FOCUS FEATURE:
New Trends in Connectomics
Functional and structural connectome properties
in the 5XFAD transgenic mouse model of
Alzheimer’s disease
1
Shelli R. Kesler
, Paul Acton
2
, Vikram Rao
1
2
, and William J. Ray
1Department of Neuro-oncology, University of Texas MD Anderson Cancer Center, Houston, TX, USA
2Neurodegeneration Consortium, Institute for Applied Cancer Science, University of Texas MD Anderson Cancer Center,
Houston, TX, USA
a n o p e n a c c e s s
j o u r n a l
Keywords: Alzheimer’s disease, Connectome, Neuroimaging, fMRI, Diffusion tensor imaging
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
N
e
N
_
UN
_
0
0
0
4
8
P
D
/
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
ABSTRACT
Neurodegeneration in Alzheimer’s disease (AD) is associated with amyloid-beta peptide
accumulation into insoluble amyloid plaques. The five-familial AD (5XFAD) transgenic
mouse model exhibits accelerated amyloid-beta deposition, neuronal dysfunction, E
cognitive impairment. We aimed to determine whether connectome properties of these
mice parallel those observed in patients with AD. We obtained diffusion tensor imaging
and resting-state functional magnetic resonance imaging data for four transgenic and four
nontransgenic male mice. We constructed both structural and functional connectomes
and measured their topological properties by applying graph theoretical analysis. Noi
compared connectome properties between groups using both binarized and weighted
networks. Transgenic mice showed higher characteristic path length in weighted structural
connectomes and functional connectomes at minimum density. Normalized clustering
and modularity were lower in transgenic mice across the upper densities of the structural
connectome. Transgenic mice also showed lower small-worldness index in higher structural
connectome densities and in weighted structural networks. Hyper-correlation of structural
and functional connectivity was observed in transgenic mice compared with nontransgenic
controls. These preliminary findings suggest that 5XFAD mouse connectomes may provide
useful models for investigating the molecular mechanisms of AD pathogenesis and testing
the effectiveness of potential treatments.
AUTHOR SUMMARY
Many connectome properties have been shown to be preserved across species, providing
potentially novel insights regarding the mechanisms of various disease processes. In this
study, we measured functional and structural connectomes in a transgenic mouse model of
Alzheimer’s disease using resting-state functional MRI and diffusion tensor imaging. Noi
showed that connectome organization was significantly altered in transgenic mice compared
with nontransgenic controls in ways that parallel what has been observed in human patients.
These findings suggest that transgenic mouse connectomes may be useful for studying the
etiology and treatment of Alzheimer’s disease.
Citation: Kesler, S. R., Acton, P., Rao,
V., & Ray, W. J. (2018). Functional and
structural connectome properties in
the 5XFAD transgenic mouse model of
Alzheimer’s disease. Network
Neuroscience, 2(2), 241–258.
https://doi.org/10.1162/netn_a_00048
DOI:
https://doi.org/10.1162/netn_a_00048
Supporting Information:
https://doi.org/10.1162/netn_a_00048
Received: 23 Giugno 2017
Accepted: 14 Febbraio 2018
Competing Interests: The authors have
declared that no competing interests
exist.
Corresponding Author:
Shelli R. Kesler
skesler@mdanderson.org
Handling Editor:
Olaf Sporns
Copyright: © 2018
Istituto di Tecnologia del Massachussetts
Pubblicato sotto Creative Commons
Attribuzione 4.0 Internazionale
(CC BY 4.0) licenza
The MIT Press
5XFAD multimodal mouse connectome
Apolipoprotein E (APOE):
A gene that encodes a protein
important for the metabolism and
transport of fats.
Amyloid precursor protein (APP):
A protein believed to be involved in
neural development and
degeneration.
Presenilin 1 (PSEN1):
A gene that encodes a protein
involved in processing APP.
Presenilin 2 (PSEN2):
A gene that encodes another protein
involved in processing APP.
5XFAD transgenic mouse model:
A mouse whose DNA has been
altered to express five genes that
are known to be associated with
Alzheimers disease.
Connectome:
The brain network.
INTRODUCTION
Alzheimer’s disease (AD) is the most common form of age-related neurodegeneration and
dementia (Risacher & Saykin, 2013). AD pathology initiates many years before diagnosis and
develops slowly in some individuals and more rapidly in others. Patients with incipient AD
initially are cognitively normal, but inevitably progress to severe dementia and death. Over 46
million people have Alzheimer’s dementia globally, and the prevalence is expected to double
every 20 years (Prince et al., 2015). There currently are no effective treatments for reversing
AD. Risk factors include age, first-degree family history, and the apolipoprotein E (APOE) e4
genotype (Green et al., 2002; Hebert et al., 2010; Saunders et al., 1993; ten Kate et al., 2016;
Wolters et al., 2017). Tuttavia, the only causative factors identified to date are mutations in
amyloid precursor protein (APP), presenilin 1 (PSEN1), or presenilin 2 (PSEN2) genes. These
mutations are rare but tend to be associated with aggressive, early onset disease and therefore
have provided unique information regarding the pathophysiology of AD (Bateman et al., 2011).
AD is associated with significant amyloid-beta peptide accumulation, which is produced
from APP by PSEN1 and PSEN2, leading to the hypothesis that it is a primary mechanism
of neurodegeneration (Hardy & Selkoe, 2002; Lloret, Fuchsberger, Giraldo, & Vina, 2015).
Neuroimaging studies of patients with AD demonstrate significant abnormalities in brain struc-
ture and function. These abnormalities are abundant in frontal and temporal regions, including
the hippocampus and prefrontal cortex, but tend to reflect widespread disruption of large-scale,
distributed networks.
In vivo functional neuroimaging of transgenic mice may yield important insights regarding
the mechanisms of AD and provide preclinical models for testing the effectiveness of candidate
drugs on preventing or reversing AD-related neuropathology. Previous studies of APP/PS1 and
ArcBeta transgenic mice have demonstrated deficits in the functional connectivity of multiple
brain regions that is associated with amyloid deposition (Bero et al., 2012; Grandjean et al.,
2014; Shah et al., 2013). The five-familial AD (5XFAD) transgenic mouse model expresses
three APP and two PSEN1 mutations on a (C57BL/6 x SJL)F1 background. These mice demon-
strate accelerated amyloid deposition and have an early onset, aggressive disease presentation.
They are particularly useful for investigating the effects of amyloid-beta deposition on neuronal
loss (Eimer & Vassar, 2013).
Few if any studies have examined connectome organization in transgenic AD mice or any
disease group. Connectomics models brain networks as graphs with nodes (regions) and edges
(connections; Bassett & Bullmore, 2006). These mathematical models of brain networks pro-
vide measurements of information-processing efficiency and resilience to pathology, among
other topological properties that are highly relevant to AD (Contreras, Goni, Risacher, Sporns,
& Saykin, 2015; Dai & Lui, 2014; Tijms, Wink et al., 2013). Connectome properties have been
shown to be preserved across species and therefore provide a unique translational bridge be-
tween preclinical and clinical studies (Gorges et al., 2017; Oh et al., 2014; Stafford et al.,
2014; van den Heuvel, Bullmore, & Sporns, 2016).
Connectome studies of patients with AD have most consistently demonstrated alterations in
measures of network integration and efficiency (Daianu et al., 2013; Fischer, Wolf, Scheurich,
Fellgiebel, & Alzheimer’s Disease Neuroimaging Initiative, 2015; Kim et al., 2016; Lo et al.,
2010; Pereira et al., 2016; Reijmer et al., 2013; Stam, Jones, Nolte, Breakspear, & Scheltens,
2007; Wang et al., 2012; Zhao et al., 2012). Additionally, studies suggest that AD patho-
genesis targets high-traffic hub regions in the brain, spreading from epicenters to secondary
Network Neuroscience
242
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
T
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
N
e
N
_
UN
_
0
0
0
4
8
P
D
.
T
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
5XFAD multimodal mouse connectome
networks as the disease progresses (Buckner et al., 2009; Dai et al., 2015; Mallio et al., 2015;
Stam et al., 2009; Zhou, Gennatas, Kramer, Mugnaio, & Seeley, 2012; Zhou & Seeley, 2014).
We compared functional and structural connectomes of 5XFAD transgenic mice with those
of nontransgenic controls. The aim of this pilot study was to determine whether 5XFAD mice
show alterations in brain networks that parallel those observed in patients with AD, including
elevated characteristic path length, reduced network efficiency, and decreased hub presence.
METHODS
Subjects
◦
5XFAD mice were purchased from the Jackson Laboratory and maintained on the B6SJLF1/J
background. Mice were maintained on a 12-hour light/dark cycle at room temperature of
75
F with unrestricted access to food and water. In total, eight male mice, 23 weeks of age,
were used in this experiment. Four mice were 5XFAD transgenic and four were nontransgenic
littermate controls. 5XFAD mice at 23 weeks of age have been shown to have cognitive deficits
that are prior to significant neuronal or synaptic loss (Eimer & Vassar, 2013; Oakley et al.,
2006). Our protocols were approved by the University of Texas MD Anderson Institutional
Animal Care and Use Committee.
Neuroimaging
We obtained in vivo resting-state functional magnetic resonance imaging (rsfMRI) data from
mice using a 7 Tesla Bruker BioSpec (Bruker BioSpin, Billerica, MA) scanner while mice were
Isoflurane was administered at 1% (mixed with O2) to keep
anesthetized with isoflurane.
the respiration rate between 80 E 120 beats per minute (Stafford et al., 2014). Mice were
secured into the head coil with a bite bar and the head was taped down to minimize motion.
We first acquired a single-shot gradient, axial echo planar imaging (EPI) functional sequence
(slice thickness = 0.5 mm, gap = 0.0 mm, repetition time [TR] = 2,000 ms, echo time [TE] =
12 ms, matrix = 80 × 64 × 32, field of view [FOV] = 20 × 16 mm, flip angle = 75
, number of
volumes = 450, averages = 1, scan time = 15 min) followed by a T2-weighted, turbo spin echo,
rapid acquisition with refocused echoes (Turbo RARE) sequence (slice thickness: 0.5 mm,
gap = 0.0 mm, TR = 4,000 ms, TE = 40.00 ms, matrix = 256 × 180, FOV = 26.600 ×
18.000 mm, flip angle = 90
, number of images = 32, scan time = 4 min and 24 S).
◦
◦
◦
Six days following rsfMRI mice were euthanized using carbon dioxide and transcardially
perfused with 20 ml of 10 U/ml heparin (Sagent Pharmaceuticals, Schaumburg, IL) in PBS pH
7.4 (Invitrogen, Carlsbad, CA) at room temperature followed by 20 ml 4% paraformaldehyde in
PBS pH 7.4 (Electron Microcopy Sciences, Hatfield, PAPÀ) at room temperature. Following per-
fusion, the heads were removed and the skin, muscle, occhi, ears, nose tip, and lower jaw were
removed to expose the skull. The skulls were then immersed in 20 ml of 4% paraformaldehyde
in PBS pH 7.4 overnight at 4
C with continuous mixing. The skulls were then transferred to
50 ml of 0.01% sodium azide (Teknova, Hollister, CA) in PBS pH 7.4 at 4
C for seven days with
continuous mixing. At the end of the seven days the skulls were transferred to 50 ml of 5 mM
Magnevist (Gadopentetate Dimeglumine; Bayer Healthcare Pharmaceuticals, Indianola, PAPÀ),
0.01% sodium azide in PBS pH 7.4 at 4
C for 24 days with continuous mixing. Following the
Magnevist treatment the skulls were transferred to 50 ml of 0.01% sodium azide in PBS pH 7.4
at 4
C and maintained in this solution with continuous mixing until the day of imaging, Quando
the skulls were transferred to Fomblin Y (Sigma-Aldrich, Saint Louis, MO). We then acquired
ex vivo diffusion tensor imaging (DTI) data using a 9.4 Tesla Bruker Avance BioSpec scanner
(fMRI was not available on this scanner at the time of this study). The following parameters
◦
◦
◦
Network Neuroscience
243
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
T
/
/
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
N
e
N
_
UN
_
0
0
0
4
8
P
D
/
.
