Cortical Tracking of Speech: Toward Collaboration
between the Fields of Signal and
Sentence Processing
Eleonora J. Beier1, Suphasiree Chantavarin1,2, Gwendolyn Rehrig1,
Fernanda Ferreira1, and Lee M. Miller1
Astratto
■ In recent years, a growing number of studies have used corti-
cal tracking methods to investigate auditory language processing.
Although most studies that employ cortical tracking stem from
the field of auditory signal processing, this approach should also
be of interest to psycholinguistics—particularly the subfield of
sentence processing—given its potential to provide insight into
dynamic language comprehension processes. Tuttavia, there
has been limited collaboration between these fields, which we
suggest is partly because of differences in theoretical background
and methodological constraints, some mutually exclusive. In this
paper, we first review the theories and methodological con-
straints that have historically been prioritized in each field and
provide concrete examples of how some of these constraints
may be reconciled. We then elaborate on how further collaboration
between the two fields could be mutually beneficial. Specifically,
we argue that the use of cortical tracking methods may help re-
solve long-standing debates in the field of sentence processing that
commonly used behavioral and neural measures (per esempio., ERPs) Avere
failed to adjudicate. Allo stesso modo, signal processing researchers who
use cortical tracking may be able to reduce noise in the neural data
and broaden the impact of their results by controlling for linguis-
tic features of their stimuli and by using simple comprehension
compiti. Overall, we argue that a balance between the methodolog-
ical constraints of the two fields will lead to an overall improved
understanding of language processing as well as greater clarity on
what mechanisms cortical tracking of speech reflects. Increased
collaboration will help resolve debates in both fields and will lead
to new and exciting avenues for research. ■
INTRODUCTION
Recent years have seen a growing interest in the cortical
tracking of speech as a potential measure of acoustic, lin-
guistic, and cognitive processing (Meyer, Sun, & Martin,
2020; Obleser & Kayser, 2019; Kösem & van Wassenhove,
2017; Meyer, 2018; see Tyler, 2020). The terms “cortical
tracking” or “speech tracking” loosely refer to continuous
neural activity that is somehow time-locked to ongoing
events in the speech signal. According to one common
interpretation, cortical tracking reflects the tendency for
neural oscillations to align, or phase-lock, with quasiperi-
odic features in the speech signal. These quasiperiodic
elements of speech can be acoustic, such as the fluctua-
tions in the amplitude envelope associated with syllables
(Doelling, Arnal, Ghitza, & Poeppel, 2014; Peelle, Gross,
& Davis, 2013) or linguistic representations generated in
the mind of the listener, such as syntactic phrase bound-
aries (Meyer, Henry, Gaston, Schmuck, & Friederici,
2017; Ding, Melloni, Zhang, Tian, & Poeppel, 2016).
Researchers adopting this approach often refer to cortical
tracking as “neural entrainment” (Obleser & Kayser,
1University of California, Davis, 2Chulalongkorn University,
Bangkok, Thailand
2019; see later sections for further discussion of terminol-
ogy and debates in this field). It has been proposed that
entrainment may contribute to improved speech pro-
cessing and language comprehension—for instance, by
instantiating temporal predictions that enable segmenta-
tion of the continuous speech signal into units at several
timescales (Keitel, Gross, & Kayser, 2018; Kösem et al.,
2018; Meyer & Gumbert, 2018; Meyer et al., 2017; Ding
et al., 2016; Zoefel & VanRullen, 2015; Doelling et al.,
2014; Peelle et al., 2013; Giraud & Poeppel, 2012;
Peelle & Davis, 2012; Ahissar et al., 2001).
Cortical tracking methods should therefore be of great
interest to researchers studying sentence processing
from a psycholinguistic perspective. Sentence processing
research makes frequent use of ERPs to draw inferences
about neural responses to isolated, discrete events such
as word onsets or sentence boundaries (Swaab, Ledoux,
Camblin, & Boudewyn, 2012; Kutas & Federmeier, 2011;
Kutas, Van Petten, & Kluender, 2006) as well as time–
frequency analyses of EEG oscillatory power at specific
bands (Prystauka & Lewis, 2019). Tuttavia, the field of
sentence processing has yet to fully incorporate cortical
tracking as a tool to investigate language processing
mechanisms continuously, rather than at discrete epochs.
© 2022 the Massachusetts Institute of Technology. Published under
a Creative Commons Attribution 4.0 Internazionale (CC BY 4.0) licenza.
Journal of Cognitive Neuroscience 33:4, pag. 574–593
https://doi.org/10.1162/jocn_a_01676
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
As we will argue, combining measures of cortical tracking
with typical psycholinguistic paradigms may help resolve
long-standing debates and distinguish between compet-
ing theories of language processing, while also making
use of continuous EEG data that are typically treated as
noise in ERP paradigms (a point we discuss further in
the section titled Contributions to Psycholinguistics).
Generalmente, there has been limited collaboration be-
tween signal processing neuroscientists who use cortical
tracking paradigms and psycholinguists, despite the fact
that both fields share the common goal of elucidating
how listeners process spoken language. To be more spe-
cific, the research areas that are most relevant for our
purposes are the study of human sentence processing,
which is a subfield of psycholinguistics, and the study
of auditory signal processing, which is often carried out
by perceptual neurophysiologists and engineers who are
interested in how the brain transforms auditory signals
and who may implement cortical tracking in their re-
search methods. These research topics intersect when
the auditory signal comprises spoken sentences. For con-
venience, we will refer to these research areas as “sen-
tence processing” and “signal processing” throughout,
with the caveat that the fields are not mutually exclusive;
there are psycholinguists who employ cortical tracking
methods to study sentence processing (per esempio., Song &
Iverson, 2018; Martin & Doumas, 2017; Meyer et al.,
2017), signal processing researchers who are interested
in the linguistic properties of the speech signal (per esempio.,
Ding et al., 2016), and investigators with clear interest
in both fields who already conduct studies that incorpo-
rate the methodological compromises we suggest later
SU (per esempio., Giraud & Poeppel, 2012; Peelle & Davis, 2012;
Obleser & Kotz, 2011).
Despite these notable examples of interaction, the two
fields remain largely independent. Part of the reason for
this limited collaboration stems from the different ways
these two fields conceptualize and define language pro-
cessazione; whereas sentence processing research focuses
on the cognitive and linguistic representations formed
during comprehension, signal processing research treats
speech as an example of a complex auditory signal and
seeks to characterize the system that transforms that in-
put signal into an output signal or response (per esempio.,
Rimmele, Morillon, Poeppel, & Arnal, 2018; Morillon &
Schroeder, 2015). An additional hurdle to collaboration
stems from the largely different methodological con-
straints that the two fields are primarily concerned with.
Some of these constraints are because of the theoretical
grounds upon which research is conducted as well as lim-
itations in the current methods for data acquisition; these
are sometimes at odds across fields and can be difficult to
reconcile. Tuttavia, we argue that most constraints are
reconcilable and that both fields would benefit from in-
corporating aspects of each other’s methodology and
theoretical constructs. Just as sentence processing re-
search would be able to answer more detailed questions
about linguistic processing by using more of the contin-
uous EEG data through cortical tracking methods, signal
processing research would gain a better understanding of
the role of cortical tracking in the processing of speech
by controlling and manipulating the linguistic features of
the stimulus, which are often underspecified in current
studies.
in questo documento, we first review the theoretical back-
ground and the methodological constraints that have his-
torically been prioritized in the study of sentence
processing and the neuroscience of auditory signal pro-
cessazione. We then explore in more detail what further col-
laboration between these two fields could bring,
highlighting how cortical tracking methods could be
used to improve our understanding of continuous sen-
tence comprehension and how paradigms from psycho-
linguistics may in turn improve our understanding of the
transformations listeners apply to speech as an input sig-
nal. Finalmente, we will provide concrete ideas for how to rec-
oncile each field’s constraints to reach these goals. Noi
conclude by arguing that, although no perfect experi-
ment can be constructed that will satisfy all constraints,
a better mutual understanding of each field’s approach
will greatly improve experimental designs in both fields
and open up new exciting avenues for research.
OVERVIEW OF RESEARCH ON SIGNAL AND
SENTENCE PROCESSING
Cortical Tracking in Signal Processing Research
Cortical tracking refers to the observed alignment of
rhythmic neural activity with an external periodic or qua-
siperiodic stimulus. It has been observed in response to
both visual and auditory stimuli (Besle et al., 2011;
Gomez-Ramirez et al., 2011; Luo, Liu, & Poeppel, 2010;
Lakatos, Karmos, Mehta, Ulbert, & Schroeder, 2008;
Lakatos et al., 2005). Although this phenomenon has re-
ceived growing interest in the past decade, there is still
considerable debate as to the neural causes of cortical
tracking and the role it may play in various aspects of cog-
nition (per esempio., Attenzione, temporal prediction, speech pro-
cessazione). Specifically, debate has centered around
whether cortical tracking results from the phase-locking
of ongoing, endogenous oscillations (Calderone, Lakatos,
Butler, & Castellanos, 2014; Doelling et al., 2014; Giraud
& Poeppel, 2012; Peelle & Davis, 2012) or whether it is
the epiphenomenal result of repeated evoked responses
(per esempio., reflexive responses to the stimulus). There is also
debate about whether cortical tracking serves a functional
role in attention and speech processing, such as by acting
as a mechanism for temporal prediction and segmenta-
zione (Kösem et al., 2018; Morillon & Schroeder, 2015;
Calderone et al., 2014; Doelling et al., 2014; Giraud &
Poeppel, 2012; Peelle & Davis, 2012; Lakatos et al.,
2008), or whether it is merely a passive response to other
mechanisms (Rimmele et al., 2018; Ding & Simone, 2014).
Beier et al.
575
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
These different viewpoints are sometimes reflected in
the terminology used to describe cortical tracking:
Studies that argue or assume that cortical tracking in-
volves the synchronization of ongoing oscillations often
use the term “neural entrainment,” as opposed to the
more neutral terms cortical/neural tracking, phase cod-
ing, speech tracking, and so on (Obleser & Kayser,
2019). in questo documento, we are agnostic about the neural or-
igins and cognitive role of this phenomenon, and we
therefore use the neutral term “cortical tracking”
throughout.
Cortical Tracking of Speech
In this section, we briefly review the empirical and theo-
retical background on cortical tracking of speech.
Interested readers are encouraged to explore reviews
by Meyer (2018), Kösem and van Wassenhove (2017),
and Obleser and Kayser (2019) for comprehensive
explanations.
Much of the empirical work on cortical tracking of
speech has been based on the view that it arises from
the phase-locking of ongoing oscillations to events in
the speech signal and that it represents an attentional
or attention-like mechanism whereby periods of high
neuronal excitability align with the temporal occurrence
of the stimulus events to maximize processing efficiency
(Giraud & Poeppel, 2012; Peelle & Davis, 2012; Lakatos
et al., 2008). The idea of cortical tracking as an attentional
mechanism has been theorized to support beat percep-
tion in music through the Dynamic Attending Theory
(Large & Jones, 1999; Large & Kolen, 1994). According
to this account, the perception of rhythm relies on the
dynamic allocation of attention to points in time when
the next beat is predicted to occur. The synchronization
of neural oscillations to the beat is therefore a mecha-
nism that allows for temporal predictions. Inoltre,
cortical tracking has been observed for imagined group-
ings of acoustically identical periodic beats (Nozaradan,
Peretz, Missal, & Mouraux, 2011), providing further evi-
dence for its role in the perception of hierarchically orga-
nized rhythmic patterns, or meter. It also indicates that
cortical tracking may reflect the segmentation of stimuli
into larger units not necessarily represented acoustically
(but see Meyer et al., 2020, and Tyler, 2020, for important
considerations on the difference between cortical track-
ing of acoustic features as opposed to endogenously gen-
erated representations).
