Disruption of Broca’s Area Alters Higher-order Chunking
Processing during Perceptual Sequence Learning
Andrea Alamia1*, Oleg Solopchuk1*, Alessandro D’Ausilio2, Violette Van Bever1,
Luciano Fadiga2,3, Etienne Olivier1,2, and Alexandre Zénon1
Astratto
■ Because Broca’s area is known to be involved in many
cognitive functions, including language, music, and action pro-
cessazione, several attempts have been made to propose a unify-
ing theory of its role that emphasizes a possible contribution
to syntactic processing. Recentemente, we have postulated that
Broca’s area might be involved in higher-order chunk process-
ing during implicit learning of a motor sequence. Chunking
is an information-processing mechanism that consists of group-
ing consecutive items in a sequence and is likely to be involved
in all of the aforementioned cognitive processes. Demon-
strating a contribution of Broca’s area to chunking during the
learning of a nonmotor sequence that does not involve lan-
guage could shed new light on its function. To address this
issue, we used offline MRI-guided TMS in healthy volunteers
to disrupt the activity of either the posterior part of Broca’s area
(left Brodmann’s area [BA] 44) or a control site just before par-
ticipants learned a perceptual sequence structured in distinct
hierarchical levels. We found that disruption of the left BA 44
increased the processing time of stimuli representing the
boundaries of higher-order chunks and modified the chunking
strategy. The current results highlight the possible role of
the left BA 44 in building up effector-independent represen-
tations of higher-order events in structured sequences. This might
clarify the contribution of Broca’s area in processing hierarchical
structures, a key mechanism in many cognitive functions, come
as language and composite actions. ■
INTRODUCTION
Mastering the myriad of behaviors that make humans dis-
tinctive, such as language, tool use, and music, relies on
the ability to learn, encode, and process perceptual, cog-
nitive, or motor sequences (Perruchet & Pacton, 2006;
Corballis, 2003; Janata & Grafton, 2003; Keele, Ivry,
Mayr, Hazeltine, & Heuer, 2003; Conway & Christiansen,
2001). Regardless of their nature, whether these se-
quences are arbitrary, hierarchically organized, or charac-
terized by some regularities leading to statistical learning
(Keele et al., 2003; Conway & Christiansen, 2001), a com-
mon mechanism known as “chunking” seems to underlie
sequence learning (Gobet et al., 2001). A chunk can be
defined as a unit in a “maximally compressed code” (Mathy
& Feldman, 2012), and chunking is regarded as a way to
facilitate learning and to optimize performance by low-
ering memory load (Penhune & Steele, 2012; Huntley,
Bor, Hampshire, Owen, & Howard, 2011; Bor, Duncan,
Wiseman, & Owen, 2003; Sakai, Kitaguchi, & Hikosaka, 2003;
Rosenbaum, Kenny, & Derr, 1983). More recently, it has
even been suggested that because chunking permits the
detection and encoding of regularities between items in
1Université catholique de Louvain, 2Fondazione Istituto Italiano di
Tecnologia, Genova, Italy, 3University of Ferrara
*These two authors contributed equally to this work.
© 2016 Istituto di Tecnologia del Massachussetts
working memory, it might also play a role in conscious-
ness (Bor & Seth, 2012). From a behavioral point of view,
chunking is characterized by a typical pattern of RTs to the
different sequence items, depending on their position in a
chunk. Infatti, retrieving a chunk comes at the cost of
longer RTs when processing the first item, possibly be-
cause it is accompanied by the retrieval of the subsequent
chunk elements (Clerget, Poncin, Fadiga, & Olivier, 2012;
Pammi et al., 2012; Verwey & Abrahamse, 2012; Sakai et al.,
2003; Verwey & Eikelboom, 2003; Koch & Hoffmann,
2000). Supposedly, chunking makes the processing and
learning of sequences more efficient ( Verwey, 2010;
Bor et al., 2003; Sakai et al., 2003; Koch & Hoffmann,
2000), although chunking is not always a good predictor
of performance or learning (Clerget et al., 2012; Wymbs,
Bassett, Mucha, Porter, & Grafton, 2012).
The neural correlates of chunking remain poorly under-
stood. Numerous brain regions have been found to be
activated during sequence learning (Penhune & Steele,
2012; Doyon et al., 2009; Ashe, Lungu, Basford, & Lu,
2006; Keele et al., 2003; Hikosaka, Nakamura, Sakai, &
Nakahara, 2002), but studies that thoughtfully examined
the neural correlates of chunking are rare. Among the
brain regions activated during sequence learning is the
ventrolateral pFC ( VLPFC), including Broca’s area, Quale
is known to be involved in diverse tasks requiring
sequence processing, including hierarchical processing
Journal of Cognitive Neuroscience 28:3, pag. 402–417
doi:10.1162/jocn_a_00911
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
D
o
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
C
8
N
0
_
7
UN
/
_
j
0
o
0
C
9
N
1
1
_
UN
P
_
D
0
0
B
9
sì
1
G
1
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
io
2
3
e
S
/
j
F
/
.
T
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
(Clerget, Andres, & Olivier, 2013; Bahlmann, Schubotz,
Mueller, Koester, & Friederici, 2009; Koechlin & Jubault,
2006; Schubotz & Fiebach, 2006; Grossman, 1980), sequence
representation (Clerget, Badets, Duqué, & Olivier, 2011;
Dominey, Hoen, Blanc, & Lelekov-Boissard, 2003),
and action sequencing (Clerget, Winderickx, Fadiga, &
Olivier, 2009; Fazio et al., 2009). Inoltre, a few neuro-
imaging studies suggest that Broca’s area is also involved
in chunk processing (Pammi et al., 2012; Wymbs et al.,
2012). In a recent TMS study, we postulated that the
posterior part of Broca’s area (the left Brodmann’s area
[BA] 44) might integrate higher-order chunks (Clerget
et al., 2012) while learning a motor sequence in a serial
RT task.
The goal of this study was to test this prediction by
seeking experimental evidence for the contribution of
the left BA 44 in encoding higher-order relations between
chunks in a nonmotor and nonlinguistic sequence. Fare
so, we took advantage of continuous theta burst stimula-
zione (cTBS; Huang, Edwards, Rounis, Bhatia, & Rothwell,
2005) applied over the left BA 44 to disrupt the neural
activity of this area in healthy participants before they
learn a perceptual sequence. The systematic relation be-
tween consecutive items of the sequence was carefully
controlled, so that it yielded a strongly structured se-
quence with different, clearly identifiable hierarchical
levels (Koch & Hoffmann, 2000) that could be reliably
chunked by most, if not all, participants. As a control,
we performed the same experiment in a separate group
of participants with cTBS applied over the vertex. Using a
between-subject design was mandatory because the
chunking pattern evolves during the course of sequence
apprendimento (Clerget et al., 2012; Wymbs et al., 2012), making
it impossible to test the same participants twice in the
same task.
METHODS
Participants
Altogether, we recruited 30 neurologically normal par-
ticipants, and the final data set (see below) included
24 participants (mean age = 23 ± 4 years, 12 women),
12 in each group. All participants were right-handed,
as assessed by the Edinburgh Handedness Inventory
(Oldfield, 1971), and had normal or corrected-to-normal
vision. Written informed consent was obtained from each
participant. The experiment was conducted according
to the Declaration of Helsinki and approved by the
ethics committee of the Université catholique de Louvain.
The participants received financial compensation to partic-
ipate in this study.
Experimental Setup and Design
The experiment took place in a quiet and dimly lit room.
The volunteers sat in an armchair in front of a personal
computer. Before participating in this study, each par-
ticipant was seen by a neurologist to minimize the poten-
tial risk of their experiencing an adverse reaction to TMS
(Keel, Smith, & Wassermann, 2001). The experiment was
controlled by a Matlab program (The MathWorks, Inc.,
Natick, MA). To avoid imprecision in measurement of RT
because of the delays inherent in Microsoft Windows,
we used a homemade device allowing us to detect key-
presses with submillisecond accuracy. Briefly, this device
contains a microcontroller (MSP430F249, Texas Instru-
menti, Inc., Dallas, TX), which combines both Video
Graphics Array and keyboard inputs; in this particular case,
the display of the imperative stimulus triggered an internal
clock, which was stopped by a keyboard event (keypress).
The microcontroller then sent both the key code and the
clock value (128 μsec temporal resolution) to the com-
puter via a USB interface.
The experiment comprised four stages: (1) a control
task block (CT1), (2) cTBS application, (3) the main task
(eight blocks), E (4) a second control task block (CT2).
Following the cTBS application, the participant remained
seated for around 5 min before moving to the experi-
mental setup in the same room.
Main Task
The design of the sequence used in this experiment
builds on a study by Kühn (2011). Participants had to
Imparare, explicitly, by trial and error, a 16-element sequence
characterized by a systematic relational structure among
the items that constituted it (Koch & Hoffmann, 2000;
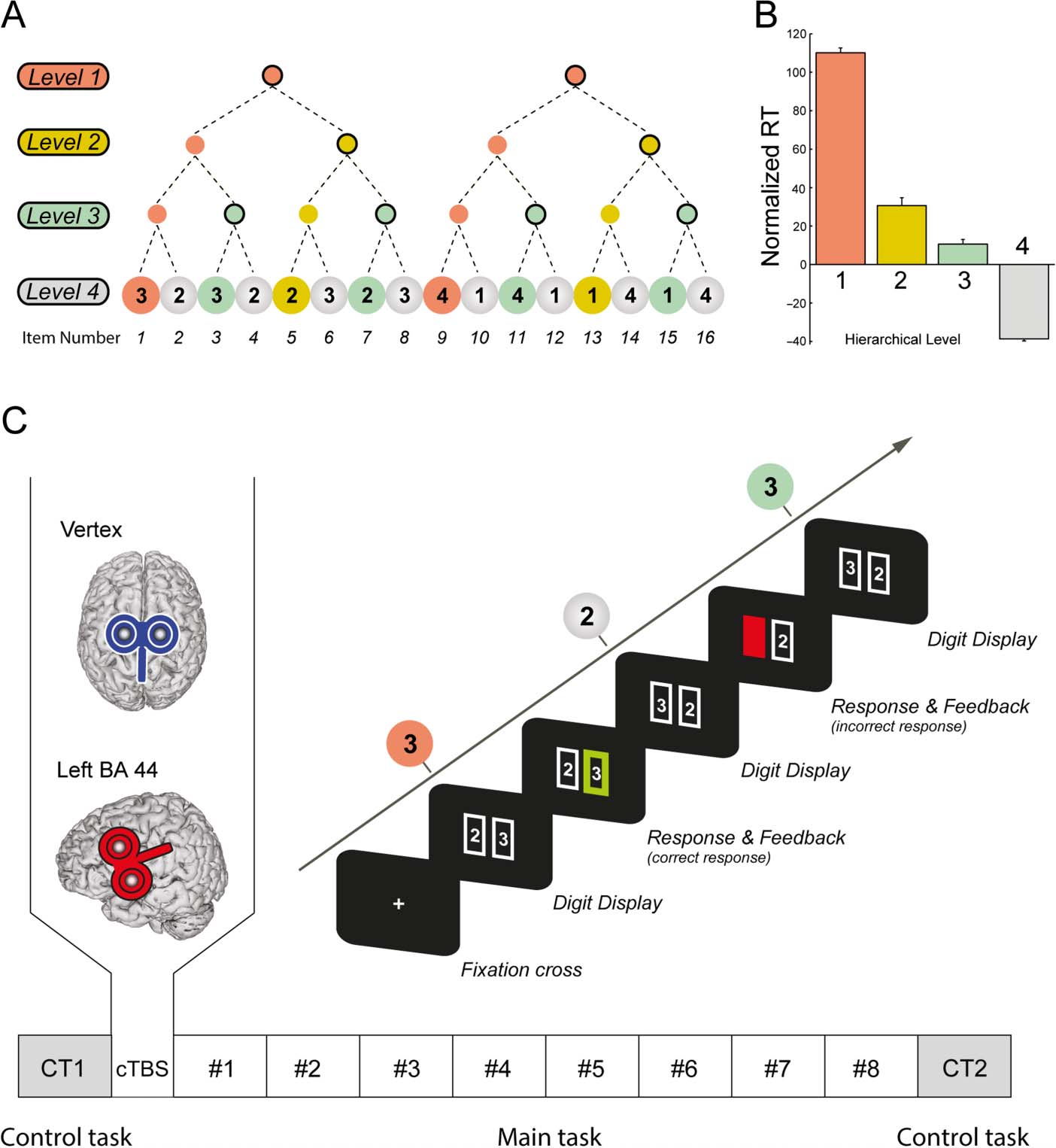
Figure 1A). The sequence was designed in such a way
that it should be chunked comparably by all participants:
Infatti, the main sequence could be parsed into two sub-
sequences of eight digits, each with the same relational
structure (32322323-41411414). These subsequences
could again be parsed into two quadruplets related to
each other by inversion (3232-2323 E 4141-1414),
and each quadruplet could be further divided into two
identical pairs (32-32, 23-23 E 41-41, 14-14). Therefore,
this sequence allowed us to distinguish among four rela-
tional levels between the stimuli, referred to as “chunk-
ing levels” hereafter. The different chunking levels are
illustrated in Figure 1A, in red (Level 1), yellow (Level
2), green (Level 3), and gray (Level 4). The statistical anal-
ysis performed on all items belonging to the same hier-
archical levels showed a strong influence of hierarchical
level on RT [see Figure 1B, main effect of Level, F(3, 66) =
13.91, P < .0001; see Results for detail], clearly demon-
strating the effectiveness of the sequence structure manip-
ulation on RT.
