Is Morality Unified? Evidence that Distinct Neural
Systems Underlie Moral Judgments of Harm,
Dishonesty, and Disgust
Carolyn Parkinson1, Walter Sinnott-Armstrong 2, Philipp E. Koralus3,
Angela Mendelovici 4, Victoria McGeer 3, and Thalia Wheatley 1
D
o
w
N
l
o
UN
D
e
D
Astratto
■ Much recent research has sought to uncover the neural basis
of moral judgment. Tuttavia, it has remained unclear whether
“moral judgments” are sufficiently homogenous to be studied
scientifically as a unified category. We tested this assumption
by using f MRI to examine the neural correlates of moral judg-
ments within three moral areas: (physical) harm, dishonesty,
E (sexual) disgust. We found that the judgment of moral wrong-
ness was subserved by distinct neural systems for each of the dif-
ferent moral areas and that these differences were much more
robust than differences in wrongness judgments within a moral
area. Dishonest, disgusting, and harmful moral transgression re-
cruited networks of brain regions associated with mentalizing, af-
fective processing, and action understanding, rispettivamente. Dorsal
medial pFC was the only region activated by all scenarios judged to
be morally wrong in comparison with neutral scenarios. Tuttavia,
this region was also activated by dishonest and harmful scenarios
judged not to be morally wrong, suggestive of a domain-general
role that is neither peculiar to nor predictive of moral decisions.
These results suggest that moral judgment is not a wholly unified
faculty in the human brain, but rather, instantiated in dissociable
neural systems that are engaged differentially depending on the
type of transgression being judged. ■
INTRODUCTION
Is it worse to cheat on an exam or to eat your dog? Contro-
sideration of these acts feels very different, yet we tend to
classify a diverse variety of acts (per esempio., theft, lying, assault,
disrespect, incest) as “morally wrong.” This common lin-
guistic label does not ensure that moral judgments of such
diverse acts are similar enough to be studied as a single
kind of judgment. Nonetheless, research investigating
moral judgments has tended to treat moral judgments per-
taining to vastly different contents as comparable or even
equivalent.
Many theorists suggest that there is a universal, innate
human moral faculty (Hauser, 2006; Dwyer, 1999; Mikhail,
Sorrentino, & Spelke, 1998; Rawls, 1971). Such theories
posit that our moral judgments are fueled neither by emo-
tion nor by conscious deliberation but by an unconscious
evaluative mechanism that makes judgments based on in-
nate, often inarticulable principles analogous to Chomskyʼs
universal grammar. Others have emphasized the causal role
of emotions in moral judgments (Wheatley & Haidt, 2005;
Haidt, 2001). Still others have suggested that emotional
responses and cognitive computations work in tandem
to give rise to moral judgments (Dupoux & Jacob, 2007;
Greene, Nystrom, Engell, Darley, & Cohen, 2004). The ex-
istence of any such shared, uniform mechanism—be it ra-
1Dartmouth College, 2Duke University, 3Princeton University,
4Rotman Institute of Philosophy, University of Western Ontario
tional, affect-driven, or both—would predict shared neural
responses across moral domains.
Some philosophers (Prinz, 2008), psychologists (Blair,
Marsh, Finger, Blair, & Luo, 2006; Moll, Zahn, de Oliveira-
Souza, Krueger, & Grafman, 2005; Haidt, 2001), and an-
thropologists (Shweder, Much, Mahapatra, & Park, 1997)
have questioned the assumption that morality is homoge-
nous enough to be treated as a unified faculty. Behavioral
data indicates that some moral judgments are rooted in
conscious reasoning whereas others are not (Cushman,
Young, & Hauser, 2006) and that psychopathy scores cor-
relate with moral judgments in some areas more than
others (Glenn, Iyer, Graham, Koleva, & Haidt, 2009). These
findings have underscored the need to adopt a more fine-
grained approach to studying morality.
Such an approach may be especially fruitful in identify-
ing the neural underpinnings of moral judgment. There is
evidence that the neural correlates of harm judgments dif-
fer depending on whether the harms are direct or indirect
(Greene et al., 2004; Greene, Sommerville, Nystrom, Darley,
& Cohen, 2001), as well as whether they are intended or
unintended (Young & Saxe, 2008; Borg, Hynes, Van Horn,
Grafton, & Sinnott-Armstrong, 2006). There is also some
evidence that different brain areas are recruited by the
judgment of scenarios involving bodily harm compared
with those that do not (Heekeren et al., 2005). Independent
studies of dishonesty (Greene & Paxton, 2009) and disgust
(Borg, Lieberman, & Kiehl, 2008; Moll, de Oliveira-Souza,
© 2011 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 23:10, pag. 3162–3180
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
2
C
3
l
1
e
0
–
3
P
1
D
6
F
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
C
2
N
_
/
UN
1
_
7
0
7
0
6
0
6
1
9
7
1
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
0
o
0
N
1
0
7
8
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
.
T
/
o
N
1
8
M
UN
sì
2
0
2
1
et al., 2005) report distinct patterns of brain activations as-
sociated with judgments of moral transgressions within
these broad categories, suggesting that these judgments
are distinct from one another and from harm judgments.
Inoltre, different morally relevant emotions such as
guilt, anger, and embarrassment are associated with disso-
ciable networks of brain regions (Immorino-Yang, McColl,
Damasio, & Damasio, 2009; Zahn et al., 2009; Kedia, Berthoz,
Wessa, Hilton, & Martinot, 2008; Moll et al., 2007; Takahashi
et al., 2004; Moll, de Oliveira-Souza, & Eslinger, 2003;
Berthoz, Armony, Blair, & Dolan, 2002; Dougherty et al.,
1999). Across these studies, activation of the dorsal medial
pFC (DMPFC), an area implicated in self-referential process-
ing (Northoff et al., 2006) and thinking about others (Amodio
& Frith, 2006), especially in ambiguous circumstances
( Jenkins & Mitchell, 2010), is most widely associated with
morally relevant processing (Moll, Zahn, et al., 2005). Im-
portantly, although independent studies have been con-
ducted within multiple moral areas (per esempio., harm, disgust,
and dishonesty), methodological differences between
studies render direct comparisons difficult.
Another obstacle to understanding the neural basis of
moral judgment is the widespread tendency to contrast
moral scenarios to each other and to neutral scenarios
without considering whether participants judge acts in
those scenarios to be morally wrong. Notably, one study
(Greene et al., 2004) found significant activation differ-
ences between those who judged the same act to be mor-
ally permissible or impermissible within the category of
physical harm. It remains to be seen whether these differ-
ences extend to other moral domains. Accordingly, we
sought to determine whether any neural substrate is com-
mon to judgments of moral transgressions (as compared
with neutral scenarios) within or across various domains,
or with judgments that an act is morally wrong (as com-
pared with not morally wrong) within or across domains.
It was predicted that judgments of different categories of
moral transgressions would be related to different patterns
of activity in the brain.
More specifically, it was predicted that disgusting moral
transgressions would evoke activity in areas associated with
affective processing, such as the amygdala, OFC, and insula
( Jabbi, Bastiaansen, & Keysers, 2008; Britton et al., 2005),
whereas dishonest transgressions would elicit activity in
areas activated when inferring othersʼ mental states, come
as the TPJ (Saxe & Kanwisher, 2003), the posterior cingulate
cortex (PCC), and the DMPFC (Rilling, Sanfey, Aronson,
Nystrom, & Cohen, 2004). In line with the bulk of fMRI stud-
ies of moral reasoning that have focused on harm (per esempio.,
Greene et al., 2001), it was predicted that the dorsolateral
pFC (DLPFC), ACC, and the aforementioned mentalizing
areas, as well as areas implicated in action understanding
(posterior STS, inferior parietal lobule [IPL], SMA; Grafton,
Arbib, Fadiga, & Rizzolatti, 1996) E, more specifically, In
processing socially informative sensory information (poste-
rior STS; Allison, Puce, & McCarthy, 2000) would show activity
in response to moral transgressions involving bodily harms.
The neural correlates of “wrong” and “not wrong” judg-
ments within and across story types were compared. As
activation differences between “wrong” and “not wrong”
judgments have only been found within the category of
harm (Greene et al., 2004) and other studies (Borg et al.,
2006) have failed to find significant differences, no specific
hypotheses were made about the neural correlates of
these judgments.
METHODS
Stimuli
Three rounds of pilot testing involving 75 participants at
Princeton University and Dartmouth College yielded 14 ac-
ceptable scenarios in each of four categories: Disgusting,
Harmful, and Dishonest moral scenarios, and Neutral (mor-
ally irrelevant) scenarios. At the start of pilot testing, partic-
ipants were told that they would read multiple scenarios
and make several ratings of each one. Participants were in-
structed to accept the details as specified in the scenario.
Per esempio, if a person is described as teasing another
persona, the participant should accept teasing as an accurate
description of the behavior. Participants were also in-
structed that if they fill in details, those details should be
what the participant takes to be most likely (per esempio., if the story
says that Jim asked Matt a question, the participant should
assume that Jim was speaking loudly enough for Matt to
hear him). Then, participants were given a sample scenario
and the following descriptions of the ratings they were to
make for each scenario:
Is the act morally wrong? This question is not about
whether the act is or should be illegal. Nor is it about
whether you would do the act or advise friends to do
Esso. The question is simply about whether that particular
act is morally wrong in the specified circumstances. In
the above example, the question is not whether it is
wrong in all circumstances to tell somebody to shut
up but whether it is wrong in the particular scenario
for Mike to tell John to shut up.
Is the act harmful? This question is not about whether
harm would result if this kind of act became common
or if other people found out about it. The question is
only about whether this act causes any harm in the par-
ticular circumstances as specified.
Is the act dishonest? This question is about whether the
act depends on intentional deception, promise break-
ing, cheating, theft, or something like that. It is not
about whether everyone in the scenario knows exactly
what is going on.
Is the act disgusting? This question is not about whether it
is morally horrendous. È, instead, about whether the act
is disgusting in the way that most people find it disgusting
to lick toilets or to have oral sex with their grandparents.
After reading the instructions, the pilot participants made
the dichotomous (yes or no) judgments listed above for
Parkinson et al.
