Decisions Made with Less Evidence Involve Higher Levels
of Corticosubthalamic Nucleus Theta Band Synchrony
Baltazar Zavala1,2, Huiling Tan1,3, Simon Little4, Keyoumars Ashkan5,
Alexander L. Green1,3, Tipu Aziz1,3, Thomas Foltynie4, Ludvic Zrinzo4,
Kareem Zaghloul2, and Peter Brown1,3
Astratto
■ The switch between automatic action selection and more
controlled forms of decision-making is a dynamic process
thought to involve both cortical and subcortical structures.
During sensory conflict, medial pFC oscillations in the theta
band (<8 Hz) drive those of the subthalamic nucleus (STN),
and this is thought to increase threshold evidence needed
for one competing response be selected over another. Here,
we were interested in testing whether STN activity also altered
by rate at which presented during a congruent
dot motion task absent any explicit sensory conflict. By having
a series randomly moving dots gradually transform con-
gruent three different rates (slow, medium, fast), we
were able show that slower increased time it took
participants make but did not alter total
amount was integrated before response.
Notably, resulted decision being made with lower
amount instantaneous slow and medium
trials. Consistent idea medial pFC–STN is
involved executing cognitive control, higher levels of
ambiguity these trials associated increased
theta band synchrony between cortex STN, with
the cortical oscillations Granger-causal STN.
These results further confirm involvement in
decision-making suggest disruption network
may underlie some unwanted deficits associated
with deep brain stimulation. ■
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
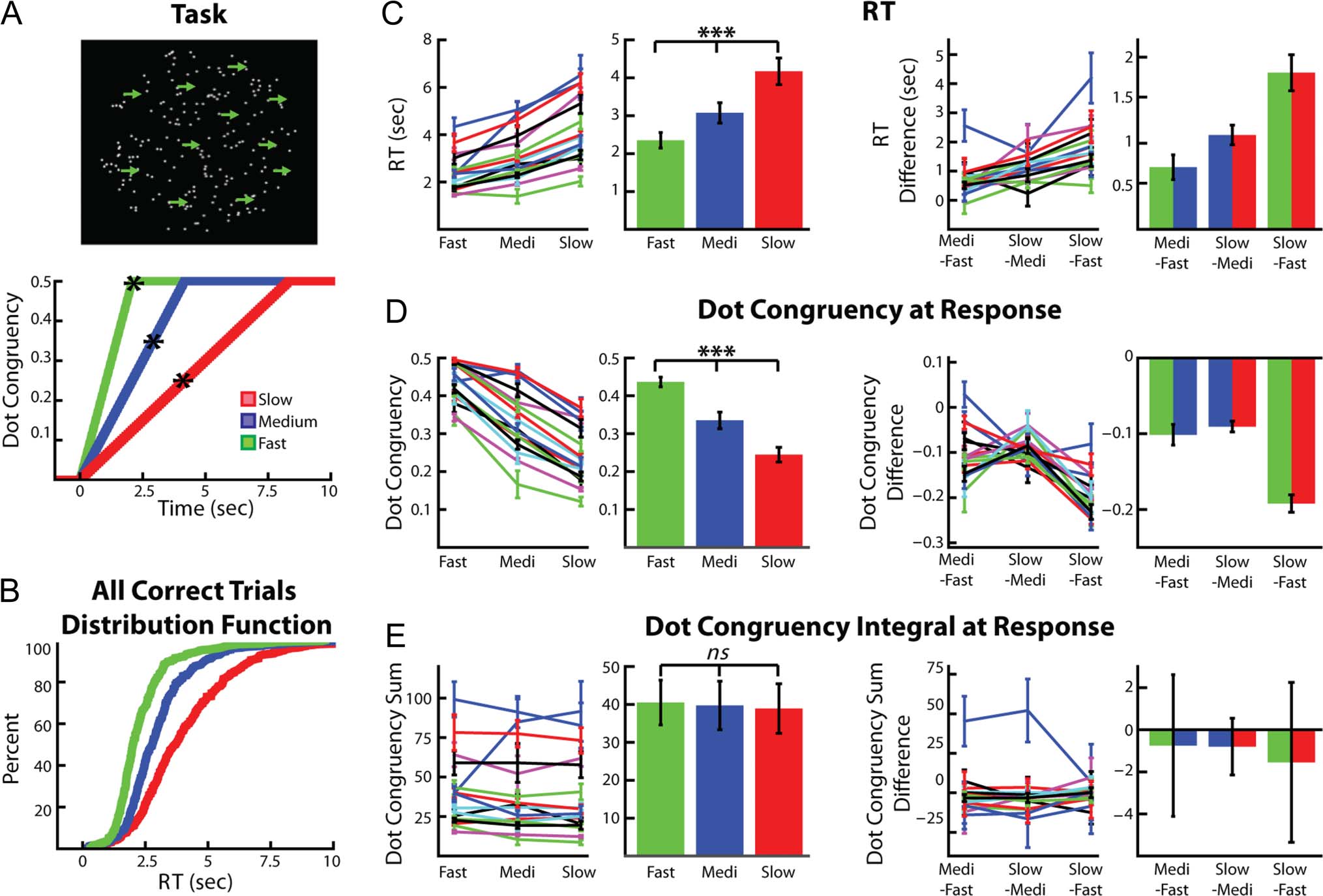
> .88). This sug-
gests that, although evidence was presented to the par-
ticipants at a slower rate during the slow and medium
trials, the longer time period of evidence integration during
these trials allowed the participants to ultimately respond
with a lower degree of instantaneous dot motion congru-
ency then they did during the fast trials. The result of this,
Tuttavia, was that the medium and slow trial responses
were made when the ambiguity in the stimulus was higher
than that experienced at the time of response for the fast
trials.
To confirm that the differences in instantaneous dot
congruency at the time of the response were not due to
the more rapid dot congruency increases during any
“nondecision time” in between when the participants
made their decision and when they executed their re-
sponse, we also calculated the average dot congruency
500 msec before the participants made their response. These
values were 36.3 ± 2.1%, 28.4 ± 2.4%, E 21.6 ± 1.2%
for fast, medium, and slow trials (ANOVA, within-subject
repeated-measures, F = 88.9, df = 2, P < .0001), respec-
tively. This suggests that the differences were not due to
dot congruency increases after the decision was made
(but before the response was executed) and that the par-
ticipants did indeed make their decision with less instan-
taneous “evidence” during the slow and medium trials
relative to the fast trials. This assumes a nondecision time
of 500 msec, which is much longer than that typically
assumed in the literature (Ratcliff & Frank, 2012; Ratcliff
& McKoon, 2008; Ratcliff & Tuerlinckx, 2002). Similarly
significant results were also observed if we assumed a non-
decision time of 1 sec (ANOVA, within-subject repeated-
measures, F = 23.4, df = 2, p < .0001).
