Estradiol and the Catechol-o-methyltransferase Gene
Interact to Predict Working Memory Performance:
A Replication and Extension
Courtney C. Louis1
, Emily Jacobs2, Mark D’Esposito3, and Jason Moser1
Astratto
■ Decades of evidence across taxa have established the impor-
tance of dopamine (DA) signaling in the pFC for successful
working memory performance. Genetic and hormonal factors
can shape individual differences in prefrontal DA tone. IL
catechol-o-methyltransferase (COMT) gene regulates basal
prefrontal DA, and the sex hormone 17β-estradiol potentiates
DA release. E. Jacobs, & M. D’Esposito [Estrogen shapes
dopamine-dependent cognitive processes: Implications for
women’s health. Journal of Neuroscience, 31, 5286–5293,
2011] investigated the moderating role of estradiol on cognition
using the COMT gene and COMT enzymatic activity as a
proxy for pFC DA tone. They found that increases in 17β-
estradiol within women at two time points during the menstrual
cycle influenced working memory performance in a COMT-
dependent manner. Here, we aimed to replicate and extend
the behavioral findings of Jacobs and D’Esposito by employing
an intensive repeated-measures design across a full menstrual
cycle. Our results replicated the original investigation. Within-
person increases in estradiol were associated with improved
performance on 2-back lure trials for participants with low
basal levels of DA ( Val/ Val carriers). The association was in
the opposite direction for participants with higher basal levels
of DA (Met/Met carriers). Our findings support the role of
estrogen in DA-related cognitive functions and further high-
light the need to consider gonadal hormones in cognitive
science research. ■
INTRODUCTION
Working memory refers to the ability to store and manip-
ulate mental representations. It is a key function involved
in goal-directed behaviors and supports other executive
functions. Since the pioneering work of Goldman-Rakic
(Brozoski, Brown, Rosvold, & Goldman, 1979), numerous
studies have established the critical role of dopamine (DA)
signaling within the pFC for regulating delay-period activ-
ity and working memory performance. DA’s influence on
pFC-dependent cognitive processes follows an inverted-U
function, such that too little or too much DA can hinder
performance, whereas moderate levels contribute to
enhanced performance (Arnsten, Wang, & Paspalas,
2015; Cools & D’Esposito, 2011).
In humans, many studies have investigated the associa-
tion between DA and cognition using the catechol-o-
methyltransferase (COMT) gene as a proxy for basal
prefrontal DA tone. The COMT gene (codon 158) gained
particular interest with respect to pFC-dependent cogni-
tive functions because of its unique role in the metaboliza-
tion of DA in the mesocortical pathway that innervates
pFC. Unlike other dopaminergic projections, ad esempio
those innervating the striatum, COMT enzyme accounts
for >60% of DA metabolism in pFC (Tunbridge et al.,
1Michigan State University, East Lansing, 2University of Califor-
nia, Santa Barbara, 3University of California, Berkeley
© 2023 Istituto di Tecnologia del Massachussetts
2019; Yavich, Forsberg, Karayiorgou, Gogos, & Männistö,
2007; Männistö & Kaakkola, 1999). Così, the COMT gene
provides a useful proxy for individual differences in pre-
frontal DA tone—that is, a person’s estimated “baseline”
position on the DA-working memory inverted-U curve.
Individuals homozygous for the Val allele have enhanced
COMT enzyme activity and less basal DA availability in
pFC, whereas those homozygous for the Met allele have
reduced COMT activity leading to more DA availability
(Tunbridge, Harrison, & Weinberger, 2006; Egan et al.,
2001). In this way, homozygous Met carriers are thought
to have enhanced DA levels under baseline conditions,
whereas homozygous Val carriers are thought to have
relatively less prefrontal DA. Empirical studies provide
support for this classification, such that Met carriers have
more COMT enzyme activity (Tunbridge et al., 2019), less
D1 binding (Slifstein et al., 2008), enhanced perfor-
mance, and enhanced cortical efficiency relative to Val
carriers (Smith, Swift-Scanlan, & Boettiger, 2014; Cools
& D’Esposito, 2011; Mier, Kirsch, & Meyer-Lindenberg,
2010; Meyer-Lindenberg et al., 2006; Egan et al., 2001).
Importantly, Tuttavia, the COMT gene is not determin-
istic of cognitive performance. Dopaminergic activity can
be influenced by many factors, including sex steroid hor-
mones. Animal studies have shown that 17β-estradiol
levels (“estradiol”) stimulate DA release and turnover
(Xiao & Becker, 1994; Becker, 1990), leading to greater
Journal of Cognitive Neuroscience 35:7, pag. 1144–1153
https://doi.org/10.1162/jocn_a_02001
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
C
N
_
UN
_
0
2
0
0
1
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
efficiency within pFC cortical circuits. A recent multimodal
PET imaging study revealed that DA synthesis capacity and
cognitive flexibility differ between hormonal contracep-
tive users and non-users (Taylor et al., 2022). Further-
more, estrogen receptors are expressed in regions that
receive major projections from midbrain DA neurons,
including pFC (Björklund & Dunnett, 2007). Some evi-
dence suggests that circulating estradiol concentrations
impact working memory performance in young adult
female populations,1 such that increased concentrations
are associated with enhanced performance (Hampson &
Morley, 2013). Note that one study found that the relation-
ship between estradiol and working memory may follow
an inverted-U curve (cioè., a quadratic effect of estradiol) In
a young adult female sample; Tuttavia, the sample size
was small (n = 8) and the finding has not been replicated
(Rosenberg & Park, 2002). Inoltre, studies examining
estradiol’s effect on verbal working memory in the ovula-
tory phase (when estradiol is heightened) are few and
mixed (Bernal & Paolieri, 2022). Very few studies have
considered Gene × hormone interactions that may shape
cognition. Theories posit that estradiol exhibits DA-agonist
effects and may therefore modulate a person’s putative
position on the inverted-U curve between DA and working
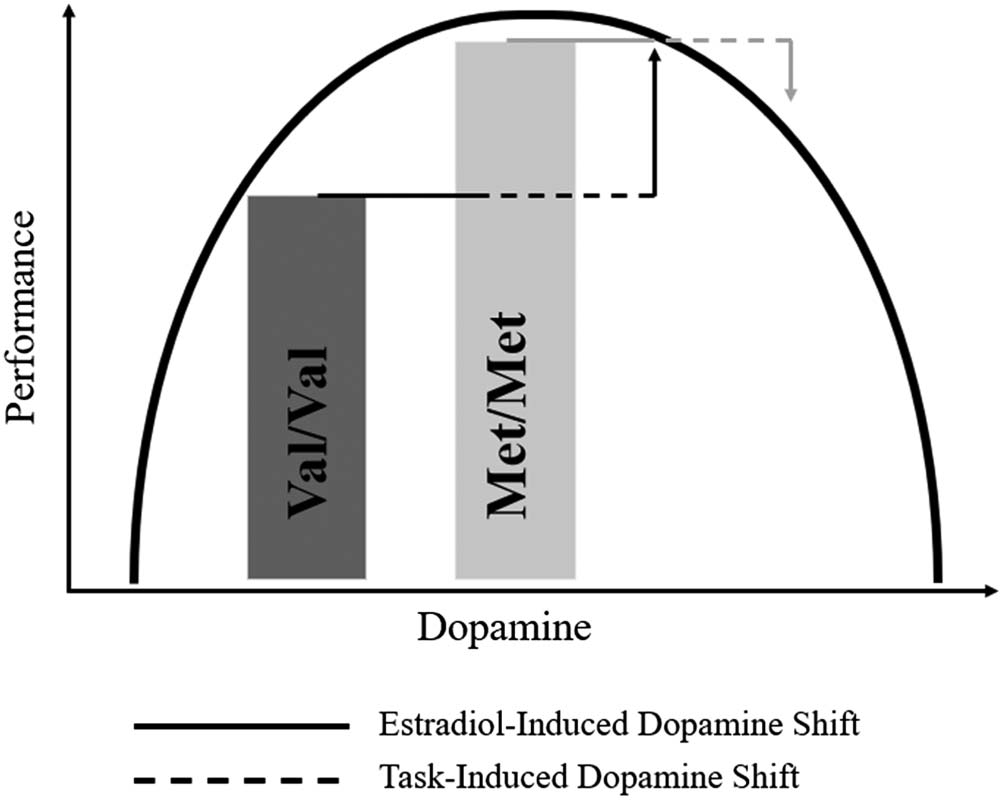
memory performance (Guarda la figura 1). Infatti, estradiol
downregulates COMT enzyme activity by inhibiting
COMT gene transcription, which may shift Val carriers
to an optimal range for performance on DA-dependent
cognitive tasks while hindering Met carriers ( Jiang, Xie,
Ramsden, & Ho, 2003).
