participants reported
using a landmark-based strategy. We ran a logistic regres-
sion to predict navigational strategy, based on age,
sex, and home environment. Sex (odds ratio = 1.415,
T(35186) = 4.32, P < .001) and age (odds ratio = 0.9929,
t(35186) = −3.49, p < .001) were both significant predic-
tors, as was the interaction between age and sex (odds
ratio = 0.9911, t(35186) = −3.52, p < .001), chi-square
statistic versus constant model: 161, p < .001. There
was no effect of home environment or any other interac-
tions. To more closely examine these effects, we ran
direct comparisons between the predictor variables using
the age bins (as described in Methods section) and inter-
preted the resultant effect sizes. We found that women and
men employed a counting-based strategy (women = 43%;
men = 40%) and a landmark-based strategy (women =
57%; men = 60%) at similar rates (phi = 0.02, CI [0.01,
0.03]; see Figure 2A). Furthermore, when exploring
strategy use differences between sexes within age bins,
we found negligible effects (all phi < 0.1). Importantly,
we did observe that overall landmark-dependent strategy
use declined with age (Figure 2B). Because of the signifi-
cant Age × Sex interaction that predicted navigational
strategy, we conducted separate regressions examining
changes in strategy with age for both male and female indi-
viduals. We found men showed a stronger decline in
landmark-dependent navigation strategy use with age
compared with women (odds ratio = 0.865, t(23766) =
−10.32, p < .001; frequency across age bins: Cramer’s
West et al.
457
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
c
n
_
a
_
0
1
9
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
c
n
_
a
_
0
1
9
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. (A) Percentage of players reporting a counting- versus landmark-dependent strategy by sex. (B) Proportion of players reporting a landmark-
dependent strategy by sex and by age bin.
V = 0.066 [0.05, 0.08], chi-square = 110.1; females: odds
ratio = 0.937, t(11426) = −3.37, p < .001; frequency
across age bins: Cramer’s V = 0.04 [0.03, 0.06], chi-
square = 19.8) (Figure 2B). Thus, sex and age do seem
to be relevant to spontaneous strategy use—however, it
should be noted that there are large sample size differ-
ences among the age and sex groups (see Table 1).
Spatial Memory Performance
Both reference and spatial working memory errors in
SHQ Virtual Radial Maze Level 2 were compared between
people who used a landmark-based or counting-based
strategy. We first examined reference memory errors
that occurred when a participant visited a radial arm that
did not contain an object, that is, a radial arm that they
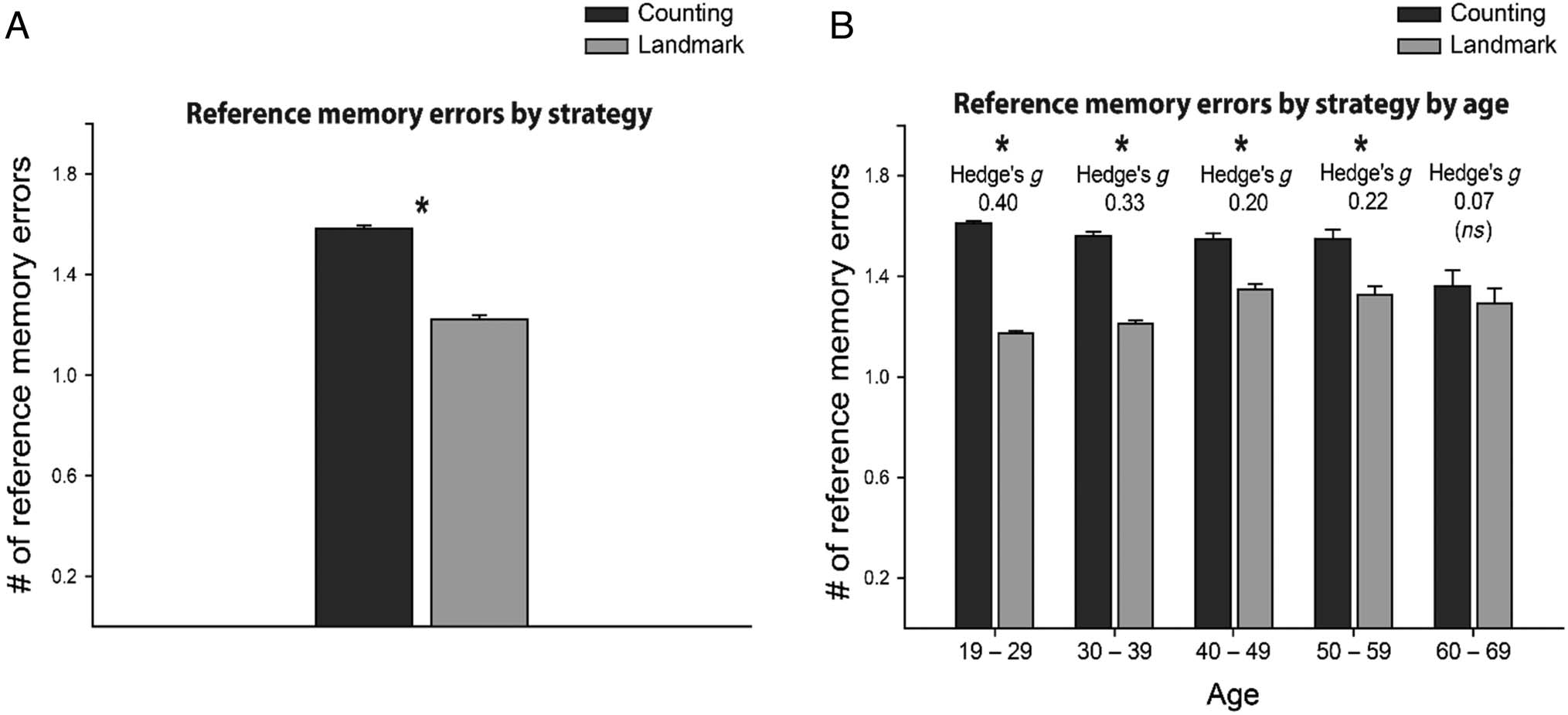
previously visited in Part 1 of that trial. People reporting
a landmark-dependent strategy made fewer overall refer-
ence memory errors (M = 1.21, SEM = 0.007) compared
with those who used a counting-based strategy (M =
1.58, SEM = 0.008; Figure 3A). We then ran a linear
regression model to predict reference memory errors
based on age, sex and navigational strategy. We found
Figure 3. (A) Reference memory errors made by players reporting using a counting versus landmark-dependent strategy. (B) Reference memory
errors by players reporting using a counting versus landmark-dependent strategy by age bin.
458
Journal of Cognitive Neuroscience
Volume 35, Number 3
a significant effect of Strategy (beta [SD] = 0.46 [0.06],
t(37577) = 8.12, p < .001), Sex (beta [SD] = −0.26
[0.05], t(37577) = −5.50, p < .001), as well as a signifi-
cant interaction between all factors (Age × Sex: [beta
(SD) = 0.006 (0.002), t(37577 ) = 4.13, p = .001];
Age × Strategy: [beta (SD) = −0.005 (0.002), t(37577) =
−2.49, p = .013]; Sex × Strategy: [beta (SD) = 0.25
(0.071), t(37577) = 3.55, p < .001]; Age × Sex × Strat-
egy: [beta (SD) = −0.006 (0.002), t(37577) = −2.62, p =
.008], F-statistic versus constant model: 173, p < .001.
