Dissociation of Frontal and Medial Temporal
Lobe Activity in Maintenance and Binding of
Sequentially Presented Paired Associates
Jena B. Hales, Sarah L. Israel, Nicole C. Swann, and James B. Brewer
D
o
w
N
l
o
UN
D
e
D
Astratto
lobe are critical
& Substructures of
the prefrontal cortex (PFC) and the
medial-temporal
for associating objects
presented over time. Previous studies showing frontal and
medial-temporal involvement in associative encoding have not
addressed the response specificity of these regions to different
aspects of the task, which include instructions to associate and
binding of stimuli. This study used a novel paradigm to
temporally separate these two components of the task by
sequential presentation of individual images with or without
fMRI was used to investigate the
associative instruction;
temporal involvement of the PFC and the parahippocampal
cortex in encoding each component. Although both regions
showed an enhanced response to the second stimulus of a
pair, only the PFC had increased activation during the delay
preceding a stimulus when associative instruction was given.
These findings present new evidence that prefrontal and
medial-temporal regions provide distinct temporal contribu-
tions during associative memory formation. &
INTRODUCTION
Animal lesion models and studies involving patients with
selective damage to structures of the medial-temporal
lobe (MTL) have demonstrated critical involvement of
this brain region in the encoding and retrieval of long-
term declarative memory, the memory for facts and
events (Squire, 1992). Multiple studies have demonstrat-
ed particular involvement of the parahippocampal cortex
(PHC) in successful memory formation (Eichenbaum,
Yonelinas, & Ranganath, 2007; Murray & Ranganath, 2007;
Gold et al., 2006; Kirwan & Stark, 2004; Davachi, Mitchell,
& Wagner, 2003; Davachi & Wagner, 2002; Brewer, Zhao,
Desmond, Glover, & Gabrieli, 1998; Henke, Buck, Weber,
& Wieser, 1997). Neuroimaging and neuropsychological
studies of patients with damage to the prefrontal cortex
(PFC) have also suggested the contribution of the PFC to
the encoding of long-term memory (LTM) (Blumenfeld &
Ranganath, 2007; Murray & Ranganath, 2007; Sperling
et al., 2003; Brewer et al., 1998; Wagner et al., 1998).
Although imaging studies have commonly reported PFC
and PHC activity in lockstep during associative encoding,
the hypothesis of the present study was that activity in
these two regions is dissociable, with PFC activity preced-
ing PHC activity, supporting a mechanism for top–down
modulation of MTL structures involved in associative
encoding.
Anatomical studies using anterograde and retrograde
tracing techniques in monkeys (Goldman-Rakic, Selemon,
University of California, San Diego
& Schwartz, 1984) and imaging methods combining
functional magnetic resonance imaging (fMRI) and diffu-
sion tensor imaging (DTI) in humans (Takahashi, Ohki, &
Kim, 2007) examined the connectivity between the PFC
and the PHC. Both studies reported direct and indirect
anatomical connections between subregions of the PFC
and PHC. Recentemente, Gazzaley and D’Esposito (2007)
examined the process of top–down modulation from
the PFC to the visual association cortex and the PHC
during scene-selective processing. The top–down influ-
ence of prefrontal activity upon parahippocampal activity
is consistent with the anatomical connectivity between
these brain regions.
A recent fMRI study examined PFC activity using an
associative memory paradigm in which two sequentially
presented words were associated when the presenta-
tion of the second word was accompanied by a relation-
al question and not associated when accompanied by
an item-specific question (Murray & Ranganath, 2007).
Greater activation of the left PHC, the dorsolateral pre-
frontal cortex (DLPFC), and the ventrolateral prefrontal
cortex (VLPFC) was observed during the encoding of
the second presented word in relational trials compared
to item-specific trials. As associative instructions were
presented concurrently with the second word, increased
activation could only be examined at that time point,
and functional specificity of PFC and MTL activity could
not be addressed. The present study, Tuttavia, was de-
signed to pursue this question of the particular involve-
ment of these two brain regions in processes recruited
for associative encoding.
D 2008 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 21:7, pag. 1244–1254
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
7
e
–
1
P
2
D
4
F
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
C
4
N
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
9
0
6
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
/
T
.
.
F
o
N
1
8
M
UN
sì
2
0
2
1
In nonhuman primates, multiunit recording data sug-
gest that PFC neurons play a role in associating tempo-
rally separate stimuli and show delay-period increases in
activity (Deco, Ledberg, Almeida, & Fuster, 2005; Fuster,
Bodner, & Kroger, 2000). Fuster et al. (2000) conducted
extracellular recordings from bilateral regions of the
dorsolateral frontal cortex while monkeys performed
an audiovisual memory task. As monkeys learned the
low tone–green and high tone–red associations, cells in
this region showed the same relationship of firing to low
and high tones as to green and red colors, rispettivamente,
with maintained activity during the delay between tones
and associated colors. The presence of delay-period ac-
tivity in medial-temporal regions, Tuttavia, has been
more controversial. Some studies report rarely seen in-
creases in delay-period activity in medial-temporal re-
gions such as the PHC in monkeys (Vidyasagar, Salzmann,
& Creutzfeldt, 1991) or the hippocampus in rats early in
the delay period (Hampson & Deadwyler, 2003), Dove-
as other studies in monkeys report the presence of
delay-period activity in medial-temporal regions ( Young,
Otto, Fox, & Eichenbaum, 1997; Cahusac, Miyashita, &
Rolls, 1989; Watanabe & Niki, 1985). Despite the dis-
agreement in the literature regarding MTL activity during
the delay period in associative tasks, there is strong elec-
trophysiological evidence for PFC activity during the
delay period in rats and monkeys.
The present study further examines the involvement
of the PFC and the PHC in the encoding of associative
memory compared to single-item memory. Rapid event-
related fMRI was used to identify the temporal involve-
ment of the PFC and the PHC in encoding sequentially
presented images with varying interstimulus intervals
(ISIs). A plus-sign presented during some ISIs instructed
participants to associate the image preceding and fol-
lowing the plus-sign as a pair. The timing separation
between the plus-sign (instructing the subject to pair
the previous image with the upcoming image) and the
presentation of the second image (at which point the
images can be associated) allowed temporal investiga-
tion of PFC and PHC involvement in associative memory
encoding. After the scan, participants completed a rec-
ognition test examining associative and single-item mem-
ory. Based on previous findings, the hypotheses were
that the PFC and the PHC would show greater activa-
tion during the encoding of paired versus unpaired
images. Prefrontal activity was expected to precede para-
hippocampal activity supporting top–down influence on
the PHC.
METHODS
Participants
Thirteen healthy volunteers (mean age = 23.69, 3 men)
recruited from the University of California—San Diego
(UCSD) community and the surrounding area were
enrolled in this study. Participants gave informed con-
sent approved by the UCSD Institutional Review Board
and had normal or corrected-to-normal vision. Twelve
additional volunteers (mean age = 25.08, 6 men) were
recruited for a behavioral pilot task.