Although studies have highlighted the powerful role of
estradiol in cognition in rodents (Shansky & Lipps, 2013;
Shansky et al., 2004; Bimonte & Denenberg, 1999; Luine,
Richards, Wu, & Beck, 1998), monkeys (Hara et al., 2014,
2016; Kromrey, Czoty, & Nader, 2015), and midlife women
(Dumas, Makarewicz, Bunn, Nickerson, & McGee, 2018;
Hampson, 2018; Jacobs & Goldstein, 2018), the literature
linking sex hormones and cognition in young adults has
yielded inconsistent results. Jacobs and D’Esposito
(2011) propose that studies that seek to draw a relation-
ship between menstrual cycle stage and working memory
performance fail to replicate because they do not account
for individual differences in basal DA levels. To test this,
their study followed a within-person design, in which 24
women (13 Val/ Val; 8 Met/Met; 3 Met/ Val) underwent
fMRI scanning at low and high estradiol phases of their
menstrual cycle, while completing a verbal n-back task
to measure working memory performance. Serum con-
centrations of 17β-estradiol were determined via liquid
chromatography mass-spectrometry. COMT enzyme activ-
ity was assessed via serum, providing an individual-level
proxy of prefrontal DA tone, in addition to COMT geno-
type. The authors found that on the 2-back condition
of cognitively demanding lure trials, Val carriers had
enhanced accuracy when estradiol was high versus low.
In contrasto, performance for Met carriers was highest
when estradiol levels were low, and reduced when estra-
diol levels were high. These results were also reflected at
the neural level. Val carriers at low estradiol levels showed
increased neural activity on high demanding trials, Quale
the authors interpreted as reduced neural efficiency.
These findings were the first to support the modulating
role of estradiol in the association between DA and
working memory function in young adult female samples.
Subsequent neuroimaging studies have extended their
findings, revealing sex-dependent effects of COMT geno-
type on prefrontal-mediated behaviors and pFC function
(Elton, Smith, Parrish, & Boettiger, 2017) as well as inter-
actions between cycle phase and basal DA on executive
functions (Hidalgo-Lopez & Pletzer, 2017).
Jacobs and D’Esposito’s findings were specific to the
2-back load condition of the task, and not the 3-back con-
dizione. They posit that this is because of a “task-induced
dopamine shift” related to the increased cognitive
demand of the 3-back condition (Guarda la figura 1). Higher
levels of cognitive demand often place stress on the
Figura 1. Figure showing the
modulatory role of estradiol on
COMT to affect working
memory performance. Increased
estradiol levels lead to reductions
in COMT enzyme protein
expression that result in more
dopamine in the pFC. IL
reduction in COMT enzyme
activity moves people along the
inverted-U curve, Perciò
enhancing performance for
those with lower basal dopamine
levels ( Val/ Val carriers), while
hurting performance for those
with higher basal levels (Met/Met
carriers). Task difficulty further
serves to lessen performance,
pushing individuals further along
the inverted U.
Louis et al.
1145
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
C
N
_
UN
_
0
2
0
0
1
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
individual and cognitive system, which can also increase
dopaminergic signaling (Arnsten, 2015; Shanmugan &
Epperson, 2014; Shansky & Lipps, 2013; Williams &
Castner, 2006). Così, interactive effects of estradiol and
DA are likely not to be consistent across levels of cogni-
tive load and, relatedly, stress. For this reason, under-
standing the COMT and estradiol interaction at various
difficulty levels may be important for specifying the con-
texts in which their interactive effects are evident.
The current study aimed to examine whether the behav-
ioral findings reported by Jacobs and D’Esposito (2011)
would replicate in a larger sample (n = 74) using the same
cognitive task. The current study built upon the prior
investigation by having up to four repeated assessments
within a person. This allowed us to test our effects with
more statistical power and use estradiol as a continuous
measure to examine its predictive value more precisely
for cognitive performance. Inoltre, our design allowed
us to assess whether an individual’s average estradiol
levels across the menstrual cycle would impact the associ-
ation between COMT and cognitive performance. IL
inclusion of between-centered estradiol levels allowed us
to delineate between- and within-person estradiol effects
more clearly. We predicted that within-person effects of
estradiol would follow those reported in Jacobs and
D’Esposito (2011), such that Val carriers would show
improved performance when estradiol is high, whereas
Met carriers would show the opposite effect. Jacobs and
D’Esposito reported that this effect is specific to the 2-back
condition, as the 3-back condition results in a “task-
induced dopamine shift” because of increased cognitive
demand and hinders performance irrespective of changes
in estradiol levels. Therefore, we did not predict that there
would be a COMT × Estradiol interaction on the 3-back
condition.
To our knowledge, no study has examined whether
between-persons levels of estradiol (cioè., average estradiol
levels between people) would impact the association
between COMT and cognitive performance in a young
adult sample. One study in postmenopausal women did
not find a COMT × Estradiol Level (between-groups)
interaction (Dumas et al., 2018) on n-back performance.
Another study completed a between-groups analysis
investigating the effect of COMT on women pre- and post-
menopause (which they defined as hormonal status).
They found that hormonal status moderated the effects
of COMT, such that there was no effect of COMT in women
premenopause, whereas there was a distinct effect of
COMT in the expected direction in women postmeno-
pause (Papaleo, Sannino, Piras, & Spalletta, 2015). Criti-
cally, these findings did not consider menstrual cycle
phase in the premenopausal women. Combined, these
findings suggest that between-persons levels of estradiol
may not evidence the expected COMT × estradiol interac-
tive effect in a young adult sample. Inoltre, because of
research indicating that within-person changes in estradiol
in particular influence DA tone, we did not expect that
between-persons levels of estradiol would moderate the
association between COMT and cognitive performance.