Briefly, when including home environment in this model,
we found no significant effects of this predictor or inter-
actions with it. To interpret these effects related to refer-
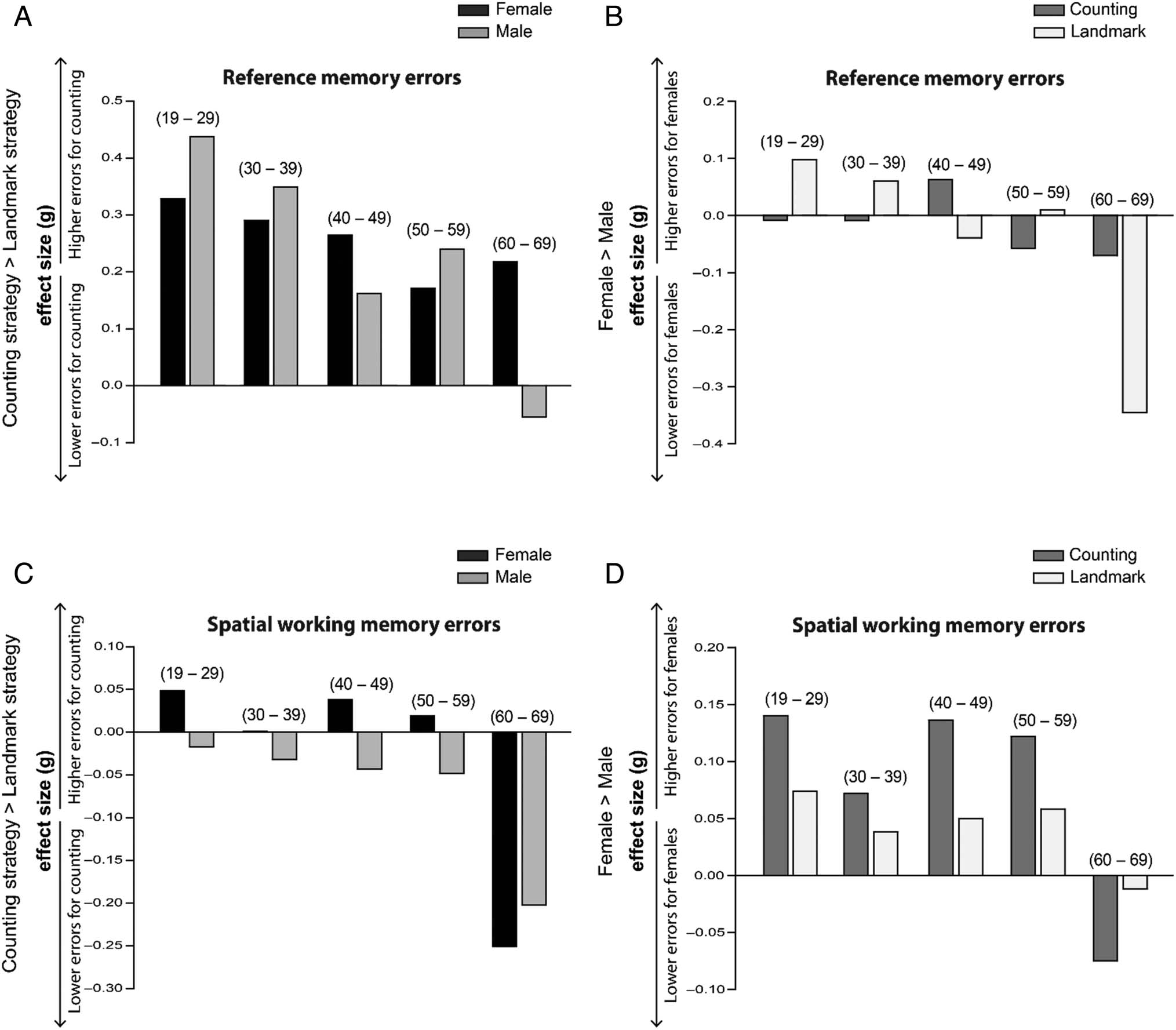
ence memory errors, we examined the effect sizes. When
we compared reference memory errors within strategy
type, males showed a small advantage when using a
landmark-dependent strategy (Hedge’s g(CI) = 0.06
[0.03, 0.09], t(22105) = 4.32, p < .001) but not when
using a counting strategy (Hedge’s g(CI) = 0 [−0.03,
0.03], t(15476) = 0.028, p > .9). Tuttavia, both woman
and men showed that landmark-dependent strategy use
was associated with fewer reference memory errors
(women Hedge’s g (CI): 0.31 [0.27, 0.34], T(12192) =
16.7, P < .001; men Hedge’s g (CI): 0.37, [0.34 0.39],
t(25389) = 28.8, p < .001). To look at the effect of
age, we first collapsed across sex (based on the above
analysis), and found there was a small to medium effect
of strategy type across all age bins showing landmark-
dependent strategy use was associated with fewer
errors, except in the oldest 60–69 age bin (all Hedge’s
g > 0.19, T(> = 1674) > = 4.44, P < .001; except last age
bin: Hedge’s g = 0.07, t(548) = 0.81, p = .42; see
Figure 3B). However, when looking at men and women
separately within each age bin, we observed an effect of
strategy use on reference memory errors in the 60–69 age
bin (see Figure 4A and 4B). To investigate this further, we
ran a factorial analysis (two-way ANOVA) of Sex and
strategy in the 60–69 age bin, and found a significant
effect of sex (F = 5.7, p = .02, eta-square = .01), with
women relying on a landmark-dependent strategy show-
ing better reference memory (M = 1.10, SEM = 0.08) than
their male landmark-dependent counterparts (M = 1.45,
SEM = 0.08; Hedge’s g (CI) = 0.34 [0.11, 0.58], t(277) =
2.87, p = .004). These landmark-dependent women also
performed better than their female counting counterparts
(M = 1.32, SEM = 0.10; Hedge’s g (C)I) = 0.22 [−0.03,
0.47], t(243) = 1.71, p = .09). This confirmed that within
the 60–69 age range, women using landmarks displayed
better reference memory performance compared with
men overall and women who used a counting strategy.
Of note, although this age category was the least popu-
lated, we have similar participant numbers across the
groups compared, and a large sample size compared with
laboratory testing (all 116 > n > 155; Vedi la tabella 1).
We next examined spatial working memory errors,
which were recorded during Part 2 when a participant
revisited an arm that they had already visited before
collecting all three stars. People reporting a landmark-based
strategy made similar working memory errors (M = 0.18,
SEM = 0.003) compared with those who used a counting-
based strategy (M = 0.18, SEM = 0.004). We ran a linear
regression model on spatial working memory errors with
the same predictors as above and found a significant
effect of Age (beta [SD] = 0.002 [0.001], T(37577) =
3.06, p = .002), and Strategy (beta [SD] = 0.058 [0.025],
T(37577) = 2.30, p = .02), and a trend for Sex (beta
[SD] = −0.037 [0.02], T(37577) = −1.81, p = .069). There
was a trend for significant interactions between Sex
and Strategy (beta [SD] = −0.054 [0.03], T(37577) =
−1.71, p = .09), and a trend toward a significant interac-
tion between Age and Strategy (beta [SD] = −0.0015
[0.0008], T(37577) = −1.81, p = .07), F-statistic versus
constant model: 15.3, P < .001. Again, we found no effect
of home environment when included in the model.