T
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
5XFAD multimodal mouse connectome
were used: spin echo, b-value = 0 and 1,000s/mm
, 20 diffusion directions with one non–
diffusion weighted image, TR = 500 ms, TE = 14.8 ms, FOV = 17 × 12.5 × 15 mm, matrix =
180 × 133 × 160, NEX = 1, δ = 3 ms, Δ = 7 ms, scan time = ~35 hr.
2
A brain mask was manually delineated in 3D for the T2 and rsfMRI volumes in FMRIB
Software Library (FSL) View v3.2.0 to remove the skull. RsfMRI data were preprocessed in
Statistical Parametric Mapping v8 including realignment and warping of the EPI volume via
the co-registered T2-weighted volume to a male C57BL/6 mouse brain template (Ma et al.,
2005). CONN Toolbox v13 software was then used to filter data to the <0.1 Hz range of
spontaneous activity (Raichle, 2011; Whitfield-Gabrieli & Ford, 2012). CONN implements
the CompCor method to remove motion and physiologic/nonneuronal artifacts. This method
involves extracting signal from white matter and cerebrospinal fluid regions using principal
component analysis and then regressing these signals out of the total fMRI signal (Behzadi,
Restom, Liau, & Liu, 2007). Functional time series were extracted from each of 32 bilateral
cortical and subcortical gray matter regions of interest to cover the entire brain (Supplementary
Figure 1; Kesler, Acton, Rao, & Ray, 2018), cross-correlated and normalized using Fisher r-to-z
transformation.
◦
DTI preprocessing was performed in FSL v5.0 (Smith et al., 2004) including eddy current
correction and tensor reconstruction. Deterministic tractography was performed in TrackVis
v0.6.1 (Wang, Benner, Sorensen, & Wedden, 2007) using an FA threshold of 0.1 and a
curvature threshold of 40
, based on the study by Chen et al. (2015). The 32 regions of inter-
est described above were warped into DTI native space via inverse transformation of the b0
volume to the mouse brain template. We determined the number of DTI streamlines connect-
ing each pair of regions, and regions were considered connected if one streamline endpoint
terminated within one region and the other endpoint terminated within the other region. A
threshold of three streamlines was applied to minimize false-positive streamlines, and each
valid edge was weighted by the average streamline fractional anisotropy (Kesler, Watson, &
Blayney, 2015).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
.
t
/
Functional and structural connectomes were constructed for each participant with N =
32 nodes, network degree of E = number of edges, and a network density of D = E/[(N ×
(N − 1))/2 ] representing the fraction of present connections to all possible connections.
Negative functional edges were zeroed given evidence that properties of negative correlation
networks are different than those of positive correlation networks (Hosseini & Kesler, 2013;
Schwarz & McGonigle, 2011). Structural connectomes were scaled to the range of 0 to 1
(Wang, Ghumare, Vandenberghe, & Dupont, 2017).
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Statistical Analysis
Connectome properties were calculated using graph theoretical analysis. Specifically, we mea-
sured characteristic path length and global/local efficiencies to test our hypothesis that these
properties would be altered in transgenic mice consistent with studies of patients with AD.
We additionally measured normalized clustering coefficient, small-worldness index, and mod-
ularity as these have also been reported in human studies of AD (Dai & He, 2014; Tijms,
Wink et al., 2013). Connectome properties were defined as previously described (Bassett &
Bullmore, 2006; Rubinov & Sporns, 2010; Sporns & Betzel, 2016). Briefly, characteristic path
length is the average shortest path length between all pairs of nodes normalized by the char-
acteristic path length of random networks. Normalized clustering coefficient is the propor-
tion of actual connections to possible connections between a node’s neighbors normalized
244
Global efficiency:
A measure of network efficiency of
information exchange based on path
lengths between regions.
Small-world:
A network organization or topology
associated with high local
connectivity and economical
large-range connectivity; that is, a
balance between segregation and
integration.
Modularity:
A measure of how well a network
can be decomposed into
nonoverlapping subnetworks or
modules.
Network Neuroscience
5XFAD multimodal mouse connectome
by the clustering coefficient of random networks. Small-worldness index is defined as nor-
malized clustering coefficient/characteristic path length. Path length and clustering coefficient
were normalized using 20 benchmark random networks (Zalesky, Fornito, & Bullmore, 2012).
Global efficiency is the inverse average shortest path length of the network, while local effi-
ciency is the inverse of the average shortest path connecting all neighbors of a node, or in
other words, the average efficiency of the local subgraphs. Modularity analysis involves de-
composing the network into nonoverlapping groups of regions (modules) that have maximal
within-group connections and minimal between-group connections. Connectome measure-
ment was conducted using Brain Connectivity Toolbox (Rubinov & Sporns, 2010).
Thresholding connectomes is necessary for removing false-positive edges and facilitating
between-group comparisons but can remove potentially valid information regarding
differences in network topology (Fornito, Zalesky, & Breakspear, 2013; van Wijk, Stam, &
Daffertshofer, 2010). Further, there tends to be a large difference in network densities between
rsfMRI- and DTI-derived connectomes. Therefore, we compared connectome properties across
multiple densities using the area under the curve (AUC; Bassett, Meyer-Lindenberg, Achard,
Duke, & Bullmore, 2006; Bassett, Nelson, Mueller, Camchong, & Lim, 2012). Specifically,
we measured connectome properties at each density from minimum connection density to
the last density associated with a small-world organization (Basset et al., 2008; Humphries &
Gurney, 2008) up to a maximum density of 0.5 (Kaiser & Hilgetag, 2006). We then measured
the AUC across this entire range as well as in a windowed manner where target windows were
determined from visual inspection of the small-worldness index curves. This approach was
based loosely on the clustering method introduced by Drakesmith et al. (2015). AUCs were
compared between groups using nonparametric permutation testing (Basset et al., 2008) using
2,000 iterations and two-tailed p values.
We also evaluated weighted networks without any thresholding. Connectome properties
from weighted networks were compared between groups using the general linear model with
network density as a covariate (Brown et al., 2011). The weighted network data are provided
in the Supplementary Information (Kesler et al., 2018).
To examine hub profiles, we determined whether the cumulative degree distribution of the
networks followed an exponentially truncated power-law indicating the presence of hub re-
gions (Achard, Salvador, Whitcher, Suckling, & Bullmore, 2006). This analysis was performed
with weighted networks and at minimum density. Power-law fitting and comparison was con-
ducted in the R statistical package v3.3.2 (R Foundation) using the “poweRlaw” library.
We supplemented hypothesis testing with exploratory analysis of regional effects using
the Network-Based Statistic Toolbox v1.2 (Zalesky, Fornito, & Bullmore, 2010). This method
identifies connected substructures, or components, within the larger network, similar to the
cluster-based thresholding approach used in traditional voxel-wise neuroimaging analyses
(Zalesky et al., 2010). Permutation testing with 2,000 permutations was then used to deter-
mine group differences in components controlling for multiple comparisons using family-wise
error (FWE). Because the network-based statistic (NBS) can be less sensitive to focal effects,
we also examined regional effects using false discovery rate (FDR; Zalesky et al., 2010). We
examined NBS using both extent and intensity; the latter improves the sensitivity of NBS to
focal effects (Zalesky et al., 2010).
We also explored the relationship between structure and function. First, network commu-
nication measures (e.g., search information of shortest paths, path transitivity) were computed
for each pair of nodes in the structural connectivity matrix for each subject. The structural
Network Neuroscience
245
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
5XFAD multimodal mouse connectome
communication measures were then entered into a multiple linear regression model to gen-
erate a predicted functional connectivity matrix for each subject (Goñi et al., 2014). In other
words, the communication measures for each structural node pair were used as the predic-
tors, and the functional connectivity between that same node pair was used the response.
The fitted responses from the linear regression were used to construct the predicted functional
connectivity matrix. Finally, a Pearson correlation was computed between the predicted and
observed functional matrices for each participant (Goñi et al., 2014). It was unknown how data
collected from two different field strengths and/or rodent neurobiology would affect structure-
function relationships, so we tested the default communication measures (shortest path length
and search information of shortest paths; Goñi et al., 2014) as well as all available measures
in the Brain Connectivity Toolbox. These included the default measures plus path transitivity,
column-wise z-scored mean first passage time, neighborhood overlap, and matching index
(Goñi et al., 2014). These are measures of information flow and community structure that
do not require global knowledge of the network’s topology (Goñi et al., 2014; Meghanathan,
2016). Between-group difference in these correlations was measured using two-tailed t test.
