High-Order Areas and Auditory Cortex Both Represent
the High-Level Event Structure of Music
Jamal A. Williams1, Elizabeth H. Margulis1, Samuel A. Nastase1, Janice Chen2,
Uri Hasson1, Kenneth A. Norman1, and Christopher Baldassano3
Astratto
■ Recent fMRI studies of event segmentation have found that
default mode regions represent high-level event structure during
movie watching. In these regions, neural patterns are relatively
stable during events and shift at event boundaries. Music, like
narratives, contains hierarchical event structure (per esempio., sections
are composed of phrases). Here, we tested the hypothesis that
brain activity patterns in default mode regions reflect the high-
level event structure of music. We used fMRI to record brain
activity from 25 participants (male and female) as they listened
to a continuous playlist of 16 musical excerpts and additionally
collected annotations for these excerpts by asking a separate
group of participants to mark when meaningful changes occurred
in each one. We then identified temporal boundaries between
stable patterns of brain activity using a hidden Markov model
and compared the location of the model boundaries to the loca-
tion of the human annotations. We identified multiple brain
regions with significant matches to the observer-identified
boundaries, including auditory cortex, medial prefrontal cortex,
parietal cortex, and angular gyrus. From these results, we con-
clude that both higher-order and sensory areas contain informa-
tion relating to the high-level event structure of music. Inoltre,
the higher-order areas in this study overlap with areas found in
previous studies of event perception in movies and audio narra-
tives, including regions in the default mode network. ■
INTRODUCTION
Recent work has demonstrated that the brain processes
information using a hierarchy of temporal receptive win-
dows, such that sensory regions represent relatively short
events (per esempio., milliseconds to seconds) and higher-order
regions represent longer events (per esempio., minutes) while
inheriting some of the lower-level structure from sensory
regions (Baldassano et al., 2017; Chen et al., 2017; Hasson,
Chen, & Honey, 2015). Per esempio, Baldassano et al.
(2017) used a hidden Markov model (HMM) to find tran-
sitions between stable patterns of neural activity in BOLD
data acquired from participants who watched an episode
of the TV series Sherlock. The HMM temporally divides
data into “events” with stable patterns of activity, punctu-
ated by “event boundaries” where activity patterns rapidly
shift to a new stable pattern. They found that, in sensory
regions such as early visual cortex, the data were best-fit by
a model with short-lasting chunks, presumably corre-
sponding to low-level perceptual changes in the episode;
by contrast, when they applied the model to data from
a higher-order area such as posterior medial cortex, IL
best-fitting model segmented the data into longer-
lasting chunks corresponding to more semantically
meaningful scene changes. Critically, human annotations
1Princeton University, 2Johns Hopkins University, 3Columbia
Università
© 2022 Istituto di Tecnologia del Massachussetts
of important scene changes most closely resembled the
model-identified boundary structure found in frontal and
posterior medial cortex, which are key hubs in the brain’s
default mode network (DMN; Raichle et al., 2001; Shulman
et al., 1997). Studies have also found that the same event-
specific neural patterns are activated in default-mode
regions by audiovisual movies and by verbal narratives
describing these events (Baldassano, Hasson, & Norman,
2018; Baldassano et al., 2017; Zadbood, Chen, Leong,
Norman, & Hasson, 2017), providing further evidence
that these regions represent the underlying meanings
of the events and not only low-level sensory information.
Jackendoff and Lerdahl (2006) suggest that music and
language are structured into meaningful events that help
people comprehend moments of tension and relaxation
between distant events. If music resembles language in
Da questa parte, then the representation of hierarchical event
structure in music (per esempio., at the level of phrases, sections,
and entire songs) and in verbal and audiovisual narratives
may be supported by similar neural substrates. Infatti,
some evidence already exists for shared neural resources
for processing music and language (Asano, Boeckx, &
Seifert, 2021; Lee, Jung, & Loui, 2019; Jantzen, Large, &
Magne, 2016; Peretz, Vuvan, Lagroi, & Armory, 2015;
Tillmann, 2012; Koelsch, 2011; Patel, 2011; Fedorenko,
Patel, Casasanto, Winawer, & Gibson, 2009; Tallal & Gaab,
2006; Koelsch et al., 2002). This connection between
music and language is also supported by recent behavioral
Journal of Cognitive Neuroscience 34:4, pag. 699–714
https://doi.org/10.1162/jocn_a_01815
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
C
N
_
UN
_
0
1
8
1
5
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
studies showing that instrumental music has the capacity
to drive shared narrative engagement across people
(Margulis, Wong, Turnbull, Kubit, & McAuley, 2021;
McAuley, Wong, Mamidipaka, Phillips, & Margulis, 2021;
Margulis, Wong, Simchy-Gross, & McAuley, 2019). Nel
current work, we test the hypothesis that DMN regions,
which represent high-level event structure in narratives,
also play a critical role in representing high-level event
structure in music.
In our paradigm, we presented fMRI participants with
examples of complex real-world music belonging to
genres familiar to our participant population: jazz and
classical. A separate group of behavioral participants were
asked to annotate meaningful events within each of the
excerpts. Using a whole-brain searchlight method, we
applied HMMs to measure event structure represented in
cortical response patterns throughout the brain. The goal
of this analysis was to identify brain regions that chunk
the stimuli in a way that matched the human annotations.
By fitting the model at each ROI and then comparing the
observed boundary structure to that of the annotators, we
show that—in a group of passive listeners—regions in the
DMN and also sensory areas are involved in representing
the high-level event structure in music (cioè., these regions
show neural pattern shifts that line up with human annota-
tions of event boundaries). We also show that these event
representations become coarser as they propagate up the
cortical processing hierarchy.
METHODS
Participants
We collected fMRI data from a total of 25 participants
(12 women, ages 21–33 years), which is roughly equal to
the number of participants recruited in recent studies on
event perception for narratives (per esempio., Baldassano et al.,
2018; Chen et al., 2017). We also recruited seven human
annotators for a separate behavioral task (described
below). Thirteen of the fMRI participants were native
English speakers. The experimental protocol was
approved by the institutional review board of Princeton
Università, and all participants gave their written informed
consent.
Stimuli
Sixteen musical excerpts were selected based on the crite-
rion that changes between subsections would likely be
recognized by people without formal music training
(per esempio., change from piano solo to drum solo). Excerpts also
had to be instrumental (cioè., lack vocals). Excerpts were
drawn from two different genres (eight classical and eight
jazz). Excerpts were then randomly selected to be trun-
cated (with the introductions kept intact) to one of four
different durations (90, 135, 180, E 225 sec), such that
there were four excerpts of each length. Inoltre,
two excerpts of each duration were sampled from each
genre. Per esempio, only two classical excerpts had a
duration of 90 sec, and only two jazz excerpts had a
duration of 90 sec. The total duration of the playlist was
approximately 45 min, and there were no breaks between
excerpts.
Experimental Design and Statistical Analysis
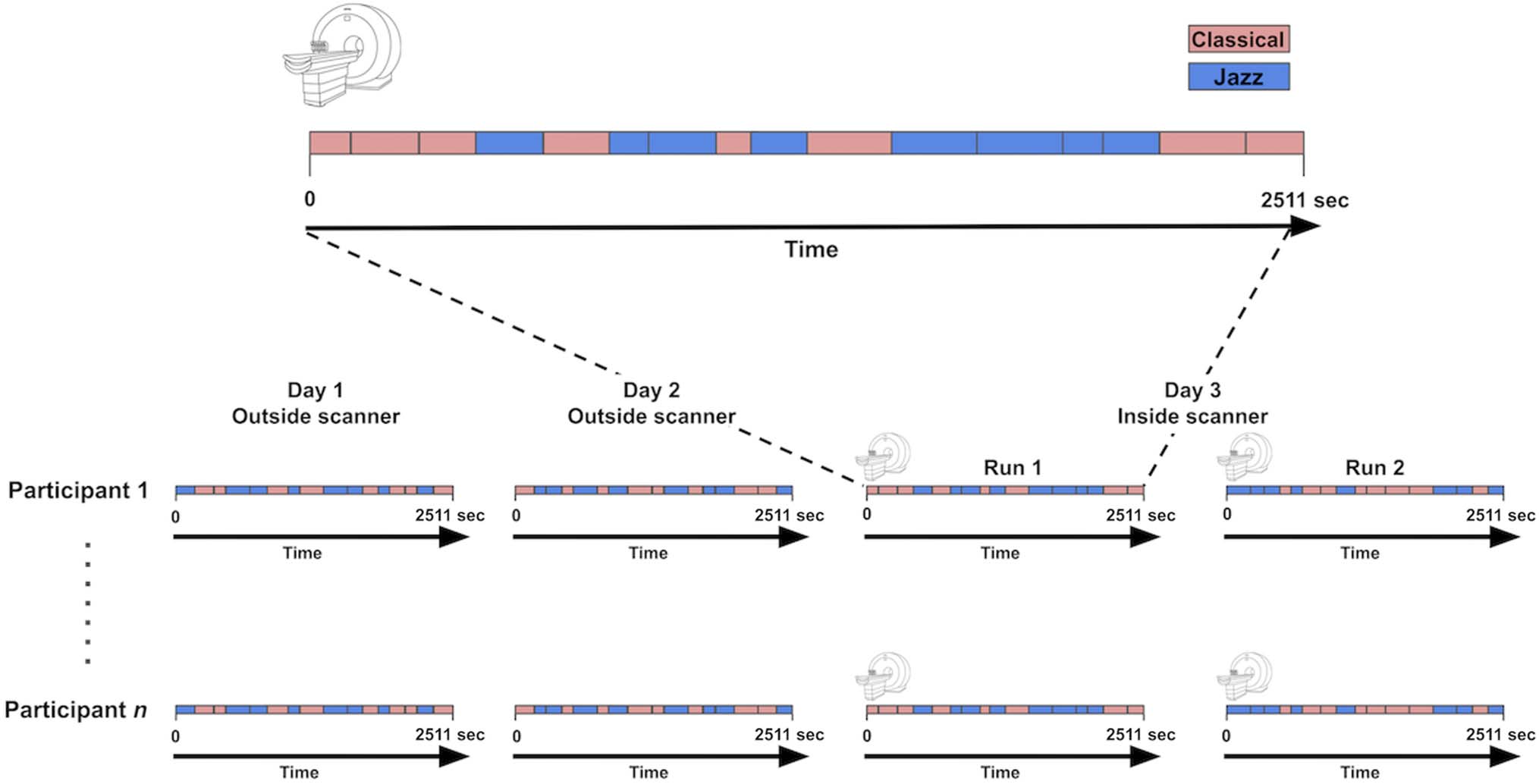
The experiment took place over three consecutive days
(Figura 1): On the first 2 days, participants heard a play-
list of 16 musical excerpts (once for each day), and on the
third day, they heard the same playlist for two separate
runs while we recorded changes in their BOLD activity
using fMRI. Altogether, each participant heard the playlist
four times. Each time that a given participant heard the
playlist, the excerpts were presented in a different order.
Tuttavia, within a given phase of the experiment (per esempio.,
the first scanner run on Day 3), the order of excerpts
was kept the same across participants. To promote stable
representations of the music, participants listened to the
playlist on each of the 2 days before scanning. During
these listening sessions, we collected ratings from partic-
ipants about their enjoyment, engagement, and familiar-
ity with each piece (only familiarity ratings are discussed
in this article); these ratings were collected immediately
after hearing each piece. Answers for each rating category
were given on a 5-point Likert scale where 1 = very
unfamiliar and 5 = very familiar. We found an increase
in average familiarity from Day 1 to Day 2, T(22) = 9.04,
P < .0001, indicating that participants remembered the
music played in the first prescan session. Two partici-
pants were excluded from this analysis because their
Day 2 ratings were lost.
