Jossinger, S., Yablonski, M., Amir, O, & Ben-Shachar, M. (2023). The role of the cerebellum in language comprehension and
production. Neurobiology of Language, Advance publication. https://doi.org/10.1162/nol_a_00098.
The contributions of the cerebellar peduncles and the frontal
aslant tract in mediating speech fluency
Jossinger S.1*, Yablonski M.1§, Amir O.2, Ben-Shachar M. 1,3*
1 The Gonda Multidisciplinary Brain Research Center, Bar-Ilan University, Ramat-Gan, Israel
2 Department of Communication Disorders, Sackler Faculty of Medicine, Tel-Aviv University,
Tel-Aviv, Israel
3 Department of English Literature and Linguistics, Bar-Ilan University, Ramat-Gan, Israel
* Corresponding author at The Gonda Multidisciplinary Brain Research Center, Bar Ilan
Università, Ramat Gan 5290002, Israel. E-mail addresses: jossins@biu.ac.il ;
michalb@mail.biu.ac.il
§ Current address at Stanford University School of Medicine and Graduate School of Education,
Stanford, CA, USA.
Abbreviated title: Cerebellar contributions to speech fluency
Ringraziamenti: This study was conducted as part of Sivan Jossinger’s doctoral dissertation,
carried out under the supervision of Prof. Michal Ben-Shachar at the Gonda Multidisciplinary
Brain Research Center, Bar-Ilan University. The analysis of speech rate data was carried out by
Gaya Noam and Michal Braun, under the supervision of Prof. Ofer Amir. We thank Yaniv Assaf,
Daniel Barazany and the team at the Strauss Center for Computational Neuroimaging for their
assistance in protocol setup and MRI setting.
Conflict of interest: Authors report no conflict of interest
Funding sources: This study is supported by the Israel Science Foundation (ISF Grant
#1083/17).
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
.
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Manuscript
Click here to access/download;Manuscript;Jossinger-et-
al_2022_NoL-SI_manuscript_R2.pdf
The contributions of the cerebellar peduncles and the frontal
aslant tract in mediating speech fluency
ABSTRACT
Fluent speech production is a complex task that spans multiple processes, from conceptual
framing and lexical access, through phonological encoding, to articulatory control. For the most
part, imaging studies portraying the neural correlates of speech fluency tend to examine clinical
populations sustaining speech impairments, and focus on either lexical access or articulatory
controllo, but not both. Here, we evaluated the contribution of the cerebellar peduncles to speech
fluency by measuring the different components of the process, in a sample of forty-five
neurotypical adults. Participants underwent an unstructured interview to assess their natural
speaking rate and articulation rate, and completed timed semantic and phonemic fluency tasks to
assess their verbal fluency. Diffusion MRI with probabilistic tractography was used to segment
the bilateral cerebellar peduncles (CPs) and frontal aslant tract (FAT), previously associated with
speech production in clinical populations. Our results demonstrate distinct patterns of white
matter associations with different fluency components. Specifically, verbal fluency is associated
with the right superior CP, whereas speaking rate is associated with the right middle CP and
bilateral FAT. No association is found with articulation rate in these pathways, in contrast to
previous findings in persons who stutter. Our findings support the contribution of the cerebellum
to aspects of speech production that go beyond articulatory control, such as lexical access,
pragmatic or syntactic generation. Further, we demonstrate that distinct cerebellar pathways
dissociate different components of speech fluency in neurotypical speakers.
1
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
/
.
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
INTRODUCTION
Humans produce about 16,000 words every day, at an astounding speed of more than 150
words per minute (Amir, 2016; Mehl et al., 2007; Rodero, 2012). Such fast and fluent speech
production depends upon complex interactions between motor, sensory and cognitive systems
underpinning different aspects of speech production, such as articulatory control, phonological
encoding, conceptual framing and lexical access (Hickok, 2012). Despite these interactions, most
studies investigating speech fluency focus on either articulatory control or lexical access, but not
both. Here, we combine the two perspectives to allow a comprehensive understanding of the
neural pathways associated with fluent speech production in neurotypical adults.
Lexical access, including lexical search, selection, and retrieval, is typically measured using
verbal fluency tasks. In these tasks, participants are asked to produce as many words as possible
within 60 seconds under a specific criterion: words beginning with a certain letter (cioè., phonemic
fluency) or a semantic category (cioè., semantic fluency). Verbal fluency is often assessed in
clinical populations such as persons with aphasia (Bose et al., 2022), dementia (Libon et al.,
2009), multiple sclerosis (Blecher et al., 2019), Parkinson’s disease (Henry & Crawford, 2004),
and more. Importantly, performance in the verbal fluency task is influenced by adequate word
selection and avoidance of repetition. Therefore, these tasks are used to assess not only lexical
knowledge, but also executive functions such as working memory and inhibition (Amunts et al.,
2020; Shao et al., 2014). Infatti, verbal fluency is a central task in neuropsychological batteries
that assess executive functions (Kramer et al., 2014; Shao et al., 2014).
Fluent speech production is also quantified using measures of speech rate. In contrasto con il
timed and highly constrained nature of verbal fluency tasks, speech rate measures, ad esempio
Speaking rate and Articulation rate, are based on a relatively naturalistic setting of spontaneous
speech samples. A grandi linee, speech rate is calculated as the number of spoken units (syllables or
parole) produced within a time unit (second or minute). Speaking rate is measured across
continuous segments of speech which may include pauses, repetitions, and revisions. Così, it is
considered a global measure of verbal output and language proficiency (Costello & Ingham,
1984; Howell et al., 1999). Articulation rate, on the other hand, is based only on fluent
utterances after excluding any kind of disfluency, and thus considered a measure of articulatory
motor control (Walker et al., 1992). Generally, speech rate (as quantified by both measures)
2
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
.
/
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
affects the speakers’ intelligibility, fluency, and communication efficiency (Amir, 2016; Sturm &
Seery, 2007). Measures of speech rate are typically used in the clinical assessment of various
speech disorders, such as apraxia of speech (Kent & Rosenbek, 1983), dysarthria (Kent et al.,
1987), and persistent developmental stuttering (Andrade et al., 2003).
Although speech fluency involves both lexical access and articulatory control, studies in the
field tend to focus either on linguistic aspects of speech or on motor aspects of speech, but not
both. In an attempt to bridge this gap, Hickok has recently suggested the Hierarchical State
Feedback Control (HSFC) model of speech production (Hickok, 2012). This model synthesizes
between psycholinguistic and motor control approaches and suggests an integrated brain
circuitry of speech production. The current understanding of the brain circuits that control fluent
speech production, Tuttavia, stems largely from studies in patients. Hence, in the current study
we wish to evaluate the different interpretations of speech fluency and evaluate the neural
substrates that stand at the base of fluent speech production in neurotypical adults.
According to computational models of speech production, the cerebellum is a key node in
different aspects of fluent speech production (Hickok, 2012; Tourville & Guenther, 2011).
Infatti, patients with cerebellar lesions exhibit impaired verbal fluency abilities and significantly
slower speaking rate and articulation rate compared to controls (Ackermann et al., 1992;
Peterburs et al., 2010). Neuroimaging data point to a significant cerebellar activation during both
verbal fluency tasks (Halari et al., 2006; Schlösser et al., 1998) and tasks that involve change in
articulation rate (Riecker et al., 2005, 2006). Naturally, the involvement of the cerebellum in
verbal fluency tasks could stem from the fact that fluency tasks use the articulatory system.
Therefore, to make a functional segregation, the contribution of the cerebellum to each of these
measures should be evaluated in the same participants.
Apart from the cerebellum, neural control of speech recruits a distributed cortical network
(Hickok, 2012; Tourville & Guenther, 2011). The cortex and the cerebellum are structurally
connected via long range white matter pathways which enable efficient communication of
signals across considerable distance. As the exclusive bridge between the cerebellum and extra-
cerebellar regions, the structural properties and organization of the cerebellar peduncles are
important for understanding the neural basis of fluent speech production.
Cerebellar input and output information is carried by three major white matter pathways
known as the cerebellar peduncles (CPs): the inferior cerebellar peduncle (ICP), the middle
3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
.
/
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
cerebellar peduncle (MCP), and the superior cerebellar peduncle (SCP). The ICP is a cerebellar
input pathway, feeding signals from the inferior olive and spinal cord into the cerebellar cortex
(Perrini et al., 2013). The ICP was shown to be implicated in articulation rate among people with
developmental stuttering. Developmental stuttering was previously associated with abnormalities
in the microstructural properties of the bilateral ICP (Connally et al., 2014; but see Jossinger et
al., 2021, 2022 for contradicting results). A recent study in young children who stutter showed
that microstructural differences in the ICP emerge early in development (Johnson et al., 2022). In
adults with developmental stuttering, but not in neurotypical speakers, the microstructural
properties of the ICP was shown to be correlated with articulation rate (Jossinger et al., 2021;
Kronfeld-Duenias et al., 2016).
The SCP is a major output pathway transmitting signals from the cerebellum into the
contralateral cerebral cortex via the thalamus. The MCP is a major input pathway feeding signals
from the cerebral cortex into the contralateral cerebellar cortex, decussating at the level of the
pontine nucleus. Together, the SCP and MCP form the cerebro-cerebellar loop which allows
transferring information from the cerebellum to the cerebral cortex and vice versa.
Computational models of speech production hypothesize that the cerebro-cerebellar projections
contribute to the feedforward control of speech by mapping between the desired speech sound
and its appropriate articulatory gesture (Tourville & Guenther, 2011). Recent data show that the
cerebro-cerebellar connectivity is also associated with speech-related cognitive demands, ad esempio
verbal working memory (Sobczak-Edmans et al., 2019) and story comprehension (Castellazzi et
al., 2018). Functional MRI (fMRI) studies have shown that verbal fluency tasks cause a
significant activation in the cerebellum, together with prefrontal and temporo-parietal areas
(Gurd et al., 2002; Hubrich-Ungureanu et al., 2002; Schlösser et al., 1998). On this basis,
cerebellar activity during verbal communication may reflect not only motor aspects of speech
production, but also cognitive demands of word generation.
The potential involvement of the cerebro-cerebellar loop in the fluent production of speech is
further supported by the anatomical connections found between the cerebellum and the prefrontal
cortex (Kelly & Strick, 2003; Middleton & Strick, 1994; Palesi et al., 2017). Within the
prefrontal cortex, feedforward control of speech involves the activation of the inferior frontal
gyrus (IFG) and the supplementary motor area (SMA) (Hickok, 2012; Tourville & Guenther,
2011). The IFG and SMA were recently shown to be connected via the frontal aslant tract (FAT)
4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
/
.
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
(Catani et al., 2012). To date, the involvement of the FAT in speech fluency was mainly studied
in clinical populations. Per esempio, Kronfeld-Duenias et al. (2016) showed that adults who
stutter demonstrate abnormal microstructure within the FAT compared to fluent speakers. Questo
structural difference was also accompanied by a correlation between the microstructural
properties of the FAT and articulation rate, association which was not evident in fluent speakers.
È interessante notare, in patients with multiple sclerosis and in patients with chronic aphasia, the FAT
was associated with verbal fluency measures (Blecher et al., 2019; Catani et al., 2013; Li et al.,
2017).
In the current study, we evaluated the contribution of the CPs and the FAT to the different
measures of speech fluency in neurotypical adults. Participants (N=45) underwent an
unstructured interview to assess their natural speaking rate and articulation rate, and completed
timed semantic and phonemic fluency tasks to assess their verbal fluency. Diffusion MRI data
was measured in the same group of participants using a single-shell high angular resolution
imaging (HARDI) sequence (Tuch et al., 2003). The automatic fiber segmentation and
quantification (AFQ) package (Yeatman et al., 2012) was adapted in order to delineate the
bilateral SCP and MCP as they decussate at the level of the inferior colliculi and pons,
rispettivamente. Microstructural properties were extracted from the CPs and the FAT, and entered
into correlation analyses with measures of speech fluency. Based on computational models of
speech production (Hickok, 2012; Tourville & Guenther, 2011), and on previous neuroimaging
reports (Castellazzi et al., 2018; Gurd et al., 2002; Hubrich-Ungureanu et al., 2002; Schlösser et
al., 1998B; Sobczak-Edmans et al., 2019), we expected that the microstructural properties of the
cerebro-cerebellar loop (cioè., SCP and MCP) will be correlated with both articulatory and verbal
aspects of speech production. Importantly, this analysis is still exploratory, due to the lack of
prior data or modelling work relating each CP to specific language functionalities. Based on
previous studies of structural connectivity in clinical populations (Blecher et al., 2019; Catani et
al., 2013; Li et al., 2017), we hypothesized that the microstructural properties of the FAT will be
associated with verbal fluency measures in the current sample of neurotypical adults. Lastly,
based on our previous findings (Jossinger et al., 2021; Kronfeld-Duenias et al., 2016),
neurotypical adults were not expected to show an association between articulation rate and the
ICP or the FAT.
