Emotional Perception: Divergence of Early and Late
Event-related Potential Modulation
Andrew H. Farkas and Dean Sabatinelli
Astratto
■ The early posterior negativity (EPN) is a mid-latency ERP
component that is reliably enhanced by emotional cues, con
a deflection beginning between 150 E 200 msec after stimu-
lus onset. The brief, bilateral occipital EPN is followed by the
centroparietal late positive potential (LPP), a long duration
slow-wave that is strongly associated with emotional arousal
ratings of scenes. A recent study suggests that the EPN is par-
ticularly sensitive to human bodies in scenes, independent of
emotional intensity. Here, we directly investigate the influence
of human body features on EPN modulation, using emotional
and neutral scenes depicting people across a range of body
exposures and orientations, in addition to scenes of pleasant,
neutro, and unpleasant animals. The results demonstrate that
the EPN is quite sensitive to human body features and weakly
related to arousal ratings, whereas the LPP is strongly modu-
lated by scenes that receive high arousal ratings. Based on these
results and relevant work on body-specific visual perception, we
speculate that modulation of the EPN may strongly reflect the
early detection of human bodies, which serves as a predictor of
emotional significance, whereas LPP modulation is more closely
associated with the extended elaborative processing of scenes
that are explicitly judged to be emotionally arousing. ■
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
J
o
C
N
_
UN
_
0
1
9
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
INTRODUCTION
The collection and interpretation of scene-evoked cortical
potentials is a powerful paradigm in the effort to under-
stand the mental processes supporting the perception of
emotional cues. Two of the most well-studied ERPs asso-
ciated with emotional processing are the early posterior
negativity (EPN) and the late positive potential (LPP).
The amplitudes of these potentials have been shown to
be enhanced by emotionally arousing, relative to neutral
content (Olofsson, Nordin, Sequeira, & Polich, 2008;
Schupp, Stockburger, et al., 2006; Pastor et al., 2008). Questo
arousal effect can be reliably found during naturalistic
scene perception, and it is also evident (although to a
reduced extent) during the perception of emotional faces
(Langeslag, Gootjes, & van Strien, 2018; Yoon, Shim, Kim,
& Lee, 2016; Herbert, Sfärlea, & Blumenthal, 2013;
Jaworska et al., 2012; Schupp, Öhman, et al., 2004), hand
gestures (Flaisch, Schupp, Renner, & Junghöfer, 2009),
and words (Schindler & Kissler, 2016; Kissler & Herbert,
2013; Herbert, Junghöfer, & Kissler, 2008; Kissler, Herbert,
Peyk, & Junghöfer, 2007).
Despite similarities in the pattern of modulation, IL
EPN and LPP represent different but related aspects of
emotional perception. The EPN and LPP are temporally
and spatially distinct, with the EPN appearing as a neg-
ative voltage deflection over left and right lateral occip-
ital sensors from 150 A 300 msec after scene onset
University of Georgia, Athens
© 2023 Istituto di Tecnologia del Massachussetts
(Schupp, Junghöfer, Weike, & Hamm, 2004; Junghöfer,
Bradley, Elbert, & Lang, 2001) whereas the LPP is a
slow-wave positivity over central-parietal electrodes with
a peak approximately 400–900 msec after scene onset
(Schuppetal.,2007;Cuthbert,Schupp,Bradley,Birbaumer,
& Lang, 2000). Studies that have attempted to localize
the neural sources of these ERPs find that the EPN is likely
driven by lateral occipital extrastriate activity ( Junghöfer
et al., 2006; Schupp, Stockburger, et al., 2006) whereas
the LPP may represent a combination of activity from
multiple dorsal and ventral visual cortical regions, COME
well as anterior cortical sources (Sabatinelli, Keil, Frank,
& Lang, 2013; Liu, Hairston, Schrier, & Fan, 2011; Moratti,
Saugar, & Strange, 2011). Taken together, these ERPs
likely represent different stages involved in the discrim-
ination and perception of emotional cues, with the EPN
possibly reflecting the initial effects of emotional con-
tent in the secondary visual cortex, whereas the LPP
indexes the activity of widespread networks of activity
associated with the conscious experience of emotional
arousal.
Some studies have identified scene factors that differen-
tially modulate the EPN and LPP. Bradley, Hamby, Löw,
and Lang (2007) used principal components analysis to
identify clusters of electrodes that distinguished simple
(figure-ground) from complex emotional scenes, locating
a posterior occipital region that overlaps with the scalp
location and latency (150–250 msec) of the EPN. This clus-
ter showed enhanced voltage negativity during simple
scene perception, and no sensitivity to emotional content.
Journal of Cognitive Neuroscience 35:6, pag. 941–956
https://doi.org/10.1162/jocn_a_01984
A centroparietal cluster of sensors showed strong emotional
modulation from 400 A 700 msec, equivalent with the LPP,
and showed no sensitivity to scene complexity. This impor-
tant article obligated all future study of the EPN to employ
scenes that are balanced for complexity across emotional
and neutral content. Particularly relevant for the current
research question, Löw, Bradley, and Lang (2013) manipu-
lated emotional scene complexity and the inclusion of
humans in scenes and assessed the impact of these factors
on the EPN, as recorded using rapid serial visual presenta-
tion of scenes at 3 Hz. This study identified the largest
EPN modulation for emotional people depicted in simple,
figure-ground scenes, followed by the EPN in response
to neutral people in figure-ground scenes, followed by the
EPN in response to emotional people in complex
scenes. Così, the presence of clearly identifiable people in
scenes was more meaningful to EPN modulation than
whether those people were engaged in an emotional act.
Further evidence for the impact of clearly identifiable
people on EPN modulation, a recent study (Farkas,
Oliver, & Sabatinelli, 2020) found that scenes of upright,
unclothed people (nudists) evoke a larger EPN than do
scenes of erotic couples, despite garnering weak arousal
ratings. Despite enhancing the EPN, nudist scenes led to
reduced LPP amplitudes relative to that evoked by erotic
scenes. This unexpected finding necessitated an
experiment that more explicitly tests the effects of human
body exposure and orientation on EPN modulation.
In the current experiment, we aimed to replicate the
sensitivity of the EPN to nudist, compared with erotic
scenes (Farkas et al., 2020), while adding new scene cate-
sanguinose, additional measures of scene features (SFs), E
targeted analyses to parcel the potential relationships
between these factors and modulation of the EPN and
LPP. We consider body exposure to be a comparatively
subordinate SF relative to emotional scene content cate-
sanguinose. We employed the nudist and erotic scenes from
our previous experiment, and added seven additional
scene categories with varied levels of body exposure
across emotional and neutral contents. We included
scenes depicting pleasant, neutro, and unpleasant animals
to provide a nonbody comparison. Our intention is to
begin to disentangle the specific effects of body exposure
from higher-level emotional features of scene perception
(pleasantness, arousal) and subordinate features (animal/
human, clothed/unclothed, upright/supine).
We quantified characteristics of the scenes using
self-report and objective image values. These SFs included
self-reported valence, arousal, body exposure, and body
orientation, as well as digitally derived aspects of each
scene, including the number of pixels containing
unclothed body parts, energy in low and high spatial
frequency bands, and Shannon’s entropy. The study
employed two parallel analyses to understand how these
SFs are associated with the modulation of the EPN and LPP.
The first was a traditional analysis in which each partici-
pant’s average ERP across the 15 exemplars of each scene
category was compared. To evaluate the contribution of
SFs more precisely, a by-scene analysis assessed which
SFs were associated with EPN and LPP amplitudes, irrespec-
tive of scene category. In the by-scene analyses, we used
multiple regression and four variable-selection or regulari-
zation ( VSR) metodi. Our research question concerned
the relative impact of multiple scenes features, many of
which have been shown to independently influence ERP
magnitude in prior studies. To simplify data interpretation,
a multi-analysis approach was chosen to define which var-
iables had the strongest association with ERP modulation
and which variables and models would be the most stable
and predictive in a new data set. We hypothesized that
scenes containing people would evoke the largest EPN
(Löw et al., 2013), and that erotic ( Weinberg & Hajcak,
2010; Schupp et al., 2007) and nudist scenes (Farkas
et al., 2020) will elicit the greatest EPN amplitudes. Anche noi
expect that scenes receiving elevated ratings of arousal will
be associated with enhanced EPN modulation (Frank &
Sabatinelli, 2019; Junghöfer et al., 2001). When analyzed
by scene exemplar, we hypothesize that scenes containing
upright, exposed bodies would better predict the EPN than
the LPP, which should have a stronger association with
average self-reported arousal for each scene (Thigpen,
Keil, & Freund, 2018; Cuthbert et al., 2000).
METHODS
Participants
Fifty-seven participants were recruited from the University
of Georgia student body and compensated with course
credit. All participants gave informed consent after receiv-
ing a description of the study approved by the University
of Georgia human subjects institutional review board.
Seven participants were excluded from the final analysis
after ERP preprocessing revealed excessive artifact leading
to the loss of more than 50% of the trials from any one
scene category. The remaining 50 participants were
between the ages of 18 E 26 years (M = 19.7 years,
SD = 1.40 years), con 28 self-reporting as female (self-
reported ethnicity: Asian 7.1%, Black 7.1%, White
85.7%) E 22 as male (Asian 9.1%, Black 13.6%, Hispanic
4.6%, Hispanic & White 4.6%, Multiracial 4.6%, White
63.6%). Participants identified as right-handed and
reported no history of mental illness. The sample size
was predetermined based on previous work using most
of the same scenes (Farkas et al., 2020).
A second online sample was recruited through Ama-
zon’s Mechanical Turk (Mturk) to rate the extent of body
exposure and body orientation of the scene stimuli. Partic-
ipants were compensated $1.50 to provide these ratings.
Participants were presented with an informed consent
approved by the University of Georgia institutional review
board. Mturk participants (n = 79) had to live in the
United States and be between the ages of 18 E 26 years.
Seven online participants consistently used the same
942
Journal of Cognitive Neuroscience
Volume 35, Numero 6
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
J
o
C
N
_
UN
_
0
1
9
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
number for every scene rating and were excluded from the
final analysis. Two participants appeared to give random
ratings and were also excluded. After exclusions, there
were 70 participants in the final online ratings sample.