It has been argued that this type of attentional mech-
anism, allowing for temporal predictions through the syn-
chronization of ongoing neural oscillations, is used for
the perception of speech as well (Meyer, 2018; Ding
et al., 2016; Doelling et al., 2014; Giraud & Poeppel,
2012; Peelle & Davis, 2012). Speech is a temporal signal
consisting of quasiperiodic events at multiple timescales.
Cortical tracking has been observed in response to many
acoustic and linguistic properties of speech, including
syllabic rate (Doelling et al., 2014; Peelle et al., 2013;
Luo & Poeppel, 2007), the presence of prosodic intona-
tional boundaries (Bourguignon et al., 2013), and the
presence of syntactic phrases (Meyer & Gumbert, 2018;
Meyer et al., 2017; Ding et al., 2016). Given that speech
requires extremely fast processing, often in noisy envi-
ronments, it would be beneficial for the listener to be
able to preallocate attention to the points in time where
important bits of signal will likely occur. In this way, IL
listener can ensure that this information will coincide
with maximal neuronal excitability and therefore more ef-
ficient processing (Meyer & Gumbert, 2018; Morillon &
Schroeder, 2015; Peelle & Davis, 2012). Speech and mu-
sic would not be unique in this respect, as temporal pre-
dictions are thought to modulate attention and facilitate
processing of events at predicted time locations more
broadly (Nobre & van Ede, 2018), and temporal predic-
tions in the auditory domain in particular have demon-
strated the involvement of delta oscillations (Stefanics
et al., 2010) similarly to the domain of language (Ding
et al., 2016).
This ability to predict where important linguistic infor-
mation is likely to occur could also support the temporal
segmentation of speech into its linguistic units (Doelling
et al., 2014; Giraud & Poeppel, 2012), such as the forma-
tion of syntactic boundaries (Meyer et al., 2017; Ding
et al., 2016). In a foundational study, Ding et al. (2016)
showed evidence of cortical tracking not only to period-
ically presented monosyllabic words but also to the two-
word phrases and the four-word sentences that these
words combined into. Importantly, syntactic boundaries
were not marked acoustically, suggesting that the ob-
served cortical tracking reflected a mental representation
rather than an acoustic property, similar to what has been
found for an imaginary meter (Nozaradan et al., 2011). If
cortical tracking plays a functional role in actively predict-
ing temporal events and in segmenting speech into units,
then it may be an essential aspect of speech perception
and language comprehension (Schwartze & Kotz, 2013;
Peelle & Davis, 2012; Kotz & Schwartze, 2010; Ghitza &
Greenberg, 2009).
Although this view has recently gained popularity,
some have pointed out that what may appear as the en-
trainment of intrinsic oscillations to speech may in fact
result from a series of evoked responses (per esempio., Nora
et al., 2020) or the by-product of attentional gain mech-
anismo (per esempio., Kerlin, Shahin, & Mugnaio, 2010; for discus-
sions, see Ding & Simone, 2014, and Kösem & van
Wassenhove, 2017). Cortical tracking would therefore
consist of a passive response that plays no functional role
in comprehension. Despite the difficulty of ruling out this
possibility, several recent studies have provided evidence
for oscillatory models that entail a more active role of cor-
tical tracking in speech comprehension (Keitel et al.,
2018; Kösem et al., 2018; Meyer & Gumbert, 2018;
Zoefel, Archer-Boyd, & Davis, 2018; Meyer et al., 2017;
Ding et al., 2016; Ding, Chatterjee, & Simone, 2014;
576
Journal of Cognitive Neuroscience
Volume 33, Numero 4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Doelling et al., 2014; Peelle et al., 2013). The idea that
cortical tracking entirely reflects evoked responses is also
inconsistent with the finding that neural oscillations per-
sist at the stimulus frequency for several cycles even after
the stimulus ends (Calderone et al., 2014). A third possi-
bility is that cortical tracking does reflect the phase-
locking of endogenous neuronal oscillations but that this
is not itself a temporal prediction mechanism; Piuttosto,
neural oscillations may constitute a processing constraint,
and phase reset is induced by subcortical structures in-
volved in the top–down temporal prediction of both pe-
riodic and aperiodic signals (Rimmele et al., 2018).
With respect to speech in particular, debate surrounds
the question of whether cortical tracking reflects only
low-level perception of acoustic properties of speech or
whether it is actively involved in top–down speech com-
prehension, tracking language-specific features beyond
acoustic features. One common way to address this ques-
tion has been to vary the degree of speech intelligibility
through acoustic manipulations, which has led to mixed
risultati (per esempio., Baltzell, Srinivasan, & Richards, 2017; Zoefel
& VanRullen, 2016; Millman, Johnson, & Prendergast,
2015; Doelling et al., 2014; Peelle et al., 2013; Howard
& Poeppel, 2010; Ahissar et al., 2001). One possibility is
that multiple mechanisms are involved, such that cortical
tracking may play different roles and track different fea-
tures depending on the frequency band and the neuroan-
atomical source (Ding & Simone, 2014; Kösem & van
Wassenhove, 2017; Zoefel & VanRullen, 2015). For in-
stance, it has been recently proposed that cortical track-
ing consists of both “entrainment proper” (phase-locking
to acoustic periodicities of the signal) and “intrinsic syn-
chronicities” reflecting the endogenous generation of lin-
guistic structure and predictions (see Meyer et al., 2020,
for a clear distinction of these terms). Così, although cor-
tical tracking of speech has recently received much atten-
zione, its source and role are still debated, and there is still
much to learn regarding its functional role in language
comprehension. We will argue that cortical tracking is a
useful tool for exploring psycholinguistic questions about
language comprehension regardless of what neural
mechanisms it may reflect and that psycholinguistic par-
adigms may in fact help elucidate the potential role(S) Di
cortical tracking in language processing and cognition
more broadly.
Psycholinguistic Issues in Sentence
Processing Research
Prossimo, we will focus on the kinds of questions that have
been asked in sentence processing research and the gen-
eral classes of theories that have been proposed to ac-
count for psycholinguistic performance. The purpose of
this section is not to provide an exhaustive review but
rather to set the stage for the discussion to follow regard-
ing how sentence processing research conceptualizes the
important considerations that go into designing empirical
studies.
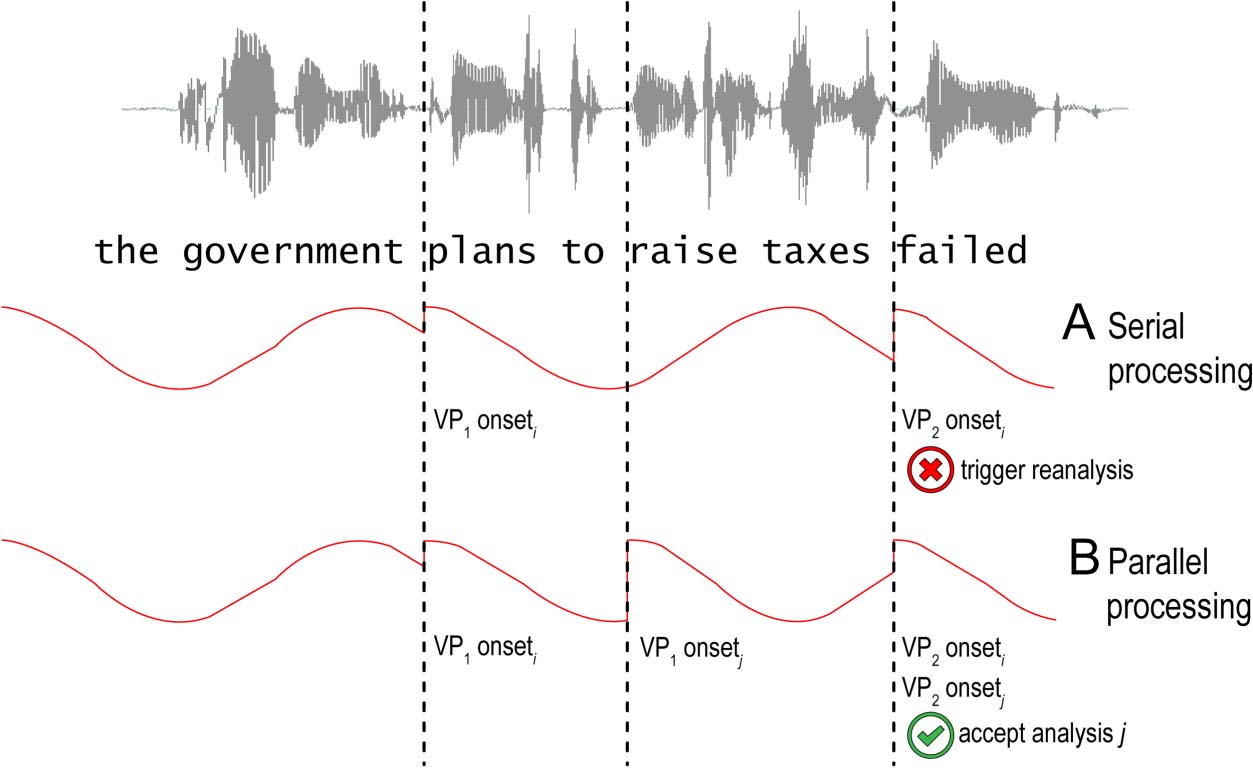
The fundamental question that sentence processing
theories try to address is how humans understand lan-
guage in real time. (Of course language production is
an important area of investigation as well but is beyond
the scope of this review.) As the written or spoken signal
unfolds, the comprehender assigns an interpretation at a
number of different levels of linguistic representation:
prosodic, syntactic, semantic, and pragmatic. A core as-
sumption is that the system is “incremental,” meaning
that interpretations are assigned as the input is received
and at all levels of representation (but see Christiansen &
Chater, 2015; Bever & Townsend, 2001). Così, upon
hearing the word “The” at the start of an utterance, IL
syllable is categorized as an instance of the word “the,” it
is assigned the syntactic category “determiner,” and a
syntactic structure is projected positing the existence of
a subject noun phrase and perhaps even an entire clause
(cioè., so-called “left-corner” parsing; Abney & Johnson,
1991; Johnson-Laird, 1983). Incremental interpretation
supports efficient processing because input is catego-
rized as it is received, which avoids the need for back-
tracking and for holding unanalyzed material in working
memory. Tuttavia, incremental interpretation will often
lead to “garden paths” at a number of levels of interpre-
tazione: Per esempio, given a sequence such as “The prin-
cipal spoke to the cafeteria…,” readers tend to spend a
long time fixating on “cafeteria” because it is implausible
as the object of “speak to,” a confusion that gets resolved
once an animate noun such as “manager” is encountered
(Staub, Rayner, Pollatsek, Hyönä, & Majewski, 2007); sim-
ilarly, there is evidence from ERPs that any word can in-
voke a late positivity similar to a P600 component with
cumulative syntactic effort, as measured by the number
of parsing steps taken to parse the sentence before en-
countering the word (Hale, Dyer, Kuncoro, & Brennan,
2018).
Debate continues regarding the most compelling
theoretical framework for explaining these and other
processing effects (for a review, see Traxler, 2014).
Tuttavia, psycholinguists do agree that comprehenders
eventually make use of all relevant information and that
processing is constrained by the architectural properties
of the overall cognitive system, including working mem-
ory constraints (per esempio., Kim, Oines, & Miyake, 2018; Huettig
& Janse, 2016; Swets, Desmet, Hambrick, & Ferreira,
2007). Recent models emphasize the need to account
for the language system’s tendency to construct shallow,
incomplete, and occasionally nonveridical representa-
tions of the input (Ferreira & Lowder, 2016; Gibson,
Bergen, & Piantadosi, 2013; Ferreira, 2003). Related ap-
proaches highlight the rational nature of comprehension,
which assume that readers and listeners optimally com-
bine the input with their rational expectations to arrive
at an optimal construction of the linguistic signal, Quale
may allow for alterations to the input in accordance with
Beier et al.