5
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
.
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
METHODS
Participants
Forty-five neurotypical adults (29 females; mean age 26.45 ± 3.72 years; Tavolo 1) were
recruited for this study. All the analyses reported here are completely new, but some data from
this sample has been reported as part of a research project focusing on associations between
well-known language pathways and word structure (Yablonski et al., 2021; Yablonski & Ben-
Shachar, 2020). All participants were right handed as estimated by the Edinburgh handedness
inventory (Oldfield, 1971; Tavolo 1), and had no history of a diagnosed speech impairment,
learning disability, or neurological condition. All participants were native Hebrew speakers who
speak and read English as L2. One participant was referred to neurological follow-up due to an
incidental finding, but otherwise had normal anatomical structure and diffusion values and was
thus not excluded from analysis. Participants were paid 200 NIS for their participation. Tutto
participants signed a written informed consent before participating in the study. This study was
approved by the Helsinki committee of the Sheba Medical Center, by the Institutional Review
Board of Tel Aviv University, and by the Ethics committee of the Faculty of Humanities in Bar-
Ilan University.
Tavolo 1. Sample characteristics (N=45).
Demographics
Gender
Age (years)
Education (years)
Handedness
Speech rate
Speaking rate (syllables/sec)
Articulation rate (syllables/sec)
Verbal fluency
Phonemic fluency (Words/min)
Semantic fluency (Words/min)
mean
SD
range
16M/29F
26.45
14.56
96.89
5.04
6.42
43.78
66.96
6
—
3.72
2.13
5.90
0.68
1.05
—
[20.23, 34.87]
[12, 20]
[80,100]
[3.49, 6.78]
[4.57, 9.08]
9.83
12.22
[17, 68]
[46, 99]
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
.
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Speech fluency assessment
Speaking task. Speaking rate and articulation rate were measured over audio recordings of
an unstructured interview. The interview took place in a quiet room, and was simultaneously
recorded with a noise-canceling microphone (Sennheiser PC21-II, Sennheiser Electronic
Corporation, Berlin, Germany) and with a digital video camera (Sony HDR-CX405, Sony
corporation of America, New York, NY, USA). Audio signals from the microphone were
digitally recorded using Audacity (https://audacityteam.org/) on a mono channel with a sampling
rate of 48 kHz (16 bit). The participant was asked to talk about a neutral topic (per esempio., a recent
travel experience, a movie, a book), for about 10 min. The experimenter (M.Y.) refrained from
interrupting, asking open questions only when the participant was having difficulty finding a
topic to talk about.
Speech rate measures. Two speech rate measures were calculated: Speaking rate and
articulation rate. Both measures were calculated over the audio recordings of the unstructured
interview. Articulation rate was calculated as the ratio between the total number of analyzed
syllables and the time it took the participant to produce them (cioè., syllables/sec), after excluding
disfluent utterances (Ambrose & Yairi, 1999; Amir, 2016; Amir & Grinfeld, 2011; Rochman &
Amir, 2013). Speaking rate was calculated in the same way, on the entire speech segment
without exclusions. For this purpose, two trained research assistants first transcribed each
interview and annotated any disfluent epochs (mostly naturally occurring disfluencies, ad esempio
hesitations, repetitions, and revisions). An utterance was defined based on three criteria: (1)
communicated an idea, (2) had a well-defined intonation contour, E (3) was grammatically
complete. Overall, a fixed number of 50 utterances were used to calculate Speaking rate, E
12.8 ± 6.36 utterances were used to calculate articulation rate.
Verbal fluency tasks. Normed Hebrew versions of the phonemic and semantic fluency tasks
were implemented (Kavé, 2006; Kavé & Knafo-Noam, 2015). Participants were asked to
produce as many words as possible within 1 minute according to a criterion: words beginning
with a certain letter (phonemic fluency tasks) or a semantic category (sematic fluency tasks).
Each task was repeated 3 times with different criteria. For the Phonemic fluency task,
participants were asked to produce words that begin with the letters Bet (/b/), Gimel (/g/), E
Shin (/ʃ/ or /s/). For the Semantic fluency task, participants were asked to produce words that
7
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
.
/
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
belong to the categories animals, fruits and vegetables, and vehicles. The administration order of
the tasks and the different items within each task was kept constant across participants, starting
with the phonemic fluency task and followed by the semantic fluency task. The experimenter
(M.Y.) read each criterion aloud, and participants’ oral responses were recorded and transcribed
offline.
Verbal fluency measures. Primo, responses in both tasks were screened according to the
guidelines described in Kavé & Knafo-Noam (2015). Accordingly, repetitions and erroneous
responses were removed. Per esempio, in the semantic fluency task, names of subcategories
(per esempio., birds) were not counted if the participant also produced specific exemplars within the
subcategory (per esempio., raven, pigeon). The total number of correct unique responses was coded per
criterion and summed across the three criteria within each task. We also calculated a
standardized verbal fluency score based on age-appropriate Hebrew norms (Kavé & Knafo-
Noam, 2015). The analysis of standardized verbal fluency scores in the current sample is
reported in a previous paper (Yablonski et al., 2021). Note, Tuttavia, that this previous paper did
not examine the associations between verbal fluency measures and the CPs, which is the focus of
the current study.
Behavioral inter-correlation analysis
Correlations between speech fluency components were calculated using the open-source R
environment for statistical analysis (R Core Team, 2013). Specifically, we calculated Spearman’s
correlations between speaking rate, articulation rate, phonemic fluency, and semantic fluency,
resulting in an inter-correlation matrix of 6 correlations overall. To account for multiple
comparisons, we controlled the false discovery rate (FDR) at a level of 5% (Benjamini &
Hochberg, 1995).
White matter analysis
MRI data acquisition
MRI scans were conducted on a 3T Siemens Magnetom Prisma scanner at the Strauss Center
for Computational Neuroimaging at Tel Aviv University, with a 64-channel head coil. The MRI
8
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
/
.
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
protocol included standard anatomical and diffusion imaging sequences, as detailed below.
Functional MRI scans were also included in the scanning protocol but are not reported here.
T1 image acquisition. High-resolution T1-weighted anatomical images were acquired using
a magnetization prepared rapid acquisition gradient echo (MPRAGE) protocol (TR=2.53 s,
TE=2.99 ms, flip angle=7º, 1 mm thick slices, matrix size: 224 ×224 ×176, voxel size: 1 ×1
×1mm).
Diffusion weighted image acquisition. A standard diffusion MRI (dMRI) protocol was
applied by means of a single-shot spin-echo diffusion-weighted echo-planar imaging (DW-EPI)
sequence (86 axial slices, each 1.7 mm thick, no gap; FOV = 204 ×204 mm, image matrix size =
120 ×120 providing a cubic resolution of 1.7 ×1.7 ×1.7 mm, TR = 4000 ms, TE = 58 ms). Sixty-
four diffusion-weighted volumes (b = 1000 s/mm2) and three reference volumes (b = 0 s/mm2)
were acquired using a standard diffusion direction matrix. Multiband acceleration was used with
slice acceleration factor of 2. Total acquisition time for the dMRI sequence was 4:48 min.
MRI data analysis
Software. Data analysis was conducted using Matlab 2012b (The Mathworks, Natick, MA).
Data preprocessing was obtained using the open source ‘mrDiffusion’ package
(https://github.com/vistalab/vistasoft/tree/master/mrDiffusion). Constrained spherical
deconvolution (CSD) diffusion model was calculated using the ‘mrTrix3’ toolbox
(https://www.mrtrix.org/; Tournier et al., 2019). Tract identification and quantification were
executed with the automatic fiber segmentation and quantification (AFQ) toolkit (
https://github.com/yeatmanlab/AFQ; Yeatman et al., 2012). Individual tracts were visually
inspected using Quench, an interactive 3D visualization tool
(http://web.stanford.edu/group/vista/cgi-bin/wiki/index.php/QUENCH; Akers, 2006).
Data preprocessing. Preprocessing was implemented in the native space of each individual,
following a published pipeline (Yablonski et al., 2021). This pipeline included several steps: (1)
Alignment of the T1 images to the anterior commissure –posterior commissure (AC-PC)
orientation; (2) Motion- and eddy-current correction of DW-EPI data (Rohde et al., 2004); (3)
Registration of the diffusion weighted volumes to the averaged non-diffusion weighted volumes
(b0); (4) Registration of the mean b0 image to the T1 image using a rigid body mutual
9
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
.
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
information maximization algorithm (implemented in SPM8; Wells et al., 1996); (5) Applying
the combined transform resulting from motion correction, eddy-current correction and
anatomical alignment to the raw diffusion data; (6) Adjusting the table of gradient directions to
fit the resampled diffusion data (Leemans & Jones, 2009).
At the voxel level, the diffusion data was modeled twice, once with the tensor model and
once with the constrained spherical deconvolution (CSD) modello. Diffusion tensors were used to
calculate, within each voxel, the commonly used diffusion metrics (per esempio., fractional anisotropy,
eccetera.), which were later used for the statistical analysis. Tensor modeling was carried out using
‘mrDiffusion’. Tensors were fit to the registered diffusion data using a robust least-square
algorithm (RESTORE), which removes outliers at the tensor estimation step (Chang et al., 2005).
Then, using tensor decomposition, we extracted the three eigenvalues and eigenvectors of the
tensor, and calculated, within each voxel, fractional anisotropy (FA) and mean diffusivity (MD).
Specifically, FA was calculated as the normalized standard deviation of the eigenvalues, and MD
was calculated as the average of the three eigenvalues (Basser & Pierpaoli, 1996).
As the tensor model is notoriously known for its inability to account for multiple orientations
within the same voxel (Jones et al., 2013), our individual tractograms were generated based on
the CSD model. The CSD model estimates the fiber orientation distribution (FOD) within each
voxel based on the spherical deconvolution between the diffusion weighted signal and an
estimation of the diffusion response function (Tournier et al., 2004, 2007). To calculate the CSD
model we used the ‘mrTrix’ toolbox (Tournier et al., 2019). Primo, to estimate the response
function we used the dhollander algorithm (implemented within the dwi2response function).
This algorithm estimates the response functions for white matter, gray matter, and cortico-spinal
fluid (CSF), separately, based on single-shell diffusion data. Prossimo, to estimate the FOD, we
applied constrained spherical deconvolution with up to eight spherical harmonics (lmax = 8) SU
the responses estimated within the white matter and the CSF. This step was carried out by the
msmt_csd algorithm (implemented within the dwi2fod function) (Jeurissen et al., 2014). Lastly,
the FODs were used to generate individual tractograms using probabilistic tractography, COME
explained below.
Fiber tractography. Individual tractograms were generated using probabilistic whole brain
tractography, with the iFOD2 tracking algorithm (implemented in the mrTrix3 function tckgen).
10
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
.
/
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
.
