Hemispheric Specialization within the Superior Anterior
Temporal Cortex for Social and Nonsocial Concepts
Gorana Pobric1, Matthew A. Lambon Ralph1, and Roland Zahn1,2
Astratto
■ Studies of semantic dementia, imaging, and repetitive TMS
have suggested that the bilateral anterior temporal lobes (ATLs)
underpin a modality-invariant representational hub within the
semantic system. Questions remain, Tuttavia, regarding func-
tional specialization across a variety of knowledge domains within
the ATL region. We investigated direct evidence for the functional
relevance of the superior ATL in processing social concepts. Using
converging evidence from noninvasive brain stimulation and
neuropsychology, we demonstrate graded differentiation of right
and left superior anterior temporal areas in social cognition.
Whereas the left superior ATL is necessary for processing both
social and nonsocial abstract concepts, social conceptual process-
ing predominates in the right superior ATL. This graded hemi-
spheric specialization is mirrored in the patient results. Our data
shed new light on the classic debate about hemispheric differ-
ences in semantic and social cognition. These results are consid-
ered in the context of models of semantic representation and the
emerging data on connectivity for left and right ATL regions. ■
INTRODUCTION
Human behavior occurs in social contexts. Human neuro-
science needs to understand which brain regions support
social behavior and how they function. Multiple lines of
evidence suggest that the anterior temporal lobes (ATLs)
are part of the network supporting semantic (Lambon
Ralph, 2013) and social cognition (Zahn et al., 2007;
Amodio & Frith, 2006; Moll, Zahn, de Oliveira-Souza,
Krueger, & Grafman, 2005).
The role of the ATL in semantic cognition has been
supported by patient investigations (Patterson, Nestor, &
Rogers, 2007; Snowden, Goulding, & Neary, 1989), neuro-
imaging studies ( Visser, Embleton, Jefferies, Parker,
& Lambon Ralph, 2010; Vandenberghe, Price, Wise,
Josephs, & Frackowiak, 1996), and neurostimulation stud-
ies (Pobric, Jefferies, & Lambon Ralph, 2007, 2010; Lambon
Ralph, Pobric, & Jefferies, 2009). Neuropsychological
evidence comes from patients with semantic dementia
(SD), who have bilateral atrophy and hypometabolism in
the ATLs. In addition to a generalized yet selective seman-
tic impairment, right ATL (rATL) neurodegeneration has
been associated with impairments of social behavior (Chan
et al., 2009), such as lack of empathy (Rankin et al., 2006),
socially inappropriate behavior (Zahn et al., 2009), and the-
ory of mind tasks (Irish, Hodges, & Piguet, 2014), although
with careful assessment, similar neuropsychiatric features
are observed in cases with predominantly left ATL (lATL)
atrophy, only less frequently (Chan et al., 2009).
Studies of patients with SD have given rise to the “hub-
and-spoke” hypothesis in which the ATLs are a key node
1University of Manchester, 2King’s College London
in the semantic network where different inputs converge
to form modality-invariant conceptual representations
(Lambon Ralph, Sage, Jones, & Mayberry, 2010; Patterson
et al., 2007; Rogers & McClelland, 2004). Given that SD is
a neurodegenerative condition, there is always the pos-
sibility that semantic dysfunction follows from pathology
extending beyond the ATL, and it is this more subtle,
widespread damage that is the cause of the patients’ se-
mantic impairment (Martin, 2007). Inoltre, because
SD is characterized by bilateral atrophy, albeit sometimes
very asymmetric in the early phase, it is not possible to
investigate the contribution of lATL and rATL in isolation.
Given these facts, the contributions of the ATL to seman-
tic processing cannot be uniquely defined on the basis
of this neuropsychological evidence alone.
Neuroimaging studies of semantic cognition have
highlighted the role of the left hemisphere in the com-
prehension and production of spoken language ( Wise,
2003). Inoltre, the importance of left anterior
temporal cortex for language processing and semantic
memory has been highlighted with PET (Devlin et al.,
2000; Vandenberghe et al., 1996), intracranial recordings
(Halgren, Baudena, Heit, Clarke, & Marinkovic, 1994),
and MEG (Marinkovic et al., 2003). Bilateral ATL activa-
tions have been observed in semantic tasks for words
(Binney, Embleton, Jefferies, Parke, & Lambon Ralph,
2010; Zahn et al., 2007), sounds, spoken names, and pic-
tures ( Visser & Lambon Ralph, 2011; Sharp, Scott, &
Wise, 2004). The neuroimaging literature of social cog-
nition has reported a right hemispheric bias for social
stimuli (Skipper, Ross, & Olson, 2011; Zahn et al.,
2007). Zahn and colleagues (2007) have suggested that
the right superior ATL (sATL) selectively represents social
© 2016 Istituto di Tecnologia del Massachussetts. Published under a
Creative Commons Attribution 3.0 Unported (CC BY 3.0) licenza
Journal of Cognitive Neuroscience 28:3, pag. 351–360
doi:10.1162/jocn_a_00902
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
C
7
N
6
_
9
UN
/
_
j
0
o
0
C
9
N
0
2
_
UN
P
_
D
0
0
B
9
sì
0
G
2
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
/
.
T
F
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
concepts. When participants were making semantic judg-
ments about social concepts (per esempio., “honor–brave”) versus
animal function concepts (per esempio., “trainable–ridden”), right
sATL regions were activated. This was independently
corroborated in a recent study (Skipper et al., 2011)—
although the peak activations were found in the lATL
rather than rATL region in another study (Ross & Olson,
2010), calling into question the strength of ATL laterality
for social conceptual processing. Inoltre, ATL activa-
tions have been reported for other high-level social verbal
and nonverbal tasks: moral judgments (Moll et al., 2005),
social versus nonsocial gestures (Straube, Verde, Jansen,
Chatterjee, & Kircher, 2010), social attribution stimuli
such as Heider and Simmel animations (Ross & Olson,
2010), and socioemotional stories (Ferstl & von Cramon,
2002). This consistency of ATL activations to high-level
social tasks irrespective of the modality of the task (verbal
or nonverbal) has led to its inclusion in the “social brain
network” (Frith & Frith, 2012; Amodio & Frith, 2006).
It is clear that diverse social stimuli and tasks evoke
activations in the ATLs. Tuttavia, the precise role of
the ATLs in the network remains controversial (Simmons,
Reddish, Bellgowan, & Martin, 2010). By combining neuro-
stimulation with patient investigation, we addressed three
key limitations in the literature: (1) Most patients with
neurodegeneration show widespread damage that is not
only confined to subregions of the ATL unilaterally; (2)
fMRI does not provide causal evidence for the necessity
of activated regions in the task of interest; E (3) A
our knowledge, no study has directly compared process-
ing of social concepts in left and right sATL.
Primo, we established the necessary role of sATL in pro-
cessing social versus nonsocial abstract concepts via TMS.
