Infection, Incest, and Iniquity: Investigating the
Neural Correlates of Disgust and Morality
Jana Schaich Borg1, Debra Lieberman2, and Kent A. Kiehl3,4
Astratto
& Disgust, an emotion related to avoiding harmful substances,
has been linked to moral
judgments in many behavioral
studies. Tuttavia, the fact that participants report feelings of
disgust when thinking about feces and a heinous crime does
not necessarily indicate that the same mechanisms mediate
these reactions. Humans might instead have separate neural
and physiological systems guiding aversive behaviors and
judgments across different domains. The present interdisci-
plinary study used functional magnetic resonance imaging
(n = 50) and behavioral assessment to investigate the bio-
logical homology of pathogen-related and moral disgust. Noi
provide evidence that pathogen-related and sociomoral acts
entrain many common as well as unique brain networks. Noi
also investigated whether morality itself
is composed of
distinct neural and behavioral subdomains. We provide evi-
dence that, despite their tendency to elicit similar ratings of
moral wrongness, incestuous and nonsexual immoral acts en-
train dramatically separate, while still overlapping, brain
networks. These results (io) provide support for the view
that the biological response of disgust is intimately tied to
immorality, (ii) demonstrate that there are at least three
separate domains of disgust, E (iii) suggest strongly that
morality, like disgust, is not a unified psychological or neuro-
logical phenomenon. &
INTRODUCTION
Disgust is an emotion traditionally defined as an aversive
state that motivates withdrawal from offensive substances,
such as animal products and certain foods (Rozin &
Fallon, 1987). Recentemente, researchers from a variety of
disciplines have taken interest in the relationship be-
tween disgust and moral behavior. Per esempio, social
psychologists have argued that disgust is an emotion
underpinning moral judgment (Haidt, Rozin, McCauley,
& Imada, 1997), adaptation-minded researchers have
proposed that disgust is a mechanism co-opted from its
original function as a pathogen avoidance system to also
guide decision-making in the moral domain (Lieberman,
Tooby, & Cosmides, 2003), clinical cognitive neuro-
scientists have addressed how impairments in disgust
might underlie psychopathic behavior (Blair, Marsh,
Finger, Blair, & Luo, 2006), and lawyers and policy-
makers have considered how disgust affects the judg-
ment and sentencing of criminals (Kahan, 1998). Despite
the purported role disgust plays in mediating the avoid-
ance of harmful substances and regulating sociomoral
judgments, the fact that subjects across a variety of
experiments report feelings of disgust when considering
both feces and a heinous crime (Mugnaio, 1997) does not
necessarily indicate that the same mechanisms mediate
1Stanford University, 2University of Hawaii, 3University of New
Mexico, 4The MIND Research Network
these reactions. Humans might instead have separate
neural and physiological systems guiding aversive judg-
ments and behaviors across distinct types of nonsocial
and social domains, all of which precipitate a feeling we
subjectively label ‘‘disgust’’ for lack of a more refined
linguistic term (Bloom, 2004).
Given the growing number of publications across aca-
demic disciplines that cite a relationship between dis-
gust and moral disapprobation,
it seems critical to
determine whether our reactions toward items such as
vomit, feces, and rotten food are indeed biologically
homologous to our reactions toward acts such as incest,
theft, and murder, or whether they are instead only lin-
guistically (but not biologically) analogous. Accordingly,
the first goal of the present interdisciplinary study was to
use functional magnetic resonance imaging (fMRI; n =
50) and behavioral assessment to begin characterizing
the proposed homology between pathogen-related and
sociomoral disgust. Drawing upon theoretical insights
and recent discoveries from evolutionary psychology,
moral psychology, and clinical cognitive neuroscience
(Koenigs et al., 2007; Schaich Borg, Hynes, Van Horn,
Grafton, & Sinnott-Armstrong, 2006; Heekeren et al., 2005;
Greene, Nystrom, Engell, Darley, & Cohen, 2004), we
converged upon another hypothesis: Like feelings of
disgust, perhaps feelings of moral wrongness do not
arise from a singular mechanism. Despite the fact
that moral wrongness is usually conceptualized as a
singular feeling or judgment, some separate neural and
D 2008 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 20:9, pag. 1529–1546
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
T
F
.
.
/
o
N
1
8
M
UN
sì
2
0
2
1
physiological systems may guide negative decisions and
aversive behaviors in response to different classes of so-
cial transgressions. Così, the second goal of the present
study was to investigate the shared and distinct neural
correlates of two different domains of sociomoral trans-
gressions: sexual immoral behaviors (incest) and non-
sexual immoral behaviors (such as cheating, stealing, E
killing).
A combination of behavioral and clinical/imaging in-
vestigations inspired the present collaboration. Cross-
culturally, disgust is the emotion most correlated with
appraisals of immorality (Scherer, 1997). For instance,
when respondents are asked to nominate acts eliciting
disgust, the majority of acts mentioned are moral of-
fenses (Haidt et al., 1997). Inoltre, conventional
transgressions (rule violations that would not be con-
sidered wrong in the absence of a cultural prohibition)
are more often judged to be moral violations (violations
that are considered wrong regardless of local conven-
zioni) if the violations are disgusting. Additionally, IL
more disgust-sensitive an individual is, the more likely he
or she is to judge a disgusting conventional transgression
as morally wrong (Nichols, 2002). It has also been shown
that American participants hypnotized to feel a flash of
disgust upon reading an arbitrary word judged acts de-
scribed using that word to be more morally wrong than
control participants, even if the act was judged by control
participants to have no moral content (Wheatley & Haidt,
2005). Most recently, it has been demonstrated that re-
calling unethical deeds inspires participants to buy and
use more cleaning products, a pattern interpreted as an
attempt to wash away feelings of impurity and disgust
(Zhong & Liljenquist, 2006). In sum, behavioral studies
demonstrate that, across cultures, reports of disgust and
moral disapproval often coincide.
Investigations constructed around evolutionary prin-
ciples also link disgust and morality. Adaptation-minded
researchers have described disgust as a mechanism that
evolved to motivate the avoidance of substances asso-
ciated with disease-causing agents in ancestral envi-
ronments (Curtis & Biran, 2001). In support, pictures of
objects cuing pathogen presence (via slimy, moist sur-
faces or colors reminiscent of body fluids) are rated as
more disgusting than the same objects presented with-
out pathogen-related cues (via furry surfaces or bright
colors rarely associated with animate substances; Curtis,
Aunger, & Rabie, 2004). Disgust is proposed to have
been co-opted for other purposes as well, in particular,
for the regulation of social judgments and behaviors.
Adaptationist logic states that these new and separate
functions would have evolved in ways specific to the
environmental pressures disgust was being co-opted to
solve. Through analyzing these pressures, adaptation-
minded researchers have predicted that disgust evolved
to solve the problem of incest, a sexual behavior with
deleterious consequences (Lieberman, Tooby, & Cosmides,
2007; Fessler & Navarrete, 2004). Counterintuitively, al-
though incest is generally considered to be immoral
in most contemporary human societies, disgust’s func-
tion in incest avoidance is argued to be distinct from
disgust’s function in nonsexual social domains, Dove
disgust is thought to mediate behaviors and judg-
ments toward sociomoral harms such as theft, violence,
cheating, and deception (Haidt et al., 1997). Therefore,
in nuanced contrast to past descriptions of disgust
in morality (ranging from Haidt et al., 1997 to Zhong
& Liljenquist, 2006), an adaptationist view predicts
that there should be separate sexual and nonsexual
moral-related disgust systems. In total, there should be
pathogen-, sexual moral– and nonsexual moral–related
disgust systems that all share a common disgust avoid-
ance mechanism, but that are also dissociable according
to the unique collection of information processing sys-
tems required by the specific adaptive problem each
type of disgust evolved to solve (per esempio., systems that detect
odors or colors to detect pathogen presence, systems
that estimate genetic relatedness to avoid incest, E
systems that assess social costs to evaluate sociomoral
harms). Recent behavioral evidence supports this novel
prediction (Lieberman et al., 2007; Simpson, Carter,
Anthony, & Overton, 2006; Fessler & Navarrete, 2003),
and provides the framework for the neural hypotheses
investigated in the current study.
Although psychologists have made progress in under-
standing how self-reports of disgust can be parsed,
clinical and cognitive neuroscientists have made prog-
ress in understanding how disgust is represented in the
brain. The brain regions associated with disgust have
been identified through a growing number of clinical
populations with selective disgust impairments (Suzuki,
Hoshino, Shigemasu, & Kawamura, 2006; Schienle et al.,
2003; Sprengelmeyer et al., 1996), indicating that disgust
is a unique biological response differentiable from other
responses, such as fear (Williams et al., 2005). Disgust is
dissociable in healthy human populations as well, E
has been correlated with activity in the thalamus, basal
ganglia, visual cortex, and sometimes, the amygdala,
anterior insula, and medial prefrontal cortex (Moll, Di
Oliveira-Souza, et al., 2005; Schafer, Schienle, & Vaitl,
2005; Stark et al., 2003, 2005; Fitzgerald et al., 2004;
Wicker et al., 2003; Zald, 2003; Phan, Wager, Taylor, &
Liberzon, 2002; Calder, Keane, Manes, Antoun, & Young,
2000; Phillips et al., 1997, 1998).
Little work has been done to investigate the neural
correlates of disgust in the context of social interactions,
but one study showed that a patient with lesions in the
left insula and the putamen scored lower than control
subjects on most pathogen-related subscales of the
Disgust Scale (Haidt, McCauley, & Rozin, 1994), Ma
higher than controls on the sex subscale (Calder et al.,
2000). Another recent study showed that Huntington’s
disease patients scored higher than controls on the sex
and hygiene subscales of the Disgust Scale, but scored
equally on all other subscales, including the food, animal
1530
Journal of Cognitive Neuroscience
Volume 20, Numero 9
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
.
/
F
.
T
o
N
1
8
M
UN
sì
2
0
2
1
natura, and body products subscales (Hayes, Stevenson,
& Coltheart, 2007). Huntington’s disease patients were
also more able to produce scenarios describing moral
disgust violations than pathogen disgust violations.
These last sets of data suggest that different types of
disgust are neurally dissociable, but this possibility is not
often discussed in the neuroscience literature, even in
the studies in which the data were originally published.