3163
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
2
C
3
l
1
e
0
–
3
P
1
D
6
F
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
C
2
N
_
/
UN
1
_
7
0
7
0
6
0
6
1
9
7
1
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
0
o
0
N
1
0
7
8
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
/
T
.
o
N
1
8
M
UN
sì
2
0
2
1
each scenario. Scenarios were considered acceptable for
use in the fMRI study if they adhered to the following cri-
teria: A Neutral scenario was deemed acceptable if 80% O
more of the pilot participants judged the associated act to
be morally “not wrong” and no more than 20% of pilot par-
ticipants deemed the act to be disgusting, harmful or dis-
honest. The moral scenarios (Disgusting, Harmful and
Dishonest scenarios) were deemed acceptable if at least
80% of the pilot participants judged the associated act as
representative of its intended moral domain (per esempio., Disgust-
ing) and no more than 20% of pilot participants deemed
the act to be representative of the other two moral do-
mains (per esempio., Harmful, Dishonest). These criteria helped
ensure that the transgression scenarios were representa-
tive of one and only one moral domain. Inoltre, IL
moral scenarios were acceptable only if no less than 30%
and no more than 70% of pilot participants considered
them to be morally “wrong.” This inherent ambiguity af-
forded sufficient response variability to contrast “wrong”
and “not wrong” responses within and across moral cate-
sanguinose. The scenarios selected for the fMRI study did not
significantly differ in length between categories (mean
lengths for all four categories ranged from 44 A 46 parole;
F(3, 55) = 1.95, ns). For further analyses of the contents
of scenarios in different categories, refer to Table A3,
Table A4, and Figure A1 of the Appendix.
For the f MRI study, each scenario was converted into
scrolling black text on a light blue background using Live-
Type software (Apple, Inc.). This ensured that participants
read the same words at approximately the same time. Each
scenario scrolled for 22 sec, and the full text remained on
the screen for 2 sec after scrolling had ceased.
f MRI Procedure
Thirty-eight naive, right-handed adults were scanned in a
3.0-T Phillips scanner. All participants were right-handed
English speakers with normal or corrected-to-normal vi-
sion. Before scanning, the experimenter obtained informed
consent and explained the task to the participant. Specifi-
cally, participants were told that they would read scrolling
text scenarios describing the actions of a person. After each
scenario, they were prompted to press a button indicating
whether they considered the main characterʼs actions to be
“wrong” or “not wrong,” and then to press a second button
indicating the confidence level of their judgment (“High”
or “Low”). See Figure 1 for a schematic representation of
a single trial.
Between reading and judging scenarios, participants
viewed a fixation cross for varying amounts of time and
pressed a button whenever the cross changed color. IL
order of the stimuli and the intervening fixation periods
was optimized for hemodynamic deconvolution by the
AFNI program RSFgen (Cox, 1996).
Post-imaging Procedure
After leaving the scanner, participants were provided with
a written copy of all of the scenarios they had viewed while
being scanned, as well as definitions of the terms “dis-
gusting,” “harmful,” and “dishonest” (the same definitions
used in pilot testing). After reading these definitions,
participants were asked if the actions described in each
scenario could be characterized as disgusting, harmful,
and/or dishonest. Inoltre, participants rated if each
Figura 1. Schematic
representation of a single trial.
Participants viewed a fixation
cross between trials for a varied
interstimulus interval of 0–12 sec.
Each trial began with a scenario,
presented via text scrolling at a
fixed rate for 22 sec. After the
scenario had stopped scrolling,
participants were asked to judge
whether the act in that scenario
was “wrong” or “not wrong” by
pressing one of two buttons.
Dopo 2 sec, subjects were then
asked to indicate whether they
had made that judgment with
high or low confidence using a
button press.
3164
Journal of Cognitive Neuroscience
Volume 23, Numero 10
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
2
C
3
l
1
e
0
–
3
P
1
D
6
F
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
C
2
N
_
/
UN
1
_
7
0
7
0
6
0
6
1
9
7
1
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
0
o
0
N
1
0
7
8
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
F
T
.
o
N
1
8
M
UN
sì
2
0
2
1
scenario evoked emotion using a Likert scale from 1
(not at all ) A 4 (intensely).
Imaging Parameters
The study was performed using a Philips Intera Achieva
3-T scanner (Philips Medical Systems, Bothell, WA) con un
SENSE (SENSEitivity Encoding) head coil. All stimuli were
presented to the participant via a Panasonic DT-4000U DLP
projector. Anatomical images were collected using a high-
resolution 3-D Magnetization-prepared rapid gradient-echo
sequence (124 slices, 1.2 mm thick, FOV = 240 mm, acqui-
sition matrix = 256 × 256). Functional images were col-
lected in seven runs using echo-planar functional images
sensitive to BOLD contrast (TR = 2000 msec, TE = 30 msec,
flip angle = 90°, 3 mm in-plane resolution). During each of
the seven functional runs, 180 sets of axial images (36 slices/
volume, 3.5 mm slice thickness with 0.5 mm skip) were col-
lected in an interleaved fashion across the whole brain. Axial
images were not angulated. See Figures A2 and A3 in the
Appendix for echo-planer magnetic resonance images of a
single participant to illustrate brain coverage.
Imaging Analysis
fMRI Preprocessing
Functional and anatomical images were analyzed with
AFNI (Cox, 1996). Transient spikes in the signal were sup-
pressed with the AFNI program 3dDespike. As the slices of
each volume were not acquired simultaneously, a timing
correction procedure was used. All volumes were motion
corrected, normalized to the standardized space of Talairach
and Tournoux (1988), and smoothed with a 6-mm FWHM
smoothing kernel before conversion to percent signal
change from the mean.
fMRI Analysis
A regression analysis using 3dREMLfit in AFNI was con-
ducted on each participantʼs preprocessed EPI volumes
to estimate the individual hemodynamic response to each
of the five stimulus conditions. Stick functions indicating
the onset times of each stimulus were convolved with a
30-sec block function. Second-order trends and the partic-
ipantʼs head motion were included as regressors at this
stage to account for signal changes due to scanner drift
and motion artifact. The convolved hemodynamic re-
sponse for each condition for each participant was sub-
mitted to a series of t tests conducted with 3dMEMA
(Chen, Saad, & Cox, 2010), an AFNI program that adopts
a mixed-effects meta-analysis approach and takes into ac-
count both within- and between-participants variability,
making it relatively robust to outliers.
Unless otherwise noted, all reported clusters survived
a statistical threshold of p < .05, corrected for multiple
comparisons as stipulated by AlphaSim within AFNI
(afni.nimh.nih.gov/pub/dist/doc/manual/AlphaSim.pdf ).
AlphaSim carries out Monte Carlo simulations by randomly
generating uncorrelated three-dimensional images using
the overall dimensions and voxel geometry of a data set,
followed by convolution with a Gaussian function to ac-
count for correlations between nearby voxels. Using the
parameters of our data set (FWHM = 6 mm and a brain-
only mask), we ran 1000 iterations over a skull-stripped
whole brain mask, which indicated that a voxel-wise
threshold of p < .005 and a cluster size threshold of
675 mm3 (25 × 27 mm3 voxels) would provide a family-
wise false positive rate of .05.
RESULTS
Evidence for Separate Systems
Behavioral Data
Consistent with the possibility that different categories of
moral judgment are subserved by different cognitive sys-
tems, postscanner questionnaire results indicated that
participants classified scenarios into only the appropriate
categories. Disgusting scenarios were rated as disgusting
but neither dishonest nor harmful, χ2(5, n = 1560) =
1113.07, p < .0001. Dishonest scenarios were rated as dis-
honest but neither disgusting nor harmful, χ2(5, n =
1560) = 1369.45, p < .0001, and Harmful scenarios were
rated as harmful but neither dishonest nor disgusting
χ2(5, n = 1560) = 1302.26, p < .0001. Neutral scenarios
were rated as neither disgusting nor harmful nor dishonest
χ2(5, n = 1560) = 1508.47, p < .0001.
Table A2 in the Appendix contains the number of
“wrong” and “not wrong” decisions for all moral and neutral
scenarios, as well as the variance of participantsʼ responses
for each scenario and for each category. A one-way ANOVA
of the three moral categories (Harm, Dishonesty, Disgust)
with response variance across participants for each scenario
as the dependent variable was performed. As Leveneʼs test
of equality of variances indicated that the assumption of
homogeneity of variance was violated (F(2, 39) = 3.56,
p < .05), the Brown–Forsythe F ratio is reported. There
were differences between moral categories with respect
to response variability, F(2, 31.42) = 6.84, p < .01. The
Games–Howell post hoc test was used to detect differences
between categories because it does not assume equal vari-
ances (Games & Howell, 1976). Disgust scenarios elicited
less variable responses than both Harm and Dishonesty
scenarios (both pʼs < .05), whereas Dishonesty and Harm
scenarios did not differ in response variability ( p = .81).
Semantic Content of Scenarios
Whereas syntactic complexity was kept consistent across
categories (see Table A4), semantic content varied by de-
sign. In the interest of ecological validity, scenarios were
constructed to resemble as closely as possible how they
might be described colloquially. As such, scenarios in each
category differed with respect to their characteristic semantic
Parkinson et al.
3165
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
t
f
o
n
1
8
M
a
y
2
0
2
1
content. In addition, Disgust scenarios were naturally more
emotionally evocative than Neutral, Dishonest, or Harmful
scenarios. These between-category differences and within-
category similarities ensured that scenarios were classified
easily into their respective moral domains and only their
respective moral domains. See Figure A1 and Table A3 in
the Appendix for analyses of emotional evocativeness and
semantic content, respectively. Also refer to the Appendix
for a full list of scenarios used in the fMRI study.
f MRI Data
To identify the neural correlates of processing different
types of moral transgressions, we conducted two sets of
analyses. In the first set of analyses, we compared brain ac-
tivity associated with the judgment of each type of moral
transgression to that of morally neutral scenarios, as well
as to transgressions in the remaining two moral categories.
A moral transgression was defined as an act in a moral sce-
nario that was judged to be morally wrong. Previous be-
havioral results indicate that individuals differ in which acts
they classify as morally wrong versus merely convention-
ally inappropriate (Haidt & Hersh, 2001). Further, there
exist significant differences in brain activity between partic-
ipants who judge the same physical harms to be either
morally permissible or impermissible (Greene et al., 2004).
Limiting the first set of analyses to moral scenarios deemed
wrong by the participants, therefore, afforded cross-subject
comparisons most relevant to participantsʼ judgments of
moral wrongness. Please refer to Table A1 for similar anal-
yses including brain activity associated with all moral sto-
ries as compared with neutral, irrespective of the associated
decision (“wrong”/“not wrong”). Importantly, because all
conditions, both moral and neutral, involved moral rea-
soning, the contrasts outlined in Table A1 do not reveal
the neural correlates of moral reasoning. Rather, these dif-
ferences in the hemodynamic response reflect differences
associated with the processing of moral scenarios relative to
neutral scenarios, irrespective of the ensuing moral judg-
ment. The following comparisons contrast the hemody-
namic responses associated with moral transgressions
compared with neutral scenarios. “Moral transgressions”
entered into the analysis for each participant include only
those scenarios that he or she deemed to be morally
“wrong.” Our primary question was whether different moral
transgressions would evoke hemodynamic responses con-
sistent with a unified or a disunified (separate systems) ac-
count of moral judgment.