/
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
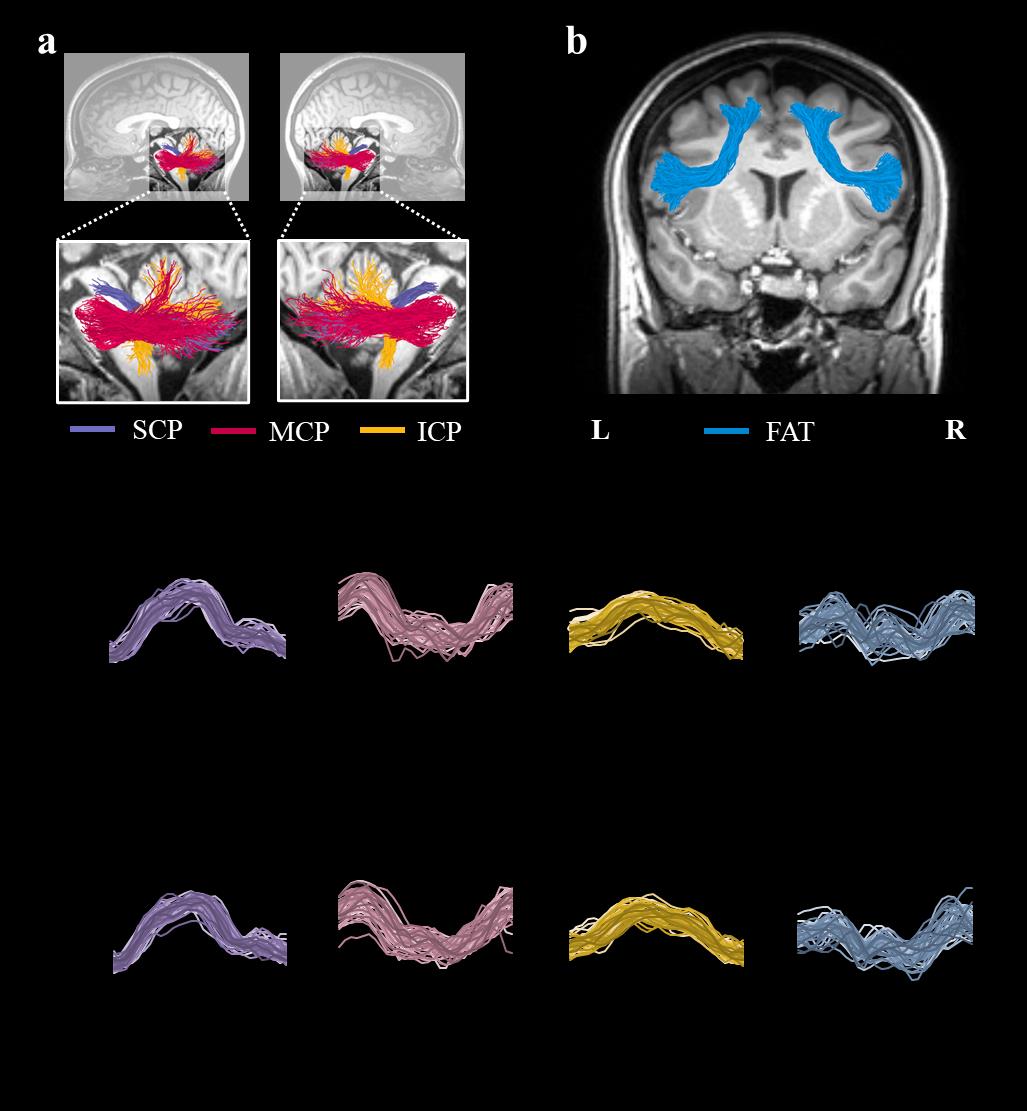
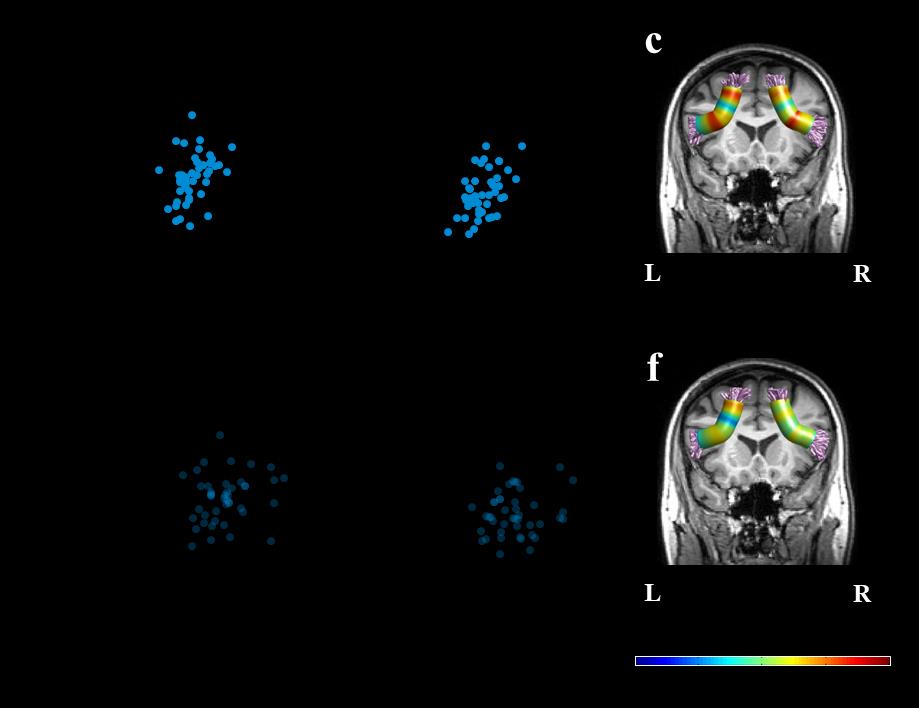
Figura 1. Tracts of interest. UN) Shown are the left and right cerebellar peduncles identified in a single
participant (female, 34), overlaid on a T1 image. The cerebellar tracts identified are the superior
cerebellar peduncle (SCP; purple), middle cerebellar peduncle (MCP; magenta), and the inferior
cerebellar peduncle (ICP; yellow). B) Shown are the left and right frontal aslant tract (FAT; blue)
identified in a single participant (female, 24), overlaid on a T1 image. c-d) Individual tract profiles
show FA values at 30 equidistant nodes along the core of the left (C) and right (D) tracts of interest.
Each subject is represented by a single line (N=45). Abbreviations: SCP – superior cerebellar peduncle;
MCP – middle cerebellar peduncle; ICP – inferior cerebellar peduncle; FAT – frontal aslant tract; L –
left; R – right; Inf. – inferior; Sup. – superior.
11
As a first step, a whole brain white matter mask was generated from each participant’s structural
T1 image. This procedure was done using the 5ttgen script, utilizing FSL tools to perform whole
brain segmentation (Smith et al., 2004). The tracking was initialized from 500,000 random seeds
within the white matter mask, and was restricted with the following parameters: (1) FOD
amplitude threshold of 0.1; (2) 45º maximum angle between successive steps; (3) 0.85 mm step
size; (4) streamline length between 50mm to 200mm. Streamlines that extended beyond the
white matter mask were truncated. The resulting whole brain tractograms were then subjected to
the automatic tract segmentation procedure.
Tract identification and segmentation. In each individual’s native space, we identified the
bilateral CPs and the bilateral FAT (Figura 1). We targeted these tracts based on previous studies
in clinical populations, directly linking the CPs (Figure 1a) and the FAT (Figure 1b) to speech
fluency (Blecher et al., 2019; Catani et al., 2013; Connally et al., 2014; Jossinger et al., 2021;
Kronfeld-Duenias et al., 2016; Li et al., 2017), and based on theoretical models of speech
production (Hickok, 2012; Tourville & Guenther, 2011). Automatic segmentation of the tracts
was carried out using AFQ. This method utilizes a multiple-ROI approach in which the
tractograms are intersected with pre-defined ROIs using logical operations (Figure S1). In
accordance with this method, the ROIs are universally defined on a template (Figure S1a), E
then back-transformed to the participant’s native space using a non-linear transformation (Figura
S1b). The individual ROIs are intersected with the tractogram (Figure S1c) to isolate the tracts of
interesse (Figure S1d). To identify the CPs we propose a new protocol which is based on several
previous studies (Bruckert et al., 2019; Palesi et al., 2015, 2017). To identify the bilateral FAT,
we used the protocol described in Kronfeld-Duenias et al. (2016), which is implemented in AFQ
(publicly available at https://github.com/yeatmanlab/AFQ/tree/master/aslant).
Automatic segmentation of the CPs was recently introduced by Bruckert et al. (2019). Questo
approach, Tuttavia, delineates only the inferior parts of the SCP and MCP, before they decussate
to the contralateral cerebral hemisphere. For this reason, the methods introduced in Bruckert et
al. (2019) are better suited to delineate the CPs based on deterministic tractography, where fibers
are less likely to decussate. D'altra parte, probabilistic tractography approaches coupled
with CSD modeling are more successful in following the CPs as they decussate. In order to
segment the resulting tracts automatically, we revised the AFQ protocol including additional
ROIs (see Fig. S2 and Table S1 for detailed MNI coordinates of the ROIs).
12
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
.
/
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
The new set of ROIs used to identify the CPs were defined on the Montreal Neurological
Institute (MNI) template (ICBM 2009a Nonlinear Asymmetric template; Fonov et al., 2011). For
the SCP, a new ROI (SCP_superior_prob) was defined on an axial slice at the level of z = -10,
encompassing a rectangle around the red nucleus (Oishi et al., 2009; Palesi et al., 2015). IL
SCP was then segmented using the newly defined ROI together with the previous SCP-ROIs
described by Bruckert et al. (2019) (Figure S2a). For the MCP, a new ROI (MCP_superior_prob)
was defined as a parallelogram on an axial slice at the level of z = -16, including all the voxels
occupied by the cerebral peduncle (Oishi et al., 2009; Palesi et al., 2017). The MCP was then
segmented using the newly defined ROI together with the previous MCP-ROI introduced by
Bruckert et al. (2019) (Figure S2b). For the ICP, a new inferior ROI (ICP_inferior_prob) era
defined to avoid fibers that enter the pons. The ICP was then segmented using the newly defined
ROI together with the previous ICP-ROI introduced by Bruckert et al. (2019) (Figure S2c).
The resulting tracts were cleaned automatically using a statistical outlier rejection algorithm
implemented in AFQ. For the cleaning procedure of the SCP, MCP and FAT, fibers were
considered outliers if they were longer than 4 standard deviations from the mean fiber length and
spatially deviated more than 4 standard deviations from the core of the tract. For the ICP, UN
relatively short tract, the length criterion was changed as previously done in (Bruckert et al.,
2019,) such that fibers longer than 1 standard deviations from the mean fiber length were
removed. The tracts are shown in Fig. S3 in 3 representative subjects.
Brain-behavior correlation analysis
The analyses were restricted to the core segment of each tract, enclosed between the two
waypoint-ROIs (Figure S2). This approach eliminates the extreme segments of the tracts which
are highly variable between participants.
Mean-tract correlations. Associations between diffusivity values and speech fluency
measures were assessed using two-tailed Spearman’s rank-order correlations. As a first step, we
calculated for each participant and each tract the average FA and average MD values across the
core of the tract (cioè., tract-FA and tract-MD, rispettivamente). Then, we assessed the simple
correlations between tract-FA and speech fluency measures. We controlled for multiple
comparisons across 8 tracts setting the FDR at a level of 5% (Benjamini & Hochberg, 1995). UN
similar analysis was conducted on tract-MD values. Separate analyses were conducted for each
behavioral measure (Xie et al., 2011).
13
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
.
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Along-tract correlations. It is well documented that diffusivity values vary significantly
along the tract (Yeatman et al., 2011, 2012). This is also true for the CPs and the FAT, come mostrato
in Figure 1c-d and Figure S4. We therefore applied a second analysis, assessing two-tailed
Spearman’s correlations between speech fluency measures and diffusivity values at 30
equidistant locations along the core of the tract (Kruper et al., 2021). To account for multiple
comparisons, significance was corrected using a non-parametric permutation test, controlling the
family-wise error (FWE) at a corrected alpha value of 0.05 (Nichols & Holmes, 2002). Clusters
were considered significant if they satisfied two criteria: (1) each node within the cluster was
significantly correlated with speech fluency at a level of alpha = 0.05 (uncorrected), E (2) IL
number of adjacent nodes composing the cluster should have been larger than a critical size,
determined by the permutation algorithm (Nichols & Holmes, 2002; Yeatman et al., 2012). FWE
correction was applied across the 30 nodes within each tract, with no further correction across
tracts, behavioral measures, or diffusion metrics (see e.g., Yablonski et al., 2021, Yeatman et al.,
2012 for similar approach).