We predicted that, if the rATL is involved in processing
social concepts, as observed in patients with rATL neuro-
degeneration, then repetitive transcranial magnetic stim-
ulation (rTMS) should generate a greater effect on social
than nonsocial concepts. If the lATL or rATL is implicated
in general semantic processing, but not general cognition
(as per patients with SD), then stimulation should impact
on performance with both social and nonsocial abstract
concepts but not on difficulty-matched nonsemantic
compiti. In the neuropsychological part of the study, we
predicted the lATL case would be impaired on processing
both social and nonsocial concepts, in keeping with gen-
eral degradation of semantic knowledge observed in pa-
tients with SD. On the basis of previous patient (Zahn
et al., 2009) and neuroimaging evidence (Skipper et al.,
2011; Zahn et al., 2007), we predicted that the rATL case
would exhibit a greater impairment on processing social
versus nonsocial concepts.
METHODS
To establish a necessary role of the ATLs in representing
semantic knowledge of social and nonsocial concepts, we
investigated convergent evidence from TMS and neuro-
psicologia. TMS is a well-established, noninvasive tech-
nique that induces electrical activation in the underlying
cortex. A long train of low-frequency stimulation tempo-
rarily suppresses neural processing and disrupts behav-
ioral tasks that rely on that cortical region (Walsh &
Cowey, 2000). Cortical sites for TMS were left and right
sATL and the occipital pole as a control site. Sites were
identified in each individual participant through coregistra-
tion with their MRI scan. In a semantic synonym judgment
task, participants were required to pick which of two con-
cepts (per esempio., “honest” vs. “eager”) is more closely related to
the probe (“sincere”). Our control task was a difficulty-
matched, number magnitude judgment assessment. Per-
formance was assessed before and after TMS stimulation.
In the neuropsychological part of the study, we used a
semantic discrimination task, which probed knowledge of
social and animal function concepts in two rare cases of
focal neurodegeneration of the lATL and rATL, rispettivamente.
Design
UN 3 × 3 × 2 repeated-measures design was used, con
site (right sATL vs. left sATL vs. occipital pole), task (non-
social synonym judgment vs. social synonym judgment vs.
magnitude judgment), and TMS (no TMS vs. rTMS stimu-
lation) as the three within-participant factors. The study
utilized rTMS, using the “virtual lesion” method in which
the train of rTMS is delivered offline (without a concur-
rent behavioral task), and then behavioral performance
is probed during the temporary refractory period and
compared with performance on the same task outside
this refractory window.
Participants
Twelve right-handed participants took part in the experi-
ment (seven women; mean age = 25.8 years, SD = 3.8 years).
All participants were strongly right-handed, with a lateral-
ity quotient above 75 (mean = 88.33, SD = 6.24) on the
Edinburgh Handedness Inventory (Oldfield, 1971) E
had normal or corrected-to-normal vision. All participants
provided written consent for participation after being
screened for adverse effects of TMS. The experiment
was approved by the local ethics committee.
Stimuli
One hundred sixty concept trials were used in the syno-
nym judgment task, E 80 number trials were used in
the number judgment task. Each trial contained three
concepts: a probe concept (per esempio., “sincere”), a target choice
(per esempio., “honest”), and an unrelated foil (per esempio., “eager”). Tutto
words within a triad were matched for frequency (mean =
28.49, SD = 11.23; Kučera & Francis, 1967) and imageability
352
Journal of Cognitive Neuroscience
Volume 28, Numero 3
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
C
7
N
6
_
9
UN
/
_
j
0
o
0
C
9
N
0
2
_
UN
P
_
D
0
0
B
9
sì
0
G
2
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
F
.
T
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
(mean = 275.71, SD = 43.72; Bird, Franklin, & Howard,
2001). All triads were divided into social and nonsocial
trials based on ratings from independent evaluators.
Item ratings were obtained for 450 words by asking 40
undergraduate students (8 men and 32 women) whose
ages ranged from 18 A 32 (mean = 25) years to rate the
words based on how social they are. Students were given
a 7-point scale (1 = very nonsocial to 7 = very social )
and asked to indicate the rating of each word. Students
were given examples of possible ratings for words not
included in the set. From that set, we created 80 social
E 80 nonsocial trials. For nonsocial trial, the mean rating
era 2.34, with SD = 0.60 and range = 1.73–2.94. For social
trial, the mean rating was 5.77, with SD = 0.79 and range =
5.02–6.56. Consequently, social and nonsocial trials were
matched on psycholinguistic variables as well as behav-
ioral measures: RTs and accuracy in pilot experiments.
The format for the number task was the same as for the
synonym judgment task: A three-digit probe number was
presented at the top of the screen, and underneath,
2 three-digit number choices were given. Participants were
required to pick which of the two numbers was closest in
value to the probe. The pilot studies revealed that making
magnitude judgments about three-digit numbers was the
best behavioral match (in terms of RTs and accuracy) for
our synonym task.
TMS
A MagStim Rapid2 (Magstim Co., Whitland, UK) stim-
ulator with two external boosters was used (maximum
output = approx. 2.2 T). Magnetic stimulation was applied
using a 50-mm figure-of-eight coil.
Selection of TMS Site
The structural T1-weighted MRI scans were coregistered with
the participant’s scalp using MRIreg (www.mccauslandcenter.
sc.edu/mricro/mricron/index.html). Immediately before
the TMS session, scalp coordinates were measured using
an Ascension Minibird (www.ascension-tech.com) mag-
netic tracking system. The right Montreal Neurological
Institute coordinates for the sATL (53, 8, −13) were ob-
tained by averaging peak coordinates from previous fMRI
studies associating the right sATL with knowledge of social
concepts (Zahn et al., 2007: 51, 15, −12; (Zahn et al., 2009:
54, 0, −3) and peak coordinates of resting state glucose
hypometabolism from a study of social concepts in fronto-
temporal lobar degeneration (FTLD; 54, 9, −24: Zahn et al.,
2009). The coordinates for the left sATL corresponded to
the homologue region (−53, 8, −13). A middle occipital
stimulation site (Oz) was also employed as a site to control
for possible nonspecific TMS effects.
Task and Procedure
Stimulation Parameters
A PC running E-Prime software (Psychology Software
Tools, Inc., Pittsburgh, PAPÀ) allowed the presentation of
stimuli and recording of the responses. In a single exper-
imental session, participants performed a synonym judg-
ment task and a number magnitude judgment task. In
a semantic synonym judgment task, participants were
required to pick which of two words (per esempio., “honest” vs.
“eager”) is more closely related to the probe (“sincere”).
The probe is presented at the top of the screen, and the
two word options are presented underneath. The par-
ticipants indicated their choice by using both hands for
pressing a single key (M or Z). Half of the targets were
associated with each key, so they used each hand for
50% of the answers. The occurrence of target location
was randomized for each participant and each session.
The experiment began with a practice block of 10 trials
for each stimulus set. Experimental trials were presented
in a random order in two blocks (80 synonyms and 40 num-
bers). Dopo 10 min of offline rTMS, another two blocks
(80 synonyms and 40 numbers) followed. This yielded
240 trials per experimental session. The blocks were ran-
domized across participants. Stimuli were presented until
the response was given (maximum duration = 3000 msec)
and followed by a blank screen (duration = 500 msec).
Computer key presses were recorded using a computer
keyboard that was placed in front of each participant.
Response latencies were recorded by the computer, E
the accuracy was checked in the analyses.
Individual active motor threshold was determined for
every participant. rTMS was delivered offline for 10 min
at 1 Hz (600 sec at 120% motor threshold level) applied
to the right sATL, left sATL, and Oz. The coil was securely
held by experimenter, centered over the site to be stim-
ulated. The average stimulation intensity during rTMS
era 64% of the machine output.