577
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
noisy channel models (Futrell, Gibson, & Levy, 2020;
Gibson et al., 2013). These models are often rooted in
computational algorithms that assign surprisal (the de-
gree to which the word is expected given the preceding
context) and entropy (the degree to which the word con-
strains upcoming linguistic content) values to each word
in a sentence, reflecting how easily a word can be inte-
grated given the left context and the overall statistics of
the language (Futrell et al., 2020). Neuroimaging evi-
dence indicates that activation in language-related brain
areas correlates with difficulty as reflected by these
information-theoretic measures (per esempio., Russo et al., 2020;
Henderson, Choi, Lowder, & Ferreira, 2016). Recent neu-
ral models of language processing locate the operation of
combining two elements into a syntactic representation
in Brodmann’s area 44 of Broca’s area, which forms a net-
work for processing of syntactic complexity in combina-
tion with the superior temporal gyrus (Fedorenko &
Blank, 2020; Zaccarella, Schell, & Friederici, 2017; for a
competing view, see Matchin & Hickok, 2020).
The theoretical debates outlined above would likely be
of interest to those who use cortical tracking to study au-
ditory signal processing, just as the cortical tracking of
speech is clearly relevant to psycholinguistic research.
Both fields have shown interest in discovering how lis-
teners might assign abstract linguistic structure to contin-
uous acoustic input as it unfolds and in determining the
neural correlates of spoken language processing.
BRIDGING THE TWO FIELDS
When spoken language is the signal, the studies of sen-
tence processing and of auditory signal processing share
the goal of characterizing the neural and cognitive archi-
tecture of language comprehension. Tuttavia, the two
areas have not extensively collaborated to address this
question. Research in sentence processing makes exten-
sive use of electrophysiological measures, particolarmente
through the use of ERPs, which have contributed greatly
to our understanding of language comprehension (Swaab
et al., 2012; Kutas & Federmeier, 2011; Kutas et al., 2006).
Beyond the now routine use of ERPs, many psycholin-
guists have also adopted time–frequency analyses of
EEG and magnetoencephalography data, as increases or
decreases in power at various frequency bands have been
found to correlate with several aspects of comprehension
(for reviews, see Prystauka & Lewis, 2019; Meyer, 2018;
Bastiaansen & Hagoort, 2006). The use of time–
frequency analyses has enabled researchers to make fuller
use of their data by including both synchronized and de-
synchronized neural activity, thus not discarding neural ac-
tivity that is not phase-locked to a stimulus (Bastiaansen,
Mazaheri, & Jensen, 2012). Yet, despite the widespread
use of ERPs and time–frequency analyses of neural oscil-
lations, the use of cortical tracking methods in particular
to answer psycholinguistic questions to date is relatively
rare. Importantly, cortical tracking implies a relationship
between the periodicities found in neural activity and
those found in the linguistic stimuli, which is different
from the types of time–frequency analyses already fre-
quently used in psycholinguistics. Strictly speaking, UN
power change in a certain frequency band does not imply
phase-locking (and by the same token, momentary phase
coherence does not imply an ongoing oscillation). As
mentioned earlier, most studies that use cortical tracking
of speech stem from the fields of signal processing, audi-
tory processing, and neuroscience.
Nonetheless, there are some notable examples of
research overlapping the methods and questions of
these two fields. Per esempio, Meyer and colleagues have
performed several experiments that measure cortical
tracking of speech using typical psycholinguistic experi-
mental designs to answer questions about syntactic
parsing, some of which we describe later in the
Contributions to Psycholinguistics section (per esempio., Meyer
& Gumbert, 2018; Meyer et al., 2017). Allo stesso modo, Martin
and Doumas (2017) have proposed a computational
model linking cortical tracking to the building of hierar-
chical representations of linguistic structure.
Tuttavia, beyond these emerging pockets of research
bridging the gap between sentence processing and signal
processing, the two fields remain largely independent.
One of the reasons for this may be that the two take very
different approaches to the study of language processing.
Although many researchers who use cortical tracking
methods are interested in signal processing more broadly
and consider speech to be one of many naturally occur-
ring complex signals (per esempio., Rimmele et al., 2018; Morillon
& Schroeder, 2015), sentence processing research typi-
cally emphasizes the linguistic properties of language
and the different levels of cognitive representations that
are generated during language processing, as discussed
in the previous section.
As is often the case in interdisciplinary research, a ma-
jor challenge lies in the discrepancies in terminology and
definitions across different literatures. In particular, stud-
ies in the two fields may sometimes even differ in their
definition of language comprehension (per esempio., the distinc-
tion between speech perception, processing, and com-
prehension; see Meyer, 2018) or may not specify the
degree or level of comprehension being assessed. Lan-
guage comprehension entails a range of cognitive pro-
cesses and levels of representations, which are sometimes
left underspecified because of shallow processing (Wang,
Bastiaansen, Yang, & Hagoort, 2011, 2012; Ferreira &
Patson, 2007; Ferreira, 2003; Sanford & Sturt, 2002;
Christianson, Hollingworth, Halliwell, & Ferreira, 2001).
Così, researchers attempting to bridge the two fields will
need to be aware of how comprehension is conceptualized
across studies.
More generally, the limited collaboration may stem
from the different methodological constraints that the
two fields typically prioritize, because of their different
theoretical backgrounds. In the following sections, we
578
Journal of Cognitive Neuroscience
Volume 33, Numero 4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
summarize some of the methodological constraints and
solutions that are often employed in signal processing
studies that use cortical tracking methods and in sen-
tence processing research and note how these con-
straints are sometimes at odds.
METHODOLOGICAL CONSTRAINTS
ACROSS FIELDS
Cortical Tracking Constraints
In cortical tracking studies, there are several experimen-
tal constraints that commonly arise. Foremost among
them is the requirement for long stretches of continuous,
varied speech. This follows from the signal processing
view that speech perception is a mathematically estima-
ble operation that transforms speech into brain re-
sponses continuously, with various responses generally
overlapping one another in time. A diverse family of tech-
niques known as system identification is well suited to
characterize such continuous input–output transforma-
zioni. In the system-identification framework, an input
signal x (per esempio., a sound) is transformed through a system
F(X) (per esempio., the brain)—which is not directly observable—
to produce an output signal y (per esempio., an EEG signal). By
presenting a range of systematically varied x inputs to
the system and measuring y, researchers can estimate
^f (X) to approximate the system that transforms input to
produzione (per esempio., the brain). Notice that these data-driven
approaches do not necessarily presume a certain relation-
ship between input (the signal) and output (the neural
risposta) and therefore may be agnostic to the specific
processes within the system that may contribute to the
transformation. Invece, they tend to approach neural
data with few a priori assumptions, to “discover” a rela-
tionship between the signal and the neural response.
This is achieved by offering the system (the listener) var-
ious instances of the input signal of interest (per esempio., spoken
sentences) and measuring how the output signal (per esempio.,
the EEG recording) responds differently at each moment.
Characterizing the speech–brain system entails modeling
this relationship.
Linguistic Stimulus Considerations
Most of the constraints signal processing researchers op-
timize for concern the stimulus, which serves as the in-
put signal. The first such constraint is that there must be
a certain degree of variability in the stimulus. Speech in-
put is often modeled by its slow (less than ∼16 Hz) power
fluctuations in the acoustic envelope. The acoustic enve-
lope reflects the perceptually salient syllabic structure of
speech and empirically relates to prominent cortical
ERPs, such as the N1 (Sanders & Neville, 2003).
Because modeling the speech–brain system is essentially
a statistical estimation problem (see Ljung, Chen, & Mu,
2020), the speech input must be varied in its properties
to sample all the possibly relevant values, and it must do
so without bias (or the analysis must explicitly correct for
any bias). With a lack of variability and naturalism in the
speech, the estimation will either be unrepresentative,
reflecting the idiosyncrasies of the specific speech cor-
pus, or it will fail to find a relationship at all.
A second constraint is that signal processing ap-
proaches may require numerous trials. Just as insufficient
stimulus variability can undermine the statistical estima-
tion described earlier, having too little data can lead to
invalid estimates or a failure to find a relationship be-
tween input and output signals. In principle, there is
no limit to the parameter space of this speech–brain sys-
tem, but here too, many instances of each parameter
must be presented to the listener. Inoltre, as in
any model estimation problem, the more “free parame-
ters” that must be characterized, the more data are usu-
ally required. System-identification approaches therefore
offer great flexibility and interpretive power, but at the
cost of acquiring more data and ensuring a statistically
balanced array of parameters, akin to using a Latin square
experimental design. A related constraint that often arises
in cortical tracking studies is the need to repeat identical
segments of speech multiple times. The motivation here
is the same as when creating a traditional, simple ERP: An
average response to multiple identical events (Dire, a tone)
will be a more representative estimate with a higher signal-
to-noise ratio than any individual response. Unsurprisingly, In
single-cell auditory neurophysiology, where a signal-
processing mindset has long dominated and influenced
many speech-tracking EEG investigators, repeated pre-
sentations are de rigueur (per esempio., in the venerable poststim-
ulus time histogram). In some cases, the data-driven
nature of system-identification techniques might compel
such averaging, but better signal-to-noise will help virtu-
ally any cortical tracking measure.
A third constraint concerns the periodicity of the audi-
tory stimulus itself. In contrast to system identification,
another class of influential cortical tracking experiments
manipulates the speech signal’s acoustic and linguistic
structures to be artificially periodic (per esempio., Ding et al.,
2016) and may result in stronger cortical tracking
(Meyer et al., 2020; Alexandrou, Saarinen, Kujala, &
Salmelin, 2018). From the signal processing perspective,
this is beneficial because it allows more straightforward
analysis of how the periodicity of speech input is
“tracked” by the brain. Specifically, frequency-domain
measures allow the investigator to focus only on those
periodicities of interest with relatively high statistical
power. Tuttavia, linguistic events are not strictly periodic
in time (Nolan & Jeon, 2014; see Beier & Ferreira, 2018,
for a discussion). This tension has further relevance to
questions of entrainment, particularly the hypothesis that
intrinsic brain oscillations become phase-reset or other-
wise temporally aligned with informative speech features.
Experimentally, to identify entrainment, it may be useful
to have well-defined periodicities as opposed to natural
Beier et al.
579
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
speech dynamics. Tuttavia, introducing such periodic-
ities limits the ability to determine whether entrainment
occurs for natural speech and, by extension, whether en-
trainment plays an active role in speech comprehension
outside the laboratory.
Behavioral Task Considerations
In cortical tracking paradigms, the main consideration re-
garding the inclusion of a behavioral task is that it should
not impede continuous EEG recording or contribute sig-
nificant noise to the data. Cortical tracking studies may
employ a passive listening paradigm (per esempio., Keitel, Ince,
Gross, & Kayser, 2017; Gross et al., 2013) to obtain
EEG recording that is not continually interrupted by mo-
tor movements (per esempio., button presses) from behavioral
tasks on the assumption that spoken language is auto-
matically processed even if there is no offline behavioral
task (per esempio., in ERP studies presenting auditory sentences;
van Berkum, 2004), an assumption that is sometimes
held in sentence processing research as well. Relatedly,
investigators may omit a behavioral task to prevent un-
natural processing strategies that may add noise to the
EEG data (for further discussion, see Hamilton & Huth,
2020).