RESULTS
Small-World Organization
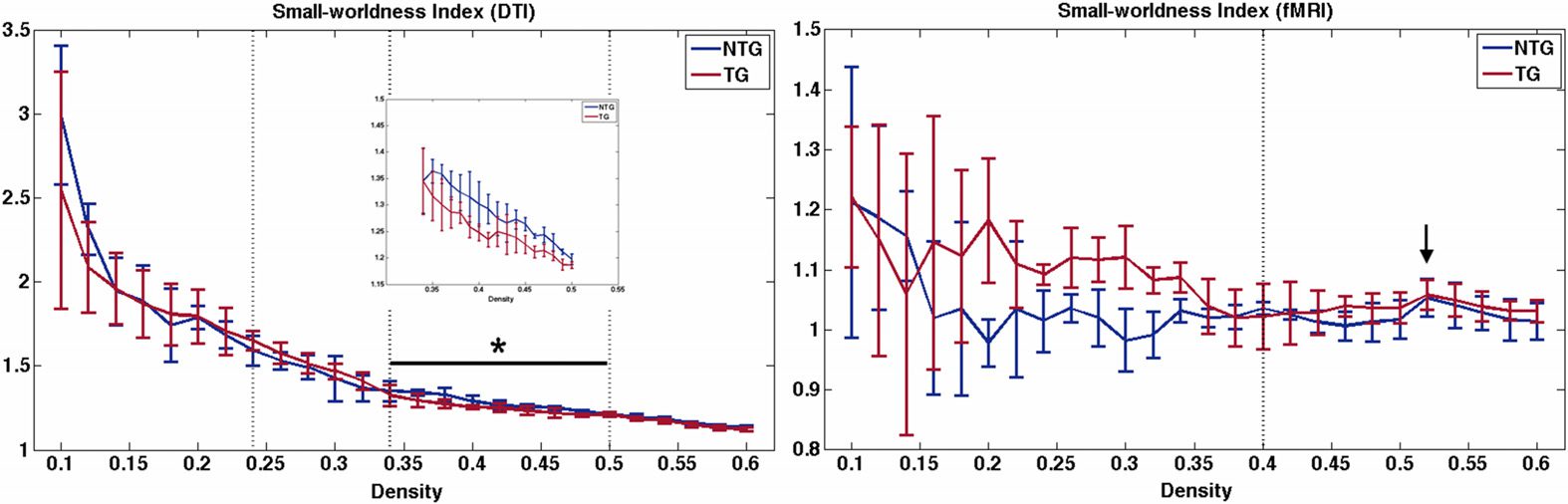
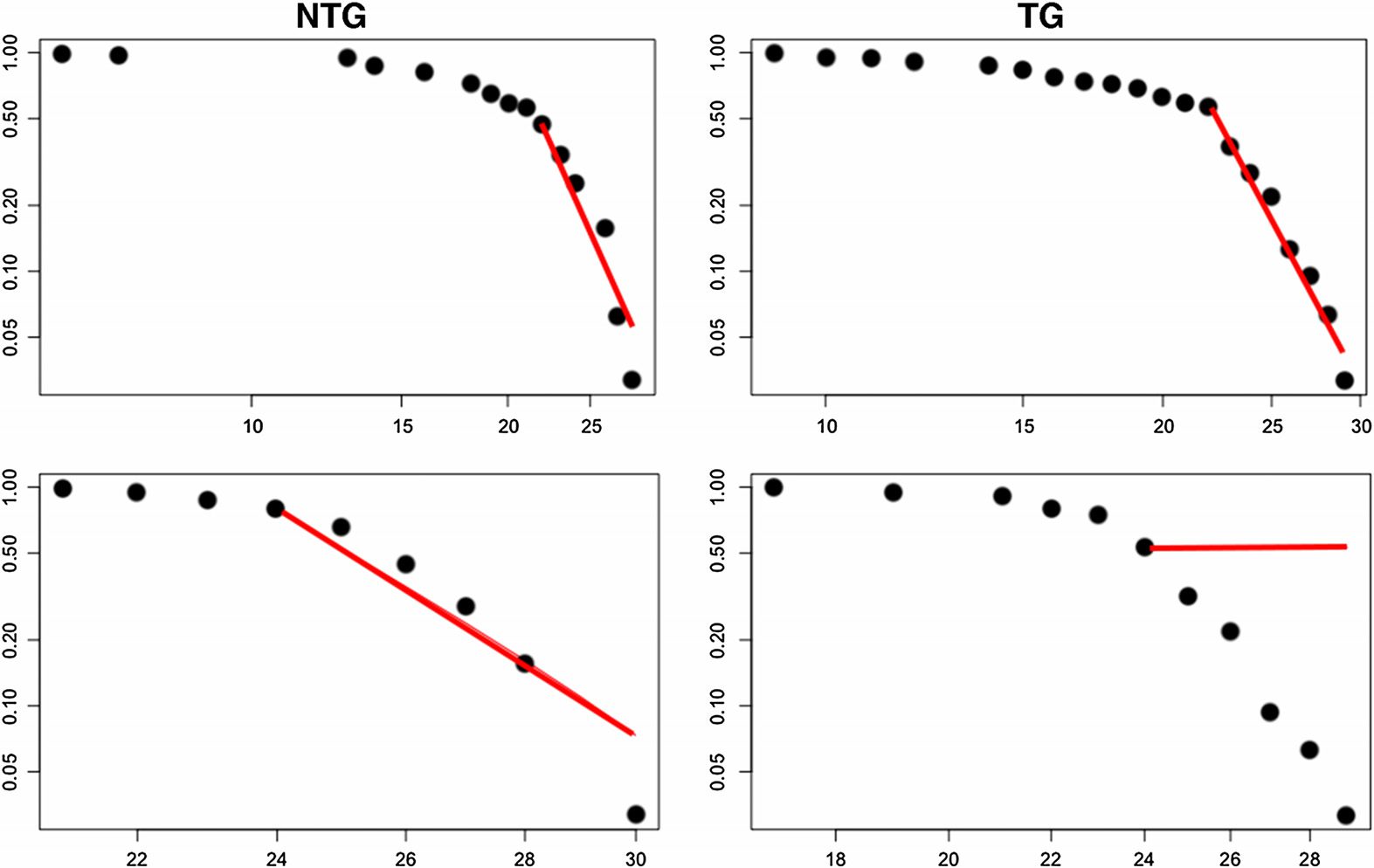
As shown in Figure 1, structural networks demonstrated expected small-world organization
defined as a small-worldness index greater than 1 (Humphries & Gurney, 2008) across multiple
densities. However, functional networks were small-world for all subjects at only one density
(0.52), which was one step above our upper density boundary. Minimum connection density
for structural networks occurred at 0.24 and at 0.4 for functional networks.
AUC Across Densities
For structural connectomes, permutation testing indicated no significant differences between
groups (p > 0.19, Figura 2) across the entire range of densities measured (0.24 A 0.5) or across
the first density window (p > 0.17, Figura 2), which was defined from minimum density to 0.34
Figura 1. Small-worldness index across network densities. For structural connectomes (DTI), ver-
tical lines indicate area under the curve (AUC) windows. Minimum connection density was 0.24
(first dotted vertical line). At a density of 0.34 (second dotted vertical line), the group curves ap-
pear to cross over, and therefore this is where we defined the first AUC window. Maximum density
was set at 0.5 (third dotted vertical line) for both modalities based on previous research. For func-
tional connectomes (fMRI), the dotted vertical line indicates minimum connection density (0.4). IL
black bar and asterisk indicate the significant AUC window, and the inset figure shows the curve
on a smaller scale for easier viewing. The arrow indicates the only density where all mice showed
small-worldness greater than 1. NTG = nontransgenic; TG = transgenic.
Network Neuroscience
246
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
N
e
N
_
UN
_
0
0
0
4
8
P
D
/
T
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
5XFAD multimodal mouse connectome
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
N
e
N
_
UN
_
0
0
0
4
8
P
D
T
.
/
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
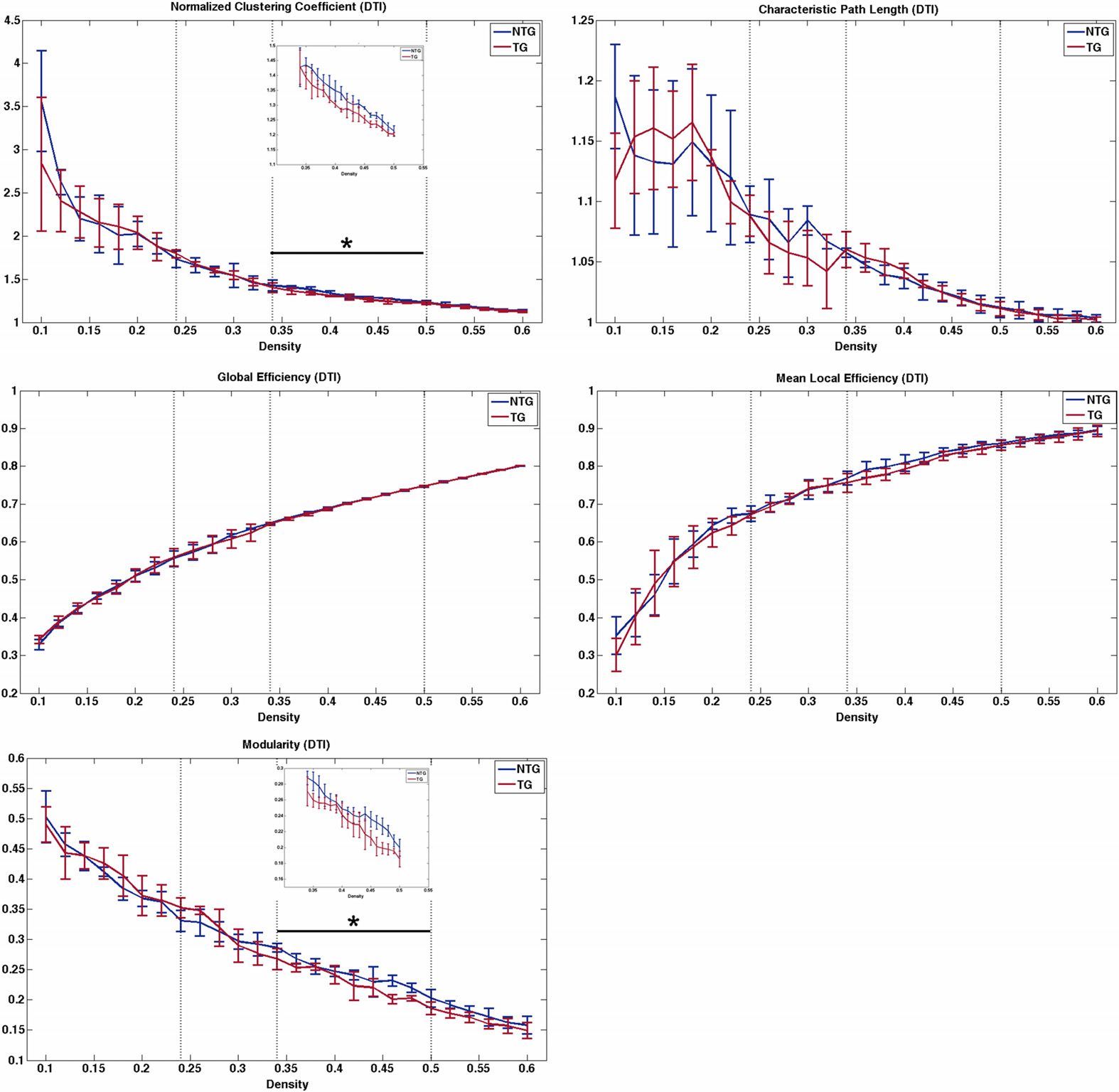
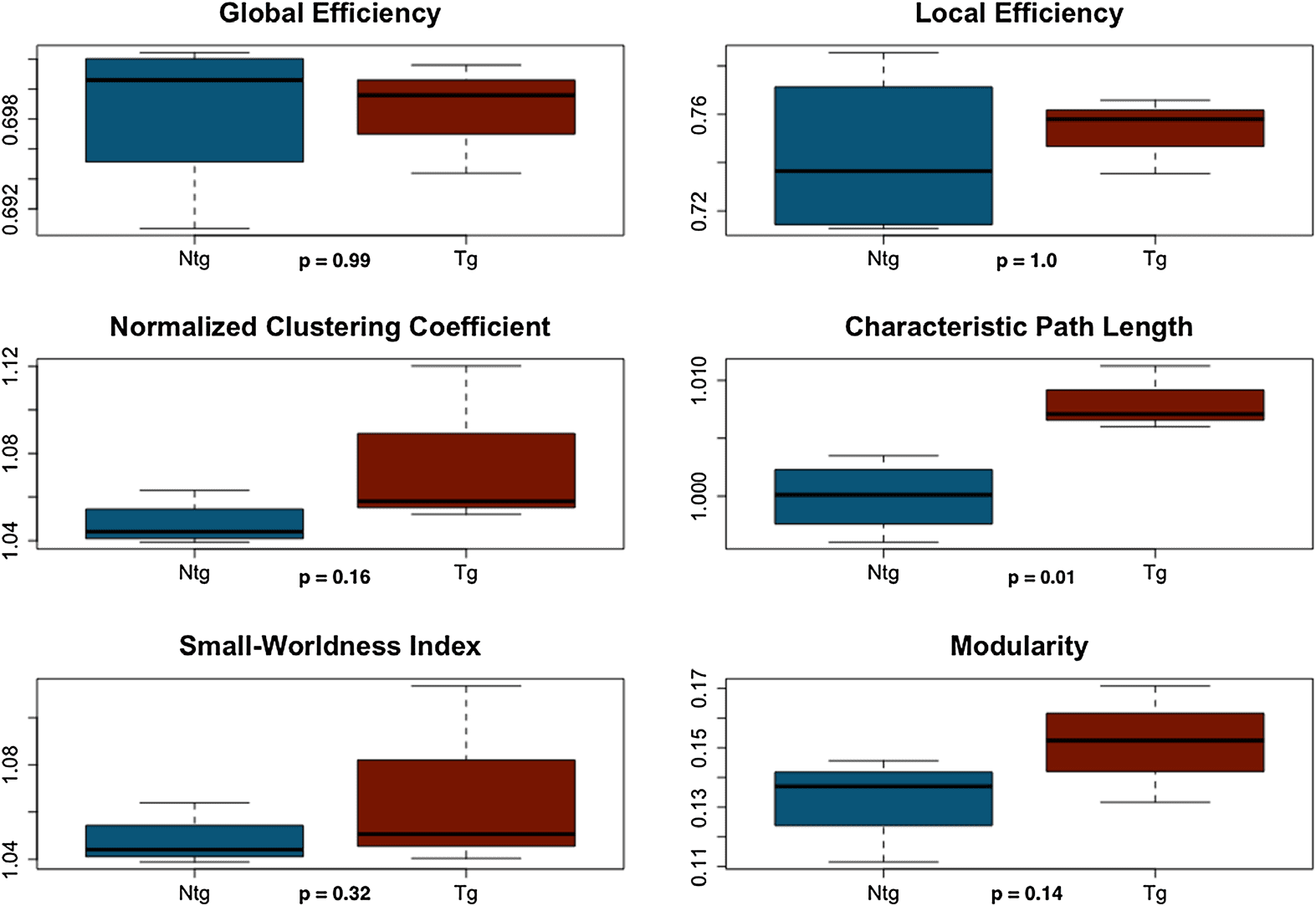
Figura 2. Structural connectome properties. Dotted vertical lines indicate area under the curve
(AUC) windows. The black bar and asterisk indicate the significant AUC window, and the inset figure
shows the curve on a smaller scale for easier viewing. NTG = nontransgenic; TG = transgenic.