The sequence was repeated five times in each block,
and the participants performed eight blocks, leading to
40 sequence repetitions across the whole experiment.
Importantly, participants were not informed about the
sequence structure. Each sequence started with the dis-
play of a central fixation cross for 2000 msec, followed by
Alamia et al.
403
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
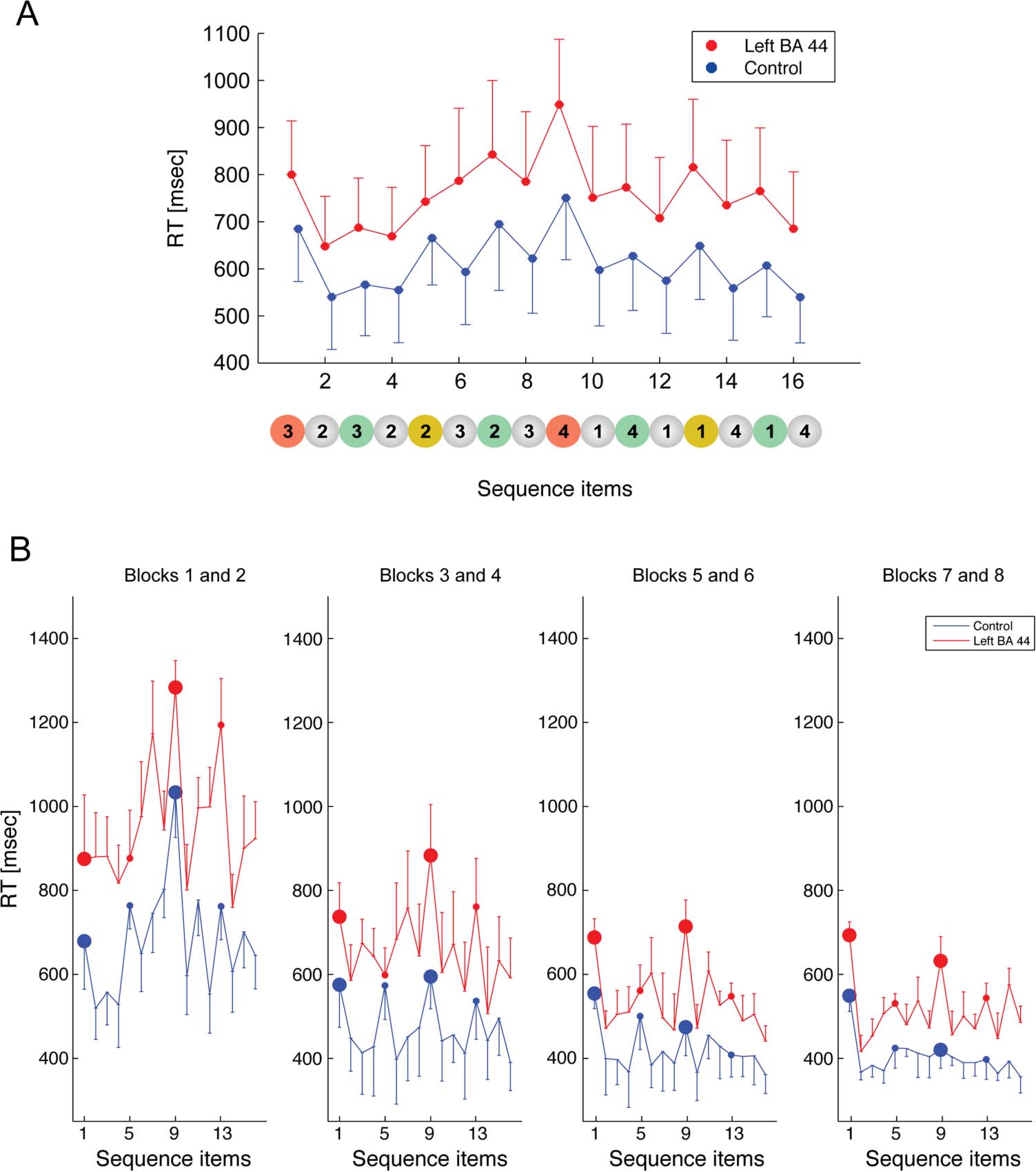
Figure 1. Sequence and
experimental procedure.
(A) The sequence to be learned
contained 16 elements and was
characterized by a particular
relational structure, so it could
be chunked consistently by
all participants according to
four distinct levels, indicated in
red (Level 1), yellow (Level 2),
green (Level 3), and gray
(Level 4). (B) Average RT
within each hierarchical level
(normalized with respect to
the mean RT of each block to
account for the learning effect).
Same color convention as in A.
(C) Time course of a sequence
and design of the experiment.
Each sequence started with the
display of a fixation cross for
3000 msec, followed by the
display of a first pair of stimuli
(digits in a rectangle). The
participant had to select the
digit belonging to the sequence
by pressing the appropriate
response key (right index or
right middle finger). This
was followed by feedback
consisting of a green rectangle
surrounding the selected digit
for a correct response or a
red mask for an incorrect one.
Then the next pair of digits
was displayed. After the
completion of one sequence
(i.e., 16 consecutives trials),
a fixation cross appeared
again for 2000 msec, indicating the onset of the next sequence. Each block comprised five sequence repetitions. The lower part of the figure
illustrates the experimental design. The experiment started with the first control task (CT1), which had the same design as the main task except
that the four digits were replaced by for four letters (A, B, C, and D) and there was no sequence to learn (“B” was always the correct response).
It was followed by the application of cTBS either over either the left BA 44 or the vertex (control group). Then, the participants performed the main
task (8 blocks × 5 sequences), and the control task was repeated (CT2) at the end of the experiment.
the simultaneous presentation of two rectangles (white
border contrasted against a black background, 9.5° width
and 13.4° height) arranged horizontally on either side of
the screen center (Figure 1C). Two digits (from 1 to 4) were
displayed on the screen, one in each rectangle: One digit
belonged to the sequence to be learned (“target” digit),
whereas the other was selected at random. Importantly,
the position of the “target” digit on the screen (left or right
side) was pseudorandomly determined in each sequence
repetition, so that only the sequence of digits was struc-
tured, not the sequence of motor responses. The two digits
were displayed until the participant pressed the response
key or for 5000 msec if no response was provided. The par-
ticipants were asked to respond to the digit presentation as
quickly as possible by pressing one response key on a com-
puter keyboard with either the right index or right middle
finger. The stimulus–response mapping was always congru-
ent (i.e., the selection of the left digit had to be indicated by
an index finger response and that of the right digit by a mid-
dle finger response). As soon as the participant provided a
response, they received visual feedback for 250 msec: The
rectangle border became green when the selected digit be-
longed to the sequence, whereas a red mask covered the
entire rectangle when the response was incorrect
(Figure 1C). Then, 500 msec after the keypress or after
the 5000-msec limit if no response was provided, the next
pair of digits was displayed on the screen. After the comple-
tion of one entire sequence (i.e., 16 consecutive trials), a
fixation cross appeared again for 2000 msec, indicating
the onset of the next sequence.
At the end of the experiment, participants were asked
to write down the sequence they learned and explain the
strategy they used to memorize it. The appropriate re-
sponse, if participants used the expected strategy, was
32-32-23-23-41-41-14-14. This test was used a posteriori
to select participants who used the anticipated chunking
404
Journal of Cognitive Neuroscience
Volume 28, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
/
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
strategy (excluded participants reported using the 32322-
323-41411-414 chunking pattern instead).
Control Task
The participants had to perform the control task once
before (CT1) and once after (CT2) the main task. The
control task design was the same as that of the main task;
that is, the number of items in each sequence (n = 16),
the number of sequence repetitions (n = 5), and the trial
design were identical. The only differences were as
follows: (1) the four digits were replaced by four letters
(A, B, C, and D), (2) the two letters displayed simulta-
neously on the screen were selected at random because
there was no sequence to learn, and (3) the target was
always the letter B. The aims of this control task were
to compare the baseline performance of the two groups
and to assess the possible effect of fatigue at the end of
the experiment.
TMS
Biphasic TMS pulses were delivered by means of a Magstim
Super Rapid stimulator (Magstim Company, Whitland, UK)
via a 70-mm-diameter figure-of-eight coil and administered
following the cTBS protocol originally described by Huang
et al. (2005): Three pulses delivered at 50 Hz every
200 msec for 40 sec, leading to a total of 600 pulses. The
stimulation intensity was set at 80% of the resting mo-
tor threshold, measured for each participant, in accor-
dance with some earlier studies from our group (Huang
& Mouraux, 2015; Zénon, Sidibé, & Olivier, 2015;
Torta et al., 2013), as well as others (Chung et al., 2012;
Goldsworthy, Pitcher, & Ridding, 2012; Nyffeler et al.,
2006). To determine the resting motor threshold, single bi-
phasic pulses were applied over the hand representation of
the left primary motor cortex, whereas motor-evoked po-
tentials were recorded from the right first dorsal interos-
seous muscle. EMG activity was recorded with surface
electrodes (Neuroline, Medicotest, Denmark), and the
signals were amplified (gain: 1K), band-pass filtered (10–
500 Hz; Neurolog Digitimer Ltd., Herefordshire, UK), and
digitized online at 1 kHz using a personal computer. Once
the optimal position of the coil was found, we determined
the minimum intensity needed to produce a 50-μV peak-
to-peak motor-evoked potential in 5 of 10 stimulations
(Rossini et al., 1994).