NEUROPSYCHOLOGICAL INVESTIGATION
Patients
Two female (age = 60 E 61 years at the time of testing,
education = 13 E 14 years) right-handed patients with
FTLD according to Lund–Manchester Consensus criteria
(Neary et al., 1998) were selected from a larger case
series because they exhibited the rare picture of pre-
dominant atrophy of the lATL versus rATL, rispettivamente.
The Lund–Manchester Consensus criteria were previ-
ously shown to correlate highly with postmortem neuro-
pathological diagnoses (Snowden et al., 2011). The study
was approved by the South Manchester NHS Research
Ethics Committee, and the patients gave written informed
consent. In addition to extensive background neuro-
psychological testing, a neurological examination, E
an extensive neuropsychiatric interview with their care-
givers, they received the social concept discrimination
task (Zahn et al., 2009). Both patients showed features of
SD as classified by a senior neuropsychiatrist (R. Z.) E
Pobric, Lambon Ralph, and Zahn
353
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
C
7
N
6
_
9
UN
/
_
j
0
o
0
C
9
N
0
2
_
UN
P
_
D
0
0
B
9
sì
0
G
2
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
F
T
.
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
senior neuropsychologist (M. UN. l. R.), but the rATL case
presented primarily with behavioral features. Data for
healthy older adults are taken from published test norms
E, for the same test, from Zahn et al. (2009).
Materials and Procedure
The semantic task consisted of two carefully matched
conditions to compare concepts describing social be-
havior (cioè., social concepts) and concepts describing
nonsocial behavior or properties of animals (cioè., animal
function concepts) of equal familiarity and descriptive-
ness (see previous description in Zahn et al., 2009). UN
prime word describing a concept (per esempio., “adventurous”)
was presented at the top of a screen, and participants
had to decide which of two concepts underneath was
more related in meaning to the prime concept at the
top. Target (per esempio., “courage”) and distracter (per esempio., “con-
trolled”) concepts were chosen from the same category
as the prime concepts (per esempio., animal function concept
prime: “trainable,” target: “ridden,” distracter: “bites”)
and had been used in our previous fMRI studies (Ross
& Olson, 2010; Zahn et al., 2007). Seventy-three items
con 80% response agreement in aged controls were
presented randomly (25 positive social concept triads,
24 negative social concept triads, E 24 animal function
concept triads).
RESULTS
RT Analyses
The RTs for all participants and all conditions were sub-
mitted to a repeated-measures ANOVA with three within-
participant factors: Task (nonsocial synonym judgment,
social synonym judgment, and magnitude judgment),
Site (right sATL, left sATL, and Oz), and TMS (no TMS
vs. rTMS). We removed 16 items from the synonym task
(11 nonsocial and 5 social) and two items from the
number task that yielded over 50% errors in the no-TMS
condition. This resulted in removal of 7.9% of items. IL
main effects of Task, Site, and TMS were not significant.
We observed a significant interaction between Task and
TMS (F = 36.108, df = 2, 22, P < .001). We also report
a significant interaction between Task and Site (F =
4.196, df = 2, 22, p = .029). Crucially, we observed a
significant three-way interaction between Task, Site, and
TMS (F = 5.259, df = 4, 44, p = .001). To explore the
three-way interaction further, we split the original anal-
ysis by task and carried out three repeated-measures
ANOVA with two within-participant factors Site (right sATL,
left sATL, and Oz) and TMS (no TMS vs. rTMS). For the
nonsocial task, main effects of Site and TMS were not sig-
nificant. However, we observed a significant interaction
between Site and TMS (F = 7.279, df = 2, 22, p = .004).
For the social task, we observed a main effect of TMS
(F = 14.994, df = 1, 11, p = .003) and a significant inter-
action between TMS and Site (F = 6.005, df = 2, 22, p =
.008). For the number task, we only observed a main
effect of the TMS (F = 10.698, df = 1, 11, p = .007).
Planned t tests on the RTs were used to compare perfor-
mance in rATL and lATL for social and nonsocial concepts.
After controlling for false discovery rate (Benjamini &
Hochberg, 1995), there was a difference for social and
nonsocial concepts between rATL and lATL. The rATL
was more involved in processing social than nonsocial
concepts, with a significant TMS effect for social concepts,
t(11) = 3.713, p < .01, but not for nonsocial concepts,
t(11) = 1.67, p > .05, whereas the lATL was involved
in both social and nonsocial concepts (T(11) = 3.63,
P < .004, and t(11) = 2.285, p < .05, respectively; see
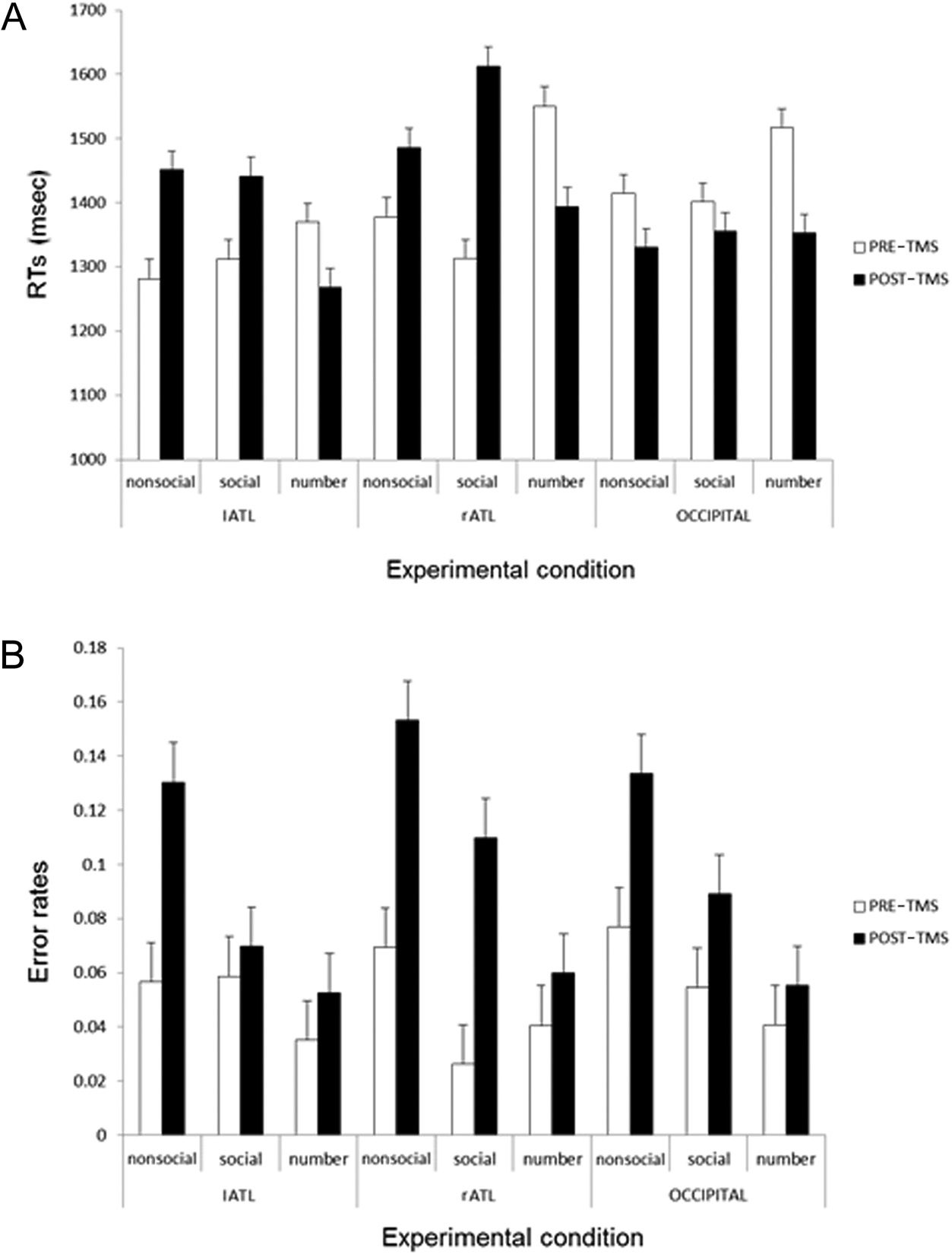
Figure 1A).