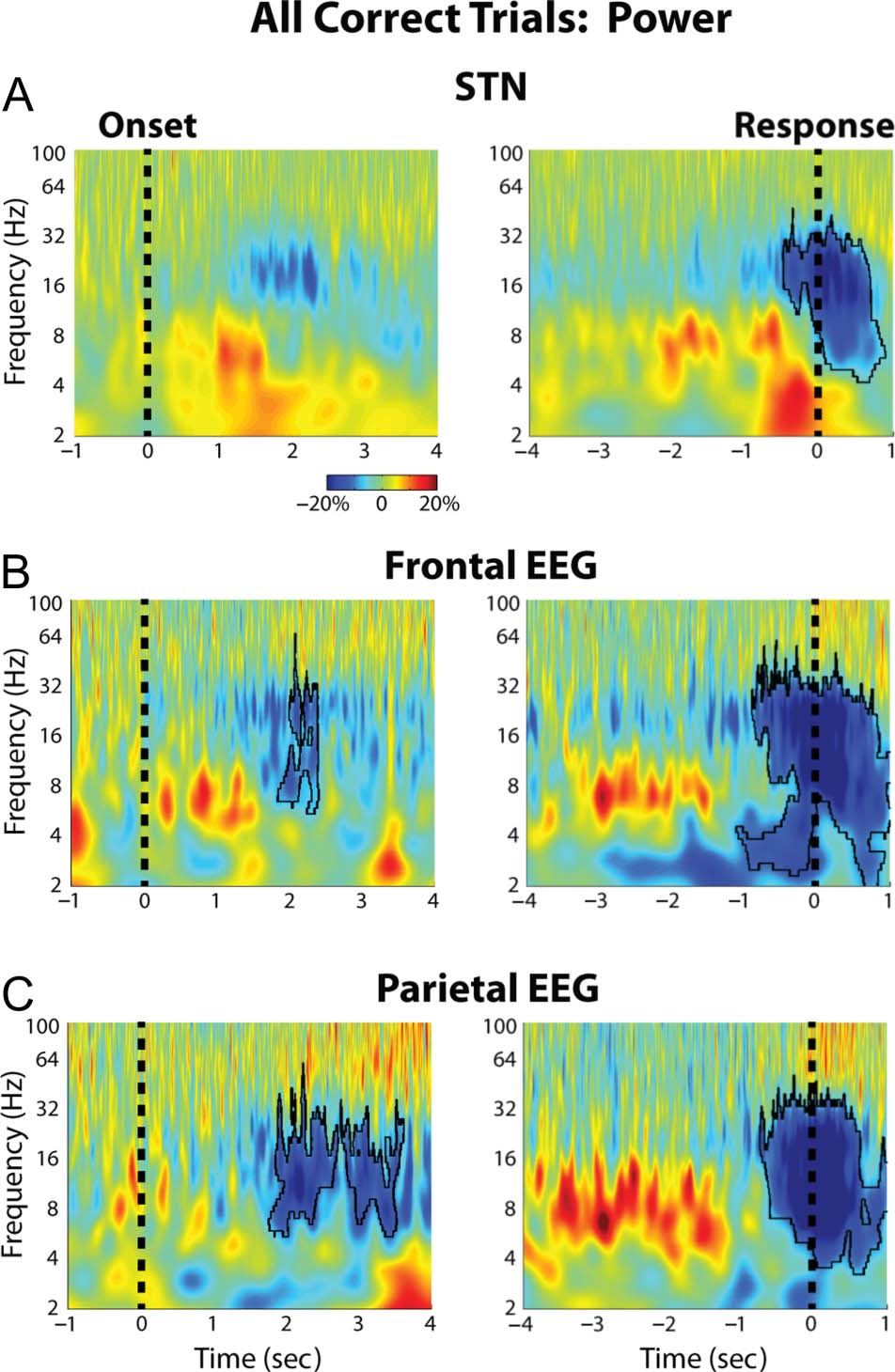
Power and ITPC Changes during the Task
Figure 2 shows the average changes in power that oc-
curred during the task for the STN and for two midline
EEG cortical recordings in the frontal (FCz-Cz) and pari-
etal cortex (Pz-Cz). Although the spectrograms suggest
an increase in STN theta and delta band activity during
the task, only the response-aligned decreases in beta band
activity survived correction for multiple comparisons ( p =
.008, permutation testing). Similarly, in the frontal and
parietal cortex, only the cue and response aligned beta
changes were significantly different from baseline ( p <
.001, permutation testing). These significant changes from
baseline were observed when all trials were averaged
together, but no significant trial type-related differences
in power were observed during the task ( p > .05, permu-
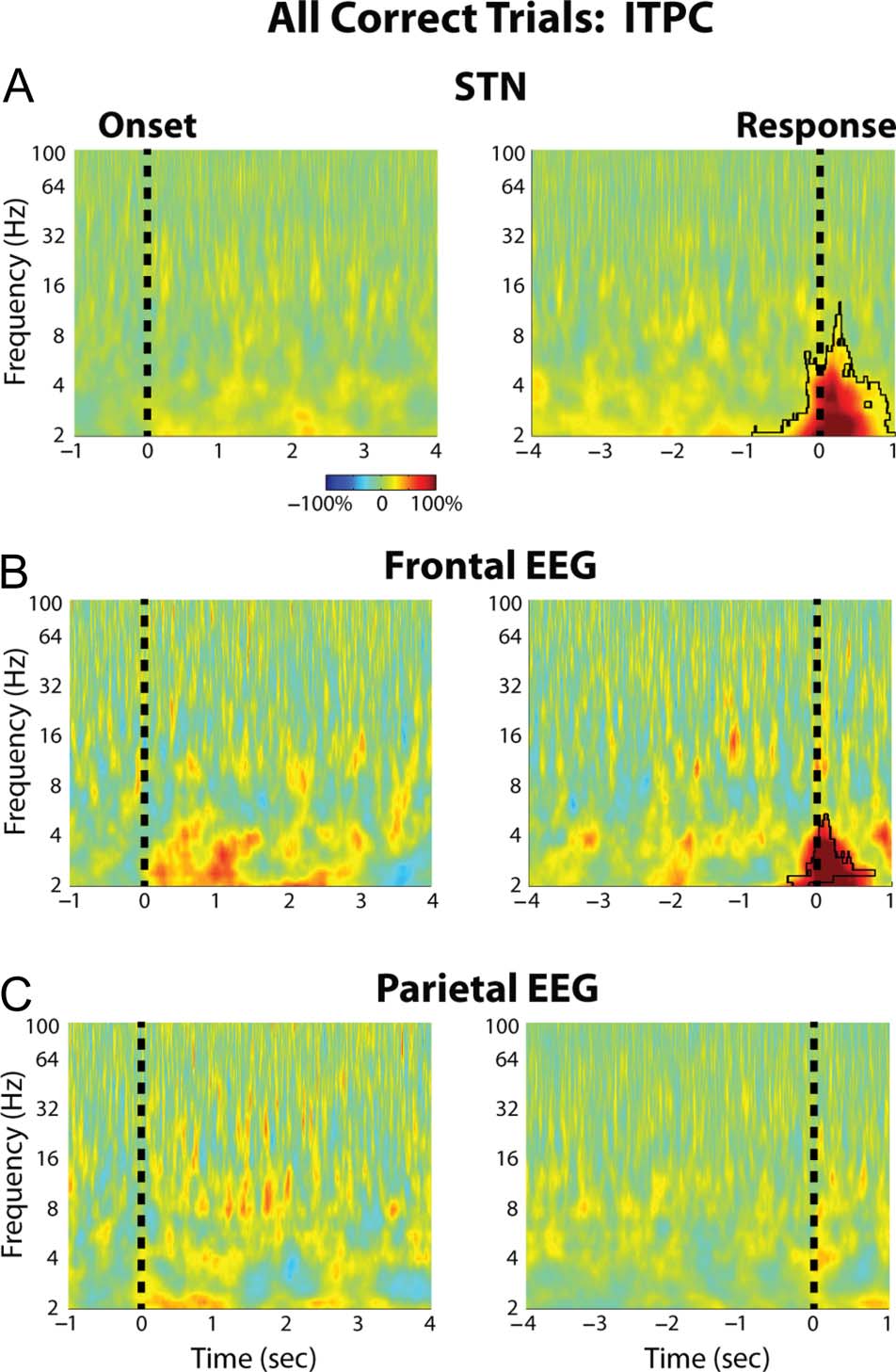
tation testing). ITPC was also analyzed during the task, E
this revealed a significant increase in delta band ITPC at the
time of the response for the STN and the midline frontal
cortex recordings (Figura 3). Once again, Tuttavia, there
were no trial type-related differences in ITPC during the
task ( p > .05, permutation testing).