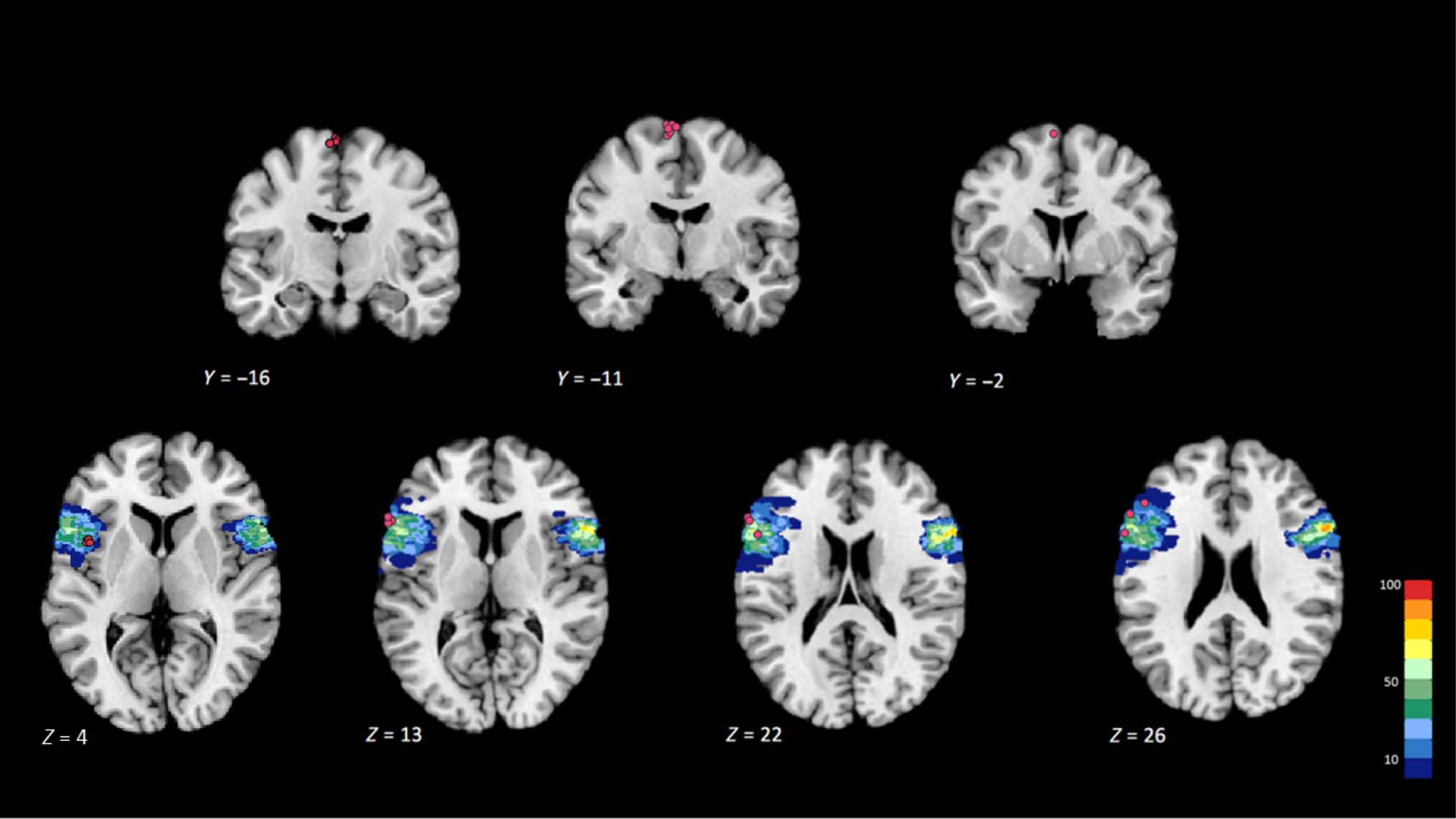
cTBS was applied either over left BA 44 (experimental
group, n = 12) or the vertex (control group, n = 12). For
both stimulation sites, the coil was placed tangentially on
the scalp with the handle pointing backward. For the
control site (vertex), it was held horizontally and main-
tained perpendicular to the midsagittal plane; for left
BA 44, the coil was pointing with an angle of about 10°
upward with respect to the horizontal plane. The coordi-
nates of the targeted stimulation sites were selected from
the literature (−46, 10, and 4 mm; x, y, z, Montreal Neuro-
logical Institute [MNI] system) for left BA 44 (Heim,
Eickhoff, & Amunts, 2009; Amunts et al., 2004) and 0,
−15, and 74 mm for the vertex (Okamoto et al., 2004).
To position the coil accurately on the scalp surface, the
selected target site was located on each individual’s struc-
tural scan, using a reverse-normalization procedure, and
then the coil was guided by means of a homemade neuro-
navigation system (Davare, Andres, Cosnard, Thonnard, &
Olivier, 2006; Zosso et al., 2006; Noirhomme et al., 2004).
When required, the coil position was then slightly ad-
justed to coincide with the “pars opercularis” of the
inferior frontal gyrus. At the end of the experiment, we
computed the actual coordinates of each individual’s
TMS site and converted them into the anatomical MNI
space (with the anterior commissure as the anatomical
reference); for the experimental group, individual TMS
sites were superimposed on the probabilistic cytoarchi-
tectonic map gathered for the left BA 44 (Eickhoff et al.,
2005, 2007; Figure 2). cTBS was tolerated well by all par-
ticipants, and no discomfort or other negative side effects
were reported.
Data and Statistical Analyses
Five participants (four in the control group and one in
the left BA 44 group) of 30 participants were discarded
because their chunking strategy differed from that antic-
ipated. An additional control participant was removed
from our data set because he executed only 10 correct
sequences out of 40, a performance much lower than
that of the other participants (correct sequence number:
27.9 ± 5.1, mean ± SD, n = 24). Hence, as mentioned
above, the final data set included 24 participants, 12 in
each group.
Because the number of correct trials differed between
participants, we used generalized linear mixed models
(GLMMs) for data analyses. The GLMM approach in-
cludes both fixed effects (the predictors or factors of
the model) and random effects, taking account of the dif-
ferences among participants (Baayen, Davidson, & Bates,
2008). The random model was designed considering all
within-subject factors and their interactions. As far as
the main task was concerned, we performed two sepa-
rate analyses, one on RT and one on response accuracy.
The RT was defined as the delay between the stimulus
presentation (digits in the main task and letters in the
control task) and the keypress response; RT values were
log-transformed to ensure the normality of their distri-
bution (log transformation led to a change in average
kurtosis from 15.98 to 3.99 and in skewness from 3.06 to
1.09); incorrect trials and RT values lower than 150 msec
were discarded from the analysis. For the second analysis,
to determine response accuracy, we computed the ratio
between the correct and all the trials; trial-by-trial per-
formance in the task was modeled as a binomial variable. In
the models used to analyze RT and accuracy, the predictors
Alamia et al.
405
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 2. TMS stimulation sites. Upper row: Projection of individual vertex TMS sites (n = 12) on three coronal sections through the MNI
single-subject template, at y = −20, −15, and −6, respectively, in anatomical MNI space (+4 in original MNI space). Lower row: Projection of
individual left BA 44 TMS sites (n = 12) on four axial sections through the MNI single-subject template, respectively, at z = 9, 18, 27, and 31 in
anatomical MNI space (−5 in original MNI space). Probabilistic maps of BA 44 were superimposed on these sections.
taken into account were Group (control vs. left BA 44),
Block (from 1 to 8), and Level (Chunking Levels 1, 2, 3,
and 4). Because the learning effect followed a decreasing
exponential trend, we considered the Block factor as a
continuous variable and log-transformed to ensure the
linearity between RT and Block and between accuracy
and Block; these log transformations increased the log-
likelihood of both GLMMs, providing evidence that they
improved the model fit.
In addition, to detect baseline difference between the
two groups and to evaluate a possible fatigue effect, we
analyzed the log-transformed RT gathered in CT1 and
CT2 by using a GLMM considering Group (control vs. left
BA 44), Block (CT1 vs. CT2), and their interaction as
factors. All the analyses were performed using SAS 9.3 soft-
ware (SAS Institute, Cary, NC).
Finally, to investigate the dynamics of the chunking
process on a trial-by-trial basis and its possible change
following left BA 44 cTBS, we used a network-based
community detection algorithm comparable to that used
recently by Grafton and colleagues (Bassett et al., 2013;
Wymbs et al., 2012). However, the modularity optimiza-
tion algorithm we used was slightly different. Indeed, in
this study, the modularity function used to detect chunks
was influenced by only one parameter, namely the inter-
sequence weight (c1), which allowed us to model the
temporal dynamics across sequences (see below for de-
tails). This parameter has been set to 0.7, such that the
intersequence weight remained comparable to the intra-
sequence weights (see below for details).
The network-based community detection algorithm we
used comprised the following steps:
(cid:1)dij−dij
(cid:1)dij
1. For each participant, each sequence was modeled as a
single-layer network (see Figure 3A): The nodes rep-
resented each element of the sequence, whereas the
weight of the links between each node was defined as
follows: wij ¼
, where wij represents the link be-
tween the ith and jth sequence elements, dij is the
difference between the RT for the ith and jth element
of the sequence, and (cid:1)dij is the largest dij value in the
sequence. The weight value was bounded between 0
and 1. It is noteworthy that there was only a link be-
tween adjacent nodes (|i − j| = 1), and therefore, for
each sequence, we ended up with a network com-
posed of 16 nodes and 15 links.
2. For each participant, the five sequences belonging to
the same block were linked together into a single net-
work, a “block network” that comprised 80 nodes.
This network was built by linking each node of the
single-layer network with its homologs in the other
four sequences; the weights of these links were de-
fined by four distinct constants, as schematically
shown in Figure 3A. These four values were linearly
dependent and were proportional to c1, the inter-
sequence weight; c2, c3, and c4 were respectively
406
Journal of Cognitive Neuroscience
Volume 28, Number 3
0.6, 0.45, and 0.3 times c1. These values were selected
so that the link between temporally-closer sequences
was higher, keeping a balance in the weight distribu-
tion; it is noteworthy that small variations of those
coefficients did not affect the results of our analysis.
This network architecture allowed us to take into
account RT changes in neighboring sequences while
identifying chunks.
3. Once the block networks were built, we ran a clustering
method for each of them, based on modularity optimiza-
tion (Blondel, Guillaume, Lambiotte, & Lefebvre, 2008).
This approach led to the identification of clusters both in
the sequence direction (the chunks) and in the orthog-
onal direction, representing the successive sequences.
4. On the basis of the clusterization performed in the
block networks in the previous step, we computed
the modularity of each sequence of the block net-
ij
(cid:2)
(cid:1)
(cid:4)
wij−Pij
(cid:3)
δ gi; gj
work, named a single-layer network. We thus ob-
tained five values per block network. The modularity
was computed using the following formula: Q ¼
X
, where wij is the element ij of
the weight matrix of the single-layer network, whereas
Pij is the element ij of the weight matrix of the null
model (see below); gi represents the chunk index of
the element i, as determined in Step 3; ∂( gi,gj) is
equal to 1 if gi = gk, indicating that the elements i
and k belong to the same chunk, otherwise it equals 0.
The null model used to compute the modularity in
each single-layer network was a matrix whose ele-
ments were equal to the node strength, that is, the
sum of the weights of each link belonging to a node
(Barrat, Barthélemy, Pastor-Satorras, & Vespignani,
2004; Newman & Girvan, 2004). This modularity
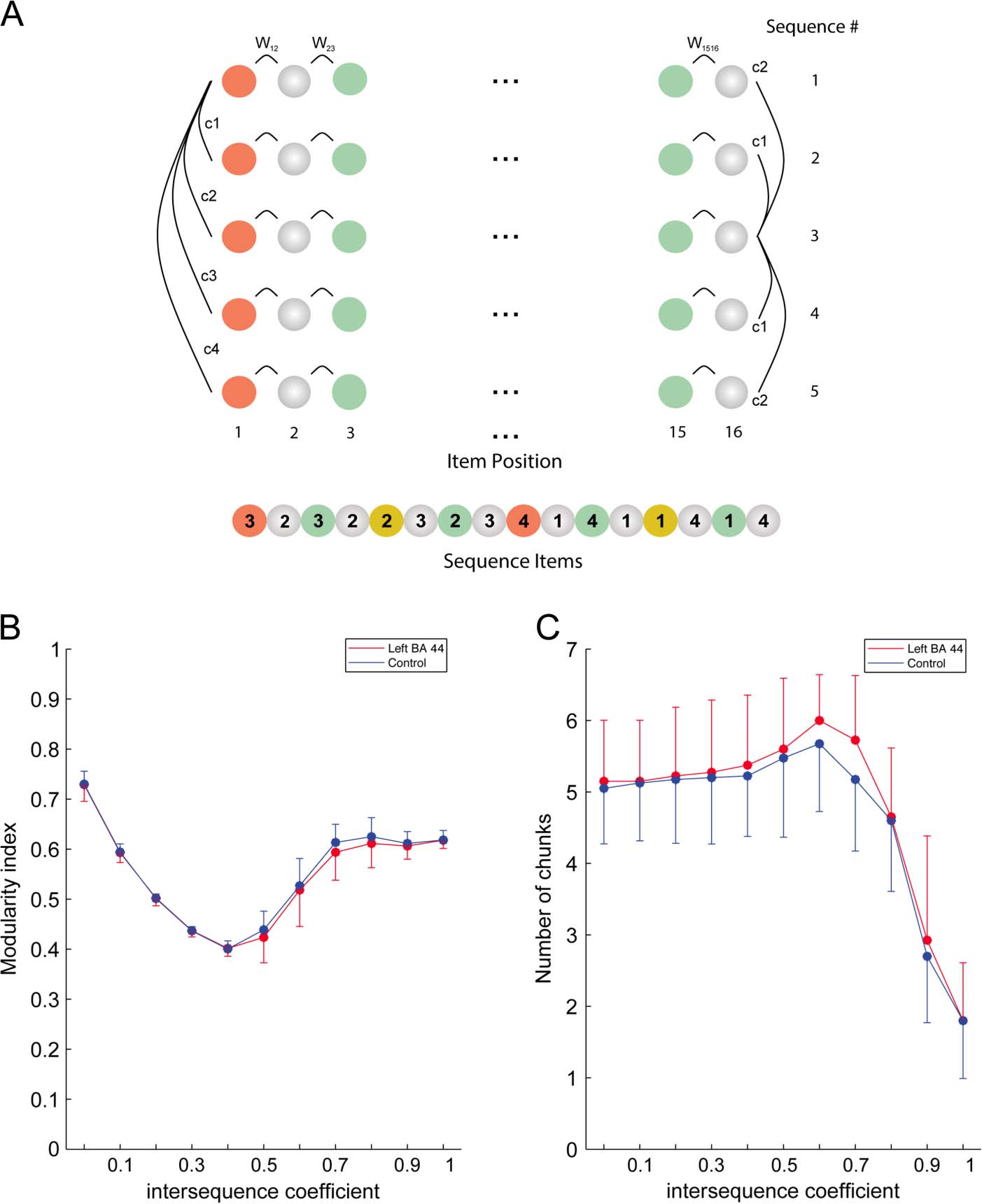
Figure 3. Network-based
community detection
algorithm. (A) Multisequence
network: In the network
analysis, each block was
modeled by combining the
five sequences to constitute
a block network. As shown
in the figure, only the links
between adjacent nodes (Wij,
intrasequence weights) and
between nodes with the same
position in the sequence (c1
to c4, intersequence weights)
were considered. The
intrasequence weights were
computed on the basis of the
RTs of each participant (see
Methods). The intersequence
weights were computed as
follows: c2 = 0.6 × c1, c3 =
0.45 × c1 and c4 = 0.3 × c1;
c1 was a constant. For
illustrative purpose, the
intersequence coefficients of
the first element of the first
row are displayed on the left
of the figure (c1, c2, c3, c4),
whereas the intersequence
coefficients of the last element
of the third row are displayed
on the right (c1, c2). (B)
Change in the modularity
index for the left BA 44 (red)
and control (blue) groups,
when varying the value of
intersequence coefficient (c1)
by steps of 0.1. Error bars
indicate SD. (C) Change in
the number of chunks for the
left BA 44 (red) and control
(blue) groups, when varying
the value of intersequence
coefficient (c1) by steps of
0.1. Error bars indicate SD. These figures show that a value of 0.7 (i.e., the value used in remaining analyses) allowed reaching an optimal balance
between the number of chunks and modularity index.