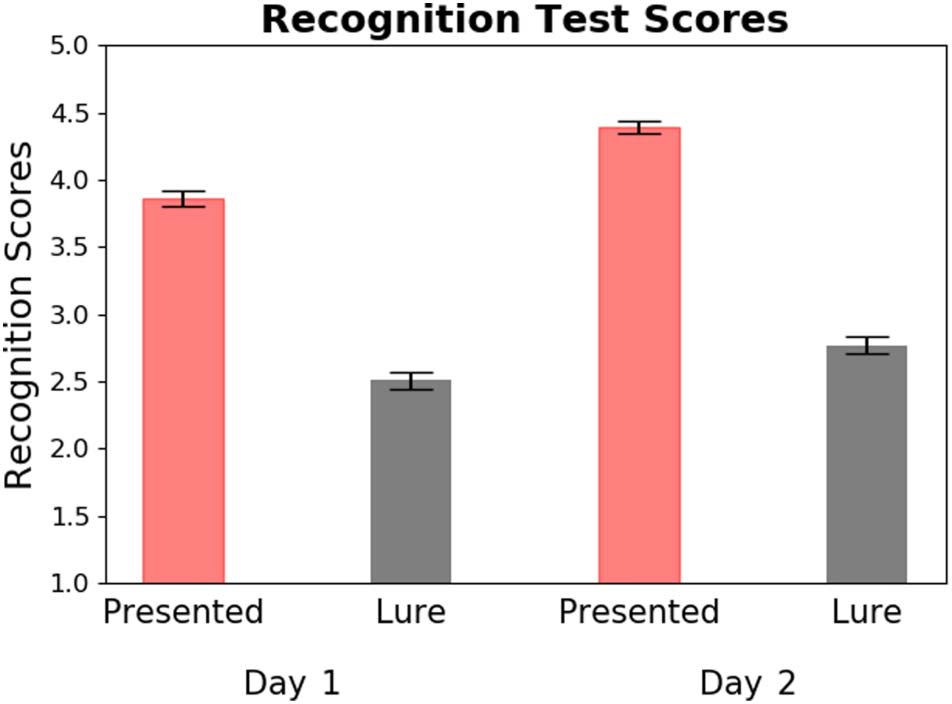
After each of these listening sessions, participants took a
short recognition test where they heard 32 randomly
drawn 3-sec clips of a piece that were either from the
actual listening session or a lure (i.e., different piece by
the same artist) and made a response using a 5-point
Likert scale indicating whether they recognized the
excerpt as having been presented previously. In addition
to the familiarity ratings across the two prescan days, this
measure helped us determine if participants had learned
the music after each behavioral listening session. Partici-
pants showed above-chance discrimination (i.e., higher
recognition scores for presented excerpts vs. lures) on
both days: Day 1: t(24) = 12.2, p < .0001; Day 2: t(24) =
15.1, p < .0001 (Figure 2).
On the third day, participants returned for the scanning
session in which they listened to the playlist twice (with
excerpts played in a different order for the two scanning
runs; as noted above, the order of excerpts within a run
was the same across participants). During each run, partic-
ipants were asked to perform a white noise detection task.
Specifically, during each excerpt, a brief (1 sec) white
noise pulse was played at a randomly chosen time point
within the middle 60% of each excerpt. The onset of
700
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. Top: Example of a 45-min scanning run, with classical excerpts depicted in pink and jazz excerpts in blue. Each block in the timeline
represents an excerpt, and block lengths reflect excerpt durations. Bottom: Overview of experiment. Participants heard the playlist four times (once
on each of the 2 days before scanning and twice on the third day while being scanned). The excerpts were presented in a different order each of the
four times that a given participant heard the playlist, but—within a given phase of the experiment (e.g., Run 1 on Day 3)—the order of excerpts was
kept the same across participants.
each noise pulse was also randomized across participants.
Participants were told to make a button response to indi-
cate that they heard the noise. This manipulation served
to keep participants attentive throughout each excerpt.
Following both scanning runs, participants took a final
recognition test and then completed a brief demo-
graphic survey.
Event Annotations by Human Observers
In a separate behavioral experiment, we asked seven dif-
ferent raters (only one rater reported having extensive
musical training) to listen to our stimuli one at a time, with
the task of pressing a button when a “meaningful” transi-
tion occurred within each piece (similar to the method
used by Sridharan, Levitin, Chafe, Berger, & Menon,
2007). The number of event boundaries identified by the
observers varied across excerpts ranging from 3 to 17
boundaries (with a mean of 7.06 and a standard deviation
of 0.91 across excerpts). It is worth noting that excerpt
durations also varied, with a range of 90–225 sec (dura-
tions were 90, 135, 190, or 225 sec) and an average dura-
tion of 157.5 sec and a standard deviation of 50.3 sec across
excerpts. A time point was considered to be an event
boundary when at least five annotators marked a boundary
within 3 sec before or after a given time point (method
used from Baldassano et al., 2017). The mean number of
consensus boundaries across excerpts acquired using this
method roughly matched the mean number of boundaries
assigned by individual participants across all of the
excerpts (with a mean of 7.98 and a standard deviation
of 2.98 across excerpts).
Figure 2. Recognition test scores for both prescan days. Plot shows
that presented excerpts were given higher recognition scores than
lures. The y-axis represents a 5-point Likert scale where 1 = not studied
and 5 = studied. Error bars represent SEM.
Scanning Parameters and Preprocessing
Imaging data were acquired on a 3-T full-body scanner
(Siemens Prisma) with a 64-channel head coil. Data were
Williams et al.
701
collected using a multiband accelerated T2-weighted EPI
sequence (release R015) provided by a C2P agreement
with University of Minnesota (Cauley, Polimeni, Bhat,
Wald, & Setsompop, 2014; Auerbach, Xu, Yacoub, Moeller,
& Uğurbil, 2013; Sotiropoulos et al., 2013; Xu et al., 2013;
Setsompop et al., 2012; Moeller et al., 2010): 72 inter-
leaved transverse slices, in-plane resolution = 2.0 mm,
slice thickness = 2.0 mm with no interslice gap, field of
view = 208 mm, base resolution = 104, repetition time
(TR) = 1000 msec, echo time (TE) = 37 msec, flip angle
(FA) = 60°, phase-encoding (PE) direction = anterior
to posterior, multiband acceleration factor = 8. Three
spin-echo volume pairs were acquired matching the
BOLD EPI slice prescription and resolution in opposing
PE directions (anterior to posterior and posterior to
anterior) for susceptibility distortion correction: TR/TE =
8000/66.60 msec, FA/refocus FA = 90°/180°, acquisition
time = 32 sec (Andersson, Skare, & Ashburner, 2003).
Additionally, a whole-brain T1-weighted volume was
collected: 3-D magnetization-prepared rapid gradient-
echo sequence, 176 sagittal slices, 1.0 mm3 resolution,
field of view = 256 mm, base resolution = 256, TR/TE =
2300/2.88 msec, inversion time = 900 msec, FA = 9°, PE
direction = anterior to posterior, IPAT mode = GRAPPA
2×, acquisition time = 5 min 20 sec.
The EPI volumes were realigned using a six-parameter
rigid-body registration (MCFLIRT; Jenkinson, Bannister,
Brady, & Smith, 2002). Given the short effective TR of 1 sec,
slice time correction was not performed. Susceptibility-
induced distortions were modeled in the opposing spin-
echo volume pairs using the FSL topup tool, and the
resulting off-resonance field output was provided as input
to distortion correct the time series of fMRI data using
the FSL applywarp tool (Andersson et al., 2003). The sus-
ceptibility distortion correction and realignment were
applied in a single interpolation step to minimize blurring.
Remaining preprocessing and coregistration steps were
performed using FEAT ( Woolrich, Behrens, Beckmann,
Jenkinson, & Smith, 2004; Woolrich, Ripley, Brady, &
Smith, 2001). This included linear detrending, high-pass
filtering (330 sec cutoff ), and spatial normalization to
the MNI152 template released with FSL.
Whole-brain Searchlight Procedure
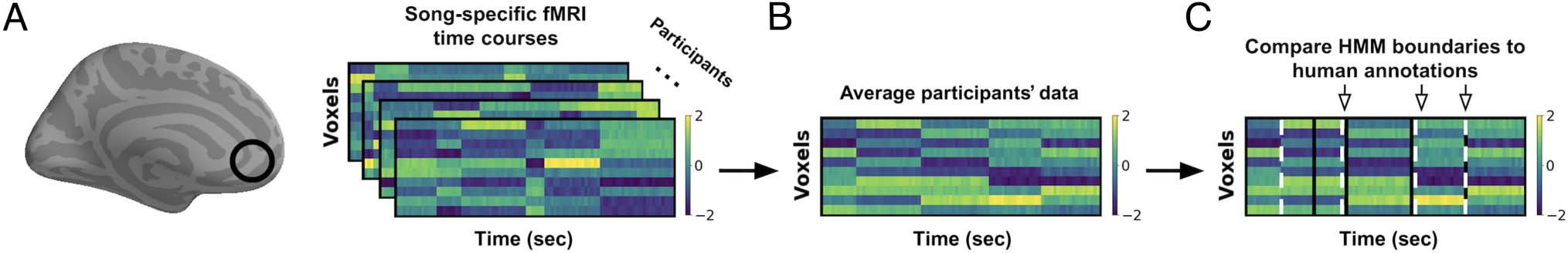
We conducted our primary analysis using a whole-brain
searchlight approach (Figure 3A). First, all participants’
volumetric data were averaged together and divided
into overlapping spherical searchlights, each with a radius
of 10 voxels and a stride of 5 voxels (Figure 3B). This
resulted in 2483 searchlights that spanned the whole
cortex in MNI space. Only searchlights containing at least
30 voxels were included in the analysis, and the mean
number of voxels per searchlight was 381.76 voxels, with
a standard deviation of 168.09 voxels. We assigned the
output value for a given searchlight to all voxels within a
5-voxel radius to account for the stride and then averaged
the values for voxels where overlap occurred. All analyses
below were run separately within each searchlight.
Event Segmentation Analysis
For each searchlight, we fit an HMM (Baldassano et al.,
2017) to the timeseries for each excerpt, setting the num-
ber of states in the HMM equal to the number of segments
specified by our human annotators for each excerpt.
Furthermore, although we provide the HMM with a spe-
cific number of events, we do not give it any information
about where these events are in the data. Therefore, the
model is unsupervised in terms of locating the boundaries
between events. We used a specialized HMM variant
developed by Baldassano et al. (2017) that is optimized
for event segmentation (i.e., identifying jumps in neural
patterns). This HMM variant seeks to model the fMRI time
series as a set of successive transitions between stable
states, where—in our variant of the HMM—the model is
not permitted to return to a state once it leaves that state.
Fitting the model to the data involves estimating the voxel
pattern for each stable event state as well as the timing of
transitions between these patterns; this HMM variant was
Figure 3. Diagram of analysis pipeline. From left to right: (A) For each participant (n = 25), voxels from an ROI were selected using a searchlight
approach; we then extracted song-specific time courses ( Voxels × TRs [TR = 1 sec]) from the selected voxels (black circle). Inflated brain image was
created using PySurfer (https://github.com/nipy/PySurfer/). (B) RT courses were averaged across participants (aligned in volumetric MNI space).
(C) An HMM was used to identify boundary time points, when there was a change in the spatial pattern of activity across voxels. HMM boundaries
(white dashed lines) and human annotations (black lines) were considered to match (downward arrows) when HMM boundaries fell within three
TRs (3 sec) of a human annotation. Then, true match scores were compared with a null distribution constructed by comparing shuffled HMM
boundaries to human annotations, resulting in a z score for each ROI.
702
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
implemented using the EventSegment function in
BrainIAK (Kumar et al., 2022).