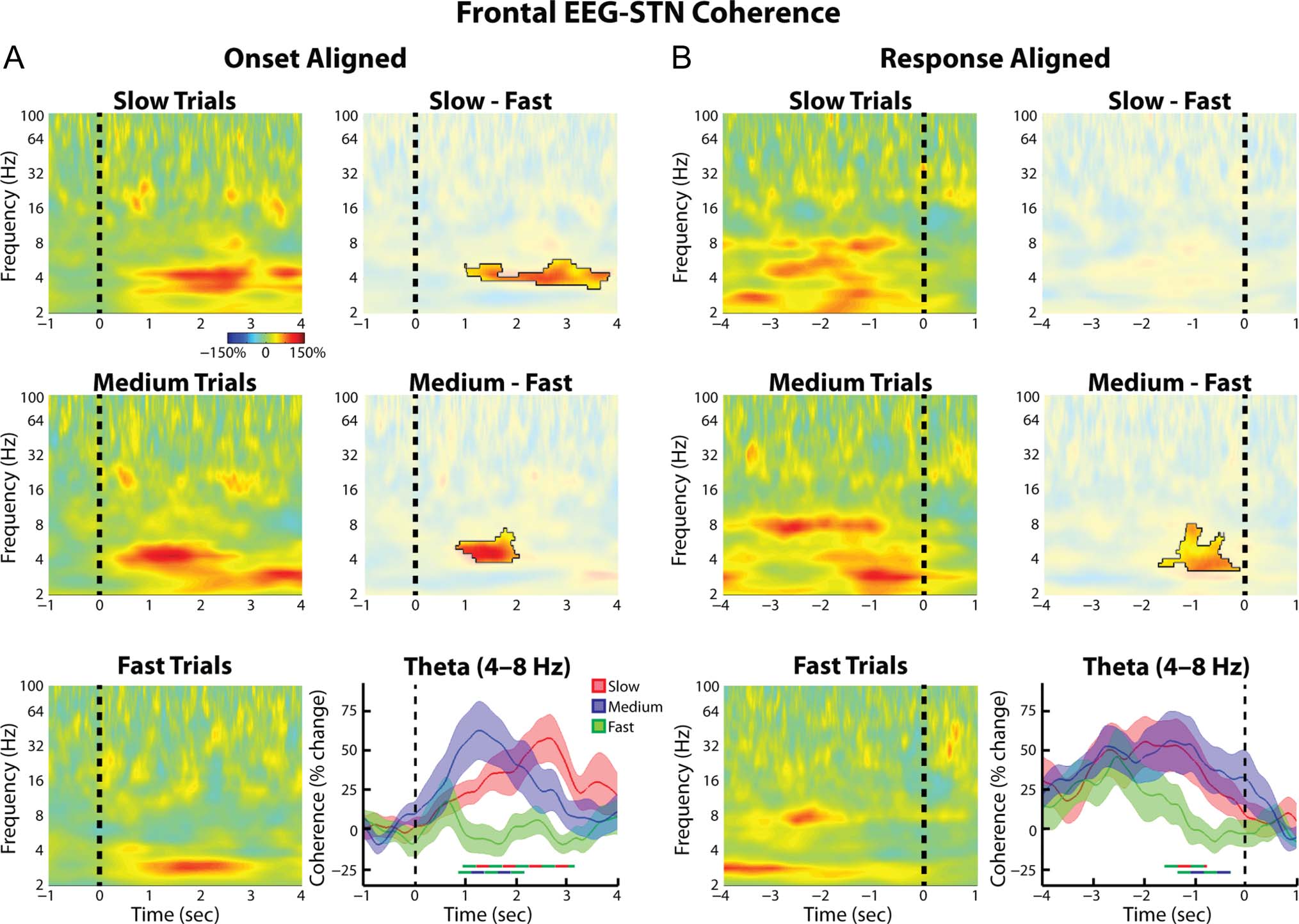
Frontal EEG–STN Synchrony Changes
during the Task
Despite not showing any trial type-related differences in
theta power or ITPC, our data revealed significant, trial type-
related differences in the level of theta band synchrony
818
Journal of Cognitive Neuroscience
Volume 28, Numero 6
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
2
8
6
8
1
1
2
0
1
7
7
1
6
/
j
o
C
N
_
UN
_
0
0
9
3
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
tion testing) and further showed that the slow trial coher-
ence remained elevated longer than the medium trial
coherence. Although this is consistent with the longer
RTs of the slow trials, it is unlikely that these differences
were due to a time on task effect in which theta coher-
ence continues to rise throughout the trial (Cohen &
Nigbur, 2013; Scherbaum & Dshemuchadse, 2013; Zavala
et al., 2013; Nachev, 2011; Yeung, Cohen, & Botvinick,
2011); in the response aligned version of these plots
(Figure 4B) coherence between the STN and the frontal
cortex increased to similar levels in both the medium
and slow trials, despite an RT difference (and thus a differ-
ence in time spent on task) of approximately 1 sec. IL
response-aligned data revealed that roughly 1.5 sec before
the response, the theta coherence during the slow and
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
2
8
6
8
1
1
2
0
1
7
7
1
6
/
j
o
C
N
_
UN
_
0
0
9
3
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 2. Group average percentage power changes in STN LFP and
cortical EEG. (UN) STN LFP power changes aligned to the congruent
dot motion onset (left) and to the response (right). Black outline
denotes time–frequency clusters determined to be significantly different
from baseline ( p = .008, permutation testing). The baseline was chosen
to be a 1-sec long time period between trials during which all dots
moved randomly in any direction. (B) Same as A but for the FCz-Cz
frontal cortical electrode ( p = .011, for the cue aligned data and
p = .001 for the response aligned data, permutation testing).
(C) Same as A but for the Pz-Cz parietal cortical electrode ( p = .017,
for the cue aligned data and p = .001 for the response aligned data,
permutation testing). There were no significant differences in induced
power between the different trial types.
between the medial frontal EEG and the STN. Spectro-
grams of the wavelet coherence aligned to the onset
of progressive dot motion congruency for all three con-
ditions are shown in Figure 4A (left). These plots re-
vealed that during the slow and medium trials, there was
an increase in (FCz-Cz)–STN theta coherence that was
significantly greater than that observed during the fast
trials ( p = .023 and p = .031 for the slow vs. fast and
medium vs. fast comparisons, rispettivamente, permutation
testing; Figure 4A, right). Comparing the theta coherence
time series (Figure 4A, bottom right) also revealed these
differences ( p = .003 and p = .004 for the slow vs. fast
and medium vs. fast comparisons, rispettivamente, permuta-
Figura 3. Group average percentage chance in STN LFP and cortical
EEG ITPC. (UN) STN LFP ITPC changes averaged across all three trials are
shown aligned to the congruent dot motion onset (left) and to the
risposta (right). Black outline denotes time–frequency clusters
determined to be significantly different from baseline ( p = .001,
permutation testing). The baseline was chosen to be a 1-sec long time
period between trials during which all dots moved randomly in any
direction. (B) Same as A but for the FCz-Cz frontal cortical electrode
( p = .006, for the response aligned data, permutation testing).
(C) Same as A but for the Pz-Cz parietal cortical electrode. There were
no significant differences in ITPC between the different trial types.
Zavala et al.