Two recent fMRI studies provide preliminary data that
common brain regions are entrained by certain types of
disgust and sociomoral sentiment (Sambataro et al.,
2006; Moll, de Oliveira-Souza, et al., 2005). Tuttavia,
in these studies, the authors investigated the emotion of
‘‘indignation’’ (Moll, de Oliveira-Souza, et al., 2005) E
facial expressions of contempt (Sambataro et al., 2006),
and it is not clear how ‘‘indignation’’ or ‘‘contempt’’
maps onto other taxonomies of disgust or relates spe-
cifically to ratings of moral wrongness. Inoltre, IL
stimuli used by Moll, de Oliveira-Souza, et al. (2005)
conflated pathogen and moral disgust by using only
moral stimuli that referenced pathogens (such as inten-
tionally putting a spider on a baby’s face or spotting a
cockroach in a restaurant). The authors also did not
differentiate incest from other types of moral transgres-
sions, which might be problematic given previous work
demonstrating that the predictors of moral sentiments
toward incest are not the same as the predictors of
moral sentiments toward other moral transgressions
(Lieberman et al., 2007). Così,
it has still not been
tested whether pathogen and moral disgust entrain
common neural systems, despite the number of disci-
plines that assume their biological commonalities. It has
also not been tested whether equally intense sentiments
of moral wrongness associated with different classes of
moral transgressions are correlated with a singular set of
psychological or neurological processes, despite the
accruing number of fMRI and lesion studies published
on moral processing. The present study was designed to
address these two main issues.
Here we investigate the common and distinct neural
and behavioral signatures of reactions toward pathogen-
related substances and sociomoral violations. Fifty male
participants were given a memory/recognition task in-
volving pathogen-related acts (pathogen), pathogen-
unrelated sociomoral acts (sociomoral), and neutral acts
(neutro) while being scanned in a 3-T magnetic reso-
nance imaging scanner. Sociomoral acts were divided
into incestuous acts (incest) and nonsexual sociomoral
acts (nonsexual moral). After being scanned, participants
provided reports of disgust, appeal, and moral wrong-
ness for each act they saw (see Methods). In light of
previous theoretical and empirical work, we made the
following predictions: (io) Pathogen-related acts and so-
ciomoral acts would be rated as more disgusting than
neutral acts, and memorizing and recalling phrases
describing these disgusting acts would activate common
neural regions including those previously shown to be
involved in disgust processing. (ii) Further, if cognitive
processing of these disgust domains also requires the
use of distinct information processing systems in addi-
tion to their common disgust system, then pathogen-
related acts and sociomoral acts should be dissociable
both in the self-report ratings they provoke and in the
brain regions they entrain. (iii) Finalmente, despite eliciting
similar ratings of moral wrongness and activating com-
mon regions of the brain previously shown to be in-
volved in moral processing—including mainly the medial
prefrontal cortex and areas around the temporal–parietal
junction (Schaich Borg et al., 2006; Heekeren et al., 2005;
Moll, Zahn, de Oliveira-Souza, Krueger, & Grafman,
2005; Greene et al., 2004; Heekeren, Wartenburger,
Schmidt, Schwintowski, & Villringer, 2003; Greene &
Haidt, 2002)—we drew from adaptationist principles to
predict that incestuous acts and nonsexual immoral acts
would, themselves, be behaviorally and neurally disso-
like disgust, might not be a
ciable. Questo è, morality,
unified psychological or neurological phenomenon in
the way it is traditionally understood and operational-
ized in behavioral and fMRI studies.
METHODS
Subjects
Subjects were recruited via advertisements in the Fall of
2004. Following previous investigations of disgust that
recruited subjects of one sex only (Schienle et al., 2002,
2006; Wicker et al., 2003; Phillips et al., 1998), we chose
to study men only (a similar study with all women is
being prepared for future publication). Fifty healthy men
(age = 25 ± 6 years) provided written, informed, IRB-
approved consent at Hartford Hospital and Yale Univer-
sity and were compensated $20/hr for their participation.
All participants were right-handed on self-report and
were able to perform the task successfully during prac-
tice sessions prior to scanning.
Experimental Task
To test implicit processing of our experimental condi-
zioni, we intentionally used a task that preoccupied
subjects with mental operations irrelevant to our con-
ditions of interest. We selected a simple memory task
shown previously to effectively probe affective process-
ing (Kiehl et al., 2005). During scanning, subjects were
presented with memorize–recall blocks of short state-
ments from one of four conditions: pathogen (P), incest
(IO), nonsexual moral (M), or neutral (N) (see below).
Each block contained statements from only one condi-
zione. Subjects memorized four statements in the ‘‘mem-
orize’’ phase of each block and reported in the ‘‘recall’’
phase whether each of four subsequent statements was
one shown in the previous memorize section or a new
statement (Figura 1). Two of the statements in each
Schaich Borg, Lieberman, and Kiehl
1531
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
.
.
T
F
/
o
N
1
8
M
UN
sì
2
0
2
1
‘‘memorize’’ phase were randomly chosen to appear in
random positions in the subsequent ‘‘recall’’ phase (par-
ticipants were not told what proportion of the state-
ments in the ‘‘recall’’ phase would be ones they had
just seen). In total, each block had six condition-related
statements: two presented in both the ‘‘memorize’’ and
‘‘recall’’ phases, two presented only in the ‘‘memorize’’
phase, and two presented only in the ‘‘recall’’ phase.
Statements were presented one at a time for 2500 msec
and had an intertrial interval of 500 msec. All statements
were equated for length and complexity. Subjects re-
sponded with their right hand, and were instructed to
press the button underneath their index finger if the
displayed statement was one they had just seen in the
‘‘memorize’’ phase or the button underneath their mid-
dle finger if the displayed statement was one they had not
seen before. Upon completion of each ‘‘recall’’ phase, IL
word ‘‘REST’’ was displayed continuously on the screen
for 5000 msec.
Two runs of 12 randomized memorize–recall–rest
blocks were presented on a visual display projected
from the back of the scanner with 30 sec of rest at the
beginning and end of each run. Of these 12 blocks, three
blocks included statements about physically repulsive
acts performed on/with a sibling of the opposite sex
(Pathogen: You sipping your sister’s urine, You eating
your sister’s scab), three included statements about
incestuous acts performed on/with a sibling of the
opposite sex (Incest: You watching your sister mastur-
bate, You fondling your sister’s nipples), three included
statements about nonsexual immoral actions performed
against a sibling of the opposite sex (Nonsexual moral:
You killing your sister’s child, You burglarizing your
sister’s home), and three included neutral statements
about actions performed on/with a sibling of the oppo-
site sex (Neutro: You holding your sister’s groceries,
You walking with your sister). In total, six blocks each of
pathogen,
incest, nonsexual moral, and neutral state-
ments were presented to participants by the end of the
second run. All stimuli across conditions were equated
for sibling presence. We analyzed all male data together,
independent of family composition given unpublished
work showing that men with sisters (n = 205) and men
without sisters (n = 87) do not provide significantly
Figura 1. An illustration of the organization of the 12 blocks in
each run: 3 neutro (N), 3 pathogen (P), 3 incest (IO), E 3 nonsexual
moral (M ).
different disgust ratings of subsets of pathogen-related
( p = .22, two-tailed) or incestuous acts ( p = .54, two-
tailed) described in the second person (DL, unpublished
dati). Two-sample t tests confirmed that brain activity of
men with sisters did not differ significantly from brain
activity of men without sisters in pathogen versus neu-
tral,
incest versus neutral, or nonsexual moral versus
neutral contrasts.
After the scanning session, subjects completed a short
survey in which they rated each of the acts presented
during the scanning session on levels of disgust, appeal,
and moral wrongness.
Imaging
fMRI data were collected on a Siemens Allegra 3-T head-
dedicated scanner equipped with 40 mT/m gradients
and a standard quadrature head coil at the Olin Neuro-
psychiatry Research Center at the Institute of Living.
Participants viewed all experimental stimuli via a mirror
on top of the head coil that reflected a screen at the rear
entrance of the magnet bore. Stimuli were displayed
on the screen using a computer-controlled projection
system. A custom visual presentation package (VAPP;
http://nilab.psychiatry.ubc.ca/vapp) controlled the tim-
ing of the experimental stimuli and recorded behavioral
dati. The functional scans were acquired using gradient-
echo echo-planar imaging (EPI) (scanning parameters:
repeat time [TR] = 1.50 sec, echo time [TE] = 27 msec,
field of view = 24 cm, acquisition matrix = 64 (cid:1) 64, flip
angle = 708, voxel size = 3.75 (cid:1) 3.75 (cid:1) 4 mm, gap =
1 mm, 29 slices, ascending acquisition). Six ‘‘dummy’’
scans were performed at the beginning of each func-
tional run to allow for longitudinal equilibrium and were
discarded before image analysis. Behavioral responses
were recorded using an MRI-compatible fiber-optic re-
sponse device (Lightwave Medical, Vancouver, BC).
Analysis
Preprocessing
fMRI data were preprocessed using Statistical Parametric
Mapping (SPM2, Wellcome Department of Imaging Neu-
roscience). Functional images were reconstructed off-
line, and the two runs were separately realigned to the
first scan of the session using INRIalign, a motion
correction algorithm unbiased by local signal changes
(Freire, Roche, & Mangin, 2002). Translation and rota-
tion corrections did not exceed 2.5 mm and 2.58,
rispettivamente, for any of the participants. After realign-
ment, a mean functional (EPI) image was computed for
each run and was subsequently matched to the SPM2
EPI template. Data were transformed into standard
Montreal Neurological Institute space using a tailored
algorithm with both linear and nonlinear components,
and this transformation was then applied to all other
1532
Journal of Cognitive Neuroscience
Volume 20, Numero 9
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
/
.
.
.
T
F
o
N
1
8
M
UN
sì
2
0
2
1
corresponding functional images (Friston et al., 1995).
Finalmente, data were spatially smoothed four times the
voxel dimensions (3 (cid:1) 3 (cid:1) 3 mm) with a full width at
half maximum Gaussian kernel (Kiehl et al., 2005), E
submitted to a fifth-order infinite impulse response
Butterworth low-pass filter of 0.16 Hz to remove any
high-frequency noise.
Data Analysis
Each individual participant’s first-level analysis modeled
the canonical hemodynamic response with temporal
derivative to each condition of interest in a block design.
The amplitude of the hemodynamic response was cal-
culated from both its nonderivative and derivative terms
(Calhoun, Stevens, Pearlson, & Kiehl, 2004) to reduce
the impact of spatially varying hemodynamic delays and
delays due to slice timing differences. First-order motion
parameters obtained from realignment were included as
confounds in each participant’s model to remove possi-
ble residual task-related motion effects. A high-pass filter
(cutoff period = 256 sec) was also incorporated into the
model to remove noise associated with low-frequency
confounds (per esempio., respiratory artifact, scanner drift). No
within-session scaling (also called proportional scaling) era
used. Contrasts from these first-level analyses were sub-
sequently entered into group-level random effects models.