Naturalistic Scenes
Nine categories of 135 naturalistic scenes were selected,
based on valence and arousal ratings from previous studies
in our laboratory. A minority of scenes were taken from
the International Affective Picture System (IAPS; Lang,
Bradley, & Cuthbert, 1997) whereas the remainder were
gathered through uncopyrighted sources and are consis-
tent in nature and composition with the IAPS. Scenes were
presented at an 800 × 600 pixel resolution at 90% JPEG
qualità (https:// JPEG.org). Luminance and complexity
were selected to be equivalent between categories, COME
measured by independent t tests with p values greater
di .20. Scenes that were outliers in luminance and
complexity were mildly edited with GNU Image Manipula-
tion Program (https://www.gimp.org/). Complexity was
measured by the file size of the JPEG images at 90%
qualità, which has been reported as a suitable measure
of complexity in human perception (Donderi, 2006). If
a scene file was too large at 90% qualità, UN 1 × 1 pixel
Gaussian blur was used, which has a negligible effect on
perceived sharpness of the content.
The nine scene categories included erotica, pleasant
animals, victorious athletes, nudists, neutral animals,
neutral people (fully clothed), threatening animals, threat-
ening people, and mutilations. An example scene for each
category can be seen in Figure 1. Erotic scenes depicted an
attractive male–female couple engaged in consensual
intercourse. The pleasant animal category contained
scenes of baby ducks, puppies, and kittens. Victorious
scenes depicted athletes in expressive moments of
success. Nudist scenes contained unclothed couples of
various ages walking on beaches or parks, not engaged
in sexual activity. The neutral animal category depicted
adult animals such as cows, squirrels, and chickens. IL
neutral people category depicted clothed people engaged
in normal activities like riding in buses or talking to each
other. The threatening people category featured one or
more persons with aggressive body positions and facial
expressions, often brandishing weapons. Threatening
animal scenes contained predators such as wolves, jaguars,
and sharks aggressively posturing or attacking. Mutilation
scenes displayed graphic injuries of open wounds or disfig-
ured body parts. The scenes were chosen to roughly equate
the amount of body exposure within each valence subset.
Erotica, nudists, and mutilations scenes often have uncov-
ered skin making up a large portion of the scene as com-
pared with the victorious athletes, neutral people, E
threatening people categories that contain clothed individ-
uals. Finalmente, the three animal categories allowed for tests of
emotional ERP modulation effects independent of any
human bodies being present in the scene.
Procedure
After providing informed consent, participants were
brought into an electrically shielded chamber for EEG
cap fitting, which lasted about 10 min. IL 135 scenes
were then presented to each participant for 2 sec each,
with an intertrial interval between 3.5 E 5.5 sec. Partici-
pants were asked to keep their eyes fixated on a small
red dot in the middle of the scene. The complete set of
135 scenes were organized into two pseudorandom sets.
Half of the participants passively viewed the first set,
whereas the other half viewed the second set. To keep
the nine categories of content evenly spread throughout
il set, every 18 scenes had to feature two scenes from
Figura 1. Example scenes from
each category. Starting from
the top and moving from left
to right, displayed here are
example scenes from the
categories of erotica, pleasant
animals, victorious athletes,
nudists, neutral animals, neutro
people, mutilations, threatening
animals, and threatening
people.
Farkas and Sabatinelli
943
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
J
o
C
N
_
UN
_
0
1
9
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
each category. There were no statistically significant cate-
gory differences between participants that saw Set 1 versus
Set 2. Scenes were presented on a 32-in. LCD monitor
that occupied 31° of the horizontal visual field of view.
Scene Self-reports
After data collection, the EEG cap was removed and partic-
ipants were seated outside of the chamber in a quiet room.
The University of Georgia, Athens (UGA) participants
reported how pleasant and aroused each scene made
them feel on a 9-point scale (with half-increments) using
the Self-Assessment Manikin (Bradley & Lang, 1994). Par-
ticipants were seated and read a script describing the rat-
ing procedure. Participants then viewed a printed version
of each scene in a binder and recorded their responses on
a worksheet. The worksheet had the Self-Assessment
Manikin figures and scales with emotion nouns at the
top of the page for participants to reference.
To quantify the additional SFs of body exposure and ori-
entation, an online sample was recruited to rate the 90
scenes depicting humans on each dimension. Participants
were instructed that the study could last an hour, but most
participants finished within 30 min. Participants accessed
the study through a Qualtrics link, which presented the
consent form and required their approval. If participants
agreed to take part in the study, they began rating each
scene in a pseudorandom order by using a click and drag
scale that ranged from 1 A 9. Each scale was displayed one
at a time underneath the scene the participant was view-
ing. The marker started in the middle of the scale on the
number 5, and participants were required to move the
marker before they could move on to the next scene
and scale. The scale allowed for precision to the tenth
decimal place (per esempio., 1.1, 1.2). For the body exposure scale,
a written question stated, “How much exposed or un-
clothed body parts are depicted in the scene?” The scale
presented below the question ranged from 1 being all
exposed body parts to 9 being no exposed body parts.
The body orientation scale stated, “How upright, O
normally oriented are the people in the scene?” with the
scale ranging from 1 indicating completely upright to 9
indicating very unusually oriented.
Quantifying Scene Features
To further quantify the extent to which exposed body
parts were present in scenes, the sum of pixels containing
exposed body parts was counted for each scene. Using
GNU Image Manipulation Program, a transparent layer
was made over each JPEG scene. On this layer, the LASSO
tool was used to manually trace the exposed body parts in
each scene. Body parts that were covered by clothes or
accessories such as arm bands and sunglasses were
excluded. After tracing, the paint bucket tool was used
to fill in the shapes with a solid color. The histogram tool
was then used to record how many pixels had been filled
by the paint bucket tool. The sum of pixels containing
exposed body parts for each scene could then be com-
pared with the total scene size.
As described above, scenes were chosen to be compa-
rable in perceptual complexity, placed in natural environ-
menti, and balanced across categories to be equivalent in
JPEG file size. The process of fully quantifying scene com-
plexity is quite complicated and beyond the scope of this
study (Donderi, 2006); Tuttavia, to expand our assess-
ment of SFs, Shannon information-entropy (from this
point simply entropy) was also calculated for each scene
using MATLAB (Mathworks.com). The entropy measure
is found by using the first order histogram frequencies
of gray values as probabilities. If every gray value is equally
likely to occur, entropy is maximized, whereas if the scene
contains only one value, then entropy is minimized. Noi
also assessed any remaining effects of JPEG file size.
Spatial frequency was assessed using the SHINE
MATLAB-based toolbox ( Willenbockel et al., 2010). Per
each scene, the relative energy was found for all possible
integers of cycles per image from the lowest frequency of
1 to the maximum of 300 (the highest frequency complete
cycle for an image with a height of 600 pixels). Low spatial
frequencies have exponentially higher relative energies
than high spatial frequencies, so energy was transformed
logarithmically with a base of 10 for analyses. Low spatial
frequency was defined as the average transformed energy
da 1 A 40 cycles, and high spatial frequency was the
average from 41 A 300 cycles. This is consistent with prior
definitions of scene spatial frequency (De Cesarei &
Codispoti, 2013). Analyses that involved spatial frequency
were also computed with thresholds at 50, 60, 70, E 80
cycles for separation between low and high spatial fre-
quencies, and the results did not meaningfully change.
EEG Data Acquisition and Reduction
Continuous EEG data were recorded using a 64-channel
BioSemi ActiveTwo system (BioSemi), which has pre-
amplified electrodes positioned according to the 10/20
system. The electrode voltage was referenced to two addi-
tional common mode electrodes (Common Mode Sense
and Driven Right Leg). The sampling rate was 512 Hz. Acti-
View acquisition software (actiview.org) was used to
ensure offsets between 50 and −50 millivolts during
EEG set up, and to monitor online data acquisition.
Offline, EEG data were segmented and preprocessed
using the Electro Magnetic Encephalography Software
analysis package for MATLAB (EMEGS; emegs.org; Peyk,
De Cesarei, & Junghöfer, 2011). Data were processed
in close accordance with the guidelines of Junghöfer,
Elbert, Tucker, and Rockstroh (2000) through a method
known as statistical correction of artifacts in dense array
studies (SCADS). The data were adjusted using a low-pass
Butterworth filter with a stopband of 40 Hz and a passband
Di 30 Hz to control for high-frequency noise. A high-pass
Butterworth filter was also implemented with a passband
944
Journal of Cognitive Neuroscience
Volume 35, Numero 6
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
J
o
C
N
_
UN
_
0
1
9
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Di 0.1 Hz and stopband of 0.05 Hz. Data were segmented
da 100 msec before to 2000 msec after scene onset.
IL 100 msec of data before scene onset was used to
baseline-adjust the ERPs. Sensors and trials were screened
for high voltage artifacts identified through an automated
analysis of each electrode per trial in EMEGS in which
the median values of maximum amplitude, standard
deviation, and the maximum first derivative were used to
find unusable trials and unreliable electrodes. Data were
then transformed to an average reference (Peyk et al.,
2011) and was again screened for artifact contaminated
trials and sensors in the same automated process. Contam-
inated sensors were removed and replaced with values
calculated through a spherical spline interpolation
allowing the least noisy and closest electrodes to contrib-
ute the most to the new replacement amplitude. IL
data were kept in an average-reference that allows for
simple visualization of the EPN, which can be difficult
to identify with other reference positions such as the
mastoid (Junghöfer et al., 2006).
After the data were preprocessed, the ERPs for each
participant were extracted for each scene and an average
was calculated for each scene category. Electrodes and
latency windows representing the EPN and LPP were
derived by a previous study that used the same scene pre-
sentation procedure (Farkas et al., 2020). The EPN was
measured by extracting voltage from the lateral-posterior
electrodes P7, PO7, O1, P8, PO8, and O2 from 150 A
300 msec after scene onset (mostrato in figura 4). The LPP
was recorded over midline parietal electrodes Cz, CPz, Pz,
CP1, and CP2 from 400 A 900 msec after scene onset
(mostrato in figura 5).
Statistical Analyses
Omnibus repeated-measures ANOVAs were used to assess
the overall effect of scene content on the ERP measures,
with paired t tests used to break down specific category
differences. This was done for the variables of valence,
arousal, rated body exposure, rated body orientation,
and EPN and LPP amplitude as μvolt change from baseline.