Alamia et al.
407
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
t
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
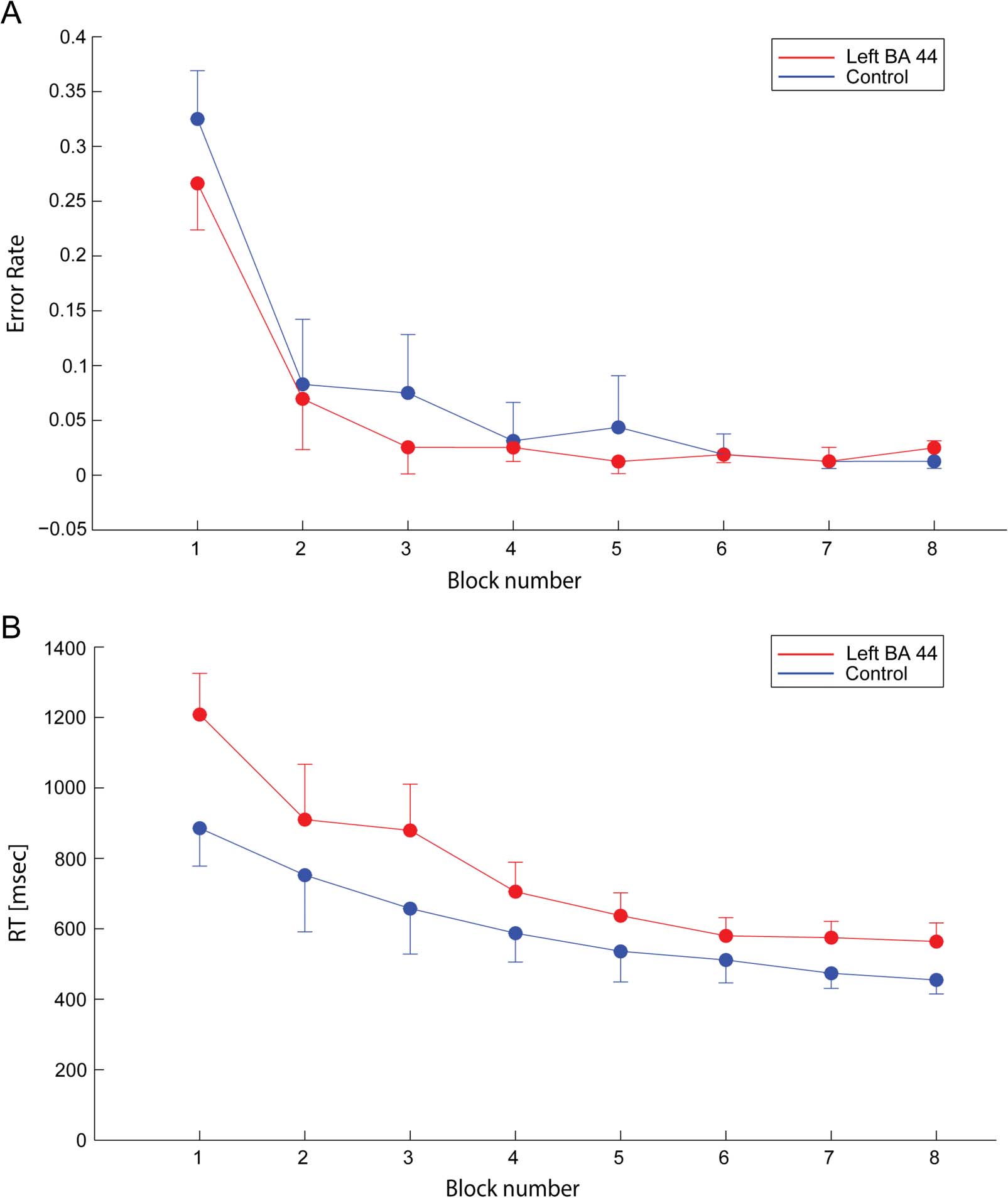
Figure 4. Error rate and RT changes across blocks. (A) Changes in error rate across blocks for the control (blue circles) and left BA 44
(red circles) groups. Error bars indicate SE. (B) Decrease in RT across block repetitions for the control (blue circles) and left BA 44 (red circles)
groups. Error bars indicate SE.
5.
optimization null model is the most commonly used
in undirected single-layer networks.
In the last step, as proposed by Wymbs et al. (2012), we
computed the “chunk magnitude,” named φ, which
was the reciprocal value of the modularity, gathered
for each individual sequence, after having normalized
the modularity values for each participant. Wymbs
et al. (2012) interpret φ as follows: a low φ value indi-
cates that the community detection algorithm was
able to detect chunks easily, suggestive of a high
modularity and evocative of what they called the
segmentation process, referring to the parsing of
the sequence into smaller groups. In contrast, a high
φ value indicates that chunks were difficult to isolate,
reminiscent of the concatenation process, corre-
sponding to the process of grouping chunks together
to build superordinate chunks. Thus, for each par-
ticipant, we ended up with 40 normalized φ values
(5 sequences × 8 blocks) on which we ran a repeated-
measures ANOVA with Group and Block as factors.
Additionally, to provide a comparison with the results
of Wymbs et al. (2012), we computed a linear regres-
sion between φ and sequence numbers for each par-
ticipant. Finally, we fitted a bilinear regression to the
408
Journal of Cognitive Neuroscience
Volume 28, Number 3
averaged φ values, using least squares estimation (nlin-
fit function in Matlab) with the following regression
function:
8
<
y ¼
:
ð
ð
Þ
ax þ b
Þ
dx þ e
(cid:6)
(cid:5)
if x < e−b
a−d
otherwise
To evaluate the consistency of our network-based
model, we computed the mean modularity for the
two groups while varying the intersequence coefficient
(c1), the only independent parameter that indicates the
strength of the link between the same items across se-
quences in the block network. This coefficient ranged
between 0 (indicating an absence of link between se-
quences in the block) and 1 (the strongest possible link
between successive sequences), with a step of 0.1, for a
total of 11 steps. We then followed the same approach
for the number of chunks (Figure 3C). For all the follow-
ing analyses, we selected the value of 0.7, which led to a
Figure 5. Chunking pattern.
(A) RT for each sequence
item (from 1 to 16) for all
correct sequences in the
control (blue lines) and left
BA 44 (red lines) groups.
Error bars indicate SE. (B) RT
for each sequence item,
as in A, but showing averaged
data over pairs of blocks.
Large and small dots represent
Level 1 and Level 2 items of
the sequence, respectively.
large modularity index and a relatively large number of
chunks (see Figure 3C) and which was close to the aver-
age value of the intrasequence coefficients (0.6934 ±
0.0245, mean ± SE), allowing to keep the block network
balanced across its two dimensions (intrasequence and
intersequence). The method we used to determine
those parameters fits with the approach followed by
Wymbs et al. (2012). This network analysis was imple-
mented by using Matlab (The MathWorks, Inc.).
RESULTS
Control Task
The GLMM analysis performed on the data gathered during
the control tasks did not reveal any significant effect neither
for the Block factor [F(1, 22) = 0.12, p = .7319] nor for the
Group factor or the Block × Group interaction [F(1, 4588) =
1.05, p = .3056 and F(1, 4588) = 2.59, p = .1076, respectively],
indicating no significant baseline difference between groups.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Alamia et al.
409
Main Task
In the main task, participants had to learn the 16-element
sequence during 40 successive repetitions (5 sequences ×
8 blocks). We first analyzed the possible influence of left
BA 44 TMS on accuracy. A binomial GLMM, considering
Block, Group, and Level as factors, indicated a significant
Block effect, F(1, 22) = 294.34, p < .0001 (see Figure 4A),
but no effect of Group, F(1, 15005) = 1.10, p = .2946 or of
Level, F(3, 66) = 2.22, p = .0937; nor were the other inter-
actions significant (all ps > .05). It is noteworthy that the
Level effect was close to reaching significance, suggesting
that the position of a given item in the sequence hierarchy
influenced the participants’ accuracy, with fewer errors be-
ing performed in response to sequence items with the low-
est hierarchical level (Level 4 items).
We then examined RT changes across blocks for the
two groups to identify the impact of left BA 44 disruption.
A GLMM analysis with Group (left BA 44 vs. vertex), Block
(Blocks 1–8), and Level (chunk level, from 1 A 4) as fixed
effect factors showed a main effect of Block on RT [F(1,
22) = 107.65, P < .0001, estimate ± SE = −0.3266 ±
0.04098] but neither a main Group effect, F(1, 13542) =
2.03, p = .1545, nor any significant two-way interactions
[see Figure 4B, Group × Block: F(1, 13542) = 0.61, p =
.4355; Block × Level: F(3, 66) = 0.56, p = .6425; Level ×
Group: F(3, 13542) = 2.07, p = .1021]. The GLMM analysis
also revealed a significant main effect of Level, F(3, 66) =
13.91, p < .0001, estimate ± SE: Level 1 = −0.1644 ±
0.03109, Level 2 = −0.01587 ± 0.03088, Level 3 = 0.05912 ±
0.02707, Level 4 = 0.1211 ± 0.02483 (see Figure 1B).
Tukey–Kramer adjusted post hoc analyses showed that
the main effect of Level resulted from a significant dif-
ference between RT for Level 1 items and for the three
subsequent levels (all t values > 4.5, all ps < .0001) and
a significant difference between Levels 2 and 4 (t value =
3.49, p < .001) and Levels 3 and 4 (t value = 2.24, p =
.0283), whereas the comparison between Levels 2 and
3 failed to reach significance (t = 1.29, p = .2027). As
illustrated in Figure 5, this analysis indicates that the
RT to the different items depended on their hierarchical
level. Specifically, in both groups, the item position in
the sequence affected RT, such that the highest hierar-
chical levels (Items 1 and 9, Level 1) led to a longer RT.