Harmful transgressions. The judgment of transgres-
sions involving physical harms compared with neutral
scenarios was associated with increased activity in the left
DLPFC, a region of DMPFC including the ACC, the SMA,
the IPL, and the posterior STS (see Table 1). Similarly, com-
pared with judgments of both Disgusting and Dishonest
transgressions, judgment of Harmful moral transgressions
was associated with increased activity in an area of the
Table 1. Talairach Coordinates for the Centers of Mass of
Significant Clusters for Each Moral Transgression versus Neutral
Scenario Comparison
Area
Hemi
BA
vxls
x
y
z
Harmful transgressions
DMPFC
DMPFC
SMA
DLPFC
DLPFC
STS
IPL
Cerebellum
L
R
L
R
L
L
L
R
Dishonest transgressions
9/32
107
6/9/10
71
−4
6
6
139 −13
8/9
72
29
39
41
17
39
32
9
39
40
58 −30
38 −45 −57
40 −55 −44
28
41 −55 −35
DMPFC
DMPFC
DLPFC
TPJ
TPJ
PCC
L
R
L
L
R
L
6/8/9
583 −14
6/8
185
14
8
119 −40
32
22
20
39/40
39/40
31
306 −46 −56
96
53
56 −56
−4 −50
Disgusting transgressions
DLPFC
DMPFC
DMPFC
IFG
IFG/FO
TP
TP
Fusiform gyrus
Amygdala
Amygdala
ACC
PCC
Thalamus
Cerebellum
All transgressions
DMPFC
L
L
R
L
L
R
L
L
L
R
L
L
R
R
p < .05, corrected.
IFG = inferior frontal gyrus.
9/10
203 −21
8/9/10
226
9/10
88
−4
7
41
51
55
47
70 −23
15 −15
46/47
222 −42
125
32
34
9
16 −22
38
36
37
24/32
7/31
129 −26
−4 −29
91 −42 −43 −16
259 −20
−4 −12
−2 −13
19
−4
31
−3 −44
−2 −16
239
137
159
150
88
38 −58 −30
9
28
2
50
29
28
30
54
35
37
13
31
44
56
42
36
35
30
35
27
23
21
27
5
3166
Journal of Cognitive Neuroscience
Volume 23, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
/
.
o
n
1
8
M
a
y
2
0
2
1
Table 2. Talairach Coordinates for the Centers of Mass of
Significant Clusters for Direct Comparisons Between Moral
Transgression Types
Area
Harm
Hemi
BA
vxls x
y
z
Harmful transgressions > Dishonest transgressions
Tavolo 2. (continued )
Area
SgACC
Parahippocampal gyrusb
Parahippocampal gyrusb
SMA
Midcingulate
IPL
IPL
Precuneus
Precuneus
l
l
l
R
R
l
6
31
40
40
7
7
33 −17
5
25 −12 −24
94 −56 −32
39
37
60 −34
9 −63
27 −7 −50
Harmful transgressions > Disgusting transgressions
DLPFC
IPL
Precuneus
Precuneus
Dishonesty
R
R
l
R
6/8/9
270
30
24
40
7
7
36
60 −40
68 −7 −53
26
7 −52
54
39
31
27
43
55
44
37
47
46
TP
Fusiform gyrus
Fusiform gyrus
Lingual gyrus
Lingual gyrus
Precuneus
PCC
PCC
Amygdala
Amygdala
FO/Insula
FOa
Dishonest transgressions > Harmful transgressions
Parahippocampal gyrusb
VMPFC
DLPFC
TPJ
TPJ
MTG
MTG
PCC
l
l
l
R
l
R
l
10/32
113 −6
49 −6
Parahippocampal gyrusb
8
39/40
39/40
21
21
33 −14
31
213 −42 −63
70
52 −60
51
38
35
Fusiform gyrus
Lingual gyrus
Lingual gyrus
166 −55 −10 −15
IPL
97
63 −32 −11
Precuneus
23/31
41 −4 −55
18
Hemi
BA
vxls x
sì
z
R
l
R
R
R
l
R
l
l
l
R
l
R
34/25
38/28
38
8
6 −11
197 −25
22 −25
38/28
124
27
39
14 −23
20 −25
60 −43 −13
35
70
111 −51 −45 −12
42
14 −85 −14
36 −14 −85 −13
38
20/37
37/20
18
18
7/19
109 −31 −63
23/30/31 227 −4 −43
23/31
81
5 −36
42
25
25
132 −22 −1 −15
96
22 −1 −15
l
R
l
R
l
l
R
l
R
l
R
13
66 −41
13/47/34 46
17
28/36/38 135 −25
28/38
111
28
4 −6
2 −10
2 −28
8 −26
37
18
18
40
7
85 −43 −43 −16
155 −9 −87 −6
46
11 −83 −5
31 −48 −27
84
11 −66
33
35
151 −23 −2 −14
139
22 −2 −14
Disgusting transgressions > Dishonest transgressions
Dishonest transgressions > Disgusting transgressions
DLPFC
DLPFC
OFC
SMA
TPJ
TPJ
Hippocampus
Disgust
R
l
R
l
R
l
l
8/9
8/9
155
34
43 −42
23
20
44
43
10/11
40
35
50 −8
6
25 −14
25
39/40
212
53 −53
39/40
155 −49 −55
41 −36 −37
54
35
30
3
Disgusting transgressions > Harmful transgressions
VMPFC
VMPFC
OFC
FO/IFGa
l
R
l
l
10/24/32
215 −4
47
0
10/24
47/11
109
7
45 −1
52 −25
34 −10
13/47/34/25 46 −17
7 −11
Amygdala
Amygdala
P < .05, corrected.
aActivation extends to sgACC.
bActivation extends to TP.
VMPFC = ventromedial prefrontal cortex; MTG = middle temporal cortex.
DLPFC that included the SMA as well as the IPL and the
anterior aspect of the precuneus (see Table 2).
Dishonest transgressions. As predicted, the judgment of
Dishonest transgressions compared with neutral scenarios
was correlated bilaterally with increased activity in the
DMPFC, the TPJ extending superiorly into the IPL and the
PCC, as well as increased activity in the left DLPFC (Table 1).
Dishonest scenarios elicited increased activity in the left
DLPFC and bilaterally in the TPJ compared with both Dis-
gusting and Harmful transgressions (Table 2).
Disgusting transgressions. The Disgusting versus Neutral
scenario contrast revealed increased activity bilaterally in the
Parkinson et al.
3167
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
.
t
o
n
1
8
M
a
y
2
0
2
1
DMPFC, amygdalae, ACC, and PCC, as well as the right tem-
poral pole (TP), left DLPFC, and a region in the left inferior
frontal cortex encompassing the frontal operculum (FO) and
the anterior aspect of the insula (see Table 1). Compared
with both Dishonest and Harm scenarios, Disgust scenarios
elicited increased activity bilaterally in the amygdalae, TPs,
and lingual gyri, in the left FO/anterior insula and fusiform
gyri, as well as the right subgenual ACC (sgACC) and pos-
terior portion of the precuneus (see Table 2).
System Overlap
To explore whether there was any activation common to
all transgression types relative to neutral scenarios, we
performed a conjunction analysis. One 28-voxel cluster
in the DMPFC was independently activated in the compar-
ison of each moral transgression type compared with neu-
tral scenarios (surviving a threshold of p < .05, corrected,
for all comparisons). To investigate whether this region
was activated specifically by moral transgressions, this
cluster was then applied as a mask to each individualʼs data
to extract the average hemodynamic response for the “not
wrong” scenarios for each category type. This secondary
ROI analysis revealed that Dishonest and Harmful sce-
narios judged to be “not wrong” also activated this region
more strongly than Neutral scenarios ( p < .005; p < .05,
respectively). See Figure 3.
Also, judgment confidence (high vs. low) was not a signif-
icant predictor of brain activity within any system, with the
exception of Disgust. Within the network of areas involved
in the judgment of disgusting transgressions, confidence
level modulated activity in the PCC. Greater judgment
confidence was associated with greater activity in this area
(Talairach coordinate of center of mass: −1, −48, 26;
30 voxels).
DISCUSSION
Separate Systems
Overview
The behavioral and imaging results reported here suggest
that multiple distinct cognitive systems support moral judg-
ment (see Figure 2). These systems are consistent with
multiple intercategory differences in semantic content
and the associated processing demands of that content.
Judgments of transgressions involving physical harm acti-
vated areas commonly associated with action understand-
ing. Judgments of dishonest transgressions evoked activity
in areas associated with representing other peopleʼs be-
liefs. Judgments of disgusting transgressions evoked ac-
tivity in areas associated with affective processing.
Harm
Secondary Analyses
No significant activation differences between “wrong” and
“not wrong” judgments within moral categories emerged.
Transgressions involving physical harms recruited areas
associated with understanding and imagining actions
(Grafton et al., 1996): the IPL, the STS, and the SMA. Addi-
tionally, the DLPFC and ACC were more activated by the
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
/
.
f
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Separable moral systems. Brain regions showing increased activity when judging moral scenarios to be “wrong” as compared to judging
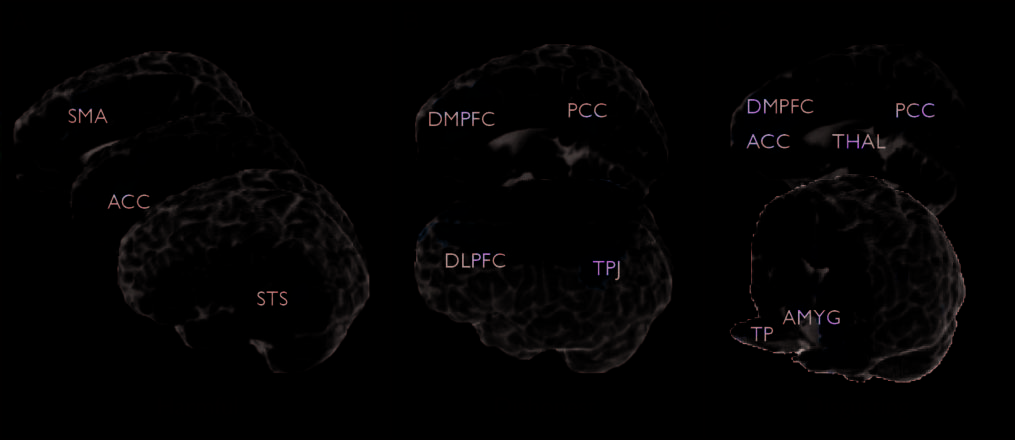
neutral scenarios to be “not wrong.” (A) The judgment of physically harmful scenarios was associated with increased activity in the dorsal ACC, SMA,
and STS. (B) The judgment of dishonest transgressions was associated with increased activity in the DMPFC and TPJ bilaterally, as well as the
left DLPFC and PCC. (C) The judgment of disgusting moral transgressions elicited increased activity in the temporal poles and amygdalae, as well
as in the DMPFC, ACC, and PCC. For significant activations not pictured, refer to Table 1. For display purposes, these group data are projected onto
a reference anatomical image (N27 AFNI brain).