All statistical tests were done with R software (r-project
.org). To address our hypotheses, we planned to use
paired t tests as a tool to make clear to the reader which
scene categories had a differential impact on ERP ampli-
tudes. Così, these are not post hoc comparisons in need
of a correction for multiple comparisons. Tuttavia, we
have included in the figure descriptions which compari-
sons would not survive Bonferroni correction (.001).
Our study question required the comparative assess-
ment of multiple SFs, which cannot be assumed to be
independent. To assess the impact of our many variables
by-scene conservatively, we used multiple regression and
four VSR regression methods to discern if and how SFs are
related to EPN and LPP modulation. Multiple regression
allowed us to use all SFs as predictors for each ERP within
the same model, where each variable acts as a covariate.
This can help to identify which variables account for com-
mon or unique variance. The VSR methods were used to
identify which SFs have the most meaningful relationships
with ERP modulation, and to minimize potential impact of
collinearity of our many variables. Whereas multiple
regression maximizes the amount of variance explained,
VSR methods use additional information to find the sim-
plest and most predictive multiple regression models for
a hypothetical future data set. In other words, the VSR
models allow us to identify which variables are most
meaningful by assessing whether these variables warranted
inclusion in the found models. By using four VSR methods,
we can assess the consistency of variable relationships that
were meaningful across models. Altogether, this multistep
approach should yield a more reliable estimate of the ways
in which our SFs are associated with EPN and LPP modula-
zione. These analyses were only used for the scenes that
depicted human content, where body-related SFs are rele-
vant to our hypotheses. The SFs used for the 90 human
scenes were valence, arousal, rated body exposure, rated
body orientation, body exposure pixels, low spatial fre-
quency, high spatial frequency, JPEG file size, and entropy.
Standard multiple regression and VSR models are
shown together in Table 2. The values for each variable
were transformed into standardized beta coefficients to
simplify interpretation of the strength of the relationship.
Statistical significance for each coefficient is presented
only for the standard multiple regression results. Per
VSR models, only those variables that warranted inclusion
in the final model list a coefficient. The four VSR regression
methods used include (1) best-subset ranked by Bayesian
information criteria (BIC), (2) best-subset ranked by cross
validation, (3) LASSO regression, E (4) sparse LASSO
regression. Best-subset regression involves computing a
multiple regression for every combination of the predictor
variables. The best multiple regression model can then
be found based on an information criterion or cross valida-
tion prediction accuracy. Our first best-subset model was
found based on BIC, which balances the variance
explained by the model with a penalty for model complex-
ità. This process was implemented with the regsubsets
function contained in the leaps package (Lumley, 2017).
The next best-subset model was found with a fivefold cross
validation procedure repeated 1000 times. In this proce-
dure, each scene was randomly assigned to one of five
groups. All possible multiple regression models were then
fit to four of the groups. Prediction accuracy was then
found based on how well the models predicted ERP ampli-
tude for the fifth test group of scenes as measured by the
root-mean-square error (RMSE), which is the absolute pre-
diction error. The process was repeated with each of the
groups acting as the test data. The fivefold procedure was
repeated 1000 times to increase the reliability of the
risultati. The model with the lowest average RMSE from
all folds through 1000 iterations is presented here as the
best model. For both best-subset regression models, we
Farkas and Sabatinelli
945
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
J
o
C
N
_
UN
_
0
1
9
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
9
4
6
J
o
tu
R
N
UN
l
o
F
C
o
G
N
io
T
io
v
e
N
e
tu
R
o
S
C
io
e
N
C
e
V
o
l
tu
M
e
3
5
,
N
tu
M
B
e
R
6
Tavolo 1. The Mean Values for Valence, Arousal, EPN, LPP, Body Exposure Rating, and Body Orientation Rating
Erotica
Sports
Victory
Neutro
Nudists
Neutro
People
Threatening
People
Mutilations
Pleasant
Animals
Neutro
Animals
Threatening
Animals
Repeated-measures
ANOVA Result
UGA sample N = 50
Valence (1–9)
5.64 (0.24)C
7.02 (0.18)B
4.80 (0.15)D
5.03 (0.48)D
2.90 (0.14)F
2.40 (0.18)G
8.09 (0.16)UN
5.86 (0.10)C
4.27 (0.15)E
Arousal (1–9)
EPN (μV)
6.52 (0.20)UN
5.86 (0.24)B
4.76 (0.15)C
3.80 (0.17)E
5.97 (0.20)B
6.15 (0.30)AB
4.97 (0.33)C
4.07 (0.18)D
6.03 (0.18)D
3.93 (0.68)B
6.89 (0.58)C
2.54 (0.64)UN
9.08 (0.58)E
7.11 (0.60)CD
7.97 (0.63)D
7.82 (0.57)D
8.87 (0.62)E
6.91 (0.64)C
LPP (μV)
4.96 (0.43)UN
0.40 (0.30)F
3.06 (0.44)B −1.54 (0.26)G
1.07 (0.34)DE
1.93 (0.44)C
1.03 (0.24)DE
0.47 (0.34)EF
1.56 (0.36)CD
Mturk sample N = 70
Body exposure
(1–9)
Body orientation
(1–9)
8.48 (0.05)B
3.85 (0.13)D
8.75 (0.04)UN
1.88 (0.10)F
2.17 (0.09)E
6.30 (0.21)C
3.63 (0.20)D
7.51 (0.15)AB
7.79 (0.21)UN
7.30 (0.13)B
5.99 (0.25)C
3.21 (0.14)E
N/A
N/A
N/A
N/A
N/A
N/A
F(8, 392), = 137.81,
p = .001; ηp
2 = .738
F(8, 392) = 25.02,
p = .001; ηp
2 = .338
F(8, 392) = 55.66,
p = .001; ηp
2 = .532
F(8, 392) = 46.75,
p = .001; ηp
2 = .488
F(5, 345) = 857.16,
p = .001; ηp
2 = .925
F(5, 345) = 167.66,
p = .001; ηp
2 = .708
Standard error for each is within the parentheses. Results of pairwise comparisons are depicted with the subscript letters. If two categories in the same row share the same letter, the categories were found
to not be statistically different at a critical p value of .05. The repeated-measures ANOVA results are presented in the final column.
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
J
o
C
N
_
UN
_
0
1
9
8
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
report how the model fits on the overall set of scenes. Questo
is because best-subset regression is a variable selection
algorithm and does not return a model with coefficients
tuned for future use. The third VSR method used was
LASSO regression, which is a regularization method that
shrinks slope coefficients by a L1 penalty scaled by a tuning
parameter. The optimum tuning parameter was found
based on a fivefold × 1000 iteration cross validation pro-
cedure. Similar to best-subset regression, this method can
remove variables from the model that do not improve pre-
diction accuracy. Tuttavia, it is computationally less
expensive and returns a model that should be more pre-
dictive in a new data set. Sparse LASSO regression is the
model with the largest L1 penalty, which was still within
1 SE of the RMSE of the best LASSO regression. The sparse
model is nearly equivalent in accuracy, but yields fewer,
more essential predictor variables. A simpler model is gen-
erally preferred because it is easier to interpret and is less
susceptible to overfitting.
RESULTS
Scene Category Analyses – Self Report
Valence was significantly different across scene categories
as found by a repeated-measures ANOVA, F(8, 392) =
137.81, P < .001; ηp
2 = .738. Overall category results
can be seen in Table 1. Results of pairwise comparisons
are depicted with the subscript letters, such that if two
categories were statistically equivalent, they share the
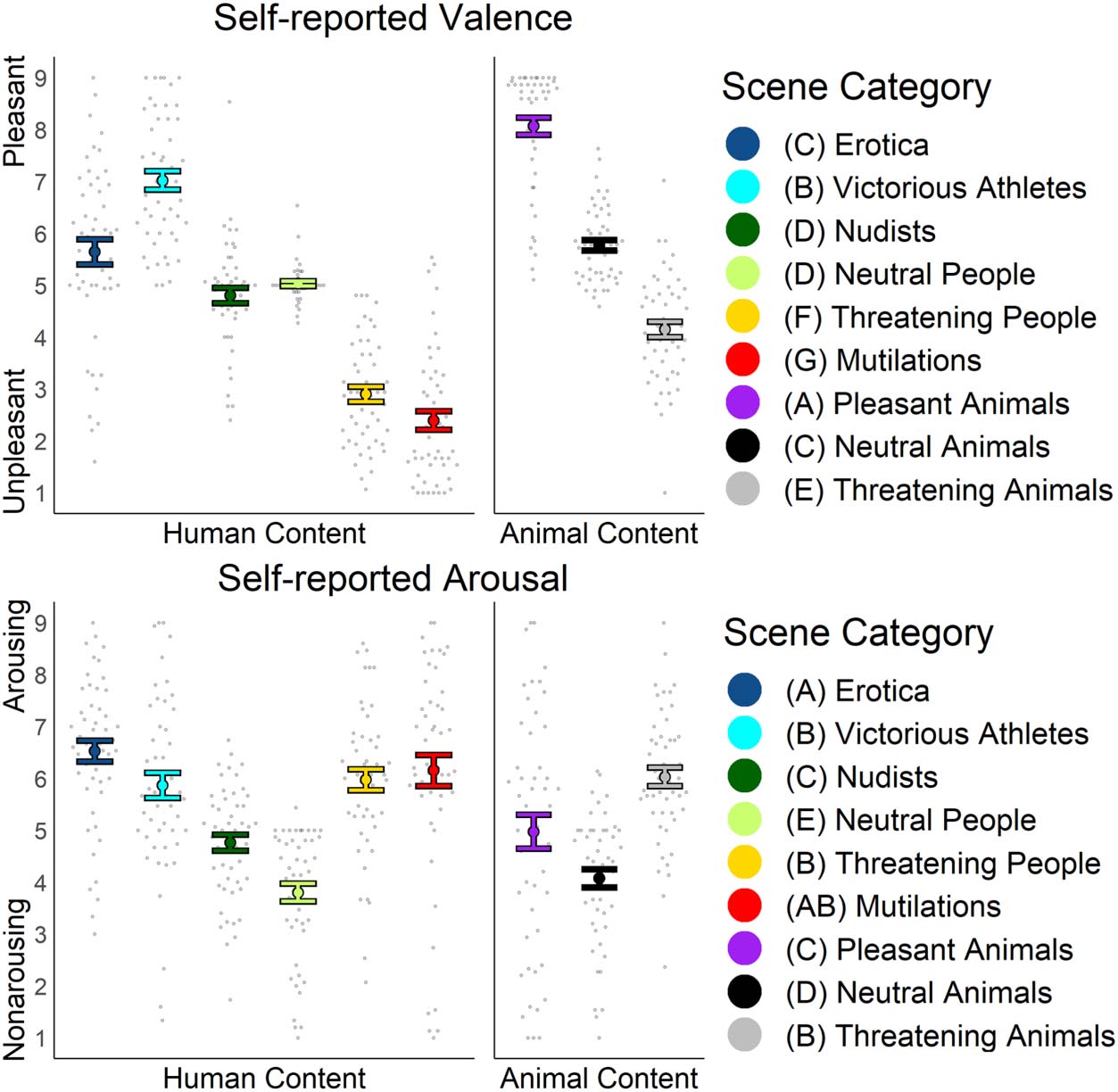
same letter. Figure 2 shows individual participant valence
and arousal self-reports as well as the overall sample mean
for each category. Pleasant animal scenes elicited the high-
est valence ratings, which differed from the second-most
pleasant category of victory scenes, t(49) = 7.28, p < .001,
d = 1.03. Victory scenes elicited more rated pleasantness
than the neutral animal category, t(49) = 6.48, p < .001,
d = 0.92, as well as erotica scenes, t(49) = 4.70, p < .001,
d = 0.67. Erotic scenes where not different than neutral
animals valence ratings, but received higher pleasantness
ratings than scenes of neutral people, t(49) = 2.43, p =
.019, d = 0.34. Valence ratings did not differ between neu-
tral people and nudist scenes. Nudist scenes elicited
higher valence ratings than threatening animal scenes,
t(49) = 2.77, p = .007, d = 0.39. However, the threatening
animals evoked less valence ratings than threatening peo-
ple scenes. Finally, mutilation scenes elicited the lowest
valence ratings as compared with the second most
unpleasant scene category of threatening people, t(49) =
2.98, p = .004, d = 0.42 (Table 2).