In some cases, Tuttavia, signal processing studies do
include a behavioral task to encourage participants to at-
tend to the auditory signal. For instance, participants may
be given probe words and asked to indicate whether they
heard those words on a previous trial (per esempio., Keitel et al.,
2018; Falk, Lanzilotti, & Schön, 2017), or they may be
asked to detect semantic anomalies in the sentence
(Meyer & Gumbert, 2018). Alternatively, participants
may be asked to count the number of words or syllables
in the presented sentences (Batterink & Paller, 2017) O
to press a button every nth-word (per esempio., fourth-word) sen-
tence (Getz, Ding, Newport, & Poeppel, 2018). To ensure
attention to the acoustic properties of the signal, cortical
tracking studies may include acoustic or temporal devia-
tion tasks, which require participants to indicate when or
whether the pitch or loudness changed in the speech
they heard (Zoefel et al., 2018; Rimmele, Golumbic,
Schröger, & Poeppel, 2015). In investigations of language
comprehension as opposed to low-level acoustic percep-
zione, an offline task is sometimes included to ensure lis-
teners interpreted the utterance successfully, such as a
self-report of the number of words participants under-
stood in the signal (Baltzell et al., 2017; Peelle et al.,
2013) or comprehension questions (Weissbart, Kandylaki,
& Reichenbach, 2020; Biau, Torralba, Fuentemilla, Di
Diego Balaguer, & Soto-Faraco, 2015). In summary, be-
cause signal processing studies are primarily concerned
with characterizing the processes that lead to compre-
hension in real time, behavioral tasks are often viewed
as tools to encourage participants to pay attention to
an input signal.
Although the experimental requirements of cortical
tracking studies tend to be rather technical in nature,
they do illustrate why signal processing research has long
placed such great emphasis on the continuous nature of
speech processing: not only because this is likely how the
brain works but also because this is mathematically inher-
ent to the most common techniques (including both
time series system-identification and frequency-domain
analyses). The constraints also reflect the data-driven
nature of signal processing approaches, which can be
theory agnostic, in part because they might require a
representative and unbiased sampling of the speech–
brain activity to achieve a valid result.
Psycholinguistic Constraints
In a typical sentence processing experiment, participants
read or listen to sentences with manipulations relevant to
the theoretical question that the experiment is designed
to evaluate (per esempio., sentences with grammatical, semantic,
or pragmatic anomalies). Researchers then compare pro-
cessing of those sentences to sentences in a baseline con-
dition that do not contain any anomaly but are otherwise
identical to the experimental sentences (cioè., “minimal
pairs” that are controlled for lexical, semantic, and syn-
tactic features). A difference in averaged behavioral
response between the experimental and baseline
conditions (per esempio., longer RTs or reading times) would in-
dicate processing effects that are because of the linguistic
manipulation. Importantly, the experiment is designed to
control for extraneous variables and to ensure that the
measures reflect specific cognitive and neural mecha-
nisms that underlie language comprehension. To address
these concerns, a set of guidelines for designing the ex-
perimental stimuli and the behavioral tasks has become
mainstream in psycholinguistics over the years.
Linguistic Stimulus Considerations
In sentence processing research, linguistic stimuli are
typically controlled for factors that are known to influ-
ence processing to rule out potential confounds. For ex-
ample, the length and frequency of words used can affect
the magnitude and timing of ERPs (Strijkers, Costa, &
Thierry, 2010; Hauk & Pulvermüller, 2004; King &
Kutas, 1998), and therefore, word frequency is either
controlled at the stimulus creation stage (per esempio., selecting
words with a similar frequency from a database) or statis-
tically controlled by including frequency in a model at the
analysis stage. Relatedly, function words (per esempio., preposi-
zioni, determiners) evoke different neural responses
than do content words (per esempio., nouns, verbs): The former
evokes a left-lateralized negative shift that the latter does
non (Brown, Hagoort, & ter Keurs, 1999). The amplitude
of the N400 response to concrete words is larger than
that for abstract words, and there is greater right-
hemisphere activity for concrete words (Kounios &
580
Journal of Cognitive Neuroscience
Volume 33, Numero 4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Holcomb, 1994). Words with a higher orthographic
neighborhood density (per esempio., the number of words with
a similar orthographic representation, such as “lose”
and “rose”) or a phonological neighborhood density
(per esempio., “cat” and “kit”) evoke greater N400 negativity than
those with a low neighborhood density ( Winsler,
Midgley, Grainger, & Holcomb, 2018; Holcomb, Grainger,
& O’Rourke, 2002).
The position of the target word in a sentence can also
affect processing and therefore must be taken into con-
sideration when designing stimuli. Per esempio, IL
N400 amplitude is larger for words that occur earlier,
and the effect is attenuated by word frequency ( Van
Petten & Kutas, 1990). Low transition probability from
word to word, or even syllable to syllable, can evoke
the N400 response (Teinonen & Huotilainen, 2012;
Kutas & Federmeier, 2011; Cunillera, Toro, Sebastián-
Gallés, & Rodríguez-Fornells, 2006). To control for tran-
sition probability, stimuli are typically cloze-normed
(Taylor, 1953). In the cloze procedure, participants read
fragments of the experimental sentences and are asked
to provide the word(S) that best completes the sentence.
The proportion of a given response out of all responses
provided is the cloze probability for that response and is
thought to index its predictability for the preceding con-
testo. Per esempio, if participants read the sentence “It
was a breezy day so the boy went outside to fly a
_____” and 90% of them responded with the word “kite”,
then the word “kite” has a cloze probability of 90% E
would be considered a highly predictable sentence con-
tinuation. A sentence that violates phrase structure rules
or is otherwise ungrammatical (per esempio., agreement errors,
such as “The doctors is late for surgery”) can trigger an
early left anterior negativity (ELAN) as well as the P600
component (Friederici & Meyer, 2004; Friederici, 2002).
By norming stimuli for acceptability, which is a proxy for
sentence grammaticality that is more accessible to naive
raters (Huang & Ferreira, 2020), stimuli that do or do
not evoke ERPs linked to structural violations can be se-
lected, depending on the research question. Inoltre,
stimuli are frequently normed for typicality, plausibility,
and naturalness. Stimuli that are atypical or implausible
will likely evoke N400 responses, and unnatural stimuli
could evoke N400 or P600 components, depending on
what aspect of the linguistic content leads raters to indi-
cate that they seem unnatural.
Behavioral Task Considerations
In addition to carefully controlling the experimental
stimuli, psycholinguistic experiments typically include
an offline behavioral task to verify that participants
comprehended the stimuli, such as employing true–false
questions (Brothers, Swaab, & Traxler, 2017), semantic
judgments ( Wang, Hagoort, & Jensen, 2018), or asking
participants to evaluate each sentence or narrative based
on their grammaticality, acceptability, or plausibility (Vedere
Myers, 2009). For research investigating lower-level lan-
elaborazione del calibro, such as acoustic representations, Rif-
searchers have used simple detection tasks such as
asking participants to monitor a particular phoneme
and to press a response key when they hear that pho-
neme, but the use of detection tasks has declined in sen-
tence processing research for various reasons (for a
revisione, see Ferreira & Anes, 1994).
Some studies of speech comprehension do not include
any explicit tasks besides passive listening (per esempio., van
Berkum, Brown, Zwitserlood, Kooijman, & Hagoort, 2005;
see van Berkum, 2004, for a discussion). These paradigms
circumvent issues associated with metalinguistic judg-
ments and may prevent unnatural processing strategies
(see Hamilton & Huth, 2020). They are also particularly
advantageous when investigating language processing
in special populations who may have impaired ability to
perform additional behavioral tasks, such as some autistic
individuals and children (Brennan, 2016). In studies of
typical language processing in adults, Tuttavia, compre-
hension tasks are often used because they allow re-
searchers to directly measure the interpretation that
participants have generated after processing the stimuli
(Ferreira & Yang, 2019). It can be argued that compre-
hension tasks are crucial because passively presenting
the linguistic stimuli to participants does not guarantee
that they have fully analyzed the linguistic material and
have generated an interpretation for it. Invece, Essi
may engage in shallow processing and come away with
an incomplete or even incorrect interpretation of the
sentence (Ferreira & Patson, 2007; Ferreira, 2003;
Christianson et al., 2001). Omitting a comprehension
task may also encourage participants to adopt idiosyn-
cratic goals during the experiment (Salverda, Brown, &
Tanenhaus, 2011). Different task demands have addition-
ally been shown to affect the extent to which people en-
gage in basic linguistic processing such as resolving
anaphors, structure building, and inferencing (Foertsch
& Gernsbacher, 1994) as well as lexical prediction
(Brothers et al., 2017). Finalmente, the behavioral task partic-
ipants engage in can systematically influence language-
related ERP components, including the N400 (Chwilla,
Brown, & Hagoort, 1995; Bentin, Kutas, & Hillyard,
1993; Deacon, Breton, Ritter, & Vaughan, 1991) and the
P600 (Schacht, Sommer, Shmuilovich, Martíenz, &
Martín-Loeches, 2014; Gunter & Friederici, 1999).
The presence and type of behavioral task will vary de-
pending on the research questions and goals of the
study. Nonetheless, it may be necessary to consider the
kind and depth of language processing that is induced in
the experiment, as motivation and strategies are known
to affect language comprehension (Ferreira & Yang,
2019; Alexopoulou, Michel, Murakami, & Meurers,
2017). Well-controlled linguistic stimuli and behavioral
tasks enable sentence processing researchers to draw
conclusions about the specific cognitive and neural
mechanisms that support language comprehension,
Beier et al.
581
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
including the time course of these processes as well as the
generated interpretation resulting from comprehension.
Reconciling Constraints across Fields
Sentence processing researchers who wish to include
cortical tracking as a method and signal processing re-
searchers who wish to employ more linguistic control
in their stimuli both face the challenge of taking into ac-
count the methodological constraints of both fields.
Some of the methodological constraints that psycholin-
guists must satisfy are difficult to reconcile with the con-
straints researchers who use cortical tracking methods
reckon with. Per esempio, psycholinguists carefully con-
trol the linguistic content of their stimuli (per esempio., surprisal,
plausibility, acceptability), which is tractable when the
number of items is relatively small, and they avoid repeat-
ing the same item in an experimental session because of
the effects of priming and learning. Tuttavia, signal pro-
cessing studies can require numerous trials, che è
sometimes achieved through repeated exposure to the
same item, which listeners could habituate to or over-
Imparare. Allo stesso modo, signal processing studies often require
long stretches of signal. It is difficult to create the num-
ber of unique experimental items following psycholin-
guistic conventions (per esempio., stimulus norming) to generate
the type of dataset that signal processing studies may re-
quire, and it is similarly difficult to control the linguistic
properties of speech in lengthy recordings. Likewise,
without an explicit comprehension task, the level of com-
prehension that took place and the participants’ motiva-
tion are unclear, but signal processing studies may need
to minimize interruptions and motor movements during
EEG recording.
All interdisciplinary work requires researchers to draw
from the methods used across multiple fields and ulti-
mately agree on a common methodology. Because some
methodological choices are mutually exclusive with
others, no experiment can realistically meet all methodo-
logical standards. Invece, it is necessary to evaluate con-
straints from each field and then prioritize those that are
most applicable to the research question at hand. Nel
event that conflicting constraints cannot be reconciled, Esso
is worth explicitly acknowledging the validity of the con-
straints that could not be applied and briefly stating why
they were not given priority (per esempio., we could not use nat-
uralistic stimuli because it was critical to our design to
eliminate prosodic cues to syntax).
Although some constraints may be mutually exclusive
—for example, the preference for more periodicity in the
acoustic signal to measure cortical tracking, which con-
flicts with the pressure for speech to sound as natural
as possible—there are steps that can be taken to recon-
cile other constraints without posing an undue burden
on researchers. Per esempio, even if stimuli cannot be
normed or constructed to control for linguistic content,
measures of some constructs (such as word frequency,
surprisal, and entropy) can be obtained for existing stim-
uli using computational models (Hamilton & Huth, 2020;
Weissbart et al., 2020; Brennan, 2016; Willems, Frank,
Nijhof, Hagoort, & van den Bosch, 2016) and then con-
trolled for statistically. Such an approach allows re-
searchers to quantify these measures for naturalistic
stimuli, which can be used to assess cortical tracking in
everyday language comprehension (Alexandrou,
Saarinen, Kujala, & Salmelin, 2020; Alexandrou et al.,
2018).