When examining the effect sizes related to spatial
working memory errors (see Figure 4C and D) there was
a negligible effect size of Sex (Hedge’s g = 0.08 [0.06,
0.11], t(37583) = 7.76, p < .001). There was a negligible
effect size of Sex within strategy types, but nevertheless,
men showed an advantage when using a landmark-
dependent strategy (Hedge’s g[CI] = 0.06 [0.03, 0.09],
t(22105) = 4.26, p < .001) and when using a counting
strategy (Hedge’s g[CI] = 0.12 [0.09, 0.15], t(15476) =
6.9, p < .001). There was no effect of Strategy when
collapsing across Sex (Hedge’s g = 0.003 [−0.018, 0.023],
t(37583) = 0.27, p = .79), and this pattern was reflected
in every age bin (all Hedge’s g < 0.03, t(> = 1674) < =
−1.04, p > 0.2) with the exception of people who were
60–69 years old, where there was a small effect of strategy
in that both women and men showing fewer errors asso-
ciated with using the counting strategy (F = 7.002, p =
.008, eta-square = 0.013, Hedge’s g = −0.22 [−0.06–
0.39], T(548) = −2.62, p = .009), women: counting: M =
0.17, SEM = 0.04; landmark: M = 0.30, SEM = 0.05;
Hedge’s g = −0.25 [−0.5, 0.001], T(243) = −1.97, p =
.051; men: counting: M = 0.20, SEM = 0.04; landmark:
M = 0.31, SEM = 0.05; Hedge’s g = −0.20 [−0.43, 0.03],
T(303) = −1.77, p = .078; see Figure 4C and 4D).
Control for Time to Complete Radial Maze Level
Although there was no time constraint or instruction to
complete the radial maze in a certain time limit, we found
that there was a general slowing of completion time (for
simplicity, we refer to this measure as “RT”) with age. Fur-
thermore, based on the diminishing effect sizes with age
displayed in Figure 4A and 4B, it appears that the cost of
using a counting strategy on reference memory error per-
formance could diminish with age. To account for the
slowed RT with age and to explore errors in the context
of ageing in more detail, we normalized errors as a function
of RT ([RT × error] + error). Così, errors would be fur-
ther penalized for slow completion time (for zero errors,
we used the value of the raw RT). A regression of the
West et al.
459
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
C
N
_
UN
_
0
1
9
5
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
C
N
_
UN
_
0
1
9
5
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
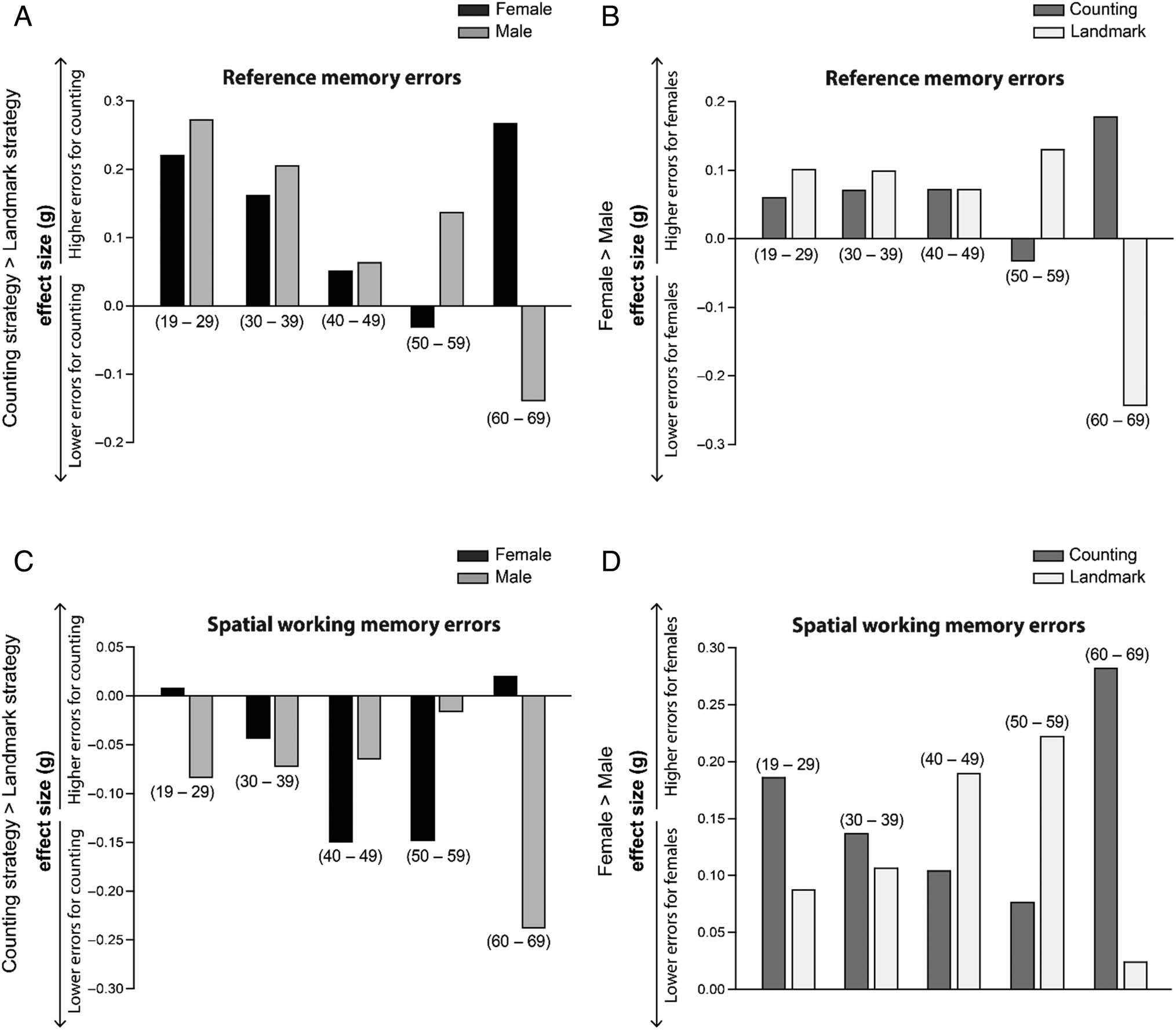
Figura 4. Effect sizes by age bin displayed for both reference memory errors (A and B) and spatial working memory errors (C and D). Age bins are
indicated in parentheses. In all figures, higher effect sizes represent higher error levels. Separate comparisons were made for navigation strategy and
sex. For navigation strategy (A and C), a higher effect size represents more errors for people using the counting strategy compared with the landmark
strategy (effect sizes for men and women are plotted separately). For sex (B and D), a higher effect size represents more errors for women compared
with men (effect sizes for the counting groups and landmark groups are plotted separately). Within the 60–69 age range, women using the landmark
strategy displayed better reference memory performance compared with men overall and women who used a counting strategy (A and B). In parallel,
60- to 69-year-olds who used the counting strategy displayed fewer spatial working memory errors overall (C and D).
normalized reference memory errors showed a signifi-
cant effect of Age (beta [SD] = 0.014 [0.003], T(37577) =
5.42, P < .001), Sex ( beta [SD] = −0.34 [0.10],
t(37577) = −3.37, p < .001) and Strategy (beta [SD] =
0.76 [0.12], t(37577) = 6.17, p < .001) and an interaction
between Age × Strategy (beta [SD] = −0.011 [0.004],
t(37577) = −2.81, p = .005). See Figure 5A and 5B for
effect sizes. Overall, the pattern of results was very similar
to the previous reference memory error analysis; how-
ever, those employing a counting strategy no longer
showed an age-related decline in errors when RT is taken
into consideration.
A regression of the normalized spatial working memory
errors showed a significant effect of Strategy (beta [SD] =
0.13 [0.06], t(37577) = 2.40, p = .02), and Age (beta
[SD] = 0.012 [0.001], t(37577) = 9.95, p < .001), as well
as a significant interaction between all factors (Age ×
Sex: (beta [SD] = −0.005 [0.001], t(37577) = −3.36, p <
.001); Age × Strategy: (beta [SD] = −0.006 [0.002],
t(37577) = −3.21, p = .001); Sex × Strategy: (beta
[SD] = −0.21 [0.070], t(37577) = −2.97, p = .003);
Age × Sex × Strategy: ( beta [SD] = 0.006 [0.002],
t(37577) = 2.53, p = .01). See Figure 5C and 5D for effect
sizes. Overall, the pattern was similar to the previous
460
Journal of Cognitive Neuroscience
Volume 35, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
c
n
_
a
_
0
1
9
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Normalized errors accounting for variations in RT with age. Effect sizes by age bin displayed for both normalized reference memory errors
(A and B) and normalized spatial working memory errors (C and D). Age bins are indicated in parentheses. In all figures, higher effect sizes represent
higher error levels. Separate comparisons were made for navigation strategy and sex. For navigation strategy (A and C), a higher effect size represents
more errors for people using the counting strategy compared with the landmark strategy (effect sizes for men and women are plotted separately).