Examining this hypothesis allowed us to further specify
whether the estradiol’s modulating effect is specific to
within-person changes.
METHODS
Participants
Data from a subset of these participants have been
reported elsewhere to answer a different research ques-
tion involving anxiety and not estradiol (Louis, D’Esposito,
& Moser, 2021). The findings below are, Perciò, risultati
from a novel set of research aims and analyses. The sample
consisted of individuals with a consistent pattern of
menstrual cycles between 22 E 32 days (considered
the typical cycle length). Inoltre, participants could
not be using hormonal contraceptives (including pill,
patch, or intrauterine device), as they affect the fluctuation
of sex steroid hormones across the menstrual cycle. Fur-
thermore, participants could not have a previous diagnosis
that affected the neuroendocrine system (per esempio., polycystic
ovary syndrome, endometriosis) or be on any psychotro-
pic medication. One hundred thirty-nine individuals were
genotyped for the COMT Val158/Met (rs4680) polymor-
phism. Of these, 74 of them were homozygous allele
carriers (33 Met/Met and 41 Val/ Val carriers), which com-
prised the final sample included in the analyses.
The mean age was 20.63 (SD = 1.60). The sample
mostly consisted of individuals who identified as White
(63%), followed by individuals who identified as Black
(24%), Asian (8%), and those who identified as more than
one race (5%). For gender identity, one participant identi-
fied as nonbinary, and the remainder of the sample iden-
tified as women. Most of the sample consisted of those
who completed partial college (55%); Tuttavia, many
had a college level education (32%), completed a high
school level of education (12%), or had a graduate level
formazione scolastica (1%). For income levels, 49% of the sample
reported an annual household income of $50,000 or less, whereas 51% of the sample reported an annual household income > $50,000. Most of the sample consisted of stu-
dents enrolled full or part time (78%), and most of the
sample reported being financially supported by someone
else in the past year (71%).
Materials
n-back
Working memory was measured with the verbal n-back
task ( Jacobs & D’Esposito, 2011; Kirchner, 1958). For each
trial, letters were presented sequentially for 1000 msec.
Participants were tasked with responding to each letter
by identifying whether the letter was presented n trials
back. The task consisted of three conditions—0-, 2-, E
3-back load. The n-back consisted of 320 trials. The 0-back
load condition consisted of 160 trials (targets: 128;
1146
Journal of Cognitive Neuroscience
Volume 35, Numero 7
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
C
N
_
UN
_
0
2
0
0
1
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
nontargets: 32), E 2- and 3-back conditions consisted of
80 trials each (targets: 52; nontargets: 16; lures: 12. For the
0-back load condition, participants were asked to identify
the letter “X” as a “target” (left button press) when it
appeared on the screen, and respond to any other letters
as nontargets (right button press). On 2- and 3-back
conditions, memory load was manipulated by asking
participants to respond to a letter based on whether the
letter presented n-trials prior. For instance, on a 2-back
load condition, a “target” (cioè., the correct response) is a
letter that was presented two trials prior, whereas a “non-
target” (cioè., incorrect response) would be a letter that was
not presented two trials back. Inoltre, 2- and 3-back
conditions included lure trials. Lure trials are those in
which a familiar letter is presented an incorrect number
of trials back. Lure trials add an additional “load” complex-
ità, as they require participants to not only remember the
sequence of letters that were presented prior, but also
require them to inhibit a prepotent response to seeing a
previously presented letter. The analyses included RTs for
correct responses only. We excluded trials with RTs that
were below 200, and we excluded observations in which
participants had an accuracy of less than 30% across all
trial types.
COMT and Estradiol Analysis
As part of the larger investigation, participants provided
daily assays of 1.8 mL of saliva using the passive drool
method across the full length of their menstrual cycle
within 30 min of waking. Participants were asked to keep
completed samples in their own personal freezer. During
in-person lab visits, participants provided their saliva sam-
ples which were then transferred to a -80F degree freezer.
All the samples were sent to Salimetrics LLC (State College,
PAPÀ) to assay estradiol levels. On a separate occasion, one
saliva sample for each participant was shipped to CD
Genomics (Shirley, NY ) to extract COMT Val158/ Met
(rs4680) polymorphisms using SNaPshot Multiplex
System for SNP Genotyping. All participants’ observed
genotype frequencies were in Hardy-Weinberg equilib-
rium (χ2= .53, df = 1 ns), indicating no significant
difference from the expected frequencies. Of the 139 par-
ticipants, 74 participants were homozygous allele carriers
(33 Met/Met, 41 Val/ Val).
Procedure and Data Analysis
As part of the larger investigation, participants provided
saliva samples daily to capture changes in hormones
across 35 days to capture the full length of the menstrual
cycle. Participants also completed four in-person labora-
tory visits that were meant to correspond with different
phases of their menstrual cycle to complete the n-back
task. In the analyses reported below, estradiol levels from
the four laboratory visits were included in the analyses.
The analyses were completed using the “lme4” package
(Bates, Mächler, Bolker, & Walker, 2014) in R Version
3.5.1. To examine whether changes in estradiol levels
moderated the association between COMT and perfor-
mance, estradiol was within-person centered. To do this,
a mean was computed for each participant and subtracted
from each of their own observations (Hoffman & Stawski,
2009). We also examined the effect of average levels of
estradiol between people, by computing a mean for each
participant, which was used to compute an overall mean
(cioè., mean of means). The overall mean was subtracted
from each participant’s mean value. This approach pro-
vides an estimate of between-persons average differences
in estradiol levels. In the final models, COMT was included
as an effects-coded predictor, estradiol levels within-
person centered and between-persons centered as main
effects, as well as a COMT × Within-Person Estradiol and
COMT × Between-Persons Estradiol interactions. We did
not include the three-way interaction. All models also
included time (up to four laboratory visits) as an effect-
coded predictor to control for practice effects, and 0-back
performance to control for their accuracy on the task. IL
final models did not include random slopes and only
included a random intercept, which calculated a mean
score (either RT or accuracy) for each participant. A
break down effects, we dummy-coded COMT. Because
we had a precise directional hypothesis for Met and Val
carriers, when breaking down interactions, we also report
the p values for a one-tailed simple slope test for the effect
of interest. For all models, assumptions were examined,
and Cook’s distance was computed to assess for leverage.
To determine whether our sample size was adequate to
answer the question of interest, we computed the effect
size for the behavioral effects reported in Jacobs and
D’Esposito (2011), which revealed a large effect for the
behavioral findings on 2-back lures (η2 = .19). Using
G*Power, we determined that for models examining a
COMT × Within-Person Centered Estradiol interaction,
we were powered to detect small effects of RTs and accu-
p’s ranging
racy on 2- and 3-back lure conditions (η2
between .02 E .03). For models examining a COMT ×
Between-Persons Estradiol interaction, we were powered
to detect small-to-medium effects of RTs and accuracy on
p’s ranging between .05
2- and 3-back load conditions (η2
E .07). We proceeded with the knowledge that we were
able to detect a small COMT × Estradiol interaction with
this sample size, which is more than adequate for the size
of the effect reported in Jacobs and D’Esposito (2011).