where the group curves crossed over. Tuttavia, transgenic mice demonstrated significantly
lower normalized clustering coefficient (p = 0.01), small-worldness index (p = 0.02), E
modularity (p = 0.03) compared with nontransgenic mice across the second density window
from 0.34 to maximum density (Figures 1 E 2). Module regions are presented in Table 1.
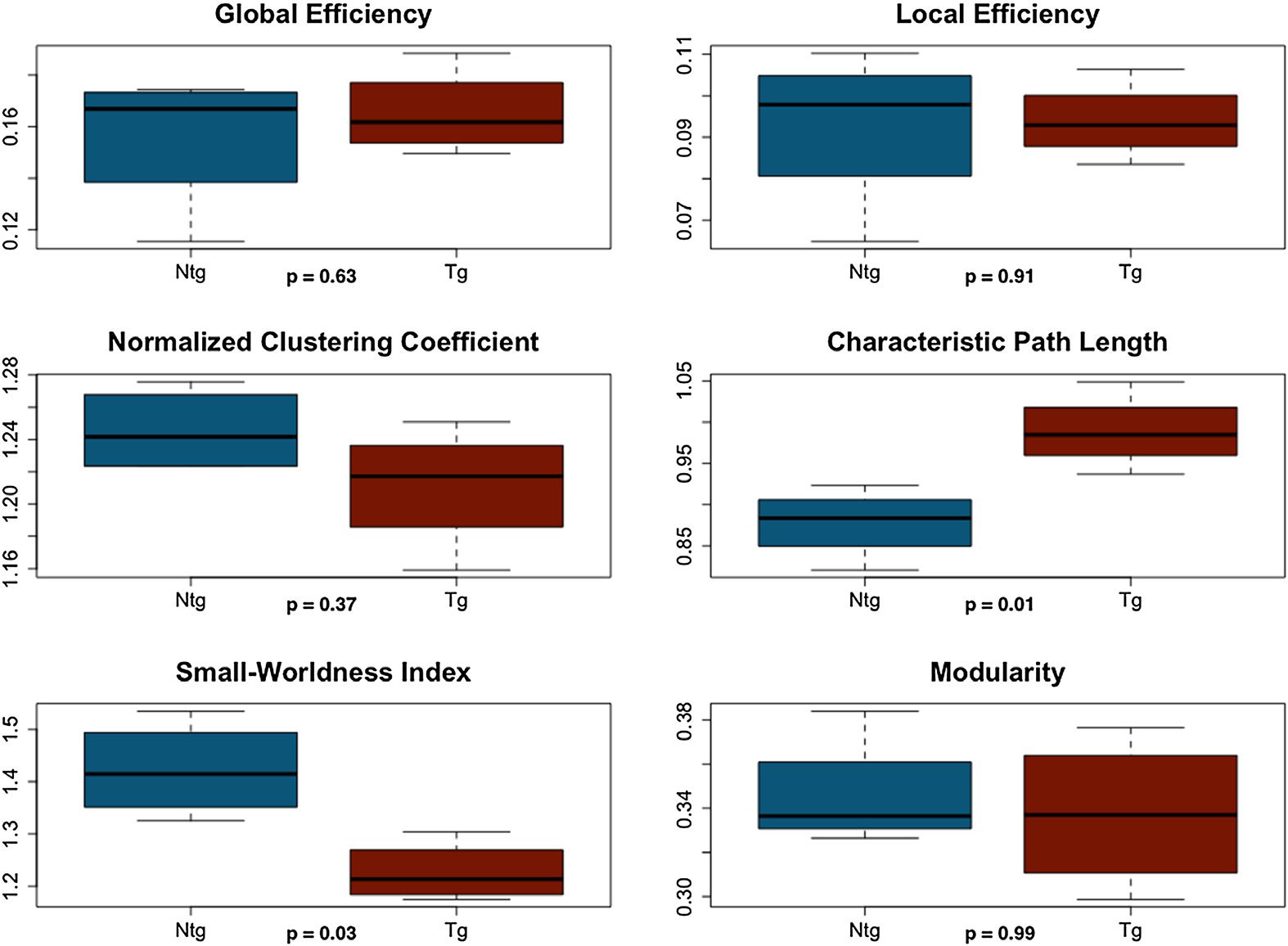
Given the above small-worldness results, we did not compare the AUCs of functional con-
nectomes between groups. It was not possible to simply exclude data since the lack of small-
worldness affected different mice at different densities. Tuttavia, at minimum connection
density, connectomes of all subjects but one in the transgenic group showed small-world or-
ganization, and therefore we compared connectome metrics at this specific density after ex-
cluding the transgenic subject. T test indicated significantly higher characteristic path length
in transgenic mice (t = 3.64 p = 0.01, Figura 3).
Weighted Networks
All weighted structural networks demonstrated small-world organization. As shown in Fig-
ure 4, general linear models covaried for density indicated that structural connectomes of
Network Neuroscience
247
5XFAD multimodal mouse connectome
Tavolo 1. Module regions
Nontransgenic
Module 1
Right amygdala
Right striatum
Right cerebellum
Right pallidum
Right hippocampus
Right inferior colliculi
Right neocortex
Left olfactory
Right olfactory
Right basal forebrain/septum
Right thalamus
Transgenic
Module 1
Left external capsule
Right external capsule
Left hypothalamus
Right hypothalamus
Left superior colliculi
Right superior colliculi
Module 2
Left external capsule
Right external capsule
Left hypothalamus
Left superior colliculi
Right superior colliculi
Module 2
Right amygdala
Right brainstem
Right striatum
Right central gray
Right cerebellum
Right pallidum
Right hippocampus
Right inferior colliculi
Right neocortex
Right olfactory
Module 3
Left amygdala
Left brainstem
Left striatum
Left central gray
Left cerebellum
Left pallidum
Left hippocampus
Left inferior colliculi
Left neocortex
Left midbrain
Left basal forebrain/septum
Left thalamus
Module 3
Left amygdala
Left brainstem
Left striatum
Left central gray
Left cerebellum
Left pallidum
Left hippocampus
Left inferior colliculi
Left neocortex
Left olfactory
Module 4
Right brainstem
Right central gray
Right hypothalamus
Right midbrain
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
/
/
T
e
D
tu
N
e
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
N
e
N
_
UN
_
0
0
0
4
8
P
D
T
/
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Right midbrain
Right basal forebrain/septum
Left midbrain
Left basal forebrain/septum
Right thalamus
Left thalamus
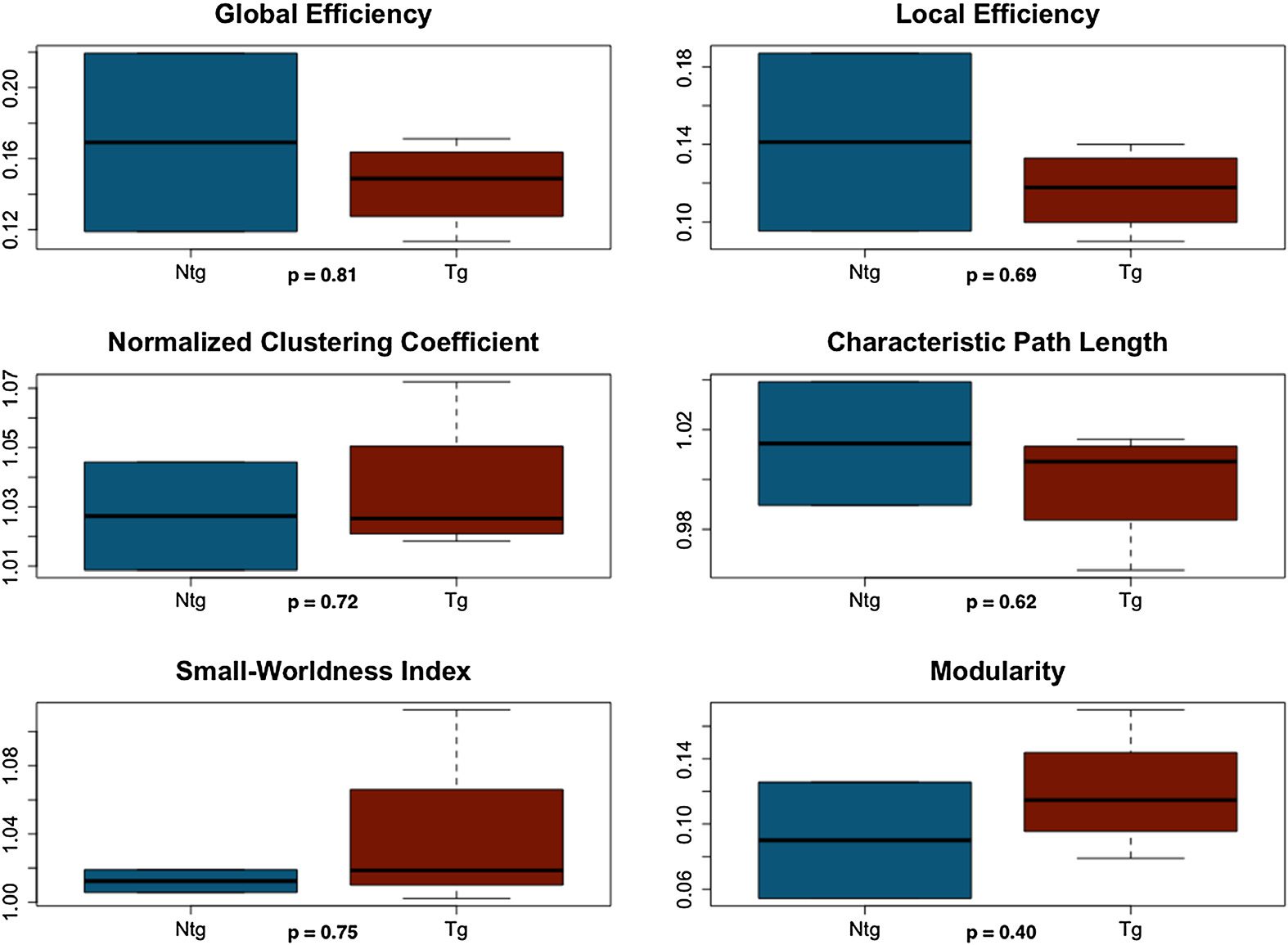
transgenic mice showed significantly higher characteristic path length (F = 15.2, p = 0.01)
and lower small-worldness index (F = 9.73, p = 0.03) compared with controls.