For sex (B and D), a higher effect size represents more errors for women compared to men (effect sizes for the counting groups and landmark
groups are plotted separately). When RT is included in the model, there are no declines in errors associated with age. Furthermore, within the
60–69 age range, women using the landmark strategy again display better reference memory performance compared with men overall and females
who used a counting strategy (A and B). In parallel, within the 60- to 69-year-old range, only men who use the counting strategy displayed fewer
spatial working memory errors (C and D).
spatial working memory error analysis; however, women
showed worse performance overall, and the advantage
seen in the counting group in the 60–69 age range was
restricted to men when RT is taken into consideration.
Radial Maze Errors in Relation to
Wayfinding Ability
We obtained wayfinding ability scores for our participants
from the larger pool of data reported in Coutrot et al.
(2018), which is a video-gaming-skill corrected value for
overall spatial navigation abilities (reported as “OPcorr”).
We compared wayfinding ability between those using a
landmark (M = 141.8, SEM = 1.42) and a counting-based
(M = 134.9, SEM = 1.71) strategy and found no difference
(Hedge’s g = 0.04 [0.01, 0.06])—the same held when
comparing within sex or across age groups. We also
found no correlation between wayfinding ability and the
number of reference or spatial working memory errors
(all r < −.05).
West et al.
461
Table 2. Proportion of Players Who Reported Using a Landmark-
dependent Strategy in Each Country Who Had at Least 500 Players
Completed the First Two SHQ Virtual Radial Maze Levels
Country
France
Spain
Greece
Poland
Brazil
Hungary
Slovakia
Czechia
Italy
Canada
Netherlands
China
United States
Australia
Germany
United Kingdom
% of Participants Reporting a
Landmark-dependent Strategy
69%
67%
67%
66%
66%
64%
61%
61%
59%
59%
57%
57%
57%
56%
55%
51%
n
919
614
2600
1511
576
1471
881
2869
1326
1133
2626
817
8579
794
3537
5593
Country and Navigational Strategy
We conducted an exploratory analysis examining by coun-
try the proportion of people who used a counting versus
landmark-based strategy. We reported the country data if
they had at least 500 participants. The data are shown in
Table 2 and reflect that there is variability among countries
with respect to the proportion of participants who com-
pleted the SHQ Virtual Radial Maze using a counting ver-
sus landmark-based strategy. No evident pattern was
observed in this sample of countries.
DISCUSSION
In the current study, we examined data collected from a
radial arm maze (SHQ Virtual Radial Maze) contained in
the SHQ mobile video game. The SHQ Virtual Radial Maze
was designed after radial arm mazes used in the human
and rodent literatures (Bohbot et al., 2007; Iaria et al.,
2003; Packard & McGaugh, 1992; Packard et al., 1989;
Olton & Papas, 1979), and allowed for the testing of our
hypotheses generated from these previous smaller studies
with a significantly larger, diverse sample of the world pop-
ulation normally inaccessible to researchers. Our results
support previous findings showing that, during ageing,
people shift away from landmark-dependent navigational
strategies, which previous studies have shown is
supported by the hippocampus, during early adulthood
in favor of stimulus–response navigational strategies later
in life (Bohbot et al., 2012). This reveals that the decreased
reliance on landmarks during ageing is not simply present
with late-life ageing, but is rather progressive from ∼30
years of age onward, similar to the general overall decline
in spatial ability previously observed with the wayfinding
task in SHQ (Coutrot et al., 2018). Previous, smaller sam-
ple studies have found no significant overall sex difference
with respect to the use of landmark-dependent or
stimulus–response navigation strategies (Konishi &
Bohbot, 2013; Konishi et al., 2013; Bohbot et al., 2007;
Iaria et al., 2003). This stands in contrast to the robust dif-
ferences in sex observed in the wayfinding task in SHQ
(Coutrot et al., 2018). It suggests that the difference
between men and women in wayfinding may not be driven
by differences in landmark or stimulus–response strate-
gies, as we found no relationship between strategy use
and wayfinding performance.
In relation to errors committed on the radial maze, we
found a more complex relationship of strategy. Reference
memory was better for people using landmark-dependent
navigation strategies overall, even in older age ranges up
until 50–59 years. Importantly, the women in the 60–69 age
group using landmark-based strategies outperformed
their male counterparts as well as their female counting
counterparts on reference memory. In contrast, more spa-
tial working memory errors were committed by the same
60–69 age group relying on landmark-based strategies.
What might cause this discrepancy between reference
and working memory errors? When there are no delays
or reorientation of view, as is the case for spatial working
memory during Part 2 of the SHQ radial maze, using a
counting strategy might be more efficient while at the
same time less flexible, as are stimulus–response strate-
gies in general. This could in turn result in fewer errors.
Indeed, response learners make fewer non-hippocampus-
dependent working memory errors in the laboratory
(Blanchette et al., 2020; Aumont, Blanchette, et al., 2019;
Bohbot et al., 2007; Iaria et al., 2003).
Our finding supports previous observations from
smaller studies that show that hippocampus-dependent
spatial learning, which relies on landmarks during naviga-
tion, declines with age. The present findings could point to
a relationship between increased landmark use (i.e., spa-
tial strategies) and preserved spatial memory performance
during ageing (Konishi et al., 2017). This might be espe-
cially important in women because compared with men
overall and women who use spatial strategies, women
who use stimulus–response strategies display the lowest
amount of gray matter in the hippocampus and the high-
est amount of gray matter in the caudate-nucleus (Sodums
& Bohbot, 2020), which is associated with greater risk of
developing AD (Persson, 2018; Apostolova et al., 2006).
Finally, we did not find any notable effects of home
environment on strategy use or errors committed. As
mentioned above, there was no effect of strategy use
462
Journal of Cognitive Neuroscience
Volume 35, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
c
n
_
a
_
0
1
9
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
on wayfinding ability, nor any relationship between way-
finding and reference or spatial working memory errors.
This suggests that these tasks within SHQ (radial maze
vs. wayfinding) measure separate skills. Note, however,
that for comparability with existing reports on SHQ, only
a basic measure wayfinding ability, that is, trajectory
length, of a few select levels was included in our analyses.
This parameter may not be the most relevant to correlate
with radial maze performance, and future studies that
focus on more sophisticated features from the trajectories
during wayfinding might be more sensitive to differences
between people using different navigation strategies.
There were limitations contained in the current study.