Stimuli
Stimuli included 256 color images of common objects
which were presented individually while the participant
was in the scanner. A plus-sign appeared between some
of the stimuli. An additional 40 novel stimuli were used
during the recognition test following the scan. Images
were acquired from Rossion and Pourtois (2004) colore
Snodgrass images (www.nefy.ucl.ac.be/facecatlab/stimuli.
htm) and Hemera object library (Hemera Technologies;
Quebec, Canada).
Experimental Procedure
While in the scanner, participants were presented with
individual
images (each remaining on the screen for
2.5 sec) followed by jittered ISIs ranging from 0.5 A
11 sec (Figure 1A). Jitter was calculated to optimize the
progetto (Dale, 1999; Dale & Buckner, 1997). Immediately
following some of the images, a plus-sign appeared in
the center of the screen for 0.5 sec. Participants were
asked to remember the presented images and,
if an
image was followed by a plus-sign, to associate the image
with the subsequent image as a pair. Participants were
given a button box and were asked to press one button
if the image represented a living object and the other
button if the image represented a nonliving object. Im-
age stimuli were presented in a series of four runs, each
lasting 362 sec and containing 64 images. Over all four
runs, 130 images were included in associated pairs and
126 images were unpaired. The presentation of stimuli
varied pseudorandomly between paired and unpaired
stimuli. For analysis purposes, but unannounced to the
participants, paired and unpaired items were presented
sequentially in multiples of two. For simplicity, stimuli
preceding a plus-sign will be denoted as ‘‘1P,’’ and the
stimuli following the plus-sign as ‘‘2P.’’ After a ‘‘2P’’
stimulus, the next image could be a ‘‘1P’’ (which would
then be followed by a plus-sign and a ‘‘2P’’), or the next
image could be an individual unpaired stimulus, Di-
noted ‘‘1U’’ for unpaired.
‘‘1U’’ was always followed
by ‘‘2U’’. This terminology is used in Figure 1 E
throughout the analysis.
Following the scan, participants completed a recogni-
tion test (Figure 1B). Participants were shown an image
and were asked to rate how well they remembered
seeing that image during the scanner presentation, 1
being ‘‘poorly’’ and 5 being ‘‘very well.’’ This question
was asked for each of the 256 images that the participant
was shown while in the scanner plus 40 additional novel
images. After rating each image, participants were shown
Hales et al.
1245
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
7
e
–
1
P
2
D
4
F
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
C
4
N
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
9
0
6
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
T
.
/
.
.
F
o
N
1
8
M
UN
sì
2
0
2
1
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
7
e
–
1
P
2
D
4
F
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
C
4
N
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
9
0
6
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
T
.
F
/
.
.
o
N
1
8
M
UN
sì
2
0
2
1
Figura 1. Experimental design. (UN) Schematic depiction of the scanner presentation of two paired and two unpaired stimuli. For the first 0.5 sec
of a 0.5- to 11-sec ISI, the associative memory instruction of a plus-sign is present between two images that should be paired (1P and 2P) E
is not present between two images that should remain unpaired (1U and 2U). (B) Schematic depiction of the recognition test conducted following
the scan. Participants were asked if they remember seeing the image in the scanner (‘‘poorly’’ if they think it is a novel item; ‘‘very well’’ if
they remember seeing the item). If the image was presented in the scanner, participants were then shown a second screen with two choice
images; they were asked to report which image (1 O 2) was the associated pair if the target image was paired or to report if the target
image was unpaired (3).
two additional images, labeled ‘‘1’’ and ‘‘2,’’ and were
instructed to identify the pair of the originally presented
image or to identify the original
image as unpaired
(option labeled ‘‘3’’). If the original image was novel,
this question was skipped all-together and the next
recognition image was presented. The postscan recog-
nition test lasted approximately 30 min.
fMRI Parameters
Participants were scanned using a 3-T GE scanner at the
Keck Center for Functional MRI at the University of
California, San Diego. Functional images were acquired
using a gradient-echo, echo-planar, T2*-weighted pulse
sequence (repetition time = 1.5 sec; one shot per rep-
etition; echo time = 30; flip angle = 908; bandwidth =
31.25 MHz). Twenty-two slices covering the brain were
obtained perpendicular to the long axis of the hippo-
campus with 4 (cid:1) 4 (cid:1) 7 mm voxels. T1-weighted
structural scans were acquired in the same plane as
the functional scans and of the same voxel size. Struc-
tural images were also acquired using high resolution
T1-weighted (1 (cid:1) 1 (cid:1) 1 mm) magnetization-prepared
rapid gradient-echo sequence.
Data Analysis
Data from each run were reconstructed using the AFNI
(Cox, 1996) suite of programs. Slices were aligned tem-
porally and then coregistered using a three-dimensional
image alignment algorithm. A threshold mask of the
functional data was used to eliminate voxels outside the
brain. A series of functional images from separate runs
were corrected for motion and concatenated. Two
general linear models were constructed using multiple
included six motion
regression analysis. Each model
regressors obtained from the registration process and
additional task-related regressors in which impulse re-
sponses were modeled from the data for each of the
stimulus conditions. The first general linear model in-
cluded regressors for 1P, 2P, 1U, 2U condition correct
linear
and incorrect responses. The second general
model included regressors for paired trials (1P and 2P
with an ISI of 3.5 sec) and unpaired trials (1U and 2U
with an ISI of 3.5 sec) (Daselaar et al., 2007; Schluppeck,
Curtis, Glimcher, & Heeger, 2006). An ISI of 3.5 sec was
selected because it was the most frequent jitter interval
and allowed sufficient measurements for analysis. In
aggiunta, parameter estimates for all delay periods be-
tween two paired images with remembered associative
properties were analyzed relative to all delay periods
between two unpaired images using repeated measures
ANOVA.
Only paired and individual unpaired images correctly
identified during the postscan recognition test were
included in the analysis of fMRI data. The hemodynamic
response function was derived from the fMRI data using
signal deconvolution and a defined time window follow-
ing stimulus onset (AFNI Software; Cox, 1996). This time
window was from 0 A 15 sec for single stimulus events,
E 0 A 21 sec for two-stimulus trials with 3.5-sec
ISIs. Standard landmarks were defined manually on the
anatomical scans. Data from the anatomical and func-
tional scans were then transformed into Talairach and
Tournoux (1998) space by AFNI using nearest-neighbor
interpolation. No spatial smoothing was performed. IL
areas under the hemodynamic response function for
the following conditions were examined using voxelwise
t tests (two-tailed) carried out across all 13 participants:
(1) 2P versus 2U, (2) trials with two paired images with
an ISI of 3.5 sec (with a plus-sign present for the first
0.5 sec of the ISI) versus trials with two unpaired images
1246
Journal of Cognitive Neuroscience
Volume 21, Numero 7
with an ISI of 3.5 sec. Given the reduced number of trials
with an ISI of 3.5 sec, all trials were included in this
analysis. A voxelwise threshold of p < .01 was used to
identify significant regional activity. Analyses were re-
stricted to clusters containing at least four voxels con-
nected by face surfaces, yielding a significance value of
p < .01 when corrected for multiple comparisons across
the whole brain. These clusters were used to create
impulse–response plots displaying the temporal charac-
teristics of the activation.