Contrasts and conjunction analyses were performed
according to previously published guidelines (Nichols,
Brett, Andersson, Wager, & Poline, 2005). The conjunc-
tion analysis (using the conjunction null hypothesis) era
calculated using a within-subject three-way analysis of
variance of the pathogen versus neutral, incest versus
neutro, and nonsexual moral versus neutral contrasts,
corrected for nonsphericity, as were subsequent analyses
comparing individual disgust conditions. All contrasts
used in these analyses were conservatively performed
on the memorize and recall phases combined together
in one block (following Kiehl et al., 2005), as we had no
a priori hypotheses about how each condition might
interact with experimental phase and there was no
significant precedent in the literature for analyzing one
phase over the other (but see Supplementary Informa-
zione: www.disgustandmorality.com). We also had no a
priori evidence proving incest acts would have different
neural correlates than nonsexual moral acts. For this
reason, all initial contrasts between pathogen disgust (P)
and sociomoral disgust (S–M) were performed using an
average of the incest (IO) and nonsexual moral (M) con-
ditions (as would be the case if other studies included
incest-related stimuli in their moral stimuli). We believed
this would be the most conservative a priori model for
our data, as well as the model most similar to what has
been applied in previous studies of disgust and moral
processing.
All data reported in this study are significant at p < .05,
FDR-corrected, or less. Figures often display contrast
maps at lower p values to facilitate visualization of
discernible clusters. Additional figures illustrating effects
at multiple p levels are posted in the Supplementary
Information.
RESULTS
Behavioral Results
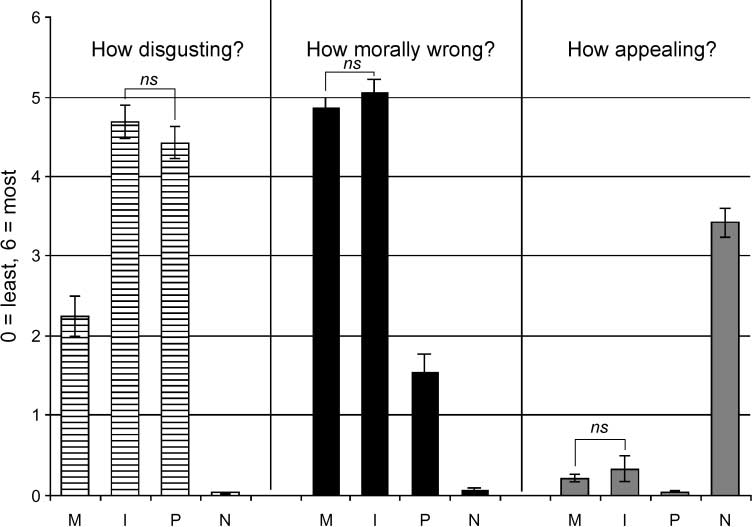
Participants correctly classified an average of 91.5% of
the phrases they saw in the ‘‘recall’’ phase as ones they
had seen in the previous ‘‘memorize’’ phase, and each
classification took about 1.2 sec. There were no signif-
icant differences in accuracy or reaction time between
conditions (Table 1). Five participants did not fill out the
postscanning surveys. From the 45 subjects who did,
pathogen acts and sociomoral acts were rated as more
disgusting than neutral acts by self-report, and within
sociomoral acts, incest acts were rated as more disgust-
ing than nonsexual moral acts. Still, incest and nonsexual
moral acts were rated as equally immoral. Neutral acts
were the only acts to be rated as appealing (Figure 2).
Pathogen and Sociomoral Disgust Activate
Common Neural Regions
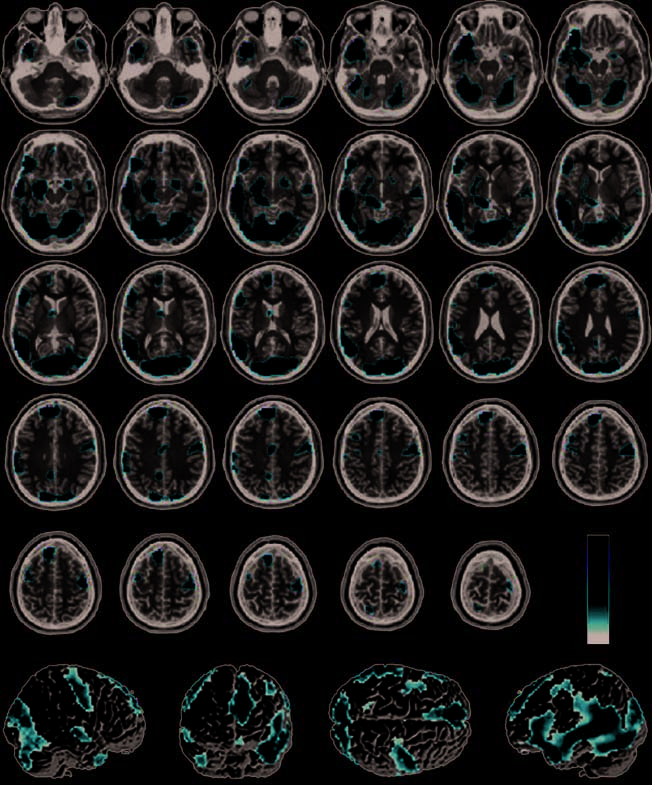
We performed a conjunction analysis to identify brain
regions that were significantly more active during the
memorizing and recognizing of both pathogen and
sociomoral acts than to neutral acts (see Methods). A
large network of brain regions was common to pro-
cessing both of these disgust-eliciting stimuli (Figure 3,
Table 2). Regions in this network included the basal
ganglia (covering the globus pallidus, putamen, and
caudate head), the amygdala, the thalamus, the para-
hippocampal gyrus, the dorsal anterior cingulate (BA 24),
the precuneus, the visual cortex (BA 17/18), and both
the precentral (BA 6) and postcentral gyri (BA 3/4/7). No
significant voxels appeared in the anterior insula.
As illustrated in Figure 3, the most significant and
extensive activity in the conjunction analysis appeared in
the medial prefrontal cortex and the left temporal lobe.
The medial prefrontal cortex activations consisted of
one extensive dorsal cluster (BA 9/10 extending dorsally
to BA 8/6) and one smaller, more ventral cluster (BA 11),
both slightly left-lateralized. Left temporal lobe activations
Table 1. Accuracy and Reaction Times for the Task
Presented in the Scanner
Type of Act
Pathogen
Incest
Nonsexual moral
Neutral
Percent Correctly
Recognized
Mean Reaction
Time (sec)
93.3
91.5
92.0
89.3
1.21
1.23
1.16
1.16
Schaich Borg, Lieberman, and Kiehl
1533
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
9
e
-
1
p
5
d
2
f
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
c
9
n
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
9
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
f
.
/
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Self-report ratings.
M = nonsexual moral acts;
I = incest acts; P = pathogen
acts; N = neutral acts. ns = not
significant.
Figure 3. Conjunction
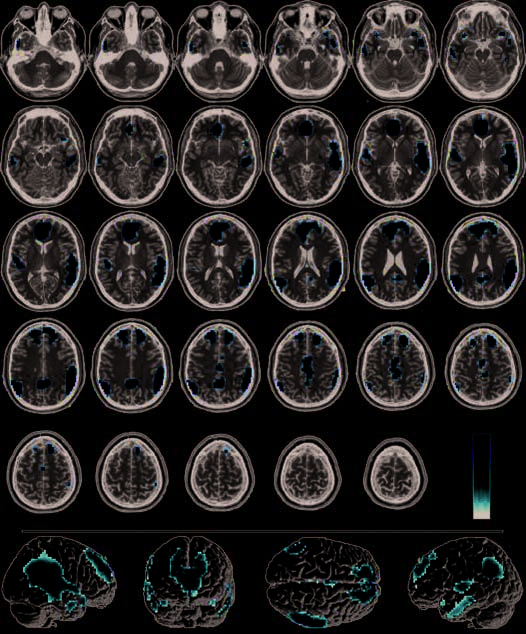
analysis of all disgust > neutro
conditions (pathogen +
incest + nonsexual moral >
neutro) overlaid on SPM2
canonical T1 brain. (FDR
corrected p < .001, cluster
minimum = 10 voxels,
neurological convention).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
9
e
-
1
p
5
d
2
f
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
c
9
n
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
9
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
t
.
/
o
n
1
8
M
a
y
2
0
2
1
1534
Journal of Cognitive Neuroscience
Volume 20, Number 9
Table 2. Conjunction Analysis of All Disgust > Neutral Contrasts
Conjunction: Pathogen + Incest + Nonsexual Moral vs. Neutro (FDR Corrected, P < .001) MNI Coordinates Brain Region Frontal Lobes BA x Medial/Superior frontal gyrus 10/9/(8, 6) Medial frontal gyrus Left middle frontal gyrus Precentral gyrus Temporal Lobes Bilateral temporal pole (into inferior frontal gyrus on left side) 11 6 6 38 47 38 Bilateral middle/superior temporal 21/22 gyrus (anterior and posterior regions) Parietal Lobes Postcentral gyrus Precuneus/post cingulate Occipital Lobes 4 3 7 31 Bilateral lingual gyrus/cuneus 17/18 Cingulate/Subcortical Anterior cingulate (dorsal) 24 Right putamen/globus pallidus Parahippocampal gyrus/thalamus (into left putamen/globus pallidus) Caudate Bilateral amygdala (cid:2)9 (cid:2)6 (cid:2)45 63 (cid:2)42 (cid:2)36 (cid:2)48 (cid:2)42 48 (cid:2)69 (cid:2)60 66 54 51 (cid:2)21 (cid:2)6 9 (cid:2)13 (cid:2)3 21 (cid:2)15 (cid:2)6 24 (cid:2)24 y 54 54 9 (cid:2)9 (cid:2)6 (cid:2)12 12 32 12 (cid:2)36 (cid:2)12 (cid:2)9 (cid:2)12 (cid:2)18 (cid:2)57 (cid:2)51 (cid:2)87 (cid:2)84 (cid:2)12 0 (cid:2)30 0 (cid:2)3 (cid:2)5 z Z t 36 (cid:2)15 54 42 60 51 (cid:2)42 (cid:2)14 (cid:2)42 0 (cid:2)12 (cid:2)6 54 60 72 33 0 0 39 (cid:2)12 (cid:2)3 12 (cid:2)15 (cid:2)15 (B)876 (K)19 (H)130 (C)480 (H)130 (H)130 (A)9017 (A)9017 (E)93 (M)2001 (M)2001 (D)146 (C)480 (C)480 ( J)18 (G)59 (A)9017 (A)9017 (I)57 (F)122 (N)59 (L)27 (F)122 (N)59 12.83 4.50 5.02 6.51 5.62 4.44 12.85 6.16 10.52 10.5 6.62 7.74 7.70 5.04 5.69 11.59 5.34 6.08 6.65 4.39 Brodmann’s area (BA), MNI coordinates, number of voxels (Z, letters indicate same cluster), and t value (t) of each cluster > 10 voxels are reported.