Error Analyses
The error rate was examined in a repeated-measures
ANOVA with Task (synonym judgment, number judgment),
Site (right sATL, left sATL, and Oz), and TMS (no TMS vs.
rTMS) as factors. There was a main effect of Task (F =
6.341, df = 2, 22, p = .007) as well as a main effect of
TMS (F = 20.982, df = 1, 11, p = .001). We also observed
a significant interaction between Task and TMS (F =
6.613, df = 2, 22, p = .011). All other main effects or in-
teractions were not significant ( p > .1). To explore the
task-and-TMS interaction further, we split the original anal-
ysis by task and carried out three repeated-measures
ANOVA with two within-participant factors Site (right
sATL, left sATL, and Oz) and TMS (no TMS vs. rTMS).
For the nonsocial task, the main effect of TMS was signif-
icant (F = 14.88, df = 1, 11, p = .003). All other main
effects or interactions were not significant ( p > .1). For
the social task, we observed the main effect of TMS
(F = 15.604, df = 1, 11, p = .002), whereas the main ef-
fect of Site and Site x-TMS interaction were not significant
( p > .1). For the number task, all main effects or interac-
tions were not significant ( p > .1; see Figure 1B).
Neuropsychological Results
Neuropsychological data were analyzed using an estab-
lished method for directly comparing two single cases
with each other (C_CTC; Crawford, Garthwaite, & Legna,
2010). We report one-tailed p values (lower performance
on social concepts relative to animal function concepts
in patients with rATL damage compared with patients
with no damage to this region). We also compared the
performance of patients with the control group by using
an established method for individual patient-based anal-
ysis (Singlims: Crawford & Garthwaite, 2002). Both the
rATL and lATL cases were impaired relative to the con-
trols on social and nonsocial concepts (all t(29)s >
2.35, all ps < .013, one-tailed). However, the difference
in impairment between social and nonsocial concepts
354
Journal of Cognitive Neuroscience
Volume 28, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
c
7
n
6
_
9
a
/
_
j
0
o
0
c
9
n
0
2
_
a
p
_
d
0
0
b
9
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. Effect of rTMS on
synonym judgment and
magnitude judgment tasks
for RTs (A) and error rates
(B). Each bar represents
the mean decision time
alongside the corresponding
standard error adjusted for
within-participant comparisons
(Loftus & Masson, 1994) for
each condition. Occipital =
occipital pole.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
c
7
n
6
_
9
a
/
_
j
0
o
0
c
9
n
0
2
_
a
p
_
d
0
0
b
9
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
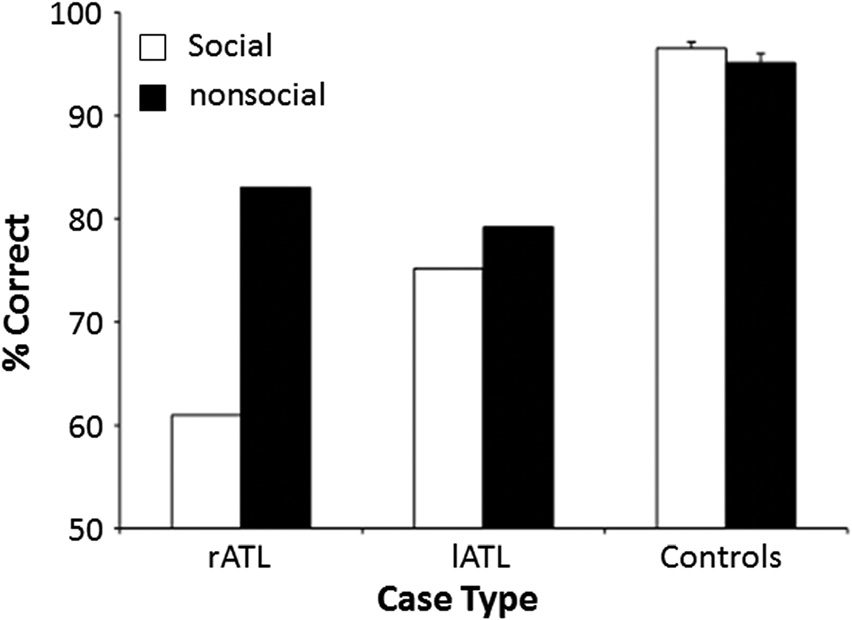
Figure 2. Overall performance of the lATL patient and rATL patient
compared with the control group on social and nonsocial concepts. Error
bars indicate SEM on the control group (n = 30) performance.
Pobric, Lambon Ralph, and Zahn
355
was significantly larger for the rATL case than for the lATL
case, t(29) = 3.0, p < .005 (see Figure 2).
The similarities between our TMS data and neuro-
psychological investigation can be observed in Figure 3.
DISCUSSION
In this study, we were able to ascertain differential con-
tributions of left and right sATL regions to social cogni-
tion and general semantic processing. The left sATL
region was implicated in both social and nonsocial con-
cepts. In contrast, the right sATL made a greater contri-
bution to social than nonsocial concepts. Our results are
in agreement with the central role of the ATL in repre-
senting conceptual knowledge (Lambon Ralph, 2013;
Patterson et al., 2007; Spitsyna, Warren, Scott, Turkheimer,
& Wise, 2006; Bozeat, Lambon Ralph, Patterson, Garrard,
& Hodges, 2000) and the importance of the right tempo-
ral lobe for social cognitive impairments in rATL neuro-
degeneration (Chan et al., 2009; Liu et al., 2004).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
c
7
n
6
_
9
a
/
_
j
0
o
0
c
9
n
0
2
_
a
p
_
d
0
0
b
9
y
0
g
2
u
.