Partial correlation. To assess the specificity of associations and dissociate between tracts
mediating lexical or articulatory components of speech fluency, significant associations were
followed up by Spearman’s partial correlations. Per esempio, associations with speaking rate
were followed up with partial correlation analyses that held constant the contribution of
articulation rate.
RESULTS
Behavioral correlations
A correlation matrix of the four speech production measures (Figure 2a, Table S2)
demonstrated significant correlations between the Speaking- and Articulation- rates (r = 0.599, P
< 10-4), and between the semantic- and phonemic- fluency scores (r = 0.548, p < 10-4; both correlations were significant when controlling the FDR at q<0.05). However, non-significant correlations were found between the verbal fluency measures and the speech rate measures (p >
0.06, uncorrected). Articulation rate was higher than speaking rate in all participants (Figure 2b;
14
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
.
/
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
.
/
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
/
.
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 2. Behavioral dissociation between verbal fluency measures and speech rate measures. UN)
Correlation matrix depicting two-tailed Spearman’s correlation coefficients between the four fluency
measures calculated: speaking rate (SR), articulation rate (AR), phonemic fluency (PF), and semantic
fluency (SF). The color saturation and size of the circles are proportional to the correlation coefficients.
Panels (B) E (C) depict the associations between speaking and articulation rate (B), and between
phonemic and semantic fluency (C). Both correlations were significant after controlling the FDR across
all behavioral measures at q < 0.05. Dashed lines delineate y=x. Abbreviations: SR – speaking rate; AR
– articulation rate; PF – phonemic fluency; SF – semantic fluency.
consistent with (Amir, 2016). Semantic fluency scores were higher than phonemic fluency scores
in 44/45 participants (Figure 2c; consistent with (Kavé, 2005)). Repeating this analysis with age
standardized scores (Kavé & Knafo-Noam, 2015) generated a similar pattern of results (see
Table S3).
Tract identification
The bilateral cerebellar peduncles and the bilateral frontal aslant tract were successfully
identified in all 45 participants (Figure 1). A qualitative examination of the tractograms (Figure
1a-b) and the individual tract profiles (Figure 1c-d) verified that the tracts were accurately
segmented and provided consistently shaped profiles in all individuals (see Figure S3 for tract
reconstruction in 3 representative participants).
Speech fluency associations within the cerebellar peduncles
To examine the relationships between speech fluency and the microstructural properties of
the cerebellar peduncles in neurotypical adults, Spearman’s correlations were first calculated
between the four measures of speech fluency (speaking rate, articulation rate, phonemic fluency,
and semantic fluency) and mean tract diffusivities (tract-FA and tract-MD; see Methods) within
15
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
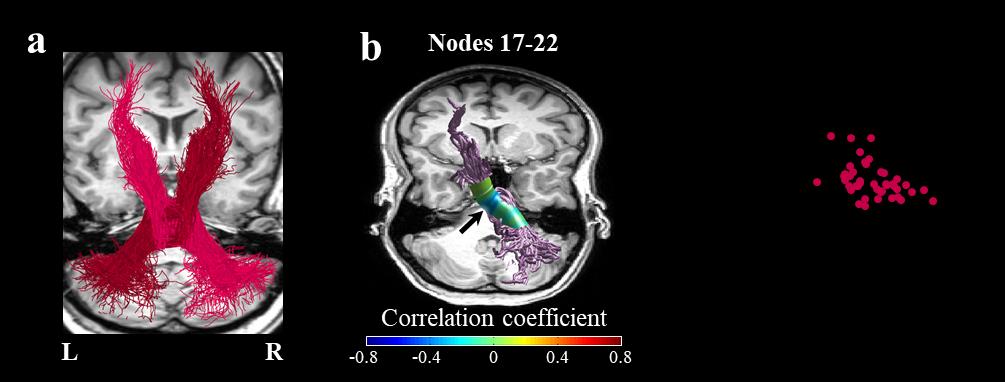
Figure 3. Mean diffusivity in the right middle cerebellar peduncle correlates with speaking rate.
a) The full trajectories of the right MCP (magenta) and the left MCP (burgundy) are shown in a
representative subject (female, 27). b) Two-tailed Spearman’s correlation coefficients are visualized in
30 nodes along the core of the right MCP. The black arrow denotes the location of the significant
cluster of nodes (nodes 17-22, p<0.05, family-wise error corrected across 30 nodes). c) A scatter plot
showing the association between speaking rate and the averaged MD in the significant cluster of nodes
within the right MCP. Abbreviations: MCP – middle cerebellar peduncle; MD – mean diffusivity; L –
left; R – right.
each of the cerebellar peduncles. No significant correlations were detected between speech
fluency and mean tract diffusivities in the cerebellar peduncles. Calculating the correlation for
the age-standardized verbal fluency measures did not change these results. See Tables S4-S5 for
a detailed list of correlation values.
To achieve enhanced sensitivity for detecting localized brain-behavior correlations, we
examined the relationships between speech fluency and local diffusivity values, node-by-node,
along the trajectory of each cerebellar peduncle (see Methods). This analysis revealed a
dissociation in the pattern of correlations within the cerebellar peduncles, such that MD within
the right MCP was significantly correlated with speaking rate (r = -0.447, p < 0.05, nodes 17-22,
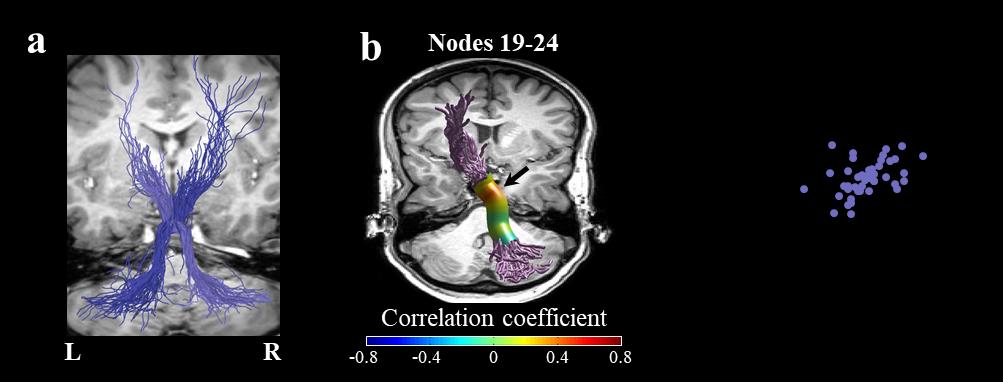
FWE corrected across 30 nodes; Figure 3), whereas FA within the right SCP was significantly
correlated with phonemic fluency (r = 0.431, p < 0.05, nodes 19-24, FWE corrected across 30
nodes; Figure 4). Note that both significant clusters were detected in the vicinity of the
decussation. We address this point in the discussion.
Importantly, these correlations remained significant when calculating partial correlations,
such that the correlation between the right MCP and speaking rate was not driven by phonemic
fluency (r = -0.451, p = 0.002), and the correlation between the right SCP and phonemic fluency
was not driven by speaking rate (r = 0.438, p = 0.003). Moreover, controlling for the contribution
16
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Fractional anisotropy in the right superior cerebellar peduncle correlates with
phonemic fluency. a) The full trajectories of the right SCP (light purple) and the left SCP (dark purple)
are shown in a representative subject (female, 27). b) Two-tailed Spearman’s correlation coefficients
are visualized in 30 nodes across the core of the right SCP. The black arrow denotes the location of the
significant cluster of nodes (nodes 19-24, p<0.05, family-wise error corrected across 30 nodes).. c) The
scatter plot shows the association between phonemic fluency and mean FA in the significant cluster of
nodes within the right SCP. Abbreviations: SCP – superior cerebellar peduncle; FA – fractional
anisotropy; L – left; R – right.
of articulation rate did not change these effects (Right MCP: r = -0.477, p = 0.001; Right SCP: r
= 0.442, p = 0.003).
No additional significant correlations were found between the microstructural properties of
the cerebellar peduncles and speech fluency measures (Table S4). Importantly, in line with a
previous report (Jossinger et al., 2021), we did not find significant correlations between
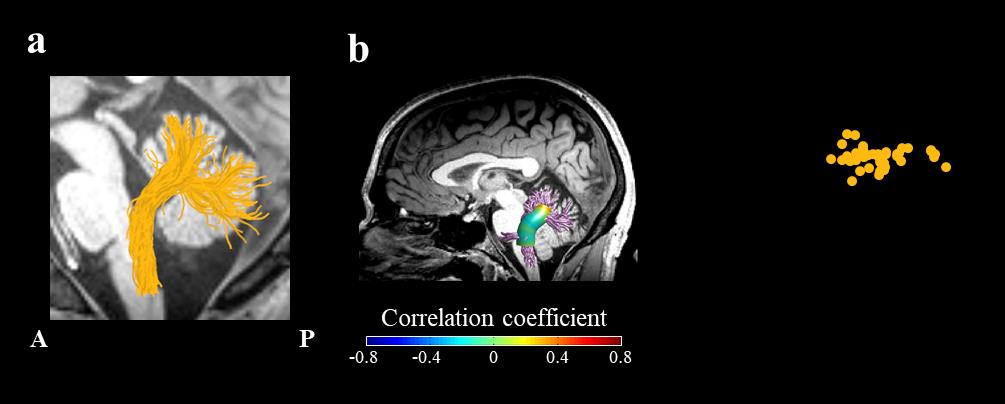
articulation rate and diffusivities within the left ICP of neurotypical adults (p > 0.5; Figura 5).
Notice that, because there was no significant cluster of nodes in the ICP, the scatter plot in
Figure 5c depicts tract-FA (rather than cluster FA) against articulation rate.
Speech fluency associations within the bilateral frontal aslant tracts
To examine the relationships between speech fluency and the microstructural properties of
the FAT in neurotypical adults, Spearman’s correlations were first calculated between the four
measures of speech fluency (speaking rate, articulation rate, phonemic fluency, and semantic
fluency) and tract diffusivities (tract-FA and tract-MD; see Methods) within the bilateral FAT
(Table S4). This analysis revealed that tract diffusivities within the bilateral FAT were
17
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
N
o
/
l
/
l
UN
R
T
io
C
e
–
P
D
F
/
D
o
io
/
l
/
.
/
1
0
1
1
6
2
N
o
_
UN
_
0
0
0
9
8
2
0
6
6
8
8
6
N
o
_
UN
_
0
0
0
9
8
P
D
/
.
l
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 5. No correlation between the FA in left inferior cerebellar peduncle and articulation rate.
UN) The trajectory of the left ICP (yellow) is shown in a representative subject (female, 24). B) Two-
tailed Spearman’s correlation coefficients are visualized in 30 nodes across the core of the left ICP. C)
Articulation rate is plotted, for each participant, against tract-FA (the mean FA across 30 nodes in the
left ICP, See Methods). Abbreviations: ICP – inferior cerebellar peduncle; FA – fractional anisotropy;
A – anterior; P – posterior.
significantly correlated with speaking rate (Figure 6a-c). Specifically, speaking rate was
positively correlated with tract-FA within the left FAT (r = 0.459, P < 0.002; Figure 6a) and with tract-FA within the right FAT (r = 0.466, p < 0.002; Figure 6b) (both correlations were significant when controlling the FDR across 8 tracts at q<0.05). In both cases, neurotypical adults who speak faster have higher FA within their left and right FAT. In line with a previous report (Kronfeld-Duenias et al., 2016) no significant correlations were found between tract-FA within the bilateral FAT and articulation rate (p > 0.1; Figure 6d-f). Inoltre, the correlations
between the bilateral FAT and speaking rate remained significant when calculating partial
correlations, controlling for the effect of articulation rate (Left FAT: r = 0.465, p = 0.0014; right
FAT: r = 0.488, p = 0.0007). Fisher’s Z test for dependent samples confirmed that the
correlations between tract-FA and speaking rate differed significantly from the correlations with
articulation rate (left FAT: Fisher’s Z = 2.396, P <0.009; right FAT: Fisher’s Z = 2.605, p <
0.006). See Tables S4-S5 for detailed correlation values between tract-FA and tract-MD values
of the FAT and all speech fluency measures.