Schaich Borg, Lieberman, and Kiehl
1535
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
/
.
.
.
F
T
o
N
1
8
M
UN
sì
2
0
2
1
spanned from the middle into the superior temporal
gyrus, and extended rostrally from the most posterior
regions of BA 21/22 all the way to the left temporal pole
(BA 38). Isolated regions of the right posterior middle
temporal gyrus and temporal pole were also more
active, but not to the same degree or extent as the same
regions in the left hemisphere. In sum, a large collection
of brain areas is active during implicit processing of both
pathogen-related and sociomoral acts, supporting be-
havioral evidence that reactions toward disgusting ob-
jects feel subjectively similar to reactions toward
sociomoral transgressions.
Pathogen Disgust and Sociomoral Disgust Activate
Distinct Neural Regions
The conjunction analysis demonstrated that many brain
regions are commonly activated by pathogen and socio-
moral disgust. The following contrasts, Tuttavia, dem-
onstrate that pathogen and sociomoral disgust are not
represented identically in the brain.
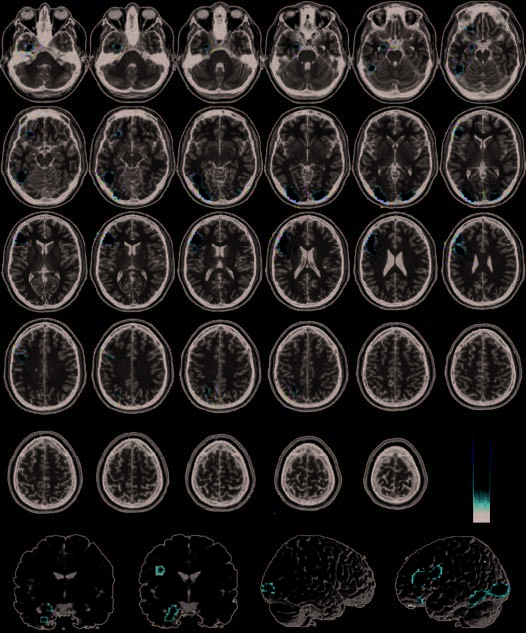
Pathogen > Sociomoral
This contrast (P > S–M) was performed to determine
what regions of the brain were uniquely active when
processing pathogen acts compared to sociomoral acts.
Activity in the left amygdala (Figura 4, Tavolo 3) was more
significant in response to pathogen acts than to socio-
moral acts. In the occipital lobes, the left fusiform gyrus
lingual gyrus (BA 18) were
(BA 37) and the bilateral
most active in response to pathogen acts as well. Nel
frontal lobes, a large cluster in the left inferior frontal
gyrus (BA 46) and a separate smaller cluster in the more
ventral region of the orbito-frontal cortex (BA 11) were
identified. Finalmente, a region of the precuneus (BA 7)
more rostral and dorsal to the region identified in the
conjunction analysis (BA 31) was uniquely associated
with the pathogen condition.
All of the brain regions identified in the P > S–M
contrast were preferentially active during processing of
disgusting acts without high levels of moral content. IL
following analysis determined what brain regions were
preferentially active during processing of disgusting acts
with high levels of moral content. It was predicted that
these brain regions should include the medial prefrontal
cortex and regions around the temporo-parietal junction,
two regions consistently shown to be involved in moral
processing (Schaich Borg et al., 2006; Heekeren et al.,
2003, 2005; Moll, Zahn, et al., 2005; Greene et al., 2004).
Sociomoral > Pathogen
Compared to pathogen and neutral acts, sociomoral acts
(incest and nonsexual moral) received very high ratings
of moral wrongness (Figura 2). We examined which
brain regions were more active during the unique pro-
cessing of sociomoral acts compared to that of path-
ogen acts (S–M > P). As predicted, significantly more
activity in large clusters of the medial prefrontal cortex
(BA 10, extending into the anterior cingulate) E
bilateral temporo-parietal junction was elicited by mor-
ally wrong acts than by acts rated not to be morally
wrong (Figura 5, Tavolo 4). Two other regions previously
identified in fMRI studies on moral processing, IL
temporal poles (BA 38) and the precuneus (BA 31),
were identified as well (Greene & Haidt, 2002). Areas of
the superior, middle, and inferior temporal gyri were
more active bilaterally in response to sociomoral acts
than to pathogen acts, as were bilateral regions of the
dorsolateral prefrontal cortex (BA 9) and a region of the
dorsal anterior cingulate (BA 24). These areas have also
previously been implicated in some types of moral
processing (but have not been identified in all morality
studies; Schaich Borg et al., 2006; Heekeren et al., 2003,
2005; Greene et al., 2004). The only brain region that
appeared in the S–M > P contrast that is not commonly
found in morality studies was the right posterior insula
(which is anatomically and functionally distinct from the
anterior insula mentioned earlier). In one study, how-
ever, this region was found to be more active in the left
hemisphere when participants read phrases that elicited
moral indignation than when they read neutral phrases
(Moll, de Oliveira-Souza, et al., 2005). Overall, Poi, IL
regions identified in the S–M > P contrast are consistent
with those identified previously to be involved in moral
processing, even though the present study uses a differ-
ent task than any other published study.
Sociomoral Disgust: A Unified Phenomenon?
The results of the S–M > P contrast illustrate that path-
ogen and sociomoral disgust are two separate (but re-
lated) physiological processes. The next experimental
question was whether sociomoral disgust, itself, is com-
posed of multiple distinct but related physiological pro-
cesses. We examined whether incest acts and nonsexual
moral acts entrain unique brain regions in addition to
their common brain networks by comparing directly the
results of an incest > neutral and nonsexual moral >
neutral contrast (see Methods), even though incest and
nonsexual moral acts are rated as equally morally wrong.
Incest > Nonsexual Moral
The incest manipulation was very powerful in this study
and the results of the incest > nonsexual moral (I > M)
contrast were highly statistically significant and extended
across many anatomical boundaries (Figura 6). IL
anatomical labels in Table 5 represent our best approx-
imation of the regions implicated in the I > M contrast,
including regions referenced by the reported coordi-
nates and the proximate brain regions the reported
clusters expanded into.
1536
Journal of Cognitive Neuroscience
Volume 20, Numero 9
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
T
.
.
F
/
o
N
1
8
M
UN
sì
2
0
2
1
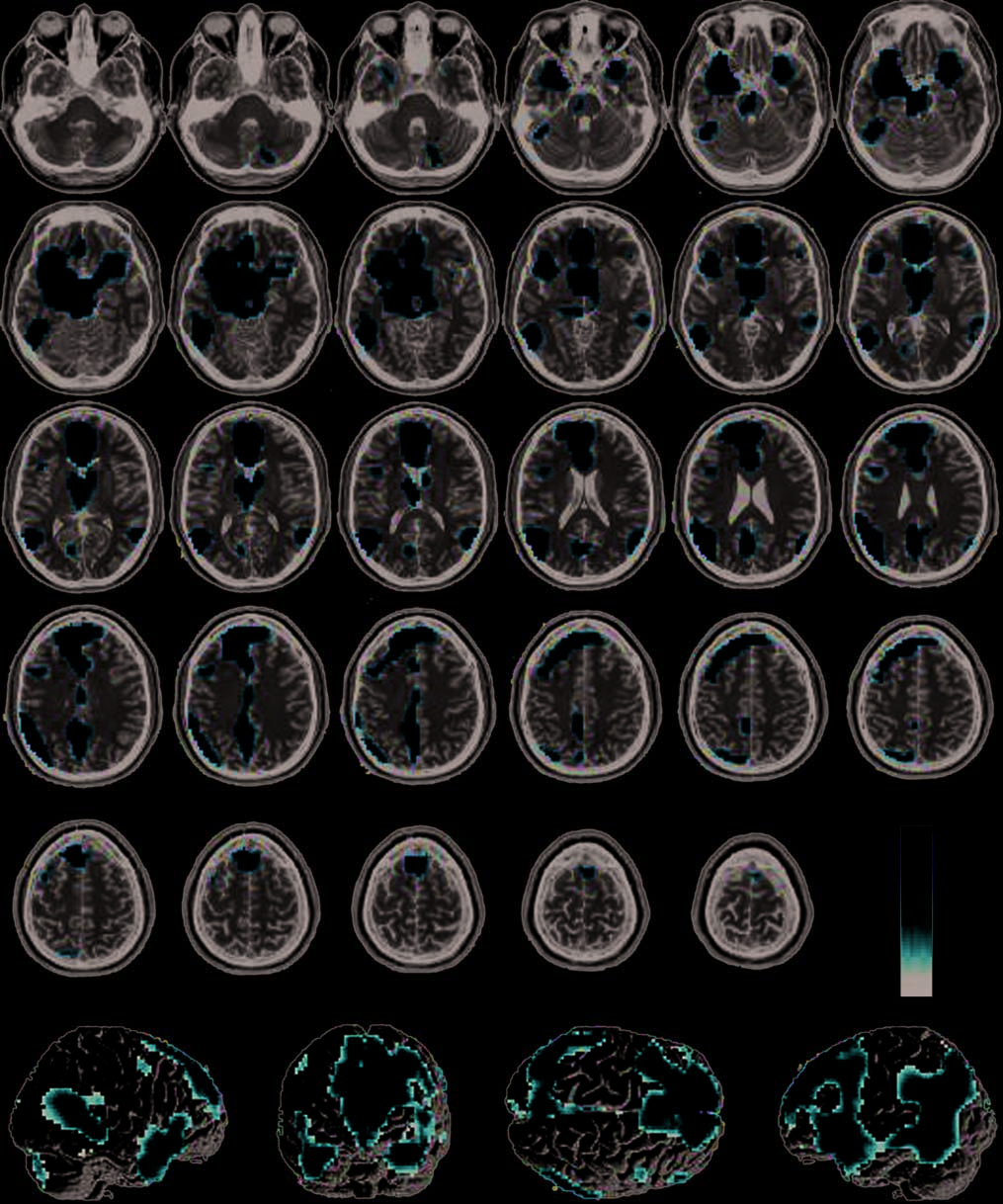
Figura 4. Regions more active
in pathogen condition than
in the sociomoral and neutral
conditions (pathogen >
sociomoral) overlaid on SPM2
canonical T1 image. (FDR
corrected p < .05, cluster
minimum = 10 voxels,
neurological convention).