RESULTS
Behavioral Pilot Task
A behavioral pilot task was conducted to evaluate
whether an instructional cue can effectively manipulate
episodic associative memory for items presented se-
quentially and to ensure that incidental associations
are not being made between proximally presented
unpaired images. Following the encoding task, which
was the same as was used for the imaging study,
participants completed a postscan recognition task sim-
ilar to that used in the current experiment, except for
that they were asked which of two images was presented
closest in time to the image they just saw during the
previous item memory question. Participants identified
the item presented adjacent in time when no associative
cue had been presented at a low rate (60 ± 4%),
significantly below their performance in identifying the
item presented adjacent in time when an associative cue
had been presented (81 ± 6%; p < .001, t = 6.623).
Behavioral Analysis
Eighty-two percent (±3%) of paired stimuli were recog-
nized with a high degree of confidence (subject re-
sponse of 4 or 5), and for those recognized items, the
correct associated pair was identified at a rate of 71%
(±4%). Unpaired items were recognized with a high
degree of confidence at a rate of 73% (±4%). Data for
correctly identified paired and unpaired items were in-
cluded in the fMRI analysis. Subjects incorrectly identi-
fied novel images as recognized at a rate of 11% (±3%).
fMRI Analysis
Based on previous studies that have found activation in
the MTL structures as well as in regions of the PFC
during the encoding of associated items (Dickerson
et al., 2007; Murray & Ranganath, 2007; Tendolkar et al.,
2007; Gold et al., 2006; Staresina & Davachi, 2006; Law
et al., 2005; Meltzer & Constable, 2005; Prince, Daselaar,
& Cabeza, 2005; Pihlajamaki et al., 2003; Sperling et al.,
2003; Yonelinas, Hopfinger, Buonocore, Kroll, & Baynes,
2001; Dolan & Fletcher, 1997; Henke et al., 1997; Rombouts
et al., 1997), analysis was focused on these brain regions.
For the present study, parahippocampal regions were
predicted to show greater activation during the encoding
of 2P stimuli (items paired with the preceding stimulus)
than during the encoding of 2U stimuli (items not paired
with the preceding stimulus and which only follow un-
paired stimuli). Prefrontal regions, specifically the DLPFC
and the VLPFC, were also predicted to show increased
activation during the encoding of 2P stimuli.
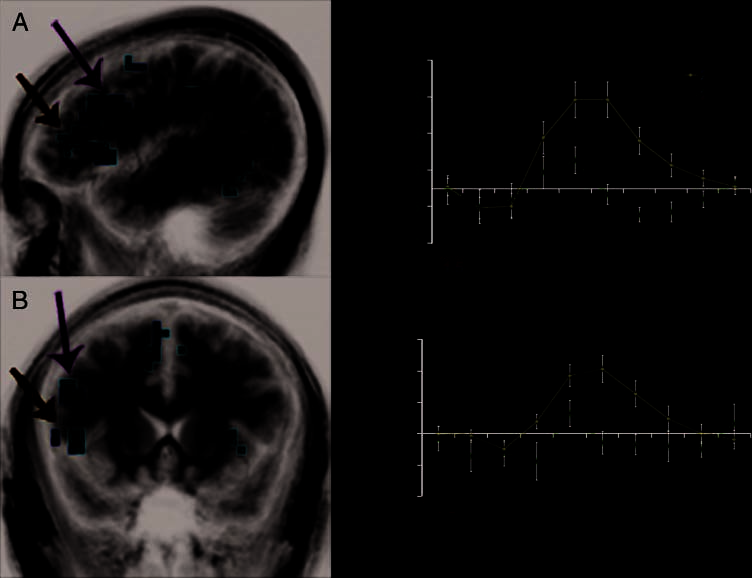
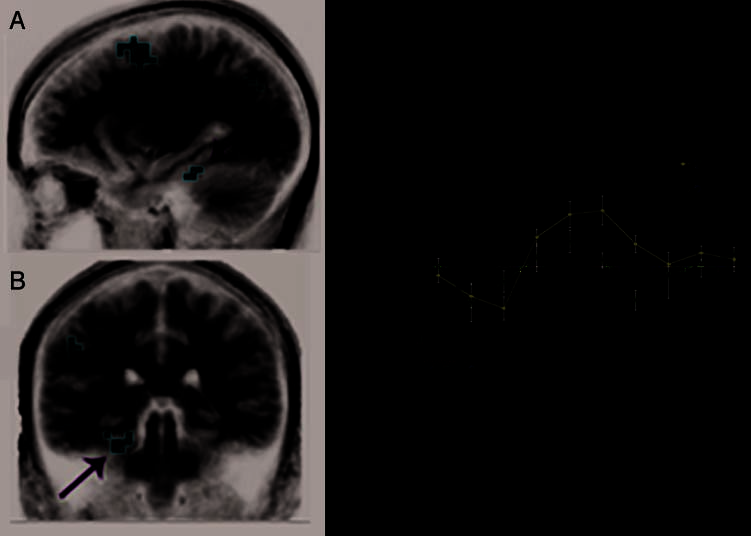
Activation during the viewing of 2P stimuli was com-
pared to activation during the viewing of 2U stimuli. This
contrast between processing 2P versus 2U stimuli re-
vealed left PHC activation ( p < .01; Figure 2A, B). The
impulse–response curve for 2U indicated PHC activity
during single-item encoding as well as during associative
encoding; however, the activity in this region was great-
er during the encoding of 2P stimuli (Figure 2C).
Previous studies reported that the DLPFC (Brodmann’s
area 9, 46) is active during encoding of individual items
(Staresina & Davachi, 2006; Brewer et al., 1998), and
such activity is further increased by associative memory
conditions (Murray & Ranganath, 2007). Consistent with
these findings, greater activation of the DLPFC during
encoding of 2P stimuli relative to the encoding of 2U
stimuli was observed ( p < .01; Figure 3A, B). Similar to
PHC involvement, the DLPFC was active during the
encoding of all remembered images; however, the acti-
vation was greatly enhanced during associative encoding
of 2P (Figure 3C). Activity in the VLPFC was also analyzed
for this contrast between the encoding of 2P stimuli and
2U stimuli. Similar to activity in the DLPFC, there was an
increase in activation in the VLPFC (Brodmann’s area 44,
45, 47) during the encoding of 2P stimuli ( p < .01;
Figure 3A, B). However, the VLPFC did not show signif-
icant activity for 2U stimuli ( p > .05). Examination of the
time course of activity in the VLPFC also showed a small
response for 2U stimuli that did not reach significance
(Figure 3D). A complete list of regions of activation for
this contrast is listed in Table 1.