3168
Journal of Cognitive Neuroscience
Volume 23, Number 10
judgment of harms than for that of neutral scenarios,
consistent with previous studies implicating these areas
in moral reasoning about physical harms (Greene et al.,
2001). Further, compared with Dishonest and Disgust-
ing transgressions, the judgment of transgressions in-
volving physical harms elicited greater activity in areas
associated with action understanding, such as the SMA
and IPL, as well as a region of the precuneus implicated
in self-centered visuospatial imagery strategies (Cavanna
& Trimble, 2005). These results suggest that in compari-
son with other types of moral transgressions, understand-
ing moral transgressions involving physical harms relies
more on processing and imagining the physical actions
described in these scenarios.
Dishonesty
The judgment of Dishonest transgressions was correlated
with increased bilateral activity in a region encompassing
the TPJ and IPL that is implicated in reasoning about mental
(Saxe & Kanwisher, 2003; Fletcher et al., 1995) and nonmen-
tal (Apperly, Samson, Chiavarino, Bickerton, & Humphreys,
2007) representations, simultaneously holding on-line multi-
ple possible perspectives (Decety & Jackson, 2004), and at-
tentional reorienting more generally (Corbetta, Patel, &
Shulman, 2008; Astafiev, Shulman, & Corbetta, 2006). Dis-
honest transgressions also preferentially activated the left
PCC and DMPFC, areas robustly associated with inferring
othersʼ mental states (Rilling et al., 2004). Similarly, compared
with judging other moral transgressions, judging Dishonest
transgressions was associated with DLPFC and TPJ activity
(Table 2). These findings suggest that understanding dishon-
est transgressions requires mentalizing, or, more generally, a
shifting of attention between deceitful actorsʼ conflicting
words and actions. Interestingly, however, Dishonest and
Harmful scenarios did not significantly differ from each other
in the frequency of belief or communication verbs (see
Table A3 in the Appendix). This suggests that the increased
bilateral activity in the TPJ for Dishonest, as compared with
Harmful, transgressions (see Table 2) does not reflect the
processing of mental state words per se but how the words,
in combination, evoke different cognitive operations such as
mentalizing. Alternatively, the bilateral activation of TPJ for
Dishonest scenarios may reflect other cognitive operations
associated with this region such as the discrimination of self
and other (Cheng, Chen, Lin, Chou, & Decety, 2010; Decety
& Sommerville, 2003). For example, the Dishonest scenar-
ios (e.g., academic dishonesty) may have hit particularly
“close to home” with our college-aged participants, thereby
evoking a need to distance oneself from the protagonist.
The PCC is also associated with social–emotional pro-
cessing (Britton et al., 2005) and the evaluation of emo-
tionally evocative stimuli (Maddock, 1999). Therefore, its
activation in the Dishonest versus Neutral contrast could
also reflect the greater emotional arousal and evaluative
processing elicited by Dishonest transgressions relative
to Neutral scenarios. The activations observed in this con-
trast could reflect a greater need for deliberation when
judging Dishonest, as compared with Neutral, scenarios.
Disgust
Disgusting transgressions were associated with increased
bilateral activity in the amygdalae, DMPFC, and PCC, areas
implicated in social–emotional processing (Britton et al.,
2005) as well as in the right TP, implicated in binding vis-
ceral affective responses to complex stimuli (Olson, Plotzker,
& Ezzyat, 2007), in understanding social norm violations
(Mendez, Anderson, & Shapira, 2005), and in social concep-
tual understanding more generally (Zahn et al., 2006). This
comparison also revealed increased activity in the anterior
portion of the left insula extending into the FO, a region as-
sociated with the experience of disgust as well as disgusting
mental imagery ( Jabbi et al., 2008). Results of this contrast
suggest that the judgment of disgusting moral transgressions
relative to neutral scenarios involves affective processing as
well as social evaluation. This result is consistent with the
higher number of emotion words contained in Disgust sce-
narios (see Table A3), as well as the higher ratings of elicited
emotion for these scenarios (see Figure A1).
Compared with Dishonest and Harmful moral trans-
gressions, judgments of Disgusting moral transgressions
were correlated with bilateral activity in areas also identi-
fied in the Disgust versus Neutral contrast that are in-
volved in general emotional processing (amygdalae, TP),
as well as emotional processing specific to disgust (left
FO/insula). These contrasts also revealed activity in the
sgACC, an area implicated in emotional arousal and inter-
oception (Critchley, 2004). Both contrasts also revealed in-
creased activity in a region of the precuneus that has been
implicated in the attribution of emotions to the self and
others (Ochsner et al., 2004), possibly indicating that emo-
tional empathy was preferentially engaged by scenarios
involving sexual disgust. The bilateral lingual and parahip-
pocampal gyri activations observed in both of these con-
trasts have previously been associated with processing
erotic stimuli (Hu et al., 2008). Finally, the activation of
the fusiform gyrus by Disgust scenarios could reflect in-
creased imageability for disgusting transgressions relative
to other transgression types, as activity in the fusiform gyrus
has been consistently associated with mental imagery (e.g.,
Ishai, Ungerleider, & Haxby, 2000). Together, these activa-
tion differences highlight the many inherent characteristics
of content and processing demands that differentiate moral
transgressions that involve sexual disgust from other types
of moral transgressions.
The differences in semantic content between moral cat-
egories almost certainly yielded concomitant differences
in cognitive operations engaged by those contents. Such
semantic and processing differences are expected in sce-
narios designed to be as natural and emblematic of their
respective categories as possible. Indeed, the moral cate-
gories used here most likely differ in a number of ways left
Parkinson et al.
3169
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
.
t
o
n
1
8
M
a
y
2
0
2
1
unspecified (e.g., imageability). These many differences
underline the fact that moral transgressions are intrinsi-
cally diverse and differ systematically for several reasons,
only some of which can be quantified by analyses of sce-
nario contents, behavioral ratings by participants, and asso-
ciated neural activity. Taken together, the diversity of content
and concomitant cognitive and emotional processes coupled
with the differences in brain activity between categories of
moral transgressions further illustrate that “moral” judgment
is best investigated as a superordinate category of judgments
that share “family resemblances” (Rosch & Mervis, 1975)
rather than any defining feature or set of features.
In addition to differences between scenarios that were
consistent across participants, in at least some cases, the
cognitive processes elicited by the same moral scenarios
likely differed somewhat between participants. For in-
stance, interesting work by Kedia et al. (2008) and Moll
et al. (2007) indicates that agency perspective affects
the neural correlates of moral processing. Although all
scenarios were written in the third person, it cannot be
ascertained whether individual participants took the per-
spective of the victim or transgressor while reading. Simi-
larly, although many studies (Immorino-Yang et al., 2009;
Zahn et al., 2009; Kedia et al., 2008; Moll et al., 2003,
2007; Takahashi et al., 2004; Berthoz et al., 2002; Dougherty
et al., 1999) indicate that experiencing different moral
emotions is associated with neural activation differences,
we cannot ascertain what specific emotion(s) partici-
pants were experiencing while reading each scenario,
which likely differed between participants for many rea-
sons, including the agency perspective that participants
took while reading a scenario. For instance in Dishonesty
scenarios, different participants could have experienced
indignation or guilt, depending on the agency perspective
they assumed while reading. Similarly, for Harm scenarios,
different participants could have differentially experienced
compassion for the victim and/or contempt for the aggressor
to varying degrees. For these reasons, characterizing the
moral emotions evoked by different transgression types is
beyond the scope of the present study. This endeavor and,
more generally, the detailed characterization of the many dif-
ferences that exist between categories of moral transgres-
sions remains an important direction for future research.
System Overlap
A region in the DMPFC was independently activated by the
contrast of each moral transgression type compared with
Neutral scenarios. This region was also activated by Dis-
honest and Harmful scenarios judged to be “not wrong”
relative to Neutral scenarios. Together, these findings sug-
gest that the activity observed in this region reflects evalua-
tive processes engaged by the moral scenarios that are
not peculiar to a particular moral decision (“wrong” or “not
wrong”). This interpretation is consistent with at least
two lines of research investigating the characteristics of
this region.
First, activity in this region is known to be modulated by
ambiguity both in social ( Jenkins & Mitchell, 2010) and
nonsocial (Volz, Schubotz, & von Cramon, 2003) contexts.
Our moral scenarios were ambiguous by design to allow
for comparisons of “wrong” and “not wrong” judgments
of the same scenarios. This ambiguity was not present in
the neutral scenarios and is reflected in the high variability
of responses to moral scenarios as compared with neutral
scenarios (see Table A2 in the Appendix). Thus, the pro-
cessing of ambiguous information represents one explana-
tion for the shared DMPFC activation observed in the
moral versus neutral comparisons.
Second, this area is consistently activated when thinking
about other people (Van Overwalle, 2009; Amodio & Frith,
2006; Gallagher & Frith, 2003; Goel, Grafman, Sadato, &
Hallett, 1995) and in self-referential processing (Northoff
et al., 2006). It is likely that the evaluation of moral trans-
gressions, as compared with Neutral scenarios, involves a
deeper evaluation of individualsʼ motives and actions and
how they relate to oneʼs own values. Of particular rele-
vance to the current study, a recent meta-analysis found
this region to be involved not only in moral judgment
tasks, but also in mentalizing and in understanding
goal-directed actions ( Van Overwalle & Baetens, 2009).
This explanation is not mutually exclusive with that of
ambiguity resolution. Indeed, the observed activity in
this region is consistent with its known role in evaluat-
ing other people in ambiguous situations ( Jenkins &
Mitchell, 2010).
Because participants were asked to employ moral rea-
soning (i.e., to answer the question, “Was it morally
wrong?”) on all trials, both moral and neutral, characteriz-
ing the neural underpinnings of moral reasoning, as com-
pared with other types of reasoning, is beyond the scope
of the current study. Rather, this study was primarily con-
cerned with characterizing the processing of what partici-
pants deemed to be moral transgressions. Nevertheless,
because moral reasoning was common to moral and neu-
tral trials, the finding that the DMPFC was activated by the
conjunction of all moral transgression types relative to
neutral scenarios (see Figure 3) and by the conjunction of
all moral story types relative to neutral stories (see Table A1
in the Appendix) suggests that some property of the moral
scenarios relative to the neutral scenarios was driving this
activation—possibly their inherent ambiguity.