Below, partial eta squared η2
p is reported to estimate effect
size with .05 representing a small effect, .1 a medium
effect, E .2 a large effect (Cohen & Taylor, 1973).
RESULTS
There was no difference in average estradiol levels
between Met (M = 1.29 pg/mL, SE = .09) and Val (M =
1.51 pg/mL, SE = .08) carriers ( p = ns).
Louis et al.
1147
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
C
N
_
UN
_
0
2
0
0
1
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
C
N
_
UN
_
0
2
0
0
1
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
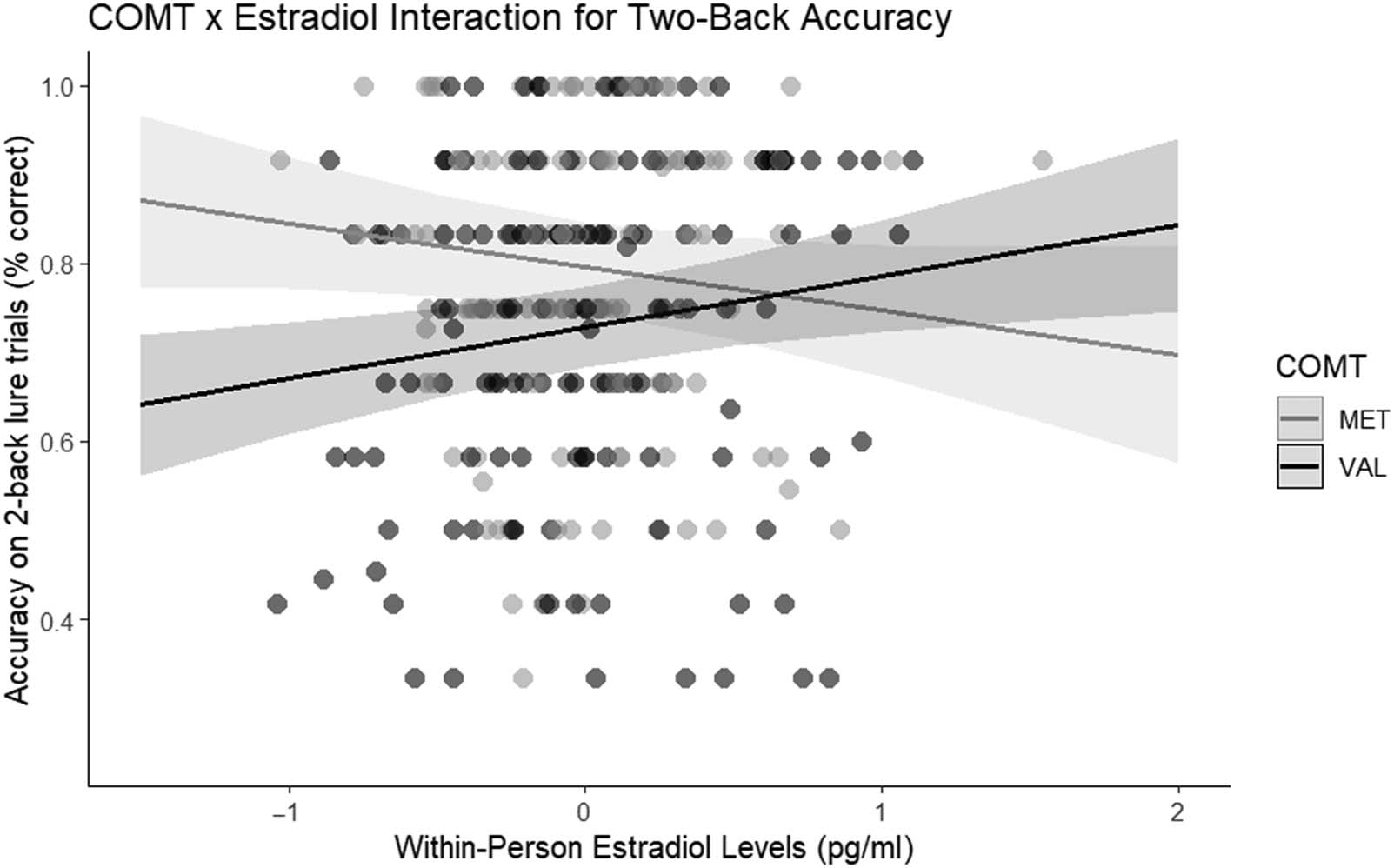
Figura 2. Line graph demonstrating the interaction between COMT and estradiol, such that Val carriers have a positive association between estradiol
and accuracy and Met carriers have a negative association. The shaded region around the fitted line depicts the 95% confidence interval.
As previously reported with this sample (Louis et al.,
2021), participants’ behavioral results replicated those
found in Jacobs and D’Esposito (2011) for load and trial
type manipulations. Load effects on RT (η2
p = .28) revealed
that participants were significantly faster on 0-back than all
other load conditions ( ps < .001), whereas 2- and 3-back
were not significantly different from each other ( p = .62).
Load effects for accuracy (η2
p = .16) demonstrated that
participants were significantly more accurate on 0-back
than other load conditions ( ps < .001). In addition, partic-
ipants were more accurate on 2-back than they were on
3-back trials ( p < .001). Results for trial type (η2
p = .29)
revealed participants were slower on lures than nontargets
and targets ( ps < .001). In addition, participants responded
more slowly to targets than nontargets ( p < .001). Similarly,
accuracy was significantly different across trial types (η2
p =
.44). Participants were significantly less accurate on lures
than nontargets and targets ( ps < .001). They were also sig-
nificantly less accurate on targets compared with nontargets
( p < .001). In summary, participants responded more
slowly and were less accurate on lure trials, replicating that
this task condition may have been more cognitively
demanding. Therefore, the analyses below solely focus on
lure trials.
Accuracy
The expected direction of COMT effects emerged ( p =
.04, η2
p = .06) such that Val carriers (M = .73, SE = .02)
were less accurate than Met carriers on 2-back lure trials
(M = .80, SE = .03). Importantly, and consistent with
hypotheses, there was a significant COMT × Within-
Person Estradiol Level interaction on 2-back lures ( p =
.003, η2
p = .05).2 This was probed further by examining
the effect of estradiol for Met and Val carriers separately.
For Val carriers, higher estradiol levels predicted higher
accuracy (b = .06, two-tailed p = .01; one-tailed p =
.005; see Figure 2). For Met carriers, on the other hand,
the effect was in the opposite direction (b = −.05, two-
tailed p = .08; one-tailed p = .04), similar to the findings
in Jacobs and D’Esposito (2011). Importantly, there was
no effect of between-centered levels of estradiol ( p =
.37) nor did it interact with COMT ( p = .29). For the
model examining these effects for three back lures, no
effects reached significance (see Table 1).
Reaction Time
For two back lures, there were no significant effects of
estradiol levels, COMT, or interactions (all ps > .14). How-
ever, there was a significant effect of RT on 0-back trials
(b = .97, P < .001), indicating a positive association
between RTs on 0-back trials and 2-back lures. For 3-back
lures, there was a significant effect of within-person estra-
diol levels (b = −40.94, p = .02), revealing that higher
within-person estradiol levels predicted faster RTs. The
effect of 0-back RTs remained for 3-back lures (b = 1.17,
p < .001). However, COMT did not interact with within-
person or between-persons estradiol levels to predict
3-back lure-RTs (both ps > .19).