Analyses were also performed comparing activity dur-
ing the encoding of trials with two paired images versus
trials with two unpaired images, allowing for the exam-
ination of activity differences during the ISI. The time
course of activity was examined beginning with the
presentation of the first paired (1P) or unpaired (1U)
image followed by a 3.5-sec ISI and the presentation of
the second image (cioè., 2P or 2U, rispettivamente). Although
ISIs varied between 0.5 E 11 sec due to jitter, trials
with 3.5-sec ISIs were used for this comparison. IL
same functional regions of interest as were previously
discussed were also predicted to be important in this
contrasto.
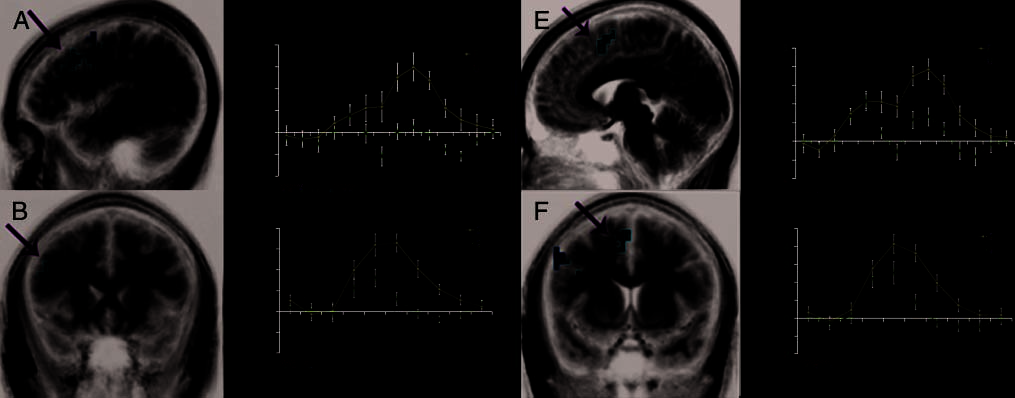
The comparison between the encoding of two paired
stimuli and two unpaired stimuli showed increased ac-
tivity in two frontal regions, the left DLPFC (Figure 4A,
B) and the left medial frontal cortex (Figure 4E, F)
during the paired trials ( P < .01). The time course of
activity in the left DLPFC showed a similar response for
Hales et al.
1247
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
7
e
-
1
p
2
d
4
f
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
c
4
n
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
f
.
/
t
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Increased activity
in the left PHC during the
encoding of the second
paired (2P) stimuli versus
the second unpaired (2U)
stimuli. Statistical activation
maps illustrating greater
activation ( p < .01) during
the encoding of 2P versus 2U
stimuli are superimposed on
sagittal (A) and coronal (B)
slices of mean anatomical scan
images across all 13 subjects;
arrows indicate the left PHC
cluster used for time-course
analysis. (C) Time course
of activity in the left PHC
beginning with the onset of
2P stimuli (blue) and 2U
stimuli (pink) demonstrates
activity during item encoding,
with increased activity during
associative encoding. The
time of stimulus presentation
is represented by the light
blue block. The y-axis
represents percent signal
change, the x-axis is time in
seconds (sec), and the error
bars represent the standard
error of the mean.
Figure 3. Increased activity
in the left DLPFC and the
left VLPFC during the encoding
of the second paired (2P)
stimuli versus the second
unpaired (2U) stimuli.
Statistical activation maps
illustrating greater activation
( p < .01) during the encoding
of 2P versus 2U stimuli are
superimposed on sagittal
(A) and coronal (B) slices of
the mean anatomical scan
images across all 13 subjects;
arrows indicate left DLPFC
(green) and left VLPFC (blue)
clusters used for time-course
analysis. (C) Time course of
activity in the left DLPFC
beginning with the onset of
2P stimuli (blue) and 2U
stimuli (pink) demonstrating
activity during item encoding,
with increased activity during
associative encoding. The time
of stimulus presentation is
represented by the light blue
block. (D) Time course of
activity in the left VLPFC for
the same comparison
demonstrates activity only
during associative encoding,
with no significant response
during the encoding of
individual items.
1248
Journal of Cognitive Neuroscience
Volume 21, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
7
e
-
1
p
2
d
4
f
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
c
4
n
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
.
f
.
/
o
n
1
8
M
a
y
2
0
2
1
Table 1. Significantly Active Brain Regions for Paired Stimuli
versus Unpaired Stimuli (2P vs. 2U)
# Volume
x
y
z
t
3904
2816
2624
(cid:2)46
(cid:2)46.6
(cid:2)4.5
14.4
22.1
11.3
25.5
5
52.4
5.54
5.49
6.65
L DLPFC (BA 9/46)
L VLPFC (BA 45)
L Superior frontal
(BA 6)
L Middle frontal
2176
(cid:2)25.6
0.2
51.6
7.18
(BA 6)
L Angular (BA 39)
1344
(cid:2)28.6 (cid:2)60.3
32.2
L Parahippocampal
896
(cid:2)26.3 (cid:2)35.3 (cid:2)14.8
4.60
5.55
(BA 36)
L Middle occipital
640
(cid:2)48.9 (cid:2)57.9 (cid:2)3
4.12
(BA 19)
R Cingulate (BA 31)
L Middle temporal
(BA 21)
R Cerebellum
R Supramarginal
(BA 40)
640
576
512
512
26.4 (cid:2)48.8
25.1
(cid:2)53 (cid:2)30.8 (cid:2)6.1
4.11
5.93
29.7 (cid:2)51.7 (cid:2)27.9
5.12
51.1 (cid:2)47.3
33.4 (cid:2)3.95
The left medial frontal cortex showed a similar in-
crease in activity following the plus-sign in the paired
trials (Figure 4E, F). Much like the left DLPFC, the
response curve for the left medial frontal cortex showed
a matched response for 1P and 1U, with divergence
occurring at the instruction to associate and a further
increase in response during the presentation of 2P
(Figure 4G). Figure 4H illustrates the larger left medial
frontal response during the encoding of 2P than during
2U (for a complete list of regions of activation for this
contrast, see Table 2).
No significant clusters were identified in the PHC using
the comparison of the above subset of trials containing
two paired or two unpaired stimuli with 3.5-sec ISIs
( p > .05). Inoltre, when ISIs spanning all delay
periods were analyzed, the left DLPFC showed a signif-
icant increase in activity during the delay period be-
tween two paired images ( P < .05, t = 2.195). However,
there was no significant difference in activity during the
delay periods between paired images and between un-
paired images in the left PHC ( p = .51, t = 0.673). An
interaction analysis between these two delay-period con-
ditions for each brain region showed a significant Re-
gion (cid:1) Condition interaction ( p < .05).