For our primary analysis, we were interested in finding
brain regions whose transition structure most closely
resembled the event boundary structure given by our
annotators (Figure 3C). After acquiring boundary esti-
mates from the HMM, we evaluated how closely in time
the boundaries found by the model matched the bound-
aries supplied by our annotators. To quantify the degree of
match, we counted the number of human-annotated
boundaries for which there was an HMM boundary within
three TRs (3 sec) of that human-annotated boundary. Note
that all human boundaries were shifted later by five TRs
(5 sec) to account for the hemodynamic lag. We created
a null model by randomly selecting time points as bound-
aries (keeping the number of events the same, as in
Baldassano et al., 2017) and computed the number of
matches for these null boundaries, repeating this process
1000 times to produce a null distribution. We computed a
z value of the real result versus the null distribution by
subtracting the average of the permuted match scores
from the true match score and dividing this difference
by the standard deviation of the permuted scores. This
procedure was repeated at every searchlight. By acquiring
z scores at each searchlight for all 32 excerpts (16 distinct
excerpts × 2 runs), we obtained 32 separate spatial maps
of z scores. Next, we averaged the two z maps corre-
sponding to each distinct excerpt (one from each run),
resulting in 16 total z maps. To summarize across the
z scores for the 16 distinct excerpts, we ran a one-sample
t test against zero to see which voxels had the most reli-
able matches across all excerpts. The resulting t values
were converted to p values and then adjusted for multi-
ple tests to control for the false discovery rate (FDR) at a
value q (Benjamini & Hochberg, 1995). To visualize the
results, each spatial map of t values was displayed on
the cortical surface (masked to include only vertices that
exhibited a significant effect). Because each analysis was
performed in volumetric space, volume data were pro-
jected to the cortical surface using the automatic volume
to surface rendering algorithm within PySurfer (https://
github.com/nipy/PySurfer/).
Controlling for Acoustic Features
To further determine whether regions of the DMN repre-
sent high-level musical event structure, as opposed to
surface-level acoustic information, we repeated the
searchlight analysis, this time regressing out musical fea-
tures extracted from each auditory stimulus before fitting
the HMM. All feature extraction was performed using
Librosa (McFee et al., 2015), a Python package developed
for audio and music analysis. These features consisted of
mel-frequency cepstral components (MFCCs; i.e., timbre
information), chromagrams (tonal information), tempo-
grams (rhythmic information), and spectrograms. For
MFCCs, the top 12 channels were extracted because these
lower-order coefficients contain most of the information
about the overall spectral shape of the source-filter trans-
fer function (Poorjam, 2018). Chromagrams consisted of
12 features, each corresponding to a distinct key in the
chromatic scale. Tempograms initially consisted of 383
features, each representing the prevalence of certain
tempi (in beats per minute) at each moment in time.
Because most of the tempo-related variance was explained
by a much smaller set of features, we reduced the 383 fea-
tures to 12 features using PCA (variance explained was
99%) to match the number of features used for MFCCs
and chromagrams. Spectrograms were extracted using
the short-time Fourier transform (STFT) and then con-
verted to a decibel-scaled spectrogram. Then, we also
used PCA to reduce the dimensionality of the spectro-
grams to 12 components, which explained 98% of the
frequency-related variance. For the final step of this analy-
sis, we applied the HMM to the residuals after the musical
features were regressed out of the neural data.
Identifying Preferred Event Timescales
After identifying brain regions with neural event bound-
aries that matched the human annotations (using the pro-
cedures described in the Event Segmentation Analysis
section above), we ran a follow-up analysis to further
probe the properties of four such regions (bilateral audi-
tory cortex, bilateral angular gyrus, bilateral medial pre-
frontal cortex [mPFC], and bilateral precuneus). Specifi-
cally, the goal of this follow-up analysis was to assess the
preferred timescales of these regions. Angular gyrus,
mPFC, and precuneus were selected (in addition to audi-
tory cortex) because activity patterns in these regions have
been found to exhibit high-level event structure in recent
studies using naturalistic stimuli such as movies (Geerligs,
van Gerven, Campbell, & Güçlü, 2021; Ben-Yakov &
Henson, 2018; Baldassano et al., 2017; Honey et al.,
2012) and spoken narratives (Lerner, Honey, Silbert, &
Hasson, 2011). In contrast to our primary event segmenta-
tion analysis (which used a fixed number of events for each
excerpt, matching the number of human-annotated events
for that excerpt), here we tried models with different num-
bers of events and assessed how well the model fit varied
as a function of the number of events. The measure of
model fit we used was the average pattern similarity
between pairs of time point-specific multivoxel patterns
falling “within” the same event, minus the average pat-
tern similarity between patterns falling “across” events
(Baldassano et al., 2017). We call this measure the “WvA
score” (short for “Within vs. Across”); higher WvA scores
indicate a better fit of the event boundaries to the data.
The ROIs for this analysis were defined by selecting voxels
within functionally defined parcellations (Schaefer et al.,
2018) corresponding to bilateral auditory cortex, bilateral
angular gyrus, bilateral mPFC, and bilateral precuneus and
then (for extra precision) intersecting these parcels with
voxels that were also significant in our primary searchlight
Williams et al.
703
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
analysis looking for neural boundaries that matched
human-annotated boundaries (q < 0.01). For each ROI,
we fit HMMs to each song with differing numbers of
events ranging from 3 to 45. For each HMM fit, we mea-
sured the maximum event duration and then identified all
pairs of time points whose temporal distance was less
than this duration. The constraint of using time points
whose distance was less than the maximum event
duration was used so that the number of within- and
across-event pairs would be roughly equal (regardless of
the number of events). The WvA score was computed as
the average spatial pattern correlation for pairs of time
points falling in the same (HMM-derived) event minus
the average correlation for pairs of time points falling in
different events. We then averaged the results across
excerpts. Note that, because the excerpts are different
lengths, a given number of events might correspond to
different average event lengths for different excerpts
(e.g., a three-event model applied to a 180-sec excerpt
has an average event length of 60 sec, but a three-event
model applied to a 90-sec excerpt would have an average
event length of 30 sec). Because our goal was to find each
area’s preferred event length, we converted our WvA
results for each excerpt to be a function of the average
event length (in seconds) rather than the number of
events and averaged these results across excerpts. Finally,
to compute the preferred event length for each ROI, we
identified the range of event lengths that were within 5%
of the maximum WvA score for that ROI; we report the
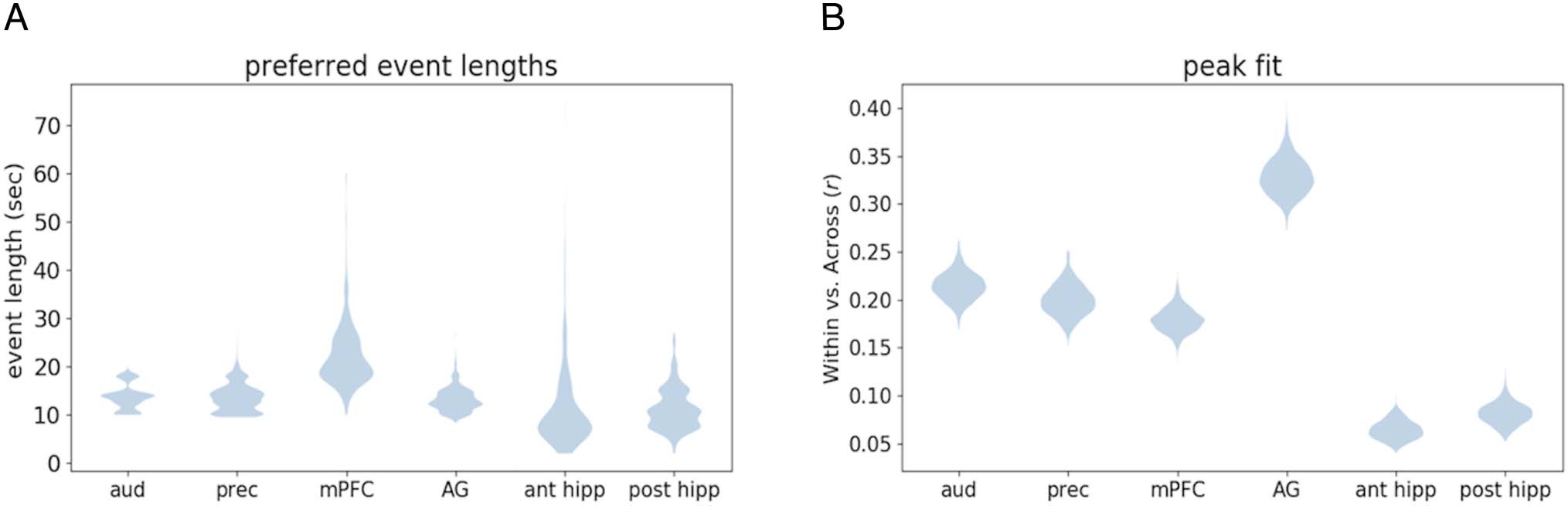
midpoint of this range as the preferred event length.
To test whether the preferred event length in auditory
cortex was shorter than that of angular gyrus, precuneus,
and mPFC, we performed a bootstrap analysis, repeating
the above analysis 1000 times for different bootstrap
resamples of the original data set. At each iteration of
the bootstrap, we applied the analysis to a sample of par-
ticipants drawn randomly with replacement from the orig-
inal data. We computed p values by finding the proportion
of bootstraps where the preferred length for auditory cor-
tex was greater than the preferred length for angular gyrus,
precuneus, and mPFC.