Figure 5. Regions more active
in the sociomoral condition than
in the pathogen and neutral
conditions (sociomoral >
pathogen) overlaid on SPM2
canonical T1 image. (FDR
corrected p < .001, cluster
minimum = 10 voxels,
neurological convention).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
9
e
-
1
p
5
d
2
f
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
c
9
n
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
9
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
f
t
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Schaich Borg, Lieberman, and Kiehl
1537
Table 3. Regions More Activated in Pathogen Condition
Compared to Sociomoral (Incest and Nonsexual Moral) and
Neutral Conditions
Pathogen > Sociomoral (FDR Corrected, P < .05)
MNI Coordinates
Brain Region
BA
x
y
z
Z
t
Frontal Lobes
Left inferior frontal
gyrus (into the
dorsolateral
prefrontal cortex)
46
9
(cid:2)48
(cid:2)45
36
6
12
27
(A)444 6.39
(A)444 4.63
Left middle frontal
11
(cid:2)27
33 (cid:2)15 (B)35
4.61
gyrus
Parietal Lobes
Precuneus/Superior
parietal lobule
7
(cid:2)24 (cid:2)66
42 (C)80
4.38
Occipital Lobes
Left fusiform gyrus
Bilateral lingual
gyrus
37
18
Cingulate/Basal Ganglia
(cid:2)48 (cid:2)51 (cid:2)15 (D)446 5.42
(cid:2)27 (cid:2)99
(cid:2)3 (D)446 5.12
27 (cid:2)99
(cid:2)3
(E)132 4.30
Left amygdala
(cid:2)24
(cid:2)3 (cid:2)24 (F)45
3.56
Uncus
20/36 (cid:2)30
(cid:2)3 (cid:2)39 (G)27
3.58
Brodmann’s area (BA), MNI coordinates, number of voxels (Z, letters
indicate same cluster), and t value (t) of each cluster > 10 voxels are
reported.
The ventral bilateral anterior cingulate (BA 32, 24), ex-
tending into the medial and superior frontal gyri (BA 10
E 9, rispettivamente), and the bilateral inferior frontal gyri
(BA 47), extending into the anterior insulae (much less
so on the right side, BA 13), were more active during the
processing of statements describing incestuous acts than
nonsexual immoral acts (Figura 6, Tavolo 5). In the tem-
poral lobes, a small region of the right anterior superior
temporal gyrus, and larger regions around the temporo-
parietal junction (extending dorsally and posteriorly, stron-
ger on the left side than on the right) were identified in
the incest > nonsexual moral (I > M) contrasto. The left
fusiform gyrus was preferentially active in response to in-
cest actions, as was the dorsal anterior cingulate (BA 24),
the precuneus/posterior cingulate (BA 7/31), and the bi-
lateral amygdalae. Finalmente, medial areas of the basal ganglia,
the thalamus, and the midbrain were more active in re-
sponse to incest acts than to nonsexual moral acts.
It is common in fMRI studies with many participants
to see clusters of activation that expand across anatom-
ical boundaries, such as those identified in the I > M
contrasto, especially in deep regions of the brain (Vedere
Figura 2 or Figure 4 in Kiehl et al., 2005, n = 100,
for examples). Ancora, it may seem surprising that such
dramatic differences can be observed between two types
of moral stimuli that are rated as equally moral wrong.
To demonstrate the robustness of the results from the
I > M contrast, we plotted histograms of each subject’s
parameter estimates for each anatomical region re-
ported in Table 4 as taken from their derivative-boosted
I > M contrast map (Calhoun et al., 2004). These histo-
grams (see Supplementary Information) demonstrated
clearly that the data for the I > M group contrast follow
normal distributions and that many subjects have very
strong responses to the incest condition compared to
the nonsexual moral condition. These results suggest
strongly that the dramatic effects we see in the I > M
contrast are not due to methodological or mechanical
complications with data acquisition, and support the
interpretation that incest acts are indeed processed very
differently in the brain than nonsexual moral acts.
To identify the brain regions in the I > M contrast
that were either unique to the incest condition or just
more active in the incest than in the nonsexual moral
condition, the results of the I > M contrast were
compared to those of the nonsexual moral > neutral
(M > N) contrasto (not shown). Voxels active in only the
I > M and not the M > N contrast were interpreted as
unique to incest. The ventral bilateral anterior cingulate,
anterior insulae, right anterior superior temporal gyrus,
and medial basal ganglia, thalami and midbrain regions
identified in the I > M contrast were all uniquely
associated with the incest condition. The anterior cin-
gulate cluster extended into the medial and superior
frontal gyri and the left insula cluster extended into
the inferior frontal gyrus, but these regions were ac-
tive in the nonsexual moral condition as well, just to a
lesser extent than in the incest condition. The rest of
the brain regions identified in the I > M contrast,
including the left fusiform gyrus, the posterior cingulate,
the dorsal anterior cingulate, the extended bilateral
temporo-parietal junction, and the amygdalae, were all
active in the nonsexual moral condition as well as the
incest condition.
Nonsexual Moral > Incest
No voxels withstood multiple comparison corrections in
the nonsexual moral > incest (M > I) contrasto.
Pathogen, Incest, and Nonsexual Moral Acts Activate
Distinct Neural Regions
The results of the I > M and M > I contrasts show that
incestuous acts are processed differently in the brain
than nonsexual immoral acts. This raises the question, È
the pathogen condition distinct from both the incest
and the nonsexual moral conditions, or just from one of
1538
Journal of Cognitive Neuroscience
Volume 20, Numero 9
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
F
T
.
.
/
.
o
N
1
8
M
UN
sì
2
0
2
1
Tavolo 4. Regions More Activated in Sociomoral (Incest and Nonsexual Moral) Conditions Compared to Pathogen and Neutral
Conditions
Sociomoral > Pathogen (FDR Corrected, P < .001) MNI Coordinates BA x Brain Region Frontal Lobes Bilateral medial frontal gyrus (extends into bilateral anterior cingulate) Bilateral superior frontal gyrus (dorsolateral prefrontal cortex) Temporal Lobes Bilateral temporal pole Left inferior temporal gyrus Middle temporal gyrus Bilateral superior temporal gyrus (extends into transverse temporal gyrus) Bilateral supramarginal/angular gyrus, inferior parietal lobule (includes temporo-parietal junction) (right extends into middle temporal gyrus, postcentral gyrus, posterior insula) Occipital Lobes Bilateral precuneus Limbic System Anterior cingulate (dorsal) 10 32 9 38 20 21 22 41 39/40 21 2/3 13 31 24 y 54 42 51 18 21 (cid:2)18 3 3 (cid:2)12 (cid:2)30 (cid:2)12 (cid:2)60 (cid:2)51 (cid:2)27 (cid:2)9 (cid:2)6 (cid:2)18 (cid:2)36 42 (cid:2)63 (cid:2)51 54 (cid:2)54 (cid:2)51 57 (cid:2)57 57 54 (cid:2)6 6 (cid:2)51 (cid:2)51 0 (cid:2)6 z 6 0 36 (cid:2)21 (cid:2)21 (cid:2)24 (cid:2)39 (cid:2)27 6 15 3 36 33 (cid:2)6 33 33 48 Z t 6.62 6.02 5.11 4.19 4.3 5.23 5.1 4.65 4.76 3.77 4.94 6.55 6.24 4.9 (A)2137 (A)2137 (A)2137 (B)19 (C)48 (D)129 (D)129 (E)61 (F)191 (F)191 (G)2044 (H)434 (G)2044 (G)2044 (G)2044 (G)2044 (I)669 (I)669 4.77 4.68 (I)669 4.47 D o w n l o a d e d l l / / / / j t t f / i t . : / / f r o m D h o t w t n p o : a / d / e m d i f t r o p m r c h . s p i l d v i e r e r c c t . h m a i r e . d u c o o m c / n j a o r c t i n c / e a - p r d t i 2 c 0 l 9 e - 1 p 5 d 2 f 9 / 1 2 9 0 3 / 7 9 0 / 5 1 2 5 o 2 c 9 n / 1 2 0 7 0 5 8 9 6 2 5 0 5 1 0 / 9 j o p c d n . b y 2 0 g 0 u 8 e . s t 2 o 0 n 1 0 0 9 7 . S p e d p f e m b y b e g r u 2 0 e 2 s 3 t / j t . / . . f Brodmann’s area (BA), MNI coordinates, number of voxels (Z, letters indicate same cluster), and t value (t) of each cluster > 10 voxels are reported.
the conditions? The answer is that the pathogen condi-
tion is unique. Although the incest and pathogen acts
were rated as equally disgusting, the incest > pathogen
contrast yielded a large network of brain areas very
similar to those identified in the sociomoral > pathogen
contrasto. Additional significant voxels were identified in
the lateral temporal poles (lt = (cid:2)33, 18, (cid:2)28; rt = 36,
18, (cid:2)27), the right inferior frontal gyrus extending into
the anterior insula (39, 21, (cid:2)21), bilateral middle tem-
poral gyrus (lt = (cid:2)61, (cid:2)27, (cid:2)16; rt = 63, (cid:2)36, 0), E
superior temporal gyrus (lt = (cid:2)54, (cid:2)18, 6; rt = 63, (cid:2)18,
3), and the right supramarginal gyrus (60, (cid:2)54, 27)
extending into inferior regions all around the temporo-
parietal junction. Fewer significant voxels were found in
the basal ganglia, and unlike the sociomoral > pathogen
contrasto, no voxels were found in the brainstem and left
fusiform gyrus. The reverse pathogen > incest contrast
did not yield any significant results, probably because
the incest manipulation was so powerful. Tuttavia, IL
pathogen > nonsexual moral contrast yielded results
very similar to the P > S–M contrast with added
significant activity in both amygdalae (lt amygdala =
24, (cid:2)3, (cid:2)24; rt amygdala = 23, (cid:2)1, (cid:2)21), brainstem (lt
brainstem = (cid:2)6, (cid:2)29, (cid:2)15; rt brainstem = 3, (cid:2)27, (cid:2)18),
o
N
1
8
M
UN
sì
2
0
2
1
Schaich Borg, Lieberman, and Kiehl
1539
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
/
.