Finally, the three-way interaction was significant [Block ×
Level × Group, F(3, 13542) = 3.06, p = .0270, estimate ±
SE for Level 1 = 0.1127 ± 0.03784, Level 2 = −0.04149 ±
0.03768, Level 3 = −0.05139 ± 0.03222, Level 4 =
−0.01987 ± 0.02901]. To explore this three-way inter-
action further, we compared the between-group differ-
ence in the slopes of the regression lines computed
between RT and blocks across the four hierarchical levels
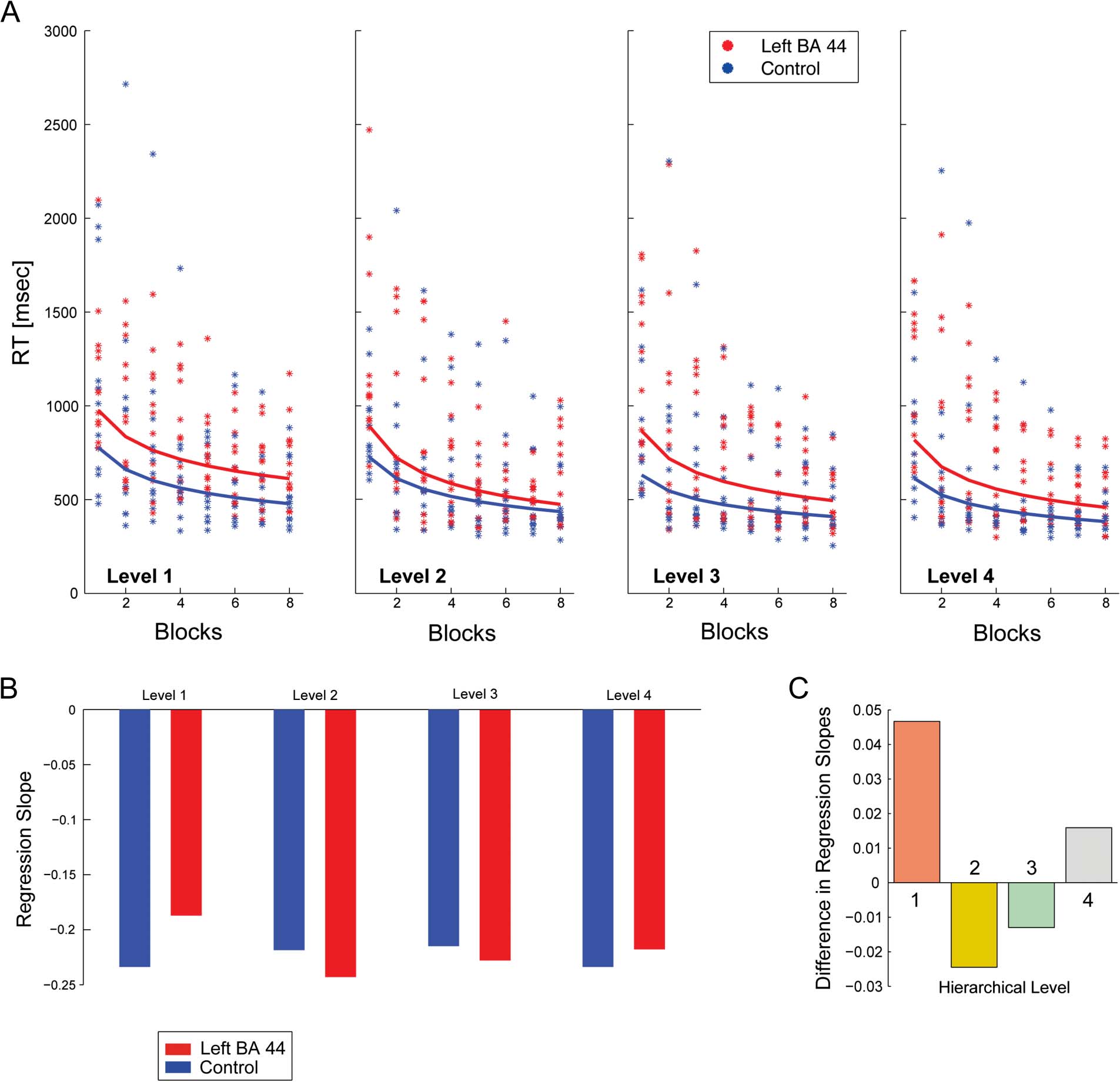
(Figure 6A, B and C). This analysis showed a significant
difference only for comparisons between Level 1 and sub-
sequent levels (Levels 1–2: estimate ± SE = 0.1157 ±
0.04818, t = 2.40, p = .0164; Level 1–3: estimate ± SE =
0.1231 ± 0.04348, t = 2.83, p = .0046; Level 1–4: estimate ±
SE = 0.09947 ± 0.04088, t = 2.43, p = .0150; all p values
Tukey–Kramer corrected); no other comparisons were
significant (all ps > .05). As illustrated in Figure 6A, for
items belonging to Levels 2–4, the computed slopes of the
regression lines for each group converged, reducing the
distance between the left BA 44 and control groups across
block repetition. In contrasto, for Level 1 items, the slopes
diverged, showing that the distance between the two groups
increased across blocks, suggesting a slower improvement
for Level 1 items for the left BA 44 group compared with
controls. Overall, these results indicate that left BA 44 cTBS
altered only the processing of higher-level chunks, in partenza
unaffected the processing of lower-level chunks.
Finalmente, to investigate chunking dynamics during learning
on a sequence-by-sequence basis and its possible alteration
following left BA 44 TMS, we used a network-based commu-
nity detection algorithm (Bassett et al., 2013; Wymbs et al.,
2012) to compute the “chunk magnitude” (φ) for each se-
quence. As a reminder, a low φ value indicates that the se-
quence was easily separable into chunks by the community
detection algorithm, reminiscent of an ongoing segmenta-
tion process, whereas a high φ value indicates that chunks
were difficult to isolate, suggesting a concatenation process
or the absence of chunks (Wymbs et al., 2012). In a first
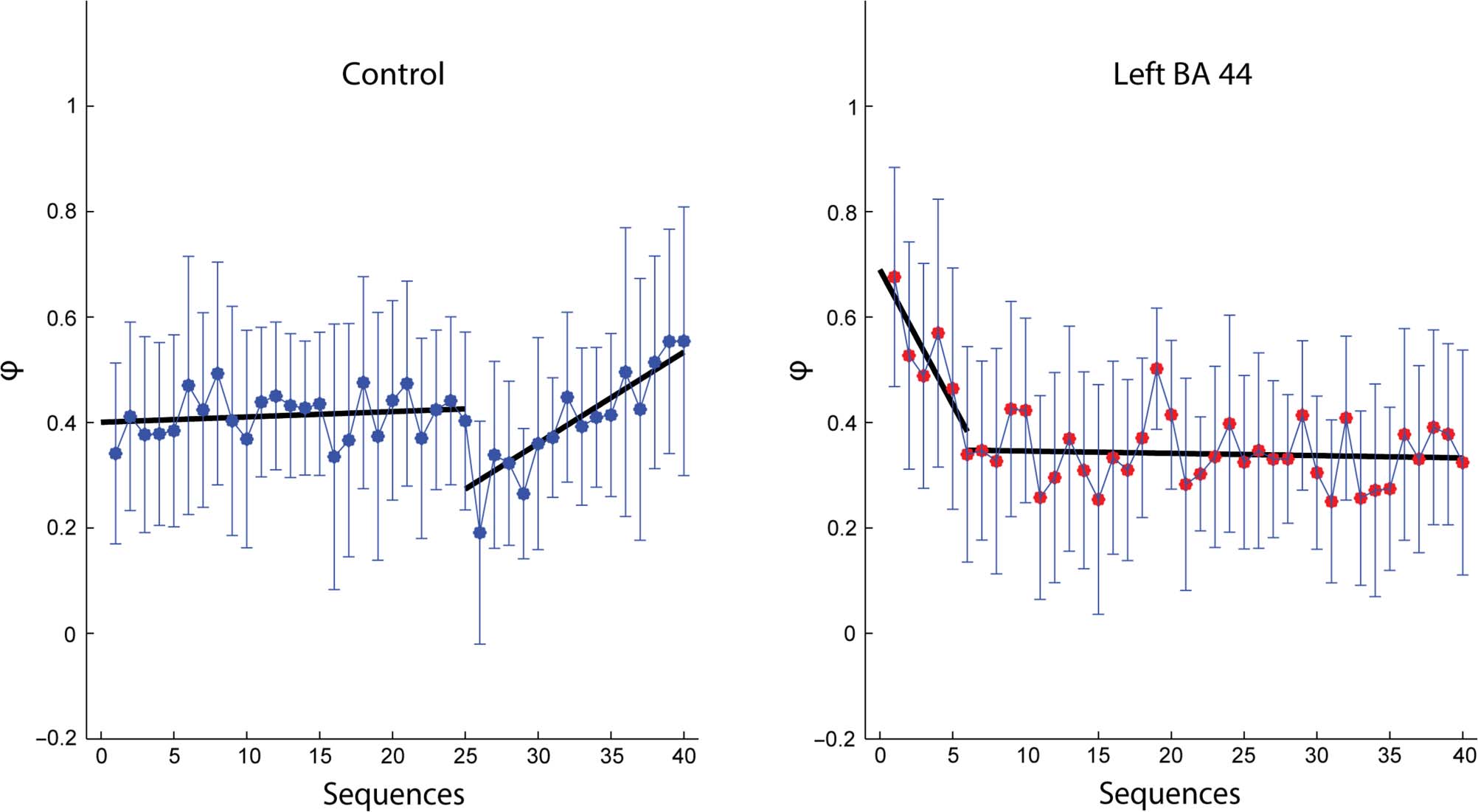
analysis, we computed the mean φ value for each sequence
(n = 40), separately for each group (12 participants per
group). The variations of φ across sequences were dramat-
ically different between groups (Figura 7). In controls, IL
mean φ values increased across sequence repetitions (lin-
ear regression between average φ and sequence number,
slope = 0.001, p = .03), in agreement with the finding of
Wymbs et al. (2012). In contrasto, following cTBS application
over left BA 44, we observed a negative slope (linear re-
gression, slope = −0.0037, p = .02). Because the linear re-
gression failed to account for the time course of φ across
sequences, we also performed a bilinear regression (Vedere
Methods). In the BA 44 group, we found a significant neg-
ative φ slope in the beginning of the sequence learning (es-
timate 95% confidence interval [CI]: −0.0933, −0.02) and a
flat φ function thereafter (estimate CI: −0.0031, 0.0012). In
the control group, the initial part of the φ function was flat
(estimate CI: −0.0018, 0.0043), whereas the later portion
had a positive slope (estimate CI: 0.0118, 0.0228). Finalmente,
this difference in φ variation between groups was
confirmed by performing a two-way, repeated-measures
ANOVA on all the φ values (n = 960, 24 participants ×
8 blocks × 5 sequences/block) with Group and Block as
factors. This analysis revealed a main effect of Block ( P <
.0001) and Group ( p = .0216) and a significant Group ×
Block interaction ( p < .0001), confirming that φ was lower
in the control group during the first blocks, whereas it
decreased in the last blocks in the BA 44 group.
DISCUSSION
This study shows that disruption of Broca’s area modifies
the processing time of higher-order chunks during the
410
Journal of Cognitive Neuroscience
Volume 28, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
.
f
t
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 6. RT change across blocks for items at different chunking levels (from 1 to 4). (A) Each dot represents the average RT for items belonging to
the same chunking level, for each participant from either the control (blue dots) or the left BA 44 (red dots) groups, computed for all correct
sequences. The BLOCK factor was modeled as a continuous log-transformed variable but displayed on a linear scale. For each chunking level, the
regression lines computed between RT and log-transformed block numbers is shown. Only the slopes of the regression lines computed for Level 1
items were significantly different between the two groups when compared with the three other levels. (B) Slope values for each hierarchical level are
displayed, in red for the left BA 44 group and in blue for the control group. (C) Slope differences for regression lines computed for each chunking
level. Same color convention as in Figure 1.
learning, by trial and error, of a highly structured per-
ceptual sequence, involving neither motor nor explicit
linguistic components. cTBS applied over the left BA 44
also changed the dynamics of chunk formation during
learning, as demonstrated by the network-based commu-
nity detection analysis showing that the evolution of the
“chunking magnitude” (φ) across sequences is strikingly
different following Broca’s area disruption.
As already mentioned, the behavioral signature of a
chunk is an increased processing time of its first boundary
item (Clerget et al., 2012; Pammi et al., 2012; Koechlin &
Jubault, 2006; Kennerley, Sakai, & Rushworth, 2004;
Rosenbaum et al., 1983), and in a strongly structured
hierarchical sequence such as the one used in the current
study, this difference in processing times is particularly
evident for stimuli occupying higher-level positions, espe-
cially Level 1 stimuli. This suggests that the relational
patterns in stimulus sequences can elicit low-level chunks,
which can be further integrated into superordinate chunks
to create a hierarchically organized sequence (Koch &
Hoffmann, 2000). In participants learning such a sequence
explicitly, we demonstrated that cTBS applied over the
Alamia et al.