RESULTS
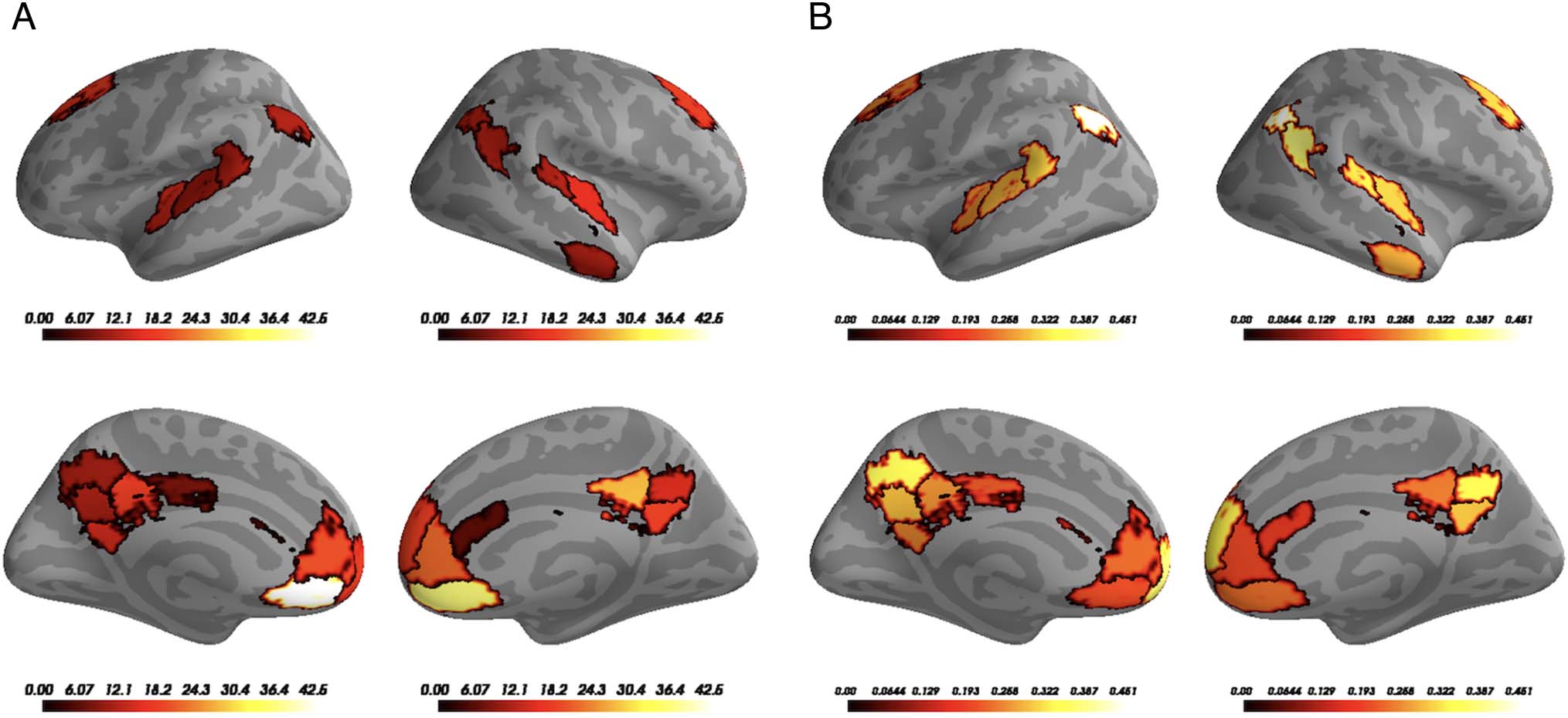
Neural Boundary Match to Behavioral Annotations
We wanted to test the hypothesis that behaviorally defined
event boundaries could be identified in higher-order corti-
cal regions, especially those overlapping with the DMN. For
this analysis, we fit an HMM to BOLD data averaged across
both runs and then compared the HMM boundaries to the
specific time points labeled as boundaries by the annota-
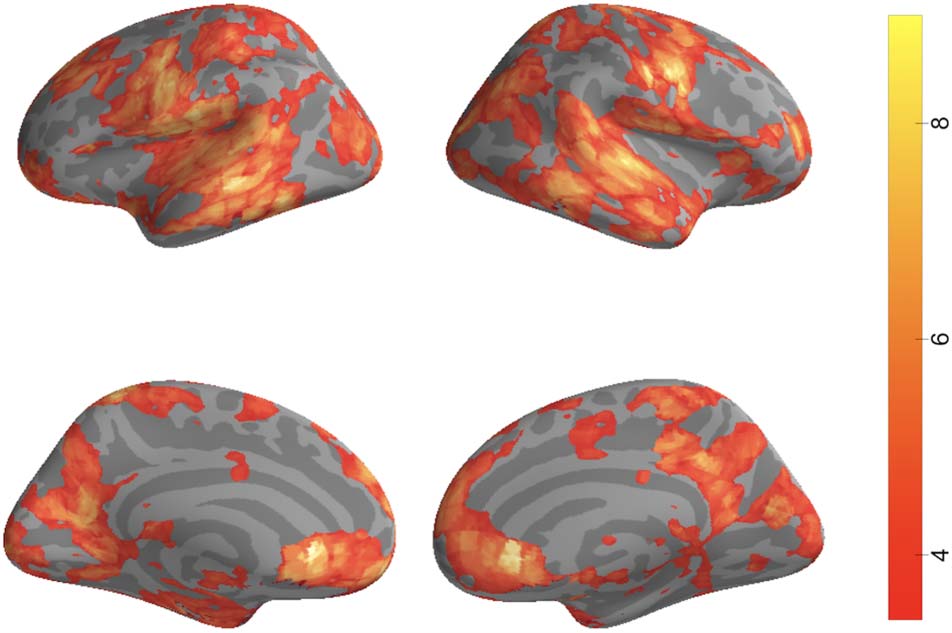
tors. We found significant matches between model bound-
aries and human annotations in auditory cortex, angular
gyrus, precuneus, and mPFC, with a greater number of
model boundaries and human boundaries having low tem-
poral distance than expected by chance (Figure 4). Results
for this analysis are split by Run 1 and Run 2 in Appendix B.
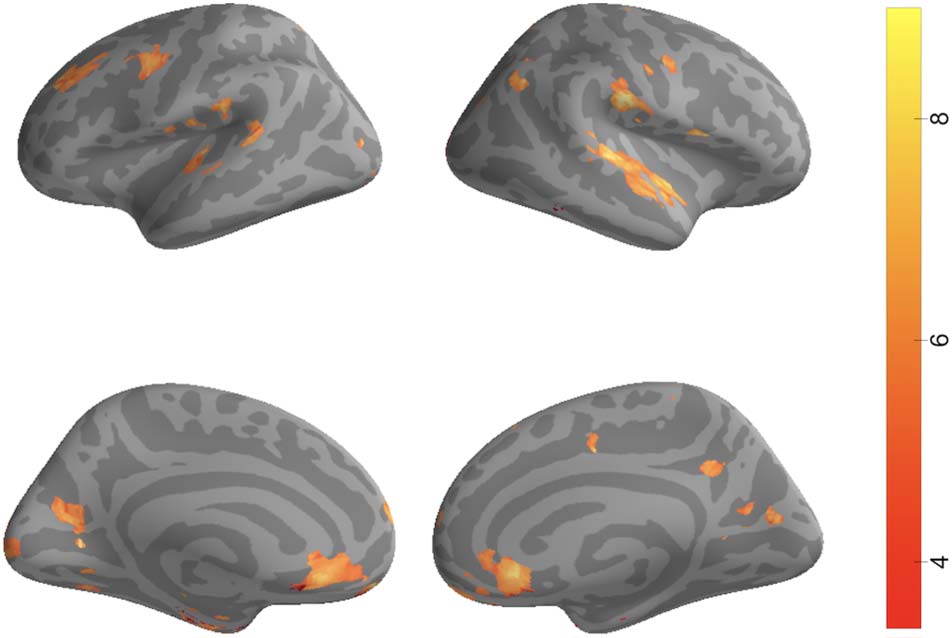
Figure 4. Distance to boundary searchlight results. For 2483
searchlights spanning the entire cortex, we tested whether the average
match between neural and annotated boundaries across all songs was
significantly greater than zero. Significant voxels overlapped with
auditory cortex as well as areas of the DMN such as precuneus, mPFC,
and angular gyrus. Results are thresholded via FDR (q < 0.01).
Influence of Acoustic Features
To determine the extent to which the neural event bound-
aries were driven by acoustic features, we also performed a
version of the searchlight analysis in which we controlled
for spectral, timbral, harmonic, and rhythmic information.
Overall, this reduced the number of searchlights passing
the q < 0.01 FDR threshold (Figure 5) compared with
the original searchlight analysis. However, searchlights in
DMN regions (precuneus, angular gyrus, and mPFC) did
pass the q < 0.01 threshold, with voxels in mPFC being
(numerically) least affected by the feature removal. When
we set a more liberal FDR threshold (q < 0.05; results
shown in Appendix A), the relationship between neural
event boundaries and human annotations was still largely
conserved in precuneus, angular gyrus, and auditory cor-
tex. This suggests that, although voxels in precuneus and
angular gyrus are more sensitive to acoustic features than
mPFC, event boundaries found in these regions do not
directly correspond to simple changes in the acoustic
features and may instead be related to more complex
representations of the event structure (e.g., nonlinear
combination of acoustic features). Notably, significant
searchlights in auditory cortex were also observed (partic-
ularly in right auditory cortex), indicating that—even in
sensory areas—the event boundaries were being driven
(at least in part) by more high-level aspects of the music.
Comparing Annotated Event Boundaries to
Changes in Acoustic Features
In a follow-up analysis, we sought to further investigate
the relationship between the event boundaries and
changes in the acoustic features by assessing how often
the behaviorally defined event boundaries occurred at
704
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
(i.e., MFCC, chromagram, tempogram, and spectrogram)
extracted from each of the excerpt audio files. One advan-
tage of using the GSBS algorithm for this analysis is that
GSBS can automatically identify the optimal number of
states that maximizes the difference between within- versus
across-event similarity. After acquiring the optimal set of
GSBS event boundaries for each excerpt, we compared
them to the human annotations by computing the probabil-
ity that a shift in an acoustic feature generated a matching
human annotation (within 3 sec). Additionally, we assessed
whether this probability was greater than what would be
predicted by chance by establishing a null distribution
whereby we shuffled the feature boundaries for each
excerpt while preserving the distances between boundaries.
We found that feature boundaries did align with human-
annotated boundaries more often than in the null model,
but that most feature changes did not result in a human-
annotated boundary ( p(annotation | chroma boundary) =
0.143 vs. null value of 0.115 [p < .001], p(annotation |
MFCC boundary) = 0.493 vs. null value of 0.299 [p < .001],
p(annotation | tempo boundary) = 0.198 vs. null value
of 0.179 [p < .05], p(annotation | spectrogram bound-
ary) = 0.160 vs. null value of 0.128 [p < .001]; illustrated
using an example excerpt in Figure 6A).
We also computed the distribution (across human-
annotated boundaries) of the number of acoustic feature
types that changed within 3 sec of each annotated bound-
ary (e.g., if chroma and tempo both changed, that would
be two feature types). We compared this distribution to a
null model that we obtained by shuffling the human-
Figure 5. Searchlight results accounting for acoustic features. We
recomputed the match between HMM-derived neural boundaries and
human annotations after regressing out acoustic features from each
participant’s BOLD data before fitting the HMM. Significant effects were
still observed in parts of the DMN as well as auditory cortex, suggesting
that boundaries detected in these areas do not necessarily depend on
acoustic features. Results are thresholded via FDR (q < 0.01).
the same time as changes in each of the acoustic features.
In other words, how often does a change in an acoustic
feature generate a human boundary? To estimate the
number and locations of state changes within each of
the excerpts, we applied the Greedy State Boundary
Search (GSBS) event segmentation model (Geerligs, van
Gerven, & Güçlü, 2021) to each of the acoustic features
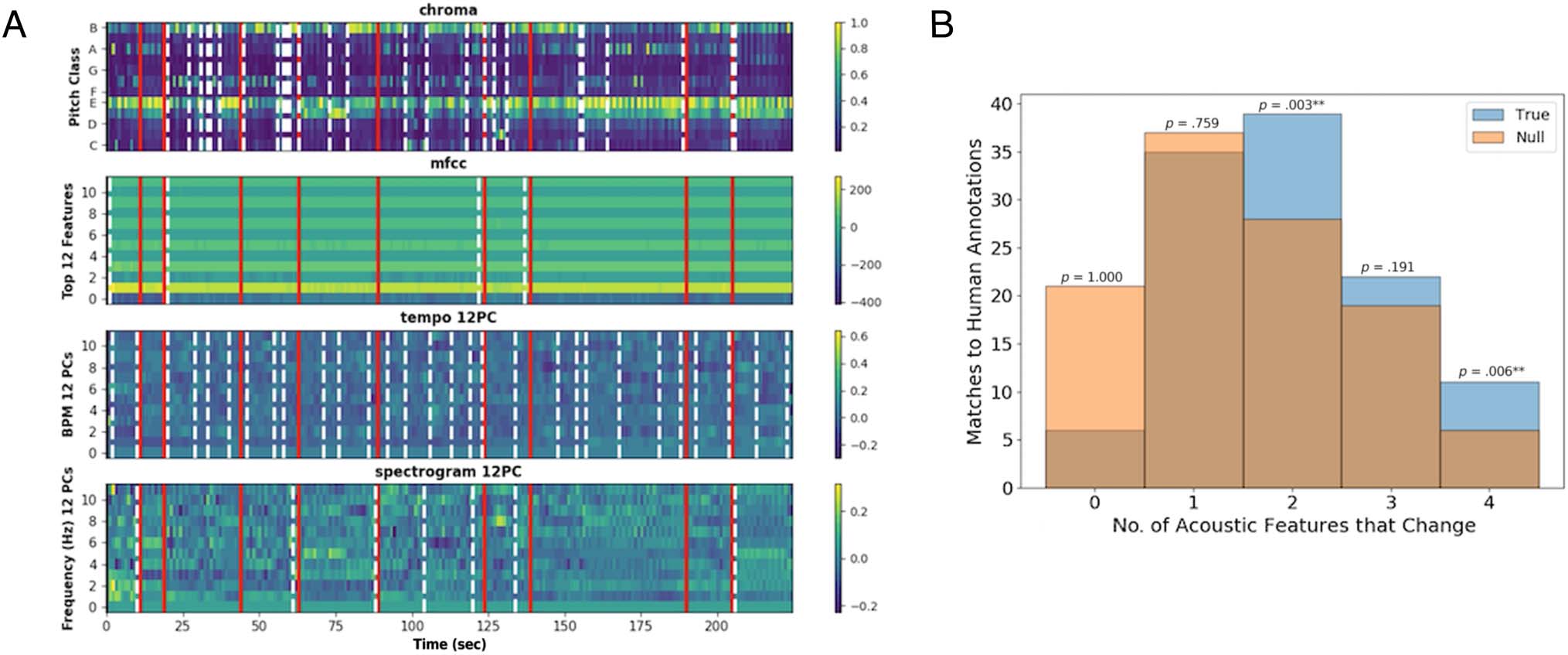
Figure 6. (A) Example of acoustic features (from My Favorite Things by John Coltrane) showing overlap between human annotations (red) and
feature boundaries (white dashed lines). For each acoustic feature, we identified time points at which changes occurred using the GSBS event
segmentation model (white dashed lines). We then compared the locations of these feature boundaries to the locations of the human annotations
(red lines); see text for results. (B) Number of acoustic features that change at human-annotated event boundaries. Counting how many acoustic
features exhibit a boundary at the same time as a human-annotated boundary (blue) versus a null distribution (orange), we find that the observed
distribution is shifted upward relative to the null distribution, such that human-annotated boundaries are more likely to occur in response to two or
more feature changes. Furthermore, some human annotations occur in the absence of any feature change.