.
.
F
T
o
N
1
8
M
UN
sì
2
0
2
1
Figura 6. Regions more active in the incest condition than in the nonsexual moral condition (incest > nonsexual moral) overlaid on SPM2
canonical T1 image. (FDR corrected p < .0001, cluster minimum = 10 voxels, neurological convention).
1540
Journal of Cognitive Neuroscience
Volume 20, Number 9
Table 5. Regions More Activated in Incest Condition Compared to the Nonsexual Moral Condition
Incest > Nonsexual Moral (FDR Corrected, P < .0001)
MNI Coordinates
Brain Region
Frontal Lobes
Medial frontal gyrus, superior frontal gyrus
Bilateral inferior frontal gyrus (into insula on
left side)
Temporal Lobes
Bilateral angular gyrus
(includes bilateral temporo-parietal junction)
(extends into superior/middle temporal gyri)
Parietal Lobes
Left supramarginal gyrus
Precuneus/posterior cingulate
Occipital Lobes
Left fusiform gyrus
Limbic System/Basal Ganglia
Anterior cingulate (ventral)
Anterior cingulate (dorsal)
Left amygdala
Left hippocampus
Bilateral caudate/globus pallidus/putamen
Bilateral thalamus
BA
x
10/9
47
13
39
22
40
7/31
(cid:2)9
27
(cid:2)27
(cid:2)51
57
63
(cid:2)57
(cid:2)6
y
54
21
21
(cid:2)60
(cid:2)66
(cid:2)57
(cid:2)42
(cid:2)54
z
Z
t
9
(cid:2)21
(cid:2)21
18
24
15
30
36
(A)4499
(A)4499
(A)4499
(B)820
(C)73
(C)73
(D)11
(E)291
9.67
7.28
7.28
6.82
5.13
5.36
4.89
5.73
37
(cid:2)45
(cid:2)48
(cid:2)18
(F)820
6.15
32/24
24
0
0
36
(cid:2)15
3
36
9.43
5.15
(A)4499
(G)22
(A)4499
(A)4499
(A)4499
(A)4499
Brodmann’s area (BA), MNI coordinates, number of voxels (Z, letters indicate same cluster), and t value (t) of each cluster > 10 voxels are reported.
and ventromedial basal ganglia (lt = (cid:2)6, 0, (cid:2)9; rt = 6,
(cid:2)2, (cid:2)6), and slightly less significant activity in the visual
cortex (see Supplementary Information for figures). IL
reverse nonsexual moral > pathogen contrast did not
yield any significant voxels. In sum, pathogen acts invoke
more activity in many brain regions than do nonsexual
moral acts, and incest acts invoke more activity in many
brain regions than do either pathogen or nonsexual
moral acts, despite the fact that incest acts are equally
disgusting as pathogen acts and equally immoral as
nonsexual moral acts.
DISCUSSION
This study is the first investigation of the common and
unique neural correlates of three separate domains of
disgust. The present results provide evidence that: (io)
common brain regions are active during processing of
both pathogen-related and sociomoral acts, (ii) pathogen-
related and sociomoral acts each entrain unique brain
regions, (iii) despite their tendency to elicit similar rat-
ings of moral disapproval, incest-related acts and non-
sexual immoral acts entrain different, but overlapping,
brain networks, E (iv) despite their tendency to elicit
similar ratings of disgust, pathogen-related acts and incest-
related acts entrain different, but again, overlapping brain
networks.
These findings call attention to a number of issues.
Primo, some researchers argue that the insula and amyg-
dala have dissociable, nonoverlapping functions with the
insula serving as the seat of disgust processing and the
amygdala the seat of fear processing (Phillips et al., 1998,
2004; Calder, 2003). The data reported here challenge
this view. The insula was only preferentially active in
Schaich Borg, Lieberman, and Kiehl
1541
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
F
.
T
.
.
/
o
N
1
8
M
UN
sì
2
0
2
1
response to incest acts (not pathogen or nonsexual
moral acts), whereas the amygdala was more active in
response to all three types of disgusting acts than to
neutral acts. A large area of the left inferior frontal
gyrus (BA 47) was identified in the conjunction analysis,
but it did not extend into the anterior insular cortex dis-
cussed in previous studies of disgust (Schafer et al., 2005;
Wright, Lui, Shapira, Goodman, & Liu, 2004; Krolak-
Salmon et al., 2003; Shapira et al., 2003; Phillips et al.,
1997, 1998). Although these results may seem surprising
to some, a growing number of studies have shown that
the insula and the amygdala are often, but not always,
involved in disgust processing (Schafer et al., 2005; Stark
et al., 2003, 2005; Phillips et al., 2004; Schienle et al.,
2002). This is consistent with a recent meta-analysis of
imaging studies that found the anterior insula to be no
more active during disgust than other emotions (Feldman
Barrett & Wager, 2007).
Nonetheless, it is possible that the lack of insula ac-
tivity in our conjunction analysis was a result of induced
tonic insula activity in the neutral condition due to
carryover effects from our highly provocative stimuli.
Exploratory analyses suggest that the insula was, Infatti,
active during the neutral condition, but not as a result of
carryover effects (see Supplementary Information). In-
sula activity was present bilaterally when the neutral
condition was compared to baseline. This activity re-
mained when the analysis was restricted to just the first
blocks of neutral stimuli, a result that is immune to
carryover effects because the first blocks of neutral
stimuli were the first acts presented to subjects. These
data suggest that the lack of insular activity in our
conjunction analyses was most likely due to an equal
presence of insular activity in the neutral condition as in
the disgust conditions, and not due to a lack of insula
activity in any condition.
If this interpretation is correct, why would the insula
respond to neutral stimuli? One possibility is that the
insula is involved in language processing (Ogar et al.,
2006; Nestor et al., 2003) and may have a second func-
tion in mapping visceral states associated with con-
scious, personal emotional experience (Damasio, 2003).
Our stimuli were short phrases describing acts in the sec-
ond person, so they required language processing and
likely elicited imagery of performing the acts described. If
the insula’s role in language or in self-monitoring trumps
its role in processing information associated with disgust,
perhaps the neutral acts activated the insula as much as
the disgusting acts because all the acts elicited the same
amount of language processing and imaginative intro-
spection. In support of this explanation, the only other
study using written acts rather than faces or pictures as
disgust stimuli did not find the anterior insula to be more
active in disgusting versus neutral statements either
(Moll, de Oliveira-Souza, et al., 2005). Inoltre, pa-
tients with Huntington’s disease, a clinical population
believed to have decreased insular volume compared to
healthy individuals (Kassubek et al., 2004), have been
shown to have intact disgust recognition in response to
linguistic stimuli, but impaired disgust recognition in
response to both facial and nonfacial visual stimuli (Hayes
et al., 2007). Therefore, the verbal nature of our stimuli
may provide the most plausible explanation for the
equivalent insula activity across all our conditions.
Alternatively, perhaps the task demands rather than
the stimuli modulated insula activity. The insula (and the
amygdala) results reported here may be due to the fact
that participants were engaged in a memory task in this
study and were not required to explicitly evaluate how
disgusting each act was until after the scanning session.
A previous study demonstrated that the amygdala—not
the insula—was preferentially active in response to
unattended pictures of disgusted faces while the insula
was only preferentially active in response to attended
pictures of disgusted faces (Anderson, Christoff, Panitz,
De Rosa, & Gabrieli, 2003). Inoltre, although the
insula is active in response to consciously perceived
disgusted facial expressions, it is not preferentially active
in response to subliminally presented disgusted facial
expressions (Phillips et al., 2004). Così, perhaps the
absence of insula activity in our conjunction analysis is
due to the fact that participants were too preoccupied
with our implicit task to actively perceive the disgusting
nature of the stimuli. Tuttavia, this interpretation is
unlikely because participants’ voluntary comments after
scanning suggested that participants actively detected
the stimuli’s disgusting nature.
One additional possibility is that the anterior insula is
simply not more involved in disgust processing than any
other type of emotional processing (Feldman Barrett &
Wager, 2007). Perhaps the insula activity identified in
past disgust studies represents emotion-related activity
in general rather than disgust-specific activity, and the
neutral stimuli used in the present study were well
matched for overall emotional engagement. Until more
research evaluates these proposed interpretations and
explanations, the present data support the view that the
insula is not always preferentially involved in disgust
processing, and the amygdala, a structure commonly as-
sociated with fear processing, can also be preferentially
involved in disgusting processing (see also Kiehl et al.,
2005 on amygdala activation to salient, not just fear-
inducing, stimuli).
A second issue raised by the results of this study is the
powerful nature of the incest manipulation. The incest
condition activated almost the entire brain (see Supple-
mentary Information), making it difficult to detect any
significant effects in either the moral > incest or the
pathogen > incest contrasts. One account could be that
the unique neural correlates of the incest condition are
due mostly to the fact that the incest acts are more
emotionally arousing (emotionally intense) than other
acts. A separate group of healthy volunteers (n = 33)
rated a subset of the incest acts to be slightly more
1542
Journal of Cognitive Neuroscience
Volume 20, Numero 9
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
/
F
.
.
.
T
o
N
1
8
M
UN
sì
2
0
2
1
arousing than a similar subset of the pathogen ( P < .01),
nonsexual moral ( p < .001), and neutral acts ( p < .001).
Still, it unlikely that the magnitude and pervasiveness of
the incest condition’s neural effects are due solely to
increased arousal because the hemodynamic signature
of the incest condition does not mimic typical arousal
effects observed in other fMRI studies. In particular, the
amygdala has been shown to be correlated specifically
with arousal (Lewis, Critchley, Rotshtein, & Dolan, 2007;
Cunningham, Raye, & Johnson, 2004), and the amygdala
was not significantly more active while processing incest
acts compared to pathogen acts. Moreover, the other
regions identified in the I > M contrast have not been
shown to be correlated with arousal ratings. It is also
useful to note that pathogen and nonsexual moral acts
were rated as equally arousing ( p = .22), suggesting that
the amygdala activation in the P > M contrast cannot be
due to differences in arousal. Nevertheless, if part of the
neural signature of the incest condition was due to an
arousal
interaction, the propensity of incest acts to
arouse more than nonsexual moral acts is a surprising
inherent difference between these two similarly immoral
conditions that may have implications for the design of
moral stimuli in future studies.