e
p
s
t
d
o
f
n
b
0
y
7
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
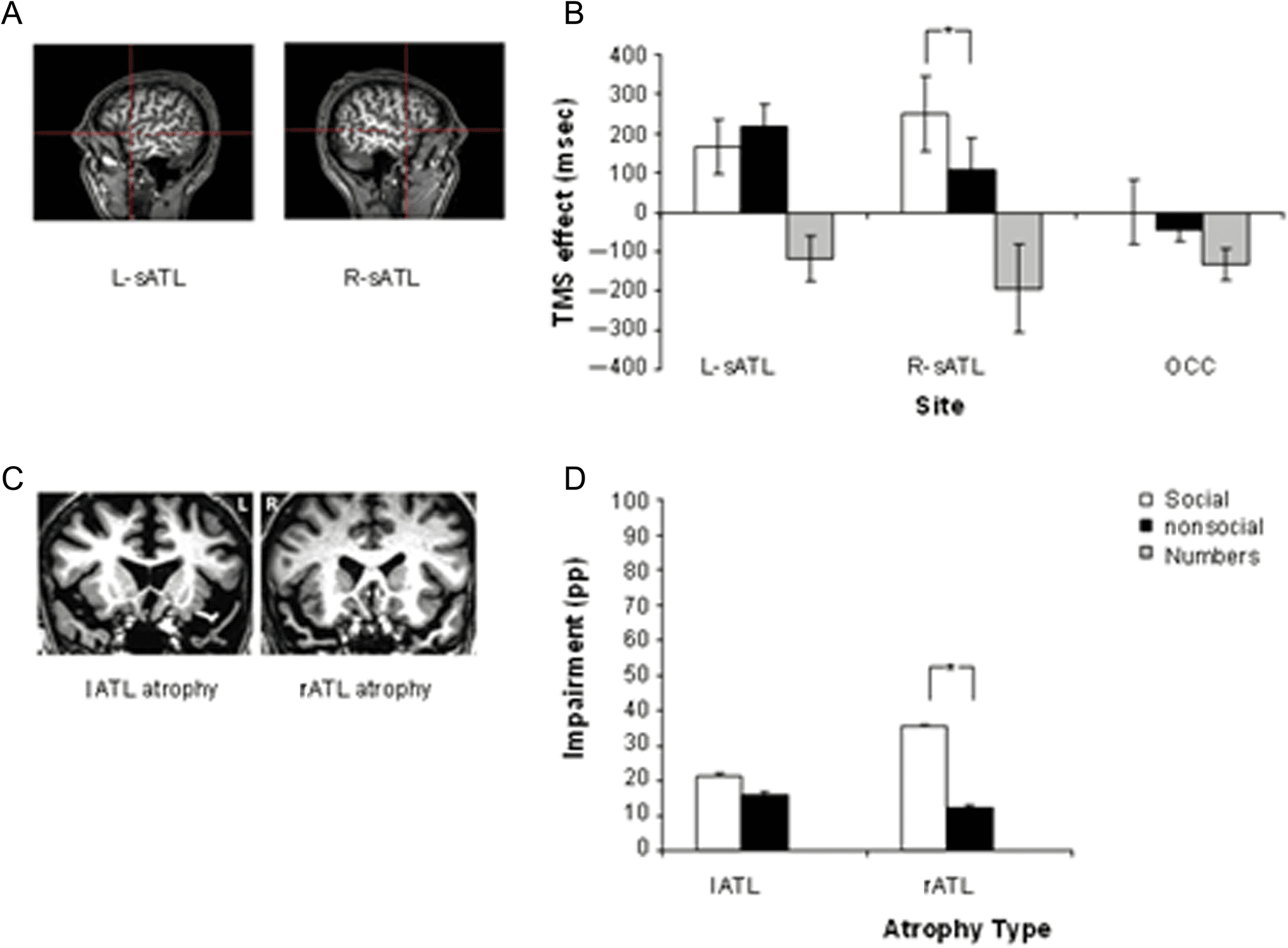
Figure 3. Stimulation sites, TMS results, lesion sites, and patient performance data on social and nonsocial concept task conditions. (A) ATL regions
that were disrupted with TMS, shown for one participant. (B) The TMS effect on semantic judgments of social and nonsocial concepts and
magnitude judgment of numbers. Each bar represents the TMS effect as RT change caused by TMS. Error bars indicate SEM adjusted to reflect
the between-condition variance used in repeated-measure designs (Loftus & Masson, 1994). (C) MRI images for patients with lATL- and rATL-
predominant SD. (D) Overall impairment in percentage points (% correct control − % correct patient) of the lATL patient and rATL patient compared
with the control group on social and nonsocial concepts. Error bars indicate SEM on the control group (n = 30) performance. L-sATL = left sATL;
R-sATL = right sATL; OCC = occipital pole; LATL atrophy = patient with predominantly lATL atrophy; RATL atrophy = patient with predominantly
rATL atrophy. Asterisk denotes significantly slower RTs after rTMS than at baseline. Asterisk denotes significantly worse performance on a task.
In a review of the role of superior temporal areas
(STS), Hein and Knight (2008) proposed that the same
brain region can support different cognitive functions
determined by a task-dependent network of connections.
Indeed, this hypothesis has been formally explored pre-
viously for the ATLs and semantic processing, using com-
putational models (Schapiro, McClelland, Welbourne,
Rogers, & Lambon Ralph, 2013; Lambon Ralph, McClelland,
Patterson, Galton, & Hodges, 2001). Most relevant to
the current findings, Lambon Ralph et al. (2001) demon-
strated that a bilateral ATL semantic system exhibits a left-
sided sensitivity to language (speech production) as a
consequence of greater connectivity of the lATL to left-
hemisphere language centers. Following the same line of
argument, the differential involvement of left and right
sATL regions in social and nonsocial conceptual process-
ing could be explained by the specific pattern of connec-
tions between the ATL regions with the rest of the brain.
Indeed, Skipper et al. (2011) have already proposed that
a greater role for the right temporal region in social pro-
cessing might reflect a right-sided pattern of connectivity
to social-processing regions.
These proposals are consistent with the emerging story
from recent white matter connectivity studies. Specifically,
the temporal pole has dense connections with fusiform
gyrus, limbic structures, and pFC (Pascual et al., 2014;
Gloor, 1997). These brain areas are involved in linguistic,
social, and emotional processing as well as decision-making
(Simmons et al., 2010; Gloor, 1997). In an in vivo probabi-
listic tractography study, Binney, Parker, and Lambon
Ralph (2012) showed that there is strong intratemporal
lobe ATL white matter connectivity, conducive to the infor-
mation integration required for the extraction of modality-
invariant, general semantic representations. Furthermore,
356
Journal of Cognitive Neuroscience
Volume 28, Number 3
there are more regionally specific connectivity patterns
to nontemporal lobe areas. Relevant to the formation of
social concepts, the sATL and temporal pole are connected
to the frontal lobe and OFC via the uncinate fasciculus
(UF), a bidirectional white matter pathway. In a recent
study, Catani and colleagues (2013) used tractography to
show that damage to UF correlated with deficits in seman-
tic processing in patients with primary progressive aphasia.
It is quite possible, therefore, that the sATL-rTMS effect on
social concepts observed in this study reflects either the
convergence and representation of socially relevant infor-
mation at the sATL site and/or the perturbation of informa-
tion transmission between ATL and orbitofrontal regions
(Quentin, Chanes, Migliaccio, Valabrègue, & Valero-Cabré,
2013; Neubert, Mars, Buch, Olivier, & Rushworth, 2010).