Williams et al.
705
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
annotated boundaries for each excerpt while preserving
the distances between boundaries. The results of this
analysis are shown in Figure 6B. The fact that the observed
distribution is shifted upward relative to the null tells us
that the probability of human boundaries coinciding with
auditory the feature changes is higher than would be
expected due to chance (χ2 = 19.54, p < .001 by permu-
tation test). The figure also shows that, though the
majority of human boundaries occurred at points where
two or more acoustic feature changes were present, some
human boundaries occurred at time points where no
acoustic feature changes were present.
Preferred Event Lengths across ROIs
How do we reconcile the role of auditory cortex in high-
level event representation (as shown in the above analy-
ses) with its well-known role in representing low-level
auditory features? Importantly, these claims are not mutu-
ally exclusive. Our analyses, which set the number of event
states in the model to equal the number of human-
annotated boundaries, show that auditory cortex has some
(statistically reliable) sensitivity to high-level events, but
this does not mean that this is the “only” event information
coded in auditory cortex or that it is the “preferred” level
of event representation.
We defined the preferred timescale of each region (ROI
selection is discussed in the Experimental Design and Sta-
tistical Analysis section) by running HMMs with different
numbers of event states and finding the average event
length (in seconds) that produced the best model fits
across songs (Figure 7A). Using a bootstrap analysis, we
found that auditory cortex’s preferred event length
(13.81 sec) was significantly shorter than the preferred
event length of mPFC (25.59 sec; p = .009) but was not
significantly shorter than the preferred length of angular
gyrus (13.36 sec; p = .664) or precuneus (14.61 sec; p =
.338). The preferred event length in mPFC was also signif-
icantly longer than the preferred event length for precu-
neus ( p = .017) and angular gyrus ( p = .004).
In addition to varying the timescale (i.e., in the best-
fitting number of events), regions could differ in the
quality of this fit; some regions may exhibit sharper event
transitions, with large pattern changes across HMM event
boundaries and highly stable patterns within events. We
therefore tested whether the model fit (maximum WvA
score) was different between the four ROIs (Figure 7B).
We found that the model fit for angular gyrus was
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
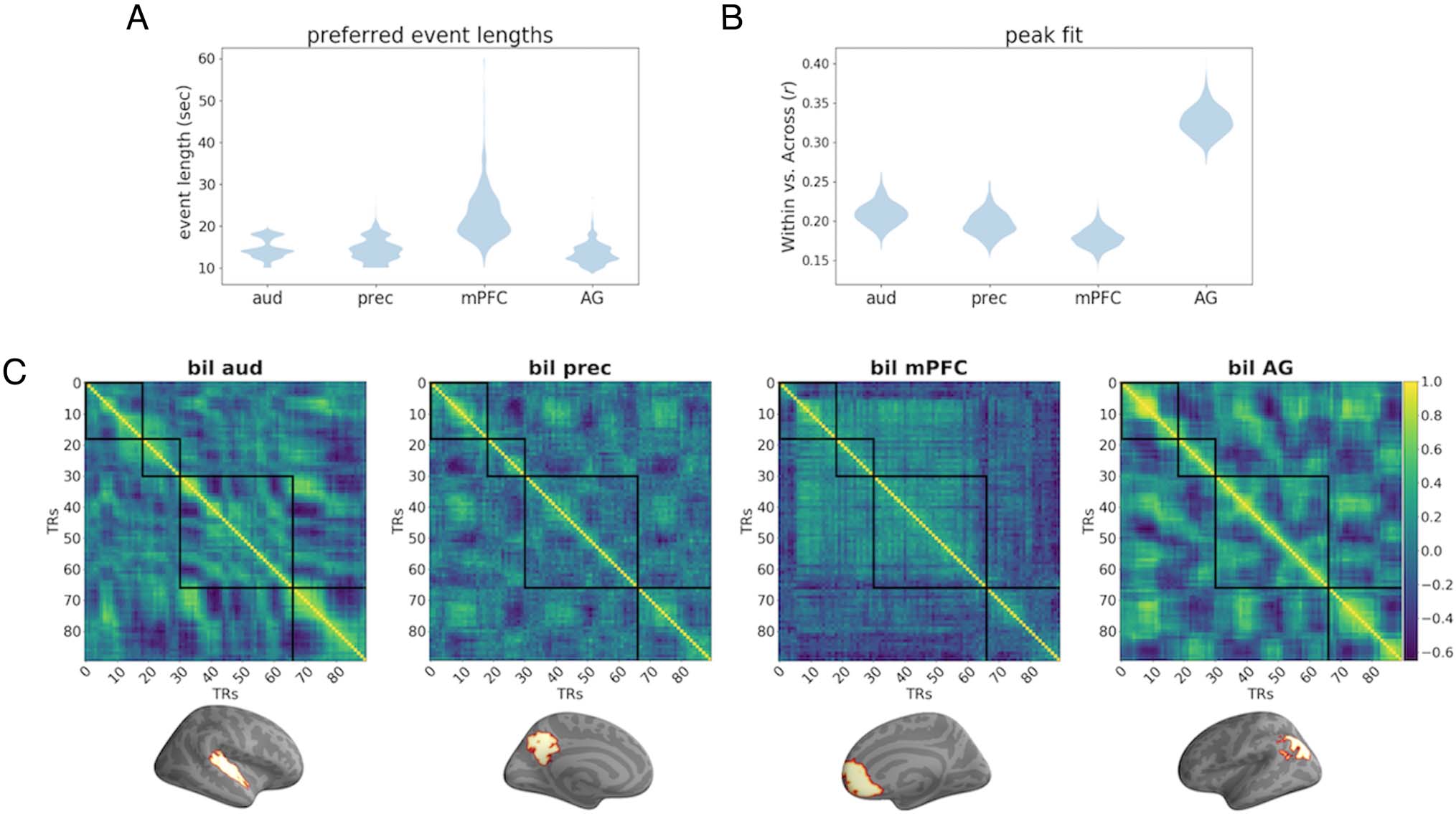
Figure 7. (A) Longer states were preferred in mPFC (average event length 25.59 sec) than in auditory cortex (13.81 sec), precuneus (14.61 sec), and
angular gyrus (13.36 sec). The preferred event length did not significantly differ between auditory cortex, precuneus, and angular gyrus. (B) The
overall within-event pattern similarity was highest in angular gyrus, suggesting that the stability of musical event representations was higher than in
other ROIs. There was no difference in within-event pattern similarity between precuneus and auditory cortex; however, pattern similarity was
significantly less in mPFC than in auditory cortex ( p < .05). (C) Similarity matrices (for the first 90 sec of the excerpt Capriccio Espagnole by Nikolai
Rimsky-Korsakov) showing pattern similarity over time for each ROI with human-annotated boundaries shown in black. mPFC exhibits the coarsest
event structure relative to auditory cortex, precuneus, and angular gyrus.
706
Journal of Cognitive Neuroscience
Volume 34, Number 4
significantly greater than auditory cortex ( p < .001), pre-
cuneus ( p < .001), and mPFC ( p < .001), indicating that
the temporal event structure was strongest in angular
gyrus. For analyses of preferred event length and model
fit in a more complete set of DMN ROIs and in hippo-
campus, see Appendices C and D, respectively.
DISCUSSION
In this study, we sought to determine whether brain areas
that have been implicated in representing high-level event
structure for narrative-based stimuli, such as movies and
spoken narratives, are also involved in representing the
high-level event structure of music in a group of passive
listeners. We provide evidence that regions of the DMN
are involved in representing the event structure of music
as characterized by human annotators. The durations of
these human-annotated events lasted on the order of a
few seconds up to over a minute.
Our results indicate that high-level structure is repre-
sented in both high-level DMN regions but also in auditory
cortex. Auditory cortex, however, may not explicitly repre-
sent high-level events at the level of human annotators;
that is, the behaviorally identified event boundaries are
likely a subset of the finer-grained event boundaries
encoded in auditory cortex. When we force the HMM to
match the number of human-annotated boundaries, the
HMM finds them, demonstrating that coding in auditory
cortex is modulated by high-level event structure. How-
ever, when we remove this constraint and allow the num-
ber of events to vary, auditory cortex prefers shorter
events on average relative to mPFC but not precuneus
and angular gyrus (Figure 7A), whereas mPFC preferred
the longest events compared with the other three ROIs.
The finding that the preferred event length of auditory cor-
tex was not significantly different from that of precuneus
and angular gyrus was surprising given the prediction that
auditory cortex, which is generally thought to respond to
fast-changing aspects of a stimulus, would represent
shorter events than higher-order brain areas (Baldassano
et al., 2017; Farbood, Heeger, Marcus, Hasson, & Lerner,
2015; Lerner et al., 2011; Hasson, Yang, Vallines, Heeger, &
Rubin, 2008); we discuss this point further in the limita-
tions section below. In addition to measuring each area’s
preferred timescale, we also measured within-event stabil-
ity across the four ROIs; here, we found that angular gyrus
exhibits the strongest within-event activity relative to pre-
cuneus, mPFC, and auditory cortex.
Next, we showed that—when we regress out acoustic
features corresponding to timbre, harmony, rhythm, and
frequency amplitude and rerun the analysis—voxels in
higher-order areas (mPFC, angular gyrus, and precuneus),
as well as auditory cortex, still significantly match with the
annotations. These results suggest that event boundaries
in these regions are not purely driven by acoustic changes
in the music but are also tracking more complex event
structure in musical pieces. These findings are consistent
with findings from Abrams et al. (2013), who found that
naturalistic music elicited reliable synchronization in
auditory cortex as well as higher-order cortical areas after
controlling for acoustic features; they concluded that this
synchronization was not purely driven by low-level acous-
tical cues and that it was likely driven by structural ele-
ments of the music that occurred over long timescales.