Another possible explanation for the pattern of activ-
ity in the incest condition is that our stimuli were written
in the second rather than third person. Adaptation-
minded researchers investigating the levels of disgust
associated with incestuous acts have encountered ceil-
ing effects when asking individuals to imagine engaging
in sexual acts with a family member compared to
imagining third-party incestuous acts (Lieberman et al.,
2007; Fessler & Navarrete, 2004). Perhaps incest stimuli
written in the third person would not have elicited such
dramatic neural activity compared to nonsexual moral
and pathogen stimuli.
As a third issue, data presented here raise the possi-
bility that the typical operationalization of ‘‘disgust’’ and
‘‘morality’’ in experimental investigations may require
revision. Neuroscientific investigations often treat disgust
as a unified psychological and neurological construct by
using disgusted faces or pictures to represent all domains
of disgust (Phillips et al., 1998, 2004; Calder, 2003). How-
ever, the data presented here show that although incest
acts and pathogen acts are rated as equally disgusting,
incest acts are rated as more immoral and elicit dramat-
ically more hemodynamic activity than pathogen acts.
These data, combined with data from other recent multi-
modal disgust studies, suggest that disgust is likely best
conceived of as a set of heterogeneous responses over-
laying a unified psychological and neurological response,
and therefore, most likely will not be fully understood
through studying only one sensory modality or one type
of disgusting stimulus.
Allo stesso modo, neuroscientific investigations have often
treated morality as a unified psychological and neuro-
logical construct (per esempio., Heekeren et al., 2003; Moll et al.,
2002), heterogeneous only insomuch as it invokes differ-
ent proportions of activity in common ‘‘emotional’’ versus
‘‘cognitive’’ systems (Schaich Borg et al., 2006; Greene
et al., 2004). The field has benefited profoundly from
these studies because they have identified a set of brain
regions including the medial prefrontal cortex and areas
around the temporo-parietal junction that is reliably
more active during processing of a broad range of moral
stimuli compared to nonmoral stimuli. These regions
appeared robustly in the results of the sociomoral >
pathogen contrast in the data presented here as well.
Together, previous and present results support the
hypothesis that the medial prefrontal cortex and areas
around the temporo-parietal junction are likely to be
involved in general moral processing across many dif-
ferent situations. Tuttavia, the results reported here
also illustrate that despite the fact that incest acts and
nonsexual moral acts were rated as equally immoral,
incest acts elicit dramatically more activity in an ex-
tensive expanse of brain regions ranging from the me-
dial prefrontal cortex itself to the basal ganglia. Questo
additional pattern of results suggests that it is impor-
tant to acknowledge the possibility that not all moral-
ity or immorality is necessarily ‘‘treated equally’’ by the
brain.
This possibility is starting to be recognized by neuro-
scientists interested in morality. One study has exam-
ined whether the presence or absence of direct bodily
harm affects the brain networks involved in moral
decision making (Heekeren et al., 2005). Tuttavia, IL
moral stimuli that included direct bodily harm were
rated as significantly more morally wrong than moral
stimuli without bodily harm, so it is not clear whether
the neurological differences identified in that study were
due to sentiments of moral wrongness or to systems
specific for detecting bodily harm. Recentemente, it was shown
that neural activity associated with processing justice-
based moral transgressions differs from that associated
with processing care-based moral transgressions. How-
ever, this study did not test whether justice- and care-
based moral transgressions are rated as equally morally
wrong, so again, it is not clear whether the differences
they report can be explained by sentiments of moral
wrongness (Robertson et al., 2007). The data reported
here, Poi, are the first to show that equally immoral
acts can be represented differently in the brain.
Heterogeneity of moral stimuli, such as that between
likely ex-
sexual and nonsexual moral transgressions,
tends to functional domains beyond those explored in
this study, especially those that require specific types of
information processing such as theory of mind or cheater-
detection (Young, Cushman, Hauser, & Saxe, 2007;
Ermer, Guerin, Cosmides, Tooby, & Mugnaio, 2006). Noi
suspect that future studies examining explicit moral
judgments made in the scanner, as opposed to after
scanning as investigated here, will evidence similar
heterogeneity as well. Although the data we provide
Schaich Borg, Lieberman, and Kiehl
1543
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
T
F
.
/
.
.
o
N
1
8
M
UN
sì
2
0
2
1
here do not speak to whether brain regions differentially
involved in moral processing are necessary for that
moral processing, they do suggest that future morality
studies might opt to use stimuli that are distributed
across the different moral domains in a controlled
maniera, and interpret data in light of the possibility
that some reported results may be specific to only some
moral domains. In other words, the results of reported
moral > nonmoral contrasts will differ depending on the
types of moral transgressions used as stimuli.
Appreciating the functional heterogeneity within dis-
gust and morality and learning more about the common-
alities and differences of neural systems that process
distinct disgusting stimuli or moral transgressions could
lead to better treatment and diagnosis of clinical pop-
ulations whose symptoms may reflect impairments in
specific moral and/or disgust domains, such as sexual
offenders, psychopaths, drug-abusers, Huntington’s dis-
ease patients, and obsessive–compulsive disorder pa-
tients. Within psychology and cognitive neuroscience,
appreciating disgusts’ heterogeneity may help us uncov-
er the true neural building blocks of basic emotions.
Allo stesso modo, exploring if and how different types of moral
transgressions can invoke different brain networks will
become increasingly important as neuroscientific data
become more influential in legal and ethical decisions. If
lawyers and ethicists continue to debate whether lesion
patients or psychiatric patients with functional deficits
should be considered culpable for their immoral actions
(Mobbs, Lau, Jones, & Frith, 2007), it will be helpful to
acknowledge that some brain regions might be involved
in only specific subsets of moral processing because
patients could conceivably be held culpable for some
types of immoral actions but not for others. Although
there is still much to explore, the data reported here lay
the groundwork for many future interdisciplinary in-
vestigations which promise to advance our knowledge
about the structure of various psychopathologies, IL
nature of disgust, and our own moral behavior.
Reprint requests should be sent to Kent A. Kiehl, The Mind
Institute, 1101 Yale Boulevard NE, MSC11 6040, Albuquerque,
NM 87106, or via e-mail: kkiehl@unm.edu.
REFERENCES
Anderson, UN. K., Christoff, K., Panitz, D., De Rosa, E., &
Gabrieli, J. D. E. (2003). Neural correlates of the automatic
processing of threat facial signals. Journal of Neuroscience,
23, 5627–5633.
Blair, J., Marsh, UN. A., Finger, E., Blair, K. S., & Luo, J. (2006).
Neuro-cognitive systems involved in morality. Philosophical
Explorations, 9, 13–27.
Bloom, P. (2004). Descartes’ baby: How the science of child
development explains what makes us human. New York:
Basic Books.
Calder, UN. J. (2003). Disgust discussed. Annals of Neurology,
53, 427–428.
Calder, UN. J., Keane, J., Manes, F., Antoun, N., & Young, UN. W.
(2000). Impaired recognition and experience of disgust
following brain injury. Nature Neuroscience, 3, 1077–1078.
Calhoun, V. D., Stevens, M. S., Pearlson, G. D., & Kiehl, K. UN.
(2004). fMRI analysis with the General Linear Model:
Removal of latency-induced amplitude bias by incorporation
of hemodynamic derivative terms. Neuroimage, 22,
252–257.
Cunningham, W. A., Raye, C. L., & Johnson, M. K. (2004).
Implicit and explicit evaluation: fMRI correlates of valence,
emotional intensity, and control in the processing of
attitudes. Journal of Cognitive Neuroscience, 16,
1717–1729.
Curtis, V., Aunger, R., & Rabie, T. (2004). Evidence that disgust
evolved to protect from risk of disease. Proceedings of
the Royal Society of London, Series B, Biological Sciences,
271, s131–s133.
Curtis, V., & Biran, UN. (2001). Dirt, disgust, and disease. Is
hygiene in our genes? Perspectives in Biology and Medicine,
44, 17–31.
Damasio, UN. (2003). Feelings of emotion and the self. Annals of
the New York Academy of Sciences, 1001, 253–261.
Ermer, E., Guerin, S., Cosmides, L., Tooby, J., & Mugnaio, M.
(2006). Theory of mind broad and narrow: Reasoning
about social exchange engages ToM areas, precautionary
reasoning does not. Social Neuroscience, 1, 196–219.
Feldman Barrett, L., & Wager, T. (2007). The neural reference
space for emotion: New meta-analytic insights. Paper
presented at the Neural Systems of Social Behavior, Austin,
TX, 2007.
Fessler, D. M. T., & Navarrete, C. D. (2003). Domain-specific
variation in disgust sensitivity across the menstrual cycle.
Evolution and Human Behavior, 24, 406–417.
Fessler, D. M. T., & Navarrete, C. D. (2004). Third-party
attitudes toward sibling incest: Evidence for Westermarck’s
hypotheses. Evolution and Human Behavior, 25,
277–294.
Fitzgerald, D. A., Posse, S., Moore, G. J., Tancer, M. E.,
Nathan, P. J., & Phan, K. l. (2004). Neural correlates of
internally-generated disgust via autobiographical recall: UN
functional magnetic resonance imaging investigation.
Neuroscience Letters, 370, 91–96.
Freire, L., Roche, A., & Mangin, J.-F. (2002). What is the
best similarity measure for motion correction in fMRI time
series? IEEE Transactions on Medical Imaging, 21,
470–484.
Friston, K. J., Ashburner, J., Frith, C. D., Poline, J.-B., Heather,
J. D., & Frackowiak, R. S. J. (1995). Spatial registration
and normalization of images. Human Brain Mapping, 3,
165–189.
Greene, J., & Haidt, J. (2002). How (and where) does moral
judgment work? Trends in Cognitive Sciences, 6, 517–523.
Greene, J. D., Nystrom, l. E., Engell, UN. D., Darley, J. M., &
Cohen, J. D. (2004). The neural bases of cognitive conflict
and control in moral judgment. Neuron, 44, 389–400.
Haidt, J., McCauley, C., & Rozin, P. (1994). Individual
differences in sensitivity to disgust: A scale sampling seven
domains of disgust elicitors. Personality and Individual
Differences, 16, 701–713.
Haidt, J., Rozin, P., McCauley, C., & Imada, S. (1997). Corpo,
psyche, e cultura: The relationship between disgust
and morality. Psychology and Developing Societies, 9,
107–131.
Hayes, C. J., Stevenson, R. J., & Coltheart, M. (2007).
Disgust and Huntington’s disease. Neuropsychologia,
45, 1135–1151.
Heekeren, H. R., Wartenburger, I., Schmidt, H., Prehn, K.,
Schwintowski, H. P., & Villringer, UN. (2005). Influence of
1544
Journal of Cognitive Neuroscience
Volume 20, Numero 9
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
F
.