The graded, differential roles of lATL and rATL in social
and general concepts may well result from the pattern of
connectivity. As noted above, the role of sATL in social
concepts (left and right) probably reflects its connectivity
between temporal cortex and OFC (Pascual et al., 2014;
Catani et al., 2013; Binney et al., 2012; Moran, Mufson, &
Mesulam, 1987). After previous proposals and computa-
tional demonstrations (Lambon Ralph et al., 2001), the
graded functional differences between left and right
probably reflect (i) a right > left bias in UF density (Highley,
Walker, Esiri, Crow, & Harrison, 2002)—which will encour-
age a rightward division of labor for social conceptual
processing—and (ii) greater left-sided language connec-
attività (both “dorsal” arcuate fasciculus and “ventral” medial
longitudinal fasciculus/extreme capsule complex language
pathways are left hemisphere biased; Parker et al., 2005).
The bilateral ATLs play a crucial role in semantic mem-
ory (Lambon Ralph et al., 2009; Patterson et al., 2007). It
is undisputed that the left hemisphere has a central role
in the comprehension and production of spoken lan-
guage ( Wise, 2003). Inoltre, imaging studies show
that abstract nouns produce greater activation almost
exclusively in the left hemisphere in superior temporal
and inferior frontal cortex (Binder, Desai, Graves, &
Conant, 2009; Spitsyna et al., 2006; Sabsevitz, Medler,
Seidenberg, & Binder, 2005). This finding has been
confirmed by neurostimulation (Hoffman, Jefferies, &
Lambon Ralph, 2010) and patient studies (Hoffman,
Pobric, & Lambon Ralph, 2012). Inoltre, patients with
lATL neurodegeneration also exhibit behavioral prob-
lems such as behavioral rigidity, disinhibition, apathy,
and obsessional behavior as well as mood changes such
as depression (Chan et al., 2009). Tuttavia, these prob-
lems are often masked and underreported given the pre-
dominant language problems associated with lATL
neurodegeneration. Although the left UF connects lATL
with pFC, it has been shown that direct stimulation of the
left uncinate does not result in general language impair-
menti (Duffau, Gatignol, Moritz-Gasser, & Mandonnet,
2009). Therefore, one possibility is that the lATL is special-
ized for verbal semantic processing because language
representations are left lateralized: Per esempio, using
a connectionist model, Lambon Ralph et al. (2001) ac-
counted for left–right asymmetries in picture naming in
terms of stronger connections between lATL and speech
output processes in the left hemisphere.
Extending this hypothesis, it is possible that the right
temporal lobe might make a greater contribution to
social semantic processing if the socially relevant infor-
mation that interacts with semantic representations
comes more strongly from right than left posterior re-
gions or if the preferential left-hemisphere language con-
nectivity induces a division of labor across the ATLs such
that the rATL defaults to social processing because of
connections to right prefrontal and orbitofrontal regions.
This is in accordance with a recent meta-analysis of func-
tional imaging studies of conceptual knowledge, Quale
argues for a graded version of the hub-and-spoke ac-
count, whereby the representation of conceptual knowl-
edge is supported by bilateral yet graded connectivity
between the ATLs and modality-specific sensory, motor,
and limbic cortices (Rice, Lambon Ralph, & Hoffman,
2015).
Although we used SD as a test case for patient perfor-
mance, these results have broader implications and are
consistent with social processing deficits in other neuro-
logical and psychiatric syndromes. Specifically, it is well
established that individuals with psychopathy have
marked difficulties in processing abstract concepts (Kiehl
et al., 2004). Subsequently, in an fMRI study, decreased
rATL activations were reported when psychopaths were
making lexical decisions about abstract concepts. In addi-
zione, in this group, ATLs are characterized by structural
abnormalities such as reduced gray matter volume, In-
cluding weaker connections from rATL to orbitofrontal
regions (Craig et al., 2009).
As well as giving insights about the role of lATL and
rATL regions in social and general semantic concepts,
the current results provide a new perspective on the rich
set of clinical results. Detailed studies of left and right
temporal lobes, FTLD (SD) cases have tended to empha-
size the fact that left-sided cases have a predominantly
language-focussed presentation; and the right-sided
patients, a social deficit (Chan et al., 2009; Zahn et al.,
2009; Rosen et al., 2005; Mugnaio, Darby, Benson, Cummings,
& Mugnaio, 1997). Consistent with the results from the cur-
rent study and implications of the white matter neuro-
anatomy reviewed above, Hornberger, Geng, and Hodges
(2011) found that patients with SD had decreased frac-
tional anisotropy values in the left and right UF, compared
with the healthy controls. Yet, only in patients with rATL
neurodegeneration did decreased fractional anisotropy
values correlate with behavioral symptoms. The clinical
literature does not support, Tuttavia, an absolute dis-
sociation between general versus social conceptual rep-
resentations in lATL versus rATL regions, rispettivamente.
Infatti, in a more formal assessment of neuropsychiatric
symptoms, Chan et al. (2009) found that both patients with
predominantly left- and right-sided ATL were reported to
Pobric, Lambon Ralph, and Zahn
357
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
C
7
N
6
_
9
UN
/
_
j
0
o
0
C
9
N
0
2
_
UN
P
_
D
0
0
B
9
sì
0
G
2
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
F
T
.
/
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
have social and behavioral impairments to different de-
grees. Our study validates these clinical studies (Chan
et al., 2009; Zahn et al., 2009) by combining approaches
from classical neuropsychology and cognitive neuro-
science; we demonstrate graded hemispheric speciali-
zation of ATL regions to social cognition and general
semantic knowledge.
Ringraziamenti
We thank Helen Beaumont, Dr. Sophie Green, and Dr. Paul
Hoffman for administering neuropsychological tests. We thank
the patients and controls for their continued support of our
research. This research was supported by MRC program grants
(G0501632 and MR/ J004146/1) to M. UN. l. R. and an MRC
Clinician Scientist Fellowship (G0902304) to R. Z.
Reprint requests should be sent to Dr. Gorana Pobric, Neuro-
science and Aphasia Research Unit, School of Psychological
Scienze, University of Manchester, Oxford Road, Manchester,
M13 9PL, United Kingdom, or via e-mail: gorana.pobric@
manchester.ac.uk.
REFERENCES
Amodio, D. M., & Frith, C. D. (2006). Meeting of minds: IL
medial frontal cortex and social cognition. Nature Reviews
Neuroscience, 4, 268–277.
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false
discovery rate: A practical and powerful approach to multiple
testing. Journal of the Royal Statistical Society, Series B,
57, 289–300.
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, l. l. (2009).
Where is the semantic system? A critical review and
meta-analysis of 120 functional neuroimaging studies.
Cerebral Cortex, 19, 2767–2796.
Binney, R. J., Embleton, K. V., Jefferies, E., Parke, G. J. M., &
Lambon Ralph, M. UN. (2010). Ventral and inferolateral aspects
of the anterior temporal lobe are crucial in semantic
memory: Evidence from a novel direct comparison of
distortion-corrected fMRI, rTMS, and semantic dementia.
Cerebral Cortex, 20, 2728–2738.
Binney, R. J., Parker, G. J., & Lambon Ralph, M. UN. (2012).
Convergent connectivity and graded specialization in
the rostral human temporal lobe as revealed by
diffusion-weighted imaging probabilistic tractography.
Journal of Cognitive Neuroscience, 10, 1998–2014.