First, there was a degree of bias selecting for those who
completed the SHQ Virtual Radial Maze because of the fact
these radial arm maze game levels were nonmandatory
(i.e., they could be skipped) and were offered as a bonus
level between each of the main parts of the game that
included the wayfinding and path integration tasks
(reported in Coutrot, 2018). Furthermore, people had to
progress beyond the first 30 levels of the game to com-
plete the first two SHQ Virtual Radial Maze levels (i.e.,
the practice level and the first experimental level) and be
included in our analysis. This likely resulted in selecting for
higher performing individuals in our analyses. Even with
this limitation, we observed significant declines in
landmark-dependent navigation strategy use with age,
which was associated with lower spatial memory perfor-
mance. Another limitation was, unlike the 4/8VM, there
was a lack of one-on-one testing and specific instruction
about how to complete the task to ensure that the partic-
ipant paid attention, understood the task, and did not sim-
ply enter the pathways randomly without any thought
about how to properly complete the task. Indeed, it is pos-
sible that that the differences we report could reflect those
between people who chose to complete the task in an
effortful fashion compared with those who did not. For
example, we cannot exclude the possibility that people
who reported using landmarks tended to be those who
pay attention to the task and notice the environment com-
pared with those who reported using a counting-only strat-
egy. We should, however, also consider the fact that 1) the
SHQ Virtual Radial Maze levels were completely optional,
2) participants had to complete 30 standard levels before
completing the experimental SHQ virtual maze level
where we ascertained whether or not people used land-
marks, and 3) people had the option to not answer the
question asking about landmark use if they wished, in
which case their data were not included in our analyses.
Therefore, we can infer that the sample of people for
which we do have complete data to include in the current
analyses were fairly motivated and likely represent behav-
ioral differences between people who were inclined to use
landmarks in some form to complete the task compared
with those who did not use landmarks and used a counting
strategy. This is especially relevant for the findings in the
oldest age group. Another important limitation is that our
assessment of navigational strategies in the SHQ Virtual
Radial Maze could not distinguish between more granular
navigational strategies that are able to be identified in the
laboratory. In the original 4/8VM, experimenters undergo
a thorough training procedure to properly administer
standardized interviews to ascertain a participant’s naviga-
tional strategy without introducing bias (see Sodums &
Bohbot, 2020, for a full description), which was not possi-
ble within SHQ. Specifically, using the 4/8VM people have
previously been categorized into distinct groups with bio-
logical differences based on navigation strategy. People
categorized as response learners included those who used
no landmarks (i.e., response-start position), those who
used a single landmark in conjunction with a full sequence
counting strategy (response-landmark), and those who
include two or more landmarks but also in conjunction
with a full sequence counting strategy from a single start-
ing point (response-landmarks). On the other hand, if
multiple landmarks were used, and a full sequence was
not used, then people could be categorized as a spatial
strategy user (Blanchette et al., 2020; Aumont, Arguin,
et al., 2019; Aumont, Blanchette, et al., 2019; Blanchette,
Amirova, Bohbot, & West, 2019; West et al., 2015, 2018;
Drisdelle et al., 2017; Bohbot, Konishi, Sodums, Dahmani,
& Bherer, 2015; Bohbot et al., 2007, 2013; Andersen et al.,
2012; Bohbot, Gupta, Banner, & Dahmani, 2011; Iaria
et al., 2003). We consider response and spatial learners
as distinct groups because of the fact that there are multi-
ple biological differences between them including basal
cortisol levels (Blanchette et al., 2020; Hussain, Hanafi,
Konishi, Brake, & Bohbot, 2016; Bohbot et al., 2011), vol-
ume of the hippocampus and caudate nucleus (West et al.,
2018; Bohbot et al., 2007; Iaria et al., 2003), and genetics
(Konishi et al., 2016; Banner, Bhat, Etchamendy, Joober, &
Bohbot, 2011). Furthermore, our experiments in the lab-
oratory are able to detect if a participant shifts their strat-
egy during the radial arm maze task; however, this process
required a detailed interview by a trained experiment and
is therefore not possible in the SHQ version of the task.
Another interesting avenue for future research would be
to include a non-self-report measure of navigational strat-
egy that could be based on, for example, dwell time that
includes landmarks in the direct field of view. This could
then be used to further evaluate the accuracy of the self-
report measures used in the current study.
Despite these observed biological differences when
comparing spatial and response learners globally as two
groups, differences in wayfinding performance between
the subcategories of response learners have also been
observed. People categorized as response-landmarks
(one or more landmarks but also in conjunction with a full
sequence counting strategy) perform better than people
who used a counting strategy from a single starting posi-
tion on a wayfinding task (Etchamendy & Bohbot, 2007).
Because of this previously identified behavioral distinction
between people categorized as using a response land-
marks and response start position strategy and our
West et al.
463
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
c
n
_
a
_
0
1
9
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
inability to categorized a more specific response learning
strategy, we combined people who reported using “Used
multiple landmarks” and “Counted from a landmark,” and
recognize that we were not able to distinguish more spe-
cific navigation strategies that we are able to ascertain using
the 4/8VM. We, however, do observe that landmark use of
some kind is associated with better memory performance
and this does significantly decline with age, which is consis-
tent with previous smaller studies. In fitting with the current
data, spatial use in the laboratory is associated with
increased landmark use, response-landmark strategies are
associated with a moderate amount of landmark use, and
people who use a counting-only strategy display the
lowest landmark use (Andersen et al., 2012).
Altogether, the results of the current study provided a
unique opportunity to validate with a very large data set
previous studies that have demonstrated reduced
landmark-dependent navigational strategy use in ageing.
Furthermore, although overall memory performance was
lower for those using non-landmark-dependent strate-
gies, memory performance depended on the memory
tested and showed a complex interaction among people
in older age ranges. Although we were not in a position to
ascertain more specific navigation strategies as we have
done previously in the laboratory, our current findings
extend previous studies that have shown a decline in
landmark-dependent spatial strategy use in ageing that
is associated with decreased hippocampal gray matter
and cognitive performance. Finally, this study contributes
to the mounting evidence in favor of mobile and out-of-lab
testing, offering larger and more diverse samples, easier
access to patients, and the continuity of research despite
mobility limitations (as exemplified by COVID-19).
Reprint requests should be sent to Greg L. West, University of
Montreal, Department of Psychology, Pavillon Marie-Victorin 90,
avenue Vincent d’Indy, Montreal QC H2V 2S9, or via e-mail:
gregory.west@umontreal.ca.
Data Availability Statement
Please contact the PIs for access to data and scripts.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender
identification of first author/ last author) publishing
in the Journal of Cognitive Neuroscience ( JoCN) during
this period were M(an)/M = .407, W(oman)/M = .32,
M/ W = .115, and W/ W = .159, the comparable propor-
tions for the articles that these authorship teams cited
were M/M = .549, W/M = .257, M/ W = .109, and W/ W =
.085 (Postle and Fulvio, JoCN, 34:1, pp. 1–3). Consequently,
JoCN encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender cita-
tion balance.
REFERENCES
Allison, S. L., Fagan, A. M., Morris, J. C., & Head, D. (2016).
Spatial navigation in preclinical Alzheimer’s disease. Journal
of Alzheimer’s Disease, 52, 77–90. https://doi.org/10.3233
/JAD-150855, PubMed: 26967209
Andersen, N. E., Dahmani, L., Konishi, K., & Bohbot, V. D.
(2012). Eye tracking, strategies, and sex differences in virtual
navigation. Neurobiology of Learning and Memory, 97,
81–89. https://doi.org/10.1016/j.nlm.2011.09.007, PubMed:
22001012
Apostolova, L. G., Dutton, R. A., Dinov, I. D., Hayashi, K. M.,
Toga, A. W., Cummings, J. L., et al. (2006). Conversion of mild
cognitive impairment to Alzheimer disease predicted by
hippocampal atrophy maps. Archives of Neurology, 63,
693–699. https://doi.org/10.1001/archneur.63.5.693, PubMed:
16682538
Aumont, E., Arguin, M., Bohbot, V., & West, G. L. (2019).
Increased flanker task and forward digit span performance in
caudate-nucleus-dependent response strategies. Brain and
Cognition, 135, 103576. https://doi.org/10.1016/j.bandc.2019
.05.014, PubMed: 31203022
Aumont, E., Blanchette, C. A., Bohbot, V. D., & West, G. L.