Weighted functional connectomes for two nontransgenic mice did not demonstrate small-
worldness, so these were excluded. There were no significant group differences (Figura 5).
Hubs: Degree Distribution
Both groups showed goodness of fit with the power-law with no between-group difference
(p = 0.68). There was no significant group difference in power-law fit for either modality at
minimum density (p > 0.343). For weighted functional networks, the transgenic group showed
poor fit with the power-law (Figura 6), and this was significantly lower than that of the control
group (P < 0.001).
Network Neuroscience
248
5XFAD multimodal mouse connectome
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Figure 3. Functional connectome properties at minimum connection density. NTG = nontrans-
genic; TG = transgenic.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Weighted structural connectome properties. NTG = nontransgenic; TG = transgenic.
Network Neuroscience
249
5XFAD multimodal mouse connectome
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
Figure 5. Weighted functional connectome properties. NTG = nontransgenic; TG = transgenic.
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Power-law fit of cumulative degree distributions. Power-law fit is shown as a plot of log
degree (x-axis) by log cumulative degree distribution (y-axis). Left column = nontransgenic (NTG),
right column = transgenic (TG), top row = weighted structural connectomes, bottom row = weighted
functional connectomes.
Network Neuroscience
250
5XFAD multimodal mouse connectome
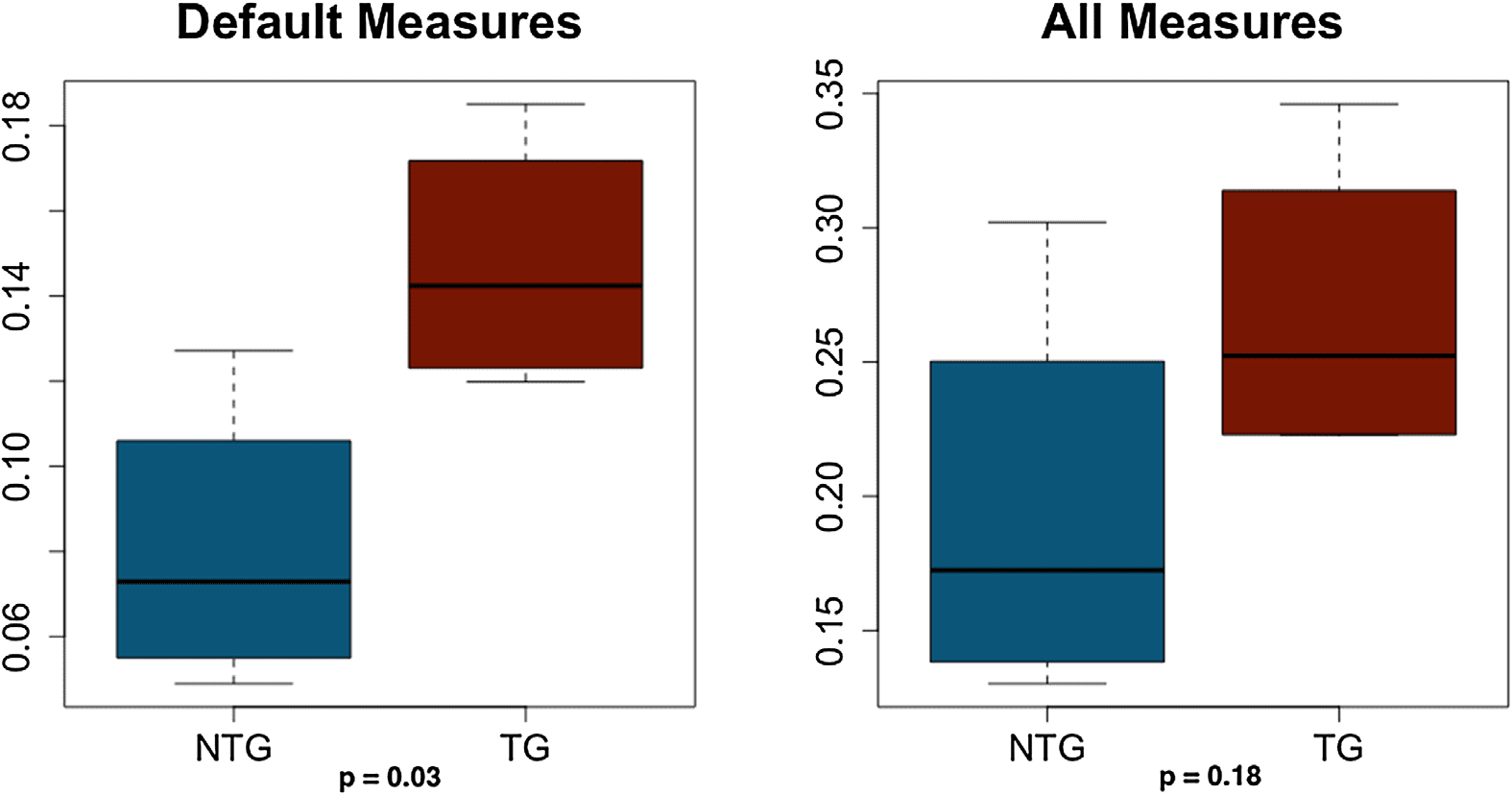
Figure 7. Correlation coefficients for structural and functional connectivity. Default measures =
shortest path length, search information of shortest path length; all measures = default measures
plus path transitivity, column-wise z-scored mean first passage time, neighborhood overlap, and
matching index. NTG = nontransgenic; TG = transgenic.
Regional Connectivity
There were no significant regional effects for structural or functional connectomes weighted
or at minimum density via NBS or FDR comparison.
Relationship Between Structure and Function
This analysis was conducted only on weighted networks without excluding any subjects. Us-
ing default communication measures resulted in significant correlations between structure and
function for five out of eight subjects (p < 0.008). Using all communication measures resulted
in significant correlations for all subjects (p < 0.004). T tests indicated that correlation coeffi-
cients based on the default model were significantly higher in the transgenic group (t = 2.92,
p = 0.03), but there was no difference in the coefficients from the all-measures model (t = 1.53,
p = 0.18, Figure 7).
DISCUSSION
To our knowledge, this is the first study to evaluate connectome organization in an AD mouse
model. Few if any studies have compared connectomes in rodent models of disease groups.
Using in vivo resting-state fMRI and ex vivo DTI, we constructed and measured functional
and structural connectomes for 5XFAD transgenic mice. This mouse model is characterized
by aggressive amyloid pathology. We evaluated connectome properties in these mice across
multiple network densities (i.e., thresholds) and also for weighted, unthresholded networks
and compared them with the connectomes of nontransgenic control mice.
Weighted DTI-based structural networks demonstrated significantly higher path length and
lower small-worldness index in transgenic mice after controlling for network density. Weighted
functional networks also demonstrated higher characteristic path length at minimum connec-
tion density. Clinical studies of patients with AD have demonstrated higher characteristic
path lengths of DTI- or fMRI-based connectomes (Lo et al., 2010; Wang et al., 2012;
Network Neuroscience
251
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
5XFAD multimodal mouse connectome
Zhao et al., 2012). Increased characteristic path length has also been observed in patients with
AD using gray matter structural connectomes (Kim et al., 2016; Pereira et al., 2016) and elec-
troencephalography (EEG)-based networks (Stam et al., 2007). In a small-world network, clus-
tering coefficient is greater than that of random networks, while the path length is comparable
to that of random networks (Bassett & Bullmore, 2006; Humphries & Gurney, 2008; Watts
& Strogatz, 1998). Characteristic path length is defined as the average shortest path length
between all pairs of network nodes divided by the mean path length of benchmark random
networks (Bassett & Bullmore, 2006; Humphries & Gurney, 2008; Watts & Strogatz, 1998;
Zalesky et al., 2012). Therefore, higher characteristic path length suggests disconnection within
the network such that longer, less efficient routes of information exchange must be taken. It
also suggests that the network is less random in terms of path length.
We also observed lower normalized clustering coefficient, small-worldness index, and
modularity, indicating that the connectomes of transgenic mice were more similar to random
networks in these properties. Lower clustering coefficient suggests lower network segrega-
tion or specialization, while lower small-worldness index reflects the overall similarity to ran-
dom networks in that segregation and integration (i.e., path length) are imbalanced (Bassett
& Bullmore, 2006; Watts & Strogatz, 1998). Previous DTI and fMRI connectome studies of
patients with AD have also demonstrated lower normalized clustering, small-worldness, and
modularity (Brier et al., 2014; Sun et al., 2014; Supekar, Menon, Rubin, Musen, & Greicius,
2008).
However, some studies have observed conflicting results, including lower characteristic
path length in DTI (Daianu et al., 2013), gray matter (Tijms, Moller et al., 2013), fMRI (Sanz-
Arigita et al., 2010), EEG (de Haan et al., 2009), and magneto-encephalogram (MEG; Stam
et al., 2009) connectomes in patients with AD. Inconsistencies in connectome findings are
a well-known issue in the literature (for reviews, see Dai & He, 2014; Tijms, Wink et al.,
2013) and often reflect differences in methodology such as imaging modality and/or choice
of thresholding method. This was part of our rationale for a multimodal study including dif-
ferent thresholding methods. Our most consistent findings included higher characteristic path
length in transgenic mice, which was noted in weighted, unthresholded DTI connectomes,
and fMRI connectomes thresholded to minimum connection density. We also observed lower
small-worldness index in transgenic mice in DTI connectomes thresholded to higher densities
and in weighted, unthresholded DTI graphs. Small-worldness index is the ratio of clustering
coefficient to path length (Bassett & Bullmore, 2006; Watts & Strogatz, 1998) and therefore,
lower values can reflect lower normalized clustering and/or higher characteristic path length.
Clinical studies have also noted decreased network efficiency in patients with AD (Daianu
et al., 2013; Fischer et al., 2015; Lo et al., 2010; Reijmer et al., 2013), which we did not ob-
serve. Efficiency and path length are related measures (Achard & Bullmore, 2007). Since this
was a preliminary study, we may have lacked power to detect differences in efficiencies. The
5XFAD mouse model used in this study is associated with accelerated amyloid-beta pathology.
Both amyloid-beta and tau are believed to synergistically drive neurodegenerative processes
involved in AD (Bloom, 2014; Lloret et al., 2015; Stancu, Vasconcelos, Terwel, & Dewachter,
It is possible that greater tau burden
2014). 5XFAD do not have significant tau pathology.
and/or some other AD-related neuropathology is more associated with impairments in con-
nectome efficiency. We evaluated the 5XFAD mice at an age prior to significant neuronal and
synaptic loss, which may have preserved network efficiency.