It is important to note that both sentence and signal
processing experiments tend to use stimuli that differ
from the kinds of sentences and utterances that occur
in everyday language. Experiments in both areas tend
to make use of read speech, which gives researchers
good control over the linguistic content of the stimuli
but is easier to comprehend (Uchanski, Choi, Braida,
Reed, & Durlach, 1996; Payton, Uchanski, & Braida,
1994), is produced at a lower speech rate (Hirose &
Kawanami, 2002; Picheny, Durlach, & Braida, 1985;
Crystal & House, 1982), and has exaggerated acoustic fea-
tures relative to spontaneous speech (Gross et al., 2013;
Finke & Rogina, 1997; Nakajima & Allen, 1993; Vedere
Alexandrou et al., 2020, for a discussion), which may af-
fect processing. Further work using naturalistic utter-
ances is required both to determine the degree to
which cortical tracking occurs when listening to sponta-
neous speech and to assess whether sentence processing
models generalize to everyday speech. Nonetheless, IL
estimable linguistic parameters available using computa-
tional models are compatible with the system-identification
techniques commonly used in signal processing research.
Parameters such as spectrotemporal, phonetic/phonemic,
or linguistic properties can be included in the statistical
modello, for instance, in approaches that elaborate on mul-
tiple regression (Sassenhagen, 2019; Di Liberto & Lalor,
2017; Crosse, Di Liberto, Bednar, & Lalor, 2016). These
can take continuous values, as with the acoustic envelope,
or categorical ones, as with phonemic class. Inoltre,
they can be modeled as changing smoothly through time
(power) or occurring only at specific, transitory moments
(per esempio., sentence onsets, syntactic boundaries, prosodic
breaks). Tuttavia, as noted before, increasing the number
of parameters also requires more data.
When the research question mandates the use of lin-
guistically controlled stimuli, as in many sentence pro-
cessing studies as well as signal processing experiments
that rely on stimuli with a fixed structure, it is worth not-
ing that the stimuli need not be generated from scratch.
It has been standard practice in psycholinguistics for
decades to share stimuli in an appendix or in other sup-
plementary materials, which means entire lists of well-
controlled stimuli that have already been normed are
readily available in published papers. Inoltre, there
are large freely available stimulus sets that researchers
have compiled for general use, Per esempio, a data set
of cloze norms for over 3,000 English sentences (Peelle
582
Journal of Cognitive Neuroscience
Volume 33, Numero 4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
et al., 2020). Audio files for prerecorded stimuli may be
found in online repositories such as the Open Science
Framework or may be made available by authors upon
request. Stimulus crowdsourcing—asking users of a
crowdsourcing platform to generate stimuli that meet
study-relevant constraints—is an additional strategy re-
searchers can employ to alleviate the burden of stimulus
generation. Tuttavia, whatever the source of the corpus,
the challenge of controlling stimuli, balancing factorial
designs, and including filler items may lead to small num-
bers of trials. Although it is possible to conduct a rigorous
study using cortical tracking methods with relatively few
stimuli (per esempio., Meyer et al., 2017, Nitems = 40), achieving
adequate statistical power might require compensation
through using extended recording times, multiple ses-
sions, or a larger number of test participants.
Inoltre, filler items are often helpful for providing
listeners with a greater variability of stimulus types while
at the same time reducing the likelihood that participants
will overlearn the stimulus structure and, as a result, en-
gage in shallow comprehension of the stimuli. This is
particularly relevant for the assumption that neural data
reflect a continuous process, as repetitive stimuli may in-
duce shallow or atypical processing of the speech signal, In
which case the function estimated by system-identification
techniques may not correspond to more naturalistic pro-
cessazione. We return to this idea in the following section.
The behavioral task constraints across sentence pro-
cessing and signal processing research can also be recon-
ciled to quantify the level of comprehension more
explicitly in cortical tracking paradigms. Adding a com-
prehension task to an experiment is a relatively easy
way to motivate participants to engage in detailed com-
prehension of the stimuli. For instance, simple yes/no
questions about the meaning of the stimuli encourage
participants to construct elaborated semantic representa-
tions rather than attending to only the surface structure.
Comprehension questions can be presented to partici-
pants in between blocks, or on the filler items only, A
avoid introducing neural responses related to decision-
making on the experimental trials and to ensure that mo-
tor movements do not interfere with the EEG recording.
If the paradigm would not allow a comprehension task to
be intermingled with the experimental trials, experi-
menters can motivate careful attention to the stimuli by
telling participants at the start of the experiment to antic-
ipate a memory test at the end of the session.
Speech is a continuous signal, and psycholinguists who
study sentence processing are ultimately interested in
understanding how listeners interpret that continuous
signal. Nevertheless, there is a tendency to examine lan-
guage processing through the analysis of discrete events,
in part because of the conventions and analysis approaches
used historically in the field (Jewett & Williston, 1971). A
understand how listeners process real-time spoken input,
sentence processing would benefit from adopting the
methodological and analysis techniques employed in the
study of signal processing for working with continuous
dati. Inoltre, it is far less common in sentence pro-
cessing work for researchers to consider the periodicity
of the signal, or variability in the amplitude envelope,
which can affect neural signals. By understanding how
these acoustic features impact EEG recordings, psycho-
linguists conducting sentence processing experiments
can better separate the relative contribution of acoustic
and linguistic properties of their experimental stimuli,
which would allow them to draw stronger conclusions
about how linguistic features (per esempio., lexical ambiguity) ulti-
mately influence comprehension.
Finalmente, although cortical tracking is an inherently tem-
poral phenomenon, linguistic attributes may strongly
affect which cortical areas are involved, and thus the
“spatial” pattern of tracking time series across EEG scalp
channels. Many neuroimaging studies using fMRI and
PET show spatial cortical activation patterns that distin-
guish lexical category or semantics (nouns vs. verbs,
concrete vs. abstract), syntax (argument structure), E
numerous other features (for examples, see Rodd, Vitello,
Woollams, & Adank, 2015; Moseley & Pulvermüller, 2014;
Price, 2012; Friederici, 2011). Insofar as the EEG scalp
activation pattern reflects (indirectly) the locations and
orientations of cortical sources, controlling such linguistic
variables should lead to more consistent and representa-
tive tracking analyses.
In summary, although many of the constraints associ-
ated with conducting signal processing and sentence pro-
cessing research may appear to be at odds, there are
reasonable compromises that can be made to reconcile
methodologies from both fields. As the difficulty of col-
laboration between these areas is partly because of meth-
odological differences, some of these solutions may
make it easier for both sentence processing and signal
processing researchers to use cortical tracking to better
understand the neural and cognitive processes underly-
ing language comprehension. In the following sections,
we further elaborate on what these two fields have to
gain from this collaboration and provide more detailed
examples of ways to incorporate each field’s methods
and standards.
Contribution to Signal Processing Research
What can the study of signal processing, using cortical
tracking methods, gain from developing stimuli that sat-
isfy certain psycholinguistic constraints? Stimuli that are
implausible, anomalous, or otherwise unnatural in some
manner elicit ERP components (per esempio., N400s, P600s,
ELAN), which will affect oscillations if they occur in
the same frequency band and therefore could contrib-
ute unwanted noise if not intentionally manipulated.
Repetition of the same (or highly similar) sentence or
the same syntactic frame throughout the study could also
have unintended processing effects. Syntactic similarity
across sentences produces structural priming, in which
Beier et al.
583
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
structural similarity between previous sentences facili-
tates processing of the current sentence (Tooley,
Traxler, & Swaab, 2009; Pickering & Ferreira, 2008;
Bock, 1986). Unintended priming effects should be
avoided because it is unclear how structural priming of
this sort might influence cortical tracking of speech or al-
ter the EEG signal in unintended ways. Another concern
is that, when a sentence template is used frequently, IL
listener can overlearn the template and employ a behav-
ioral strategy that undermines the study. Per esempio, if
the task is to detect a word and the target word repeatedly
occurs in the same location in the sentence, listeners could
successfully circumvent the intended purpose of the com-
prehension task by attending only to the target region of
the sentence. A shallow processing strategy of this sort
would allow for high performance on the task without
the need to comprehend the sentence. This problem could
be avoided by including filler items with different sentence
structures and varying the location of the target word
within the experimental items, when possible. When
the syntactic structure is highly predictable because of
overlearning, it may additionally attenuate the EEG sig-
nal (Tooley et al., 2009).
Importantly, controlling for the linguistic aspects of
stimuli may also help researchers determine whether cor-
tical tracking reflects evoked responses or intrinsic oscil-
lations. If stimuli are controlled such that we can
determine when and where larger ERPs should occur,
variation introduced by ERPs may be more readily disso-
ciated from variation because of intrinsic oscillations.
Through the availability of computational models, many
linguistic factors can be controlled for statistically
(Hamilton & Huth, 2020; Weissbart et al., 2020;
Brennan, 2016; Willems et al., 2016), which would allow
researchers to use lengthy, naturalistic auditory stimuli
that are often required in signal processing experiments
and still account for linguistic constraints. Controlling for
linguistic factors that are known to induce processing dif-
ficulty or to otherwise affect language processing will yield
cleaner data and will provide greater context for interpret-
ing variations in the EEG signal.
In addition to carefully controlling the stimuli, it may
be worthwhile to include an explicit comprehension task
to ensure participants engage in detailed comprehension
while listening to the stimuli, especially if the research
aim is to test the role of cortical tracking in comprehen-
sion. As discussed in the Psycholinguistic Constraints sec-
zione, listeners’ strategies for comprehension can vary
depending on the goal and the task demands (Vedere
Ferreira & Yang, 2019), and shallow processing can
sometimes lead to underspecified or even incorrect rep-
resentations (Ferreira & Patson, 2007; Sanford & Sturt,
2002), thus potentially adding noise to the neural data
corresponding to these cognitive processes. It is impor-
tant to acknowledge that naturalistic paradigms have
numerous advantages (see Hamilton & Huth, 2020;
Brennan, 2016), and it is indeed not necessary to include
a comprehension task if the study’s goal does not pertain
to higher-level language comprehension per se (per esempio., using
cortical tracking methods to investigate the sequential
grouping of syllables into words, not sentence or discourse-
level comprehension). Nevertheless, even in this case, it is
worthwhile to consider how task effects impact EEG data
because any neural response that has not been accounted
for has the potential to add noise. As mentioned previously,
language-related ERPs have been shown to vary depending
on the level of processing induced by different tasks (per esempio.,
Chwilla et al., 1995; Bentin et al., 1993).
In selecting a behavioral task that addresses compre-
hension, there are a number of considerations regarding
the kind of processing that is induced by the task. When
appropriate, comprehension questions are ideal because
they enable researchers to quantify the level of compre-
hension that took place, and they may be a better alter-
native to self-reported intelligibility because they
circumvent unconscious biases. Word detection and
anomaly detection tasks are useful in encouraging partic-
ipants to attend to the sentences, but participants may
not necessarily engage in detailed comprehension be-
cause these tasks tap into memory for the surface struc-
ture rather than the overall meaning of the sentence.
Temporal or acoustic deviation tasks, in which partici-
pants indicate when or whether the pitch, loudness, O
timing changed in the speech, have similar limitations
to detection tasks because they only index attention to
the acoustic properties of the speech signal, piuttosto che
tapping into the processing of the linguistic content of
the speech stream. Inoltre, ERPs have been shown
to be influenced by whether participants are instructed to
pay attention to speech rhythm or syntax (Schmidt-
Kassow & Kotz, 2009UN, 2009B), which may also lead to
overall noisier and potentially misleading EEG data.