18
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. The bilateral frontal aslant tract is associated with speaking rate, but not with
articulation rate. Scatter plots show the association between the number of syllables per second and
tract-FA within the left FAT (a, d) and the right FAT (b, e). Tract-FA in the left and right FAT
significantly correlated with speaking rate (blue; a-b, q<0.05, FDR corrected across all tracts), but not
with articulation rate (light blue; d-e). For completeness, Spearman’s correlation coefficients between
FA and speaking rate, and between FA and articulation rate are visualized in 30 nodes along the left
and right FAT (panels c and f, respectively). The correlations along the tracts did not pass FWE
correction for 30 nodes. Abbreviations: FAT – frontal aslant tract; FA – fractional anisotropy; L – left;
R – right.
For completeness, we also calculated the correlations with speech fluency measures along the
trajectory of the bilateral FAT (see Fig. 6c for Speaking rate and 6f for Articulation rate). In line
with tract-FA results (6a-b), several nodes showed positive local correlations with Speaking rate
(Fig. 6c). However, these associations did not survive the FWE correction. Significant negative
correlation was detected between speaking rate and MD within the right FAT (nodes 10-24; r = -
0.425, p < 0.05 FWE correction across 30 nodes; Figure S4). This correlation remained
significant when calculating partial correlation, controlling for the effect of articulation rate (r = -
0.325, p = 0.03). No other significant correlations were found between diffusivities within the
bilateral FAT and speech fluency measures (Table S4).
19
DISCUSSION
The goal of the current study was to evaluate the contribution of the CPs and the FAT to the
fluent production of speech in neurotypical adults. Our findings demonstrate that the cerebellar
peduncles dissociate between different speech fluency measures, such that the right MCP was
associated with speaking rate while the right SCP was associated with phonemic fluency. These
correlations were not driven by articulation rate. Further, we found that the bilateral FAT is also
associated with speaking rate, but not with articulation rate. These findings shed light on the
functional contributions of distinct subsystems involved in speech production. We discuss the
findings concerning different aspects of speech fluency as they relate to the CPs and the FAT in
the following sections.
Dissociation between verbal fluency and speech rate measures
The production of fast and fluent speech requires interaction between several processes,
including high-level linguistic components, such as lexical access and phonological encoding,
and motor components which are crucial for articulatory control. In the current study we
assessed these different components by using different types of tasks that require overt
production of speech: verbal fluency tasks and an unstructured interview. Our data show a
behavioral dissociation between verbal fluency measures and speech rate measures (Figure 2).
Moreover, we found that specific fluency measures mapped unto different white matter tracts.
Together, these findings suggest that different fluency measures may reflect different aspects of
speech production.
Task differences
The tasks we used to evaluate speech fluency differ in several aspects. First, an unstructured
interview is a relatively naturalistic setting of speech production, while verbal fluency tasks
measure speech production in a more artificial way. Second, producing a free narrative involves
conceptualizing the message and projecting a syntactic structure, while a verbal fluency task
elicits a list of words, mostly unconnected nouns, based on a predetermined criterion. Third, each
task may involve different cognitive demands beyond speech production. For example, verbal
fluency tasks require the use of executive functions such as response selection, the inhibition of
irrelevant responses, and more (Friedman et al., 1998; Kramer et al., 2014). Spontaneously
speaking during an unstructured interview, on the other hand, may be effected by the social and
20
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
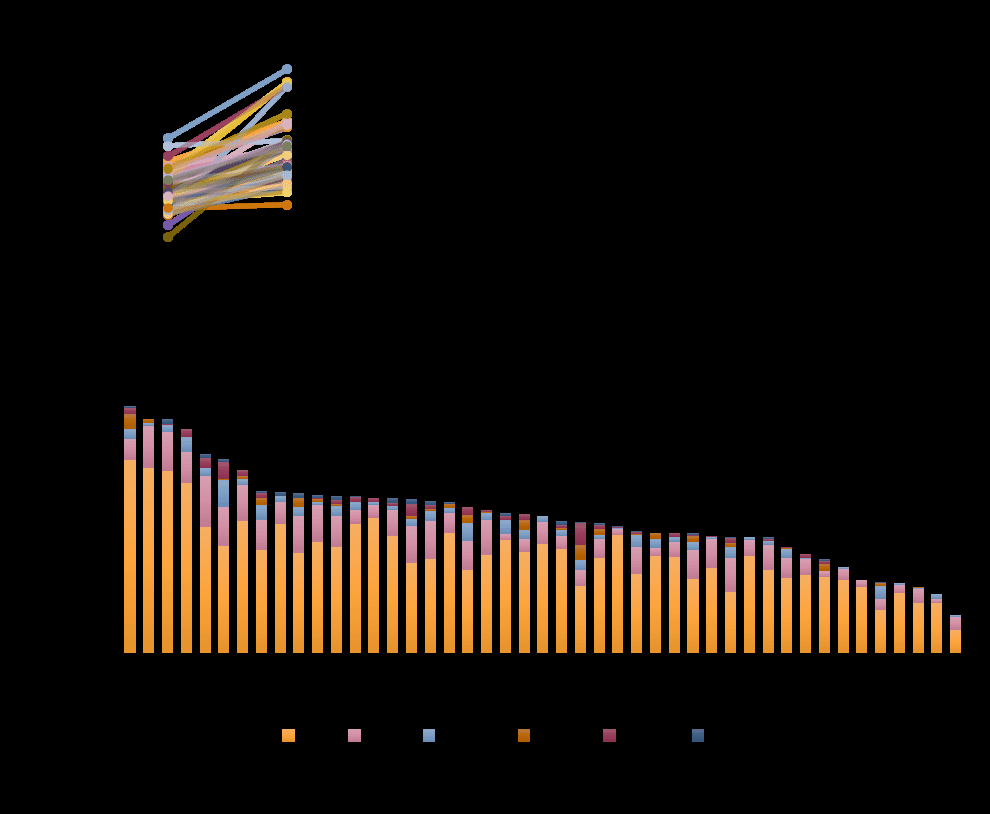
Figure 7. Analysis of individual speaking rate. (a) Individual speaking rates and articulation rates.
Each line represents a single participant. In all participants, speaking rate is slower compared to
articulation rate. (b-c) Speaking rate, but not articulation rate, is significantly associated with the
percent of dysfluencies (r=-0.51, p = 0.0003 compared with r = 0.09, p = 0.56), such that slower
speakers demonstrate more dysfluencies while they speak. (d) Distribution of dysfluencies by type. In
our sample of neurotypical speakers, most dysfluencies included interjections (I%; e.g., “um”) and
revisions (R%; e.g., “so I was, we were…”). Other dysfluencies that occurred at much lower rates were
part-word repetitions (PW%; e.g., “In, in..”), stuttered syllables (SS%; e.g., “The, the..”), phrase
repetitions (PR%; e.g., “..because the flight was… the flight was”), and dysrhythmic phonation (DP%;
e.g., “looooooves).
emotional state of the speaker (Amir, 2016). The extent to which each of the two speech fluency
tasks used in this study relies on other cognitive functions, which are not speech-related, remains
to be studied directly in future studies.
21
What does speaking rate actually measure?
Despite the considerable correlation between articulation rate and speaking rate (Figure 2b),
the two metrics are thought to represent different aspects of speech production. Speaking rate,
which is calculated over continuous segments of speech, including disfluencies of various
sources, is considered a global measure of verbal output and language proficiency (Costello &
Ingham, 1984). Articulation rate, in contrast, is calculated over the same speech samples but after
excluding disfluent segments, thus thought to reduce linguistic effects and to represent
articulatory motor control (Walker et al., 1992).
To better understand the sources of the differences between speaking and articulation rates,
we conducted a more elaborate analysis of the individual speech segments, as described in
Figure 7. As expected, in all participants, speaking rate is slower compared to articulation rate
(Figure 7a). This difference may reflect naturally occurring disfluencies, which are included in
the calculation of speaking rate, not articulation rate. In line with this view, speaking rate, but not
articulation rate, is negatively correlated with the frequency of dysfluencies, such that slower
speakers demonstrate more dysfluencies (compare Figure 7b and 7c). In our sample of
neurotypical adults, the vast majority of dysfluencies included interjections (e.g., “um”) and
revisions (e.g., “so I was, we were…”) (see Figure 7d). Such naturally occurring dysfluencies are
commonly observed in neurotypical speakers and may reflect difficulties in formulating the
message at different levels, e.g., the conceptual, syntactic or pragmatic levels.
Cerebellar internal models of speech and language
The cerebellum has long been viewed as a motor control structure. Over the past two
decades, however, evidence points to a more general role for the cerebellum in the modulation of
higher-level cognitive processes (Schmahmann, 2010). Our findings support this view by
showing that the white matter pathways that connect the cerebellum to the cerebrum are
important for the modulation of both speaking rate and phonemic fluency. Importantly, these
correlations remain significant after we partial out the effect of articulation rate, a proxy for
motor control. Therefore, the selective association patterns highlight the unique contribution of
the cerebellum to linguistic, rather than motor components of speech, in neurotypical adults.
22
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cerebellar internal models
Studies focusing on the cerebellar computations postulate that it encodes “internal models”,
corresponding to the neural representations that we generate for the external world (Ito, 2008;
Kawato, 1999; Shadmehr & Krakauer, 2008; Wolpert et al., 1998). This concept developed from
contemporary theories of motor control, arguing that cerebellar internal models capture the
causal relationships between a voluntary movement and its consequence. Internal models are
used to predict the outcome of an action before the arrival of the actual sensory feedback from
the environment. This mechanism is often used to explain how fast and coordinated movements
can be carried out accurately, regardless of the slow nature of sensory feedback loops (Shadmehr
& Krakauer, 2008; Wolpert et al., 1998). Indeed, current models of speech production argue that
cerebellar internal models are used to transform motor-to-sensory information which is crucial
for articulation, thus enabling fast and fluent production of speech (Guenther, 2006; Hickok,
2012).
Internal models of language in the cerebellum
The accumulative evidence implicating the cerebellum in high-level cognitive functions
raises the possibility that cerebellar internal models are utilized to simulate mental
representations, similarly to the manner in which they operate on motor behaviors (Ito, 2008).
With respect to language processing, cerebellar internal models are hypothesized to store
memories of sequential linguistic events, such as phonological, semantic and syntactic
sequences, which are used to predict upcoming linguistic events during speech comprehension
(Argyropoulos, 2016). By showing an association between the cerebero-cerebellar pathways and
speech production, we may speculate that the cerebellum functions not only as an internal model
during speech comprehension but may also contribute to prediction of linguistic elements during
speech production.
Cerebro-cerebellar pathways mediate speech fluency in neurotypical adults
The hypotheses regarding the involvement of the cerebellum in cognition and language are
largely based on the anatomical connections between the cerebellum and the cerebral cortex. In
the late 1900’s it was suggested that in humans, the increased size of the cerebellum and cerebral
cortex led to the formation of new cerebro-cerebellar pathways that may contribute to high-level
functions such as language (Leiner, 2010; Leiner et al., 1986). Connections between the
23
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
cerebellum and the prefrontal cortex were indeed traced in both monkeys (Middleton & Strick,
1994) and humans (Palesi et al., 2015, 2017). Recently, the cerebro-cerebellar pathways were
implicated in various reading tasks that require overt production of speech (Bruckert et al., 2020;
Travis et al., 2015). This is the first study, however, that shows a direct link between the
microstructural properties of the cerebro-cerebellar pathways and speech fluency that goes
beyond articulation, providing an anatomical and functional support to the hypotheses made in
the 1980’s (Leiner et al., 1986) .
The cerebro-cerebellar pathways dissociate between speech fluency measures
Our results point to distinct patterns of association between speech fluency measures and the
microstructural properties of the cerebro-cerebellar pathways. Specifically, we found that the
right SCP is associated with phonemic fluency (but not with speaking rate, see Figure 4 and
accompanying text), while the right MCP is associated with speaking rate (but not with
phonemic fluency, see Figure 3 and accompanying text). The difference in the functionalities
associated with each peduncle is in line with the difference in the type of signals conveyed by
each: the SCP is mainly an output pathway, transmitting signals from the cerebellar deep nuclei
into the contralateral cerebral hemisphere, while the MCP is mainly an input pathway, feeding
signals from the cerebral cortex into the contralateral cerebellar hemisphere. We may speculate
that the involvement of the right SCP in phonemic fluency reflects the retrieval of phonological
sequences from the cerebellar internal model, while the involvement of the right MCP in
spontaneous speaking rate reflects the evaluation of the linguistic content against its internal
representation. Future neurophysiological investigations will be essential to directly test this
interpretation.