L Inferior temporal
448
(cid:2)50.3 (cid:2)51.7 (cid:2)13.6
4.90
(BA 20)
L Inferior parietal
448
(cid:2)48.5 (cid:2)32.1
35.5
5.55
DISCUSSION
(BA 40)
R Insula (BA 47)
L Inferior parietal
(BA 40)
L Caudate
R Supramarginal
(BA 40)
384
320
256
256
31.4
17.3
2.5
(cid:2)42 (cid:2)49.6
45.9
4.03
3.68
(cid:2)14
10.2
4.1
3.25
54 (cid:2)51.1
20.2 (cid:2)4.18
L Supramarginal
256
(cid:2)41 (cid:2)43
35.2
3.92
(BA 40)
L Precentral (BA 6)
256
(cid:2)46
(cid:2)0.7
48.8
4.74
1P and 1U (4.5–7.5 sec). In the unpaired trial, the
activation decreased during the ISI and then increased
during the presentation of 2U with a time course similar
to that of 1U. In contrast, the paired trial showed
sustained DLPFC activity throughout the ISI and rising
further with the onset of 2P (Figure 4C). The time
course of activity during paired and unpaired trials
diverged at 7.5 sec, corresponding to the instruction to
associate the 1P stimulus with the following stimulus.
The larger activation during the encoding of 2P relative
to that of 2U, seen in Figure 4C, was the result of
increased size of response in addition to the increase
in baseline (revealed when all
jittered ISI trials are
analyzed with separate covariates for paired and un-
paired ISIs modeled as sustained responses; Figure 4D).
The present study is the first to examine temporal
contributions of the PFC and the PHC in associative
memory encoding by separating the associative instruc-
tion from the time at which binding may occur. Activity
in the PFC and the PHC was analyzed while subjects
were instructed to encode sequentially presented stim-
uli as paired or as separate items. Contrasts between the
encoding of 2P and 2U stimuli and the encoding of
paired and unpaired trials with a 3.5-sec ISI were exam-
ined. The left PHC and DLPFC were active for all
correctly encoded stimuli, with increased activity during
2P encoding versus 2U encoding. In contrast, the left
VLPFC was significantly active during 2P, but not during
2U encoding.
Declarative Memory Encoding with
Associative Instruction
In the present study, participants were instructed only
to associate two stimuli when a plus-sign intervened; all
other stimuli were to be remembered as single items.
Although it is possible that associations can develop
between sequentially presented images with or without
associative instruction, episodic associative memory was
improved by the presence of the cue. In addition, sub-
sequent recognition of individual stimuli was improved
by the presence of the cue (paired items recognized at
a rate of 82%, and unpaired items recognized at a rate
of 73%; p < .01). Thus, the instruction to associate
Hales et al.
1249
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
7
e
-
1
p
2
d
4
f
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
c
4
n
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
.
.
t
/
o
n
1
8
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
7
e
-
1
p
2
d
4
f
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
c
4
n
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
f
/
.
.
Figure 4. Initiation of activity increase in the left DLPFC and the left medial frontal cortex at onset of associative memory instruction. Statistical
activation maps illustrating greater activation ( p < .01) during the encoding of two paired images (with a 3.5-sec ISI) versus two unpaired images
(with a 3.5-sec ISI) are superimposed on sagittal (A, E) and coronal (B, F) slices of mean anatomical scan images across all 13 subjects; arrows
indicate the left DLPFC (A, B) and left medial frontal (E, F) clusters used for time-course analysis. (C, G) Time courses of activity in the left
DLPFC (C) and the left medial frontal cortex (G) beginning with the onset of the first image of two paired images (blue) and the first image of
two unpaired images (pink) demonstrate divergence at the onset of associative instruction. The time of stimulus presentation is represented
by the light blue block, and the time of associative instruction presentation is represented by the green block. (D, H ) Time courses of activity
in the left DLPFC (D) and the left medial frontal cortex (H) during the presentation of 2P (blue) and 2U (pink) stimuli illustrate the enhanced
response to the second stimulus in the associated condition. The time of stimulus presentation is represented by the light blue block.
modulates episodic memory performance along with its
enhancement of brain activity.
Using sequential presentation of single images, se-
mantic information was balanced across stimuli. How-
ever, the instruction to associate may engage verbal
processes when nameable stimuli are used. It is possible
that using nonverbal stimuli could result in different
patterns or degrees of left frontal lobe activation. The
left lateralization reported in the present study with
nameable stimuli
is, on one hand, similar to that re-
ported in other encoding studies using verbal stimuli
(Blumenfeld & Ranganath, 2007; Murray & Ranganath,
2007; Sperling et al., 2003; Wagner et al., 1998). On the
other hand, the data are also in agreement with the
revised Hemispheric Encoding/Retrieval Asymmetry
model (Habib, Nyberg, & Tulving, 2003), which would
predict left-sided activation for encoding regardless of
stimulus type. The paradigm presented here could be
adapted to address such questions through the use of
nonverbal stimuli.
Increased PHC Activity during
Associative Encoding
The involvement of particular MTL substructures in
various aspects of long-term memory is debated in the
literature (Eichenbaum et al., 2007). The present study
showed PHC activity during encoding of
individual
images and pairs of associated images; however, this
region showed selectivity through an increased response
during associative encoding relative to individual-item
encoding. These results complement other studies dem-
onstrating PHC involvement in item encoding with
enhanced activity during associative encoding (Murray
Table 2. Significantly Active Brain Regions for Two Paired
Stimuli (3.5-sec ISI) versus Two Unpaired Stimuli (3.5-sec ISI)
# Volume
x
y
z
t
4480
3584
1280
(cid:2)47.3
7.5
35.3
5.93
(cid:2)32.6 (cid:2)56.4
35.5
5.82
(cid:2)5.4
5.3
52.5
6.71
L DLPFC (BA 9)
L Angular (BA 39)
L Medial frontal
(BA 6)
L Middle temporal
704
(cid:2)47 (cid:2)46.6
5.8
4.50
(BA 21)
L Fusiform (BA 37)
R Cuneus (BA 19)
R Superior temporal
(BA 39)
R Cerebellum
L Middle frontal
(BA 6)
o
n
1
8
M
a
y
2
0
2
1
512
512
512
384
384
(cid:2)37.4 (cid:2)43.3 (cid:2)8.6
4.05
19 (cid:2)83.9
30.7 (cid:2)3.79
32.1 (cid:2)52.7
31.6
5.18
1 (cid:2)38.1 (cid:2)14.8
3.91
(cid:2)27.3 (cid:2)7.1
46.2
3.80
L Middle temporal
320
(cid:2)39.5 (cid:2)50.2
10.9
5.39
(BA 39)
L Superior frontal
320
(cid:2)31.6
48.8
17.7
3.68
(BA 10)
1250
Journal of Cognitive Neuroscience
Volume 21, Number 7
& Ranganath, 2007; Kirwan & Stark, 2004). Using a
different paradigm where three words were presented
concurrently under instructions to repeat the words
throughout the trial or to order the words according
to their desirability, different patterns of brain activity
were reported (Davachi & Wagner, 2002). Bilateral
hippocampus was active for both encoding tasks, where-
as the right entorhinal and bilateral parahippocampal
gyri were more active during the repetition task. Al-
though only the reordering task is described as using
relational processing, both tasks could involve associa-
tive encoding. The cognitive strategies adopted to per-
form each type of task, however, will differ. The current
study, which requires the association of two nameable
visual stimuli, involves a cognitive strategy that is per-
haps more similar to the repetition condition than to the
reorder condition of the previous study (Davachi &
Wagner, 2002). Therefore, the presence of parahippo-
campal activity in both the present study and in the
repetition task in the previous study could reflect a
common strategy.