In summary, signal processing research using cortical
tracking can reap various benefits from designing stimuli
and behavioral tasks that fulfill the previously described
psycholinguistic constraints. If cortical tracking of speech
potentially serves a functional role in speech comprehen-
sion, it would be crucial to ensure that the electrophysi-
ological recordings reflect comprehension of the
linguistic material, in which participants build syntactic
structures, commit to a sentence interpretation, resolve
anaphors and ambiguity, and make inferences when ap-
plicable. To this aim, including comprehension questions
yields a direct measure of linguistic processing and en-
courages a more detailed analysis of the sentence struc-
ture and meaning. Comprehension tasks also provide an
explicit goal of comprehension for participants and pre-
vent idiosyncratic goals and strategies, which reduce
noise in the data from these extraneous factors. Nel
data analysis stage, the use of information-theoretic mea-
sures in statistical control can be easily implemented to
account for systematic noise concerning syntactic and se-
mantic processing. A key advantage of this computational
approach is that it can be used on large stretches of
584
Journal of Cognitive Neuroscience
Volume 33, Numero 4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
naturalistic uncontrolled stimuli, bolstering the goal of in-
vestigating naturalistic language processing that is an
emerging trend in both signal processing and sentence
processing research (see Alexandrou et al., 2018, 2020;
Hamilton & Huth, 2020; Alday, 2019; Brennan, 2016).
More generally, the computational modeling approach
can also elucidate the role of cortical tracking in instan-
tiating temporal predictions, as information-theoretic
modeling can identify the rich linguistic information in
the signal that is coded by the brain. Signal processing
researchers who are interested in using cortical tracking
to study predictive coding can benefit from quantifying
the depth of processing that took place because predic-
tive processing will depend on how deeply the linguistic
material was processed, which is in turn influenced by
the presence and type of behavioral task (for further
discussion, see Kuperberg & Jaeger, 2016). Overall, IL
endeavor of studying auditory signal processing can be
greatly augmented by accounting for linguistic aspects
in the stimuli when spoken language constitutes the
signal and by employing behavioral tasks that enable
explicit assessment of the depth of comprehension that
took place.
Contributions to Psycholinguistics
Sentence processing research has long studied syntactic
ambiguity to differentiate between contrasting theoreti-
cal accounts of cognitive parsing mechanisms. In a recent
study, Meyer et al. (2017) presented ambiguous sen-
tences such as “The client sued the murderer with the
corrupt lawyer” that either did or did not include a disam-
biguating prosodic break before the prepositional
phrase. Cortical tracking in delta-band oscillations re-
flected syntactic phrase groupings, which frequently—
but not always—corresponded to the prosodic grouping
(Bögels, Schriefers, Vonk, & Chwilla, 2011; Clifton, Carlson,
& Frazier, 2002; Cutler, Dahan, & van Donselaar, 1997;
Shattuck-Hufnagel & Turk, 1996; Ferreira, 1993), generat-
ing new evidence that syntactic grouping biases can over-
ride acoustic grouping cues. Cortical tracking methods
could be applied further using temporarily ambiguous sen-
tences to help differentiate between sentence parsing
models.
Per esempio, Ding et al. (2016) found that listeners
showed cortical tracking to syntactic phrase boundaries
(per esempio., cortical tracking reflects the subject noun phrase
and verb phrase boundary). If tracking of syntactic
boundaries generalizes beyond the stimulus materials
that Ding et al. used, then using cortical tracking to tem-
porarily ambiguous sentences should reveal the parsing
mechanisms at play. Consider the temporarily ambiguous
garden-path sentence “The government plans to raise
taxes failed.” The sentence fragment “The government
plans to raise taxes” is ambiguous because the subject
of the sentence is ambiguous (1).1 “The government”
could be the subject of the verb “plans” (1UN), or “The
government plans” could be the subject of a sentence
in which “government plans” is a compound noun (1B).
1. UN) [S [NP The government] [VP plans …]
B) [S [NP The government plans] [VP …]
Before the disambiguating word (“failed”), either
interpretation of the sentence is viable. Garden-path
effects suggest that comprehenders initially assume the
structure in 1a (MacDonald, 1993; Frazier & Rayner,
1987). The structure is initially favored at least in part
because “plans” occurs more frequently as a verb (59
occurrences) than as a noun (two occurrences) in this
particolare contesto (Corpus of Contemporary American
English; Davies, 2008).
Sentence processing theories disagree with respect to
whether multiple structures are considered simulta-
neously and on where in the sentence the parser will en-
counter difficulty. Serial processing models (per esempio., Frazier,
1987; Frazier & Fodor, 1978) build only one structure at a
time, and reanalysis only occurs when the parser at-
tempts to integrate a syntactic unit that is not compatible
with the structure. In the sentence under consideration,
encountering the verb “failed” would trigger reanalysis.
Parallel processing models (per esempio., MacDonald, Pearlmutter,
& Seidenberg, 1994; Trueswell & Tanenhaus, 1994), COME
the name implies, generate multiple structures and nar-
row down the field of candidates as the parser encoun-
ters more and more disambiguating information, Quale
means the parser should encounter the greatest difficulty
during the ambiguous region of the sentence (before
“failed”).
Under a parallel processing model, during the tem-
porarily ambiguous region of a sentence, at least two
competing parses (1a and 1b) are actively under consider-
ation. Crucially, the syntactic phrase boundaries differ be-
tween the two structures early on in the sentence. Noi
would expect to see cortical tracking to phrase boundaries
corresponding to each of the competing parses during the
ambiguous portion of the sentence as the parser considers
multiple viable candidates. In contrasto, under a serial
processing model, only one parse (per esempio., 1UN) would be con-
sidered at a time, and the delta-band oscillatory phase
should indicate the parse under consideration. We would
therefore predict cortical tracking to syntactic phrase
boundaries that are consistent with the parse under
consideration only, and we would expect delta-band oscil-
latory phase reset to occur once contradictory evidence is
encountered. Così, cortical tracking methods provide us
with a unique opportunity to resolve some theoretical
issues that have proven difficult to disentangle using
common behavioral methods such as the recording of
eye movements during reading (Figura 1).
Inoltre, signal processing studies that compare
cortical tracking to attended versus unattended speech
suggest that we might be able to study depth of
Beier et al.
585
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. Illustration of
predicted delta-band oscillation
responses under (UN) serial and
(B) parallel processing accounts
of sentence parsing. Cortical
tracking of verb phrase
boundaries indicated by a reset
in oscillatory phase. IL
approximation of delta
oscillations at 1 Hz is simplified
(relative to an actual EEG
recording) for clarity.
processing by measuring the degree of cortical tracking
of speech. There is evidence that listeners employ shal-
low processing to efficiently construct a “good-enough”
interpretation of the sentence (Ferreira & Patson, 2007;
Ferreira, 2003; Christianson et al., 2001). Per esempio,
Ferreira (2003) presented listeners with sentences de-
scribing transitive events that either were plausible or im-
plausible, and had active or passive syntax, and found a
tendency for listeners to transform implausible passive
sentences (per esempio., “The dog was bitten by the man”) into
actives (per esempio., “The dog bit the man”), thereby “correcting”
the noncanonical nature of both the syntax and meaning
of the sentence. The degree of cortical tracking to
speech may be able to predict whether or not the listener
used a heuristic strategy when processing the sentence.
Specifically, we might expect weak cortical tracking to
“The dog was bitten by the man” to predict a listener
arriving at the incorrect but more felicitous “The dog
bit the man” interpretation.
Cortical tracking would supplement not only behavioral
methods but also the measures of neural activity already
in use in the field of sentence processing. Cortical track-
ing goes beyond the use of ERPs to study language
processing in that it can reveal processes occurring
continuously, rather than being constrained by neural re-
sponses to discrete events. This could facilitate the pro-
cess of generating linguistic stimuli, which are often
required to be built around specific target words in many
current designs; using cortical tracking methods, sentence
processing researchers may be able to expand to more
naturalistic stimuli. Cortical tracking methods also go be-
yond the time–frequency analyses currently in use in sen-
tence processing research by observing neural activity
that is phase-aligned to periodicities in the stimuli. As
we have shown, this property may be exploited to mea-
sure how comprehenders deal with stimuli presenting
ambiguous structures. Whereas the types of time–
frequency analyses already in use add an invaluable piece
to our understanding of language comprehension
(Prystauka & Lewis, 2019), cortical tracking tools will un-
doubtedly add to the types of linguistic questions and
paradigms that can be addressed through the recording
of EEG and magnetoencephalography data.
In summary, there are exciting opportunities to inves-
tigate psycholinguistic theories by studying cortical track-
ing of speech and to use psycholinguistic methods to
further elucidate the relationship between cortical track-
ing and cognitive processes associated with language
processing and comprehension. As we have argued, cor-
tical tracking may help resolve long-standing debates
such as whether parsing occurs in a serial or parallel fash-
ion, which have been left unresolved by behavioral
methods and the measures of neural activity currently
employed in this field.
Conclusione
The fields of sentence and signal processing both seek to
understand how listeners process speech, yet collabora-
tion between the two fields has been limited. We out-
lined several barriers to collaboration, with the primary
ones being the different methods used across fields as
well as differences in the constraints that experiments
in each field must satisfy. Although some of those con-
straints are at odds with each other, many can be recon-
ciled. We advocate for further collaboration across fields,
which would require researchers in each area to acknowl-
edge the experimental constraints of the other and to in-
tegrate interdisciplinary methods in their own work,
whenever possible. We believe both sentence processing
and signal processing research would benefit as a result,
because (1) avoiding linguistic stimulus confounds would
help determine whether cortical tracking reflects evoked
586
Journal of Cognitive Neuroscience
Volume 33, Numero 4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
responses or neural entrainment, (2) psycholinguists
could pursue research questions that current methods
(per esempio., ERPs) are not well suited to address, E (3) lan-
guage processing models in psycholinguistics could be
better informed by incorporating findings from signal
processing work. More broadly, both fields would be able
to make a fuller use of their data. Signal processing re-
searchers could reduce unwanted noise by controlling
and manipulating linguistic features of their stimuli that
are often overlooked and ensuring that full comprehen-
sion takes place. Sentence processing researchers could
better interpret real-time processing by measuring con-
tinuous neural activity corresponding to the structure
of the stimuli, rather than limiting themselves to observa-
tions of neural responses to discrete events such as par-
ticular target words. Further collaboration will give rise to
new and exciting scientific discoveries of interest to both
research communities.
Ringraziamenti
We thank the two anonymous reviewers for their insightful
comments, which helped us greatly improve our paper.
Opinions, interpretations, conclusions, and recommendations
are those of the authors and are not necessarily endorsed by
the Department of Defense.
Reprint requests should be sent to Eleonora J. Beier, Psychology
Department, University of California, Davis, One Shields Ave.,
Davis, CA 95616-5270, or via e-mail: ejbeier@ucdavis.edu.
Funding Information
We acknowledge support from the National Science
Foundation (http://dx.doi.org/10.13039/100000001) GRFP
number 1650042 awarded to E. J. B., National Science
Foundation (http://dx.doi.org/10.13039/100000001) grant
BCS-1650888 awarded to F. F., and National Institutes of
Health grant 1R01HD100516 awarded to F. F. This work
was also supported by the Office of the Assistant
Secretary of Defense for Health Affairs (http://dx.doi.org
/10.13039/100000005) through the Hearing Restoration
Research Program, under award no. W81XWH-20-1-0485,
to L. M. M.; the National Institutes of Health (http://dx
.doi.org/10.13039/100000002), with grant no. R56
AG053346-02 awarded to G. R.; and the Chulalongkorn
Università (http://dx.doi.org/10.13039/501100002873), con
grant no. CU_GIF_62_01_38_01 awarded to S. C.
Diversity in Citation Practices
A retrospective analysis of the citations in every article
published in this journal from 2010 A 2020 has revealed
a persistent pattern of gender imbalance: Although the
proportions of authorship teams (categorized by esti-
mated gender identification of first author/last author)
publishing in the Journal of Cognitive Neuroscience
( JoCN ) during this period were M(an)/ M = .408,
W(oman)/M = .335, M/W = .108, and W/W = .149, IL
comparable proportions for the articles that these author-
ship teams cited were M/M = .579, W/M = .243, M/W =
.102, and W/W = .076 (Fulvio et al., JoCN, 33:1, pag. 3–7).