Functional lateralization within the cerebellum
Our findings demonstrate a functional lateralization within the cerebellum, by associating
linguistic aspects of speech production with microstructural properties of the right SCP and right
MCP. The concept of the “lateralized linguistic cerebellum”, implicating the right cerebellar
hemisphere in linguistic operations (Marien et al., 2001), has been well established in previous
studies. For example, tasks that require lexical access and word retrieval were shown to activate
the inferior lateral part of the right cerebellar hemisphere in neurotypical adults (Petersen et al.,
1988, 1989). Further, patients with damage to the right cerebellar hemisphere were shown to
24
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
have a specific deficit in tasks that require semantic retrieval (Fiez et al., 1992) or grammatical
production (Silveri et al., 1994; Zettin et al., 1997). The right cerebellar hemisphere is
reciprocally connected to the left cerebral hemisphere via the right SCP and the right MCP. Our
findings provide an independent support in healthy adults for the involvement of the right
cerebellum in speech and language.
No significant correlations between the CPs and articulation rate in neurotypical adults
Articulation rate was not associated with the microstructural properties in any of the CPs
among our sample of neurotypical adults (Table S4). Of particular relevance is the non-
significant association between articulation rate and diffusivities within the left ICP (Figure 5), in
agreement with our previous findings in an independent, smaller sample of neurotypical adults
(Jossinger et al., 2021). In that study, articulation rate was associated with FA along the left ICP
only in a group of adults with persistent developmental stuttering and not in age-matched fluent
speakers (see Fig. 6 in Jossinger et al., 2021). The ICP, transmitting sensory feedback signals
from the periphery to the cerebellum, was repeatedly implicated in encoding motor errors during
various motor tasks, including reaching movements (Shadmehr, 2017) and locomotion (Jossinger
et al., 2020), which are crucial for the formation and updating of speech-related internal models
(Tourville & Guenther, 2011). It was suggested that adults who stutter rely too heavily on
sensory feedback while they speak due to a noisy or insufficient internal model (Hickok et al.,
2011; Max et al., 2004). In neurotypical adults, on the other hand, a well-functioning internal
model may be sufficient for producing fast and fluent speech, thus diminishing the speakers’
dependency on sensory feedback in order to produce adequate articulatory output.
An alternative explanation for the lack of correlation between articulation rate and the
microstructural properties of the ICP may rest in the type of task used to evaluate articulation
rate. In clinical populations, diagnosed with various speech disorders, articulation rate is
typically calculated over audio recordings of an unstructured interview. In neurotypical speakers,
however, natural speaking may be insufficient to challenge the articulatory system in order to
reveal its underlying neural mechanisms. To cope with this limitation, future studies could use
the Diadochokinetic rate (DDK), a task designed to stress the articulatory system by measuring
how quickly a person can produce a series of alternating sounds.1
1 We are thankful to an anonymous NOL Reviewer for this idea.
25
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The involvement of the frontal aslant tract in speech fluency
The FAT connects two cerebral areas important for speech production: the posterior inferior
frontal gyrus (also known as “Broca’s area”) with the pre-supplementary motor area (pre-SMA)
and SMA (Catani et al., 2013). Accumulative evidence from the last decade, mostly from clinical
data, suggests that the FAT is a key pathway for speech fluency, as detailed below. The current
study supplies further anatomical evidence for the involvement of the FAT in the fluent
production of speech.
The FAT mediates speaking rate in neurotypical adults
Our results show a positive correlation between diffusivity of the bilateral FAT and speaking
rate in neurotypical adults (Figure 6a and b; Figure S4). Specifically, we found that adults who
speak faster have higher mean FA within their left and right FAT, and lower MD within their
right FAT. The involvement of the FAT in speaking was previously demonstrated in patients.
Electrical stimulation of the FAT during neural surgery was shown to elicit speech arrest (Fujii et
al., 2015; Kinoshita et al., 2015; Vassal et al., 2014). In patients with primary-progressive
aphasia and in patients with multiple sclerosis, the microstructural properties of the FAT were
correlated with measures of speech rate and verbal fluency (Blecher et al., 2019; Catani et al.,
2013; Li et al., 2017). To the best of our knowledge, this is the first study that shows a direct
involvement of the bilateral FAT in speech production among healthy subjects.
No significant correlation between the FAT and articulation rate in neurotypical adults
Although speaking rate and articulation rate are highly correlated (Figure 2b), the bilateral
FAT was associated with speaking rate, but not with articulation rate, and the difference between
these correlation coefficients was significant. These findings are in line with previous results
showing a significant association between the left FAT and articulation rate only in adults with
persistent developmental stuttering, not in neurotypical adults (Kronfeld-Duenias et al., 2016). A
recent study, investigating the same sample of neurotypical adults reported here, found that the
bilateral FAT was also implicated in a morphological task that requires overt speech, beyond the
contribution of verbal fluency scores (Yablonski et al., 2021). Taken together, these findings
suggest that, in neurotypical adults, the FAT contributes to higher-level aspects of language
production, rather than to articulatory motor control per-se.
26
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Interpreting the results in terms of tissue properties
Both FA and MD are modulated by multiple biological factors, such as axonal density,
directional coherence, axonal diameter and myelin content, which may influence FA and MD in
opposite directions (Assaf & Pasternak, 2008; Beaulieu, 2002; Jones et al., 2013). For example,
tight axonal packing or elevated myelination which lead to better neuronal communication,
would manifest in elevated FA but reduced MD in the same voxel. On the other hand, wider
axonal diameter, promoting efficient conduction (Liewald et al., 2014), would result in reduced
FA and elevated MD (Barazany et al., 2009; Horowitz et al., 2015). Broadly speaking, however,
FA and MD are not negatively correlated across different pathways, and may therefore reflect
different components of the underlying tissue (De Santis et al., 2014; Uddin et al., 2019).
In our data, better phonemic fluency and faster speaking rate are associated with higher FA
values in the right SCP and the bilateral FAT, respectively. Such positive correlations may be
mediated by axonal packing or myelination: elevated axonal packing (or myelin content) would
lead to both higher FA and better conductance, hence better performance. In addition to the
positive correlations observed with FA metrics, negative correlations between MD and speaking
rate were detected in the right MCP and the right FAT. Following the same line of reasoning
developed earlier, one scenario that could lead to this pattern of results suggests that tightly
packed axons, which are linked to better performance, also restrict the overall diffusivity in a
voxel, thus causing a reduction in the local MD values in association with improved
performance.
In sum, the directions of the associations we found are internally consistent, and are
compatible with some explanations in terms of tissue properties, and not with others. These
explanations may be further tested with advanced quantitative MRI measures that are sensitive to
the different biological factors comprising the tissue (Assaf et al., 2008; Assaf & Basser, 2005;
Berman et al., 2019; Filo et al., 2019; Mezer & Yeatman, 2014).
Limitations
The current study takes a tract-based approach, in which we test a small number of pre-
defined hypotheses using individualized tractography methods in native space. This approach
limits the number of hypotheses tested, as adding more tracts to the analysis would increase the
chances of false positive results. Here, we focused on the bilateral CPs and FAT that were
previously associated with different aspects of speech fluency in clinical populations (Jossinger
27
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
et al., 2021; Kronfeld-Duenias et al., 2016). However, we do not rule out the possibility that
other white matter tracts may also contribute to various aspects of speech production.
Specifically, the basal ganglia were previously shown to be implicated in speech fluency, in both
clinical and neurotypical populations (Booth et al., 2007; Chang & Guenther, 2020; Zenon &
Olivier, 2014). Future studies will be needed to test whether basal ganglia connections underpin
speech fluency in neurotypical speakers.
A second limitation concerns the location of the correlations along the right SCP and right
MCP, which were found in the vicinity of the decussation. At the point of decussation, voxels are
occupied by both right and left tracts, making it difficult to attribute the findings to either the left
or the right tract. Considering that voxels in the decussation of the CPs contain two fiber
orientations (as shown by Jeurissen et al., 2013), such voxels likely contain primarily the two
crossing CPs (e.g., left and right SCP), uncontaminated by other tracts. Nonetheless, crossing
fibers are known to affect (typically reduce) FA values (Fig. 1c-d). The FA drop near the
decussation is not sufficient, however, to generate a significant correlation, as the significant
results with FA at the vicinity of decussation were specific to the right SCP and were not shown
in other pathways that contain decussating fibers. The effect of crossing fibers on MD in our data
is less dramatic: MD profiles (particularly in the MCP and ICP, see Fig. S4c-f) are generally
flatter compared to FA profiles (Fig. 1c-d). Therefore, the fact that we find a significant
correlation with MD in the right MCP suggests that this effect is not driven only by crossing
fibers.
Lastly, the current study, as any other study that identifies correlations in a sample of adults,
cannot be conclusive about the direction of causality that underlies the effects. Future
developmental and longitudinal studies will help in determining whether the variability in the
microstructure of the CPs and the FAT derives the variability in speech fluency seen in
adulthood, or perhaps the differences in microstructure arise as a result of other cognitive and
motor tasks involved in speech fluency.
Conclusions
In conclusion, our findings support the involvement of the cerebellum in aspects of speech
production that go beyond articulatory control, such as lexical access, pragmatic or syntactic
generation. Using complex modeling and probabilistic tracking enabled us to follow the
trajectory of the cerebro-cerebellar pathways as they decussate, and to detect novel associations
28
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
with speech fluency in these pathways. By evaluating multiple measures of speech fluency, our
study makes an important contribution to the understanding of the neural basis of speech
production in neurotypical adults.
REFERENCES
Ackermann, H., Vogel, M., Petersen, D., & Poremba, M. (1992). Speech deficits in ischaemic
cerebellar lesions. Journal of Neurology, 239, 223–227.
Akers, D. (2006). CINCH. Proceedings of the 19th Annual ACM Symposium on User Interface
Software and Technology - UIST ’06, 33.
Ambrose, N. G., & Yairi, E. (1999). Normative disfluency data for early childhood stuttering.
Journal of Speech, Language, and Hearing Research : JSLHR, 42(4), 895–909.
Amir, O. (2016). Speaking Rate among Adult Hebrew Speakers: A Preliminary Observation.
Annals of Behavioural Science, 2(1), 1–9.
Amir, O., & Grinfeld, D. (2011). Articulation rate in childhood and adolescence: Hebrew
speakers. Language and Speech, 54(2), 225–240.
Amunts, J., Camilleri, J. A., Eickhoff, S. B., Heim, S., & Weis, S. (2020). Executive functions
predict verbal fluency scores in healthy participants. Scientific Reports, 10(1), 1–11.
Andrade, C. R. F., Maluf Cervone, L., & Chiarion Sassi, F. (2003). Relationship between the
stuttering severity index and speech rate. Sao Paulo Medical Journal, 121(2), 81–84.
Argyropoulos, G. P. D. (2016). The cerebellum, internal models and prediction in ‘non-motor’
aspects of language: A critical review. Brain and Language, 161, 4–17.
Assaf, Y., & Basser, P. J. (2005). Composite hindered and restricted model of diffusion
(CHARMED) MR imaging of the human brain. NeuroImage, 27(1), 48–58.
Assaf, Y., Blumenfeld-Katzir, T., Yovel, Y., & Basser, P. J. (2008). AxCaliber: A method for
measuring axon diameter distribution from diffusion MRI. Magnetic Resonance in
Medicine, 59(6), 1347–1354.
Assaf, Y., & Pasternak, O. (2008). Diffusion tensor imaging (DTI)-based white matter mapping
in brain research: A review. Journal of Molecular Neuroscience, 34(1), 51–61.
Barazany, D., Basser, P. J., & Assaf, Y. (2009). In vivo measurement of axon diameter
29
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
distribution in the corpus callosum of rat brain. Brain, 132(5), 1210–1220.