PFC Activity and Dissociation of Substructures
Results from the current study showed increased DLPFC
and VLPFC activity during the encoding of stimuli under
associative conditions and revealed that enhancement
of DLPFC activity begins at associative memory instruc-
tion. The noted further increase in DLPFC and VLPFC
activity during 2P stimuli agrees with the present litera-
ture. The DLPFC is also active during the encoding of
unpaired stimuli, whereas the VLPFC does not signifi-
cantly respond to unpaired stimuli. This dissociation
differs from previous results examining regional speci-
ficity within the PFC.
Previous studies have examined dissociations between
regions of the PFC in relational and item-specific mem-
ory encoding (Murray & Ranganath, 2007; Blumenfeld &
Ranganath, 2006). In an fMRI study using pairs of se-
quentially presented words, the second word was ac-
companied by a question prompting the participant
to (1) relate the two words together (‘‘relational trial’’)
or (2) semantically evaluate the second word (‘‘item-
specific trial’’) (Murray & Ranganath, 2007). A dorsal–
ventral dissociation was reported in lateral PFC activation.
Both regions showed increased activation for encoding
relational words versus item-specific words. VLPFC ac-
tivity also predicted both successful relational and item-
specific encoding, whereas DLPFC activity only predicted
successful relational encoding. An earlier study exam-
ined the function of the DLPFC in long-term memory
formation using a paradigm where three words were
presented with the instruction either to rehearse the
words or to reorder them according to the weight of
the object (Blumenfeld & Ranganath, 2006). Based on
results showing increased DLPFC activity during the en-
coding of reorder trials relative to rehearse trials and for
the encoding of reorder trials where words were subse-
quently remembered, this study concluded that the
DLPFC is involved in encoding organizational informa-
tion. There are, however, several key differences between
the current study and previous studies examining subre-
gional contributions of the PFC to long-term memory.
The purpose of the present study was to examine the
contributions of the PFC and the PHC in the encoding of
pairs of associated images versus the encoding of un-
paired images. Differences in activation between remem-
bered compared to forgotten images were not the focus
of the current study, and will be a topic of future inves-
tigation. Only correctly encoded images, as determined
by the recognition task, were included in the analysis. A
design optimized to examine subsequent memory-related
activity might reveal different results. For example, ac-
tivity seen in the DLPFC for the encoding of unpaired
images might not differ based upon subsequent memory
performance. Such results would then support previous
findings of DLPFC activity predicting successful associa-
tive, and not individual-item, encoding.
Results from the current study show that the VLPFC
does not significantly respond to the encoding of sub-
sequently remembered unpaired images. These obser-
vations appear to differ from those of previous studies,
which report VLPFC involvement in successful encod-
ing of relational and item-specific memory (Murray &
Ranganath, 2007) and memory for word rehearsal and
reordered words (Blumenfeld & Ranganath, 2006). How-
ever, small differences in VLPFC cluster location may be
relevant. The location of DLPFC activity (BA 46, 9) in the
present study is very similar to the location of DLPFC
activity in the previous studies, but the peak location of
VLPFC activity (BA 45) is more anterior in the present
study. One study separated the VLPFC into two different
clusters, the anterior VLPFC (BA 47, 45), with a location
similar to the current study, and the posterior VLPFC
(6, 44), and although both clusters were predictive of
subsequent memory for reorder trials, only the posterior
cluster was predictive of subsequent memory for re-
hearse trials (Blumenfeld & Ranganath, 2006). Another
study that also reports VLPFC activation predictive of
subsequent memory for item-specific trials also de-
scribes a VLPFC cluster that appears more posterior
than the VLPFC cluster in the present study (Murray &
Ranganath, 2007).
The points discussed earlier in the discussion con-
cerning the differences between three-word reordering/
rehearsal paradigms and the present paradigm regarding
activation in the PHC are also relevant when discussing
dissociations in PFC activity. Rehearsing and reorder-
ing words may each involve associative memory, with
reordering implementing additional working memory
components. Rehearsal could establish a phonological
association, whereas reordering may create visual and
spatial associations. Although both types of trials may
involve associative memory formation, each may utilize
Hales et al.
1251
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
7
e
-
1
p
2
d
4
f
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
c
4
n
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
.
.
/
f
o
n
1
8
M
a
y
2
0
2
1
different organizational strategies resulting in differential
VLPFC activity. In contrast, the DLPFC has been shown
to be involved in task switching (Loose, Kaufmann, Tucha,
Auer, & Lange, 2006; Vanderhasselt, De Raedt, Baeken,
Leyman, & D’haenen, 2006; Smith, Taylor, Brammer, &
Rubia, 2004; Sylvester et al., 2003). The above studies
examining associative memory formation, as well as the
present study, require a switch in task as instructed by a
cue, which may contribute to the overlapping activity of
the DLPFC despite the differences in study design.
Top–Down Influence of the PFC on PHC Activity
The sequential presentation of stimuli and an interven-
ing plus-sign allowed for temporal separation of the
neural activity related to (1) instructions to associate
and (2) presentation of the second stimulus required to
form the association. Following the plus-sign, the left
DLPFC and the medial frontal cortex showed a sustained
increase in activation relative to ISIs without a plus-sign
(during which, activity in these regions returned to base-
line; Figure 4C, G). Left PHC activity was not significantly
different during the ISIs in paired and unpaired con-
ditions ( p = .51). These results suggest that the left
DLPFC and the left medial frontal cortex are involved in
maintaining 1P in working memory to create the associ-
ation once 2P is presented.
When 2P is presented, increased activity is observed in
the left DLPFC, in the medial frontal cortex, and in the
PHC compared to the response to 2U. The left VLPFC is
also active during the encoding of 2P, but does not show
a significant response to 2U (Figure 3D). These results
suggest that the left DLPFC, VLPFC, medial frontal cor-
tex, and PHC are involved in associating the two paired
stimuli. The left DLPFC and the medial frontal cortex
also show increased activity in the paired trials starting at
the plus-sign and continuing through the ISI (blank
screen) and 2P, whereas the left PHC and the VLPFC
show increased activity beginning at the presentation of
2P. The dynamics of encoding activation across the
DLPFC/medial frontal cortex and the PHC/ VLPFC dem-
onstrate the temporal characteristics of functional inter-
action between these regions in associative encoding.