Consequently, JoCN encourages all authors to consider
gender balance explicitly when selecting which articles to
cite and gives them the opportunity to report their article’s
gender citation balance. The authors of this article report
its proportions of citations by gender category to be as
follows: M/M = .433, W/M = .133, M/W = .167, E
W/W = .267.
Note
1. For ease of explanation, we opted to show simplified syn-
tactic structures and only the relevant syntactic phrase bound-
aries in this example.
REFERENCES
Abney, S. P., & Johnson, M. (1991). Memory requirements and
local ambiguities of parsing strategies. Journal of
Psycholinguistic Research, 20, 233–250. DOI: https://doi.org
/10.1007/BF01067217
Ahissar, E., Nagarajan, S., Ahissar, M., Protopapas, A., Mahncke,
H., & Merzenich, M. M. (2001). Speech comprehension is
correlated with temporal response patterns recorded from
auditory cortex. Proceedings of the National Academy of
Scienze, U.S.A., 98, 13367–13372. DOI: https://doi.org
/10.1073/pnas.201400998, PMID: 11698688, PMCID:
PMC60877
Alday, P. M. (2019). M/EEG analysis of naturalistic stories: UN
review from speech to language processing. Language,
Cognition and Neuroscience, 34, 457–473. DOI: https://
doi.org/10.1080/23273798.2018.1546882
Alexandrou, UN. M., Saarinen, T., Kujala, J., & Salmelin, R. (2018).
Cortical tracking of global and local variations of speech
rhythm during connected natural speech perception.
Journal of Cognitive Neuroscience, 30, 1704–1719. DOI:
https://doi.org/10.1162/jocn_a_01295, PMID: 29916785
Alexandrou, UN. M., Saarinen, T., Kujala, J., & Salmelin, R. (2020).
Cortical entrainment: What we can learn from studying
naturalistic speech perception. Language, Cognition and
Neuroscience, 35, 681–693. DOI: https://doi.org/10.1080
/23273798.2018.1518534
Alexopoulou, T., Michel, M., Murakami, A., & Meurers, D.
(2017). Task effects on linguistic complexity and accuracy:
A large-scale learner corpus analysis employing natural
language processing techniques. Language Learning, 67,
180–208. DOI: https://doi.org/10.1111/lang.12232
Baltzell, l. S., Srinivasan, R., & Richards, V. M. (2017). IL
effect of prior knowledge and intelligibility on the
cortical entrainment response to speech. Journal of
Neurophysiology, 118, 3144–3151. DOI: https://doi.org
/10.1152/jn.00023.2017, PMID: 28877963, PMCID:
PMC5814715
Bastiaansen, M., Mazaheri, A., & Jensen, O. (2012). Beyond
ERPs: Oscillatory neuronal dynamics. In S. J. Luck & E. S.
Kappenman (Eds.), The Oxford handbook of event-related
potential components (pag. 31–49). New York: Oxford
Stampa universitaria.
Beier et al.
587
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Bastiaansen, M., & Hagoort, P. (2006). Oscillatory neuronal
dynamics during language comprehension. Progress in Brain
Research, 159, 179–196. DOI: https://doi.org/10.1016/S0079
-6123(06)59012-0, PMID: 17071231
Batterink, l. J., & Paller, K. UN. (2017). Online neural monitoring
of statistical learning. Cortex, 90, 31–45. DOI: https://doi.org
/10.1016/j.cortex.2017.02.004, PMID: 28324696, PMCID:
PMC5438777
Beier, E. J., & Ferreira, F. (2018). The temporal prediction of
stress in speech and its relation to musical beat perception.
Frontiers in Psychology, 9, 431. DOI: https://doi.org
/10.3389/fpsyg.2018.00431, PMID: 29666600, PMCID:
PMC5892344
Bentin, S., Kutas, M., & Hillyard, S. UN. (1993).
Electrophysiological evidence for task effects on semantic
priming in auditory word processing. Psychophysiology,
30, 161–169. DOI: https://doi.org/10.1111/j.1469-8986
.1993.tb01729.x, PMID: 8434079
Besle, J., Schevon, C. A., Mehta, UN. D., Lakatos, P., Goodman, R. R.,
McKhann, G. M., et al. (2011). Tuning of the human neocortex
to the temporal dynamics of attended events. Journal of
Neuroscience, 31, 3176–3185. DOI: https://doi.org/10.1523
/JNEUROSCI.4518-10.2011, PMID: 21368029, PMCID:
PMC3081726
Bever, T. G., & Townsend, D. J. (2001). Some sentences on our
consciousness of sentences. In D. J. Townsend & T. G. Bever
(Eds.), Language, brain, and cognitive development: Essays
in honor of Jacques Mehler (pag. 143–155). Cambridge, MA:
CON Premere.
Biau, E., Torralba, M., Fuentemilla, L., de Diego Balaguer, R., &
Soto-Faraco, S. (2015). Speaker’s hand gestures modulate
speech perception through phase resetting of ongoing
neural oscillations. Cortex, 68, 76–85. DOI: https://doi.org
/10.1016/j.cortex.2014.11.018, PMID: 25595613
Bock, J. K. (1986). Syntactic persistence in language production.
Cognitive Psychology, 18, 355–387. DOI: https://doi.org
/10.1016/0010-0285(86)90004-6
Bögels, S., Schriefers, H., Vonk, W., & Chwilla, D. J. (2011).
Pitch accents in context: How listeners process accentuation
in referential communication. Neuropsychologia, 49, 2022–2036.
DOI: https://doi.org/10.1016/j.neuropsychologia.2011.03.032,
PMID: 21458470
Bourguignon, M., De Tiège, X., Op de Beeck, M., Ligot, N.,
Paquier, P., Van Bogaert, P., et al. (2013). The pace of
prosodic phrasing couples the listener’s cortex to the
reader’s voice. Human Brain Mapping, 34, 314–326. DOI:
https://doi.org/10.1002/hbm.21442, PMID: 22392861,
PMCID: PMC6869855
Brennan, J. (2016). Naturalistic sentence comprehension in the
brain. Language and Linguistics Compass, 10, 299–313.
DOI: https://doi.org/10.1111/lnc3.12198
Brothers, T., Swaab, T. Y., & Traxler, M. J. (2017). Goals and
strategies influence lexical prediction during sentence
comprehension. Journal of Memory and Language, 93,
203–216. DOI: https://doi.org/10.1016/j.jml.2016.10.002
Brown, C. M., Hagoort, P., & ter Keurs, M. (1999).
Electrophysiological signatures of visual lexical processing:
Open- and closed-class words. Journal of Cognitive
Neuroscience, 11, 261–281. DOI: https://doi.org/10.1162
/089892999563382, PMID: 10402255
Calderone, D. J., Lakatos, P., Butler, P. D., & Castellanos, F. X.
(2014). Entrainment of neural oscillations as a modifiable
substrate of attention. Trends in Cognitive Sciences, 18,
300–309. DOI: https://doi.org/10.1016/j.tics.2014.02.005,
PMID: 24630166, PMCID: PMC4037370
Christiansen, M. H., & Chater, N. (2015). The language faculty
that wasn’t: A usage-based account of natural language
recursion. Frontiers in Psychology, 6, 1182. DOI: https://doi
.org/10.3389/fpsyg.2015.01182, PMID: 26379567, PMCID:
PMC4550780
Christianson, K., Hollingworth, A., Halliwell, J. F., & Ferreira, F.
(2001). Thematic roles assigned along the garden path linger.
Cognitive Psychology, 42, 368–407. DOI: https://doi.org
/10.1006/cogp.2001.0752, PMID: 11368528
Chwilla, D. J., Brown, C. M., & Hagoort, P. (1995). The N400 as a
function of the level of processing. Psychophysiology, 32,
274–285. DOI: https://doi.org/10.1111/j.1469-8986.1995
.tb02956.x, PMID: 7784536
Clifton, C., Jr., Carlson, K., & Frazier, l. (2002). Informative
prosodic boundaries. Language and Speech, 45, 87–114.
DOI: https://doi.org/10.1177/00238309020450020101,
PMID: 12613557
Crosse, M. J., Di Liberto, G. M., Bednar, A., & Lalor, E. C. (2016).
The multivariate temporal response function (mTRF)
toolbox: A MATLAB toolbox for relating neural signals to
continuous stimuli. Frontiers in Human Neuroscience, 10,
604. DOI: https://doi.org/10.3389/fnhum.2016.00604, PMID:
27965557, PMCID: PMC5127806
Crystal, T. H., & House, UN. S. (1982). Segmental durations in
connected speech signals: Preliminary results. Journal of the
Acoustical Society of America, 72, 705–716. DOI: https://doi
.org/10.1121/1.388251, PMID: 7130529
Cunillera, T., Toro, J. M., Sebastián-Gallés, N., & Rodríguez-
Fornells, UN. (2006). The effects of stress and statistical cues on
continuous speech segmentation: An event-related brain
potential study. Brain Research, 1123, 168–178. DOI:
https://doi.org/10.1016/j.brainres.2006.09.046, PMID:
17064672
Cutler, A., Dahan, D., & van Donselaar, W. (1997). Prosody in
the comprehension of spoken language: A literature review.
Language and Speech, 40, 141–201. DOI: https://doi.org
/10.1177/002383099704000203, PMID: 9509577
Davies, M. (2008). The corpus of contemporary American
English (COCA): 560 million words, 1990–present. Accessed
May 14, 2019: https://corpus.byu.edu/coca/.
Deacon, D., Breton, F., Ritter, W., & Vaughan, H. G., Jr. (1991).
The relationship between N2 and N400: Scalp distribution,
stimulus probability, and task relevance. Psychophysiology,
28, 185–200. DOI: https://doi.org/10.1111/j.1469-8986.1991
.tb00411.x, PMID: 1946885
Di Liberto, G. M., & Lalor, E. C. (2017). Indexing cortical
entrainment to natural speech at the phonemic level:
Methodological considerations for applied research. Hearing
Research, 348, 70–77. DOI: https://doi.org/10.1016/j.heares
.2017.02.015, PMID: 28246030
Ding, N., Chatterjee, M., & Simone, J. Z. (2014). Robust cortical
entrainment to the speech envelope relies on the spectro-
temporal fine structure. Neuroimage, 88, 41–46. DOI:
https://doi.org/10.1016/j.neuroimage.2013.10.054, PMID:
24188816, PMCID: PMC4222995
Ding, N., Melloni, L., Zhang, H., Tian, X., & Poeppel, D. (2016).
Cortical tracking of hierarchical linguistic structures in
connected speech. Nature Neuroscience, 19, 158–164. DOI:
https://doi.org/10.1038/nn.4186, PMID: 26642090, PMCID:
PMC4809195
Ding, N., & Simone, J. Z. (2014). Cortical entrainment to
continuous speech: Functional roles and interpretations.
Frontiers in Human Neuroscience, 8, 311. DOI: https://
doi.org/10.3389/fnhum.2014.00311, PMID: 24904354,
PMCID: PMC4036061
Doelling, K. B., Arnal, l. H., Ghitza, O., & Poeppel, D. (2014).
Acoustic landmarks drive delta–theta oscillations to enable
speech comprehension by facilitating perceptual parsing.