To further determine how much event boundaries were
driven by changes in acoustic features, we ran a follow-up
analysis where we first identified event transitions in each
of the acoustic features corresponding to timbre, tonality,
rhythm, and frequency amplitudes for each excerpt using
an unsupervised algorithm (GSBS); then, we computed
the probability that a human annotation was generated
by changes in each of the different types of acoustic fea-
tures. We found that the probability of human-annotated
boundaries coinciding with acoustic feature changes was
higher than the rate expected because of chance, but
the relationship was complex: Although changes in each
of the individual acoustic feature types were significantly
related to the occurrence of annotated boundaries, none
of these features came close to fully predicting the anno-
tated boundaries, and although the majority of annotated
boundaries occurred at time points where two or more
acoustic features changed, some annotated boundaries
did not correspond to changes in any of the acoustic
features that we tracked. This adds further support to
the possibility that boundaries marking the shift between
large-scale segments within the DMN and auditory areas
could be driven by a complex shift in a combination of
the acoustic properties and/or possibly emotional (Daly
et al., 2015) or narrative (Margulis et al., 2019, 2021;
McAuley et al., 2021) changes within the excerpts, rather
than a change in a single feature.
Importantly, our findings of high-level coding in audi-
tory cortex converge with other recent work demonstrat-
ing that hierarchical neural representations of music are
distributed across primary and nonprimary auditory cortex
(Landemard et al., 2021) and that higher-order represen-
tations of music in these areas may even support complex
behaviors such as genre recognition in humans (Kell,
Yamins, Shook, Norman-Haignere, & McDermott, 2018).
Our study contributes to this growing literature by
showing that auditory cortex codes for musical event rep-
resentations at intermediate timescales (∼14 sec). Note
also that auditory cortex coding for these intermediate-
scale events is not mutually exclusive with it “also” coding
for shorter-timescale events. When discussing limitations
of our study below (limitation point number 4), we pro-
vide some reasons why our design was not ideal for detect-
ing neural coding of short-timescale events.
In our study, we provide strong evidence for the involve-
ment of mPFC in representing high-level musical event
structure. Recent fMRI studies of naturalistic stimulus pro-
cessing (i.e., audiovisual movies) have shown that mPFC
may perform event segmentation and integration during
continuous memory formation (Antony et al., 2021; Liu,
Williams et al.
707
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Shi, Cousins, Kohn, & Fernández, 2021) and that events in
this region can last up to hundreds of seconds (Geerligs
et al., 2021; Chen et al., 2017; Hasson et al., 2015). We also
show that the preferred event length in mPFC was ∼25 sec
(which was roughly equal to the preferred timescale found
for mPFC in the study by Geerligs et al., 2021, in which a
movie was used rather than music), adding further support
to the hypothesis that mPFC plays an important role in
representing long-timescale information in naturalistic
stimuli. Furthermore, our findings go beyond the assump-
tion that areas of the DMN only represent long-timescale
information for narrative-based stimuli and instead suggest
that areas of the DMN represent long-timescale informa-
tion across a range of naturalistic stimuli, including
music. The recruitment of mPFC during music process-
ing has also been found in a previous study (Blood &
Zatorre, 2001). Specifically, Blood and Zatorre showed
that activity in vmPFC was correlated with pleasure
response ratings to music, suggesting that frontal areas,
which represent long-timescale event structure for music,
may also play a role in processing reward and affect in
response to music.
Our findings that precuneus, mPFC, and angular gyrus
were involved in representing high-level musical event
structure contrast with those in Farbood et al. (2015),
who found that regions that responded reliably to stories
did not respond reliably to music. Furthermore, in their
study, there was minimal overlap between voxels in angu-
lar gyrus and mPFC that responded to stories and voxels
that responded to music. In our study, we show that, at
a regional level, these areas are indeed involved in repre-
senting the high-level event structure in music. One major
way in which our studies differed was our use of an HMM
to detect evidence of musical event structure in higher-
order areas. The HMM is optimized to detect periods of
relative stability punctuated by shifts in response patterns,
which one would expect for an area encoding high-level
event structure (i.e., there should be stability within events
and changes across events). Temporal intersubject corre-
lation analysis (the analysis method used in the study by
Farbood et al., 2015) is designed to pick up on “any” kind
of reliable temporal structure and is not specifically
designed to detect the “stability punctuated by shifts”
structure that we associate with event cognition, making
it less sensitive to this kind of structure when it is present.
This highlights one of the advantages of using HMMs for
detecting meaningful brain activity related to the temporal
dynamics of naturalistic stimuli, such as music.
Our study had several limitations:
1.
In our feature regression analysis, the acoustic fea-
tures we selected may not represent the full range
of acoustic dynamics occurring throughout each
excerpt. Previous studies using encoding models to
examine brain activity evoked by music employed a
range of acoustic features, such as the modulation
transfer function (Norman-Haignere, Kanwisher, &
McDermott, 2015; Patil, Pressnitzer, Shamma, &
Elhilali, 2012) as well as music-related models repre-
senting mode, roughness, root mean square energy,
and pulse clarity (Nakai, Koide-Majima, & Nishimoto,
2021; Toiviainen, Alluri, Brattico, Wallentin, & Vuust,
2014; Alluri et al., 2012). However, the types of infor-
mation captured by these features are also roughly
captured by the features used in this study. For exam-
ple, features representing roughness and root mean
square capture timbral information, whereas pulse
clarity captures rhythmic information. On the other
hand, although these features capture some informa-
tion related to the ones used in this study, they may
nonetheless still be useful for capturing additional
information not fully captured by our features. Future
work is needed to determine how higher-order areas
are affected by a larger set of acoustic features.
2. Another caveat is that we only scanned participants
listening to prefamiliarized musical stimuli—as such,
it is unclear whether the observed pattern of DMN
results (showing engagement of these regions in
long-timescale segmentation) would extend to unfa-
miliar musical stimuli. Consistent with this view, the
work by Castro et al. (2020) showed that familiar
music engaged DMN more strongly than unfamiliar
music. However, a study by Taruffi, Pehrs, Skouras,
and Koelsch (2017) showed that DMN was engaged
for unfamiliar music, particularly for sad music com-
pared with happy music. Future work investigating
high-level musical event structure representation
can address this by scanning participants while they
listen to both unfamiliar and familiar stimuli.
3. The white noise detection task that participants per-
formed may have influenced DMN responding. The
DMN has been shown to activate during mind-
wandering or stimulus-independent thought (Mason
et al., 2007). Because the white noise was spectrally
distinct from the music, participants could conceiv-
ably perform the white noise detection task without
attending to the music, leaving room for them to
mind-wander in between white noise bursts; conse-
quently, some of the DMN responding could (in prin-
ciple) have been driven by mind-wandering instead of
music listening. However, stimulus-independent
mind-wandering cannot explain our key finding that
neural event boundaries in DMN regions align with
the annotated event boundaries—this result clearly
demonstrates that these DMN areas are tracking
structural aspects of the music.
It is possible that our estimates of preferred event
length for different ROIs were biased by the range
of event lengths present in our stimulus set. In par-
ticular, a lack of short (vs. long) events may have
resulted in an upward bias in our estimates of pre-
ferred event length. This bias, however, cannot
explain the relative differences that we observed
between ROIs’ preferred timescales, such as mPFC
4.
708
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
preferring longer events than auditory cortex, precu-
neus, and angular gyrus. However, the relative scar-
city of short events may have impaired our ability
to resolve timescale differences between regions at
the short end of the timescale continuum; in partic-
ular, this might help to explain why we did not
observe significant differences in preferred time-
scales between primary auditory cortex (which, based
on prior work, we expected to have a short timescale
preference) and DMN regions. Future work can shed
light on this by using stimuli with a broader range of
event lengths. However, even if we include stimuli
with shorter events, our ability to detect these more
rapid event transitions may be inherently limited by
the slow speed of the fMRI BOLD response.
Conclusion
In this study, we sought to determine whether certain
regions in the DMN, which have been shown to be
involved in representing the high-level event structure
in narratives, were also involved in representing the
high-level event structure in real-world music. Recent
fMRI work, not using music, has shown that HMMs can
help us understand how the brain represents large-scale
event structure. By using HMMs to segment fMRI
response patterns over time according to the event struc-
ture provided by a separate group of human annotators,
we found that areas of the DMN were indeed involved in
representing the high-level event structure (e.g., phrases,
sections) in music in a group of passive listeners. Of par-
ticular importance are the findings that mPFC has a
chunking response that is close to that of human
observers and survives the boundary alignment search-
light analysis even after controlling for acoustic features.
This suggests that mPFC plays an important role in high-
level event representation not only for movies and stories
(Geerligs et al., 2021; Baldassano et al., 2017; Chen et al.,
2017; Hasson et al., 2015; Lerner et al., 2011) but also for
instrumental music.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
APPENDIX A
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure A1. Distance to boundary regression results at q < 0.05. Plots show distance to boundary regression results in which we regress out MFCCs,
chromagrams, tempograms, and spectrograms. Results are FDR corrected at q < 0.05. These results show that, although many voxels in the DMN are
not significant at the q < 0.01 threshold (Figure 5), many DMN voxels do survive when we threshold the regression results at q < 0.05. This suggests
that, though many voxels in the DMN are somewhat sensitive to acoustic features (because many of these voxels do not survive at q < 0.01 in the
nonregression distance to boundary results), activity in these areas is not solely driven by low-level acoustic features.
Williams et al.
709
APPENDIX B
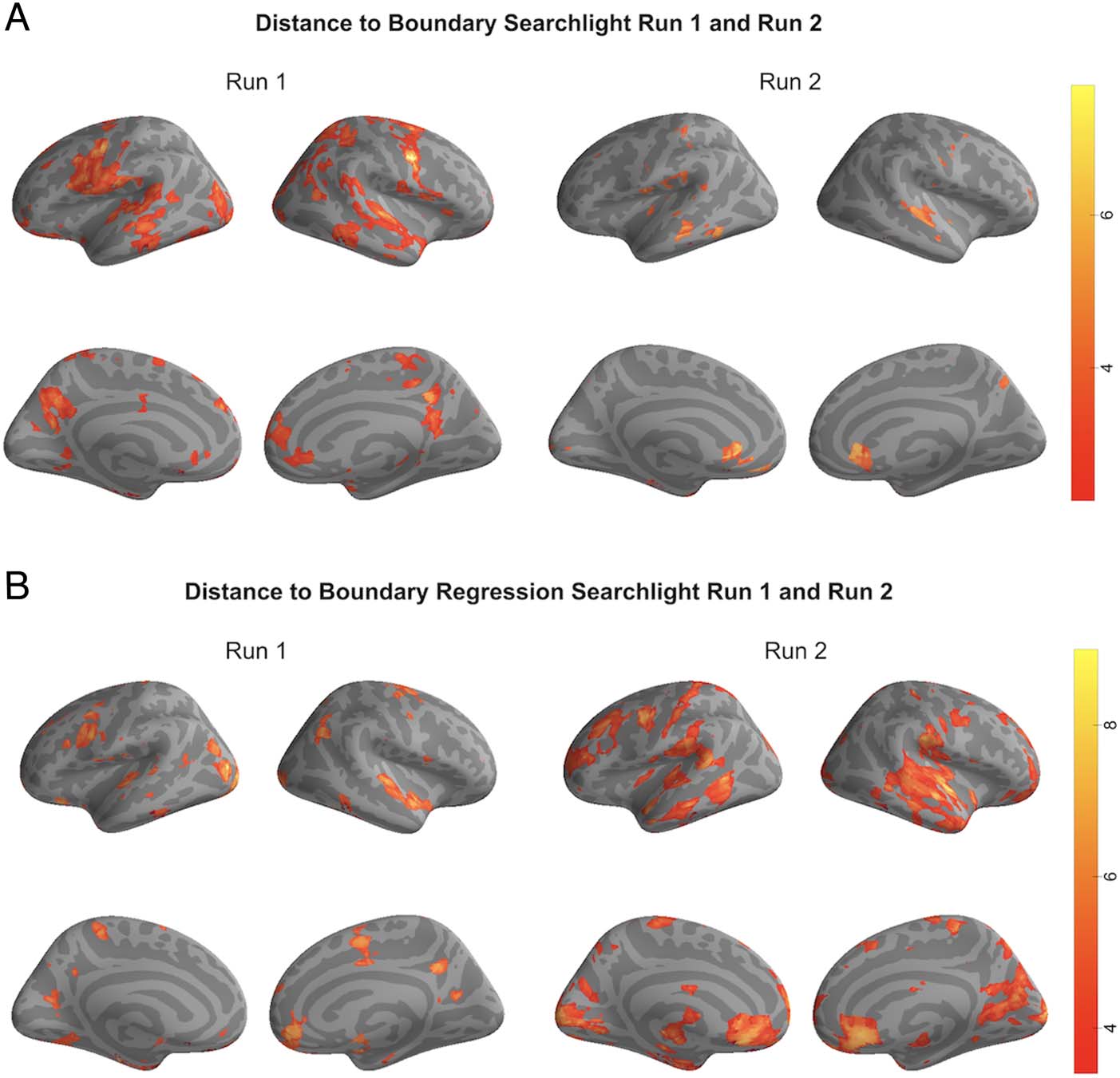
Figure B1. (A) Distance to
boundary searchlight Run 1
and Run 2. Searchlight maps
for each run separately showing
regions where significant
matches between human
annotations and HMM
boundaries were observed
(FDR corrected q < 0.01).