Bird, H., Franklin, S., & Howard, D. (2001). Age of acquisition
and imageability ratings for a large set of words, including
verbs and function words. Behavior Research Methods,
Instruments, & Computers, 33, 73–79.
Bozeat, S., Lambon Ralph, M. A., Patterson, K., Garrard, P.,
& Hodges, J. R. (2000). Non-verbal semantic impairment
in semantic dementia. Neuropsychologia,
38, 1207–1215.
Catani, M., Mesulam, M. M., Jakobsenm, E., Malik, F.,
Martersteck, A., Wieneke, C., et al. (2013). A novel frontal
pathway underlies verbal fluency in primary progressive
aphasia. Brain, 136, 2619–2628.
Chan, D., Anderson, V., Pijnenburg, Y., Whitwell, J., Barnes, J.,
Scahill, R., et al. (2009). The clinical profile of right temporal
lobe atrophy. Brain, 132, 1287–1298.
Craig, M. C., Catani, M., Deeley, Q., Latham, R., Daly, E., Kanaan,
R., et al. (2009). Altered connections on the road to
psychopathy. Molecular Psychiatry, 14, 946–953.
Crawford, J. R., & Garthwaite, P. H. (2002). Investigation of
the single case in neuropsychology: Confidence limits on the
abnormality of test scores and test score differences.
Neuropsychologia, 40, 1196–1208.
Crawford, J. R., Garthwaite, P. H., & Legna, l. T. (2010).
Inferential methods for comparing two single cases.
Cognitive Neuropsychology, 27, 377–400.
Devlin, J. T., Russell, R. P., Davis, M. H., Price, C. J., Wilson,
J., Moss, H. E., et al. (2000). Susceptibility-induced loss of
signal: Comparing PET and fMRI on a semantic task.
Neuroimage, 11, 589–600.
Duffau, H., Gatignol, P., Moritz-Gasser, S., & Mandonnet, E.
(2009). Is the left uncinate fasciculus essential for language?
A cerebral stimulation study. Journal of Neurology, 256,
382–389.
Ferstl, E. C., & von Cramon, D. Y. (2002). What does
the frontomedian cortex contribute to language
processing: Coherence or theory of mind? Neuroimage,
17, 1599–1612.
Frith, C. D., & Frith, U. (2012). Mechanisms of social cognition.
Annual Review of Psychology, 63, 287–313.
Gloor, P. (1997). The temporal lobe and the limbic system.
Oxford: Oxford University Press.
Halgren, E., Baudena, P., Heit, G., Clarke, M., & Marinkovic, K.
(1994). Spatiotemporal stages in face and word-processing:
1. Depth recorded potentials in the human occipital and
parietal lobes. Journal of Physiology-Paris, 88, 1–50.
Hein, G., & Knight, R. T. (2008). Superior temporal sulcus—It’s
my area: Or is it? Journal of Cognitive Neuroscience,
20, 2125–2136.
Highley, J. R., Walker, M. A., Esiri, M. M., Crow, T. J., &
Harrison, P. J. (2002). Asymmetry of the uncinate
fasciculus: A post-mortem study of normal subjects
and patients with schizophrenia. Cerebral Cortex,
11, 1218–1224.
Hoffman, P., Jefferies, E., & Lambon Ralph, M. UN. (2010).
Ventrolateral prefrontal cortex plays an executive regulation
role in comprehension of abstract words: Convergent
neuropsychological and rTMS evidence. Journal of
Neuroscience, 46, 15450–15456.
Hoffman, P., Pobric, G., & Lambon Ralph, M. UN. (2012). Brain
networks in semantic association tasks. Aphasiology,
26, 1119–1130.
Hornberger, M., Geng, J., & Hodges, J. R. (2011). Convergent
grey and white matter evidence of orbitofrontal cortex
changes related to disinhibition in behavioural variant
frontotemporal dementia. Brain, 134, 2502–2512.
Irish, M., Hodges, J. R., & Piguet, O. (2014). Right anterior
temporal lobe dysfunction underlies theory of mind
impairments in semantic dementia. Brain, 137, 1241–1253.
Kiehl, K. A., Smith, UN. M., Mendrek, A., Forster, B. B., Hare,
R. D., & Liddle, P. F. (2004). Temporal lobe abnormalities
in semantic processing by criminal psychopaths as revealed
by functional magnetic resonance imaging. Psychiatry
Research, 130, 297–312.
Kučera, H., & Francis, W. (1967). Computational analysis of
present day American English. Providence, RI: Brown
Stampa universitaria.
Lambon Ralph, M. UN. (2013). Neurocognitive insights on
conceptual knowledge and its breakdown. Philosophical
Transactions of the Royal Society of London, Series B,
Biological Sciences, 369, 20120392.
Lambon Ralph, M. A., McClelland, J. L., Patterson, K.,
Galton, C. J., & Hodges, J. R. (2001). No right to speak?
The relationship between object naming and semantic
impairment: Neuropsychological evidence and a
computational model. Journal of Cognitive Neuroscience,
13, 341–356.
358
Journal of Cognitive Neuroscience
Volume 28, Numero 3
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
C
7
N
6
_
9
UN
/
_
j
0
o
0
C
9
N
0
2
_
UN
P
_
D
0
0
B
9
sì
0
G
2
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
.
/
F
T
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
Lambon Ralph, M. A., Pobric, G., & Jefferies, E. (2009).
Conceptual knowledge is underpinned by the temporal
pole bilaterally: Convergent evidence from rTMS. Cerebral
Cortex, 19, 832–838.
Lambon Ralph, M. A., Sage, K., Jones, R. W., & Mayberry, E. J.
(2010). Coherent concepts are computed in the anterior
temporal lobes. Proceedings of the National Academy of
Scienze, U.S.A., 107, 2717–2722.
Liu, W., Mugnaio, B. L., Kramer, J. H., Rankin, K., Wyss-Coray, C.,
Gearhart, R., et al. (2004). Behavioral disorders in the
frontal and temporal variants of frontotemporal dementia.
Neurology, 62, 742–748.
Loftus, G. R., & Masson, M. E. J. (1994). Using confidence
intervals in within-subject designs. Psychological Bulletin &
Review, 1, 476–490.
Marinkovic, K., Dhond, R. P., Dale, UN. M., Glessner, M., Carr, V.,
& Halgren, E. (2003). Spatiotemporal dynamics of
modality-specific and supramodal word processing. Neuron,
38, 487–497.
Martin, UN. (2007). The representation of object concepts in
the brain. Annual Review of Psychology, 58, 25–45.
Mugnaio, B. L., Darby, A., Benson, D. F., Cummings, J. L., & Mugnaio,
M. H. (1997). Aggressive, socially disruptive and antisocial
behaviour associated with fronto-temporal dementia.
British Journal of Psychiatry, 170, 150–154.
Moll, J., Zahn, R., de Oliveira-Souza, R., Krueger, F., & Grafman,
J. (2005). The neural basis of human moral cognition.
Nature Reviews Neuroscience, 6, 799–809.
Moran, M. A., Mufson, E. J., & Mesulam, M. M. (1987). Neural
inputs into the temporopolar cortex of the rhesus monkey.
Journal of Comparative Neurology, 256, 88–103.
Neary, D., Snowden, J. S., Gustafson, L., Passant, U., Stuss, D.,
Black, S., et al. (1998). Frontotemporal lobar degeneration:
A consensus on clinical diagnostic criteria. Neurology,
51, 1546–1554.