411
left BA 44 only increases the processing time of stimuli
marking the boundaries of superordinate chunks, leaving
lower-level chunk processing unchanged, compared with
control. This finding corroborates the view that Broca’s
area plays a critical role in processing sequences (Fitch
& Martins, 2014; Opitz & Friederici, 2007; Schubotz &
Fiebach, 2006; Tettamanti & Weniger, 2006) and that its
contribution is all the more important in hierarchically
complex sequences, as previously suggested for both
linguistic (Grodzinsky & Santi, 2008; Rogalsky, Matchin,
& Hickok, 2008; Santi & Grodzinsky, 2007; Fiebach,
Schlesewsky, Lohmann, Von Cramon, & Friederici, 2005)
and nonlinguistic tasks (Clerget et al., 2013; Koechlin &
Jubault, 2006).
In the current study, it could be argued that cTBS
applied over Broca’s area interfered with higher-order
chunking processing because it requires more cognitive
resources, such as increased working memory or execu-
tive control demands. Indeed, when processing a higher-
order chunk, lower-order items need to be retrieved and
held in a buffer. Although a growing number of studies
have investigated the possible involvement of Broca’s
area in working memory in both linguistic and non-
linguistic tasks, a consensus on its role is still lacking.
On the one hand, some studies have concluded that
Broca’s area is involved in language processing via its
contribution to working memory (Nee et al., 2013;
Rogalsky et al., 2008; Fiebach et al., 2005; Smith, Jonides,
& Koeppe, 1996) or via complex interactions between
syntactic and working memory processes (Santi &
Grodzinsky, 2007; Kaan & Swaab, 2002). Some authors
have even suggested that Broca’s area implements a
“working memory buffer” shared among numerous cog-
nitive systems (including music, action, sequence process-
ing, and language), which could explain its involvement
in so many different cognitive tasks (Fitch & Martins,
2014). On the other hand, it is clear that several processes
operate on information stored in working memory,
namely encoding, maintenance, and retrieval ( Jonides
et al., 2008), and we are still a long way from fully under-
standing the contribution, if any, of the frontal cortex
and Broca’s area in particular to these mechanisms dur-
ing sequence learning. An important goal for future re-
search will be to investigate the relationship between
chunking and working memory, because these two pro-
cesses might be flip sides of the same coin. Another
potential confound in the interpretation of our results is
that the participants may have relied on inner speech to
maintain the sequence in memory (Baddeley, Gathercole,
& Papagno, 1998). Therefore, it could be argued that the
disruption of BA 44 impacted the task performance
through its effect on language. An argument against this
hypothesis is that TMS disruption led to a specific alter-
ation of higher-order chunking performance, whereas it
seems that alterations of linguistic processes should have
rather disrupted the global sequence learning perfor-
mance, irrespective of the hierarchical level. In addition,
we showed in a previous study that BA 44 disruption led
to behavioral alterations in an implicit learning task, in
which no inner speech could be involved (Clerget et al.,
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
/
t
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 7. Change in “chunking magnitude” across sequences. The φ values computed for each sequence (1–40) and averaged across participants are
shown for the two groups. A bilinear regression was computed for each set of group data. Error bars indicate SE.
412
Journal of Cognitive Neuroscience
Volume 28, Number 3
2012). However, excluding formally this alternative inter-
pretation will require further experiments.
In the current study, the contribution of left BA 44 to
chunking was also evidenced by a community detection
network analysis, showing that the two groups of par-
ticipants adopted a strikingly different chunking strategy
during sequence learning, as shown by the evolution of
the chunking magnitude (φ) across sequences. In the
control group, we found, in accordance with the work
of Grafton and colleagues ( Wymbs et al., 2012), that φ
increases gradually during sequence learning. In the
framework of motor sequence learning, chunking has
been theorized as a twofold dynamic process, relying
on distinct mechanisms. The first chunking operation,
called “segmentation,” leads to the parsing of the se-
quence into shorter clusters or “chunks” ( Wymbs et al.,
2012; Sakai et al., 2003; Verwey & Eikelboom, 2003;
Verwey, 2001; Koch & Hoffmann, 2000) and is character-
ized by a high modularity and therefore by a low φ value
(Wymbs et al., 2012). The second chunking process, the
“concatenation,” occurs probably at a later stage of learn-
ing and consists of integrating those elementary chunks
into superordinate chunks ( Wymbs et al., 2012; Sakai
et al., 2003; Koch & Hoffmann, 2000; Verwey, 1996), lead-
ing to a decrease in modularity, revealed by a higher φ
value ( Wymbs et al., 2012). The progressive φ increase
we observed in our control participants can thus be con-
strued as evidence for a gradual occurrence of conca-
tenation, although we found that φ did not increase
monotonically across sequences as reported originally
(Wymbs et al., 2012), but rather was augmented abruptly
in the second half of the training session, possibly cor-
responding to the moment when the participants reached
a plateau in terms of both error and RT (see Figure 4).
Interestingly, we found that left BA 44 cTBS altered the
φ change across sequence repetitions (see Figure 7): first,
cTBS yielded a much higher φ value than in controls at
the beginning of the training session, followed by a rapid
decrease during the first half of the experiment; second, φ
values remained more or less constant (or were even
slightly decreased) during the second half of the training
session. To understand this chunking strategy, it is impor-
tant to keep in mind that a high φ value suggests a diffi-
culty for the community detection algorithm in isolating
chunks, and although a high φ value at the end of training
has been regarded as evidence for concatenation (see
above), it could also be explained by a lack of or a very
low number of chunks in the sequence. Our interpreta-
tion is that following BA 44 disruption, the whole pro-
gression of the chunking processing was delayed, with
segmentation occurring later and concatenation failing
to happen within the time frame of the experiment. Alter-
natively, it could be proposed that the delayed pattern of
φ change across blocks in the BA 44 group was caused by
a disruption of learning rather than chunking specifically.
The observation that RT alterations were level specific
argues strongly against this hypothesis. However, it must
be pointed out that our task was designed to highlight the
role of BA 44 in chunking specifically and was very easy to
learn. Therefore, we cannot exclude that this region may
be important in other, more global, aspects of sequence
learning, as shown in a previous study (Clerget et al.,
2012). Finally, it is noteworthy that one limit of the model
used in the current study is that it assumes a constant
intersequence weight, whereas in fact it is likely to in-
crease over time. More complex models, accounting for
these changes over time, have been proposed recently
(Mucha, Richardson, Macon, Porter, & Onnela, 2010).
However, given that, in the present task, the sequences
were learned very quickly, we believe that using such a
model would not impact significantly on our results.
Regarding the neural correlates of chunking, although
many functional imaging studies have sought to identify
structures involved in sequence learning (Ashe et al., 2006;
Rhodes, Bullock, Verwey, Averbeck, & Page, 2004; Grafton,
Hazeltine, & Ivry, 1998; Toni, Krams, Turner, & Passingham,
1998), only very few studies have explicitly tried to identify
the neural correlates of chunking (Ruitenberg, Verwey,
Schutter, & Abrahamse, 2014; Pammi et al., 2012; Wymbs
et al., 2012; Kennerley et al., 2004; Verwey, Lammens, &
van Honk, 2002). An exception is the BG, for which a con-
tribution to chunking has already been demonstrated
( Wymbs et al., 2012; Tremblay et al., 2010; Boyd et al.,
2009; Tanji, 2001; Graybiel, 1998). As far as the involvement
of cortical areas in chunking is concerned, in a recent func-
tional imaging study Pammi and colleagues investigated
the effect of sequence complexity on the BOLD signal in
a frontoparietal network, including the VLPFC, and tried
to relate these changes to chunking (Pammi et al., 2012).
Increasing the sequence complexity led, in the early stages,
to a larger activation in bilateral VLPFC, including BA 44 and
BA 45. Then, in the intermediate learning stage, this acti-
vation moved leftward and more dorsally to the dorso-
lateral pFC. Because this increase in sequence complexity
was also accompanied by the emergence of chunking, the
authors speculated that these changes in BOLD signals in
VLPFC might be related to the encoding and execution of
chunks (Pammi et al., 2012). Moreover, Wymbs and col-
leagues interpreted the negative correlation that they
found between φ and the BOLD signal in the left fronto-
parietal network, including the left inferior frontal sulcus,
as evidence for its involvement in chunk segmentation,
contrasting with the positive correlation between φ and
activation of the bilateral sensorimotor putamen, inter-
preted as illustrative of its involvement in concatenation
(Wymbs et al., 2012). This study certainly implicates the left
inferior frontal gyrus in the chunking process but does not
allow us to isolate clearly its specific contribution to either
segmentation or concatenation. On the basis of the time
course of φ following cTBS, it seems that left BA 44 dis-
ruption delayed the sequence segmentation, and pre-
vented further concatenation. One possible explanation
is that concatenation did not occur in this study because
of delayed segmentation, the experiment duration being
Alamia et al.
413
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
too short to see the emergence of concatenation. The dif-
ficulty in reconciling these results illustrates the limitations
inherent in the decryption of the neural correlates of the
chunking process with functional neuroimaging tech-
niques because of the fluctuating contribution of different
brain structures during training (Pammi et al., 2012; Bassett
et al., 2011; Bapi, Miyapuram, Graydon, & Doya, 2006;
Sakai et al., 1998) and because chunking is, by definition,
a dynamic process (Clerget et al., 2012; Pammi et al., 2012),
which is generally highly variable between subjects
(Wymbs et al., 2012; Kennerley et al., 2004). Moreover, it
is likely that segmentation and concatenation are parallel
processes overlapping in time during learning, making it
even more difficult to disentangle them and to pinpoint
their respective neural correlates using functional MRI.
Despite decades of research, the role of Broca’s area is
still unclear and remains a source of debate. Indeed, the
finding that Broca’s area is involved in so many different
cognitive functions (Fitch & Martins, 2014; Clos, Amunts,
Laird, Fox, & Eickhoff, 2013; Fedorenko, Duncan, &
Kanwisher, 2012; Fadiga, Craighero, & D’Ausilio, 2009;
Grodzinsky & Santi, 2008; Nishitani, Schürmann, Amunts,
& Hari, 2005) has been seen as an argument for the
view that it implements a general, effector-independent
mechanism such as working memory, hierarchy, syntac-
tic processing, or possibly cognitive control. In contrast,
the contribution of Broca’s area to such a wide range of
cognitive tasks might imply functional heterogeneity and
its parcellation into specialized subregions (Fedorenko
et al., 2012), a view substantiated by a recent study of
the anatomical segregation of BA 44 and 45 (Amunts
et al., 2010). Our finding that Broca’s area plays a role
in processing higher-order chunks, supports the “broad-
spectrum mechanism” hypothesis, and is reminiscent of
the conclusions of Koechlin and Jubault (2006), who
proposed a model in which the anterior (BA 45) and pos-
terior (BA 44) parts of Broca’s area—and of its right
homolog—constitute, with the dorsal premotor cortex
(BA 6), a rostrocaudally organized network for processing
hierarchically structured sequences. One key conclusion
of that study was that motor behavior shares some simi-
larities with language and that a complex action can be
viewed as a chain of subordinate movements, which
need to be combined according to certain rules to reach
a given goal (Botvinick, 2008; Dominey et al., 2003;
Dehaene & Changeux, 1997). Sequence learning in par-
ticular appears to rely on cognitive processes that are
shared with language (Granena, 2013; Nemeth et al.,
2011; Conway, Karpicke, & Pisoni, 2007). On the basis of
the results of the current study, it is tempting to hypothe-
size that Broca’s area could be involved in higher-order
chunk processing whenever a serial stream of information
has to be processed (Pallier, Devauchelle, & Dehaene,
2011; Bapi, Doya, & Harner, 2000). Moreover, because
many studies on perception, learning, and cognition sup-
port the view that chunking is a universal information-
processing mechanism (Kurby & Zacks, 2008; Gobet et al.,
2001), it is tempting to hypothesize that chunking is a basic
mechanism that makes Broca’s area central to a large num-
ber of different cognitive and sensorimotor domains. Ver-
ifying the validity of this assumption will require further
experiments to prove that left BA 44 cTBS alters higher-
order chunking in tasks from different domains.
As already mentioned, besides Broca’s area, the other
structure commonly considered to be involved in chunk-
ing is the BG ( Jin, Tecuapetla, & Costa, 2014; Wymbs
et al., 2012; Boyd et al., 2009; Graybiel, 1998). Interestingly,
in addition to their well-known contribution to motor
sequence processing, BGs are also involved in language
syntax (Ackermann, Hage, & Ziegler, 2014; Chan, Ryan, &
Bever, 2013; Vargha-Khadem, Gadian, Copp, & Mishkin,
2005; Watkins & Dronkers, 2002; Alcock, Passingham,
Watkins, & Vargha-Khadem, 2000) and working memory
(Cools, 2011; Cools & D’Esposito, 2011; Baier et al.,
2010), a combination of functions strikingly similar to those
assigned to Broca’s area. Details are still lacking on how
Broca’s area interacts with the BG (Ullman, 2006), but this
further highlights the possible connections between
working memory, syntax, and chunking (Zénon & Olivier,
2014). It also emphasizes the tight interaction between
action-related and higher cognitive functions (Mendoza &
Merchant, 2014; Andres, Olivier, & Badets, 2008; Olivier,
Davare, Andres, & Fadiga, 2007), supporting the view that
the latter have invaded evolutionarily older motor brain
circuits during the course of evolution (Dehaene & Cohen,
2007).