Basser, P. J., & Pierpaoli, C. (1996). Microstructural and Physiological Features of Tissues
Elucidated by Quantitative-Diffusion-Tensor MRI. Journal of Magnetic Resonance, Series
B, 111(3), 209–219.
Beaulieu, C. (2002). The basis of anisotropic water diffusion in the nervous system - a technical
review. NMR in Biomedicine, 15(7–8), 435–455.
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery rate: A practical and
powerful approach to multiple testing. Journal of the Royal Statistical Society Series B
(Methodological), 57(1), 289–300.
Berman, S., Filo, S., & Mezer, A. A. (2019). Modeling conduction delays in the corpus callosum
using MRI-measured g-ratio. NeuroImage, 195(November 2018), 128–139.
Blecher, T., Miron, S., Schneider, G. G., Achiron, A., & Ben-Shachar, M. (2019). Association
between white matter microstructure and verbal fluency in patients with multiple sclerosis.
Frontiers in Psychology, 10(JULY).
Booth, J. R., Wood, L., Lu, D., Houk, J. C., & Bitan, T. (2007). The role of the basal ganglia and
cerebellum in language processing. Brain Research, 1133(1), 136–144.
Bose, A., Patra, A., Antoniou, G. E., Stickland, R. C., & Belke, E. (2022). Verbal fluency
difficulties in aphasia: A combination of lexical and executive control deficits. International
Journal of Language & Communication Disorders.
Bruckert, L., Shpanskaya, K., McKenna, E. S., Borchers, L. R., Yablonski, M., Blecher, T., Ben-
Shachar, M., Travis, K. E., Feldman, H. M., & Yeom, K. W. (2019). Age-Dependent White
Matter Characteristics of the Cerebellar Peduncles from Infancy Through Adolescence. The
Cerebellum, 18(3), 372–387.
Bruckert, L., Travis, K. E., Mezer, A. A., Ben-Shachar, M., & Feldman, H. M. (2020).
Associations of Reading Efficiency with White Matter Properties of the Cerebellar
Peduncles in Children. Cerebellum, 19(6), 771–777.
Castellazzi, G., Bruno, S. D., Toosy, A. T., Casiraghi, L., Palesi, F., Savini, G., D’Angelo, E., &
Wheeler-Kingshott, C. A. M. G. (2018). Prominent changes in cerebro-cerebellar functional
connectivity during continuous cognitive processing. Frontiers in Cellular Neuroscience,
12(October), 1–15.
30
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Catani, M., Dell’Acqua, F., Vergani, F., Malik, F., Hodge, H., Roy, P., Valabregue, R., &
Thiebaut de Schotten, M. (2012). Short frontal lobe connections of the human brain. Cortex,
48(2), 273–291.
Catani, M., Mesulam, M. M., Jakobsen, E., Malik, F., Martersteck, A., Wieneke, C., Thompson,
C. K., Thiebaut De Schotten, M., Dell’Acqua, F., Weintraub, S., & Rogalski, E. (2013). A
novel frontal pathway underlies verbal fluency in primary progressive aphasia. Brain,
136(8), 2619–2628.
Chang, L. C., Jones, D. K., & Pierpaoli, C. (2005). RESTORE: Robust estimation of tensors by
outlier rejection. Magnetic Resonance in Medicine, 53(5), 1088–1095.
Chang, S. E., & Guenther, F. H. (2020). Involvement of the Cortico-Basal Ganglia-
Thalamocortical Loop in Developmental Stuttering. Frontiers in Psychology, 10(January).
Connally, E. L., Ward, D., Howell, P., & Watkins, K. E. (2014). Disrupted white matter in
language and motor tracts in developmental stuttering. Brain and Language, 131, 25–35.
Costello, J. M., & Ingham, R. (1984). Assessment strategies for stuttering. In R. F. Curlee & W.
H. Perkins (Eds.), Nature and treatment of stuttering: New directions (pp. 303–333).
College Hill Press.
De Santis, S., Assaf, Y., Evans, C. J., & Jones, D. K. (2014). Improved precision in CHARMED
assessment of white matter through sampling scheme optimization and model parsimony
testing. Magnetic Resonance in Medicine, 71(2), 661–671.
Fiez, J. A., Petersen, S., Cheney, M. K., & Raichle, M. E. (1992). Imparied non-motor learning
and error detection associated with cerebellar damage. Brain, 115(1), 155–178.
Filo, S., Shtangel, O., Salamon, N., Kol, A., Weisinger, B., Shifman, S., & Mezer, A. A. (2019).
Disentangling molecular alterations from water-content changes in the aging human brain
using quantitative MRI. Nature Communications, 10(1), 3403.
Fonov, V., Evans, A. C., Botteron, K., Almli, C. R., McKinstry, R. C., & Collins, D. L. (2011).
Unbiased average age-appropriate atlases for pediatric studies. NeuroImage, 54(1), 313–
327.
Friedman, L., Kenny, J. T., Wise, A. L., Wu, D., Stuve, T. A., Miller, D. A., Jesberger, J. A., &
Lewin, J. S. (1998). Brain Activation During Silent Word Generation Evaluated with
Functional MRI. Brain and Language, 64(2), 231–256.
31
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Fujii, M., Maesawa, S., Motomura, K., Futamura, M., Hayashi, Y., Koba, I., & Wakabayashi, T.
(2015). Intraoperative subcortical mapping of a language-associated deep frontal tract
connecting the superior frontal gyrus to Broca’s area in the dominant hemisphere of patients
with glioma. Journal of Neurosurgery, 122(6), 1390–1396.
Guenther, F. H. (2006). Cortical interactions underlying the production of speech sounds.
Journal of Communication Disorders, 39(5), 350–365.
Gurd, J. M., Amunts, K., Weiss, P. H., Zafiris, O., Zilles, K., Marshall, J. C., & Fink, G. R.
(2002). Posterior parietal cortex is implicated in continuous switching between verbal
fluency tasks: an fMRI study with clinical implications. Brain : A Journal of Neurology,
125(Pt 5), 1024–1038.
Halari, R., Sharma, T., Hines, M., Andrew, C., Simmons, A., & Kumari, V. (2006). Comparable
fMRI activity with differential behavioural performance on mental rotation and overt verbal
fluency tasks in healthy men and women. Experimental Brain Research, 169(1), 1–14.
Henry, J. D., & Crawford, J. R. (2004). Verbal fluency deficits in Parkinson’s disease: A meta-
analysis. Journal of the International Neuropsychological Society, 10(4), 608–622.
Hickok, G. (2012). Computatinal neuroanatomy of speech production. Nature Reviews
Neuroscience., 13(2), 135-145.
Hickok, G., Houde, J., & Rong, F. (2011). Sensorimotor Integration in Speech Processing:
Computational Basis and Neural Organization. Neuron, 69(3), 407–422.
Horowitz, A., Barazany, D., Tavor, I., Bernstein, M., Yovel, G., & Assaf, Y. (2015). In vivo
correlation between axon diameter and conduction velocity in the human brain. Brain
Structure and Function, 220(3), 1777–1788.
Howell, P., Au-Yeung, J., & Pilgrim, L. (1999). Utterance rate and linguistic properties as
determinants of lexical dysfluencies in children who stutter. Journal of the Acoustical
Society of America, 105, 481–490.
Hubrich-Ungureanu, P., Kaemmerer, N., Henn, F. A., & Braus, D. F. (2002). Lateralized
organization of the cerebellum in a silent verbal fluency task: A functional magnetic
resonance imaging study in healthy volunteers. Neuroscience Letters, 319(2), 91–94.
Ito, M. (2008). Control of mental activities by internal models in the cerebellum. Nature Reviews
Neuroscience, 9(4), 304–313.
32
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Jeurissen, B., Leemans, A., Tournier, J. D., Jones, D. K., & Sijbers, J. (2013). Investigating the
prevalence of complex fiber configurations in white matter tissue with diffusion magnetic
resonance imaging. Human Brain Mapping, 34(11), 2747–2766.
Jeurissen, B., Tournier, J.-D., Dhollander, T., Connelly, A., & Sijbers, J. (2014). Multi-tissue
constrained spherical deconvolution for improved analysis of multi-shell diffusion MRI
data. NeuroImage, 103, 411–426.
Johnson, C. A., Liu, Y., Waller, N., & Chang, S. E. (2022). Tract profiles of the cerebellar
peduncles in children who stutter. Brain Structure and Function, 0123456789.
Jones, D. K., Knösche, T. R., & Turner, R. (2013). White matter integrity, fiber count, and other
fallacies: The do’s and don’ts of diffusion MRI. NeuroImage, 73, 239–254.
Jossinger, S., Kronfeld-Duenias, V., Zislis, A., Amir, O., & Ben-Shachar, M. (2021). Speech rate
association with cerebellar white-matter diffusivity in adults with persistent developmental
stuttering. Brain Structure and Function, 226(3), 801–816.
Jossinger, S., Mawase, F., Ben-Shachar, M., & Shmuelof, L. (2020). Locomotor Adaptation Is
Associated with Microstructural Properties of the Inferior Cerebellar Peduncle. The
Cerebellum, 19(3), 370–382.
Jossinger, S., Sares, A., Zislis, A., Sury, D., Gracco, V., & Ben-Shachar, M. (2022). White
matter correlates of sensorimotor synchronization in persistent developmental stuttering.
Journal of Communication Disorders, 95(November 2021), 106169.
Kavé, G. (2006). The development of naming and word fluency: evidence from Hebrew-
speaking children between ages 8 and 17. Developmental Neuropsychology, 29(3), 493–
508.
Kavé, G., & Knafo-Noam, A. (2015). Lifespan development of phonemic and semantic fluency:
Universal increase, differential decrease. Journal of Clinical and Experimental
Neuropsychology, 37(7), 751–763.
Kawato, M. (1999). Internal models for motor control and trajectory planning. Current Opinion
in Neurobiology, 9(6), 718–727.
Kelly, R. M., & Strick, P. L. (2003). Cerebellar loops with motor cortex and prefrontal cortex of
a nonhuman primate. The Journal of Neuroscience : The Official Journal of the Society for
Neuroscience, 23(23), 8432–8444.
33
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Kent, R. D., Kent, J. F., & Rosenbek, J. C. (1987). Maximum Performance Tests of Speech
Production. Journal of Speech and Hearing Disorders, 52(4), 367–387.
Kent, R. D., & Rosenbek, J. C. (1983). Acoustic Patterns of Apraxia of Speech. Journal of
Speech, Language, and Hearing Research, 26(2), 231–249.
Kinoshita, M., de Champfleur, N. M., Deverdun, J., Moritz-Gasser, S., Herbet, G., & Duffau, H.
(2015). Role of fronto-striatal tract and frontal aslant tract in movement and speech: an
axonal mapping study. Brain Structure and Function, 220(6), 3399–3412.
Kramer, J. H., Mungas, D., Possin, K. L., Rankin, K. P., Boxer, A. L., Rosen, H. J., Bostrom, A.,
Sinha, L., Berhel, A., & Widmeyer, M. (2014). NIH EXAMINER: Conceptualization and
Development of an Executive Function Battery. Journal of the International
Neuropsychological Society, 20(1), 11–19.
Kronfeld-Duenias, V., Amir, O., Ezrati-Vinacour, R., Civier, O., & Ben-Shachar, M. (2016). The
frontal aslant tract underlies speech fluency in persistent developmental stuttering. Brain
Structure and Function, 221(1), 365–381.
Kruper, J., Yeatman, J. D., Richie-Halford, A., Bloom, D., Grotheer, M., Caffarra, S., Kiar, G.,
Karipidis, I. I., Roy, E., Chandio, B. Q., Garyfallidis, E., & Rokem, A. (2021). Evaluating
the Reliability of Human Brain White Matter Tractometry. Aperture Neuro, 2021(1), 1–26.
Leemans, A., & Jones, D. K. (2009). The B -matrix must be rotated when correcting for subject
motion in DTI data. Magnetic Resonance in Medicine, 61(6), 1336–1349.
Leiner, H. C. (2010). Solving the mystery of the human cerebellum. Neuropsychology Review,
20(3), 229–235.
Leiner, H. C., Leiner, A. L., & Dow, R. S. (1986). Does the cerebellum contribute to mental
skills? Behavioral Neuroscience, 100(4), 443–454.