Neubert, F. X., Mars, R. B., Buch, E. R., Olivier, E., & Rushworth,
M. F. (2010).Cortical and subcortical interactions during
action reprogramming and their related white matter
pathways. Proceedings of the National Academy of Sciences,
U.S.A., 107, 13240–13245.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsycholgia,
9, 97–113.
Parker, G. J., Luzzi, S., Alexander, D. C., Wheeler-Kingshott,
C. A., Ciccarelli, O., & Lambon Ralph, M. UN. (2005).
Lateralization of ventral and dorsal auditory-language
pathways in the human brain. Neuroimage,
24, 656–666.
Pascual, B., Masdeu, J. C., Hollenbeck, M., Makris, N., Insausti,
R., Ding, S. L., et al. (2014). Large-scale brain networks of
the human left temporal pole: A functional connectivity MRI
study. Cerebral Cortex, 34, 14864–14873.
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do
you know what you know? The representation of semantic
knowledge in the human brain. Nature Reviews Neuroscience,
8, 976–987.
Pobric, G., Jefferies, E., & Lambon Ralph, M. UN. (2007). Anterior
temporal lobes mediate semantic representation: Mimicking
semantic dementia by using rTMS in normal participants.
Proceedings of the National Academy of Sciences, U.S.A.,
104, 20137–20141.
Pobric, G., Jefferies, E., & Lambon Ralph, M. UN. (2010).
Category-specific vs. category-general semantic impairment
induced by transcranial magnetic stimulation. Current
Biology, 20, 964–968.
Quentin, R., Chanes, L., Migliaccio, R., Valabrègue, R., &
Valero-Cabré, UN. (2013). Fronto-tectal white matter
connectivity mediates facilitatory effects of non-invasive
neurostimulation on visual detection. Neuroimage,
82, 344–354.
Rankin, K. P., Gorno-Tempini, M. L., Allison, S. C., Stanley,
C. M., Glenn, S., Weiner, M. W., et al. (2006). Structural
anatomy of empathy in neurodegenerative disease. Brain,
129, 2945–2956.
Rice, G. E., Lambon Ralph, M. A., & Hoffman, P. (2015).
The roles of left versus right anterior temporal lobes in
conceptual knowledge: An ALE meta-analysis of 97 functional
neuroimaging studies. Cerebral Cortex, 25, 4374–4391.
Rogers, T. T., & McClelland, J. l. (2004). Semantic cognition: UN
parallel distributed processing approach. Cambridge, MA:
CON Premere.
Rosen, H. J., Allison, S. C., Schauer, G. F., Gorno-Tempini, M. L.,
Weiner, M. W., & Mugnaio, B. l. (2005). Neuroanatomical
correlates of behavioural disorders in dementia. Brain,
128, 2612–2625.
Ross, l. A., & Olson, IO. R. (2010). Social cognition and
anterior temporal poles. Neuroimage, 49, 3452–3462.
Sabsevitz, D. S., Medler, D. A., Seidenberg, M., & Binder, J. R.
(2005). Modulation of the semantic system by word
imageability. Neuroimage, 27, 188–200.
Schapiro, UN. C., McClelland, J. L., Welbourne, S. R., Rogers, T. T.,
& Lambon Ralph, M. UN. (2013). Why bilateral damage is worse
than unilateral damage to the brain. Journal of Cognitive
Neuroscience, 12, 107–123.
Sharp, D. J., Scott, S. K., & Wise, R. J. (2004). Retrieving
meaning after temporal lobe infarction: The role of the basal
language area. Annals of Neurology, 56, 836–846.
Simmons, W. K., Reddish, M., Bellgowan, P. S., & Martin,
UN. (2010). The selectivity and functional connectivity
of the anterior temporal lobes. Cerebral Cortex,
20, 813–825.
Skipper, l. M., Ross, l. A., & Olson, IO. R. (2011). The sensory
and categorical topography of the anterior temporal lobe.
Neuropsychologia, 49, 3419–3429.
Snowden, J. S., Goulding, P. J., & Neary, D. (1989). Semantic
dementia: A form of circumscribed cerebral atrophy.
Behavioural Neurology, 2, 167–182.
Snowden, J. S., Thompson, J. C., Stopford, C. L., Richardson,
UN. M., Gerhard, A., Neary, D., et al. (2011). The clinical
diagnosis of early-onset dementias: Diagnostic accuracy
and clinicopathological relationships. Brain,
134, 2478–2492.
Spitsyna, G., Warren, J. E., Scott, S. K., Turkheimer, F. E., &
Wise, R. J. S. (2006). Converging language streams in the
human temporal lobe. Journal of Neuroscience, 26,
7328–7336.
Straube, B., Verde, A., Jansen, A., Chatterjee, A., & Kircher, T.
(2010). Social cues, mentalizing and the neural processing of
speech accompanied by gestures. Neuropsychologia, 48,
382–393.
Vandenberghe, R., Price, C., Wise, R., Josephs, O., & Frackowiak,
R. S. J. (1996). Functional anatomy of a common semantic
system for words and pictures. Nature, 383, 254–256.
Visser, M., Embleton, K. V., Jefferies, E., Parker, G. J., &
Lambon Ralph, M. UN. (2010). The inferior, anterior
temporal lobes and semantic memory clarified: Novel
evidence from distortion-corrected fMRI. Neuropsychologia,
48, 1689–1696.
Visser, M., & Lambon Ralph, M. UN. (2011). Differential
contributions of bilateral ventral anterior temporal
lobe and left anterior superior temporal gyrus to
semantic processes. Journal of Cognitive Neuroscience,
23, 3121–3131.
Walsh, V., & Cowey, UN. (2000). Transcranial magnetic
stimulation and cognitive neuroscience. Nature Reviews
Neuroscience, 1, 73–79.
Pobric, Lambon Ralph, and Zahn
359
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
C
7
N
6
_
9
UN
/
_
j
0
o
0
C
9
N
0
2
_
UN
P
_
D
0
0
B
9
sì
0
G
2
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
F
.
/
T
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
Wise, R. J. (2003). Language systems in normal and aphasic
human subjects: Functional imaging studies and
inferences from animal studies. British Medical Bulletin,
65, 95–119.
Zahn, R., Moll, J., Iyengar, V., Huey, E. D., Tierney, M.,
Krueger, F., et al. (2009). Social conceptual impairments
in frontotemporal lobar degeneration with right anterior
temporal hypometabolism. Brain, 132, 604–616.
Zahn, R., Moll, J., Krueger, F., Huey, E. D., Garrido, G., &
Grafman, J. (2007). Social concepts are represented in the
superior anterior temporal cortex. Atti del
National Academy of Sciences, U.S.A., 104, 6430–6435.
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
3
2
8
3
/
5
3
1
/
1
3
9
5
5
1
0
/
6
1
3
7
0
8
o
4
C
7
N
6
_
9
UN
/
_
j
0
o
0
C
9
N
0
2
_
UN
P
_
D
0
0
B
9
sì
0
G
2
tu
.
e
P
S
T
D
o
F
N
B
0
sì
7
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
/
F
.
T
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
360
Journal of Cognitive Neuroscience
Volume 28, Numero 3