Network Neuroscience
252
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
5XFAD multimodal mouse connectome
Lower cerebrospinal fluid (CSF) levels of amyloid-beta have been associated with higher
path length and lower clustering of the gray matter structural connectome in human partic-
ipants; low levels of amyloid-beta in the CSF indicate higher amyloid plaque burden in the
brain (Tijms et al., 2015). Gray matter and DTI-based connectome properties show moderate
convergence (Gong, He, Chen, & Evans, 2012). Accordingly, our findings provide indirect
support for amyloid-beta effects on connectome properties of the 5XFAD transgenic mouse
connectome. We did not have molecular assays available for analysis in this study. Future AD
transgenic mouse connectome studies could provide unique insights regarding the molecular
mechanisms underlying impairments in various connectome properties associated with AD.
For example, Golgi staining and longitudinal fluorescent microscopy could be used to exam-
ine neuronal morphology and survival rates in impaired connectome regions. Evaluating the
role of mitochondrial dysfunction (Lin & Beal, 2006) via Seahorse flux technology (Brand &
Nicholls, 2011) is another potential application.
Previous research has demonstrated lower connectivity among regions involved in specific
brain networks of patients with AD, including the default mode network (Bai et al., 2011;
Damoiseaux, Prater, Miller, & Greicius, 2012; Lehmann et al., 2013; Simic, Babic, Borovecki,
& Hof, 2014). Relevant subnetworks have been shown to be present in mice both structurally
and functionally (Liska, Galbusera, Schwarz, & Gozzi, 2015; Stafford et al., 2014), but we
did not find any significant regional connectome differences between transgenic mice and
controls. This may again reflect limited statistical power and/or may indicate diffuse regional
effects. Modularity was lower at higher densities in structural connectomes of transgenic mice,
suggesting fewer dissociable networks. Transgenic mice appeared to lack separation between
certain sensorimotor/homeostatic regions and other networks.
Both functional and structural connectome topologies showed the expected goodness of fit
with a power-law distribution. The power-law fit is believed to reflect the brain network’s hub
organization wherein the majority of information processing is handled by a small number
of core regions (Achard et al., 2006). This is consistent with other studies showing presence
of hubs in the mouse brain (Liska et al., 2015; Rubinov, Ypma, Watson, & Bullmore, 2015).
There was no difference in power-law fit between the groups for structural connectomes, but
transgenic mice showed a significantly poorer power-law fit in weighted functional connec-
tomes. Previous studies have suggested that AD pathogenesis may selectively target certain
hub regions (Dai et al., 2015; Stam et al., 2009; Xie & He, 2011; Yao et al., 2010; Zhou et al.,
2012).
Despite problematic functional connectomes that did not show adequate small-world char-
acteristics, functional connectivity was predicted from structural connectivity. Transgenic mice
tended to show hyper-correlation of structural and functional networks compared with
controls. Such hyper-correlation has been noted in patients with neurologic disorders (Kesler
et al., 2017; Rudie et al., 2012; Wirsich et al., 2016). However, Sun et al. (2014) observed
lower structure-function coupling in connectomes of patients with AD. Few studies have ex-
amined both structural and functional connectomes in AD, and therefore further investigation
regarding the relationship between structure and function is required.
There are several limitations to consider for this preliminary, pilot study. It is unclear why
functional connectomes failed to demonstrate expected small-worldness. This could be the
results of anesthesia, which has been shown to attenuate intrinsic functional networks
(Boveroux et al., 2010; Peltier et al., 2005). The effects of anesthesia on connectome organi-
zation are currently unknown. MRI field strength could play a role, although a previous study
Network Neuroscience
253
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
5XFAD multimodal mouse connectome
was also conducted at 7 Tesla (Liska et al., 2015). Another drawback is that fMRI and DTI
were acquired at different MRI field strengths. This study is also limited by the small sample,
which may have reduced our power to detect certain effects. However, we used a conservative
statistical approach, including permutation analysis with a large number of permutations and
correction for multiple comparisons where appropriate. Currently there is no standard regard-
ing the parcellation scheme for connectome analyses, and therefore a different approach may
yield alternate results. DTI-based connectomes have particular limitations, as they have been
shown to correspond poorly with neuron tracer data (Calabrese, Badea, Cofer, Qi, & Johnson,
2015). However, as noted above, DTI connectome properties have been shown to differentiate
patient groups and therefore seem to provide valuable insights regarding the effects of AD on
brain networks.
In conclusion, we demonstrated preliminary evidence that connectome properties of 5XFAD
transgenic mice show some correspondence with results observed in patients with AD. There
were several innovative aspects of this study, including connectome measurement in an AD
mouse model, multimodal connectome measurement, and the use of different network thresh-
olding methods. Future studies in mice could allow us to better understand the molecular
mechanisms underlying connectome disruption in AD. These models could also aid in drug
discovery and preclinical trials for AD by providing outcome measurements of connectome
organization.
ACKNOWLEDGMENTS
The authors would like to thank the faculty and staff of the MD Anderson Small Animal Imaging
Facility as well as Robia Pautler, PhD, and others at the Baylor College of Medicine Small
Animal MRI.
AUTHOR CONTRIBUTIONS
Shelli R. Kesler: Conceptualization; Data curation; Formal analysis; Funding acquisition; Inves-
tigation; Methodology; Project administration; Resources; Software; Supervision; Validation;
Visualization; Writing – original draft; Writing – review & editing. Paul Acton: Data curation;
Methodology; Resources; Writing – review & editing. Vikram Rao: Formal analysis; Method-
ology; Writing – review & editing. William J. Ray: Conceptualization; Data curation; Funding
acquisition; Investigation; Methodology; Project administration; Resources; Supervision; Vali-
dation; Visualization; Writing – review & editing.
FUNDING INFORMATION
This research was funded by the Neurodegeneration Consortium, the MD Anderson Founda-
tion, and the National Institutes of Health (1R03CA191559, 1R01NR014195, 1R01CA172145:
SK). The sponsors had no role in the design, implementation, analysis, or interpretation of the
study.
REFERENCES
Achard, S., & Bullmore, E. (2007). Efficiency and cost of economi-
cal brain functional networks. PLoS Computational Biology, 3(2),
e17. https://doi.org/10.1371/journal.pcbi.0030017
functional network with highly connected association cortical
hubs. Journal of Neuroscience, 26(1), 63–72. https://doi.org/10.
1523/JNEUROSCI.3874-05.2006
Achard, S., Salvador, R., Whitcher, B., Suckling, J., & Bullmore,
E. (2006). A resilient, low-frequency, small-world human brain
Bai, F., Watson, D. R., Shi, Y., Wang, Y., Yue, C., YuhuanTeng,
(2011). Specifically progressive deficits of brain
. . . Zhang, Z.
Network Neuroscience
254
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
5XFAD multimodal mouse connectome
functional marker in amnestic type mild cognitive impairment.
PLoS ONE, 6(9), e24271. https://doi.org/10.1371/journal.pone.
0024271
Bassett, D. S., & Bullmore, E. (2006). Small-world brain networks.
The Neuroscientist, 12(6), 512–523. https://doi.org/10.1177/
1073858406293182
Bassett, D. S., Bullmore, E., Verchinski, B. A., Mattay, V. S.,
Weinberger, D. R., & Meyer-Lindenberg, A.
(2008). Hierarchi-
cal organization of human cortical networks in health and
Journal of Neuroscience, 28(37), 9239–9248.
schizophrenia.
https://doi.org/28/37/9239 [pii] 10.1523/JNEUROSCI.1929-08.
2008
Bassett, D. S., Meyer-Lindenberg, A., Achard, S., Duke, T.,
& Bullmore, E. (2006). Adaptive reconfiguration of fractal small-
world human brain functional networks. Proceedings of the
National Academy of Sciences, 103(51), 19518–19523. https://
doi.org/10.1073/pnas.0606005103
Bassett, D. S., Nelson, B. G., Mueller, B. A., Camchong, J., & Lim,
K. O.
(2012). Altered resting state complexity in schizophre-
nia. NeuroImage, 59(3), 2196–2207. https://doi.org/10.1016/j.
neuroimage.2011.10.002
Bateman, R. J., Aisen, P. S., De Strooper, B., Fox, N. C., Lemere,
C. A., Ringman, J. M., . . . Xiong, C. (2011). Autosomal-dominant
Alzheimer’s disease: A review and proposal for the prevention
of Alzheimer’s disease. Alzheimer’s Research and Therapy, 3(1),
1–1. https://doi.org/10.1186/alzrt59
Behzadi, Y., Restom, K., Liau, J., & Liu, T. T. (2007). A component
based noise correction method (compcor) for BOLD and perfu-
sion based fMRI. NeuroImage, 37(1), 90–101.
Bloom, G. S.
Bero, A. W., Bauer, A. Q., Stewart, F. R., White, B. R., Cirrito,
J. R., Raichle, M. E., . . . Holtzman, D. M. (2012). Bidirectional
relationship between functional connectivity and amyloid-beta
deposition in mouse brain. Journal of Neuroscience, 32(13),
4334–4340. https://doi.org/10.1523/JNEUROSCI.5845-11.2012
(2014). Amyloid-beta and tau: The trigger and
JAMA Neurology,
bullet
71(4), 505–508. https://doi.org/10.1001/jamaneurol.2013.5847
Boveroux, P., Vanhaudenhuyse, A., Bruno, M.-A., Noirhomme, Q.,
Lauwick, S., Luxen, A., . . . Phillips, C.
(2010). Breakdown of
within- and between-network resting state functional magnetic
resonance imaging connectivity during propofol-induced loss of
consciousness. Journal of the American Society of Anesthesiolo-
gists, 113(5), 1038–1053.
in Alzheimer disease pathogenesis.
Brand, Martin D., & Nicholls, David G.
(2011). Assessing mito-
chondrial dysfunction in cells. Biochemical Journal, 435(Pt. 2),
297–312. https://doi.org/10.1042/BJ20110162
Brier, M. R., Thomas, J. B., Fagan, A. M., Hassenstab, J., Holtzman,
D. M., Benzinger, T. L., . . . Ances, B. M.
(2014). Functional
connectivity and graph theory in preclinical Alzheimer’s disease.
Neurobiology of Aging, 35(4), 757–768. https://doi.org/10.1016/
j.neurobiolaging.2013.10.081
Brown, J. A., Terashima, K. H., Burggren, A. C., Ercoli, L. M.,
Miller, K. J., Small, G. W., & Bookheimer, S. Y. (2011). Brain net-
work local interconnectivity loss in aging APOE-4 allele carri-
ers. Proceedings of the National Academy of Sciences, 108(51),
20760–20765. https://doi.org/10.1073/pnas.1109038108
Buckner, R. L., Sepulcre, J., Talukdar, T., Krienen, F. M., Liu, H.,
Hedden, T., . . . Johnson, K. A. (2009). Cortical hubs revealed by
intrinsic functional connectivity: Mapping, assessment of stabil-
ity, and relation to alzheimer’s disease. Journal of Neuroscience,
29(6), 1860–1873. https://doi.org/10.1523/JNEUROSCI.5062-
08.2009
Calabrese, E., Badea, A., Cofer, G., Qi, Y., & Johnson, G. A. (2015).
A diffusion MRI tractography connectome of the mouse brain
and comparison with neuronal tracer data. Cerebral Cortex,
25(11), 4628–4637. https://doi.org/10.1093/cercor/bhv121
Chen, H., Liu, T., Zhao, Y., Zhang, T., Li, Y., Li, M., . . . Liu, T.
(2015). Optimization of large-scale mouse brain connectome via
joint evaluation of DTI and neuron tracing data. NeuroImage,
115,202–213.https://doi.org/10.1016/j.neuroimage.2015.04.050
Contreras, J. A., Goni, J., Risacher, S. L., Sporns, O., & Saykin, A. J.