(B) Distance to boundary
regression searchlight Run 1
and Run 2. Searchlight maps for
each run separately showing
regions where significant
matches between human
annotations and HMM
boundaries were observed after
regressing out acoustic features
(FDR corrected q < 0.01).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
710
Journal of Cognitive Neuroscience
Volume 34, Number 4
APPENDIX C
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure C1. (A) Preferred event lengths across finer set of DMN and auditory ROIs. We sought to further determine the set of event lengths preferred
within each ROI using a finer set of parcellations (Schaefer 300 as opposed to Schaefer 100). We attempted to threshold this image by only including
ROIs with significant model fits (determined via bootstrapping). Nothing survived our threshold criteria; therefore, we are reporting unthresholded
results. Subregions of DMN preferred a variety of event lengths, which was not obvious when using a coarser set of parcellations. For example,
although mPFC obtained from the Schaefer 100 parcellation set shows a preference for the longest event lengths (∼25 sec), when evaluating this for a
finer set of mPFC ROIs (Schaefer 300), we can see that mPFC subregions prefer a variety of event lengths ranging from 6 to 40 sec. (B) Model fits also
vary greatly for the same set of DMN and auditory ROIs.
APPENDIX D
Figure D1. (A) Anterior hippocampus preferred event length did not significantly differ from auditory cortex, precuneus, mPFC, angular gyrus, or
posterior hippocampus. Posterior hippocampus preferred event length did not significantly differ from auditory cortex, precuneus, angular gyrus, or
anterior hippocampus, but was significantly less than mPFC ( p < .05). (B) Our measure of model fit (i.e., the difference between within-event and
across-event pattern similarity) was significantly lower in hippocampal ROIs than in other DMN ROIs (auditory cortex, p < .001; precuneus, p < .001;
mPFC, p < .001; angular gyrus, p < .001), whereas model fit in posterior hippocampus was greater than in anterior hippocampus ( p < .05).
Williams et al.
711
Acknowledgments
We thank Mark A. Pinsk for contributing to the Scanning Param-
eters and Preprocessing section of the article, Benson Deverett
for helping with the stimulus presentation script in Python,
Elizabeth McDevitt for suggestions on the figures, Sara Chuang
for helping with stimulus selection, and the members of the
Hasson, Pillow, and Norman labs for their comments and
support.
Reprint requests should be sent to Jamal A. Williams, Princeton
Neuroscience Institute and Department of Psychology, Prince-
ton University, Princeton, NJ 08544, or via e-mail: jamalawilliams
@gmail.com.
Author Contributions
Jamal A. Williams: Conceptualization, Data curation, Formal
analysis, Funding acquisition, Investigation, Methodology,
Project administration, Supervision, Validation, Visualization,
Writing—Original Draft, Writing—Review & Editing.
Elizabeth H. Margulis: Supervision, Writing—Original
Draft, Writing—Review & Editing. Samuel A. Nastase:
Writing—Original Draft, Writing—Review & Editing.
Janice Chen: Conceptualization, Methodology, Supervi-
sion, Writing—Review & Editing. Uri Hasson: Conceptual-
ization, Funding Acquisition, Methodology, Supervision.
Kenneth A. Norman: Conceptualization, Funding Acquisi-
tion, Methodology, Project Administration, Supervision,
Writing—Original Draft, Writing—Review & Editing.
Christopher Baldassano: Conceptualization, Formal
analysis, Methodology, Project Administration, Software,
Supervision, Writing—Original Draft, Writing—Review &
Editing.
Funding Information
This work was supported by National Institute of Mental
Health (https://dx.doi.org/10.13039/100000025), grant
number: R01 MH112357-01 to U. H. and K. A. N. and
National Institute of Neurological Disorders and Stroke
(https://dx.doi.org/10.13039/100000065), grant number:
F99 NS118740-01 to J. W.
Data Availability
The fMRI data used in this study have been publicly
released on OpenNeuro (https://openneuro.org/datasets
/ds004007/versions/1.0.2).
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender identi-
fication of first author/last author) publishing in the Journal
of Cognitive Neuroscience ( JoCN) during this period were
M(an)/M = .407, W(oman)/M = .32, M/W = .115, and
W/W = .159, the comparable proportions for the articles
that these authorship teams cited were M/M = .549,
W/M = .257, M/W = .109, and W/W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. The
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .612; W/M = .224;
M/ W = .122; W/ W = .041.
REFERENCES
Abrams, D. A., Ryali, S., Chen, T., Chordia, P., Khouzam, A.,
Levitin, D. J., et al. (2013). Inter-subject synchronization of
brain responses during natural music listening. European
Journal of Neuroscience, 37, 1458–1469. https://doi.org/10
.1111/ejn.12173, PubMed: 23578016
Alluri, V., Toiviainen, P., Jääskeläinen, I. P., Glerean, E., Sams,
M., & Brattico, E. (2012). Large-scale brain networks emerge
from dynamic processing of musical timbre, key and rhythm.
Neuroimage, 59, 3677–3689. https://doi.org/10.1016/j
.neuroimage.2011.11.019, PubMed: 22116038
Andersson, J. L. R., Skare, S., & Ashburner, J. (2003). How to
correct susceptibility distortions in spin-echo echo-planar
images: Application to diffusion tensor imaging.
Neuroimage, 20, 870–888. https://doi.org/10.1016/S1053
-8119(03)00336-7, PubMed: 14568458
Antony, J. W., Hartshorne, T. H., Pomeroy, K., Gureckis, T. M.,
Hasson, U., McDougle, S. D., et al. (2021). Behavioral,
physiological, and neural signatures of surprise during
naturalistic sports viewing. Neuron, 109, 377–390.
https://doi.org/10.1016/j.neuron.2020.10.029, PubMed:
33242421
Asano, R., Boeckx, C., & Seifert, U. (2021). Hierarchical control
as a shared neurocognitive mechanism for language and
music. Cognition, 216, 104847. https://doi.org/10.1016/j
.cognition.2021.104847, PubMed: 34311153
Auerbach, E. J., Xu, J., Yacoub, E., Moeller, S., & Uğurbil, K.
(2013). Multiband accelerated spin-echo echo planar imaging
with reduced peak RF power using time-shifted RF pulses.
Magnetic Resonance in Medicine, 69, 1261–1267. https://doi
.org/10.1002/mrm.24719, PubMed: 23468087
Baldassano, C., Chen, J., Zadbood, A., Pillow, J. W., Hasson, U.,
& Norman, K. A. (2017). Discovering event structure in
continuous narrative perception and memory. Neuron,
95, 709–721. https://doi.org/10.1016/j.neuron.2017.06.041,
PubMed: 28772125
Baldassano, C., Hasson, U., & Norman, K. A. (2018).
Representation of real-world event schemas during narrative
perception. Journal of Neuroscience, 38, 9689–9699.
https://doi.org/10.1523/JNEUROSCI.0251-18.2018, PubMed:
30249790
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false
discovery rate: A practical and powerful approach to multiple
testing. Journal of the Royal Statistical Society, Series B:
Methodological, 57, 289–300. https://doi.org/10.1111/j.2517
-6161.1995.tb02031.x
Ben-Yakov, A., & Henson, R. N. (2018). The hippocampal film
editor: Sensitivity and specificity to event boundaries in
continuous experience. Journal of Neuroscience, 38,
10057–10068. https://doi.org/10.1523/JNEUROSCI.0524-18
.2018, PubMed: 30301758
Blood, A. J., & Zatorre, R. J. (2001). Intensely pleasurable
responses to music correlate with activity in brain regions
712
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
implicated in reward and emotion. Proceedings of the
National Academy of Sciences, U.S.A., 98, 11818–11823.
https://doi.org/10.1073/pnas.191355898, PubMed: 11573015
Castro, M., L’héritier, F., Plailly, J., Saive, A.-L., Corneyllie, A.,
Tillmann, B., et al. (2020). Personal familiarity of music and its
cerebral effect on subsequent speech processing. Scientific
Reports, 10, 14854. https://doi.org/10.1038/s41598-020-71855-5,
PubMed: 32908227
Cauley, S. F., Polimeni, J. R., Bhat, H., Wald, L. L., & Setsompop,
K. (2014). Interslice leakage artifact reduction technique for
simultaneous multislice acquisitions. Magnetic Resonance in
Medicine, 72, 93–102. https://doi.org/10.1002/mrm.24898,
PubMed: 23963964
Chen, J., Leong, Y. C., Honey, C. J., Yong, C. H., Norman, K. A.,
& Hasson, U. (2017). Shared memories reveal shared
structure in neural activity across individuals. Nature
Neuroscience, 20, 115–125. https://doi.org/10.1038/nn.4450,
PubMed: 27918531
Daly, I., Williams, D., Hallowell, J., Hwang, F., Kirke, A., Malik,
A., et al. (2015). Music-induced emotions can be predicted
from a combination of brain activity and acoustic features.
Brain and Cognition, 101, 1–11. https://doi.org/10.1016/j
.bandc.2015.08.003, PubMed: 26544602
Farbood, M. M., Heeger, D. J., Marcus, G., Hasson, U., & Lerner,
Y. (2015). The neural processing of hierarchical structure in
music and speech at different timescales. Frontiers in
Neuroscience, 9, 157. https://doi.org/10.3389/fnins.2015
.00157, PubMed: 26029037
Fedorenko, E., Patel, A., Casasanto, D., Winawer, J., & Gibson,
E. (2009). Structural integration in language and music:
Evidence for a shared system. Memory & Cognition, 37, 1–9.
https://doi.org/10.3758/MC.37.1.1, PubMed: 19103970
Geerligs, L., van Gerven, M., Campbell, K. L., & Güçlü, U. (2021).
A nested cortical hierarchy of neural states underlies event
segmentation in the human brain. bioRxiv, 2021.02.05.429165.
https://doi.org/10.1101/2021.02.05.429165
Geerligs, L., van Gerven, M., & Güçlü, U. (2021). Detecting
neural state transitions underlying event segmentation.
Neuroimage, 236, 118085. https://doi.org/10.1016/j
.neuroimage.2021.118085, PubMed: 33882350
Hasson, U., Chen, J., & Honey, C. J. (2015). Hierarchical process
memory: Memory as an integral component of information
processing. Trends in Cognitive Sciences, 19, 304–313.
https://doi.org/10.1016/j.tics.2015.04.006, PubMed: 25980649
Hasson, U., Yang, E., Vallines, I., Heeger, D. J., & Rubin, N.