A repeated-measures ANOVA was significant for the
self-reports of emotional arousal across the nine scene
categories, F(8, 392) = 25.02, p < .001; ηp
2 = .338. Erotic
and mutilation scenes elicited the highest ratings of
arousal and were not statistically different. Erotica scenes
evoked more rated arousal than threatening animal
scenes, t(49) = 2.51, p = .016, d = 0.35; threatening people
scenes, t(49) = 2.23, p = .030, d = 0.32; and victorious
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
Figure 2. Self-reported valence
and arousal for each category.
Each gray dot is the category
average for a participant (n =
50). Colored dots and error bars
are the overall category average
and standard error for all
participants. Letters on the key
indicate which categories were
statistically different by paired
t tests. If two categories share
the same letter, there was not a
significant difference between
the categories at critical p value
of .05. If corrected for multiple
comparisons to a critical p value
of .001, valence ratings would
not be different between
the pairs of erotica versus
neutral people, nudists versus
threatening animals, and
mutilation versus threatening
people. Using the same
threshold, arousal ratings would
not have differed for erotica
versus threatening animals,
erotica versus threatening
people, erotica versus victorious
athletes, victorious athletes
versus pleasant animals,
and neutral people versus
neutral animals.
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Farkas and Sabatinelli
947
9
4
8
J
o
u
r
n
a
l
o
f
C
o
g
n
i
t
i
v
e
N
e
u
r
o
s
c
i
e
n
c
e
V
o
l
u
m
e
3
5
,
N
u
m
b
e
r
6
Table 2. Multiple Regression and VSR Models Result for the 90 Human Scenes
Multiple Regression Standardize Beta Coefficients
Model Statistics
Valence
Arousal
JPEG File
Size
Entropy
Body
Exposure
Rating
Body
Orientation
Rating
Body
Exposure
Pixels
Low-spatial
Frequency
High-spatial
Frequency
F
p
R 2
adj
BIC
RMSE
EPN
Standard multiple
−.17
−.14
.01
.10
−.48**
−.28*
−.14
−.23
−.13
11.90 < .001
.52
410
−.74
−.64
−.49
−.46
.10
.08
−.27
−.37
−.22
−.10
.22
.19
−.01
−.12
−.10
392
396
1.93
2.08
2.16
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
−.00
.33**
−.04
.11
.78**
−.07
−.20*
−.18
.06
29.58 < .001
.74
317
.36
.38
.32
.23
.69
.79
.66
.57
−.15
−.04
−.04
−.05
.01
297
297
1.14
1.21
1.25
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Model and variable statistical significance is only applicable for the standard multiple regression. For Models 2 through 5, variables can be deemed important if they warranted inclusion in the final reported
model.
* p < .05.
** p < .001.
regression
Best-subset
regression (BIC)
Best-subset
regression (CV)
−.17
LASSO regression
−.15
−.10
Sparse LASSO
regression
LPP
Standard multiple
regression
Best-subset
regression
(BIC)
Best-subset
regression (CV)
LASSO regression
Sparse LASSO
regression
athlete scenes, t(49) = 2.49, p = .016, d = 0.35. Mutila-
tion, threatening animals, threatening people, and victori-
ous athletes scenes were not different in self-reported
arousal. Pleasant animal scenes evoked less arousal than
victorious athlete scenes, t(49) = 2.32, p = .024, d =
0.33, but were not different in rated arousal from nudist
scenes. Nudist scenes elicited more arousal than neutral
animal scenes, t(49) = 3.39, p = .001, d = 0.48. Lastly,
participants reported that neutral animal scenes were
more arousing than the neutral people scenes, t(49) =
2.45, p = .018, d = 0.35.
The Mturk sample reported the six human scene cate-
gories had a different amount of exposed body parts as
found through a repeated-measures ANOVA, F(5, 345) =
857.16, p < .001; ηp
2 = .925. Mturk ratings of body expo-
sure and body orientation can be seen in Table 1 and
Figure 3. The nudist category was perceived as having
the most exposed body parts and were significantly differ-
ent than erotic scenes, t(69) = 6.30, p < .001, d = 0.75.
Erotic scenes were perceived as having more exposed
body parts than mutilation scenes, t(69) = 10.93, p <
.001, d = 1.31. Participants felt mutilation scenes had
more body part exposure than victorious athlete scenes,
t(69) = 12.25, p < .001, d = 1.46. In turn, victorious ath-
lete scenes were rated as including less exposed body than
threatening people scenes, t(69) = 19.30, p < .001, d =
2.31. Threatening people scenes were reported as more
clothed than the final scene category of neutral people,
t(69) = 6.51, p < .001, d = 0.78.
Mturk reports of body orientation were different
between scene categories, F(5, 345) = 167.66, p < .001,
ηp
2 = .708. Participants rated the nudist category as having
the most normally oriented depicted people, but the
nudists scenes were not statistically different to the second
highest rated category of victorious athletes. Nudist
scenes were more normally oriented than neutral people
scenes, t(69) = 2.16, p = .034, d = 0.26, but victorious ath-
lete scenes were rated as not different than the neutral
people category. The neutral people were more normally
oriented than the threatening people scenes, t(69) = 5.26,
p < .001, d = 0.63. The threatening people category was
rated higher than the erotic scenes, t(69) = 7.77, p < .001,
d = 0.93. Finally, the erotic scenes were reported as more
normally oriented as the people in the mutilation scenes,
t(69) = 2.18, p = .033, d = 0.26.
Scene Category Analyses
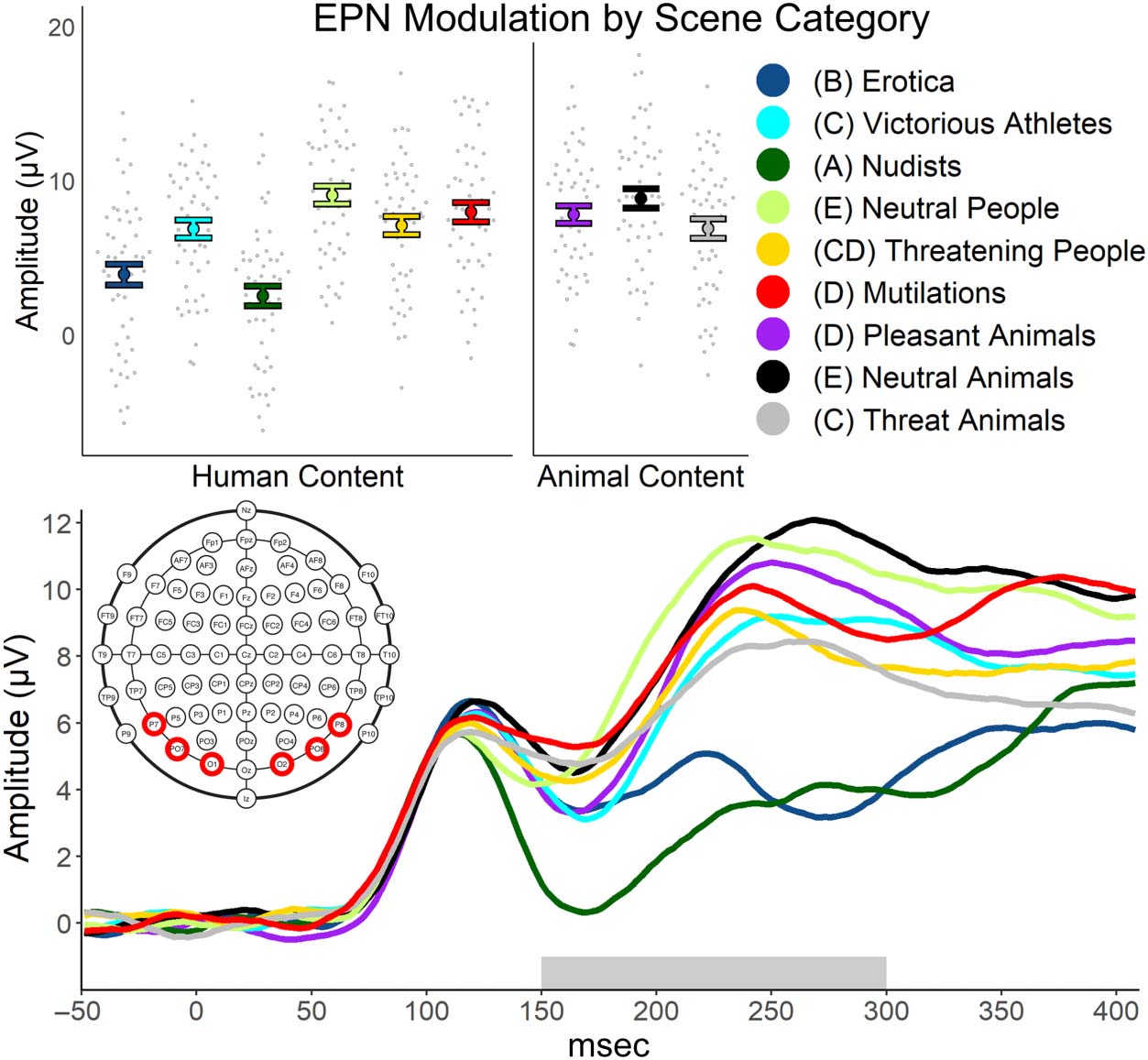
EPN. A repeated-measures ANOVA found significant
modulation of the EPN across the nine scene categories,
F(8, 392) = 55.66, p < .001; ηp
2 = .532. The EPN waveforms
can be seen in Figure 4, whereas the amplitudes are shown
in Table 1, including significant contrast effects, indicated
by different subscript letters. Nudist scenes modulated
the EPN the most and were significantly different than
modulation evoked by erotic scenes, t(49) = 4.41, p <
.001, d = 0.62. EPN modulation from erotic scenes were
greater than modulation elicited by victorious athletes,
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
Figure 3. Perceived body
exposure and body orientation
per each category from the
Mturk sample (n = 70). Each
gray dot represents the average
rating for each participant,
whereas the colored dots and
error bars represent the overall
average and standard error,
respectively. In the legend, if
categories do not share a letter,
they were statistically different
at a critical p value of .05.