(2015). The structural and functional connectome and predic-
tion of risk for cognitive impairment in older adults. Current
Behavioral Neuroscience Reports, 2(4), 234–245. https://doi.org/
10.1007/s40473-015-0056-z
Dai, Z., & He, Y. (2014). Disrupted structural and functional brain
connectomes in mild cognitive impairment and Alzheimer’s
disease. Neuroscience Bulletin, 30(2), 217–232. https://doi.org/
10.1007/s12264-013-1421-0
Dai, Z., Yan, C., Li, K., Wang, Z., Wang, J., Cao, M., . . . He, Y.
(2015).
Identifying and mapping connectivity patterns of brain
network hubs in alzheimer’s disease. Cerebral Cortex, 25(10),
3723–3742. https://doi.org/10.1093/cercor/bhu246
Daianu, M., Jahanshad, N., Nir, T. M., Toga, A. W., Jack, C. R., Jr.,
Weiner, M. W., & Thompson, P. M., for the Alzheimer’s Disease
Neuroimaging Initiative. (2013). Breakdown of brain connectiv-
ity between normal aging and Alzheimer’s disease: A structural
k-core network analysis. Brain Connectivity, 3(4), 407–422.
https://doi.org/10.1089/brain.2012.0137
Damoiseaux, J. S., Prater, K. E., Miller, B. L., & Greicius, M. D.
(2012). Functional connectivity tracks clinical deterioration in
Alzheimer’s disease. Neurobiology of Aging, 33(4), 828.e19–
828.e30. https://doi.org/10.1016/j.neurobiolaging.2011.06.024
de Haan, W., Pijnenburg, Y. A. L., Strijers, R. L. M., van der Made,
Y., van der Flier, W. M., Scheltens, P., & Stam, C. J. (2009). Func-
tional neural network analysis in frontotemporal dementia and
Alzheimer’s disease using EEG and graph theory. BMC Neuro-
science, 10, 101. https://doi.org/10.1186/1471-2202-10-101
Drakesmith, M., Caeyenberghs, K., Dutt, A., Lewis, G., David, A. S.,
& Jones, D. K. (2015). Overcoming the effects of false positives
and threshold bias in graph theoretical analyses of neuroimag-
ing data. NeuroImage, 118, 313–333. https://doi.org/10.1016/j.
neuroimage.2015.05.011
Eimer, W. A., & Vassar, R. (2013). Neuron loss in the 5XFAD mouse
model of Alzheimer’s disease correlates with intraneuronal Aβ42
accumulation and Caspase-3 activation. Molecular Neurodegen-
eration, 8(1), 2. https://doi.org/10.1186/1750-1326-8-2
Fischer, F. U., Wolf, D., Scheurich, A., Fellgiebel, A., & Alzheimer’s
(2015). Altered whole-brain
Disease Neuroimaging Initiative.
white matter networks in preclinical Alzheimer’s disease. Neu-
roImage: Clinical, 8, 660–666. https://doi.org/10.1016/j.nicl.
2015.06.007
Network Neuroscience
255
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
5XFAD multimodal mouse connectome
Fornito, A., Zalesky, A., & Breakspear, M. (2013). Graph analysis of
the human connectome: Promise, progress, and pitfalls. Neuro-
Image, 80, 426–444. https://doi.org/10.1016/j.neuroimage.2013.
04.087
Gong, G., He, Y., Chen, Z. J., & Evans, A. C. (2012). Convergence
and divergence of thickness correlations with diffusion connec-
tions across the human cerebral cortex. NeuroImage, 59(2),
1239–1248. https://doi.org/10.1016/j.neuroimage.2011.08.017
Goñi, J., van den Heuvel, M. P., Avena-Koenigsberger, A., Velez de
Mendizabal, N., Betzel, R. F., Griffa, A., . . . Sporns, O. (2014).
Resting-brain functional connectivity predicted by analytic mea-
sures of network communication. Proceedings of the National
Academy of Sciences, 111(2), 833–838. https://doi.org/10.1073/
pnas.1315529111
Gorges, M., Roselli, F., Müller, H.-P., Ludolph, A. C., Rasche, V.,
& Kassubek, J. (2017). Functional connectivity mapping in the
animal model: Principles and applications of resting-state fMRI.
Frontiers in Neurology, 8(200). https://doi.org/10.3389/fneur.
2017.00200
Grandjean, J., Schroeter, A., He, P., Tanadini, M., Keist, R., Krstic,
D., . . . Rudin, M. (2014). Early alterations in functional connec-
tivity and white matter structure in a transgenic mouse model of
cerebral amyloidosis. Journal of Neuroscience, 34(41), 13780–
13789. https://doi.org/10.1523/JNEUROSCI.4762-13.2014
Green, R. C., Cupples, L., Go, R., Benke, K. S., Edeki, T., Griffith,
(2002). Risk of dementia among White
P. A., . . . Farrer, L. A.
and African American relatives of patients with Alzheimer
disease. JAMA, 287(3), 329–336. https://doi.org/10.1001/jama.
287.3.329
Hardy,
J.
J., & Selkoe, D.
(2002). The amyloid hypothesis of
Alzheimer’s disease: Progress and problems on the road to thera-
peutics. Science, 297(5580), 353–356. https://doi.org/10.1126/
science.1072994
Hebert, L. E., Bienias, J. L., Aggarwal, N. T., Wilson, R. S., Bennett,
(2010). Change in risk of
D. A., Shah, R. C., & Evans, D. A.
Alzheimer disease over time. Neurology, 75(9), 786–791. https://
doi.org/10.1212/WNL.0b013e3181f0754f
Hosseini, S. M., & Kesler, S. R. (2013). Comparing connectivity pat-
tern and small-world organization between structural correlation
and resting-state networks in healthy adults. NeuroImage, 78,
402–414. https://doi.org/10.1016/j.neuroimage.2013.04.032
Humphries, M. D., & Gurney, K.
(2008). Network “small-world-
ness”: A quantitative method for determining canonical net-
work equivalence. PLoS One, 3(4), e0002051. https://doi.org/10.
1371/journal.pone.0002051
Kaiser, M., & Hilgetag, C. C. (2006). Nonoptimal component place-
ment, but short processing paths, due to long-distance projec-
tions in neural systems. PLoS Computational Biology, 2(7), e95.
https://doi.org/10.1371/journal.pcbi.0020095
Kesler, S. R., Acton, P., Rao, V., & Ray, W. J.
(2018). Supplemen-
tal material for “Functional and structural connectome properties
in the 5XFAD transgenic mouse model of Alzheimer’s disease.”
Network Neuroscience, 2(2), 241–258. https://doi.org/10.1162/
netn_a_00048
Kesler, S. R., Adams, M., Packer, M., Rao, V., Henneghan, A. M.,
Blayney, D. W., & Palesh, O.
(2017). Disrupted brain network
functional dynamics and hyper-correlation of structural and func-
tional connectome topology in patients with breast cancer prior
to treatment. Brain and Behavior, 7(3), e00643. https://doi.org/
10.1002/brb3.643
Kesler, S. R., Watson, C. L., & Blayney, D. W. (2015). Brain network
alterations and vulnerability to simulated neurodegeneration in
breast cancer. Neurobiology of Aging, 36(8), 2429–2442. https://
doi.org/10.1016/j.neurobiolaging.2015.04.015
Kim, H. J., Shin, J. H., Han, C. E., Kim, H. J., Na, D. L., Seo, S. W.,
. . . Alzheimer’s Disease Neuroimaging Initiative. (2016). Using
individualized brain network for analyzing structural covariance
of the cerebral cortex in Alzheimer’s patients. Frontiers in Neuro-
science, 10, 394. https://doi.org/10.3389/fnins.2016.00394
Lehmann, M., Madison, C. M., Ghosh, P. M., Seeley, W. W.,
Mormino, E., Greicius, M. D., . . . Rabinovici, G. D. (2013). In-
trinsic connectivity networks in healthy subjects explain clinical
variability in Alzheimer’s disease. Proceedings of the National
Academy of Sciences, 110(28), 11606–11611. https://doi.org/
10.1073/pnas.1221536110
Lin, M. T., & Beal, M. F. (2006). Mitochondrial dysfunction and ox-
idative stress in neurodegenerative diseases. Nature, 443(7113),
787–795. https://doi.org/10.1038/nature05292
Liska, A., Galbusera, A., Schwarz, A. J., & Gozzi, A. (2015). Func-
tional connectivity hubs of the mouse brain. NeuroImage. https://
doi.org/10.1016/j.neuroimage.2015.04.033
Lloret, A., Fuchsberger, T., Giraldo, E., & Vina, J.
(2015). Molec-
ular mechanisms linking amyloid beta toxicity and Tau hyper-
phosphorylation in Alzheimer’s disease. Free Radical Biology
and Medicine, 83, 186–191. https://doi.org/10.1016/j.freeradbio
med.2015.02.028
Lo, C. Y., Wang, P. N., Chou, K. H., Wang, J., He, Y., & Lin, C. P.
(2010). Diffusion tensor tractography reveals abnormal topolog-
ical organization in structural cortical networks in Alzheimer’s
disease. Journal of Neuroscience, 30(50), 16876–16885. https://
doi.org/10.1523/JNEUROSCI.4136-10.2010
Ma, Y., Hof, P. R., Grant, S. C., Blackband, S. J., Bennett, R., Slatest,
L., . . . Benveniste, H. (2005). A three-dimensional digital atlas
database of the adult C57BL/6J mouse brain by magnetic reso-
nance microscopy. Neuroscience, 135(4), 1203–1215. https://
doi.org/10.1016/j.neuroscience.2005.07.014
Mallio, C. A., Schmidt, R., de Reus, M. A., Vernieri, F., Quintiliani,
L., Curcio, G., . . . van den Heuvel, M. P.
(2015). Epicentral
disruption of structural connectivity in Alzheimer’s disease. CNS
Neuroscience and Therapeutics, 21(10), 837–845. https://doi.
org/10.1111/cns.12397
Meghanathan, N.
(2016). A greedy algorithm for neighborhood
overlap-based community detection. Algorithms, 9(1). https://
doi.org/10.3390/a9010008
(2006).
Oakley, H., Cole, S. L., Logan, S., Maus, E., Shao, P., Craft, J.,
Intraneuronal beta-amyloid aggregates,
. . . Vassar, R.
neurodegeneration, and neuron loss in transgenic mice with five
familial Alzheimer’s disease mutations: Potential factors in amy-
loid plaque formation. Journal of Neuroscience, 26(40), 10129–
10140. https://doi.org/10.1523/JNEUROSCI.1202-06.2006
Oh, S. W., Harris, J. A., Ng, L., Winslow, B., Cain, N., Mihalas, S.,
. . . Zeng, H.
(2014). A mesoscale connectome of the mouse
brain. Nature, 508(7495), 207–214. https://doi.org/10.1038/
nature13186
Network Neuroscience
256
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
5XFAD multimodal mouse connectome
Peltier, S. J., Kerssens, C., Hamann, S. B., Sebel, P. S., Byas-Smith,
M., & Hu, X.
(2005). Functional connectivity changes with
concentration of sevoflurane anesthesia. NeuroReport, 16(3),
285–288.
Pereira, J. B., Mijalkov, M., Kakaei, E., Mecocci, P., Vellas, B.,
Tsolaki, M., . . . Westman, E. (2016). Disrupted network topology
in patients with stable and progressive mild cognitive impairment
and Alzheimer’s disease. Cerebral Cortex, 26(8), 3476–3493.
https://doi.org/10.1093/cercor/bhw128
Prince, M., Wimo, A., Guerchet, M., Ali, G.-C., Wu, Y.-T., & Prina,
M. (2015). World Alzheimer report 2015: The global impact of
dementia. Retrieved from http://www.worldalzreport2015.org/
Raichle, M. E. (2011). The restless brain. Brain Connectivity, 1(1),
3–12. https://doi.org/10.1089/brain.2011.0019
Reijmer, Y. D., Leemans, A., Caeyenberghs, K., Heringa, S. M.,
Koek, H. L., & Biessels, G.