(2019). Caudate nucleus-dependent navigation strategies are
associated with increased risk-taking and set-shifting
behavior. Learning & Memory, 26, 101–108. https://doi.org
/10.1101/lm.048306.118, PubMed: 30898972
Banner, H., Bhat, V., Etchamendy, N., Joober, R., & Bohbot,
V. D. (2011). The brain-derived neurotrophic factor Val66Met
polymorphism is associated with reduced functional
magnetic resonance imaging activity in the hippocampus and
increased use of caudate nucleus-dependent strategies in a
human virtual navigation task. European Journal of
Neuroscience, 33, 968–977. https://doi.org/10.1111/j.1460
-9568.2010.07550.x, PubMed: 21255124
Barnes, C. A., Nadel, L., & Honig, W. K. (1980). Spatial memory
deficit in senescent rats. Canadian Journal of Psychology,
34, 29–39. https://doi.org/10.1037/h0081022, PubMed:
7388694
Bates, S. L., & Wolbers, T. (2014). How cognitive aging affects
multisensory integration of navigational cues. Neurobiology
of Aging, 35, 2761–2769. https://doi.org/10.1016/j
.neurobiolaging.2014.04.003, PubMed: 24952995
Blanchette, C. A., Amirova, J., Bohbot, V. D., & West, G. L.
(2019). Autistic traits in neurotypical individuals are
associated with increased landmark use during navigation.
PsyCh Journal, 8, 137–146. https://doi.org/10.1002/pchj.230,
PubMed: 30294869
Blanchette, C. A., Kurdi, V., Fouquet, C., Schachar, R., Boivin,
M., Hastings, P., et al. (2020). Opposing effects of cortisol on
learning and memory in children using spatial versus
response-dependent navigation strategies. Neurobiology of
Learning and Memory, 169, 107172. https://doi.org/10.1016/j
.nlm.2020.107172, PubMed: 31978550
Bohbot, V. D., Del Balso, D., Conrad, K., Konishi, K., & Leyton,
M. (2013). Caudate nucleus-dependent navigational
strategies are associated with increased use of addictive
drugs. Hippocampus, 23, 973–984. https://doi.org/10.1002
/hipo.22187, PubMed: 23939925
Bohbot, V. D., Gupta, M., Banner, H., & Dahmani, L. (2011).
Caudate nucleus-dependent response strategies in a virtual
464
Journal of Cognitive Neuroscience
Volume 35, Number 3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
c
n
_
a
_
0
1
9
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
navigation task are associated with lower basal cortisol and
impaired episodic memory. Neurobiology of Learning and
Memory, 96, 173–180. https://doi.org/10.1016/j.nlm.2011.04
.007, PubMed: 21539927
Bohbot, V. D., Iaria, G., & Petrides, M. (2004). Hippocampal
function and spatial memory: Evidence from functional
neuroimaging in healthy participants and performance of
patients with medial temporal lobe resections.
Neuropsychology, 18, 418–425. https://doi.org/10.1037/0894
-4105.18.3.418, PubMed: 15291720
Bohbot, V. D., Konishi, K., Sodums, D., Dahmani, L., & Bherer,
L. (2015). Hippocampus and cortical plasticity following a
virtual spatial memory intervention program promote
spontaneous hippocampus-dependent navigation strategies
in healthy older adults. Paper presented at the annual
meeting of the Society for Neuroscience. Chicago, IL.
Bohbot, V. D., Lerch, J., Thorndycraft, B., Iaria, G., & Zijdenbos,
A. P. (2007). Gray matter differences correlate with
spontaneous strategies in a human virtual navigation task.
Journal of Neuroscience, 27, 10078–10083. https://doi.org/10
.1523/JNEUROSCI.1763-07.2007, PubMed: 17881514
Bohbot, V. D., McKenzie, S., Konishi, K., Fouquet, C., Kurdi, V.,
Schachar, R., et al. (2012). Virtual navigation strategies from
childhood to senescence: Evidence for changes across the
life span. Frontiers in Aging Neuroscience, 4, 28. https://doi
.org/10.3389/fnagi.2012.00028, PubMed: 23162463
Chang, Q., & Gold, P. E. (2003). Switching memory systems
during learning: Changes in patterns of brain acetylcholine
release in the hippocampus and striatum in rats. Journal of
Neuroscience, 23, 3001–3005. https://doi.org/10.1523
/JNEUROSCI.23-07-03001.2003, PubMed: 12684487
Cialone, C., Tenbrink, T., & Spiers, H. J. (2018). Sculptors,
architects, and painters conceive of depicted spaces
differently. Cognitive Science, 42, 524–553. https://doi.org/10
.1111/cogs.12510, PubMed: 28656679
Coughlan, G., Coutrot, A., Khondoker, M., Minihane, A. M.,
Spiers, H., & Hornberger, M. (2019). Toward personalized
cognitive diagnostics of at-genetic-risk Alzheimer’s disease.
Proceedings of the National Academy of Sciences, U.S.A.,
116, 9285–9292. https://doi.org/10.1073/pnas.1901600116,
PubMed: 31015296
Coughlan, G., Flanagan, E., Jeffs, S., Bertoux, M., Spiers, H.,
Mioshi, E., et al. (2018). Diagnostic relevance of spatial
orientation for vascular dementia: A case study. Dementia &
Neuropsychologia, 12, 85–91. https://doi.org/10.1590/1980
-57642018dn12-010013, PubMed: 29682239
Coughlan, G., Puthusseryppady, V., Lowry, E., Gillings, R.,
Spiers, H., Minihane, A. M., et al. (2020). Test-retest reliability
of spatial navigation in adults at-risk of Alzheimer’s disease.
PLoS One, 15, e0239077. https://doi.org/10.1371/journal.pone
.0239077, PubMed: 32960930
Coutrot, A., Manley, E., Goodroe, S., Gahnstrom, C., Filomena, G.,
Yesiltepe, D., et al. (2022). Entropy of city street networks
linked to future spatial navigation ability. Nature, 604, 104–110.
https://doi.org/10.1038/s41586-022-04486-7, PubMed: 35355009
Coutrot, A., Schmidt, S., Coutrot, L., Pittman, J., Hong, L.,
Wiener, J. M., et al. (2019). Virtual navigation tested on a
mobile app is predictive of real-world wayfinding navigation
performance. PLoS One, 14, e0213272. https://doi.org/10
.1371/journal.pone.0213272, PubMed: 30883560
Coutrot, A., Silva, R., Manley, E., de Cothi, W., Sami, S., Bohbot,
V. D., et al. (2018). Global determinants of navigation ability.
Current Biology, 28, 2861–2866. https://doi.org/10.1016/j.cub
.2018.06.009, PubMed: 30100340
Dahmani, L., & Bohbot, V. D. (2020). Habitual use of GPS
negatively impacts spatial memory during self-guided
navigation. Scientific Reports, 10, 6310. https://doi.org/10
.1038/s41598-020-62877-0, PubMed: 32286340
Donix, M., Burggren, A. C., Suthana, N. A., Siddarth, P.,
Ekstrom, A. D., Krupa, A. K., et al. (2010). Longitudinal
changes in medial temporal cortical thickness in normal
subjects with the APOE-4 polymorphism. Neuroimage, 53,
37–43. https://doi.org/10.1016/j.neuroimage.2010.06.009,
PubMed: 20541611
Driscoll, I., Hamilton, D. A., Petropoulos, H., Yeo, R. A., Brooks,
W. M., Baumgartner, R. N., et al. (2003). The aging
hippocampus: Cognitive, biochemical and structural findings.
Cerebral Cortex, 13, 1344–1351. https://doi.org/10.1093
/cercor/bhg081, PubMed: 14615299
Drisdelle, B. L., Konishi, K., Diarra, M., Bohbot, V. D., Jolicoeur,
P., & West, G. L. (2017). Electrophysiological evidence
for enhanced attentional deployment in spatial learners.