Regardless of the exact mechanisms involved, the pres-
ent results suggest that the overlapping activation in
DMPFC reflects aspects of processing moral transgres-
sions that are not peculiar to the decision of moral wrong-
ness. This suggests that this region is not the seat of a
unified moral faculty in the strongest sense—that is, a fac-
ulty that is dedicated to judgments of moral transgressions
in particular. The present results are consistent with a more
general role of this region in the evaluation of human behav-
ior, of which moral action is a subcategory. Thus, lesions or
disruptions to this region would likely result in deficits or dif-
ferences in making moral judgments due to the disruption
3170
Journal of Cognitive Neuroscience
Volume 23, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
/
t
o
n
1
8
M
a
y
2
0
2
1
methods (e.g., theory of mind; Saxe, 2009), no network
seems to be common to the judgment of a diverse variety
of moral transgressions. Separate systems were found to
characterize different kinds of moral judgment. The only
region of anatomical overlap between these systems
(DMPFC) is robustly associated more generally with
self-referential processing, thinking about other people
and processing ambiguous information.
It is likely that moral judgment is even more multi-
dimensional than what is suggested here, given that
there remain other domains of morality that were not
examined in the current study (e.g., disrespect, betrayal
of an in-group, fairness). These results suggest that, just
as disparate systems are now understood to subserve
aspects of cognitive faculties once thought to be mono-
lithic (e.g., memory, attention), distinct systems subserve
different types of moral judgment. Future research may
benefit from working toward a taxonomy of these sys-
tems as Haidt and Graham (2007) have suggested. Such
a taxonomic approach should help reduce the risks of
false negatives and overgeneralization. The risk of false
negatives is present whenever diverse stimuli are com-
bined within a single experimental condition. Lumping
together diverse moral transgressions risks failing to detect
the component processes activated by one type of trans-
gression that are not also activated by other types of trans-
gressions. Similarly, caution should be taken in generalizing
neural and cognitive processes across moral domains. Con-
flation of different types of moral judgment for analysis
likely contributes to inconsistency in the current literature
regarding, for instance, whether emotion causes (Haidt,
2001) or emerges as a mere consequence of (Huebner,
Dwyer, & Hauser, 2009) moral judgment and whether the
principles that guide moral judgments are consistent ac-
ross demographic groups (Hauser, Cushman, Young, Jin,
& Mikhail, 2007) or vary as a function of socioeconomic
status and political affiliation (Haidt & Graham, 2007).
The current results are consistent with the hypothesis
that morality is not a natural kind in the mind but, rather,
is comprised of multiple dissociable mental processes.
That said, there could exist a cognitive process common
and peculiar to moral judgments that is not detectable
using the current methods. The hemodynamic differ-
ences observed here appear to be modulated primarily
by differences in the semantic content of moral scenarios,
and the general cognitive processes they engage, rather
than their perceived moral wrongness. Irrespective of
whether subsequent studies find evidence for a unified
moral faculty, future research using f MRI and standard
paradigms will likely benefit from taking an increasingly
fine-grained approach—from isolating distinct categories
of moral behavior for study, and from investigating the
specific component processes that contribute to different
types of moral judgments. In so doing, we will be most likely
to generate consistent and precise findings across studies,
and to gain a more accurate and thorough understanding
of the manifold judgments, we term “moral.”
Parkinson et al.
3171
Figure 3. System overlap. One 28-voxel cluster in the DMPFC was
independently activated by the judgment of each moral transgression
type (Harmful, Disgusting, Dishonest) relative to the judgment of
neutral scenarios.
of evaluative processes (e.g., ambiguity resolution, men-
talizing) that are not specific to the domain of morality.
Wrong versus Not Wrong Judgments
Although we had hoped to compare “wrong” and “not
wrong” judgments within categories, we lacked sufficient
power to do so. Although our sample was relatively large
(n = 38), subtle activation differences between “wrong”
and “not wrong” judgments may have been diluted by
the length of the written scenarios used, which limited
the number of exemplars per condition and precluded
isolating the specific time at which an individual reached
a moral judgment for each scenario. Future studies em-
ploying designs with greater temporal constraints may
find more significant differences between “wrong” and
“not wrong” moral judgments. However, these differ-
ences appear to be subtle in comparison with differences
between moral transgression types.
Differences between aspects of the content of the sce-
narios (such as whether they depict physical violence, de-
ceit, or sexual acts that elicit disgust) were much more
robust. This suggests that previously observed patterns of
activity that characterize moral judgment may reflect, at
least in part, processing of characteristics of these scenarios
independent of whether acts are judged morally wrong.
Conclusion
These results provide empirical support for philosophical
arguments against the existence of a functional or ana-
tomical module common and peculiar to all moral judg-
ments (Prinz, 2008; Sinott-Armstrong, 2008). Although
consistent networks of brain regions have been reliably
implicated in some dimensions of cognition using diverse
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
1
8
M
a
y
2
0
2
1
APPENDIX 1: SCENARIOS BY CATEGORY
Harmful Scenarios
1. Francis is writing a very important paper that is due tomorrow. Alex talks to him repeatedly and plays music loudly to distract
Francis, even after Francis asks him to leave the room. Finally, in frustration, Francis punches Alex hard enough to bruise him badly.
2. Peter sees a car coming straight at him. He knows the driver is trying to kill him. His only escape is to run behind a crowd, so the
car will hit them instead of him. He runs behind the crowd, and the car hits three people before it stops.
3. Beth has two young sons who often play very rudely, cursing at each other. After more than 3 hr of trying to control them, Beth
finally hits them on their hands with a wooden ruler hard enough to make their hands bruise.
4. Elaine and five other passengers are up in a hot air balloon with a big hole, so it is falling rapidly. The only way to prevent a life-
threatening crash is to throw a passenger overboard. Elaine pushes out the heaviest passenger, who hits the ground hard and
breaks his legs.
5. Susan owns a small camera store and lives in an apartment above it. One evening she hears something in the store. When she
goes downstairs, she sees someone stealing her cameras. She pulls her gun and shoots the thief in the back before he can leave.
6. After their ship sinks, Elmer and 12 other passengers are in an overcrowded lifeboat that is about to sink. The only way to keep it
afloat is to force two passengers overboard. After drawing lots Elmer pushes out two passengers, who soon drown.
7. Jen is walking upstairs to class with her friend Tom who is teasing her relentlessly. Jen asks Tom to stop, but he keeps teasing her
and playfully pushes her. Jen pushes him back hard, which knocks him down the stairs. Tom breaks his arm in the fall.
8. Ferdinand lives alone in the country. Late one night, he hears some noises in his living room. He runs downstairs with his gun
and sees an intruder wearing a ski mask across the room. Without warning, he shoots and kills the intruder.
9. Allen sees an assassin in a crowd of people aiming at him. To protect himself, he shoots at the assassin with both barrels of a
double barreled shotgun that he just brought back from a hunting trip. He hits the assassin and a bystander in the crowd.
10. Laura is in a library with her young daughter, Mandy, who is being disruptive. Laura tries to reason with Mandy and warns her several
times, but Mandy keeps making noise. When a librarian tells them to leave, Laura forcefully drags Mandy out by the arm, hurting her.
11. Mike has a few beers at a party. His friend, Alice, tells him that heʼs too drunk to drive. Mike still insists on driving home, so Alice
hits him to make him drop his car keys. Mike falls and bangs his head, causing a concussion.
12. Matthew and Nick get into a heated argument. Nick suggests they settle it by a fistfight. Matthew is reluctant, but Nick insists, so
Matthew finally agrees. Matthew is much bigger and stronger, so he quickly wins by punching Nick in the side, breaking two ribs.
13. Kim and Olivia have an argument. Olivia suggests that they resolve their dispute by wrestling, although she knows Kim is on a
wrestling team. Kim agrees and quickly wins with a special move she learned from her wrestling coach. In the process, she badly
dislocates Oliviaʼs shoulder.
14. Greg is driving when he sees another car out of control heading directly toward an unsuspecting pedestrian. The pedestrian
canʼt get out of the way, so Greg slams his car into the other car, saving the pedestrian, but paralyzing the driver of the other car.
Dishonest Scenarios
1. While paying for groceries, Andrew forgets to remove an expensive package of meat from the bottom of his cart. The store clerk
does not notice. Discovering his mistake in the parking lot, Andrew decides that he will not go back to pay for the meat.
2. Helen borrows her boyfriendʼs car to go shopping. She accidentally bumps into a pole and scrapes the side of the car. She knows her
boyfriend will be furious if he finds out, so she tells him that she found it that way when she came out of the store.
3. Jeff is watching his favorite TV show. His friend calls him from the airport, which is 45 min away, and asks for a ride. Jeff doesnʼt
want to miss his show, so he tells his friend that he let someone borrow his car for the night.
4. Belinda borrows her friend Samanthaʼs jacket and accidentally leaves $40 in the pocket and a stain on the sleeve. Samantha finds the money when she takes the coat to get cleaned. Belinda hasnʼt mentioned the money and Samantha is broke, so she decides to keep the $40.
5. When his grades come, Ed sees the recorded grade in math is A. He later bumps into his math teacher, who tells him that he was
sorry to have to give Ed an A—because he was just below the cut off. Ed says nothing.
6. Claire canʼt afford the ink cartridges that her kids need to print their school assignments. The office where Claire works has plenty
of ink cartridges and often discards extras when new printers arrive. When nobody is looking, Claire takes an ink cartridge without
asking.
3172
Journal of Cognitive Neuroscience
Volume 23, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
f
.
t
o
n
1
8
M
a
y
2
0
2
1
. (continued )
Dishonest Scenarios (continued )
7. Ellenʼs daughter needs medicine to relieve her chronic pain, which is available only at CVS (a pharmacy). CVS buys the medicine cheaply
in large quantities and never sells out. They charge much more than Ellen can afford. The manager at CVS refuses to help, so Ellen steals
some medicine.
8. In the morning Taryn promised her mother that she would come over for dinner. She doesnʼt feel like leaving her apartment, however, so
she calls her mother and tells her that she isnʼt feeling well. Her mother, although disappointed, tells her she that she should stay home.