(2013). Disruption of cere-
J.
bral networks and cognitive impairment in Alzheimer disease.
Neurology, 80(15), 1370–1377. https://doi.org/10.1212/WNL.
0b013e31828c2ee5
Risacher, S. L., & Saykin, A. J.
(2013). Neuroimaging and other
biomarkers for Alzheimer’s disease: The changing landscape of
early detection. Annual Review of Clinical Psychology, 9, 621–
648. https://doi.org/10.1146/annurev-clinpsy-050212-185535
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003
(2015).
Rubinov, M., Ypma, R. J., Watson, C., & Bullmore, E. T.
Wiring cost and topological participation of
the mouse
brain connectome. Proceedings of the National Academy of
Sciences, 112(32), 10032–10037. https://doi.org/10.1073/pnas.
1420315112
J., Schoonheim, M. M., Damoiseaux,
Rudie, J. D., Brown, J. A., Beck-Pancer, D., Hernandez, L. M.,
Dennis, E. L., Thompson, P. M., . . . Dapretto, M. (2012). Altered
functional and structural brain network organization in autism.
NeuroImage: Clinical, 2, 79–94. https://doi.org/10.1016/j.nicl.
2012.11.006
Sanz-Arigita, E.
J. S.,
Rombouts, S. A., Maris, E., Barkhof, F., . . . Stam, C. J.
(2010).
Loss of “small-world” networks in Alzheimer’s disease: Graph
analysis of fMRI resting-state functional connectivity. PLoS One,
5(11), e13788. https://doi.org/10.1371/journal.pone.0013788
Saunders, A. M., Strittmatter, W. J., Schmechel, D., St. George-
Hyslop, P. H., Pericak-Vance, M. A., Joo, S. H., . . . Roses,
A. D. (1993). Association of apolipoprotein E allele ∈4 with
late-onset familial and sporadic alzheimer’s disease. Neurology,
43(8), 1467–1472.
Schwarz, A. J., & McGonigle, J. (2011). Negative edges and soft
thresholding in complex network analysis of resting state func-
tional connectivity data. NeuroImage, 55(3), 1132–1146. https://
doi.org/10.1016/j.neuroimage.2010.12.047
Shah, D., Jonckers, E., Praet, J., Vanhoutte, G., Delgado y Palacios,
R., Bigot, C., et al.
(2013). Resting state fMRI reveals dimin-
ished functional connectivity in a mouse model of amyloidosis.
PLoS ONE, 8(12), e84241. https://doi.org/10.1371/journal.pone.
0084241
Simic, G., Babic, M., Borovecki, F., & Hof, P. R. (2014). Early failure
of the default-mode network and the pathogenesis of Alzheimer’s
disease. CNS Neuroscience and Therapeutics, 20(7), 692–698.
https://doi.org/10.1111/cns.12260
Smith, S. M., Jenkinson, M., Woolrich, M. W., Beckmann, C. F.,
Behrens, T. E., Johansen-Berg, H., . . . Matthews, P. M. (2004).
Advances in functional and structural MR image analysis and
implementation as FSL. NeuroImage, 23(Suppl. 1), S208–S219.
https://doi.org/10.1016/j.neuroimage.2004.07.051
Sporns, O., & Betzel, R. F. (2016). Modular brain networks. Annual
Review of Psychology, 67, 613–640. https://doi.org/10.1146/
annurev-psych-122414-033634
Stafford, J. M., Jarrett, B. R., Miranda-Dominguez, O., Mills, B. D.,
Cain, N., Mihalas, S., . . . Fair, D. A. (2014). Large-scale topol-
ogy and the default mode network in the mouse connectome.
Proceedings of the National Academy of Sciences, 111(52),
18745–18750. https://doi.org/10.1073/pnas.1404346111
J., de Haan, W., Daffertshofer, A.,
Jones, B. F.,
.
I., van Cappellen van Walsum, A. M.,
Manshanden,
Scheltens, P.
(2009). Graph theoretical analysis of magne-
toencephalographic functional connectivity in Alzheimer’s dis-
ease. Brain, 132(Pt. 1), 213–224. https://doi.org/10.1093/brain/
awn262
Stam, C.
.
.
Stam, C. J., Jones, B. F., Nolte, G., Breakspear, M., & Scheltens,
P. (2007). Small-world networks and functional connectivity in
Alzheimer’s disease. Cerebral Cortex, 17(1), 92–99. https://doi.
org/10.1093/cercor/bhj127
Stancu, I.-C., Vasconcelos, B., Terwel, D., & Dewachter, I. (2014).
Models of β-amyloid induced Tau-pathology: The long and
“folded” road to understand the mechanism. Molecular Neuro-
degeneration, 9(1), 51. https://doi.org/10.1186/1750-1326-9-51
Sun, Y., Yin, Q., Fang, R., Yan, X., Wang, Y., Bezerianos, A., . . .
(2014). Disrupted functional brain connectivity and its
Sun, J.
association to structural connectivity in amnestic mild cognitive
impairment and Alzheimer’s disease. PLoS One, 9(5), e96505.
https://doi.org/10.1371/journal.pone.0096505
Supekar, K., Menon, V., Rubin, D., Musen, M., & Greicius, M. D.
(2008). Network analysis of intrinsic functional brain connec-
tivity in Alzheimer’s disease. PLoS Computational Biology, 4(6),
e1000100. https://doi.org/10.1371/journal.pcbi.1000100
ten Kate, M., Sanz-Arigita, E. J., Tijms, B. M., Wink, A. M., Clerigue,
M., Garcia-Sebastian, M., . . . Villanua, J. (2016). Impact of
APOE-ε4 and family history of dementia on gray matter atro-
phy in cognitively healthy middle-aged adults. Neurobiology of
Aging, 38, 14–20.
Tijms, B. M., Kate, M. T., Wink, A. M., Visser, P. J., Ecay, M.,
Clerigue, M., . . . Barkhof, F. (2015). Gray matter network disrup-
tions and amyloid beta in cognitively normal adults. Neurobiol-
ogy of Aging. https://doi.org/10.1016/j.neurobiolaging.2015.10.
015
Tijms, B. M., Moller, C., Vrenken, H., Wink, A. M., de Haan, W.,
van der Flier, W. M., . . . Barkhof, F. (2013). Single-subject grey
matter graphs in Alzheimer’s disease. PLoS One, 8(3), e58921.
https://doi.org/10.1371/journal.pone.0058921
Tijms, B. M., Wink, A. M., de Haan, W., van der Flier, W. M., Stam,
C. J., Scheltens, P., & Barkhof, F.
(2013). Alzheimer’s disease:
Connecting findings from graph theoretical studies of brain net-
works. Neurobiology of Aging, 34(8), 2023–2036. https://doi.
org/10.1016/j.neurobiolaging.2013.02.020
Network Neuroscience
257
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
5XFAD multimodal mouse connectome
van den Heuvel, M. P., Bullmore, E. T., & Sporns, O. (2016). Com-
parative connectomics. Trends in Cognitive Sciences, 20(5), 345–
361. https://doi.org/10.1016/j.tics.2016.03.001
van Wijk, B. C. M., Stam, C. J., & Daffertshofer, A. (2010). Com-
paring brain networks of different size and connectivity density
using graph theory. PLoS ONE, 5(10), e13701.
Wang, J., Zuo, X., Dai, Z., Xia, M., Zhao, Z., Zhao, X., et al. (2012).
Disrupted functional brain connectome in individuals at risk
for Alzheimer’s disease. Biological Psychiatry. https://doi.org/10.
1016/j.biopsych.2012.03.026
Wang, R., Benner, T., Sorensen, A. G., & Wedden, V. J. (2007).
Diffusion Toolkit: A software package for diffusion imaging data
processing and tractography. Proceedings of the International
Society for Magnetic Resonance in Medicine, 15, 3720.
Wang, Y., Ghumare, E., Vandenberghe, R., & Dupont, P.
(2017).
Comparison of different generalizations of clustering coefficient
and local efficiency for weighted undirected graphs. Neural
Computation, 29(2), 313–331. https://doi.org/10.1162/NECO_
a_00914
Watts, D. J., & Strogatz, S. H. (1998). Collective dynamics of “small-
world” networks. Nature, 393(6684), 440–442. https://doi.org/
10.1038/30918
Whitfield-Gabrieli, S., & Ford, J. M. (2012). Default mode network
activity and connectivity in psychopathology. Annual Review of
Clinical Psychology, 8, 49–76. https://doi.org/10.1146/annurev-
clinpsy-032511-143049
Wirsich, J., Perry, A., Ridley, B., Proix, T., Golos, M., Benar, C.,
. . . Guye, M.
(2016). Whole-brain analytic measures of net-
work communication reveal increased structure-function corre-
lation in right temporal lobe epilepsy. NeuroImage: Clinical, 11,
707–718. https://doi.org/10.1016/j.nicl.2016.05.010
Wolters, F. J., van der Lee, S. J., Koudstaal, P. J., van Duijn, C. M.,
Hofman, A., Ikram, M. K., . . . Ikram, M. A.
(2017). Parental
family history of dementia in relation to subclinical brain
disease and dementia risk. Neurology, 88(17), 1642–1649.
Xie, T., & He, Y. (2011). Mapping the Alzheimer’s brain with
connectomics. Frontiers in Psychiatry, 2, 77. https://doi.org/10.
3389/fpsyt.2011.00077
Yao, Z., Zhang, Y., Lin, L., Zhou, Y., Xu, C., & Jiang, T. (2010).
Abnormal cortical networks in mild cognitive impairment
and Alzheimer’s disease. PLoS Computational Biology, 6(11),
e1001006. https://doi.org/10.1371/journal.pcbi.1001006
Zalesky, A., Fornito, A., & Bullmore, E. (2012). On the use of
correlation as a measure of network connectivity. NeuroImage,
60(4), 2096–2106. https://doi.org/10.1016/j.neuroimage.2012.
02.001
Zalesky, A., Fornito, A., & Bullmore, E. T. (2010). Network-based
statistic: Identifying differences in brain networks. NeuroImage,
53(4), 1197–1207. https://doi.org/10.1016/j.neuroimage.2010.
06.041
Zhao, X., Liu, Y., Wang, X., Liu, B., Xi, Q., Guo, Q., . . . Wang,
P.
(2012). Disrupted small-world brain networks in moderate
Alzheimer’s disease: A resting-state fMRI study. PLoS One, 7(3),
e33540. https://doi.org/10.1371/journal.pone.0033540
Zhou, J., Gennatas, E. D., Kramer, J. H., Miller, B. L., & Seeley,
W. W.
(2012). Predicting regional neurodegeneration from
the healthy brain functional connectome. Neuron, 73(6), 1216–
1227. https://doi.org/10.1016/j.neuron.2012.03.004
Zhou,
J., & Seeley, W. W.
(2014). Network dysfunction in
Alzheimer’s disease and frontotemporal dementia: Implications
for psychiatry. Biological Psychiatry, 75(7), 565–573. https://doi.
org/10.1016/j.biopsych.2014.01.020
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
0
2
0
2
2
4
1
1
0
9
2
2
0
7
n
e
n
_
a
_
0
0
0
4
8
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
258