1148
Journal of Cognitive Neuroscience
Volume 35, Numero 7
Tavolo 1. Model Results Examining the Effect of COMT, Estradiol Levels (within- and between-centered), and their Interaction on
2- and 3-back Lure Accuracy
Dependent Variable
Fixed Effect
Estimate
Standard Error
Two-back accuracy
–
Intercept
Estradiol levels (within-centered)
Estradiol levels (between-centered)
COMT
COMT × Estradiol Levels
(within-centered)
COMT × Estradiol Levels
(between-centered)
Zero-back accuracy (within-centered)
–
.73
.06
−.03
.07
−.11
.07
−.95
–
.02
.02
.04
.03
.03
.07
.51
Variance Components
Variance
Standard Deviation
Intercept
Residual
.02
.01
.13
.12
Dependent Variable
Fixed Effect
Estimate
Standard Error
Three-back accuracy
–
Intercept
Estradiol levels (within-centered)
Estradiol levels (between-centered)
COMT
COMT × Estradiol Levels
(within-centered)
COMT × Estradiol Levels
(between-centered)
Zero-back accuracy (within-centered)
–
.66
.04
−.06
.02
−.05
.08
−.54
–
.02
.03
.03
.03
.04
.06
.61
T
–
32.12
2.64
−.90
2.05
−3.03
1.06
−1.87
–
T
–
33.52
1.36
−1.94
.52
−1.14
1.43
−.89
P (two-tailed)
–
.000*
.01
.37
.04*
.003*
.29
.06
–
P (two-tailed)
–
.000*
.18
.06
.60
.26
.16
.37
Variance Components
Variance
Standard Deviation
–
–
Intercept
Residual
.009
.02
.10
.14
COMT was included as a dummy-coded variable, and Val is the base in the models presented in the table. Laboratory visits (cioè., time) were also
included in this model as an effects-coded predictor to control for practice effects ( P < .001).
DISCUSSION
The main aim of the current study was to determine
whether the behavioral findings from Jacobs and
D’Esposito (2011), that estradiol moderates the effect of
COMT on working memory performance, replicate in
another young adult sample. The original study tracked
women at two timepoints, whereas our study design con-
sisted of up to four repeated assessments across the
menstrual cycle that allowed for testing these effects with
more statistical power, investigating the predictive value of
estradiol by using it as a continuous predictor, and examining
both within- and between-persons differences of estradiol on
behavioral performance. Our results replicated Jacobs and
D’Esposito (2011) such that on 2-back lure trials, Val carriers
had enhanced performance when endogenous estradiol
levels were elevated. We found an effect in the opposite
direction for Met carriers, as hypothesized. Furthermore,
Louis et al.
1149
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
c
n
_
a
_
0
2
0
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
consistent with the original investigation, we did not find this
moderation effect at higher loads. These findings add to addi-
tional reports of Cycle × COMT interactions on behavior
(Wu et al., 2019; Smith, Sierra, Oppler, & Boettiger, 2014).
These findings strengthen the case that rhythmic
changes in sex hormone production within a person over
time shape cognitive functions that are sensitive to dopa-
minergic signaling in the pFC. In line with the signal-to-
noise hypothesis, DA plays a critical role in “sculpting”
mental representations in pFC and dampening noise or
distraction (Arnsten et al., 2015). Val/ Val individuals typi-
cally show enhanced COMT activity and reduced prefron-
tal DA relative to Met homozygotes. Some of estradiol’s
“pro-dopaminergic” effects likely arise via the hormone’s
ability to downregulate COMT activity. At the behavioral
level, across both studies, Val carriers exhibited a relative
boost in working memory performance when endoge-
nous estradiol levels are elevated, perhaps because of
changes in their ability to gate distractions. Similarly, at
the neural level, Jacobs and D’Esposito found that individ-
uals with low pFC DA levels (indexed by high COMT
enzyme activity) displayed exaggerated working
memory-related BOLD responses in the dorsolateral
pFC. Individuals with elevated pFC DA had reduced work-
ing memory-related pFC activity sustained across task
blocks, a putative marker of heightened cortical efficiency
(Green, Kraemer, DeYoung, Fossella, & Gray, 2013; Gray,
Chabris, & Braver, 2003). This pattern of results is consis-
tent with the broader COMT literature, in which Val car-
riers consistently show exaggerated task-evoked BOLD
responses on 2-back conditions relative to Met carriers
(Egan et al., 2001). The specificity of this effect to moder-
ate working memory loads (e.g., 2-back conditions) sug-
gests that there might be a particular context in which
estradiol’s moderating effect can be seen. When cognitive
demands exceed this level (e.g., 3-back load), participants
may experience additional task-induced DA release that
results in a further shift along the inverted-U (see
Figure 2). Our findings suggest that estradiol’s modulating
role may depend on task parameters, such as cognitive
load. The heightened cognitive demand on the 3-back
condition may be similar to an increase in acute stress.
Therefore, heightened estradiol and stress may increase
DA signaling on 3-back conditions, pushing individuals
further along the “inverted U.”
On the other hand, we did not find that between-
persons differences in estradiol influenced the COMT-
working memory performance association on 2-back lure
trials. Few studies have investigated the differential effects
of within- versus between-persons effects of hormones on
cognition. It is unknown whether individuals with higher
average levels of estradiol across the menstrual cycle also
exhibit elevated dopaminergic tone. Our test of between-
persons estradiol levels and COMT is a between-subjects
test, examining how individuals may perform in compari-
son to each other across the length of the cycle. Perhaps
regardless of average estradiol levels, individuals adapt to
task demands in ways that result in comparable levels of
performance. Although our findings did not examine
imaging data, it would be useful for future studies to exam-
ine whether there are COMT × between-estradiol interac-
tive effects on pFC activity. Importantly, our findings imply
that relative increases in estradiol may acutely alter perfor-
mance because of changes in dopaminergic action. This
has important implications for future study designs, disag-
gregating within- and between-persons estrogenic effects.
In contrast to the original findings on 3-back lure trials,
however, we found that within-person increases in estra-
diol levels negatively predicted RT overall, such that partic-
ipants responded more quickly when estradiol levels were
higher. The emergence of this finding could be because of
differences in our study design and analytic approach.
Importantly, this finding also coincides with previous
studies demonstrating that estradiol may lead to faster
RTs (Amunts, Camilleri, Eickhoff, Heim, & Weis, 2020;
Ho, Gilger, & Brink, 1986), and work indicating that pro-
cessing speed declines during perimenopause (Greendale
et al., 2009). Such a finding does not exclude a role for DA
but suggests it may not depend on where people start at
baseline (i.e., COMT). Typically, when individuals have
improved processing speed without changes in accuracy,
it is interpreted as enhanced processing efficiency. Indeed,
findings suggest that estradiol may facilitate faster perfor-
mance in high load contexts, perhaps because of its DA-
agonist effects that lead to efficient neural activation and
synaptic transmission (Del Río et al., 2018; Barth, Villringer,
& Sacher, 2015; Jacobs & D’Esposito, 2011). Importantly
however, individuals still performed less favorably on
3-back lures than 2-back lure conditions. Therefore,
although participants may be more efficient during this
context, it is not resulting in better performance overall.