If corrected for multiple
comparisons to a critical p value
of .001, all category differences
in body exposure ratings would
still be statistically significant.
Using the same threshold, body
orientation ratings would not
have statistically differed for
nudists versus neutral people
and erotica versus mutilation
scenes.
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Farkas and Sabatinelli
949
Figure 4. The top depicts
the average participant and
category EPN amplitude by
category. The bottom displays
the EPN waveform by category.
In the legend, if categories do
not share a letter, they were
statistically different at a critical
p value of .05. If corrected for
multiple comparisons to a
critical p value of .001, the EPN
would not have statistically
differed for pleasant animals
versus victorious athletes,
pleasant animals versus
threatening animals, pleasant
animals versus neutral animals,
mutilation versus neutral
animals, and mutilation versus
neutral people.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
t(49) = 8.00, p < .001, d = 1.13; threatening animals,
t(49) = −7.50, p < .001, d = 1.06; and threatening people
scenes, t(49) = 8.59, p < .001, d = 1.21. However, the
three categories of victorious athletes, threatening
animals, and threatening people did not differ significantly
in EPN modulation. Pleasant animal scenes modulated
the EPN less than victorious athlete scenes, t(49) = 2.61,
p = .012, d = 0.37, and the threatening animal scenes,
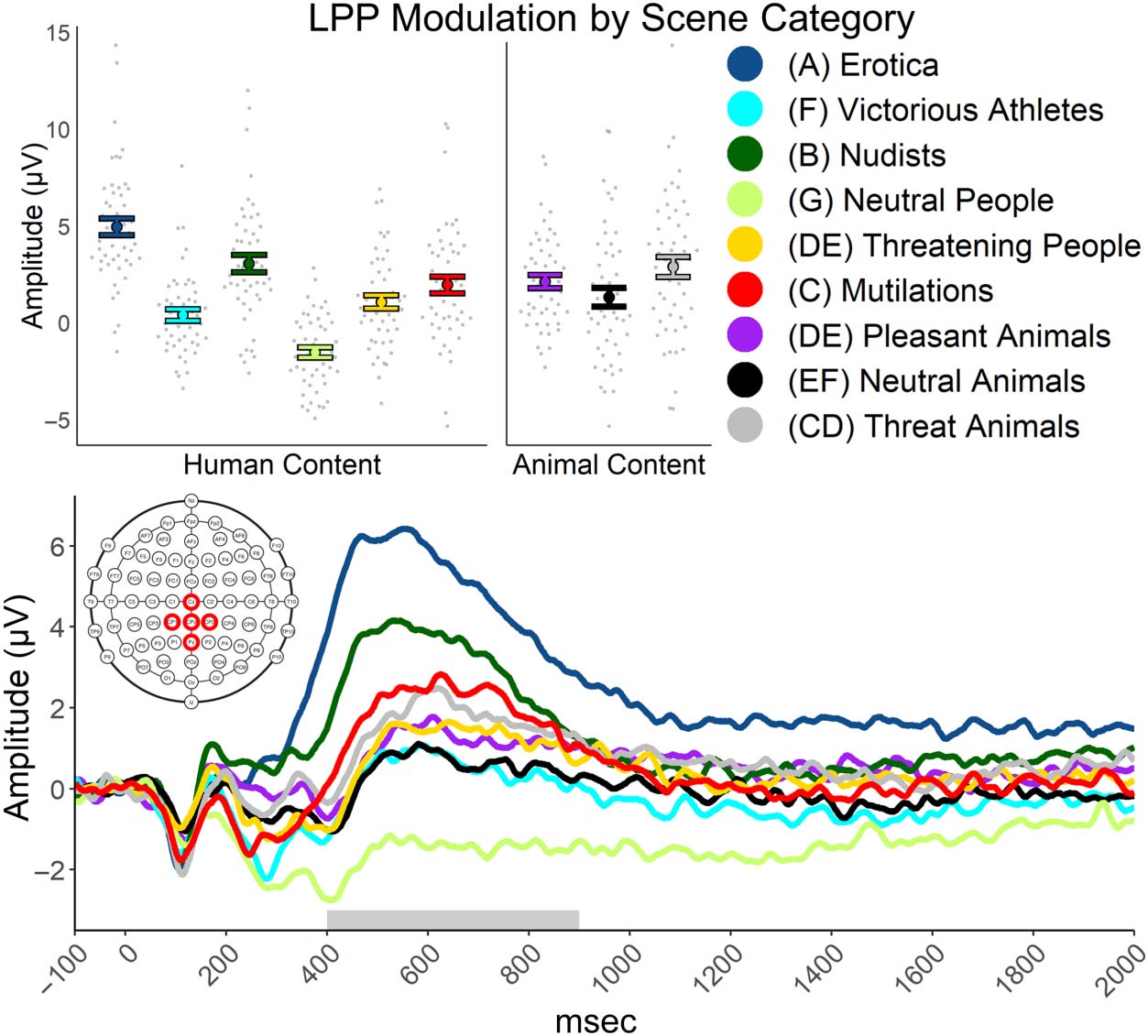
Figure 5. The top depicts
the average participant and
category LPP amplitude by
category. The bottom displays
the LPP waveform by category.
In the legend, if categories do
not share a letter, they were
statistically different at a critical
p value of .05. If corrected for
multiple comparisons to a
critical p value of .001, the LPP
would not have statistically
differed for nudists versus
mutilation, nudists versus
threatening animals, mutilation
versus threatening people,
mutilation versus pleasant
animals, threatening animals
versus neutral animals,
threatening animals versus
victorious athletes, and
threatening people versus
victorious athletes.
950
Journal of Cognitive Neuroscience
Volume 35, Number 6
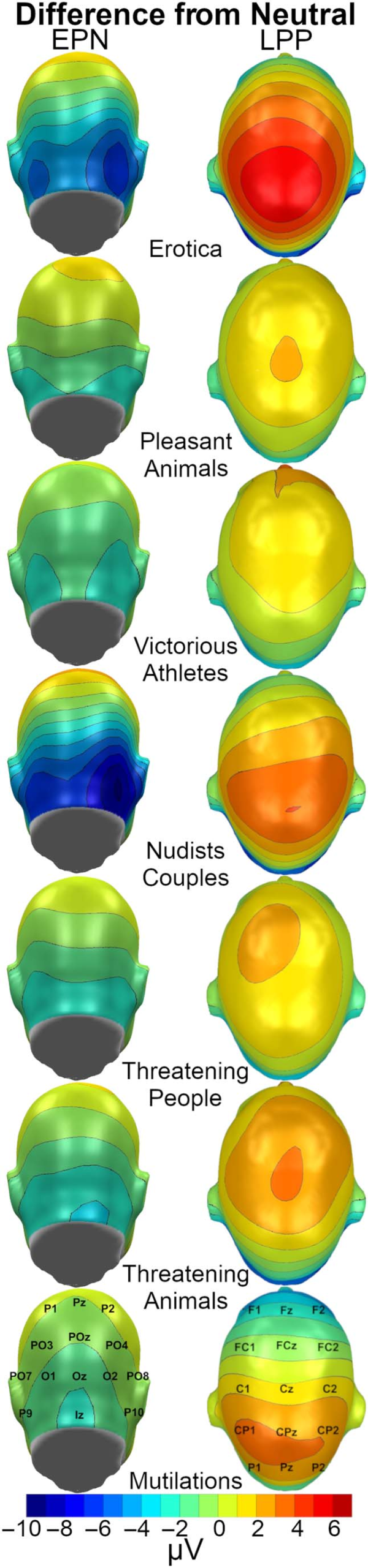
Figure 6. Scene category
topographies for the EPN
(150–300 msec) and LPP
(400–900 msec) visualized as
the difference in microvolts
from the neutral people
and animal categories. Each
color represents a range of
1 microvolt, such that the
darkest blue represents a
microvoltage difference
between −10 and −9.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Farkas and Sabatinelli
951
t(49) = 2.54, p = .014, d = 0.36. However, the pleasant
animal, threatening people, and mutilation scenes did
not differ in EPN amplitude significantly. Mutilation
scenes modulated the EPN more than neutral animals,
t(49) = 2.34, p = .023, d = 0.33, and neutral people scenes,
t(49) = 2.45, p = .018, d = 0.35. Neutral animal and neu-
tral people scenes did not differ in EPN modulation.