9. Alison deposits a check for $156 in her bank. When her monthly statement arrives, it lists a deposit of $165 instead of $156. Alison knows that this is a mistake, but she decides not to tell anyone at the bank about this error. 10. Every month Alvin has his paychecks deposited automatically into his bank account. When his monthly statement arrives, it lists a deposit of $3530 instead of $3503. Alvin knows that this is an error, but he does not mention the mistake to his employer. 11. Suzy takes Joe to an expensive restaurant that she can barely afford. When the bill comes, she notices that the charges do not include the bottle of wine they drank. She leaves a good tip but does not correct the mistake on the bill. 12. Priscillaʼs house contains a room that she uses mainly for guests and occasionally as an office. For tax purposes, she claims the room is an office, counts it as a business expense, and takes a deduction. She knows that she will never be caught. 13. During a closed-book take-home French exam, Sammy catches a glimpse of an answer on a conjugation table on his wall, which he forgot to take down. His grade wonʼt affect other studentsʼ grades, so he decides not tell the teacher that he looked at the table. 14. Esther asks to borrow Debbieʼs car to go dancing when Esther really needs to study for an important test. Debbie worries about Esther failing, but she does not want to argue or anger Esther, so she tells Esther that her car is broken, although it isnʼt. Disgusting Scenarios 1. Jackʼs aunt is dying of cancer at home. She asks Jack to have sex with her after she dies as a sign of his love. Jack says he will think about it. After she dies, Jack copulates with his auntʼs dead body. There are no ill effects. 2. Phil is visiting his 67-year-old grandmother. As she is baking, he comes up behind her and kisses her passionately. They are both aroused and start to rub up against each other until they climax. This encounter never creates problems for either of them. 3. Tim is hiking one afternoon when he discovers a freshly dead male coyote. He has not seen anyone on the trail all day, so he has anal intercourse with the coyote body, using a condom. Afterwards, he continues his hike and never suffers any ill effects. 4. While taking a walk, Miles finds a recently deceased female deer in his backyard. He gets a condom and has sexual intercourse with the still warm doe carcass. Afterwards, he buries the deer and cleans off. He never suffers any ill effects. 5. James and Holly are brother and sister. After they both graduate from college, they share an apartment in a large building. When nobody else is around, they sometimes touch each othersʼ genitals passionately. This activity never creates any problem for either of them. 6. After college, Leah moved in to live with her grandfather, who is 70 and healthy. One evening Leah and her grandfather fondle each other sexually. They both feel closer afterward but never do it again. Neither suffers any ill effects. 7. Janeʼs father asks her to stimulate his penis right after he dies to see whether he gets an erection. She never promises, but after he dies Jane stimulates his penis for several minutes with her hand. Jane suffers no ill effects, and she feels sexually aroused. 8. Larry and Max are brothers who enjoy boating together. One night, when they are alone on their boat, they have anal intercourse with each other. Both of them enjoy it, and the memory of that experience makes them feel closer. Neither of them ever suffers any ill effects. 9. Katie and David are adult siblings camping in the woods one evening. For fun, they have sexual intercourse. They use multiple forms of birth control and never do it again. Afterwards, they feel a stronger connection, and neither of them ever suffers any ill effects at all. 10. Fred goes to a large chain supermarket once a week and buys a fresh whole chicken. At home, he thoroughly cleans the chicken and rubs butter all over it. Then he has sexual intercourse with it, using a condom. He does this only once. 11. Ann and Bob are adult siblings. When they are alone, they like to kiss each other on the mouth passionately, using their tongues. They do this only a few times, and neither of them ever suffers any ill effects from doing it. 12. While preparing a tenderloin roast for his dinner, Frank cuts a hole into the roast and has sexual intercourse with it, using a condom. Afterwards, he cleans and eats the roast. He does it again a few times but never suffers any ill effects. 13. Once in a while, when he is going to eat at home alone, William obtains sheep penises from an oriental meat market. He makes sure that they are still fresh. Then he cooks the sheep penises thoroughly and simulates oral sex with them while he eats them. 14. Ursula occasionally buys leftover pig sex organs at a butcher shop in a large grocery store. After she takes them home, she plays with these sex organs by inserting the male organs repeatedly into the female organs, slowly at first and then faster to simulate sexual intercourse. Parkinson et al. 3173 D o w n l o a d e d l l / / / / j f / t t i t . : / / f r o m D o h w t t n p o : a / d / e m d i f r t o p m r c h . s p i l d v i r e e r c t c . m h a i e r d . u c o o m c n / j a o r t c i c n e / - a p r d t i 2 c 3 l 1 e 0 - 3 p 1 d 6 f 2 / 1 2 9 3 4 / 2 1 6 0 0 / 1 3 1 o 6 c 2 n _ / a 1 _ 7 0 7 0 6 0 6 1 9 7 1 p / d j o b c y n g _ u a e _ s 0 t 0 o 0 n 1 0 7 8 . S p d e f p e b m y b e g r u 2 e 0 s 2 t 3 / j / t f . o n 1 8 M a y 2 0 2 1 Neutral Scenarios 1. Amy buys a lemon-lime soda at a convenience store for $1.21 after tax. She notices a bowl marked “For our customers: take a
penny, leave a penny” next to the register. She takes a penny so that she can pay the exact amount.
2. Patrick is a lawyer who usually eats dinner with his friends on Mondays. One Monday, he needs to stay late in his office to prepare
for an important hearing the next morning. He apologizes to his friends in advance and meets them again the next week.
3. Paulʼs father was in an automobile accident in which he suffered compound fractures in both legs. His recovery period coincides with
Paulʼs summer holidays. Paul was planning to stay home and take it easy anyway, so he is happy to help his dad recover at home.
4. Melissa is driving to work. The traffic in her lane is moving very slowly, and the cars in the lane next to her are moving faster. She is
late to work, so after looking in her mirror and signaling properly she changes into the faster lane.
5. After lunch, Angelaʼs bill is only for $12, but all she has is a $20 bill. Angela needs to get back to work, so she does not have time
to wait for her change. She puts the $20 bill on the table and leaves.
D
o
w
n
l
o
a
d
e
d
6. Alexis goes to a fancy French restaurant for the first time in her life. She sees vichyssoise (cold potato soup) on the menu. Not
knowing what it is, she orders vichyssoise and eats it. She does not like it very much, but she is glad she tried it.
7. Lindsay and Barry are friends who agree to have lunch on Wednesday at noon. On Wednesday morning, Lindsay feels ill. The only
appointment that is available with her doctor is at noon. Lindsay calls Barry, who says that it is fine to have lunch on Thursday
instead.
8. Lucia is waiting to check out in a long line at a supermarket. She is late for a meeting, so when she notices that there is a shorter
line at another register, she leaves her line and gets in the shorter one.
9. Oscar likes to work out by lifting weights. He believes that Gatorade contains essential vitamins and minerals that will make his
workouts more effective, so he drinks Gatorade before and after working out every day. Otherwise, his diet is completely normal.
10. Julie lives next to a church that is open all night for prayers. Her grandmother is very sick, so Julie goes into the church well after
midnight when nobody else is there and prays for her grandmother until the sun comes up.
11. Rochelle and Bill have been married happily for several years, and they love each other very much. Now they are ready to have
children of their own, so they have sexual intercourse several times at night without using any kind of contraception at all.
12. Marthaʼs next-door neighbors tell Martha that she may come and use their outdoor hot tub whenever she wants without asking
them for permission. One day, Martha visits her neighbors and asks anyway whether she may use their outdoor hot tub.
13. Pennie and Lydia enjoy going for walks in the park together on Sundays. One Sunday, Lydia is away on vacation in Europe. Pennie
decides to go for a walk in the park anyway, without Lydia. She has a lot of fun, although she misses Lydia.
14. Jake asks Antonio whether he may borrow Antonioʼs van to pick up a heavy box from a hardware store. Antonio consents, so Jake
uses the van and then returns it to Antonio with a full tank of gas and no damage to the vehicle.
Table A1. Talairach Coordinates for the Centers of Mass of Significant Clusters for Each Story Type versus Neutral Comparison
Area
Hemi
BA
vxls
Harmful Stories
SMA
DMPFC
DMPFC
DLPFC
DLPFC
STS
Dishonest Stories
DMPFC
DMPFC
TPJ
TPJ
L
L
R
L
R
L
L
R
L
R
6
8/32
9/10
8
9
39
6/8/9/10/32
6/8/9
39/40
40
119
55
38
44
24
44
1069
411
298
61
x
−14
−4
5
−29
23
−44
−14
12
−46
55
y
17
32
52
32
50
−56
33
39
−58
−53
z
55
31
25
38
31
16
40
41
34
35
3174
Journal of Cognitive Neuroscience
Volume 23, Number 10
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
f
t
/
o
n
1
8
M
a
y
2
0
2
1
Table A1. (continued )
Area
ITS
Precuneus
Cerebellum
Disgust Stories
DMPFC
DMPFC
DLPFCa
IFG
SMA
MFG
ACC
sgACC
TP
TP
TP
IPL
Fusiform gyrus
PCC
Amygdala
Amygdala
Cerebellum
All Moral Stories
DMPFC
DMPFC
Hemi
BA
L
L
R
L
R
L
L
R
L
R
R
R
R
L
L
L
L
L
R
R
L
R
21
31
8/9/10/32
9/10
10/13/44/46/47
34/47
6
9
24
25/34
38
38
28/38
7/40
20/37
23/31
9/32
9/10
vxls
58
48
39
721
94
313
277
24
27
26
161
54
31
185
28
118
166
165
112
154
44
28
x
−52
−5
43
−9
7
−43
−13
7
−42
5
10
29
39
−26
−35
−43
−3
−22
21
−6
−4
6
y
−4
−50
−58
41
55
31
−3
15
13
31
−7
10
20
5
−59
−43
−43
−3
−2
−59
33
53
z
−22
31
−37
30
22
7
−3
56
26
15
−6
−24
−22
−25
42
−16
26
−14
−13
−31
30
24
p < .001, corrected.
aCluster includes FO and anterior insula.
Table A2. Frequency and Variance of “Wrong” and “Not Wrong” Judgments Decisions by Scenario, Sorted in Decreasing Order
of Frequency of Wrongness Judgments
Category
Scenario Number of “Wrong” Judgments Number of “Not Wrong” Judgments Missed Trials
Variance
Disgust
Disgust
Disgust
Disgust
Disgust
Disgust
Disgust
Disgust
1
2
3
4
5
6
7
8
31
31
31
30
30
29
29
29
6
7
6
8
8
8
8
9
1
0
1
0
0
1
1
0
0.14
0.15
0.14
0.17
0.17
0.17
0.17
0.18
Parkinson et al.