Increases in PFC activity during the delay period
under associative instruction supports results from elec-
trophysiology studies using nonhuman primates (Deco
et al., 2005; Fuster et al., 2000). Fuster et al. (2000)
reported PFC neuronal activity in the delay period dur-
ing the association of tones and colors. Similarly, the
present study shows increased PFC activity in the delay
period during the association of two visual stimuli using
human functional imaging (Figure 4). Electrophysiolog-
ical evidence of MTL activity in rats and nonhuman
primates during the delay period is less consistent, with
some studies reporting the presence of MTL activity
( Young et al., 1997; Cahusac et al., 1989; Watanabe &
Niki, 1985) and others reporting very rare MTL activity
(Hampson & Deadwyler, 2003; Vidyasagar et al., 1991).
Such discrepancies in MTL delay-period activity may be
the result of subtle differences in tasks. Nevertheless,
two of the studies using delayed nonmatch-to-sample in
rats also reported divergent results. Further study is
required to examine the circumstances in which sus-
tained MTL delay-period activity may be present in rats,
monkeys, and humans. In the current study, no signif-
icant increase in MTL activity was observed during the
delay period between paired stimuli.
Previous studies have shown direct and indirect ana-
tomical connections between the PFC and the PHC
using anterograde and retrograde tracing techniques in
rhesus monkeys (Goldman-Rakic et al., 1984) and using
DTI and fMRI in humans (Takahashi et al., 2007). Fur-
thermore, top–down modulation from the PFC to the
PHC has been examined using human imaging tech-
niques, including fMRI, electroencephalography (EEG),
and transcranial magnetic stimulation (TMS). Gazzaley
and D’Esposito (2007) employed a visual working mem-
ory task for scenes with constant sensory input for all
conditions to control bottom–up processing and to iso-
late top–down mechanisms of enhancement and suppres-
sion. Event-related fMRI and EEG measured enhanced
activity relative to passive baseline in scene-specific
visual association areas (parahippocampal/lingual gyrus)
when subjects were told to remember scenes and to
ignore faces. When opposite instructions were given,
these regions showed suppressed activity. This effect
demonstrated top–down modulation of the PHC. Fur-
ther research is examining whether the PFC is critical for
modulating PHC activity; preliminary results using re-
petitive TMS to disrupt PFC activity and studies using
working memory tasks that challenge PFC function
suggest that disrupted PFC activity results in deficits in
top–down suppression (Gazzaley & D’Esposito, 2007).
The current study shows that PFC and PHC responses
to a visual stimulus change depending upon the pres-
ence or absence of preceding associative instruction.
Enhanced prefrontal activity at the presentation of asso-
ciative instruction and the resulting enhancement of PFC
and PHC activity during the following stimulus corre-
spond to improved subsequent memory for that item as
well as for the association. These findings reveal that
frontal and medial-temporal regions subserve different
functions in maintaining and binding visual stimuli into
long-term associative memory.
Acknowledgments
This work was supported by the National Institute of Neuro-
logical Disorders and Stroke K23 NS050305, General Electric
Medical Foundation, and the University of California, San Diego
Departments of Neurosciences and Radiology.
The authors thank Sanja Kovacevic and Anders Dale for meth-
odological input and Larry Squire and Sonia Ancoli-Israel for
their insightful comments and helpful feedback.
1252
Journal of Cognitive Neuroscience
Volume 21, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
7
e
-
1
p
2
d
4
f
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
c
4
n
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
f
.
/
o
n
1
8
M
a
y
2
0
2
1
Reprint requests should be sent to James B. Brewer, Human
Memory Laboratory, 8950 Villa La Jolla Drive C212, La Jolla, CA
92037, or via e-mail: jbrewer@ucsd.edu.
REFERENCES
Blumenfeld, R. S., & Ranganath, C. (2006). Dorsolateral
prefrontal cortex promotes long-term memory formation
through its role in working memory organization. Journal of
Neuroscience, 26, 916–925.
Blumenfeld, R. S., & Ranganath, C. (2007). Prefrontal cortex
and long-term memory encoding: An integrative review of
findings from neuropsychology and neuroimaging.
Neuroscientist, 13, 280–291.
Brewer, J. B., Zhao, Z., Desmond, J. E., Glover, G. H., &
Gabrieli, J. D. (1998). Making memories: Brain activity that
predicts how well visual experience will be remembered.
Science, 281, 1185–1187.
Cahusac, P. M. B., Miyashita, Y., & Rolls, E. T. (1989).
Responses of hippocampal formation neurons in the
monkey related to delayed spatial response and
object-place memory tasks. Behavioral Brain Research,
33, 299–240.
Cox, R. W. (1996). AFNI: Software for analysis and
visualization of functional magnetic resonance
neuroimages. Computers and Biomedical Research, 29,
162–173.
Dale, A. M. (1999). Optimal experimental design for
event-related fMRI. Human Brain Mapping, 8, 109–114.
Dale, A. M., & Buckner, R. L. (1997). Selective averaging of
rapidly presented individual trials using fMRI. Human Brain
Mapping, 5, 329–340.
Daselaar, S. M., Rice, H. J., Greenberg, D. L., Cabeza, R., LaBar,
K. S., & Rubin, D. C. (2007). The spatiotemporal dynamics of
autobiographical memory: Neural correlates of recall,
emotional intensity, and reliving. Cerebral Cortex, 18,
217–229.
Davachi, L., Mitchell, J. P., & Wagner, A. D. (2003). Multiple
routes to memory: Distinct medial temporal lobe
processes build item and source memories. Proceedings
of the National Academy of Sciences, U.S.A., 100,
2157–2162.
Davachi, L., & Wagner, A. D. (2002). Hippocampal
contributions to episodic encoding: Insights from relational
and item-based learning. Journal of Neurophysiology, 88,
982–990.
Deco, G., Ledberg, A., Almeida, R., & Fuster, J. (2005). Neural
dynamics of cross-modal and cross-temporal associations.
Experimental Brain Research, 166, 325–336.
Dickerson, B. C., Miller, S. L., Greve, D. N., Dale, A. M.,
Albert, M. S., Schacter, D. L., et al. (2007). Prefrontal–
hippocampal–fusiform activity during encoding predicts
intraindividual differences in free recall ability: An
event-related functional–anatomic MRI study.
Hippocampus, 17, 1060–1070.
Dolan, R. J., & Fletcher, P. C. (1997). Dissociating prefrontal
and hippocampal function in episodic memory encoding.
Nature, 388, 582–585.
Eichenbaum, H., Yonelinas, A. P., & Ranganath, C. (2007). The
medial temporal lobe and recognition memory. Annual
Review of Neuroscience, 30, 123–152.
Fuster, J. M., Bodner, M., & Kroger, J. K. (2000). Cross-modal
and cross-temporal association in neurons of frontal cortex.
Nature, 405, 347–351.
Gazzaley, A., & D’Esposito, M. (2007). Top–down modulation
and normal aging. Annals of the New York Academy of
Sciences, 1097, 67–83.
Gold, J. J., Smith, C. N., Bayley, P. J., Shrager, Y., Brewer,
J. B., Stark, C. E. L., et al. (2006). Item memory, source
memory, and the medial temporal lobe: Concordant
findings from fMRI and memory-impaired patients.