819
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
2
8
6
8
1
1
2
0
1
7
7
1
6
/
j
o
C
N
_
UN
_
0
0
9
3
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 4. Group average normalized changes in (frontal EEG)–STN LFP wavelet coherence. (UN, left) Coherence spectrogram for the slow (top),
medium (middle), and fast (bottom) trials. Changes were found relative to a 1-sec long time period between trials during which all dots moved
randomly in any direction. (UN, right). Differences between slow and fast trials were significant (top, p = .023, permutation testing, unmasked area)
as were those between the medium and the fast trials (middle, p = .031). Time series of percent change in theta band (4–8 Hz) coherence are
also shown (bottom, mean ± SEM ), with thick lines denoting significant time points between two trial types (color of line indicates which two trial
types were compared, p = .003 for slow vs. fast comparison, p = .004 for medium vs. fast comparison, permutation testing). (B) Same as A but
for response aligned data. (B, bottom right) Both slow and medium trials showed significantly higher theta coherence in the second preceding the
risposta ( p = .039 for slow vs. fast comparison, p = .009 for medium vs. fast comparison, permutation testing).
medium trials became significantly greater than that ob-
served during the fast trials. During the time point cen-
tered 1 sec prior to the response, both medium and
slow trials demonstrated coherence values that were sig-
nificantly greater than baseline ( p = .010 and p = .013,
rispettivamente, one sample t test), but the fast trials did
non ( p > .95, one-sample t test). A one-way repeated-
measures ANOVA between trial types during this time
period revealed a significant effect of Trial type (F =
6.16, df = 2, p = .006). When the individual time points
were considered, coherence values in slow trials were
found to be significantly greater than in fast trials from
−1.60 to −0.77 sec, and those in medium trials were found
to be significantly greater from −1.34 to −0.30 sec ( p =
.039 and p = .009, rispettivamente, permutation testing).
Notably, these differences disappeared by the time the re-
sponse was executed, further arguing against a time on task
interpretation in which the theta coherence would be
expected to continue rising until the end of the response.
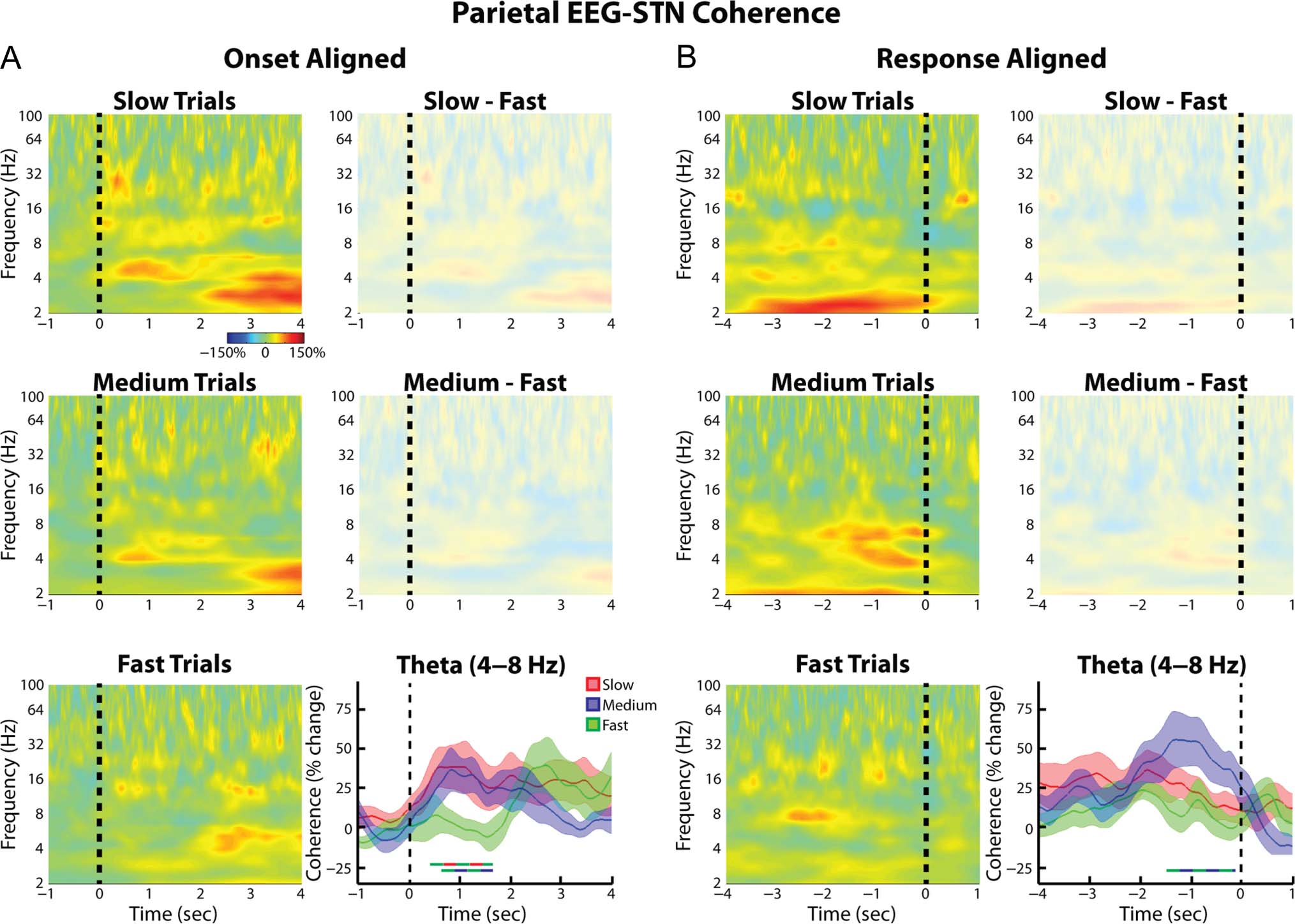
When the (Pz-Cz)–STN coherence was analyzed, simi-
lar trends as those seen for the FCz-Cz–STN connectivity
were observed (Figura 5). Although no differences in the
(Pz-Cz)–STN coherence multiple comparisons survived
correction at the spectrogram level, restricting the analy-
sis to the theta band did reveal some time points that
showed significantly higher coherence values during
the slow and medium trials relative to the fast trials
(Figure 5A). When the time period centered 1 sec prior
to the response was considered, a significant effect of
Trial type was observed (ANOVA, within-subject repeated-
measures, F = 3.74, df = 2, p = .04). In line with this, Quando
we directly tested for an effect of Cortical electrode and
Trial type, we did not observe a significant interaction be-
tween the two factors (ANOVA, within-subject repeated
820
Journal of Cognitive Neuroscience
Volume 28, Numero 6
measures, Trial type × EEG electrode: Trial type, F = 5.91,
df = 2, p = .008; EEG electrode, F = 0, df = 2, p = .96;
interaction F = 2.29, df = 2, p = .12).