3175
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
f
t
.
o
n
1
8
M
a
y
2
0
2
1
Table A2. (continued )
Category
Scenario Number of “Wrong” Judgments Number of “Not Wrong” Judgments Missed Trials
Variance
Disgust
Disgust
Disgust
Disgust
Disgust
Disgust
Disgust total
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty
Dishonesty total
Harm
Harm
Harm
Harm
Harm
Harm
Harm
Harm
Harm
Harm
Harm
Harm
Harm
Harm
Harm total
9
10
11
12
13
14
1
2
3
4
5
6
7
8
9
10
11
12
13
14
1
2
3
4
5
6
7
8
9
10
11
12
13
14
27
26
26
22
21
20
382
29
28
28
27
24
23
23
22
22
22
20
15
13
10
306
28
27
25
25
23
23
22
21
20
17
16
14
14
9
284
9
12
12
16
17
18
144
9
9
10
10
13
14
14
16
15
14
16
23
24
24
2
0
0
0
0
0
6
0
1
0
1
1
1
1
0
1
2
2
0
1
4
211
15
9
11
13
11
15
15
15
15
16
19
22
23
23
28
1
0
0
2
0
0
1
2
2
2
0
1
1
1
235
13
0.19
0.22
0.22
0.24
0.25
0.25
0.20
0.18
0.18
0.19
0.20
0.23
0.24
0.24
0.24
0.24
0.24
0.25
0.24
0.23
0.21
0.24
0.18
0.21
0.23
0.21
0.24
0.24
0.24
0.24
0.25
0.25
0.24
0.24
0.24
0.18
0.30
3176
Journal of Cognitive Neuroscience
Volume 23, Number 10
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
2
c
3
l
1
e
0
-
3
p
1
d
6
f
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
c
2
n
_
/
a
1
_
7
0
7
0
6
0
6
1
9
7
1
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
0
o
0
n
1
0
7
8
.
S
p
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
f
t
o
n
1
8
M
a
y
2
0
2
1
Table A2. (continued )
Category
Scenario Number of “Wrong” Judgments Number of “Not Wrong” Judgments Missed Trials
Variance
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral
Neutral total
1
2
3
4
5
6
7
8
9
10
11
12
13
14
3
1
1
1
1
1
0
0
0
0
0
0
0
0
8
35
37
37
37
37
37
38
37
38
38
38
36
38
37
520
0
0
0
0
0
0
0
1
0
0
0
2
0
1
4
0.07
0.03
0.03
0.03
0.03
0.03
0.00
0.00
0.00
0.00
0.00
0.00
0.00
0.00
0.01
Table A3. Verb Frequency by Scenario Category
Scenario Category
Action
Belief
Perception
Desire
Emotion
Communication
Neutral
Harm
Dishonest
Disgust
61
71
72
70
2
2
4
0
4
7
4
2
3
3
5
0
9
4
5
15
7
14
15
3
Each scenario was parsed into the five verb categories described by Bartsch and Wellman (1989). These categories included action (e.g., eats, makes),
belief (e.g., thinks, knows), desire (e.g., wants, tries), emotion (e.g., suffers, feels), and communication (e.g., asks, tells). Action verbs were more fre-
quently represented than any other category. Chi square analyses revealed that the frequency of action, belief, and desire verbs did not differ significantly
across categories (χ2 < 4.64; all ps > .20). The frequency of emotion and communication verbs did differ by scenario category (χ2(3) = 9.06, p = .028
and χ2(3) = 10.13, p = .017, rispettivamente).
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
2
C
3
l
1
e
0
–
3
P
1
D
6
F
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
C
2
N
_
/
UN
1
_
7
0
7
0
6
0
6
1
9
7
1
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
0
o
0
N
1
0
7
8
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
.
T
/
o
N
1
8
M
UN
sì
2
0
2
1
Table A4. Reading Ease and Grade Level by Scenario Category
Scenario Category
Flesch Reading Ease
Flesch–Kincaid Grade
Neutro
Harm
Dishonest
Disgust
68.77 (9.82)
72.62 (14.48)
70.94 (12.77)
70.02 (12.79)
7.69 (1.92)
6.63 (2.63)
7.26 (2.03)
6.66 (2.26)
Both measures (Kincaid, Fishburne, Rogers, & Chissom, 1975; Flesch, 1948) calculate textual difficulty with formulas that use sentence length and number
of syllables per word. These measures are highly correlated in an inverse direction: The higher the reading ease, the lower is the grade level required to
understand the text. Each cell lists the mean with standard deviations in parentheses. There were no significant differences between scenario categories
for reading ease (F(3, 52) = .23, p = .87) or grade level (F(3, 52) = .74, p = .53).
Parkinson et al.
3177
Figure A1. Emotion ratings by
scenario category. After the
scanning session, participants
indicated if each scenario
evoked emotion using a scale
ranging from 1 (not at all ) A
4 (intensely). Error bars indicate
standard error.
Figure A2. Ventral frontal
coverage of sample subject.
Slices depict three planes of
the same Talairach coordinate:
(UN) y = 47, (B) x = 1, (C) z = 0.
Figure A3. Temporal pole
coverage of sample subject.
Slices are from Talairach
coordinates: (UN) x = −44,
(B) z = −22, (C) x = 46.
Ringraziamenti
We thank Liane Young, Joshua Greene, Fiery Cushman, Bertram
Malle, Rebecca Saxe, and the Moral Psychology Research Group
for helpful discussion and feedback and Daniel Leopold, Christina
Chick, Cameron Shirazi, Anne Krendl, and Cecilia Tilli for their
contributions to this project. We also wish to thank Dartmouth
Università, Princeton University, the Australian National University
and the Rotman Institute of Philosophy for institutional support.
Reprint requests should be sent to Thalia Wheatley, Psicologico
and Brain Science, Dartmouth College, Hanover, NH 03755, or via
e-mail: thalia.p.wheatley@dartmouth.edu.
REFERENCES
Amodio, D. M., & Frith, C. (2006). Meeting of minds: IL
medial frontal cortex and social cognition. Nature Reviews
Neuroscience, 7, 268–277.
Apperly, IO. A., Samson, D., Chiavarino, C., Bickerton, W., &
Humphreys, G. W. (2007). Testing the domain-specificity
of a theory of mind deficit in brain-injured patients: Evidence
for consistent performance on non-verbal, “reality-unknown”
false belief and false photograph tests. Cognition, 103,
300–321.
Astafiev, S. V., Shulman, G. L., & Corbetta, M. (2006).
Visuospatial reorienting signals in the human temporo-
parietal junction are independent of response selection.
European Journal of Neuroscience, 23, 591–596.
Bartsch, K., & Wellman, H. M. (1989). Young childrenʼs
attribution of action to beliefs and desires. Child
Development, 60, 946–964.
Allison, T., Puce, A., & McCarthy, G. (2000). Social perception
from visual cues: Role of the STS region. Trends in Cognitive
Scienze, 4, 267–278.
Berthoz, S., Armony, J. L., Blair, R. J. R., & Dolan, R. J. (2002). An
f MRI study of intentional and unintentional (embarrassing)
violations of social norms. Brain, 125, 1696–1708.
3178
Journal of Cognitive Neuroscience
Volume 23, Numero 10
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
2
C
3
l
1
e
0
–
3
P
1
D
6
F
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
C
2
N
_
/
UN
1
_
7
0
7
0
6
0
6
1
9
7
1
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
0
o
0
N
1
0
7
8
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
F
T
.
o
N
1
8
M
UN
sì
2
0
2
1
Blair, J., Marsh, UN. A., Finger, E., Blair, K. S., & Luo, J. (2006).
Neurocognitive systems involved in morality. Philosophical
Explorations, 9, 13–26.
Borg, J. S., Hynes, C., Van Horn, J., Grafton, S., & Sinnott-
Armstrong, W. (2006). Consequences, action, and intention
as factors in moral judgment: An fMRI investigation. Journal
of Cognitive Neuroscience, 18, 803–817.
Borg, J. S., Lieberman, D., & Kiehl, K. UN. (2008). Infection, incest
and iniquity: Investigating the neural correlates of disgust and
morality. Journal of Cognitive Neuroscience, 20, 1529–1546.
Britton, J. C., Phan, K. L., Taylor, S. F., Welsh, R. C., Berridge,
K. C., & Liberzon, IO. (2005). Neural correlates of social and
nonsocial emotions: An fMRI study. Neuroimage, 31, 397–409.
Cavanna, UN. E., & Trimble, M. R. (2005). The precuneus: UN
review of its functional anatomy and behavioral correlates.
Brain, 129, 564–583.
Chen, G., Saad, Z. S., & Cox, R. W. (2010). Modeling multilevel
variance components and outliers in group analysis. In
Proceedings of the 16th Annual Meeting of the Organization
for Human Brain Mapping (P. 43). Minneapolis, MN:
Organization for Human Brain Mapping.
Cheng, Y., Chen, C. Y., Lin, C. P., Chou, K. H., & Decety, J.
(2010). Love hurts: An fMRI study. Neuroimage, 51, 923–929.
Corbetta, M., Patel, G., & Shulman, G. l. (2008). IL
reorienting system of the human brain: From environment
to theory of mind. Neuron, 58, 306–324.
Cox, R. W. (1996). AFNI: Software for analysis and visualization
of functional magnetic resonance neuroimages. Computers
and Biomedical Research, 29, 162–173.
Critchley, H. D. (2004). The human cortex responds to an
interoceptive challenge. Proceedings of the National
Academy of Sciences, U.S.A., 101, 6333–6334.
Cushman, F., Young, L., & Hauser, M. (2006). The role of
conscious reasoning and intuition in moral judgment:
Testing three principles of harm. Psychological Science,
17, 1082–1089.
Decety, J., & Jackson, P. l. (2004). The functional architecture
of human empathy. Behavioral and Cognitive Neuroscience
Reviews, 3, 71–100.
Decety, J., & Sommerville, J. UN. (2003). Shared representations
between self and others: A social cognitive neuroscience
view. Trends in Cognitive Sciences, 7, 527–533.
Dougherty, D. D., Shin, l. M., Alpert, N. M., Pitman, R. K., Orr,
S. P., Lasko, M., et al. (1999). Anger in healthy men: A PET
study using script-driven imagery. Biological Psychiatry,
46, 466–472.
Dupoux, E., & Jacob, P. (2007). Universal moral grammar: UN
critical appraisal. Trends in Cognitive Science, 11, 373–378.
Dwyer, S. J. (1999). Moral competence. In K. Murasugi &
R. Stainton (Eds.), Philosophy and linguistics
( pag. 169–190). Boulder, CO: Westview Press.
Flesch, R. (1948). A new readability yardstick. Journal of
Applied Psychology, 32, 221–233.
Fletcher, P. C., Happe, F., Baker, S. C., Dolan, R. J., Frackowiak,
R. S. J., & Frith, C. D. (1995). Other minds in the brain:
A functional imaging study of “theory of mind” in story
comprehension. Cognition, 57, 109–128.