LPP. A repeated-measures ANOVA found a difference in LPP
modulation across the nine scene categories, F(8, 392) =
46.75, p < .001; ηp
2 = .488. The LPP waveforms can be seen
in Figure 5, whereas the amplitudes are shown in Table 1,
including contrast effects. Erotic scenes modulated the
LPP the most and was statistically different than the next
most potent scene category of nudists, t(49) = 6.11, p <
.001, d = 0.86. Nudist scenes modulated the LPP more
than mutilation, t(49) = 2.94, p = .005, d = 0.42, and
threatening animal scenes, t(49) = 3.21, p = .002, d =
0.45. However, mutilation and threatening animal scenes
statistically did not differ in LPP amplitude. Mutilation
scenes evoked more LPP modulation than threatening
people scenes, t(49) = 2.85, p = .006, d = 0.40, and
pleasant animals scenes, t(49) = 2.14, p = .037, d =
0.30. Threatening animal modulation was not different
to the modulation of threatening people and pleasant
animal categories. Neutral animal scenes elicited less LPP
modulation than the threatening animal scenes, t(49) =
2.82, p = .007, d = 0.40, but the neutral animals were
not different than threatening people and happy animal
scenes. Victorious athlete scenes modulated the LPP less
than threatening people, t(49) = 2.15, p = .037, d = 0.30,
and threatening animal scenes, t(49) = 3.29, p = .002, d =
0.46, whereas the victorious athlete scenes were not
different to modulation by neutral animal scenes. Neutral
people scenes modulated the LPP less than all other cate-
gories including the next closest category of victorious
athletes, t(49) = 7.30, p < .001, d = 1.03 (Figure 6).
By Scene Analyses
EPN. The EPN standard multiple regression was signifi-
cant, F(9, 80) = 11.90, p < .001, R2
adj = .52. Notably, rated
arousal was not significantly associated with EPN modula-
tion (β = −.17, p = .164). Scenes, which were rated with
high body exposure (β = −.48, p < .001) and upright ori-
entation (β = −.28, p = .024) were associated with greater
EPN modulation. The best-subset regression based on BIC
included only rated body exposure (β = −.74) and body
orientation (β = −.27) as meaningful. The best-subset
regression based on prediction accuracy (RMSE =1.93)
included arousal (β = −.17), rated body exposure (β =
−.64), rated body orientation (β = −.37), low-spatial fre-
quency (β = .22), and high-spatial frequency (β = −.12)
as meaningful. The most predictive LASSO regression
(RMSE = 2.08) included all predictor variables except
JPEG file size. The sparse LASSO regression (RMSE =
2.16) included body exposure ratings (β = −.46), entropy
(β = .08), body orientation ratings (β = −.01), and low-
spatial frequency (β = .01) as meaningful.
LPP. A standard multiple regression of LPP modulation
across the 90 human scene set was significant, F(9, 80) =
adj = .74. Higher rated arousal (β =
29.58, p < .001, R2
.33, p < .001) and rated body exposure (β = .78, p <
.001) was associated with a larger LPP amplitude, whereas
the sum of body exposure pixels (β = −.20, p = .024) had a
negative relationship with the LPP. The best-subset model
based on BIC included rated body exposure (β = .69) and
rated arousal (β = .36). The best-subset model based on
prediction accuracy (RMSE = 1.14) included arousal (β =
.38), rated body exposure (β = .79), and body exposure
pixels (β = −.15). The most predictive LASSO regression
(RMSE = 1.21) included rated arousal (β = .32), body expo-
sure ratings (β = .66), body orientation ratings (β = −.04),
body exposure pixels (β = −.04), low-spatial frequency
(β = −.05), and high-spatial frequency (β = .01). The
sparse LASSO regression (RMSE = 1.25) included body
exposure ratings (β = .57) and arousal ratings (β = .23).
DISCUSSION
In this study, we directly assessed the extent to which the
emotion-modulated EPN may be sensitive to the presence
of exposed body parts in scenes, independent from rated
arousal. Across the wide range of scenes selected, the
results replicated our initial study (Farkas et al., 2020)
and supported the hypothesis that highly exposed bodies
and upright body orientation together are strong predic-
tors of EPN amplitude, whereas the rated arousal of scenes
was weakly predictive. This pattern contrasts with modu-
lation of the LPP, which consistently reflected scene
arousal ratings.
The scene category effect size differences of the EPN
and LPP suggest a different hierarchy of sensitivity. Nudist
scenes clearly elicited the largest EPN, showing a
medium/large 0.62 effect size over erotica, the second-
most effective category. This difference is equivalent to
the 0.61 effect size between the remaining emotional
scene categories and neutral scenes. Averaged together,
the nudist and erotica scenes showed a very large 1.38
effect size compared with all other emotional categories.
Conversely, erotic scenes elicited the largest LPP, showing
a 0.86 effect size over nudists, the second-most effective
category. This difference is similar in size to the 1.00 effect
between the remaining emotional scene categories and
neutral scenes. Averaged together, LPP modulation to
the nudist and erotica scenes showed a 1.26 effect size dif-
ference as compared with all other emotional categories.
The regression models fit to the 90 individual scenes
showed that body exposure ratings were a reliable predic-
tor of EPN and LPP amplitude, whereas standardized beta-
coefficients indicate that body exposure ratings had the
strongest relationship with the EPN. Notably, rated arousal
was not significantly related to EPN modulation in the
952
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
standard multiple regression, and was selected in just two
of the four VSR models. For LPP modulation, rated arousal
and body exposure ratings were consistently strong
predictors.
These data suggest that the EPN may reflect the early
activation of brain networks that function to enhance the
perceptual processing of classes of stimuli that reliably
predict relevant outcomes (Frank & Sabatinelli, 2017;
Bradley, 2009; Lang, Bradley, & Cuthbert, 1998). Specifi-
cally, scenes clearly featuring people may enhance EPN
modulation because people are associated with aversive
and appetitive experience, and it may be that the more
clearly a person is depicted in a scene (e.g., upright,
unclothed), the larger the EPN, possibly reflecting reen-
trant feedback from downstream ventral visual structures
(Kravitz, Saleem, Baker, Ungerleider, & Mishkin, 2013).
After this potentially meaningful cue is further processed
across the brain, LPP modulation strongly reflects the
rated emotional intensity of the scene, while being less
sensitive to clearly visible people. This might explain the
pronounced sensitivity of the EPN to nudist, relative to
erotic scenes, whereas the LPP shows the reverse pattern.
Despite the apparent bias toward bodies, the EPN
remains sensitive to nonbody emotional cues as well. This
is evident in the EPN modulation by pleasant (D = 0.39)
and unpleasant (D = 0.63) animals scenes relative to neu-
tral animal scenes in the current study, and in published
work using emotional faces (Schupp, Öhman, et al.,
2004), hand gestures (Flaisch et al., 2009), and even words
(Kissler et al., 2007).
Body-specific Activity in the Visual System
There is considerable evidence that body percepts have a
privileged status in the human visual system. Studies with
newborns have found that upright bodies draw attention
more than inverted bodies or other stimuli (Filippetti,
Johnson, Lloyd-Fox, Dragovic, & Farroni, 2013). fMRI
studies have identified a region of lateral occipital cortex
that is particularly sensitive to line-drawings of bodies
relative to tools (Peelen & Downing, 2007). In the ERP
literature, the perception of a headless body on a blank
background can elicit a potential similar to the N170
elicited by faces. Although this body-evoked N170 appears
at a more anterior scalp location to the EPN and dissipates
by 210 msec (Thierry et al., 2006), it has been shown that
a greater proportion of skin exposure increases the ampli-
tude of this potential (Alho, Salminen, Sams, Hietanen, &
Nummenmaa, 2015; Hietanen, Kirjavainen, & Nummenmaa,
2014; Hietanen & Nummenmaa, 2011). These findings
suggest that the human visual system is especially sensi-
tive to body cues.
If bodies elicit a pronounced EPN amplitude because the
visual system is tuned to identify these cues, then the EPN
could be a useful metric of other features for which the
visual system shows a similar processing bias. In other
words, the EPN may be less an index of emotional
perception, and more a measure of the degree of early
recognition of features that predict emotional percepts.
Consistent with this distinction, snakes also appear to show
particularly efficient processing in the visual system (Soares,
Lindstrom, Esteves, & Öhman, 2014; Öhman & Mineka,
2001), and also elicit larger EPN amplitudes than would
be expected as compared with arousal ratings and LPP mod-
ulation (Van Strien, Eijlers, Franken, & Huijding, 2014; Van
Strien, Franken, & Huijding, 2014). Naturally, those cues
that predict emotional percepts will most often be emo-
tional, and thus elicit similar EPN and LPP modulation, in
keeping with rated arousal. Because of its early latency
and broad reactivity to naturalistic scenes, the EPN may
enable research to identify the nature and extent of these
visual features, and thus help to define the subtle mecha-
nisms of emotional perception in large human samples.
Limitations and Future Directions
Although this work expanded the variety of scene stimuli
and feature quantification from earlier efforts, it could still
benefit from more heterogeneity of scenes, additional
measures of experienced arousal, and more precise calcu-
lations of exposed body parts. To understand how specific
scene content and features affected EPN modulation, we
curated scenes that were quite similar in the content
within each category. Although this was useful and inten-
tional for the main objectives of this study, it is possible
that discrete content categories affect our by-scene analy-
ses. Potentially, this could lead to what is referred to as a
Simpson’s paradox, in which the overall relationship does
not reflect associations within each category. This concern
could be addressed in future studies by evaluating the rela-
tionship between perceived body exposure and EPN
amplitude for a larger, more diverse scene set. Separately,
the LPP results suggest that nudist scenes may have been
more arousing than the participants self-reported. This is a
difficult issue, but future studies could include physiolog-
ical measures of arousal such as skin conductance or pupil
diameter. Lastly, although the sum of pixels that contained
exposed body parts was associated with EPN modulation,
it did not capture the effect as well as self-reported body
exposure. Although we aimed to keep the action of a
scene consistently placed in the middle, and within a stan-
dard depth of field, the distance of body parts from the
camera lens does vary across scenes, and may have an
impact on the utility of this index. Future studies could
also investigate if there are intermediate-level visual fea-
tures of a body that are especially potent modulators of
the EPN. A computer simulation study suggests that small
fragments of images, such as a part of a face, can provide
useful information for accurate classification (Ullman,
Vidal-Naquet, & Sali, 2002). Using a similar approach with
fragmented pictures of bodies may reveal which features
are most influential in body classification, and if the EPN is
modulated by specific body features, or the recognition of
a full body.