Experimental Brain Research, 235, 1387–1395. https://doi
.org/10.1007/s00221-017-4884-9, PubMed: 28229169
Ekstrom, A. D., & Ranganath, C. (2018). Space, time, and
episodic memory: The hippocampus is all over the cognitive
map. Hippocampus, 28, 680–687. https://doi.org/10.1002
/hipo.22750, PubMed: 28609014
Etchamendy, N., & Bohbot, V. D. (2007). Spontaneous
navigational strategies and performance in the virtual town.
Hippocampus, 17, 595–599. https://doi.org/10.1002/hipo
.20303, PubMed: 17546682
Etchamendy, N., Konishi, K., Pike, G. B., Marighetto, A., &
Bohbot, V. D. (2012). Evidence for a virtual human analog of
a rodent relational memory task: A study of aging and fMRI in
young adults. Hippocampus, 22, 869–880. https://doi.org/10
.1002/hipo.20948, PubMed: 21656872
Gardner, R. S., Gold, P. E., & Korol, D. L. (2020). Inactivation of
the striatum in aged rats rescues their ability to learn a
hippocampus-sensitive spatial navigation task. Neurobiology
of Learning and Memory, 172, 107231. https://doi.org/10
.1016/j.nlm.2020.107231, PubMed: 32305514
Guderian, S., Dzieciol, A. M., Gadian, D. G., Jentschke, S.,
Doeller, C. F., Burgess, N., et al. (2015). Hippocampal volume
reduction in humans predicts impaired allocentric spatial
memory in virtual-reality navigation. Journal of
Neuroscience, 35, 14123–14131. https://doi.org/10.1523
/JNEUROSCI.0801-15.2015, PubMed: 26490854
Harris, M. A., & Wolbers, T. (2014). How age-related strategy
switching deficits affect wayfinding in complex environments.
Neurobiology of Aging, 35, 1095–1102. https://doi.org/10
.1016/j.neurobiolaging.2013.10.086, PubMed: 24239438
Head, D., & Isom, M. (2010). Age effects on wayfinding and route
learning skills. Behavioural Brain Research, 209, 49–58.
https://doi.org/10.1016/j.bbr.2010.01.012, PubMed: 20085784
Hentschke, H., & Stuttgen, M. C. (2011). Computation of
measures of effect size for neuroscience data sets. European
Journal of Neuroscience, 34, 1887–1894. https://doi.org/10
.1111/j.1460-9568.2011.07902.x, PubMed: 22082031
Hussain, D., Hanafi, S., Konishi, K., Brake, W. G., & Bohbot,
V. D. (2016). Modulation of spatial and response strategies by
phase of the menstrual cycle in women tested in a virtual
navigation task. Psychoneuroendocrinology, 70, 108–117.
https://doi.org/10.1016/j.psyneuen.2016.05.008, PubMed:
27213559
Iaria, G., Petrides, M., Dagher, A., Pike, B., & Bohbot, V. D.
(2003). Cognitive strategies dependent on the hippocampus
and caudate nucleus in human navigation: Variability and
change with practice. Journal of Neuroscience, 23,
5945–5952. https://doi.org/10.1523/JNEUROSCI.23-13-05945
.2003, PubMed: 12843299
Jagust, W., Gitcho, A., Sun, F., Kuczynski, B., Mungas, D., &
Haan, M. (2006). Brain imaging evidence of preclinical
Alzheimer’s disease in normal aging. Annals of Neurology,
59, 673–681. https://doi.org/10.1002/ana.20799, PubMed:
16470518
West et al.
465
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
c
n
_
a
_
0
1
9
5
6
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Jessen, F., Wiese, B., Bachmann, C., Eifflaender-Gorfer, S.,
Haller, F., Kolsch, H., et al. (2010). Prediction of dementia by
subjective memory impairment: Effects of severity and
temporal association with cognitive impairment. Archives of
General Psychiatry, 67, 414–422. https://doi.org/10.1001
/archgenpsychiatry.2010.30, PubMed: 20368517
Klein, D. A., Steinberg, M., Galik, E., Steele, C., Sheppard,
J. M., Warren, A., et al. (1999). Wandering behaviour in
community-residing persons with dementia. International
Journal of Geriatric Psychiatry, 14, 272–279. https://doi.org
/10.1002/(sici)1099-1166(199904)14:4<272::aid-gps896>3.0
.co;2-P, PubMed: 10340188
Konishi, K., Bhat, V., Banner, H., Poirier, J., Joober, R., &
Bohbot, V. D. (2016). APOE2 is associated with spatial
navigational strategies and increased gray matter in the
hippocampus. Frontiers in Human Neuroscience, 10, 349.
https://doi.org/10.3389/fnhum.2016.00349, PubMed: 27468260
Konishi, K., & Bohbot, V. D. (2013). Spatial navigational
strategies correlate with gray matter in the hippocampus of
healthy older adults tested in a virtual maze. Frontiers in
Aging Neuroscience, 5, 1. https://doi.org/10.3389/fnagi.2013
.00001, PubMed: 23430962
Konishi, K., Etchamendy, N., Roy, S., Marighetto, A., Rajah, N., &
Bohbot, V. D. (2013). Decreased functional magnetic
resonance imaging activity in the hippocampus in favor of the
caudate nucleus in older adults tested in a virtual navigation
task. Hippocampus, 23, 1005–1014. https://doi.org/10.1002
/hipo.22181, PubMed: 23929534
Konishi, K., McKenzie, S., Etchamendy, N., Roy, S., & Bohbot,
V. D. (2017). Hippocampus-dependent spatial learning is
associated with higher global cognition among healthy older
adults. Neuropsychologia, 106, 310–321. https://doi.org/10
.1016/j.neuropsychologia.2017.09.025, PubMed: 28963056
Laczo, M., Martinkovic, L., Lerch, O., Wiener, J. M., Kalinova, J.,
Matuskova, V., et al. (2022). Different profiles of spatial
navigation deficits in Alzheimer’s disease biomarker-positive
versus biomarker-negative older adults with amnestic mild
cognitive impairment. Frontiers in Aging Neuroscience, 14,
886778. https://doi.org/10.3389/fnagi.2022.886778, PubMed:
35721017
Lerch, J. P., Yiu, UN. P., Martinez-Canabal, A., Pekar, T., Bohbot,
V. D., Frankland, P. W., et al. (2011). Maze training in mice
induces MRI-detectable brain shape changes specific to the
type of learning. Neuroimage, 54, 2086–2095. https://doi.org
/10.1016/j.neuroimage.2010.09.086, PubMed: 20932918
Lester, UN. W., Moffat, S. D., Wiener, J. M., Barnes, C. A., &
Wolbers, T. (2017). The aging navigational system. Neuron,
95, 1019–1035. https://doi.org/10.1016/j.neuron.2017.06.037,
PubMed: 28858613
Lupien, S. J., de Leon, M., de Santi, S., Convit, A., Tarshish, C.,
Nair, N. P., et al. (1998). Cortisol levels during human aging
predict hippocampal atrophy and memory deficits. Nature
Neuroscience, 1, 69–73. https://doi.org/10.1038/271,
PubMed: 10195112
Mapstone, M., Steffenella, T. M., & Duffy, C. J. (2003). UN
visuospatial variant of mild cognitive impairment: Getting lost
between aging and AD. Neurology, 60, 802–808. https://doi
.org/10.1212/01.wnl.0000049471.76799.Di, PubMed:
12629237
McDonald, R. J., & White, N. M. (1993). A triple dissociation
of memory systems: Hippocampus, amygdala, and dorsal
striatum. Behavioral Neuroscience, 107, 3–22. https://doi.org
/10.1037/0735-7044.107.1.3, PubMed: 8447956
Moffat, S. D., Elkins, W., & Resnick, S. M. (2006). Age
differences in the neural systems supporting human
allocentric spatial navigation. Neurobiology of Aging, 27,
965–972. https://doi.org/10.1016/j.neurobiolaging.2005.05
.011, PubMed: 15982787
Nemmi, F., Boccia, M., & Guariglia, C. (2017). Does aging affect
the formation of new topographical memories? Evidence
from an extensive spatial training. Neuropsychology,
Development, and Cognition. Section B, Aging,
Neuropsychology and Cognition, 24, 29–44. https://doi.org
/10.1080/13825585.2016.1167162, PubMed: 27045346
Nicolle, M. M., Prescott, S., & Bizon, J. l. (2003). Emergence of a
cue strategy preference on the water maze task in aged
C57B6 x SJL F1 hybrid mice. Apprendimento & Memory, 10,
520–524. https://doi.org/10.1101/lm.64803, PubMed:
14657263
O’Dwyer, L., Lamberton, F., Matura, S., Tanner, C., Scheibe, M.,
Mugnaio, J., et al. (2012). Reduced hippocampal volume in
healthy young ApoE4 carriers: An MRI study. PLoS One, 7,
e48895. https://doi.org/10.1371/journal.pone.0048895,
PubMed: 23152815
O’Keefe, J., & Nadel, l. (1978). The hippocampus as a
cognitive map. Clarendon Press.