Our study should also be interpreted with some limita-
tions. Although our behavioral study had a larger sample
than the initial investigation, more efforts to replicate in
larger samples would be of great utility. In addition,
whereas Jacobs and D’Esposito (2011) found a large effect
for 2-back accuracy (η2 =.19), the effect was small in our
study. The discrepancy in effect size may be because of
methodological differences in our study design, including
using saliva (instead of serum), and having more repeated
measures. We also conducted a multilevel model to exam-
ine our interactions of interest, which accounts for multi-
ple sources of variance. In addition, we used estradiol as a
continuous predictor, in contrast to the 2011 investigation
that compared group differences in accuracy scores.
Therefore, methodological and analytic differences could
explain the discrepancy in effect sizes.
Replication of the findings in Jacobs and D’Esposito
(2011) offers additional support for the importance of con-
sidering estradiol’s influence on dopamine-dependent
cognitive functions. Indeed, recent work has called for a
more holistic understanding of cognition by including
the sex hormone milieu (Taylor, Pritschet, & Jacobs,
2021; Beltz & Moser, 2020). Our study supports the notion
1150
Journal of Cognitive Neuroscience
Volume 35, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
c
n
_
a
_
0
2
0
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
that empirical efforts probing DA’s influence on cognition
should include a role for estradiol (Colzato & Hommel,
2014). Importantly, our results indicate that the role of
estradiol may depend on task conditions, including overall
difficulty, which has implications for how we broadly
understand estradiol’s role in cognitive performance.
The current findings, therefore, have important implica-
tions for the generalizability of our knowledge of working
memory, in that we must consider for whom, and in what
contexts, certain effects may or may not be seen.
Acknowledgments
The authors would like to thank Courtney Callahan, Megan
Wright, Teona Velehorschi, and Alycia Winters for their invalu-
able role in the administration of this study, as well as all the
participants for their vital contribution to this research.
Reprint requests should be sent to Courtney C. Louis, Depart-
ment of Psychology, Michigan State University, 157F Psychology
Building, East Lansing, Michigan 48824, United States, or via
e-mail: louiscou@msu.edu.
Data Availability Statement
Data and code can be made available upon request.
Author Contributions
Courtney C. Louis: Data curation; Formal analysis;
Funding acquisition; Investigation; Project administration;
Visualization; Writing—Original draft; Writing—Review &
editing. Emily Jacobs: Conceptualization; Funding acqui-
sition; Methodology; Supervision; Writing—Review &
editing. Mark D’Esposito: Conceptualization; Funding
acquisition; Methodology; Supervision; Writing—Review
& editing. Jason Moser: Conceptualization; Formal analy-
sis; Funding acquisition; Investigation; Methodology;
Project administration; Resources; Software; Supervision;
Visualization; Writing—Review & editing.
Funding Information
National Institute of Mental Health (https://dx.doi.org/10
.13039/100000025), grant numbers: 1R01MH108511 and
1F31MH125604-01.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN ) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. The
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .261; W/M = .348;
M/ W = .087; W/ W = .304.
Notes
It is critical to make the distinction between sex and gen-
1.
der; neither of which are binary, nor do they have to overlap.
“Female” is used in this text to refer those who are assigned
female sex at birth. Importantly, however, the term “female”
for those who are assigned female at birth can be elusive, as
sex can refer to many things, including genitals, hormones,
chromosol makeup, among other things. Therefore, we would
like to be clear that in this text, we solely use “female” to refer
to those who experience menstrual cycles. Furthermore, we
use the term “women” to refer to those who identify as such.
We do not intent to extend “female” to signify any other biolog-
ical, social, or identity related factors in this text.
2. Because Jacobs and D’Esposito (2011) found and reported
an interaction between Met and Val homozygous carriers, our
main aims only focused on these two groups. However, we also
tested a model with all three groups (41 Val/ Val, 33 Met/Met,
and 65 Val/Met). Our results remained unchanged. There was
a significant interaction between COMT × Estradiol ( p =
.02). The effects revealed that there was a significant effect of
estradiol for Val carriers (b = .06, two-tailed p = .01, one-tailed
p = .005), trending in the opposite direction for Met carriers
(b = −.04, two-tailed p = .15, one-tailed p = .075) and no
effect for Val/Met carriers ( p = .27).
REFERENCES
Amunts, J., Camilleri, J. A., Eickhoff, S. B., Heim, S., & Weis, S.
(2020). Executive functions predict verbal fluency scores in
healthy participants. Scientific Reports, 10, 11141. https://doi
.org/10.1038/s41598-020-65525-9, PubMed: 32636406
Arnsten, A. F. T. (2015). Stress weakens prefrontal networks:
Molecular insults to higher cognition. Nature Neuroscience,
18, 1376–1385. https://doi.org/10.1038/nn.4087, PubMed:
26404712
Arnsten, A. F., Wang, M., & Paspalas, C. D. (2015). Dopamine’s
actions in primate prefrontal cortex: Challenges for treating
cognitive disorders. Pharmacological Reviews, 67, 681–696.
https://doi.org/10.1124/pr.115.010512, PubMed: 26106146
Barth, C., Villringer, A., & Sacher, J. (2015). Sex hormones affect
neurotransmitters and shape the adult female brain during
hormonal transition periods. Frontiers in Neuroscience, 9, 37.
https://doi.org/10.3389/fnins.2015.00037, PubMed: 25750611
Bates, D., Mächler, M., Bolker, B., & Walker, S. (2014). Fitting
linear mixed-effects models using lme4. arXiv:1406.5823.
https://doi.org/10.48550/arXiv.1406.5823
Becker, J. B. (1990). Estrogen rapidly potentiates amphetamine-
induced striatal dopamine release and rotational behavior
during microdialysis. Neuroscience Letters, 118, 169–171.
https://doi.org/10.1016/0304-3940(90)90618-J, PubMed:
2125712
Beltz, A. M., & Moser, J. S. (2020). Ovarian hormones: A long
overlooked but critical contributor to cognitive brain
structures and function. Annals of the New York Academy of
Sciences, 1464, 156–180. https://doi.org/10.1111/nyas.14255,
PubMed: 31639230
Louis et al.
1151
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
c
n
_
a
_
0
2
0
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bernal, A., & Paolieri, D. (2022). The influence of estradiol and
progesterone on neurocognition during three phases of the
menstrual cycle: Modulating factors. Behavioural Brain
Research, 417, 113593. https://doi.org/10.1016/j.bbr.2021
.113593, PubMed: 34560130
Bimonte, H. A., & Denenberg, V. H. (1999). Estradiol facilitates
performance as working memory load increases.
Psychoneuroendocrinology, 24, 161–173. https://doi.org/10
.1016/S0306-4530(98)00068-7, PubMed: 10101725
Björklund, A., & Dunnett, S. B. (2007). Dopamine neuron
systems in the brain: An update. Trends in Neurosciences,
30, 194–202. https://doi.org/10.1016/j.tins.2007.03.006,
PubMed: 17408759
Brozoski, T. J., Brown, R. M., Rosvold, H. E., & Goldman, P. S.