Proceedings of the National Academy of Sciences, U.S.A.,
103, 9351–9356.
Goldman-Rakic, P. S., Selemon, L. D., & Schwartz, M. L.
(1984). Dual pathways connecting the dorsolateral
prefrontal cortex with the hippocampal formation and
parahippocampal cortex in the rhesus monkey.
Neuroscience, 12, 719–743.
Habib, R., Nyberg, L., & Tulving, E. (2003). Hemispheric
asymmetries of memory: The HERA model revisited. Trends
in Cognitive Sciences, 7, 241–245.
Hampson, R. E., & Deadwyler, S. A. (2003). Temporal firing
characteristics and the strategic role of subicular neurons in
short-term memory. Hippocampus, 13, 529–541.
Henke, K., Buck, A., Weber, B., & Wieser, H. G. (1997). Human
hippocampus establishes associations in memory.
Hippocampus, 7, 249–256.
Kirwan, C. B., & Stark, C. E. (2004). Medial temporal lobe
activation during encoding and retrieval of novel face–name
pairs. Hippocampus, 14, 919–930.
Law, J. R., Flanery, M. A., Wirth, S., Yanike, M., Smith, A. C.,
Frank, L. M., et al. (2005). Functional magnetic resonance
imaging activity during the gradual acquisition and
expression of paired-associate memory. Journal of
Neuroscience, 25, 5720–5729.
Loose, R., Kaufmann, C., Tucha, O., Auer, D. P., & Lange, K. W.
(2006). Neural networks of response shifting: Influence of
task speed and stimulus material. Brain Research, 1090,
146–155.
Meltzer, J. A., & Constable, R. T. (2005). Activation of human
hippocampal formation reflects success in both encoding
and cued recall of paired associates. Neuroimage, 24,
384–397.
Murray, L. J., & Ranganath, C. (2007). The dorsolateral
prefrontal cortex contributes to successful relational
memory encoding. Journal of Neuroscience, 27, 5515–5522.
Pihlajamaki, M. H., Tanila, H., Hanninen, T., Kononen, M.,
Mikkonen, M., Jalkanen, V., et al. (2003). Encoding of novel
picture pairs activates the perirhinal cortex: An fMRI study.
Hippocampus, 13, 67–80.
Prince, S. E., Daselaar, S. M., & Cabeza, R. (2005). Neural
correlates of relational memory: Successful encoding and
retrieval of semantic and perceptual associations. Journal of
Neuroscience, 25, 1203–1210.
Rombouts, S. A., Machielsen, W. C., Witter, M. P., Barkhof, F.,
Lindeboom, J., & Scheltens, P. (1997). Visual association
encoding activates the medial temporal lobe: A functional
magnetic resonance imaging study. Hippocampus, 7,
594–601.
Rossion, B., & Pourtois, G. (2004). Revisiting Snodgrass and
Vanderwart’s object pictorial set: The role of surface detail in
basic-level object recognition. Perception, 33, 217–236.
Schluppeck, D., Curtis, C. E., Glimcher, P. W., & Heeger, D. J.
(2006). Sustained activity in topographic areas of human
posterior parietal cortex during memory-guided saccades.
Journal of Neuroscience, 26, 5098–5108.
Smith, A. B., Taylor, E., Brammer, M., & Rubia, K. (2004).
Neural correlates of switching set as measured in fast,
event-related functional magnetic resonance imaging.
Human Brain Mapping, 21, 247–256.
Sperling, R., Chua, E., Cocchiarella, A., Rand-Giovannetti, E.,
Poldrack, R., Schacter, D. L., et al. (2003). Putting names to
faces: Successful encoding of associative memories activates
the anterior hippocampal formation. Neuroimage, 20,
1400–1410.
Hales et al.
1253
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
7
e
-
1
p
2
d
4
f
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
c
4
n
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
.
/
t
f
o
n
1
8
M
a
y
2
0
2
1
Squire, L. R. (1992). Memory and the hippocampus: A synthesis
from findings with rats, monkeys, and humans.
Psychological Review, 99, 195–231.
the right dorsolateral prefrontal cortex on intentional
set switching. Experimental Brain Research, 172,
561–565.
Staresina, B. P., & Davachi, L. (2006). Differential encoding
Vidyasagar, T. R., Salzmann, E., & Creutzfeldt, O. D. (1991).
mechanisms for subsequent associative recognition and free
recall. Journal of Neuroscience, 26, 9162–9172.
Sylvester, C. Y., Wagner, T. D., Lacey, S. C., Hernandez, L.,
Nichols, T. E., Smith, E. E., et al. (2003). Switching attention
and resolving interference: fMRI measures of executive
functions. Neuropsychologia, 41, 357–370.
Takahashi, E., Ohki, K., & Kim, D. S. (2007). Diffusion
tensor studies dissociated two fronto-temporal
pathways in the human memory system. Neuroimage,
34, 827–838.
Talairach, J., & Tournoux, P. (1998). A co-planar stereotaxic
atlas of the human brain. New York: Thieme.
Tendolkar, I., Arnold, J., Petersson, K. M., Weis, S.,
Brockhaus-Dumke, A., van Eijndhoven, P., et al. (2007).
Probing the neural correlates of associative memory
formation: A parametrically analyzed event-related
functional MRI study. Brain Research, 1142, 159–168.
Vanderhasselt, M. A., De Raedt, R., Baeken, C., Leyman, L.,
& D’haenen, H. (2006). The influence of rTMS over
Unit activity in the hippocampus and the parahippocampal
temporobasal association cortex related to memory and
complex behaviour in the awake monkey. Brain Research,
544, 269–278.
Wagner, A. D., Schacter, D. L., Rotte, M., Koutstaal, W., Maril,
A., Dale, A. M., et al. (1998). Building memories:
Remembering and forgetting of verbal experiences as
predicted by brain activity. Science, 281, 1188–1191.
Watanabe, T., & Niki, H. (1985). Hippocampal unit activity and
delayed response in the monkey. Brain Research, 325,
241–254.
Yonelinas, A. P., Hopfinger, J. B., Buonocore, M. H., Kroll, N. E.,
& Baynes, K. (2001). Hippocampal, parahippocampal and
occipital–temporal contributions to associative and item
recognition memory: An fMRI study. NeuroReport, 12,
359–363.
Young, B. J., Otto, T., Fox, G. D., & Eichenbaum, H. (1997).
Memory representation within the parahippocampal region.
Journal of Neuroscience, 17, 5183–5195.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
7
e
-
1
p
2
d
4
f
4
/
1
2
9
1
3
/
7
7
7
/
0
1
9
2
o
4
c
4
n
/
1
2
0
7
0
6
9
0
1
2
4
1
7
0
9
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
0
9
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
.
.
.
f
t
o
n
1
8
M
a
y
2
0
2
1
1254
Journal of Cognitive Neuroscience
Volume 21, Number 7