.
/
T
o
N
1
8
M
UN
sì
2
0
2
1
bodily harm on neural correlates of semantic and moral
decision-making. Neuroimage, 24, 887–897.
Heekeren, H. R., Wartenburger, I., Schmidt, H., Schwintowski,
H. P., & Villringer, UN. (2003). An fMRI study of simple ethical
decision-making. NeuroReport, 14, 1215–1219.
Kahan, D. M. (1998). ‘‘The anatomy of disgust’’ in criminal law.
Michigan Law Review, 96, 1621–1657.
Kassubek, J., Juengling, F. D., Kioschies, T., Henkel, K.,
Karitzky, J., Kramer, B., et al. (2004). Topography of cerebral
atrophy in early Huntington’s disease: A voxel based
morphometric MRI study. Journal of Neurology,
Neurosurgery, and Psychiatry, 75, 213–220.
Kiehl, K. A., Stevens, M. C., Laurens, K. R., Pearlson, G.,
Calhoun, V. D., & Liddle, P. F. (2005). An adaptive
reflexive processing model of neurocognitive function:
Supporting evidence from a large scale (n = 100) fMRI
study of an auditory oddball task. Neuroimage, 25,
899–915.
Koenigs, M., Young, L., Adolphs, R., Tranel, D., Cushman, F.,
Hauser, M., et al. (2007). Damage to the prefrontal
cortex increases utilitarian moral judgements. Nature, 446,
908–911.
Krolak-Salmon, P., Henaff, M.-A., Isnard, J., Tallon-Baudry, C.,
Guenot, M., Vighetto, A., et al. (2003). An attention
modulated response to disgust in human ventral anterior
insula. Annals of Neurology, 53, 446–453.
Lewis, P., Critchley, H., Rotshtein, P., & Dolan, R. (2007).
Neural correlates of processing valence and arousal in
affective words. Cerebral Cortex, 17, 742–748.
Lieberman, D., Tooby, J., & Cosmides, l. (2003). Does morality
have a biological basis? An empirical test of the factors
governing moral sentiments related to incest. Proceedings of
the Royal Society of London, Series B, Biological Sciences,
270, 819–826.
Lieberman, D., Tooby, J., & Cosmides, l. (2007). IL
architecture of human kin detection. Nature, 445, 727–731.
Mugnaio, W. IO. (1997). The anatomy of disgust. Cambridge, MA:
Stampa dell'Università di Harvard.
Mobbs, D., Lau, H. C., Jones, O. D., & Frith, C. D. (2007). Legge,
responsibility, and the brain. PLoS Biology, 5.
Moll, J., de Oliveira-Souza, R., Eslinger, P. J., Bramati, IO. E.,
Mourao-Miranda, J., Andreiuolo, P. A., et al. (2002).
The neural correlates of moral sensitivity: A functional
magnetic resonance imaging investigation of basic
and moral emotions. Journal of Neuroscience, 22,
2730–2736.
Moll, J., de Oliveira-Souza, R., Moll, F. T., Ignacio, F. A., Bramati,
IO. E., Caparelli-Daquer, E. M., et al. (2005). The moral
affiliations of disgust: A functional MRI study. Cognitive and
Behavioral Neurology, 18, 68–78.
Moll, J., Zahn, R., de Oliveira-Souza, R., Krueger, F., & Grafman,
J. (2005). The neural basis of human moral cognition.
Nature Reviews Neuroscience, 6, 799–809.
Nestor, P. J., Graham, N. L., Fryer, T. D., Williams, G. B.,
Patterson, K., & Hodges, J. R. (2003). Progressive non-fluent
aphasia is associated with hypometabolism centred on the
left anterior insula. Brain, 126, 2406–2418.
Nichols, S. (2002). Norms with feeling: Towards a
psychological account of moral judgment. Cognition, 84,
221–236.
Nichols, T., Brett, M., Andersson, J., Wager, T., & Poline, J.-B.
(2005). Valid conjunction inference with the minimum
statistic. Neuroimage, 25, 653–660.
Ogar, J., Willock, S., Baldo, J., Wilkins, D., Ludy, C., & Dronkers,
N. (2006). Clinical and anatomical correlates of apraxia of
speech. Brain and Language, 97, 343–350.
Phan, K. L., Wager, T., Taylor, S. F., & Liberzon, IO. (2002).
Functional neuroanatomy of emotion: A meta-analysis of
emotion activation studies in PET and fMRI. Neuroimage,
16, 331–348.
Phillips, M. L., Williams, l. M., Heining, M., Herba, C. M.,
Russell, T., Andrew, C., et al. (2004). Differential neural
responses to overt and covert presentations of facial
expressions of fear and disgust. Neuroimage, 21, 1484–1496.
Phillips, M. L., Young, UN. W., Scott, S. K., Calder, UN. J., Andrew,
C., Giampietro, V., et al. (1998). Neural responses to facial
and vocal expressions of fear and disgust. Atti del
Royal Society of London, Series B, Biological Sciences, 265,
1809–1817.
Phillips, M. L., Young, UN. W., Senior, C., Brammer, M., Andrew,
C., Calder, UN. J., et al. (1997). A specific neural substrate
for perceiving facial expressions of disgust. Nature, 389,
495–498.
Robertson, D., Snarey, J., Ousley, O., Harenski, K., Bowman, F.,
Gilkey, R., et al. (2007). The neural processing of moral
sensitivity to issues of justice and care. Neuropsychologia,
45, 755–766.
Rozin, P., & Fallon, UN. E. (1987). A perspective on disgust.
Psychological Review, 94, 23–41.
Sambataro, F., Dimalta, S., Di Giorgio, A., Taurisano, P., Blasi,
G., Scarabino, T., et al. (2006). Preferential responses
in amygdala and insula during presentation of facial
contempt and disgust. European Journal of Neuroscience,
24, 2355–2362.
Schafer, A., Schienle, A., & Vaitl, D. (2005). Stimulus type and
design influence hemodynamic responses towards visual
disgust and fear elicitors. International Journal of
Psychophysiology, 57, 53–59.
Schaich Borg, J., Hynes, C., Van Horn, J., Grafton, S., &
Sinnott-Armstrong, W. (2006). Consequences, action,
and intention as factors in moral judgments: An fMRI
investigation. Journal of Cognitive Neuroscience, 18,
803–817.
Scherer, K. R. (1997). The role of culture in emotion-antecedent
appraisal. Journal of Personality and Social Psychology, 73,
902–922.
Schienle, A., Schafer, A., Hermann, A., Walter, B., Stark, R., &
Vaitl, D. (2006). fMRI responses to pictures of mutilation
and contamination. Neuroscience Letters, 393, 174–178.
Schienle, A., Schafer, A., Stark, R., Walter, B., Franz, M., & Vaitl,
D. (2003). Disgust sensitivity in psychiatric disorders: UN
questionnaire study. Journal of Nervous and Mental
Disease, 191, 831–834.
Schienle, A., Stark, R., Walter, B., Blecker, C., Ott, U., Kirsch, P.,
et al. (2002). The insula is not specifically involved in disgust
processing: An fMRI study. NeuroReport, 13, 2023–2026.
Shapira, N. A., Liu, Y., Lui, UN. G., Bradley, M. M., Lessig,
M. C., James, G. A., et al. (2003). Brain activation by
disgust-inducing pictures in obsessive–compulsive
disorder. Biological Psychiatry, 54, 751–756.
Simpson, J., Carter, S., Anthony, S. H., & Overton, P. G. (2006).
Is disgust a homogeneous emotion? Motivation and
Emotion, 30, 31–41.
Sprengelmeyer, R., Young, UN. W., Calder, UN. J., Karnat, A.,
Lange, H., Homberg, V., et al. (1996). Loss of disgust:
Perception of faces and emotions in Huntington’s disease.
Brain, 119, 1647–1665.
Stark, R., Schienle, A., Sarlo, M., Palomba, D., Walter, B., &
Vaitl, D. (2005). Influences of disgust sensitivity on
hemodynamic responses towards a disgust-inducing
film clip. International Journal of Psychophysiology, 57,
61–67.
Stark, R., Schienle, A., Walter, B., Kirsch, P., Sammer, G., Ott,
U., et al. (2003). Hemodynamic responses to fear and
disgust-inducing pictures: An fMRI study. Internazionale
Journal of Psychophysiology, 50, 225–234.
Schaich Borg, Lieberman, and Kiehl
1545
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
/
F
.
.
.
T
o
N
1
8
M
UN
sì
2
0
2
1
Suzuki, A., Hoshino, T., Shigemasu, K., & Kawamura, M.
Wright, P., Lui, G., Shapira, N., Goodman, W., & Liu, Y.
(2006). Disgust-specific impairment of facial expression
recognition in Parkinson’s disease. Brain, 129, 707–717.
Wheatley, T., & Haidt, J. (2005). Hypnotic disgust makes
moral judgments more severe. Psychological Science, 16,
780–784.
Wicker, B., Keysers, C., Plailly, J., Royet, J.-P., Gallese, V., &
Rizzolatti, G. (2003). Both of us disgusted in my insula: IL
common neural basis of seeing and feeling disgust. Neuron,
40, 655–664.
Williams, l. M. C. A., Das, P., Liddell, B., Olivieri, G., Peduto, A.,
Brammer, M. J., et al. (2005). BOLD, sweat and fears: fMRI
and skin conductance distinguish facial fear signals.
NeuroReport, 16, 49–52.
(2004). Disgust and the insula: fMRI responses to pictures
of mutilation and contamination. NeuroReport, 15,
2347–2351.
Young, L., Cushman, F., Hauser, M., & Saxe, R. (2007). IL
neural basis of the interaction between theory of mind and
moral judgment. Proceedings of the National Academy of
Scienze, U.S.A., 104, 8235–8240.
Zald, D. H. (2003). The human amygdala and the emotional
evaluation of sensory stimuli. Brain Research Reviews, 41,
88–123.
Zhong, C., & Liljenquist, K. (2006). Washing away your sins:
Threatened morality and physical cleansing. Scienza, 313,
1451–1452.
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
9
e
–
1
P
5
D
2
F
9
/
1
2
9
0
3
/
7
9
0
/
5
1
2
5
o
2
C
9
N
/
1
2
0
7
0
5
8
9
6
2
5
0
5
1
0
/
9
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
8
e
.
S
T
2
o
0
N
1
0
0
9
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
/
.
F
T
.
o
N
1
8
M
UN
sì
2
0
2
1
1546
Journal of Cognitive Neuroscience
Volume 20, Numero 9