(1979). Cognitive deficit caused by regional depletion of
dopamine in prefrontal cortex of rhesus monkey. Science,
205, 929–932. https://doi.org/10.1126/science.112679,
PubMed: 112679
Cohen, E. R., & Taylor, B. N. (1973). The 1973 least-squares
adjustment of the fundamental constants. Journal of
Physical and Chemical Reference Data, 2, 663–734. https://
doi.org/10.1063/1.3253130
Colzato, L. S., & Hommel, B. (2014). Effects of estrogen on
higher-order cognitive functions in unstressed human
females may depend on individual variation in dopamine
baseline levels. Frontiers in Neuroscience, 8, 65. https://doi
.org/10.3389/fnins.2014.00065, PubMed: 24778605
Cools, R., & D’Esposito, M. (2011). Inverted-U–shaped dopamine
actions on human working memory and cognitive control.
Biological Psychiatry, 69, e113–e125. https://doi.org/10.1016
/j.biopsych.2011.03.028, PubMed: 21531388
Del Río, J. P., Alliende, M. I., Molina, N., Serrano, F. G., Molina,
S., & Vigil, P. (2018). Steroid hormones and their action in
women’s brains: The importance of hormonal balance.
Frontiers in Public Health, 6, 141. https://doi.org/10.3389
/fpubh.2018.00141, PubMed: 29876339
Dumas, J. A., Makarewicz, J. A., Bunn, J., Nickerson, J., &
McGee, E. (2018). Dopamine-dependent cognitive processes
after menopause: The relationship between COMT genotype,
estradiol, and working memory. Neurobiology of Aging, 72,
53–61. https://doi.org/10.1016/j.neurobiolaging.2018.08.009,
PubMed: 30212711
Egan, M. F., Goldberg, T. E., Kolachana, B. S., Callicott, J. H.,
Mazzanti, C. M., Straub, R. E., et al. (2001). Effect of COMT
Val108/158 met genotype on frontal lobe function and risk for
schizophrenia. Proceedings of the National Academy of
Sciences, U.S.A., 98, 6917–6922. https://doi.org/10.1073/pnas
.111134598, PubMed: 11381111
Elton, A., Smith, C. T., Parrish, M. H., & Boettiger, C. A. (2017).
COMT Val158Met polymorphism exerts sex-dependent
effects on fMRI measures of brain function. Frontiers in
Human Neuroscience, 11, 578. https://doi.org/10.3389
/fnhum.2017.00578, PubMed: 29270116
Gray, J. R., Chabris, C. F., & Braver, T. S. (2003). Neural
mechanisms of general fluid intelligence. Nature
Neuroscience, 6, 316–322. https://doi.org/10.1038/nn1014,
PubMed: 12592404
Green, A. E., Kraemer, D. J. M., DeYoung, C. G., Fossella, J. A.,
& Gray, J. R. (2013). A gene–brain–cognition pathway:
Prefrontal activity mediates the effect of COMT on cognitive
control and IQ. Cerebral Cortex, 23, 552–559. https://doi.org
/10.1093/cercor/bhs035, PubMed: 22368081
Greendale, G. A., Huang, M.-H., Wight, R. G., Seeman, T.,
Luetters, C., Avis, N. E., et al. (2009). Effects of the
menopause transition and hormone use on cognitive
performance in midlife women. Neurology, 72, 1850–1857.
https://doi.org/10.1212/WNL.0b013e3181a71193, PubMed:
19470968
Hampson, E. (2018). Estrogens, aging, and working memory.
Current Psychiatry Reports, 20, 109. https://doi.org/10.1007
/s11920-018-0972-1, PubMed: 30306352
Hampson, E., & Morley, E. E. (2013). Estradiol concentrations
and working memory performance in women of reproductive
age. Psychoneuroendocrinology, 38, 2897–2904. https://doi
.org/10.1016/j.psyneuen.2013.07.020, PubMed: 24011502
Hara, Y., Yuk, F., Puri, R., Janssen, W. G. M., Rapp, P. R., &
Morrison, J. H. (2014). Presynaptic mitochondrial morphology
in monkey prefrontal cortex correlates with working memory
and is improved with estrogen treatment. Proceedings of the
National Academy of Sciences, U.S.A., 111, 486–491. https://
doi.org/10.1073/pnas.1311310110, PubMed: 26791219
Hara, Y., Yuk, F., Puri, R., Janssen, W. G. M., Rapp, P. R., &
Morrison, J. H. (2016). Estrogen restores multisynaptic
boutons in the dorsolateral prefrontal cortex while
promoting working memory in aged rhesus monkeys.
Journal of Neuroscience, 36, 901–910. https://doi.org/10
.1523/JNEUROSCI.3480-13.2016, PubMed: 26791219
Hidalgo-Lopez, E., & Pletzer, B. (2017). Interactive effects of
dopamine baseline levels and cycle phase on executive
functions: The role of progesterone. Frontiers in Neuroscience,
11, 403. https://doi.org/10.3389/fnins.2017.00403, PubMed:
28751855
Ho, H. Z., Gilger, J. W., & Brink, T. M. (1986). Effects of
menstrual cycle on spatial information-processes. Perceptual
and Motor Skills, 63, 743–751. https://doi.org/10.2466/pms
.1986.63.2.743, PubMed: 3808858
Hoffman, L., & Stawski, R. S. (2009). Persons as contexts:
Evaluating between-person and within-person effects in
longitudinal analysis. Research in Human Development, 6,
97–120. https://doi.org/10.1080/15427600902911189
Jacobs, E., & D’Esposito, M. (2011). Estrogen shapes
dopamine-dependent cognitive processes: Implications for
women’s health. Journal of Neuroscience, 31, 5286–5293.
https://doi.org/10.1523/JNEUROSCI.6394-10.2011, PubMed:
21471363
Jacobs, E. G., & Goldstein, J. M. (2018). The middle-aged brain:
Biological sex and sex hormones shape memory circuitry.
Current Opinion in Behavioral Sciences, 23, 84–91. https://
doi.org/10.1016/j.cobeha.2018.03.009, PubMed: 30271832
Jiang, H., Xie, T., Ramsden, D. B., & Ho, S. L. (2003). Human
catechol-O-methyltransferase down-regulation by estradiol.
Neuropharmacology, 45, 1011–1018. https://doi.org/10.1016
/S0028-3908(03)00286-7, PubMed: 14573393
Kirchner, W. K. (1958). Age differences in short-term retention
of rapidly changing information. Journal of Experimental
Psychology, 55, 352–358. https://doi.org/10.1037/h0043688,
PubMed: 13539317
Kromrey, S. A., Czoty, P. W., & Nader, M. A. (2015). Relationship
between estradiol and progesterone concentrations and
cognitive performance in normally cycling female cynomolgus
monkeys. Hormones and Behavior, 72, 12–19. https://doi.org
/10.1016/j.yhbeh.2015.04.017, PubMed: 25921587
Louis, C. C., D’Esposito, M., & Moser, J. S. (2021). Investigating
interactive effects of worry and the catechol-o-
methyltransferase gene (COMT) on working memory
performance. Cognitive, Affective, & Behavioral
Neuroscience, 21, 1153–1163. https://doi.org/10.3758/s13415
-021-00922-9, PubMed: 34173216
Luine, V. N., Richards, S. T., Wu, V. Y., & Beck, K. D. (1998).