To test whether the trial-dependent differences in cor-
tical STN theta connectivity were symmetrical or biased
in one direction, we repeated the synchrony analysis
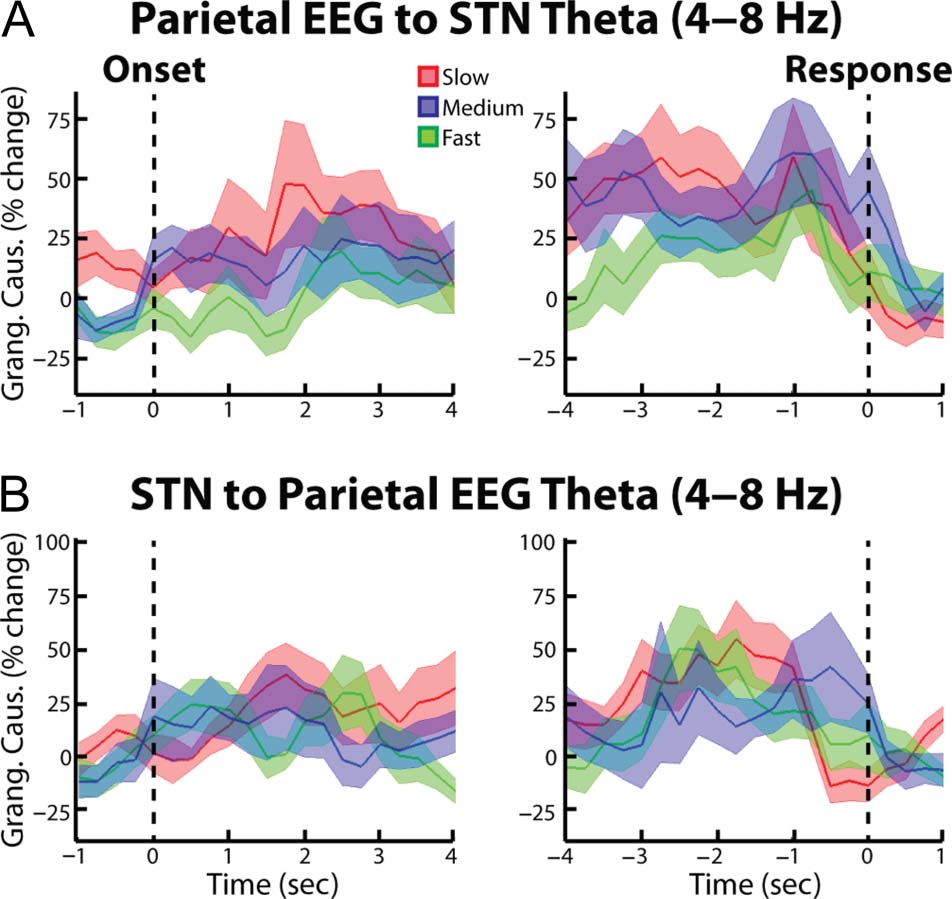
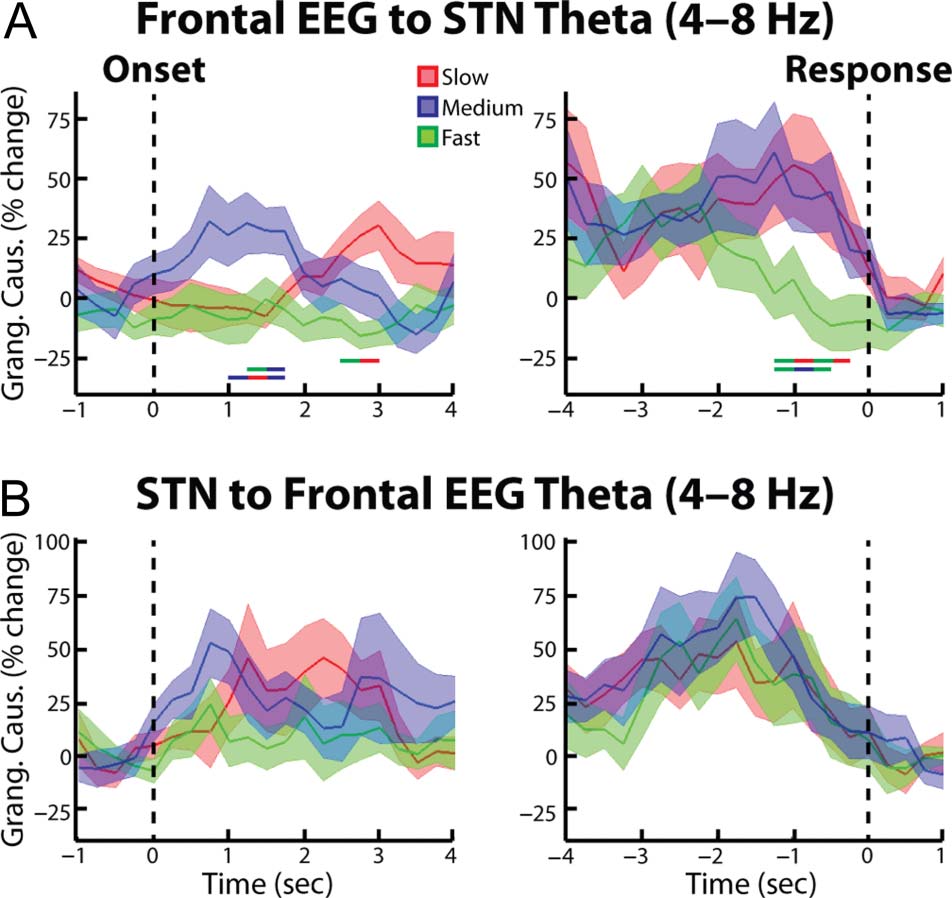
using Granger causality analytical techniques. When we
analyzed the spectral Granger causality in the frontal
cortex–STN direction, we also observed significantly high
synchrony during the slow and medium trials relative to
the fast trials ( p = .021 and p = .047 for the cue aligned
slow vs. fast and medium vs. fast comparisons, respec-
tively; Figure 6A). In contrasto, the STN–cortex spectral
Granger causality did not reveal any significant differ-
ences between trial types (Figure 6B). Together these
results suggest that the coherence differences reported in
the preceding paragraph likely reflect cortex–STN inputs.
When we repeated the Granger causality analysis for the
posterior cortex recordings (Pz-Cz), we did not reveal any
significant differences in cortex–STN and STN–cortex
spectral Granger causality (Figura 7). In accordance with
the wavelet coherence results, Tuttavia, when we directly
tested for an effect of cortical electrode and trial type, we
did not observe a significant interaction between the two
factors (ANOVA, within-subject repeated measures, Trial
type × EEG electrode: Trial type F = 2.78, df = 2, p =
.08; EEG electrode F = 1.34, df = 2, p = .27; interaction
F = 1.54, df = 2, p = .23).
DISCUSSION
Previous studies have demonstrated that tasks involving
high levels of conflict are associated with increased theta
band activity in the mPFC and in the STN (Zavala, Damera,
et al., 2015; Cohen & van Gaal, 2014; Zavala et al., 2013,
2014; Brittain et al., 2012; Cavanagh et al., 2011, 2012;
Cohen & Cavanagh, 2011). Our previous work using a
related paradigm showed that, when a similar number
of dots were moving in conflicting directions, theta band
activity of the STN became synchronized to that of the
mPFC (Zavala et al., 2014). But are such dynamic changes
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
2
8
6
8
1
1
2
0
1
7
7
1
6
/
j
o
C
N
_
UN
_
0
0
9
3
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 5. Group average normalized changes in (parietal EEG)–STN LFP wavelet coherence. Same as Figure 4, but for Pz-Cz electrode. Although no
significant differences across the whole spectrogram were observed, when the analysis was restricted to the theta band, both the slow and medium
trials showed significantly higher theta band phase synchrony relative to the fast trials ( p = .012 E .033, rispettivamente, permutation testing).
Zavala et al.
821
your horses” role the STN is thought to play during time
periods of conflict (Frank et al., 2007). Once other brain
structures had integrated enough evidence to make a de-
cision, cortico-STN coherence returned to baseline levels
and the movement was processed and executed.
What is the evidence that our three experimental con-
ditions engendered different levels of cognitive control?
A priori, one might have expected that there would be
a specific threshold of dot congruency below which par-
ticipants would not be able to perceive congruent dot
motion and above which the participants would know
with a high degree of certainty the direction in which
the dots were moving. If this were the case, all three trial
types would have required identical dot congruency at
the time of the response. Tuttavia, instead, during the
slowest trials, participants were able to make decisions
with less evidence on the screen. This suggests that par-
ticipants may have integrated dot motion congruency
evidence over time until the integral exceeded the same
threshold of integrated evidence for fast, medium, E
slow trial types. With respect to task uncertainty, IL
critical feature though is that the level of dot motion
congruency up to and including the time of the response
was less for medium and slow trials than that for fast
trials. We believe that the higher level of ambiguity during
slow and medium trials (responses were made with an
average of 24% E 33% dot congruency, rispettivamente)
could potentially require a higher level of cognitive con-
trol than that required during the fast trials (44% average
dot congruency at the time of the response). We propose
that for this reason we observed elevated theta band
Figura 7. Group average percent change in theta band (4–8 Hz)
spectral Granger causality between parietal EEG and STN LFP. Same as
Figura 6, but for Pz-Cz electrode. No significant differences were
observed ( p > .05, permutation testing).