Acknowledgments
This work was performed at the Institute of Neuroscience of
the Université catholique de Louvain (Brussels, Belgium); it was
supported by grants from the ARC (Actions de Recherche
Concertées, Communauté Française de Belgique), the Fondation
Médicale Reine Elisabeth (FMRE), and the Fonds de la Recherche
Scientifique (FNRS-FDP) to E. O., by EU grants Poeticon++ and
Siempre, and by the E.R. Region-University fund to L. F. A. A. is
a Research Fellow at the FNRS and A. Z. is a Senior Research
Associate supported by INNOVIRIS. The authors appreciate the
assistance of Emeline Clerget in running some subjects. We are
also grateful to Benvenuto Jacob for his help in programming,
to Laurence Dricot for processing the TMS site locations, and
to Vincent Blondel for his advice on data analysis.
Reprint requests should be sent to Prof. Etienne Olivier, Institute
of Neuroscience, Université catholique de Louvain, 53, Avenue
Mounier, COSY-B1.53.04, 1200 Brussels, Belgium, or via e-mail:
etienne.olivier@uclouvain.be.
REFERENCES
Ackermann, H., Hage, S. R., & Ziegler, W. (2014). Brain
mechanisms of acoustic communication in humans
and nonhuman primates: An evolutionary perspective.
Behavioral and Brain Sciences, 37, 1–84.
Alcock, K. J., Passingham, R. E., Watkins, K. E., & Vargha-
Khadem, F. (2000). Oral dyspraxia in inherited speech and
language impairment and acquired dysphasia. Brain and
Language, 75, 17–33.
414
Journal of Cognitive Neuroscience
Volume 28, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
/
f
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Amunts, K., Lenzen, M., Friederici, A. D., Schleicher, A.,
Morosan, P., Palomero-Gallagher, N., et al. (2010). Broca’s
region: Novel organizational principles and multiple receptor
mapping. PLoS Biology, 8, e1000489.
Amunts, K., Weiss, P. H., Mohlberg, H., Pieperhoff, P., Eickhoff,
S., Gurd, J. M., et al. (2004). Analysis of neural mechanisms
underlying verbal fluency in cytoarchitectonically defined
stereotaxic space—The roles of Brodmann areas 44 and 45.
Neuroimage, 22, 42–56.
Andres, M., Olivier, E., & Badets, A. (2008). Actions, words, and
numbers: A motor contribution to semantic processing?
Current Directions in Psychological Science, 17, 313–317.
Ashe, J., Lungu, O. V., Basford, A. T., & Lu, X. (2006). Cortical
control of motor sequences. Current Opinion in
Neurobiology, 16, 213–221.
Baayen, R. H., Davidson, D. J., & Bates, D. M. (2008).
Mixed-effects modeling with crossed random effects for
subjects and items. Journal of Memory and Language,
59, 390–412.
Baddeley, A., Gathercole, S., & Papagno, C. (1998). The
phonological loop as a language learning device.
Psychological Review, 105, 158–173.
Bahlmann, J., Schubotz, R. I., Mueller, J. L., Koester, D., &
Friederici, A. D. (2009). Neural circuits of hierarchical
visuo-spatial sequence processing. Brain Research,
1298, 161–170.
Baier, B., Karnath, H.-O., Dieterich, M., Birklein, F., Heinze, C.,
& Müller, N. G. (2010). Keeping memory clear and
stable—The contribution of human basal ganglia and
prefrontal cortex to working memory. Journal of
Neuroscience, 30, 9788–9792.
Bapi, R. S., Doya, K., & Harner, A. M. (2000). Evidence for
effector independent and dependent representations and
their differential time course of acquisition during motor
sequence learning. Experimental Brain Research,
132, 149–162.
Bapi, R. S., Miyapuram, K. P., Graydon, F. X., & Doya, K. (2006).
fMRI investigation of cortical and subcortical networks in the
learning of abstract and effector-specific representations of
motor sequences. Neuroimage, 32, 714–727.
Barrat, A., Barthélemy, M., Pastor-Satorras, R., & Vespignani, A.
(2004). The architecture of complex weighted networks.
Proceedings of the National Academy of Sciences, U.S.A.,
101, 3747–3752.
Bassett, D. S., Porter, M. A., Wymbs, N. F., Grafton, S. T.,
Carlson, J. M., & Mucha, P. J. (2013). Robust detection of
dynamic community structure in networks. Chaos, 23, 1–16.
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T. (2011). Dynamic reconfiguration of
human brain networks during learning. Proceedings of the
National Academy of Sciences, U.S.A., 108, 7641–7646.
Blondel, V. D., Guillaume, J.-L., Lambiotte, R., & Lefebvre, E.
(2008). Fast unfolding of communities in large networks.
Journal of Statistical Mechanics: Theory and Experiment,
P10008. doi:10.1088/1742-5468/2008/10/P10008.
Bor, D., Duncan, J., Wiseman, R. J., & Owen, A. M. (2003).
Encoding strategies dissociate prefrontal activity from
working memory demand. Neuron, 37, 361–367.
Bor, D., & Seth, A. K. (2012). Consciousness and the prefrontal
parietal network: Insights from attention, working memory,
and chunking. Frontiers in Psychology, 3, 1–14.
Chan, S.-H., Ryan, L., & Bever, T. G. (2013). Role of the striatum
in language: Syntactic and conceptual sequencing. Brain
and Language, 125, 283–294.
Chung, H.-K., Tsai, C.-H., Lin, Y.-C., Chen, J.-M., Tsou, Y.-A.,
Wang, C.-Y., et al. (2012). Effectiveness of theta-burst
repetitive transcranial magnetic stimulation for treating
chronic tinnitus. Audiology & Neuro-Otology, 17, 112–120.
Clerget, E., Andres, M., & Olivier, E. (2013). Deficit in complex
sequence processing after a virtual lesion of left BA 45.
PLoS One, 8, 1–9.
Clerget, E., Badets, A., Duqué, J., & Olivier, E. (2011). Role of
Broca’s area in motor sequence programming: A cTBS study.
NeuroReport, 22, 965–969.
Clerget, E., Poncin, W., Fadiga, L., & Olivier, E. (2012). Role of
Broca’s area in implicit motor skill learning: Evidence from
continuous theta-burst magnetic stimulation. Journal of
Cognitive Neuroscience, 24, 80–92.
Clerget, E., Winderickx, A., Fadiga, L., & Olivier, E. (2009). Role
of Broca’s area in encoding sequential human actions: A
virtual lesion study. NeuroReport, 20, 1496–1499.
Clos, M., Amunts, K., Laird, A. R., Fox, P. T., & Eickhoff, S. B.
(2013). Tackling the multifunctional nature of Broca’s region
meta-analytically: Co-activation-based parcellation of area 44.
Neuroimage, 83, 174–188.
Conway, C. M., & Christiansen, M. H. (2001). Sequential
learning in non-human primates learning fixed sequences.
Trends in Cognitive Sciences, 5, 539–546.
Conway, C. M., Karpicke, J., & Pisoni, D. B. (2007). Contribution
of implicit sequence learning to spoken language processing:
Some preliminary findings with hearing adults. Journal of
Deaf Studies and Deaf Education, 12, 317–334.
Cools, R. (2011). Dopaminergic control of the striatum for
high-level cognition. Current Opinion in Neurobiology,
21, 402–407.
Cools, R., & D’Esposito, M. (2011). Inverted-U-shaped dopamine
actions on human working memory and cognitive control.
Biological Psychiatry, 69, e113–e125.
Corballis, M. C. (2003). From mouth to hand: Gesture, speech,
and the evolution of right-handedness. Behavioral and
Brain Sciences, 26, 199–208; discussion 208–260.
Davare, M., Andres, M., Cosnard, G., Thonnard, J.-L., & Olivier,
E. (2006). Dissociating the role of ventral and dorsal
premotor cortex in precision grasping. Journal of
Neuroscience, 26, 2260–2268.
Dehaene, S., & Changeux, J. P. (1997). A hierarchical neuronal
network for planning behavior. Proceedings of the National
Academy of Sciences, U.S.A., 94, 13293–13298.
Dehaene, S., & Cohen, L. (2007). Cultural recycling of cortical
maps. Neuron, 56, 384–398.
Dominey, P. F., Hoen, M., Blanc, J. M., & Lelekov-Boissard, T.
(2003). Neurological basis of language and sequential
cognition: Evidence from simulation, aphasia, and ERP
studies. Brain and Language, 86, 207–225.
Doyon, J., Bellec, P., Amsel, R., Penhune, V., Monchi, O.,
Carrier, J., et al. (2009). Contributions of the basal ganglia and
functionally related brain structures to motor learning.
Behavioural Brain Research, 199, 61–75.
Eickhoff, S. B., Paus, T., Caspers, S., Grosbras, M. H., Evans,
A. C., Zilles, K., et al. (2007). Assignment of functional
activations to probabilistic cytoarchitectonic areas revisited.
Neuroimage, 36, 511–521.
Botvinick, M. M. (2008). Hierarchical models of behavior and
Eickhoff, S. B., Stephan, K. E., Mohlberg, H., Grefkes, C.,
prefrontal function. Trends in Cognitive Sciences,
12, 201–208.
Boyd, L. A., Edwards, J. D., Siengsukon, C. S., Vidoni, E. D.,
Wessel, B. D., & Linsdell, M. A. (2009). Motor sequence
chunking is impaired by basal ganglia stroke. Neurobiology of
Learning and Memory, 92, 35–44.
Fink, G. R., Amunts, K., et al. (2005). A new SPM toolbox
for combining probabilistic cytoarchitectonic maps and
functional imaging data. Neuroimage, 25, 1325–1335.
Fadiga, L., Craighero, L., & D’Ausilio, A. (2009). Broca’s area
in language, action, and music. Annals of the New York
Academy of Sciences, 1169, 448–458.
Alamia et al.
415
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Fazio, P., Cantagallo, A., Craighero, L., D’Ausilio, A., Roy, A. C.,
Pozzo, T., et al. (2009). Encoding of human action in Broca’s
area. Brain, 132, 1980–1988.
Fedorenko, E., Duncan, J., & Kanwisher, N. (2012). Language-
selective and domain-general regions lie side by side within
Broca’s area. Current Biology, 22, 2059–2062.
Fiebach, C. J., Schlesewsky, M., Lohmann, G., Von Cramon,
D. Y., & Friederici, A. D. (2005). Revisiting the role of Broca’s
area in sentence processing: Syntactic integration versus
syntactic working memory. Human Brain Mapping,
24, 79–91.
Fitch, W. T., & Martins, M. D. (2014). Hierarchical processing
in music, language, and action: Lashley revisited. Annals of
the New York Academy of Sciences, 1316, 87–104.
Gobet, F., Lane, P. C. R., Croker, S., Cheng, P. C., Jones, G.,
Oliver, I., et al. (2001). Chunking mechanisms in human
learning, Trends in Cognitive Sciences, 5, 236–243.
Goldsworthy, M. R., Pitcher, J. B., & Ridding, M. C. (2012).
A comparison of two different continuous theta burst
stimulation paradigms applied to the human primary motor
cortex. Clinical Neurophysiology, 123, 2256–2263.
Grafton, S. T., Hazeltine, E., & Ivry, R. B. (1998). Abstract and
effector-specific representations of motor sequences
identified with PET. Journal of Neuroscience, 18, 9420–9428.
Granena, G. (2013). Individual differences in sequence learning
ability and second language acquisition in early childhood
and adulthood. Language Learning, 63, 665–703.
Keele, S. W., Ivry, R., Mayr, U., Hazeltine, E., & Heuer, H.
(2003). The cognitive and neural architecture of sequence
representation. Psychological Review, 110, 316–339.
Kennerley, S. W., Sakai, K., & Rushworth, M. F. S. (2004).
Organization of action sequences and the role of the
pre-SMA. Journal of Neurophysiology, 91, 978–993.
Koch, I., & Hoffmann, J. (2000). Patterns, chunks, and
hierarchies in serial reaction-time tasks. Psychological
Research, 63, 22–35.