(2008). A hierarchy of temporal receptive windows in
human cortex. Journal of Neuroscience, 28, 2539–2550.
https://doi.org/10.1523/jneurosci.5487-07.2008, PubMed:
18322098
Honey, C. J., Thesen, T., Donner, T. H., Silbert, L. J., Carlson,
C. E., Devinsky, O., et al. (2012). Slow cortical dynamics
and the accumulation of information over long timescales.
Neuron, 76, 668. https://doi.org/10.1016/j.neuron.2012.10.024
Jackendoff, R., & Lerdahl, F. (2006). The capacity for music:
What is it, and what’s special about it? Cognition, 100, 33–72.
https://doi.org/10.1016/j.cognition.2005.11.005, PubMed:
16384553
Jantzen, M. G., Large, E. W., & Magne, C. (2016). Overlap of
neural systems for processing language and music. Frontiers
in Psychology, 7, 876. https://doi.org/10.3389/fpsyg.2016
.00876, PubMed: 27378976
Jenkinson, M., Bannister, P., Brady, M., & Smith, S. (2002).
Improved optimization for the robust and accurate linear
registration and motion correction of brain images.
Neuroimage, 17, 825–841. https://doi.org/10.1006/nimg.2002
.1132, PubMed: 12377157
Kell, A. J. E., Yamins, D. L. K., Shook, E. N., Norman-Haignere,
S. V., & McDermott, J. H. (2018). A task-optimized neural
network replicates human auditory behavior, predicts brain
responses, and reveals a cortical processing hierarchy.
Neuron, 98, 630–644. https://doi.org/10.1016/j.neuron.2018
.03.044, PubMed: 29681533
Koelsch, S. (2011). Toward a neural basis of music perception—
A review and updated model. Frontier in Psychology, 2, 110.
https://doi.org/10.3389/fpsyg.2011.00110, PubMed: 21713060
Koelsch, S., Gunter, T. C., v. Cramon, D. Y., Zysset, S.,
Lohmann, G., & Friederici, A. D. (2002). Bach speaks: A
cortical “language-network” serves the processing of music.
Neuroimage, 17, 956–966. https://doi.org/10.1006/nimg.2002
.1154, PubMed: 12377169
Kumar, M., Anderson, M. J., Antony, J. W., Baldassano, C.,
Brooks, P. P., Cai, M. B., et al. (2022). BrainIAK: The brain
imaging analysis kit. Aperture Neuro, 1. https://doi.org/10
.52294/31bb5b68-2184-411b-8c00-a1dacb61e1da
Landemard, A., Bimbard, C., Demené, C., Shamma, S., Norman-
Haignere, S., & Boubenec, Y. (2021). Distinct higher-order
representations of natural sounds in human and ferret
auditory cortex. eLife, 10, e65566. https://doi.org/10.7554
/eLife.65566, PubMed: 34792467
Lee, D. J., Jung, H., & Loui, P. (2019). Attention modulates
electrophysiological responses to simultaneous music and
language syntax processing. Brain Sciences, 9, 305. https://
doi.org/10.3390/brainsci9110305, PubMed: 31683961
Lerner, Y., Honey, C. J., Silbert, L. J., & Hasson, U. (2011).
Topographic mapping of a hierarchy of temporal receptive
windows using a narrated story. Journal of Neuroscience, 31,
2906–2915. https://doi.org/10.1523/jneurosci.3684-10.2011,
PubMed: 21414912
Liu, W., Shi, Y., Cousins, J. N., Kohn, N., & Fernández, G.
(2021). Hippocampal-medial prefrontal event segmentation
and integration contribute to episodic memory formation.
Cerebral Cortex. https://doi.org/10.1101/2020.03.14.990002,
PubMed: 34398213
Margulis, E. H., Wong, P. C. M., Simchy-Gross, R., & McAuley,
J. D. (2019). What the music said: Narrative listening across
cultures. Palgrave Communications, 5, 146. https://doi.org
/10.1057/s41599-019-0363-1
Margulis, E. H., Wong, P. C. M., Turnbull, C., Kubit, B. M. &
McAuley, J. D. (2021). Narratives imagined in response to
music reveal culture-bounded intersubjectivity. Proceedings
of the National Academy of Sciences, U.S.A., 119,
e2110406119.
Mason, M. F., Norton, M. I., Van Horn, J. D., Wegner, D. M.,
Grafton, S. T., & Macrae, C. N. (2007). Wandering minds: The
default network and stimulus-independent thought. Science,
315, 393–395. https://doi.org/10.1126/science.1131295,
PubMed: 17234951
McAuley, J. D., Wong, P. C. M., Mamidipaka, A., Phillips, N., &
Margulis, E. H. (2021). Do you hear what I hear? Perceived
narrative constitutes a semantic dimension for music.
Cognition, 212, 104712. https://doi.org/10.1016/j.cognition
.2021.104712, PubMed: 33848700
McFee, B., Raffel, C., Liang, D., Ellis, D., McVicar, M.,
Battenberg, E., et al. (2015). librosa: Audio and music
signal analysis in python. Proceedings of the 14th Python
in Science Conference. https://doi.org/10.25080/majora
-7b98e3ed-003
Moeller, S., Yacoub, E., Olman, C. A., Auerbach, E., Strupp, J.,
Harel, N., et al. (2010). Multiband multislice GE-EPI at 7 tesla,
with 16-fold acceleration using partial parallel imaging with
application to high spatial and temporal whole-brain fMRI.
Magnetic Resonance in Medicine, 63, 1144–1153. https://doi
.org/10.1002/mrm.22361, PubMed: 20432285
Nakai, T., Koide-Majima, N., & Nishimoto, S. (2021).
Correspondence of categorical and feature-based
representations of music in the human brain. Brain and
Williams et al.
713
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Behavior, 11, e01936. https://doi.org/10.1002/brb3.1936,
PubMed: 33164348
Norman-Haignere, S., Kanwisher, N. G., & McDermott, J. H.
(2015). Distinct cortical pathways for music and speech
revealed by hypothesis-free voxel decomposition. Neuron,
88, 1281–1296. https://doi.org/10.1016/j.neuron.2015.11.035,
PubMed: 26687225
Patel, A. D. (2011). Why would musical training benefit the
neural encoding of speech? The OPERA hypothesis. Frontiers
in Psychology, 2, 142. https://doi.org/10.3389/fpsyg.2011
.00142, PubMed: 21747773
Patil, K., Pressnitzer, D., Shamma, S., & Elhilali, M. (2012).
Music in our ears: The biological bases of musical timbre
perception. PLoS Computational Biology, 8, e1002759.
https://doi.org/10.1371/journal.pcbi.1002759, PubMed:
23133363
Peretz, I., Vuvan, D., Lagroi, M.-E., & Armory, J. L. (2015). Neural
overlap in processing music and speech. Philosophical
Transactions of the Royal Society of London, Series B:
Bilogical Sciences, 370, 20140090. https://doi.org/10.1098
/rstb.2014.0090, PubMed: 25646513
Poorjam, A. H.. (2018). Re: Why we take only 12–13 MFCC
coefficients in feature extraction?. Retrieved from https://
www.researchgate.net/post/ Why_we_take_only_12-13
_MFCC_coefficients_in_feature_extraction
/5b0fd2b7cbdfd4b7b60e9431/citation/download
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. L. (2001). A default mode of
brain function. Proceedings of the National Academy of
Sciences, U.S.A., 98, 676–682. https://doi.org/10.1073/pnas.98
.2.676, PubMed: 11209064
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo,
X.-N., Holmes, A. J., et al. (2018). Local–global parcellation
of the human cerebral cortex from intrinsic functional
connectivity MRI. Cerebral Cortex, 28, 3095–3114. https://doi
.org/10.1093/cercor/bhx179, PubMed: 28981612
Setsompop, K., Gagoski, B. A., Polimeni, J. R., Witzel, T.,
Wedeen, V. J., & Wald, L. L. (2012). Blipped-controlled
aliasing in parallel imaging for simultaneous multislice echo
planar imaging with reduced g-factor penalty. Magnetic
Resonance in Medicine, 67, 1210–1224. https://doi.org/10
.1002/mrm.23097, PubMed: 21858868
Shulman, G. L., Fiez, J. A., Corbetta, M., Buckner, R. L., Miezin,
F. M., Raichle, M. E., et al. (1997). Common blood flow
changes across visual tasks: II. Decreases in cerebral cortex.
Journal of Cognitive Neuroscience, 9, 648–663. https://doi
.org/10.1162/jocn.1997.9.5.648, PubMed: 23965122
Sotiropoulos, S. N., Moeller, S., Jbabdi, S., Xu, J., Andersson,
J. L., Auerbach, E. J., et al. (2013). Effects of image
reconstruction on fiber orientation mapping from
multichannel diffusion MRI: Reducing the noise floor
using SENSE. Magnetic Resonance in Medicine, 70,
1682–1689. https://doi.org/10.1002/mrm.24623, PubMed:
23401137
Sridharan, D., Levitin, D. J., Chafe, C. H., Berger, J., & Menon, V.
(2007). Neural Dynamics of event segmentation in music:
converging evidence for dissociable ventral and dorsal
networks. Neuron, 55, 521–532. https://doi.org/10.1016/j
.neuron.2007.07.003, PubMed: 17678862
Tallal, P., & Gaab, N. (2006). Dynamic auditory processing,
musical experience and language development. Trends in
Neurosciences, 29, 382–390. https://doi.org/10.1016/j.tins
.2006.06.003, PubMed: 16806512
Taruffi, L., Pehrs, C., Skouras, S., & Koelsch, S. (2017). Effects of
sad and happy music on mind-wandering and the default
mode network. Scientific Reports, 7, 14396. https://doi.org/10
.1038/s41598-017-14849-0, PubMed: 29089542
Tillmann, B. (2012). Music and language perception: Expectations,
structural integration, and cognitive sequencing. Topics in
Cognitive Science, 4, 568–584. https://doi.org/10.1111/j.1756
-8765.2012.01209.x, PubMed: 22760955
Toiviainen, P., Alluri, V., Brattico, E., Wallentin, M., & Vuust, P.
(2014). Capturing the musical brain with Lasso: Dynamic
decoding of musical features from fMRI data. Neuroimage,
88, 170–180. https://doi.org/10.1016/j.neuroimage.2013.11
.017, PubMed: 24269803
Woolrich, M. W., Behrens, T. E. J., Beckmann, C. F., Jenkinson,
M., & Smith, S. M. (2004). Multilevel linear modelling for
fMRI group analysis using Bayesian inference. Neuroimage,
21, 1732–1747. https://doi.org/10.1016/j.neuroimage.2003.12
.023, PubMed: 15050594
Woolrich, M. W., Ripley, B. D., Brady, M., & Smith, S. M. (2001).
Temporal autocorrelation in univariate linear modeling of
fMRI data. Neuroimage, 14, 1370–1386. https://doi.org/10
.1006/nimg.2001.0931, PubMed: 11707093
Xu, J., Moeller, S., Auerbach, E. J., Strupp, J., Smith, S. M.,
Feinberg, D. A., et al. (2013). Evaluation of slice accelerations
using multiband echo planar imaging at 3T. Neuroimage, 83,
991–1001. https://doi.org/10.1016/j.neuroimage.2013.07.055,
PubMed: 23899722
Zadbood, A., Chen, J., Leong, Y. C., Norman, K. A., & Hasson, U.
(2017). How we transmit memories to other brains:
Constructing shared neural representations via
communication. Cerebral Cortex, 27, 4988–5000.
https://doi.org/10.1093/cercor/bhx202, PubMed: 28922834
714
Journal of Cognitive Neuroscience
Volume 34, Number 4
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
4
4
6
9
9
1
9
9
6
6
9
5
/
j
o
c
n
_
a
_
0
1
8
1
5
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3