Olton, D. S., & Papas, B. C. (1979). Spatial memory and
hippocampal function. Neuropsychologia, 17, 669–682.
https://doi.org/10.1073/pnas.0611233104, PubMed: 17296931
O’Shea, A., Cohen, R. A., Porges, E. C., Nissim, N. R., & Woods,
UN. J. (2016). Cognitive aging and the hippocampus in older
adults. Frontiers in Aging Neuroscience, 8, 298. https://doi
.org/10.3389/fnagi.2016.00298, PubMed: 28008314
Packard, M. G., Hirsh, R., & White, N. M. (1989). Differential
effects of fornix and caudate nucleus lesions on two radial
maze tasks: Evidence for multiple memory systems. Journal
of Neuroscience, 9, 1465–1472. https://doi.org/10.1523
/JNEUROSCI.09-05-01465.1989, PubMed: 2723738
Packard, M. G., & McGaugh, J. l. (1992). Double dissociation of
fornix and caudate nucleus lesions on acquisition of two
water maze tasks: Further evidence for multiple memory
systems. Behavioral Neuroscience, 106, 439–446. https://doi
.org/10.1037/0735-7044.106.3.439, PubMed: 1616610
Packard, M. G., & McGaugh, J. l. (1996). Inactivation of
hippocampus or caudate nucleus with lidocaine differentially
affects expression of place and response learning.
Neurobiology of Learning and Memory, 65, 65–72. https://
doi.org/10.1006/nlme.1996.0007, PubMed: 8673408
Passini, R., Rainville, C., Marchand, N., & Joanette, Y. (1995).
Wayfinding in dementia of the Alzheimer type: Planning
abilities. Journal of Clinical and Experimental
Neuropsychology, 17, 820–832. https://doi.org/10.1080
/01688639508402431, PubMed: 8847388
Persson, K. (2018). Finding of increased caudate nucleus in
patients with Alzheimer’s disease. Acta Neurologica
Scandinavica, 137, 224–232. https://doi.org/10.1111/ane
.12800, PubMed: 28741672
Pievani, M., Galluzzi, S., Thompson, P. M., Rasser, P. E., Bonetti,
M., & Frisoni, G. B. (2011). APOE4 is associated with greater
atrophy of the hippocampal formation in Alzheimer’s disease.
Neuroimage, 55, 909–919. https://doi.org/10.1016/j
.neuroimage.2010.12.081, PubMed: 21224004
Raz, N., Rodrigue, K. M., Head, D., Kennedy, K. M., & Acker, J. D.
(2004). Differential aging of the medial temporal lobe: A study
of a five-year change. Neurology, 62, 433–438. https://doi.org
/10.1212/01.wnl.0000106466.09835.46, PubMed: 14872026
Ritter, A., Hawley, N., Banks, S. J., & Mugnaio, J. B. (2017). IL
association between Montreal Cognitive Assessment memory
scores and hippocampal volume in a neurodegenerative
disease sample. Journal of Alzheimer’s Disease, 58, 695–699.
https://doi.org/10.3233/JAD-161241, PubMed: 28453481
Schinazi, V. R., Nardi, D., Newcombe, N. S., Shipley, T. F., &
Epstein, R. UN. (2013). Hippocampal size predicts rapid
learning of a cognitive map in humans. Hippocampus, 23,
515–528. https://doi.org/10.1002/hipo.22111, PubMed:
23505031
466
Journal of Cognitive Neuroscience
Volume 35, Numero 3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
C
N
_
UN
_
0
1
9
5
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Small, S. A., Tsai, W. Y., DeLaPaz, R., Mayeux, R., & Stern, Y.
(2002). Imaging hippocampal function across the human
life span: Is memory decline normal or not? Annals of
Neurology, 51, 290–295. https://doi.org/10.1002/ana.10105,
PubMed: 11891823
Sodums, D. J., & Bohbot, V. D. (2020). Negative correlation
between grey matter in the hippocampus and caudate
nucleus in healthy aging. Hippocampus, 30, 892–908. https://
doi.org/10.1002/hipo.23210, PubMed: 32384195
Spiers, H. J., Burgess, N., Hartley, T., Vargha-Khadem, F., &
O’Keefe, J. (2001). Bilateral hippocampal pathology impairs
topographical and episodic memory but not visual pattern
matching. Hippocampus, 11, 715–725. https://doi.org/10
.1002/hipo.1087, PubMed: 11811666
Spiers, H. J., Coutrot, A., & Hornberger, M. (2021). Explaining
world-wide variation in navigation ability from millions of
people: Citizen science Project Sea Hero Quest. Topics in
Cognitive Science, 15, 120–138. https://doi.org/10.1111/tops
.12590, PubMed: 34878689
Stangl, M., Kanitscheider, I., Riemer, M., Fiete, I., & Wolbers, T.
(2020). Sources of path integration error in young and aging
humans. Nature Communications, 11, 2626. https://doi.org
/10.1038/s41467-020-15805-9, PubMed: 32457293
Thambisetty, M., Wan, J., Carass, A., An, Y., Prince, J. L., &
Resnick, S. M. (2010). Longitudinal changes in cortical
thickness associated with normal aging. Neuroimage, 52,
1215–1223. https://doi.org/10.1016/j.neuroimage.2010.04
.258, PubMed: 20441796
West, G. L., Drisdelle, B. L., Konishi, K., Jackson, J., Jolicoeur, P., &
Bohbot, V. D. (2015). Habitual action video game playing is
associated with caudate nucleus-dependent navigational
strategies. Proceedings of the Biological Sciences, 282, 20142952.
https://doi.org/10.1098/rspb.2014.2952, PubMed: 25994669
West, G. L., Konishi, K., Diarra, M., Benady-Chorney, J.,
Drisdelle, B. L., Dahmani, L., et al. (2018). Impact of video
games on plasticity of the hippocampus. Molecular
Psychiatry, 23, 1566–1574. https://doi.org/10.1038/mp.2017
.155, PubMed: 28785110
Zhong, J. Y., & Moffat, S. D. (2016). Age-related differences in
associative learning of landmarks and heading directions in a
virtual navigation task. Frontiers in Aging Neuroscience, 8, 122.
https://doi.org/10.3389/fnagi.2016.00122, PubMed: 27303290
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
3
4
5
2
2
0
6
9
3
9
1
/
j
o
C
N
_
UN
_
0
1
9
5
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
West et al.
467