Koechlin, E., & Jubault, T. (2006). Broca’s area and the
hierarchical organization of human behavior. Neuron,
50, 963–974.
Kühn, A. (2011). Neuronale Korrelate der Vorhersage
hierarchischer Stimulussequenzen. Rheinisch-Westfälischen
Technischen Hochschule Aachen.
Kurby, C. A., & Zacks, J. M. (2008). Segmentation in the
perception and memory of events. Trends in Cognitive
Sciences, 12, 72–79.
Mathy, F., & Feldman, J. (2012). What’s magic about magic
numbers? Chunking and data compression in short-term
memory. Cognition, 122, 346–362.
Mendoza, G., & Merchant, H. (2014). Motor system evolution
and the emergence of high cognitive functions. Progress in
Neurobiology, 122, 73–93.
Mucha, P. J., Richardson, T., Macon, K., Porter, M. A., & Onnela,
J.-P. (2010). Community structure in time-dependent,
multiscale, and multiplex networks. Science, 328, 876–879.
Graybiel, A. M. (1998). The basal ganglia and chunking of action
Nee, D. E., Brown, J. W., Askren, M. K., Berman, M. G.,
repertoires. Neurobiology of Learning and Memory,
136, 119–136.
Grodzinsky, Y., & Santi, A. (2008). The battle for Broca’s region.
Trends in Cognitive Sciences, 12, 474–480.
Grossman, M. (1980). A central processor for hierarchically-
structured material: Evidence from Broca’s aphasia.
Neuropsychologia, 18, 299–308.
Heim, S., Eickhoff, S. B., & Amunts, K. (2009). Different roles of
cytoarchitectonic BA 44 and BA 45 in phonological and
semantic verbal fluency as revealed by dynamic causal
modelling. Neuroimage, 48, 616–624.
Hikosaka, O., Nakamura, K., Sakai, K., & Nakahara, H. (2002).
Central mechanisms of motor skill learning. Current Opinion
in Neurobiology, 12, 217–222.
Huang, G., & Mouraux, A. (2015). MEP latencies predict
the neuromodulatory effect of cTBS delivered to the
ipsilateral and contralateral sensorimotor cortex. Plos One,
10, e0133893.
Huang, Y. Z., Edwards, M. J., Rounis, E., Bhatia, K. P., &
Rothwell, J. C. (2005). Theta burst stimulation of the human
motor cortex. Neuron, 45, 201–206.
Huntley, J., Bor, D., Hampshire, A., Owen, A., & Howard, R.
(2011). Working memory task performance and chunking in
early Alzheimer’s disease. British Journal of Psychiatry, 198,
398–403.
Janata, P., & Grafton, S. T. (2003). Swinging in the brain: Shared
neural substrates for behaviors related to sequencing and
music. Nature Neuroscience, 6, 682–687.
Jin, X., Tecuapetla, F., & Costa, R. M. (2014). Basal ganglia
subcircuits distinctively encode the parsing and
concatenation of action sequences. Nature Neuroscience,
17, 423–430.
Jonides, J., Lewis, R. L., Nee, D. E., Lustig, C. A., Berman, M. G.,
& Moore, K. S. (2008). The mind and brain of short-term
memory. Annual Review of Psychology, 59, 193–224.
Kaan, E., & Swaab, T. Y. (2002). The brain circuitry of syntactic
comprehension. Trends in Cognitive Sciences, 6, 350–356.
Keel, J. C., Smith, M. J., & Wassermann, E. M. (2001). A safety
screening questionnaire for transcranial magnetic
stimulation. Clinical Neurophysiology, 112, 720.
Demiralp, E., Krawitz, A., et al. (2013). A meta-analysis of
executive components of working memory. Cerebral Cortex,
23, 264–282.
Nemeth, D., Janacsek, K., Csifcsak, G., Szvoboda, G., Howard,
J. H., & Howard, D. V. (2011). Interference between sentence
processing and probabilistic implicit sequence learning.
PLoS One, 6, 6–11.
Newman, M., & Girvan, M. (2004). Finding and evaluating
community structure in networks. Physical Review E,
69, 026113.
Nishitani, N., Schürmann, M., Amunts, K., & Hari, R. (2005).
Broca’s region: From action to language. Physiology, 20,
60–69.
Noirhomme, Q., Ferrant, M., Vandermeeren, Y., Olivier, E.,
Macq, B., & Cuisenaire, O. (2004). Registration and real-time
visualization of transcranial magnetic stimulation with 3-D
MR images. IEEE Transactions on Bio-Medical Engineering,
51, 1994–2005.
Nyffeler, T., Wurtz, P., Lüscher, H. R., Hess, C. W., Senn, W.,
Pflugshaupt, T., et al. (2006). Repetitive TMS over the human
oculomotor cortex: Comparison of 1-Hz and theta burst
stimulation. Neuroscience Letters, 409, 57–60.
Okamoto, M., Dan, H., Sakamoto, K., Takeo, K., Shimizu, K.,
Kohno, S., et al. (2004). Three-dimensional probabilistic
anatomical cranio-cerebral correlation via the international
10-20 system oriented for transcranial functional brain
mapping. Neuroimage, 21, 99–111.
Oldfield, R. C. (1971). The assessment and analysis of
handedness: The Edinburgh inventory. Neuropsychologia,
9, 97–113.
Olivier, E., Davare, M., Andres, M., & Fadiga, L. (2007). Precision
grasping in humans: From motor control to cognition.
Current Opinion in Neurobiology, 17, 644–648.
Opitz, B., & Friederici, A. D. (2007). Neural basis of processing
sequential and hierarchical syntactic structures. Human
Brain Mapping, 28, 585–592.
Pallier, C., Devauchelle, A.-D., & Dehaene, S. (2011). Cortical
representation of the constituent structure of sentences.
Proceedings of the National Academy of Sciences, U.S.A.,
108, 2522–2527.
416
Journal of Cognitive Neuroscience
Volume 28, Number 3
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
f
/
t
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Pammi, V. S. C., Miyapuram, K. P., Ahmed, Samejima, K.,
Bapi, R. S., & Doya, K. (2012). Changing the structure
of complex visuo-motor sequences selectively activates the
fronto-parietal network. Neuroimage, 59, 1180–1189.
Penhune, V. B., & Steele, C. J. (2012). Parallel contributions
of cerebellar, striatal and M1 mechanisms to motor
sequence learning. Behavioural Brain Research, 226,
579–591.
Perruchet, P., & Pacton, S. (2006). Implicit learning and
statistical learning: One phenomenon, two approaches.
Trends in Cognitive Sciences, 10, 233–238.
Rhodes, B. J., Bullock, D., Verwey, W. B., Averbeck, B. B.,
& Page, M. P. A. (2004). Learning and production of
movement sequences: Behavioral, neurophysiological, and
modeling perspectives. Human Movement Science, 23,
699–746.
Rogalsky, C., Matchin, W., & Hickok, G. (2008). Broca’s area,
sentence comprehension, and working memory: An fMRI
study. Frontiers in Human Neuroscience, 2, 14.
Rosenbaum, D. A., Kenny, S. B., & Derr, M. A. (1983).
Hierarchical control of rapid movement sequences. Journal
of Experimental Psychology. Human Perception and
Performance, 9, 86–102.
Rossini, P. M., Barker, A. T., Berardelli, A., Caramia, M. D.,
Caruso, G., Cracco, R. Q., et al. (1994). Non-invasive
electrical and magnetic stimulation of the brain,
spinal cord and roots: Basic principles and procedures
for routine clinical application. Report of an IFCN committee.
Electroencephalography and Clinical Neurophysiology,
91, 79–92.
Toni, I., Krams, M., Turner, R., & Passingham, R. E. (1998).
The time course of changes during motor sequence learning:
A whole-brain fMRI study. Neuroimage, 61, 50–61.
Torta, D. M. E., Legrain, V., Algoet, M., Olivier, E., Duque, J.,
& Mouraux, A. (2013). Theta burst stimulation applied
over primary motor and somatosensory cortices produces
analgesia unrelated to the changes in nociceptive
event-related potentials. PLoS One, 8, 1–15.
Tremblay, P. L., Bedard, M. A., Langlois, D., Blanchet, P. J.,
Lemay, M., & Parent, M. (2010). Movement chunking during
sequence learning is a dopamine-dependant process: A study
conducted in Parkinson’s disease. Experimental Brain
Research, 205, 375–385.
Ullman, M. T. (2006). Is Broca’s area part of a basal ganglia
thalamocortical circuit? Cortex, 42, 480–485.
Vargha-Khadem, F., Gadian, D. G., Copp, A., & Mishkin, M.
(2005). FOXP2 and the neuroanatomy of speech and
language. Nature Reviews Neuroscience, 6, 131–138.
Verwey, W., & Abrahamse, E. L. (2012). Distinct modes of
executing movement sequences: Reacting, associating,
and chunking. Acta Psychologica, 140, 274–282.
Verwey, W. B. (1996). Buffer loading and chunking in sequential
keypressing. Journal of Experimental Psychology: Human
Perception and Performance, 22, 544–562.
Verwey, W. B. (2001). Concatenating familiar movement
sequences: The versatile cognitive processor. Acta
Psychologica, 106, 69–95.
Verwey, W. B. (2010). Diminished motor skill development in
elderly: Indications for limited motor chunk use. Acta
Psychologica, 134, 206–214.
Ruitenberg, M. F. L., Verwey, W. B., Schutter, D. J. L. G., &
Verwey, W. B., & Eikelboom, T. (2003). Evidence for lasting
Abrahamse, E. L. (2014). Cognitive and neural foundations
of discrete sequence skill: A TMS study. Neuropsychologia,
56, 229–238.
Sakai, K., Hikosaka, O., Miyauchi, S., Takino, R., Sasaki, Y., &
Pütz, B. (1998). Transition of brain activation from frontal to
parietal areas in visuomotor sequence learning. Journal of
Neuroscience, 18, 1827–1840.
Sakai, K., Kitaguchi, K., & Hikosaka, O. (2003). Chunking during
human visuomotor sequence learning. Experimental Brain
Research, 152, 229–242.
Santi, A., & Grodzinsky, Y. (2007). Working memory and
syntax interact in Broca’s area. Neuroimage, 37, 8–17.
Schubotz, R. I., & Fiebach, C. J. (2006). Integrative models of
Broca’s area and the ventral premotor cortex. Cortex,
42, 461–463.
Smith, E. E., Jonides, J., & Koeppe, R. A. (1996). Dissociating
verbal and spatial working memory using PET. Cerebral
Cortex, 6, 11–20.
Tanji, J. (2001). Sequential organization of multiple movements:
Involvement of cortical motor areas. Annual Review of
Neuroscience, 24, 631–651.
Tettamanti, M., & Weniger, D. (2006). Broca’s area: A
supramodal hierarchical processor? Cortex, 42, 491–494.
sequence segmentation in the discrete sequence-production
task. Journal of Motor Behavior, 35, 171–181.
Verwey, W. B., Lammens, R., & van Honk, J. (2002). On the role
of the SMA in the discrete sequence production task: A TMS
study. Transcranial magnetic stimulation. Neuropsychologia,
40, 1268–1276.
Watkins, K. E., & Dronkers, N. F. (2002). Behavioural analysis
of an inherited speech and language disorder: Comparison
with acquired aphasia. Brain, 125, 452–464.
Wymbs, N. F., Bassett, D. S., Mucha, P. J., Porter, M. A., &
Grafton, S. T. (2012). Differential recruitment of the
sensorimotor putamen and frontoparietal cortex during
motor chunking in humans. Neuron, 74, 936–946.
Zénon, A., & Olivier, E. (2014). Contribution of the basal ganglia
to the spoken language: Is speech production like the other
motor skills? Behavioral and Brain Sciences, 37, 576.
Zénon, A., Sidibé, M., & Olivier, E. (2015). Disrupting the
supplementary motor area makes physical effort appear
less effortful. Journal of Neuroscience, 35, 8737–8744.
Zosso, D., Noirhomme, Q., Davare, M., MacQ, B., Olivier, E.,
Thiran, J., et al. (2006). Normalization of transcranial
magnetic stimulation points by means of atlas registration.
European Signal Processing Conference, 2–6.
Alamia et al.
417
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
8
/
3
2
8
4
/
0
3
2
/
1
4
9
0
5
2
0
/
7
1
0
7
7
8
o
4
c
8
n
0
_
7
a
/
_
j
0
o
0
c
9
n
1
1
_
a
p
_
d
0
0
b
9
y
1
g
1
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
i
2
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1