Figura 6. Group average percent change in theta band (4–8 Hz) spectral
Granger causality between frontal EEG and STN LFP. (UN) EEG–STN
spectral Granger causality (mean ± SEM ) aligned to the congruent
dot motion onset (left). Changes were found relative to a 1-sec long
time period between trials during which all dots moved randomly in any
direction. Thick lines denoting significant time points between two trial
types are shown (color of line indicates which two trial types were
compared, p = .021 for slow vs. fast comparison, p = .047 for medium
vs. fast comparison, p = .020 for slow vs. medium comparison,
permutation testing). (UN, right) Same as (UN, left) but for data aligned
to the response ( p = .014 for slow vs. fast comparison, p = .009 for
medium vs. fast comparison, permutation testing. (B) Same as A,
but for the STN–frontal EEG direction. No significant differences
were observed.
in functional connectivity related to conflict per se, O
the increased need for cognitive control in difficult tasks?
The novel dot motion discrimination task we used
here, in which the rate at which randomly moving dots
became congruent was set to three different speeds,
allowed us to investigate the mechanisms the brain uses
to make sensorimotor decisions in which evidence is
incrementally presented. Our results suggest that cortical
STN theta synchrony is not only involved in inhibiting re-
sponses when stimuli simultaneously activate two or
more conflicting motor commands but also when indi-
viduals are asked to make decisions during higher levels
of uncertainty. The time course of the theta band coher-
ence during the medium and slow trials indicated that, COME
early as 1.5 sec before the response, cortico-STN networks
became synchronized. This suggests that the participants,
either consciously or subconsciously (O’Connell et al.,
2012), became aware of the congruent dot motion this
early, but it took up to an additional 1.5 sec to gather
enough evidence to confidently make their decision. Noi
posit that, during this time period, increased synchrony
between the cortex and the STN acts to prevent the par-
ticipants from making a response, paralleling the “hold
822
Journal of Cognitive Neuroscience
Volume 28, Numero 6
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
2
8
6
8
1
1
2
0
1
7
7
1
6
/
j
o
C
N
_
UN
_
0
0
9
3
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
coherence between activities in medial EEG electrodes
and the STN during the slow and medium trials.
Intriguingly, unlike our previous results involving ex-
plicit stimulus conflict (Zavala et al., 2013, 2014), the dif-
ferences in cortico-STN theta coherence took place
without any trial type-related differences in power or
ITPC. When we analyzed the average changes in power
across all trials, the only change from baseline that we
observed was the classic, movement-related beta band
power decrease. We also observed a significant delta
band ITPC increase in the STN and in the medial frontal
EEG electrodes that was similar to that observed in our
previous work involving both gradual dot congruency
increases (Zavala et al., 2014) and discrete stimuli presen-
tations (Zavala et al., 2013). Taken together, the relative
timing of the cortico-STN theta coherence differences
between trials and the response-related changes in beta
power and delta ITPC changes across all trials suggests
that theta band activity may play a role in inhibiting move-
ment whereas beta band suppression and delta band
ITPC may be involved in facilitating movement. As we
and others have previously argued (Kelly & O’Connell,
2013, 2015; Zavala et al., 2014; O’Connell et al., 2012),
the intentional use of gradually evolving continuous stim-
uli, as opposed to discretely presented stimuli, allows us
to separate movement and stimulus-related activity from
activity related to the decision-making process.
This study has several possible limitations that should
be considered. Primo, all recordings were made in patients
with PD, which has been consistently shown to involve
abnormal STN activity (reviewed in Hammond, Bergman,
& Brown, 2007). Tuttavia, all recordings were made with
patients on their dopaminergic medication in an attempt
to approximate physiological functioning as closely as
possible. Secondo, LFP and EEG recordings can be subject
to volume conduction of electrical signals. To mitigate
this effect, only bipolar signals were analyzed. Never-
theless, volume conduction does diminish the spatial reso-
lution of our results and may have contributed to why
we did not observe significant trial type-by-electrode inter-
actions when we compared (FCz-Cz)–STN coupling to
(Pz-Cz)–STN coupling. Although cortical oscillations are
thought to be related to structures in the frontal cortex
(Womelsdorf, Johnston, et al., 2010; Womelsdorf, Vinck,
et al., 2010; Tsujimoto et al., 2006; Wang et al., 2005;
Gevins et al., 1997), our data do not allow us to rule
out the possibility that other more posterior cortical
areas may also be involved. Third, we should acknowl-
edge the effects of any eye movements potentially not
rejected through visual inspection of the raw data, par-
ticularly as the spectral changes of interest were at low
frequencies. Finalmente, it should be noted that there are
several possible alternative interpretations of the results
we have presented. For instance, it could be argued that
there is still a decision to be made between which direc-
tion to take and that there was dot movement in both
directions (albeit just stochastic in one) presenting con-
flict even in the current paradigm. At any given time,
instantaneous dot motion congruency was less in the
medium and slow trials affording the greatest conflict
with the stochastic movement. Accordingly, it might be
argued that conflict was present and greater in the medi-
um and slow trials. Tuttavia, this interpretation would
not explain why participants made choices at points with
different levels of instantaneous dot motion congruency
in the different trial types. The alternative hypothesis,
that dot motion congruency is integrated, accounts for
this feature and yet means that trials did not differ with
respect to the level of explicit conflict at the time of re-
sponse selection. Tuttavia, conflict induced need for
delay could be reconciled with responses being made
at different levels of instantaneous dot motion congru-
ency if task behavior were governed by time-on-task,
either because some sort of temporal discounting was
at play or the participants were impulsive. In questo caso,
the increasing cost of delaying a response might mean
that participants acted despite having resolved less con-
flict in medium and slow trials. But against this, the error
rate was no different between trials, again consistent
with the interpretation that participants acted on inte-
grated evidence, which was similar at the time of decision.
Taken together, our data support the idea that en-
hanced theta synchrony between the STN and midline
electrodes may increase the net antikinetic action of
the STN. Questo, Tuttavia, may be a general operational
principle when choosing what to do and precisely when
to do it is difficult, regardless of whether this is due to the
simultaneous activation of conflicting actions or due to
sensory uncertainty. Accordingly, the results presented
here provide further insights into the processes that
mitigate response inhibition (Bastin et al., 2014; Benis
et al., 2014; Alegre et al., 2013; Brittain et al., 2012; Ray
et al., 2012; Kühn et al., 2004) and more “higher-level”
decisions (Zaghloul et al., 2012; Cavanagh et al., 2011;
Fumagalli et al., 2011), as well as help us understand some
of the impulsivity related deficits associated with neuro-
pathology (Rustamov et al., 2013; Van Meel, Heslenfeld,
Oosterlaan, & Sergeant, 2007; Fitzgerald et al., 2005), dopa-
minergic medication (Coulthard et al., 2012; Rodriguez-
Oroz et al., 2011), and DBS (Antoniades et al., 2014; Hälbig
et al., 2009; Frank et al., 2007; Frank, 2006).
Ringraziamenti
B. Z. is supported by the Rhodes Trust and the National Institutes
of Health Oxford-Cambridge fellowship. M. H., T. F., and L. Z. are
funded by the Department of Health, National Institute for Health
Research, University College London Biomedical Research
Center; The Monument Trust; and Parkinson’s Appeal for Deep
Brain Stimulation. P. B. and H. T. are funded by the Medical
Research Council, and P. B. is further funded by the Department
of Health, National Institute for Health Research, Oxford Biomed-
ical Research Centre. S. l. is funded by the Wellcome Trust. Questo
work was partly carried out in the National Institute for Health
Research cognitive health Clinical Research Facility, Oxford.
Zavala et al.