Estradiol enhances learning and memory in a spatial memory
task and effects levels of monoaminergic neurotransmitters.
Hormones and Behavior, 34, 149–162. https://doi.org/10
.1006/hbeh.1998.1473, PubMed: 9799625
Männistö, P. T., & Kaakkola, S. (1999). Catechol-O-
methyltransferase (COMT): Biochemistry, molecular biology,
pharmacology, and clinical efficacy of the new selective
1152
Journal of Cognitive Neuroscience
Volume 35, Number 7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
c
n
_
a
_
0
2
0
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
COMT inhibitors. Pharmacological Reviews, 51, 593–628.
PubMed: 10581325
Meyer-Lindenberg, A., Nichols, T., Callicott, J. H., Ding, J.,
Kolachana, B., Buckholtz, J., et al. (2006). Impact of complex
genetic variation in COMT on human brain function.
Molecular Psychiatry, 11, 867–797. https://doi.org/10.1038/sj
.mp.4001860, PubMed: 16786032
Mier, D., Kirsch, P., & Meyer-Lindenberg, A. (2010). Neural
substrates of pleiotropic action of genetic variation in COMT:
A meta-analysis. Molecular Psychiatry, 15, 918–927. https://
doi.org/10.1038/mp.2009.36, PubMed: 19417742
Papaleo, F., Sannino, S., Piras, F., & Spalletta, G. (2015). Sex-
dichotomous effects of functional COMT genetic variations on
cognitive functions disappear after menopause in both health
and schizophrenia. European Neuropsychopharmacology,
25, 2349–2363. https://doi.org/10.1016/j.euroneuro.2015.10
.005, PubMed: 26560201
Rosenberg, L., & Park, S. (2002). Verbal and spatial functions
across the menstrual cycle in healthy young women.
Psychoneuroendocrinology, 27, 835–841. https://doi.org/10
.1016/S0306-4530(01)00083-X, PubMed: 12183218
Shanmugan, S., & Epperson, C. N. (2014). Estrogen and the
prefrontal cortex: Towards a new understanding of estrogen’s
effects on executive functions in the menopause transition.
Human Brain Mapping, 35, 847–865. https://doi.org/10.1002
/hbm.22218, PubMed: 23238908
in the frontal cortex interact to affect target detection
under high working memory load. Journal of Cognitive
Neuroscience, 26, 395–407. https://doi.org/10.1162/jocn_a
_00501, PubMed: 24144248
Taylor, C. M., Furman, D. J., Berry, A. S., White, R. L., Jagust, W. J.,
D’Esposito, M., et al. (2022). Striatal dopamine synthesis and
cognitive flexibility differ between hormonal contraceptive
users and non-users. bioRxiv. https://doi.org/10.1101/2022.10
.20.513082
Taylor, C. M., Pritschet, L., & Jacobs, E. G. (2021). The scientific
body of knowledge—Whose body does it serve? A spotlight
on oral contraceptives and women’s health factors in
neuroimaging. Frontiers in Neuroendocrinology, 60, 100874.
https://doi.org/10.1016/j.yfrne.2020.100874, PubMed: 33002517
Tunbridge, E. M., Harrison, P. J., & Weinberger, D. R. (2006).
Catechol-o-methyltransferase, cognition, and psychosis:
Val158Met and beyond. Biological Psychiatry, 60, 141–151.
https://doi.org/10.1016/j.biopsych.2005.10.024, PubMed:
16476412
Tunbridge, E. M., Narajos, M., Harrison, C. H., Beresford, C.,
Cipriani, A., & Harrison, P. J. (2019). Which dopamine
polymorphisms are functional? Systematic review and
meta-analysis of COMT, DAT, DBH, DDC, DRD1–5, MAOA,
MAOB, TH, VMAT1, and VMAT2. Biological Psychiatry, 86,
608–620. https://doi.org/10.1016/j.biopsych.2019.05.014,
PubMed: 31303260
Shansky, R. M., Glavis-Bloom, C., Lerman, D., McRae, P.,
Williams, G. V., & Castner, S. A. (2006). Under the curve: Critical
Benson, C., Miller, K., et al. (2004). Estrogen mediates sex
differences in stress-induced prefrontal cortex dysfunction.
Molecular Psychiatry, 9, 531–538. https://doi.org/10.1038/sj
.mp.4001435, PubMed: 14569273
Shansky, R. M., & Lipps, J. (2013). Stress-induced cognitive
dysfunction: Hormone-neurotransmitter interactions in the
prefrontal cortex. Frontiers in Human Neuroscience, 7,
123. https://doi.org/10.3389/fnhum.2013.00123, PubMed:
23576971
Slifstein, M., Kolachana, B., Simpson, E. H., Tabares, P., Cheng,
B., Duvall, M., et al. (2008). COMT genotype predicts
cortical-limbic D1 receptor availability measured with
[11C]NNC112 and PET. Molecular Psychiatry, 13, 821–827.
https://doi.org/10.1038/mp.2008.19, PubMed: 18317466
Smith, C. T., Sierra, Y., Oppler, S. H., & Boettiger, C. A. (2014).
Ovarian cycle effects on immediate reward selection bias in
humans: A role for estradiol. Journal of Neuroscience, 34,
5468–5476. https://doi.org/10.1523/JNEUROSCI.0014-14
.2014, PubMed: 24741037
Smith, C. T., Swift-Scanlan, T., & Boettiger, C. A. (2014).
Genetic polymorphisms regulating dopamine signaling
issues for elucidating D1 receptor function in working
memory. Neuroscience, 139, 263–276. https://doi.org/10
.1016/j.neuroscience.2005.09.028, PubMed: 16310964
Wu, C., Ding, Y., Chen, B., Gao, Y., Wang, Q., Wu, Z., et al.
(2019). Both Val158Met polymorphism of catechol-O-
methyltransferase gene and menstrual cycle affect prepulse
inhibition but not attentional modulation of prepulse
inhibition in younger-adult females. Neuroscience, 404,
396–406. https://doi.org/10.1016/j.neuroscience.2019.02.001,
PubMed: 30742958
Xiao, L., & Becker, J. B. (1994). Quantitative microdialysis
determination of extracellular striatal dopamine concentration
in male and female rats: Effects of estrous cycle and
gonadectomy. Neuroscience Letters, 180, 155–158. https://doi
.org/10.1016/0304-3940(94)90510-X, PubMed: 7700570
Yavich, L., Forsberg, M. M., Karayiorgou, M., Gogos, J. A., &
Männistö, P. T. (2007). Site-specific role of catechol-O-
methyltransferase in dopamine overflow within prefrontal
cortex and dorsal striatum. Journal of Neuroscience, 27,
10196–10209. https://doi.org/10.1523/JNEUROSCI.0665-07
.2007, PubMed: 17881525
Louis et al.
1153
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
5
7
1
1
4
4
2
1
2
2
8
7
3
/
/
j
o
c
n
_
a
_
0
2
0
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3