Farkas and Sabatinelli
953
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Consistent with prior work that manipulated scene
complexity (Nordström & Wiens, 2012; Löw et al., 2013;
Bradley et al., 2007), a subset of our analyses suggest that
spatial frequency of scenes modulated EPN amplitude,
despite the fact that the scene set was composed to be
equivalent in JPEG file size across categories. The use of
additional indices of scene complexity will be needed to
resolve this issue more fully, but we are encouraged by
the relationship between body exposure ratings and
ERP modulation, which strongly surpassed the modest
relationship between low spatial frequency and the ERPs.
Considering that the original EPN study (Junghöfer et al.,
2001) employed nearly the entire IAPS set (1000+
scenes) and did not identify any reliable relationships with
scene color, luminance, or spatial frequency, we are rea-
sonably confident that the nudist-erotica EPN effect is not
a result of low-level perceptual differences.
Conclusion
These data suggest a refinement in a common interpreta-
tion of what the EPN component represents in visual per-
ception. Researchers have generally concluded that the
EPN is modulated primarily by arousing content, much like
the LPP (Frank & Sabatinelli, 2019; Sabatinelli et al., 2013;
Olofsson et al., 2008; Schupp, Flaisch, Stockburger, &
Junghöfer, 2006). The current findings suggest that scenes
depicting unclothed bodies in upright postures elicit a
greater EPN independent of arousal ratings and LPP
amplitude. Thus, in addition to reflecting motivational
relevance in general, this EPN sensitivity might be a result
of a learned or phylogenetic tendency to attend to human
bodies. This ability to distinguish the impact of SFs on early
and late stage emotion-modulated ERPs could enable future
studies to identify other visual features that show this dis-
tinction, thus providing a means to differentiate the mech-
anisms of emotional perception with noninvasive measures.
Acknowledgments
We sincerely thank Heather DiGregorio, Hannah Myers, and
Ansley Quinn Delaney for their help on data acquisition and
management.
Reprint requests should be sent to Andrew H. Farkas, Depart-
ment of Psychology, University of Georgia, Athens, GA 30602,
or via e-mail: andrew.farkas@uga.edu.
Data Availability Statement
Data and experimental materials from this study will be
made available upon request to the authors.
Author Contributions
Andrew H. Farkas: Conceptualization; Data curation;
Formal analysis; Investigation; Methodology; Project
administration; Software; Supervision; Validation;
Visualization; Writing—Original draft; Writing—Review &
editing. Dean Sabatinelli: Conceptualization; Formal anal-
ysis; Funding acquisition; Methodology; Project adminis-
tration; Resources; Supervision; Validation; Visualization;
Writing—Review & editing.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 to 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender iden-
tification of first author/last author) publishing in the Jour-
nal of Cognitive Neuroscience ( JoCN) during this period
were M(an)/M = .407, W(oman)/M = .32, M/ W = .115,
and W/ W = .159, the comparable proportions for the arti-
cles that these authorship teams cited were M/M = .549,
W/M = .257, M/ W = .109, and W/ W = .085 (Postle and
Fulvio, JoCN, 34:1, pp. 1–3). Consequently, JoCN encour-
ages all authors to consider gender balance explicitly when
selecting which articles to cite and gives them the oppor-
tunity to report their article’s gender citation balance. The
authors of this article report its proportions of citations by
gender category to be as follows: M/M = .521; W/M = .271;
M/ W = .125; W/ W = .083.
REFERENCES
Alho, J., Salminen, N., Sams, M., Hietanen, J. K., & Nummenmaa,
L. (2015). Facilitated early cortical processing of nude human
bodies. Biological Psychology, 109, 103–110. https://doi.org
/10.1016/j.biopsycho.2015.04.010, PubMed: 25960070
Bradley, M. M. (2009). Natural selective attention: Orienting and
emotion. Psychophysiology, 46, 1–11. https://doi.org/10.1111
/j.1469-8986.2008.00702.x, PubMed: 18778317
Bradley, M. M., Hamby, S., Löw, A., & Lang, P. J. (2007). Brain
potentials in perception: Picture complexity and emotional
arousal. Psychophysiology, 44, 364–373. https://doi.org/10
.1111/j.1469-8986.2007.00520.x, PubMed: 17433095
Bradley, M. M., & Lang, P. J. (1994). Measuring emotion: The
Self-Assessment Manikin and the semantic differential.
Journal of Behavior Therapy and Experimental Psychiatry,
25, 49–59. https://doi.org/10.1016/0005-7916(94)90063-9,
PubMed: 7962581
Caplar, N., Tacchella, S., & Birrer, S. (2017). Quantitative
evaluation of gender bias in astronomical publications from
citation counts. Nature Astronomy, 1, 1–5. https://doi.org/10
.1038/s41550-017-0141
Cuthbert, B. N., Schupp, H. T., Bradley, M. M., Birbaumer, N., &
Lang, P. J. (2000). Brain potentials in affective picture
processing: Covariation with autonomic arousal and affective
report. Biological Psychology, 52, 95–111. https://doi.org/10
.1016/S0301-0511(99)00044-7, PubMed: 10699350
De Cesarei, A., & Codispoti, M. (2013). Spatial frequencies
and emotional perception. Reviews in the Neurosciences,
24, 89–104. https://doi.org/10.1515/revneuro-2012-0053,
PubMed: 23183741
Dion, M. L., Sumner, J. L., & Mitchell, S. M. (2018). Gendered
citation patterns across political science and social science
methodology fields. Political Analysis, 26, 312–327. https://
doi.org/10.7910/DVN/R7AQT1
954
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Donderi, D. C. (2006). An information theory analysis of visual
complexity and dissimilarity. Perception, 35, 823–835. https://
doi.org/10.1068/p5249, PubMed: 16836047
Dworkin, J. D., Linn, K. A., Teich, E. G., Zurn, P., Shinohara,
R. T., & Bassett, D. S. (2020). The extent and drivers of
gender imbalance in neuroscience reference lists. Nature
Neuroscience, 23, 918–926. https://doi.org/10.1038/s41593
-020-0658-y 5, PubMed: 32561883
Farkas, A. H., Oliver, K. I., & Sabatinelli, D. (2020). Emotional
and feature-based modulation of the early posterior
negativity. Psychophysiology, 57, e13484. https://doi.org/10
.1111/psyp.13484, PubMed: 31573679
Filippetti, M. L., Johnson, M. H., Lloyd-Fox, S., Dragovic, D., &
Farroni, T. (2013). Body perception in newborns. Current
Biology, 23, 2413–2416. https://doi.org/10.1016/j.cub.2013.10
.017, PubMed: 24268410
Flaisch, T., Schupp, H. T., Renner, B., & Junghöfer, M. (2009).
Neural systems of visual attention responding to emotional
gestures. Neuroimage, 45, 1339–1346. https://doi.org/10
.1016/j.neuroimage.2008.12.073, PubMed: 19349245
Frank, D. W., & Sabatinelli, D. (2017). Primate visual perception:
Motivated attention in naturalistic scenes. Frontiers in
Psychology, 8, 226. https://doi.org/10.3389/fpsyg.2017.00226,
PubMed: 28265250
Frank, D. W., & Sabatinelli, D. (2019). Hemodynamic and
electrocortical reactivity to specific scene contents in
emotional perception. Psychophysiology, 56, e13340. https://
doi.org/10.1111/psyp.13340, PubMed: 30776100
Fulvio, J. M., Akinnola, I., & Postle, B. R. (2021). Gender
(im)balance in citation practices in cognitive neuroscience.
Journal of Cognitive Neuroscience, 33, 3–7. https://doi.org
/10.1162/jocn_a_01643, PubMed: 33078992
Herbert, C., Junghöfer, M., & Kissler, J. (2008). Event related
potentials to emotional adjectives during reading.
Psychophysiology, 45, 487–498. https://doi.org/10.1111/j.1469
-8986.2007.00638.x, PubMed: 18221445
Herbert, C., Sfärlea, A., & Blumenthal, T. (2013). Your emotion
or mine: Labeling feelings alters emotional face perception—
An ERP study on automatic and intentional affect labeling.
Frontiers in Human Neuroscience, 7, 378. https://doi.org/10
.3389/fnhum.2013.00378, PubMed: 23888134
Hietanen, J. K., Kirjavainen, I., & Nummenmaa, L. (2014).
Additive effects of affective arousal and top-down attention
on the event-related brain responses to human bodies.
Biological Psychology, 103, 167–175. https://doi.org/10.1016/j
.biopsycho.2014.09.003, PubMed: 25224182
Hietanen, J. K., & Nummenmaa, L. (2011). The naked truth:
The face and body sensitive N170 response is enhanced for
nude bodies. PLoS One, 6, e24408. https://doi.org/10.1371
/journal.pone.0024408, PubMed: 22110574
Jaworska, N., Thompson, A., Shah, D., Fisher, D., Ilivitsky, V.,
& Knott, V. (2012). Acute tryptophan depletion effects on the
vertex and late positive potentials to emotional faces in individuals
with a family history of depression. Neuropsychobiology, 65,
28–40. https://doi.org/10.1159/000328992, PubMed: 22094352
Junghöfer, M., Bradley, M. M., Elbert, T. R., & Lang, P. J. (2001).
Fleeting images: A new look at early emotion discrimination.
Psychophysiology, 38, 175–178. https://doi.org/10.1111/1469
-8986.3820175, PubMed: 11347862
Junghöfer, M., Elbert, T., Tucker, D. M., & Rockstroh, B. (2000).