823
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
2
8
6
8
1
1
2
0
1
7
7
1
6
/
j
o
C
N
_
UN
_
0
0
9
3
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Reprint requests should be sent to Professor Peter Brown,
Nuffield Department of Clinical Neurology, University of Oxford,
Level 6, West Wing, John Radcliffe Hospital, Oxford, OX3 9DU,
UK, or via e-mail: peter.brown@ndcn.ox.ac.uk.
REFERENCES
Alegre, M., Lopez-Azcarate, J., Obeso, I., Wilkinson, L., Rodriguez-
Oroz, M. C., Valencia, M., et al. (2013). The subthalamic
nucleus is involved in successful inhibition in the stop-signal
task: A local field potential study in Parkinson’s disease.
Experimental Neurology, 239, 1–12.
Antoniades, C. A., Bogacz, R., Kennard, C., FitzGerald, J. J., Aziz,
T., & Verde, UN. l. (2014). Deep brain stimulation abolishes
slowing of reactions to unlikely stimuli. Journal of Neuroscience,
34, 10844–10852.
Bastin, J., Polosan, M., Benis, D., Goetz, L., Bhattacharjee, M.,
Piallat, B., et al. (2014). Inhibitory control and error monitoring
by human subthalamic neurons. Translational Psychiatry,
4, e439.
Benis, D., David, O., Lachaux, J.-P., Seigneuret, E., Krack, P.,
Fraix, V., et al. (2014). Subthalamic nucleus activity dissociates
proactive and reactive inhibition in patients with Parkinson’s
disease. Neuroimage, 91, 273–281.
Bogacz, R., & Gurney, K. (2007). The basal ganglia and cortex
implement optimal decision making between alternative
actions. Calcolo neurale, 19, 442–477.
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004). Conflict
monitoring and anterior cingulate cortex: An update. Trends
in Cognitive Sciences, 8, 539–546.
Brittain, J.-S., Watkins, K. E., Joundi, R. A., Ray, N. J., Holland,
P., Verde, UN. L., et al. (2012). A role for the subthalamic
nucleus in response inhibition during conflict. Journal of
Neuroscience, 32, 13396–13401.
Brovelli, A., Ding, M., Ledberg, A., Chen, Y., Nakamura, R., &
Bressler, S. l. (2004). Beta oscillations in a large-scale
sensorimotor cortical network: Directional influences revealed
by Granger causality. Proceedings of the National Academy
of Sciences, U.S.A., 101, 9849–9854.
Cavanagh, J. F., & Shackman, UN. J. (2015). Frontal midline theta
reflects anxiety and cognitive control: Meta-analytic evidence.
Journal of Physiology (Paris), 109, 3–15.
Cavanagh, J. F., Wiecki, T. V., Cohen, M. X., Figueroa, C. M.,
Samanta, J., Sherman, S. J., et al. (2011). Subthalamic nucleus
stimulation reverses mediofrontal influence over decision
threshold. Nature Neuroscience, 14, 1462–1467.
Cavanagh, J. F., Zambrano-Vazquez, L., & Allen, J. J. B. (2012).
Theta lingua franca: A common midfrontal substrate for
action monitoring processes. Psychophysiology, 49, 220–238.
Cohen, M. X. (2014). Analyzing neural time series data:
Theory and practice. Cambridge, MA: CON Premere.
Cohen, M. X., & Cavanagh, J. F. (2011). Single-trial regression
elucidates the role of prefrontal theta oscillations in response
conflict. Frontiers in Perception Science, 2, 30.
Cohen, M. X., & Gulbinaite, R. (2014). Five methodological
challenges in cognitive electrophysiology. Neuroimage,
85, 702–710.
Cohen, M. X., & Nigbur, R. (2013). Reply to “Higher response
time increases theta energy, conflict increases response
time.” Clinical Neurophysiology, 124, 1479–1481.
Cohen, M. X., & van Gaal, S. (2014). Subthreshold muscle
twitches dissociate oscillatory neural signatures of conflicts
from errors. Neuroimage, 86, 503–513.
Coulthard, E. J., Bogacz, R., Javed, S., Mooney, l. K., Murphy,
G., Keeley, S., et al. (2012). Distinct roles of dopamine and
subthalamic nucleus in learning and probabilistic decision
making. Brain, 135, 3721–3734.
Ding, M., Bressler, S. L., Yang, W., & Liang, H. (2000).
Short-window spectral analysis of cortical event-related
potentials by adaptive multivariate autoregressive
modeling: Data preprocessing, model validation, E
variability assessment. Biological Cybernetics, 83, 35–45.
Ding, M., Chen, Y., & Bressler, S. l. (2006). Granger causality:
Basis theory and application to neuroscience. In B. Schelter,
M. Winterhalder, & J. Timmer (Eds.), Handbook of time
series analysis: Recent theoretical developments and
applications (pag. 437–460). Berlin: Wiley-VCH. Available
at: www.ccs.fau.edu/∼bressler/pdf/HTSA06.pdf.
Fitzgerald, K. D., Welsh, R. C., Gehring, W. J., Abelson, J. L., Himle,
J. A., Liberzon, I., et al. (2005). Error-related hyperactivity
of the anterior cingulate cortex in obsessive-compulsive
disorder. Biological Psychiatry, 57, 287–294.
Foltynie, T., & Hariz, M. IO. (2010). Surgical management of
Parkinson’s disease. Expert Review of Neurotherapeutics,
10, 903–914.
Frank, M. J. (2006). Hold your horses: A dynamic computational
role for the subthalamic nucleus in decision making.
Neural Networks, 19, 1120–1136.
Frank, M. J., Samanta, J., Moustafa, UN. A., & Sherman, S. J.
(2007). Hold your horses: Impulsivity, deep brain stimulation,
and medication in parkinsonism. Scienza, 318, 1309–1312.
Fumagalli, M., Giannicola, G., Rosa, M., Marceglia, S., Lucchiari,
C., Mrakic-Sposta, S., et al. (2011). Conflict-dependent
dynamic of subthalamic nucleus oscillations during moral
decisions. Social Neuroscience, 6, 243–256.
Gardner, W. UN. (1992). A unifying view of coherence in signal
processing. Signal Processing, 29, 113–140.
Gevins, A., Smith, M. E., McEvoy, L., & Yu, D. (1997).
High-resolution EEG mapping of cortical activation related
to working memory: Effects of task difficulty, type of
processing, and practice. Cerebral Cortex, 7, 374–385.
Geweke, J. (1982). Measurement of linear dependence and
feedback between multiple time series. Journal of the
American Statistical Association, 77, 304–313.
Granger, C. (1969). Investigating causal relations by econometric
models and cross-spectral methods. Econometrica, 37,
424–438.
Verde, N., Bogacz, R., Huebl, J., Beyer, A.-K., Kühn, UN. A., &
Heekeren, H. R. (2013). Reduction of influence of task
difficulty on perceptual decision making by STN deep brain
stimulation. Current Biology, 23, 1681–1684.
Hälbig, T. D., Tse, W., Frisina, P. G., Hollander, E., Shapiro, H.,
Tagliati, M., et al. (2009). Subthalamic deep brain stimulation
and impulse control in Parkinson’s disease. European
Journal of Neurology, 16, 493–497.
Hammond, C., Bergman, H., & Brown, P. (2007). Pathological
synchronization in Parkinson’s disease: Networks, models
and treatments. Trends in Neurosciences, 30, 357–364.
Kamiński, M. J., & Blinowska, K. J. (1991). A new method of
the description of the information flow in the brain
structures. Biological Cybernetics, 65, 203–210.
Kelly, S. P., & O’Connell, R. G. (2013). Internal and external
influences on the rate of sensory evidence accumulation
in the human brain. Journal of Neuroscience, 33,
19434–19441.