Li, M., Zhang, Y., Song, L., Huang, R., Ding, J., Fang, Y., Xu, Y., & Han, Z. (2017). Structural
connectivity subserving verbal fluency revealed by lesion-behavior mapping in stroke
patients. Neuropsychologia, 101(January), 85–96.
Libon, D. J., McMillan, C., Gunawardena, D., Powers, C., Massimo, L., Khan, A., Morgan, B.,
Farag, C., Richmond, L., Weinstein, J., Moore, P., Coslett, H. B., Chatterjee, A., Aguirre,
G., & Grossman, M. (2009). Neurocognitive contributions to verbal fluency deficits in
frontotemporal lobar degeneration. Neurology, 73(7), 535–542.
34
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Liewald, D., Miller, R., Logothetis, N., Wagner, H. J., & Schüz, A. (2014). Distribution of axon
diameters in cortical white matter: an electron-microscopic study on three human brains and
a macaque. Biological Cybernetics, 108(5), 541–557.
Marien, P., Engelborghs, S., Fabbro, F., & De Deyn, P. P. (2001). The lateralized linguistic
cerebellum: A review and a new hypothesis. Brain and Language, 79(3), 580–600.
Max, L., Guenther, F., & Gracco, V. (2004). Unstable or insufficiently activated internal models
and feedback-biased motor control as sources of dysfluency: A theoretical model of
stuttering. Contemporary Issues in Communication Science and Disorders, 31, 105–122.
Mehl, M. R., Vazire, S., Ramírez-Esparza, N., Slatcher, R. B., & Pennebaker, J. W. (2007). Are
women really more talkative than men? Science, 317(5834), 82.
Mezer, A., & Yeatman, J. D. (2014). Quantifying the local tissue volume and composition in
individual brains with MRI. Pediatric Nephrology, 29(7), 1231–1238.
Middleton, F. a, & Strick, P. L. (1994). Anatomical evidence for cerebellar and basal ganglia
involvement in higher cognitive function. Science, 266(5184), 458–461.
Nichols, T. E., & Holmes, A. P. (2002). Nonparametric permutation tests for functional
neuroimaging: A primer with examples. Human Brain Mapping, 15(1), 1–25.
Oishi, K., Faria, A., Jiang, H., Li, X., Akhter, K., Zhang, J., Hsu, J. T., Miller, M. I., van Zijl, P.
C. M., Albert, M., Lyketsos, C. G., Woods, R., Toga, A. W., Pike, G. B., Rosa-Neto, P.,
Evans, A., Mazziotta, J., & Mori, S. (2009). Atlas-based whole brain white matter analysis
using large deformation diffeomorphic metric mapping: Application to normal elderly and
Alzheimer’s disease participants. NeuroImage, 46(2), 486–499.
Oldfield, R. C. (1971). The assessment and analysis of handedness: The Edinburgh inventory.
Neuropsychologia, 9(1), 97–113.
Palesi, F., De Rinaldis, A., Castellazzi, G., Calamante, F., Muhlert, N., Chard, D., Tournier, J.
D., Magenes, G., D’Angelo, E., & Wheeler-Kingshott, C. A. M. G. (2017). Contralateral
cortico-ponto-cerebellar pathways reconstruction in humans in vivo: Implications for
reciprocal cerebro-cerebellar structural connectivity in motor and non-motor areas.
Scientific Reports, 7(1), 1–13.
Palesi, F., Tournier, J. D., Calamante, F., Muhlert, N., Castellazzi, G., Chard, D., D’Angelo, E.,
& Wheeler-Kingshott, C. A. M. (2015). Contralateral cerebello-thalamo-cortical pathways
35
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
with prominent involvement of associative areas in humans in vivo. Brain Structure and
Function, 220(6), 3369–3384.
Perrini, P., Tiezzi, G., Castagna, M., & Vannozzi, R. (2013). Three-dimensional microsurgical
anatomy of cerebellar peduncles. Neurosurgical Review, 36(2), 215–225.
Peterburs, J., Bellebaum, C., Koch, B., Schwarz, M., & Daum, I. (2010). Working memory and
verbal fluency deficits following cerebellar lesions: Relation to interindividual differences
in patient variables. Cerebellum, 9(3), 375–383.
Petersen, S. E., Fox, P. T., Posner, M. I., Mintun, M., & Raichle, M. E. (1988). Positron emission
tomographic studies of the cortical anatomy of single-word processing. Nature, 331(6157),
585–589.
Petersen, S. E., Fox, P. T., Posner, M. I., Mintun, M., & Raichle, M. E. (1989). Positron
Emission Tomographic Studies of the Processing of Singe Words. Journal of Cognitive
Neuroscience, 1(2), 153–170.
R Core Team. (2013). R: A language and environment for statistical computing. R Foundation
for Statistical Computing, Vienna, Austria.
Riecker, A., Kassubek, J., Gröschel, K., Grodd, W., & Ackermann, H. (2006). The cerebral
control of speech tempo: Opposite relationship between speaking rate and BOLD signal
changes at striatal and cerebellar structures. NeuroImage, 29(1), 46–53.
Riecker, A., Mathiak, K., Wildgruber, D., Erb, M., Hertrich, I., Grodd, W., & Ackermann, H.
(2005). fMRI reveals two distinct cerebral networks subserving speech motor control.
Neurology, 64(4), 700–706.
Rochman, D., & Amir, O. (2013). Examining in-session expressions of emotions with
speech/vocal acoustic measures: An introductory guide. Psychotherapy Research, 23(4),
381–393.
Rodero, E. (2012). A comparative analysis of speech rate and perception in radio bulletins. Text
and Talk, 32(3), 391–411.
Rohde, G. K., Barnett, A. S., Basser, P. J., Marenco, S., & Pierpaoli, C. (2004). Comprehensive
approach for correction of motion and distortion in diffusion-weighted MRI. Magnetic
Resonance in Medicine, 51(1), 103–114.
Schlösser, R., Hutchinson, M., Joseffer, S., Rusinek, H., Saarimaki, A., Stevenson, J., Dewey, S.
36
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
L., & Brodie, J. D. (1998). Functional magnetic resonance imaging of human brain activity
in a verbal fluency task. Journal of Neurology, Neurosurgery & Psychiatry , 64(4), 492–
498.
Schmahmann, J. D. (2010). The role of the cerebellum in cognition and emotion: Personal
reflections since 1982 on the dysmetria of thought hypothesis, and its historical evolution
from theory to therapy. Neuropsychology Review, 20(3), 236–260.
Shadmehr, R. (2017). Learning to Predict and Control the Physics of Our Movements. The
Journal of Neuroscience, 37(7), 1663–1671.
Shadmehr, R., & Krakauer, J. W. (2008). A computational neuroanatomy for motor control.
Experimental Brain Research, 185(3), 359–381.
Shao, Z., Janse, E., Visser, K., & Meyer, A. S. (2014). What do verbal fluency tasks measure?
Predictors of verbal fluency performance in older adults. Frontiers in Psychology, 5(JUL),
1–10.
Silveri, M. C., Leggio, M. G., & Molinari, M. (1994). The cerebellum contributes to linguistic
production. Neurology, 44(11), 2047–2047.
Sobczak-Edmans, M., Lo, Y. C., Hsu, Y. C., Chen, Y. J., Kwok, F. Y., Chuang, K. H., Tseng, W.
Y. I., & Chen, S. H. A. (2019). Cerebro-Cerebellar Pathways for Verbal Working Memory.
Frontiers in Human Neuroscience, 12(January), 1–13.
Sturm, J., & Seery, C. H. (2007). Speech and articulatory rates of school-age children in
conversation and narrative contexts. Language, Speech, and Hearing Services in Schools,
38(1), 47–59.
Tournier, J. D., Calamante, F., & Connelly, A. (2007). Robust determination of the fibre
orientation distribution in diffusion MRI: Non-negativity constrained super-resolved
spherical deconvolution. NeuroImage, 35(4), 1459–1472.
Tournier, J. D., Calamante, F., Gadian, D. G., & Connelly, A. (2004). Direct estimation of the
fiber orientation density function from diffusion-weighted MRI data using spherical
deconvolution. NeuroImage, 23(3), 1176–1185.
Tournier, J. D., Smith, R., Raffelt, D., Tabbara, R., Dhollander, T., Pietsch, M., Christiaens, D.,
Jeurissen, B., Yeh, C.-H., & Connelly, A. (2019). MRtrix3: A fast, flexible and open
software framework for medical image processing and visualisation. NeuroImage, 202,
37
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
/
.
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
116137.
Tourville, J. A., & Guenther, F. H. (2011). The DIVA model: A neural theory of speech
acquisition and production. Language and Cognitive Processes, 26(7), 952–981.
Travis, K. E., Leitner, Y., Feldman, H. M., & Ben-Shachar, M. (2015). Cerebellar white matter
pathways are associated with reading skills in children and adolescents. Human Brain
Mapping, 36(4), 1536–1553.
Tuch, D. S., Reese, T. G., Wiegell, M. R., & Wedeen, V. J. (2003). Diffusion MRI of Complex
Neural Architecture. Neuron, 40(5), 885–895.
Uddin, M. N., Figley, T. D., Solar, K. G., Shatil, A. S., & Figley, C. R. (2019). Comparisons
between multi-component myelin water fraction, T1w/T2w ratio, and diffusion tensor
imaging measures in healthy human brain structures. Scientific Reports, 9(1), 2500.
Vassal, F., Boutet, C., Lemaire, J.-J., & Nuti, C. (2014). New insights into the functional
significance of the frontal aslant tract: An anatomo–functional study using intraoperative
electrical stimulations combined with diffusion tensor imaging-based fiber tracking. British
Journal of Neurosurgery, 28(5), 685–687.
Walker, J. F., Archibald, L. M. D., Cherniak, S. R., & Fish, V. G. (1992). Articulation Rate in 3-
and 5-Year-Old Children. Journal of Speech, Language, and Hearing Research, 35(1), 4–
13.
Wells, W., Viola, P., Atsumi, H., Nakajima, S., & Kikinis, R. (1996). Multi-modal volume
registration by maximization of mutual information. Medical Image Analysis, 1(1), 35–51.
Wolpert, D. M., Miall, R. C., & Kawato, M. (1998). Internal models in the cerebellum. Trends in
Cognitive Sciences, 2(9), 338–347.
Xie, J., Cai, T. T., Maris, J., & Li, H. (2011). Optimal false discovery rate control for dependent
data. Statistics and Its Interface, 4(4), 417–430.
Yablonski, M., & Ben-Shachar, M. (2020). Sensitivity to word structure in adult Hebrew readers
is associated with microstructure of the ventral reading pathways. Cortex, 128, 234–253.
Yablonski, M., Menashe, B., & Ben-Shachar, M. (2021). A general role for ventral white matter
pathways in morphological processing: Going beyond reading. NeuroImage, 226(June
2020), 117577.
Yeatman, J. D., Dougherty, R. F., Myall, N. J., Wandell, B. A., & Feldman, H. M. (2012). Tract
38
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
.
/
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Profiles of White Matter Properties: Automating Fiber-Tract Quantification. PLoS ONE,
7(11), e49790.
Yeatman, J. D., Dougherty, R. F., Rykhlevskaia, E., Sherbondy, A. J., Deutsch, G. K., Wandell,
B. A., & Ben-Shachar, M. (2011). Anatomical Properties of the Arcuate Fasciculus Predict
Phonological and Reading Skills in Children. Journal of Cognitive Neuroscience, 23(11),
3304–3317.
Zenon, A., & Olivier, E. (2014). Contribution of the basal ganglia to spoken language: Is speech
production like the other motor skills? Behavioral and Brain Sciences, 37(6), 576–576.
Zettin, M., Cappa, S. F., D’amico, A., Rago, R., Perino, C., Perani, D., & Fazio, F. (1997).
Agrammatic speech production after a right cerebellar haemorrhage. Neurocase, 3(5), 375–
380.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
d
o
i
/
l
/
.
/
1
0
1
1
6
2
n
o
_
a
_
0
0
0
9
8
2
0
6
6
8
8
6
n
o
_
a
_
0
0
0
9
8
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
39