Statistical control of artifacts in dense array EEG/MEG
studies. Psychophysiology, 37, 523–532. https://doi.org/10
.1111/1469-8986.3740523, PubMed: 10934911
Junghöfer, M., Sabatinelli, D., Bradley, M. M., Schupp, H. T.,
Elbert, T. R., & Lang, P. J. (2006). Fleeting images:
Rapid affect discrimination in visual cortex. NeuroReport,
17, 225–229. https://doi.org/10.1097/01.wnr.0000198437
.59883.bb, PubMed: 16407776
Kissler,J.,Herbert,C.,Peyk,P.,&Junghöfer,M.(2007).Buzzwords:
Early cortical responses to emotional words during reading.
Psychological Science, 18, 475–480. https://doi.org/10.1111/j
.1467-9280.2007.01924.x, PubMed: 17576257
Kissler, J., & Herbert, C. (2013). Emotion, Etmnooi, or
Emitoon?–faster lexical access to emotional than to neutral
words during reading. Biological Psychology, 92, 464–479.
https://doi.org/10.1016/j.biopsycho.2012.09.004, PubMed:
23059636
Kravitz, D. J., Saleem, K. S., Baker, C. I., Ungerleider, L. G., &
Mishkin, M. (2013). The ventral visual pathway: An expanded
neural framework for the processing of object quality. Trends
in Cognitive Sciences, 17, 26–49. https://doi.org/10.1016/j.tics
.2012.10.011, PubMed: 23265839
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (1997).
International Affective Picture System (IAPS): Technical
manual and affective ratings. NIMH Center for the Study of
Emotion and Attention.
Lang, P. J., Bradley, M. M., & Cuthbert, B. N. (1998).
Emotion, motivation, and anxiety: Brain mechanisms and
psychophysiology. Biological Psychiatry, 44, 1248–1263.
https://doi.org/10.1016/S0006-3223(98)00275-3, PubMed:
9861468
Langeslag, S. J., Gootjes, L., & van Strien, J. W. (2018). The
effect of mouth opening in emotional faces on subjective
experience and the early posterior negativity amplitude.
Brain and Cognition, 127, 51–59. https://doi.org/10.1016/j
.bandc.2018.10.003, PubMed: 30316954
Liu, X., Hairston, J., Schrier, M., & Fan, J. (2011). Common and
distinct networks underlying reward valence and processing
stages: A meta-analysis of functional neuroimaging studies.
Neuroscience & Biobehavioral Reviews, 35, 1219–1236.
https://doi.org/10.1016/j.neubiorev.2010.12.012, PubMed:
21185861
Löw, A., Bradley, M. M., & Lang, P. J. (2013). Perceptual
processing of natural scenes at rapid rates: Effects of
complexity, content, and emotional arousal. Cognitive,
Affective, & Behavioral Neuroscience, 13, 860–868. https://
doi.org/10.3758/s13415-013-0179-1, PubMed: 23780520
Lumley, T. (2017). Leaps: Regression subset selection. R package
version 3.0. https://CRAN.R-project.org/package=leaps
Maliniak, D., Powers, R., & Walter, B. F. (2013). The gender
citation gap in international relations. International
Organization, 67, 889–922. https://doi.org/10.1017
/S0020818313000209
Mitchell, S. M., Lange, S., & Brus, H. (2013). Gendered citation
patterns in international relations journals. International
Studies Perspectives, 14, 485–492. https://doi.org/10.1111
/insp.12026
Moratti, S., Saugar, C., & Strange, B. A. (2011). Prefrontal-
occipitoparietal coupling underlies late latency human
neuronal responses to emotion. Journal of Neuroscience,
31, 17278–17286. https://doi.org/10.1523/JNEUROSCI.2917
-11.2011, PubMed: 22114294
Nordström, H., & Wiens, S. (2012). Emotional event-related
potentials are larger to figures than scenes but are similarly
reduced by inattention. BMC Neuroscience, 13, 49.
https://doi.org/10.1186/1471-2202-13-49, PubMed: 22607397
Öhman, A., & Mineka, S. (2001). Fears, phobias, and
preparedness: Toward an evolved module of fear and fear
learning. Psychological Review, 108, 483. https://doi.org/10
.1037//0033-295X.108.3.483, PubMed: 11488376
Olofsson, J. K., Nordin, S., Sequeira, H., & Polich, J. (2008).
Affective picture processing: An integrative review of ERP
findings. Biological Psychology, 77, 247–265. https://doi.org
/10.1016/j.biopsycho.2007.11.006, PubMed: 18164800
Pastor, M. C., Bradley, M. M., Löw, A., Versace, F., Moltó, J., &
Lang, P. J. (2008). Affective picture perception: Emotion,
Farkas and Sabatinelli
955
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
context, and the late positive potential. Brain Research,
1189, 145–151. https://doi.org/10.1016/j.brainres.2007.10.072,
PubMed: 18068150
Peelen, M. V., & Downing, P. E. (2007). The neural basis of
visual body perception. Nature Reviews Neuroscience, 8,
636–648. https://doi.org/10.1038/nrn2195, PubMed: 17643089
Peyk, P., De Cesarei, A., & Junghöfer, M. (2011).
ElectroMagnetoEncephalography software: Overview and
integration with other EEG/MEG toolboxes. Computational
Intelligence and Neuroscience, 2011, 861705. https://doi.org
/10.1155/2011/861705, PubMed: 21577273
Sabatinelli, D., Keil, A., Frank, D. W., & Lang, P. J. (2013).
Emotional perception: Correspondence of early and late
event-related potentials with cortical and subcortical
functional MRI. Biological Psychology, 92, 513–519. https://
doi.org/10.1016/j.biopsycho.2012.04.005, PubMed: 22560889
Schindler, S., & Kissler, J. (2016). Selective visual attention to
emotional words: Early parallel frontal and visual activations
followed by interactive effects in visual cortex. Human Brain
Mapping, 37, 3575–3587. https://doi.org/10.1002/hbm.23261,
PubMed: 27218232
Schupp, H. T., Junghöfer, M., Weike, A. I., & Hamm, A. O.
(2004). The selective processing of briefly presented affective
pictures: An ERP analysis. Psychophysiology, 41, 441–449.
https://doi.org/10.1111/j.1469-8986.2004.00174.x, PubMed:
15102130
Schupp, H. T., Öhman, A., Junghöfer, M., Weike, A. I.,
Stockburger, J., & Hamm, A. O. (2004). The facilitated
processing of threatening faces: An ERP analysis. Emotion,
4, 189–200. https://doi.org/10.1037/1528-3542.4.2.189,
PubMed: 15222855
Schupp, H. T., Flaisch, T., Stockburger, J., & Junghöfer, M.
(2006). Emotion and attention: Event-related brain potential
studies. Progress in Brain Research, 156, 31–51. https://doi
.org/10.1016/S0079-6123(06)56002-9, PubMed: 17015073
Schupp, H. T., Stockburger, J., Codispoti, M., Junghöfer, M.,
Weike, A. I., & Hamm, A. O. (2006). Stimulus novelty and
emotion perception: The near absence of habituation in the
visual cortex. NeuroReport, 17, 365–369. https://doi.org/10
.1097/01.wnr.0000203355.88061.c6, PubMed: 16514360
Schupp, H. T., Stockburger, J., Codispoti, M., Junghöfer, M.,
Weike, A. I., & Hamm, A. O. (2007). Selective visual
attention to emotion. Journal of Neuroscience, 27,
1082–1089. https://doi.org/10.1523/JNEUROSCI.3223-06
.2007, PubMed: 17267562
Soares, S. C., Lindstrom, B., Esteves, F., & Öhman, A. (2014).
The hidden snake in the grass: Superior detection of snakes
in challenging attentional conditions. PLoS One, 9, e114724.
https://doi.org/10.1371/journal.pone.0114724, PubMed:
25493937
Thierry, G., Pegna, A. J., Dodds, C., Roberts, M., Basan, S., &
Downing, P. (2006). An event-related potential component
sensitive to images of the human body. Neuroimage, 32,
871–879. https://doi.org/10.1016/j.neuroimage.2006.03.060,
PubMed: 16750639
Thigpen, N. N., Keil, A., & Freund, A. M. (2018). Responding to
emotional scenes: Effects of response outcome and picture
repetition on reaction times and the late positive potential.
Cognition and Emotion, 32, 24–36. https://doi.org/10.1080
/02699931.2016.1266305, PubMed: 27922339
Ullman, S., Vidal-Naquet, M., & Sali, E. (2002). Visual features
of intermediate complexity and their use in classification.
Nature Neuroscience, 5, 682–687. https://doi.org/10.1038
/nn870, PubMed: 12055634
Van Strien, J. W., Eijlers, R., Franken, I. H. A., & Huijding, J.
(2014). Snake pictures draw more early attention than
spider pictures in non-phobic women: Evidence from
event-related brain potentials. Biological Psychology, 96,
150–157. https://doi.org/10.1016/j.biopsycho.2013.12.014,
PubMed: 24374241
Van Strien, J. W., Franken, I. H., & Huijding, J. (2014). Testing
the snake-detection hypothesis: Larger early posterior
negativity in humans to pictures of snakes than to pictures
of other reptiles, spiders and slugs. Frontiers in Human
Neuroscience, 8, 691. https://doi.org/10.3389/fnhum.2014
.00691, PubMed: 25237303
Weinberg, A., & Hajcak, G. (2010). Beyond good and evil: The
time-course of neural activity elicited by specific picture
content. Emotion, 10, 767–782. https://doi.org/10.1037
/a0020242, PubMed: 21058848
Willenbockel, V., Sadr, J., Fiset, D., Horne, G. O., Gosselin, F., &
Tanaka, J. W. (2010). Controlling low-level image properties:
The SHINE toolbox. Behavior Research Methods, 42,
671–684. https://doi.org/10.3758/BRM.42.3.671, PubMed:
20805589
Yoon, S., Shim, M., Kim, H. S., & Lee, S. H. (2016).
Enhanced early posterior negativity to fearful faces in
patients with anxiety disorder. Brain Topography, 29,
262–272. https://doi.org/10.1007/s10548-015-0456-0,
PubMed: 26476635
956
Journal of Cognitive Neuroscience
Volume 35, Number 6
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
5
6
9
4
1
2
0
8
2
9
2
5
/
j
o
c
n
_
a
_
0
1
9
8
4
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3