Kelly, S. P., & O’Connell, R. G. (2015). The neural processes
underlying perceptual decision making in humans: Recente
progress and future directions. Journal of Physiology (Paris),
109, 27–37.
Krain, UN. L., Wilson, UN. M., Arbuckle, R., Castellanos, F. X., &
Milham, M. P. (2006). Distinct neural mechanisms of risk and
ambiguity: A meta-analysis of decision-making. Neuroimage,
32, 477–484.
Kühn, UN. A., Williams, D., Kupsch, A., Limousin, P., Hariz, M.,
Schneider, G.-H., et al. (2004). Event-related beta
824
Journal of Cognitive Neuroscience
Volume 28, Numero 6
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
2
8
6
8
1
1
2
0
1
7
7
1
6
/
j
o
C
N
_
UN
_
0
0
9
3
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
desynchronization in human subthalamic nucleus correlates
with motor performance. Brain, 127, 735–746.
Lachaux, J.-P., Lutz, A., Rudrauf, D., Cosmelli, D., Le Van Quyen,
M., Martinerie, J., et al. (2002). Estimating the time-course
of coherence between single-trial brain signals: An
introduction to wavelet coherence. Clinical Neurophysiology,
32, 157–174.
Maris, E., & Oostenveld, R. (2007). Nonparametric statistical
testing of EEG- and MEG-data. Journal of Neuroscience
Methods, 164, 177–190.
Mushtaq, F., Bland, UN. R., & Schaefer, UN. (2011). Uncertainty
and cognitive control. Frontiers in Psychology, 2, 249.
Nachev, P. (2011). The blind executive. Neuroimage,
57, 312–313.
O’Connell, R. G., Dockree, P. M., & Kelly, S. P. (2012). UN
supramodal accumulation-to-bound signal that determines
perceptual decisions in humans. Nature Neuroscience,
15, 1729–1735.
Peirce, J. W. (2007). PsychoPy—Psychophysics software in
Python. Journal of Neuroscience Methods, 162, 8–13.
Pesaran, B. (2008). Short course III, presented at 2008 Società
for Neuroscience Annual Meeting. In P. Mitra (Ed.), Neural
signal processing: Quantitative analysis of neural activity.
Washington, DC: Society for Neuroscience.
Ratcliff, R., & Frank, M. J. (2012). Reinforcement-based decision
making in corticostriatal circuits: Mutual constraints by
neurocomputational and diffusion models. Neural
Computation, 24, 1186–1229.
Ratcliff, R., & McKoon, G. (2008). The diffusion decision model:
Theory and data for two-choice decision tasks. Neural
Computation, 20, 873–922.
Ratcliff, R., & Tuerlinckx, F. (2002). Estimating parameters of
the diffusion model: Approaches to dealing with contaminant
reaction times and parameter variability. Psychonomic
Bulletin & Review, 9, 438–481.
Ray, N. J., Brittain, J.-S., Holland, P., Joundi, R. A., Stein, J. F.,
Aziz, T. Z., et al. (2012). The role of the subthalamic nucleus
in response inhibition: Evidence from local field potential
recordings in the human subthalamic nucleus. Neuroimage,
60, 271–278.
Rodriguez-Oroz, M. C., López-Azcárate, J., Garcia-Garcia, D.,
Alegre, M., Toledo, J., Valencia, M., et al. (2011).
Involvement of the subthalamic nucleus in impulse
control disorders associated with Parkinson’s disease.
Brain, 134, 36–49.
Ruiz, M. H., Huebl, J., Schönecker, T., Kupsch, A., Yarrow, K.,
Krauss, J. K., et al. (2014). Involvement of human internal
globus pallidus in the early modulation of cortical
error-related activity. Cerebral Cortex, 24, 1502–1517.
Rustamov, N., Rodriguez-Raecke, R., Timm, L., Agrawal, D.,
Dressler, D., Schrader, C., et al. (2013). Absence of
congruency sequence effects reveals neurocognitive
inflexibility in Parkinson’s disease. Neuropsychologia,
51, 2976–2987.
Scherbaum, S., & Dshemuchadse, M. (2013). Higher response
time increases theta energy, conflict increases response
time. Clinical Neurophysiology, 124, 1477–1479.
Schlögl, A., & Supp, G. (2006). Analyzing event-related EEG data
with multivariate autoregressive parameters. Progress in
Brain Research, 159, 135–147.
Seth, UN. K. (2010). A MATLAB toolbox for Granger causal
connectivity analysis. Journal of Neuroscience Methods,
186, 262–273.
Stern, E. R., Gonzalez, R., Welsh, R. C., & Taylor, S. F. (2010).
Updating beliefs for a decision: Neural correlates of uncertainty
and underconfidence. Journal of Neuroscience,
30, 8032–8041.
Tsujimoto, T., Shimazu, H., & Isomura, Y. (2006). Direct recording
of theta oscillations in primate prefrontal and anterior
cingulate cortices. Journal of Neurophysiology, 95, 2987–3000.
Van Meel, C. S., Heslenfeld, D. J., Oosterlaan, J., & Sergeant,
J. UN. (2007). Adaptive control deficits in attention-deficit/
hyperactivity disorder (ADHD): The role of error processing.
Psychiatry Research, 151, 211–220.
Wang, C., Ulbert, I., Schomer, D. L., Marinkovic, K., & Halgren,
E. (2005). Responses of human anterior cingulate cortex
microdomains to error detection, conflict monitoring,
stimulus-response mapping, familiarity, and orienting.
Journal of Neuroscience, 25, 604–613.
White, T. P., Engen, N. H., Sørensen, S., Overgaard, M., &
Shergill, S. S. (2014). Uncertainty and confidence from the
triple-network perspective: Voxel-based meta-analyses. Brain
and Cognition, 85, 191–200.
Womelsdorf, T., Johnston, K., Vinck, M., & Everling, S. (2010).
Theta-activity in anterior cingulate cortex predicts task rules
and their adjustments following errors. Atti del
National Academy of Sciences, U.S.A., 107, 5248–5253.
Womelsdorf, T., Vinck, M., Leung, l. S., & Everling, S. (2010).
Selective theta-synchronization of choice-relevant information
subserves goal-directed behavior. Frontiers in Human
Neuroscience, 4, 210.
Yeung, N., Cohen, J. D., & Botvinick, M. M. (2011). Errors of
interpretation and modeling: A reply to Grinband et al.
Neuroimage, 57, 316–319.
Zaghloul, K. A., Weidemann, C. T., Lega, B. C., Jaggi, J. L.,
Baltuch, G. H., & Kahana, M. J. (2012). Neuronal activity in the
human subthalamic nucleus encodes decision conflict during
action selection. Journal of Neuroscience, 32, 2453–2460.
Zavala, B., Brittain, J.-S., Jenkinson, N., Ashkan, K., Foltynie, T.,
Limousin, P., et al. (2013). Subthalamic nucleus local field
potential activity during the Eriksen flanker task reveals a
novel role for theta phase during conflict monitoring.
Journal of Neuroscience, 33, 14758–14766.
Zavala, B., Damera, S., Dong, J. W., Lungu, C., Brown, P., &
Zaghloul, K. UN. (2015). Human subthalamic nucleus theta and
beta oscillations entrain neuronal firing during sensorimotor
conflict. Cerebral Cortex, bhv244.
Zavala, B., Zaghloul, K., & Brown, P. (2015). The subthalamic
nucleus, oscillations, and conflict. Movement Disorders,
30, 328–338.
Zavala, B. A., Tan, H., Little, S., Ashkan, K., Hariz, M., Foltynie,
T., et al. (2014). Midline frontal cortex low-frequency activity
drives subthalamic nucleus oscillations during conflict.
Journal of Neuroscience, 34, 7322–7333.
Zavala et al.
825
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
2
8
6
8
1
1
2
0
1
7
7
1
6
/
j
o
C
N
_
UN
_
0
0
9
3
4
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3