Gallagher, H. L., & Frith, C. D. (2003). Functional imaging
“theory of mind.” Trends in Cognitive Sciences, 7, 77–83.
Games, P. A., & Howell, J. F. (1976). Pairwise multiple
comparison procedure with unequal Nʼs and/or variances:
A Monte Carlo study. Journal of Educational and
Behavioral Statistics, 1, 113–124.
Glenn, UN. L., Iyer, R., Graham, J., Koleva, S., & Haidt, J. (2009).
Are all types of morality compromised in psychopathy?
Journal of Personality Disorders, 23, 384–398.
Goel, V., Grafman, J., Sadato, M., & Hallett, M. (1995). Modeling
other minds. NeuroReport, 6, 1741–1746.
Grafton, S. T., Arbib, M. A., Fadiga, L., & Rizzolatti, G.
(1996). Localization of grasp representations in humans by
positron emission tomography: 2. Observation compared with
imagination. Experimental Brain Research, 112, 103–111.
Greene, J. D., Nystrom, l. E., Engell, UN. D., Darley, J. M., &
Cohen, J. D. (2004). The neural basis of cognitive conflict
and control in moral judgment. Neuron, 44, 389–400.
Greene, J. D., & Paxton, J. M. (2009). Patterns of neural activity
associated with honest and dishonest moral decisions.
Proceedings of the National Academy of Sciences,
U.S.A., 106, 12506–12511.
Greene, J. D., Sommerville, R. B., Nystrom, l. E., Darley, J. M., &
Cohen, J. D. (2001). An fMRI investigation of emotional
engagement in moral judgment. Scienza, 293, 2105–2108.
Haidt, J. (2001). The emotional dog and its rational tail: A social
intuitionist approach to moral judgment. Psicologico
Review, 108, 814–834.
Haidt, J., & Graham, J. (2007). When morality opposes justice:
Conservatives have moral intuitions that liberals may not
recognize. Social Justice Research, 20, 98–116.
Haidt, J., & Hersh, M. (2001). Sexual morality: The cultures and
emotions of conservatives and liberals. Journal of Applied
Social Psychology, 31, 191–221.
Hauser, M., Cushman, F., Young, L., Jin, R. K. X., & Mikhail, J.
(2007). A dissociation between moral judgments and
justifications. Mind & Language, 22, 1–21.
Hauser, M. D. (2006). Moral minds: How nature designed
our universal sense of right and wrong. New York:
Ecco/Harper Collins.
Heekeren, H. R., Wartenburger, I., Schmidt, H., Prehn, K.,
Schwintowski, H.-P., & Villringer, UN. (2005). Influence
of bodily harm on neural correlates of semantic and
moral decision-making. Neuroimage, 24, 887–897.
Eh, S., Wei, N., Wang, Q., Yan, L., Wei, E., Zhang, M., et al.
(2008). Patterns of brain activation during visually evoked
sexual arousal differ between homosexual and heterosexual
men. American Journal of Neuroradiology, 29, 1890–1896.
Huebner, B., Dwyer, S., & Hauser, M. (2009). The role of
emotion in moral psychology. Trends in Cognitive
Scienze, 13, 1–6.
Immorino-Yang, M. H., McColl, A., Damasio, H., & Damasio, UN.
(2009). The neural correlates of admiration and compassion.
Proceedings of the National Academy of Sciences, U.S.A.,
106, 8021–8026.
Ishai, A., Ungerleider, l. G., & Haxby, J. V. (2000). Distributed
neural systems for the generation of visual images. Neuron,
28, 979–990.
Jabbi, M., Bastiaansen, J., & Keysers, C. (2008). A common
anterior insula representation of disgust observation,
experience and imagination shows divergent functional
connectivity pathways. PLoS ONE, 3, e2939.
Jenkins, UN. C., & Mitchell, J. P. (2010). Mentalizing under
uncertainty: Dissociated neural responses to ambiguous
and unambiguous mental state inferences. Cerebral Cortex,
20, 404–410.
Kedia, G., Berthoz, S., Wessa, M., Hilton, D., & Martinot, J.
(2008). An agent harms a victim: A functional magnetic
resonance study on specific moral emotions. Journal of
Cognitive Neuroscience, 20, 1788–1798.
Kincaid, J. P., Fishburne, R. P., Jr., Rogers, R. L., & Chissom, B. S.
(1975). Derivation of new readability formulas (Automated
Readability Index, Fog Count and Flesch Reading Ease
Formula) for Navy enlisted personnel. Research Branch
Report (pag. 8–75). Millington, TN: Naval Technical Training,
U. S. Naval Air Station, Memphis, TN.
Maddock, R. J. (1999). The retrosplenial cortex and emotion:
New insights from functional neuroimaging of the human
brain. Trends in Neurosciences, 22, 310–316.
Parkinson et al.
3179
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
2
C
3
l
1
e
0
–
3
P
1
D
6
F
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
C
2
N
_
/
UN
1
_
7
0
7
0
6
0
6
1
9
7
1
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
0
o
0
N
1
0
7
8
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
.
F
T
o
N
1
8
M
UN
sì
2
0
2
1
Mendez, M. F., Anderson, E., & Shapira, J. S. (2005). An
Rosch, E., & Mervis, C. B. (1975). Family resemblances:
investigation of moral judgment in frontotemporal dementia.
Cognitive and Behavioral Neurology, 18, 193–197.
Studies in the internal structure of categories. Cognitive
Psychology, 7, 573–605.
Mikhail, J., Sorrentino, C. M., & Spelke, E. S. (1998). Toward
a universal moral grammar. In M. UN. Gernsbacher & S. UN.
Derry (Eds.), Proceedings of the Twentieth Annual
Conference of the Cognitive Science Society (P. 1250).
Mahwah, NJ: Lawrence Erlbaum Associates.
Moll, J., de Oliveira-Souza, R., & Eslinger, P. J. (2003). Morals
and the human brain: A working model. NeuroReport,
14, 299–305.
Moll, J., de Oliveira-Souza, R., Garrido, G. J., Bramati, IO. E.,
Caparelli-Daquer, E. M. A., Paiva, M. l. M. F., et al. (2007). IL
self as a moral agent: Linking the neural basis of social agency
and moral sensitivity. Social Neuroscience, 2, 336–352.
Moll, J., de Oliveira-Souza, R., Moll, F. T., Ignacio, F. A.,
Bramati, IO. E., Caparelli-Daquer, E., et al. (2005). The moral
affiliations of disgust: A functional MRI study. Cognitive
and Behavioral Neurology, 18, 68–78.
Moll, J., Zahn, R., de Oliveira-Souza, R., Krueger, F., & Grafman, J.
(2005). The neural basis of human moral cognition. Nature
Reviews Neuroscience, 6, 799–809.
Saxe, R. (2009). The happiness of the fish: Evidence for a
common theory of oneʼs own and othersʼ actions. In
K. Markman, B. Klein, & J. Suhr (Eds.), The handbook
of imagination and mental simulation (pag. 257–266).
New York: Taylor & Francis.
Saxe, R., & Kanwisher, N. (2003). People thinking about
thinking people: The role of the temporo-parietal junction
in “theory of mind.” Neuroimage, 19, 1835–2842.
Shweder, R. A., Much, N. C., Mahapatra, M., & Park, l. (1997).
The “big three” of morality (autonomy, community, E
divinity), and the “big three” explanations of suffering. In
UN. Brandt & P. Rozin (Eds.), Morality and health
(pag. 119–169). New York: Routledge.
Sinott-Armstrong, W. (2008). Is moral phenomenology unified?
Phenomenology and the Cognitive Sciences, 7, 85–97.
Takahashi, H., Yahata, N., Koeda, M., Matsuda, T., Asai, K., &
Okubo, Y. (2004). Brain activation associated with evaluative
processes of guilt and embarrassment: An f MRI study.
Neuroimage, 23, 967–974.
Northoff, G., Heinzel, A., de Greck, M., Bermpohl, F.,
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
Dobrowolny, H., & Panksepp, J. (2006). Self-referential
processing in our brain—A meta-analysis of imaging studies
on the self. Neuroimage, 31, 440–457.
Ochsner, K. N., Knierim, K., Ludlow, D. H., Haneline, J.,
Ramachandran, T., Glover, G., et al. (2004). Reflecting upon
feelings: An fMRI study of neural systems supporting the
attribution of emotion to self and others. Journal of
Cognitive Neuroscience, 16, 1746–1772.
Olson, IO. R., Plotzker, A., & Ezzyat, Y. (2007). The enigmatic
temporal pole: A review of findings on social and emotional
processing. Brain, 130, 1718–1731.
Prinz, J. J. (2008). Is morality innate? In W. Sinnott-Armstrong
(Ed.), Moral psychology (pag. 267–406). Cambridge, MA:
CON Premere.
Rawls, J. (1971). A theory of justice. Cambridge, MA: Harvard
Stampa universitaria.
Rilling, J. K., Sanfey, UN. G., Aronson, J. A., Nystrom, l. E., &
Cohen, J. D. (2004). The neural correlates of theory of mind
within interpersonal interactions. Neuroimage, 22, 1694–1703.
atlas of the human brain. New York: Thieme.
Van Overwalle, F. (2009). Social cognition and the brain: UN
meta-analysis. Human Brain Mapping, 30, 585–594.
Volz, K. G., Schubotz, R. I., & von Cramon, D. Y. (2003).
Predicting events of varying probability: Uncertainty
investigated by fMRI. Neuroimage, 19, 271–280.
Wheatley, T., & Haidt, J. (2005). Hypnotic disgust makes
moral judgments more severe. Psychological Science, 16,
780–784.
Young, L., & Saxe, R. (2008). The neural basis of belief encoding
and integration in moral judgment. Neuroimage, 40,
1912–1920.
Zahn, R., Moll, J., Krueger, F., Huey, E. D., Garrido, G., &
Grafman, J. (2006). Social concepts are represented in the
superior anterior temporal cortex. Atti del
National Academy of Sciences, U.S.A., 104, 6430–6435.
Zahn, R., Moll, J., Paiva, M., Garrido, G., Krueger, F., Huey, E. D.,
et al. (2009). The neural basis of human social values:
Evidence from functional MRI. Cerebral Cortex, 19, 276–283.
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
R
T
o
P
M
R
C
H
.
S
P
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
2
C
3
l
1
e
0
–
3
P
1
D
6
F
2
/
1
2
9
3
4
/
2
1
6
0
0
/
1
3
1
o
6
C
2
N
_
/
UN
1
_
7
0
7
0
6
0
6
1
9
7
1
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
0
o
0
N
1
0
7
8
.
S
P
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
F
.
T
o
N
1
8
M
UN
sì
2
0
2
1
3180
Journal of Cognitive Neuroscience
Volume 23, Numero 10