Neuroimage, 85, 761–768. DOI: https://doi.org/10.1016
/j.neuroimage.2013.06.035, PMID: 23791839, PMCID:
PMC3839250
588
Journal of Cognitive Neuroscience
Volume 33, Numero 4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Falk, S., Lanzilotti, C., & Schön, D. (2017). Tuning neural phase
entrainment to speech. Journal of Cognitive Neuroscience,
29, 1378–1389. DOI: https://doi.org/10.1162/jocn_a_01136,
PMID: 28430043
Fedorenko, E., & Blank, IO. UN. (2020). Broca’s area is not a natural
kind. Trends in Cognitive Sciences, 24, 270–284. DOI:
https://doi.org/10.1016/j.tics.2020.01.001, PMID: 32160565,
PMCID: PMC7211504
Ferreira, F. (1993). Creation of prosody during sentence
production. Psychological Review, 100, 233–253. DOI:
https://doi.org/10.1037/0033-295X.100.2.233, PMID: 8483983
Ferreira, F. (2003). The misinterpretation of noncanonical
sentences. Cognitive Psychology, 47, 164–203. DOI: https://
doi.org/10.1016/s0010-0285(03)00005-7, PMID: 12948517
Ferreira, F., & Anes, M. (1994). Why study spoken language?
In M. UN. Gernsbacher (Ed.), Handbook of psycholinguistics
(pag. 33–56). San Diego, CA: Academic Press.
Ferreira, F., & Lowder, M. W. (2016). Prediction, informazione
structure, and good-enough language processing. In B. H.
Ross (Ed.), The psychology of learning and motivation
(Vol. 65, pag. 217–247). New York: Academic Press.
Ferreira, F., & Patson, N. D. (2007). The ‘good enough’
approach to language comprehension. Language and
Linguistics Compass, 1, 71–83. DOI: https://doi.org/10.1111
/j.1749-818X.2007.00007.x
Ferreira, F., & Yang, Z. (2019). The problem of comprehension
in psycholinguistics. Discourse Processes, 56, 485–495. DOI:
https://doi.org/10.1080/0163853X.2019.1591885
Finke, M., & Rogina, IO. (1997). Wide context acoustic modeling
in read vs. spontaneous speech. In 1997 IEEE International
Conference on Acoustics, Speech, and Signal Processing
(Vol. 3, pag. 1743–1746). Los Alamitos, CA: IEEE Computer
Society Press.
Foertsch, J., & Gernsbacher, M. UN. (1994). In search of complete
comprehension: Getting “minimalists” to work. Discourse
Processes, 18, 271–296. DOI: https://doi.org/10.1080
/01638539409544896, PMID: 25520530, PMCID:
PMC4266472
Frazier, l. (1987). Sentence processing: A tutorial review. In M.
Coltheart (Ed.), Attention and performance 12: IL
psychology of reading (pag. 559–586). Hillsdale, NJ: Erlbaum.
Frazier, L., & Fodor, J. D. (1978). The sausage machine: A new
two-stage parsing model. Cognition, 6, 291–325. DOI:
https://doi.org/10.1016/0010-0277(78)90002-1
Frazier, L., & Rayner, K. (1987). Resolution of syntactic category
ambiguities: Eye movements in parsing lexically ambiguous
sentences. Journal of Memory and Language, 26, 505–526.
DOI: https://doi.org/10.1016/0749-596X(87)90137-9
Friederici, UN. D. (2002). Towards a neural basis of auditory
sentence processing. Trends in Cognitive Sciences, 6, 78–84.
DOI: https://doi.org/10.1016/s1364-6613(00)01839-8, PMID:
15866191
Friederici, UN. D. (2011). The brain basis of language processing:
From structure to function. Physiological Reviews, 91,
1357–1392. DOI: https://doi.org/10.1152/physrev.00006.2011,
PMID: 22013214
Friederici, UN. D., & Meyer, M. (2004). The brain knows the
difference: Two types of grammatical violations. Brain
Research, 1000, 72–77. DOI: https://doi.org/10.1016
/j.brainres.2003.10.057, PMID: 15053954
Futrell, R., Gibson, E., & Levy, R. P. (2020). Lossy-context
surprisal: An information-theoretic model of memory effects
in sentence processing. Cognitive Science, 44, e12814.
DOI: https://doi.org/10.1111/cogs.12814, PMID: 32100918,
PMCID: PMC7065005
Getz, H., Ding, N., Newport, E. L., & Poeppel, D. (2018).
Cortical tracking of constituent structure in language
acquisition. Cognition, 181, 135–140. DOI: https://doi.org
/10.1016/j.cognition.2018.08.019, PMID: 30195135, PMCID:
PMC6201233
Ghitza, O., & Greenberg, S. (2009). On the possible role of
brain rhythms in speech perception: Intelligibility of time-
compressed speech with periodic and aperiodic insertions of
silence. Phonetica, 66, 113–126. DOI: https://doi.org/10.1159
/000208934, PMID: 19390234
Gibson, E., Bergen, L., & Piantadosi, S. T. (2013). Rational
integration of noisy evidence and prior semantic expectations
in sentence interpretation. Proceedings of the National
Academy of Sciences, U.S.A., 110, 8051–8056. DOI: https://
doi.org/10.1073/pnas.1216438110, PMID: 23637344,
PMCID: PMC3657782
Giraud, A.-L., & Poeppel, D. (2012). Cortical oscillations and
speech processing: Emerging computational principles and
operations. Nature Neuroscience, 15, 511–517. DOI: https://
doi.org/10.1038/nn.3063, PMID: 22426255, PMCID:
PMC4461038
Gomez-Ramirez, M., Kelly, S. P., Molholm, S., Sehatpour, P.,
Schwartz, T. H., & Foxe, J. J. (2011). Oscillatory sensory
selection mechanisms during intersensory attention
to rhythmic auditory and visual inputs: A human
electrocorticographic investigation. Journal of Neuroscience,
31, 18556–18567. DOI: https://doi.org/10.1523
/JNEUROSCI.2164-11.2011, PMID: 22171054, PMCID:
PMC3298747
Gunter, T. C., & Friederici, UN. D. (1999). Concerning the
automaticity of syntactic processing. Psychophysiology, 36,
126–137. DOI: https://doi.org/10.1017/s004857729997155x,
PMID: 10098388
Gross, J., Hoogenboom, N., Thut, G., Schyns, P., Panzeri, S.,
Belin, P., et al. (2013). Speech rhythms and multiplexed
oscillatory sensory coding in the human brain. PLoS Biology,
11, e1001752. DOI: https://doi.org/10.1371/journal.pbio
.1001752, PMID: 24391472, PMCID: PMC3876971
Hale, J., Dyer, C., Kuncoro, A., & Brennan, J. (2018). Finding
syntax in human encephalography with beam search. In
Proceedings of the 56th Annual Meeting of the Association
for Computational Linguistics ( Volume 1: Documenti lunghi)
(pag. 2727–2736). Melbourne, Australia: Associazione per
Linguistica computazionale. DOI: https://doi.org/10.18653/v1
/P18-1254
Hamilton, l. S., & Huth, UN. G. (2020). The revolution will not be
controlled: Natural stimuli in speech neuroscience.
Language, Cognition and Neuroscience, 35, 573–582. DOI:
https://doi.org/10.1080/23273798.2018.1499946, PMID:
32656294, PMCID: PMC7324135
Hauk, O., & Pulvermüller, F. (2004). Effects of word length and
frequency on the human event-related potential. Clinical
Neurophysiology, 115, 1090–1103. DOI: https://doi.org
/10.1016/j.clinph.2003.12.020, PMID: 15066535
Henderson, J. M., Choi, W., Lowder, M. W., & Ferreira, F.
(2016). Language structure in the brain: A fixation-related
fMRI study of syntactic surprisal in reading. Neuroimage, 132,
293–300. DOI: https://doi.org/10.1016/j.neuroimage.2016.02
.050, PMID: 26908322
Hirose, K., & Kawanami, H. (2002). Temporal rate change of
dialogue speech in prosodic units as compared to read
speech. Speech Communication, 36, 97–111. DOI: https://
doi.org/10.1016/S0167-6393(01)00028-0
Holcomb, P. J., Grainger, J., & O’Rourke, T. (2002). An
electrophysiological study of the effects of orthographic
neighborhood size on printed word perception. Journal of
Cognitive Neuroscience, 14, 938–950. DOI: https://doi.org
/10.1162/089892902760191153, PMID: 12191460
Howard, M. F., & Poeppel, D. (2010). Discrimination of speech
stimuli based on neuronal response phase patterns
depends on acoustics but not comprehension. Journal of
Beier et al.
589
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neurophysiology, 104, 2500–2511. DOI: https://doi.org
/10.1152/jn.00251.2010, PMID: 20484530, PMCID:
PMC2997028
Huang, Y., & Ferreira, F. (2020). The application of signal
detection theory to acceptability judgments. Frontiers in
Psychology, 11, 73. DOI: https://doi.org/10.3389/fpsyg.2020
.00073, PMID: 32082223, PMCID: PMC7005104
Huettig, F., & Janse, E. (2016). Individual differences in working
memory and processing speed predict anticipatory spoken
language processing in the visual world. Language,
Cognition and Neuroscience, 31, 80–93. DOI: https://doi
.org/10.1080/23273798.2015.1047459
Jewett, D. L., & Williston, J. S. (1971). Auditory-evoked far fields
averaged from the scalp of humans. Brain, 94, 681–696.
DOI: https://doi.org/10.1093/brain/94.4.681, PMID: 5132966
Johnson-Laird, P. N. (1983). Mental models: Towards a
cognitive science of language, inference, E
consciousness. Cambridge, MA: Stampa dell'Università di Harvard.
Keitel, A., Gross, J., & Kayser, C. (2018). Perceptually relevant
speech tracking in auditory and motor cortex reflects distinct
linguistic features. PLoS Biology, 16, e2004473.
DOI: https://doi.org/10.1371/journal.pbio.2004473, PMID:
29529019, PMCID: PMC5864086
Keitel, A., Ince, R. UN. A., Gross, J., & Kayser, C. (2017). Auditory
cortical delta-entrainment interacts with oscillatory power in
multiple fronto-parietal networks. Neuroimage, 147, 32–42.
DOI: https://doi.org/10.1016/j.neuroimage.2016.11.062,
PMID: 27903440, PMCID: PMC5315055
Kerlin, J. R., Shahin, UN. J., & Mugnaio, l. M. (2010). Attentional gain
control of ongoing cortical speech representations in a
“cocktail party.” Journal of Neuroscience, 30, 620–628. DOI:
https://doi.org/10.1523/JNEUROSCI.3631-09.2010, PMID:
20071526, PMCID: PMC2832933
Kim, UN. E., Oines, L., & Miyake, UN. (2018). Individual differences
in verbal working memory underlie a tradeoff between
semantic and structural processing difficulty during language
comprehension: An ERP investigation. Journal of
Experimental Psychology: Apprendimento, Memory, E
Cognition, 44, 406-420. DOI: https://doi.org/10.1037
/xlm0000457, PMID: 28933902
King, J. W., & Kutas, M. (1998). Neural plasticity in the dynamics
of human visual word recognition. Neuroscience Letters, 244,
61–64. DOI: https://doi.org/10.1016/s0304-3940(98)00140-2,
PMID: 9572585
Kösem, A., Bosker, H. R., Takashima, A., Meyer, A., Jensen,
O., & Hagoort, P. (2018). Neural entrainment determines
the words we hear. Current Biology, 28, 2867–2875. DOI:
https://doi.org/10.1016/j.cub.2018.07.023, PMID:
30197083
Kösem, A., & van Wassenhove, V. (2017). Distinct contributions
of low- and high-frequency neural oscillations to speech
comprehension. Language, Cognition and Neuroscience,
32, 536–544. DOI: https://doi.org/10.1080/23273798.2016
.1238495
Kotz, S. A., & Schwartze, M. (2010). Cortical speech processing
unplugged: A timely subcortico-cortical framework. Trends
in Cognitive Sciences, 14, 392–399. DOI: https://doi.org
/10.1016/j.tics.2010.06.005, PMID: 20655802
Kounios, J., & Holcomb, P. J. (1994). Concreteness effects in
semantic processing: ERP evidence supporting dual-coding
theory. Journal of Experimental Psychology: Apprendimento,
Memory, and Cognition, 20, 804–823. DOI: https://doi.org
/10.1037/0278-7393.20.4.804, PMID: 8064248
Kuperberg, G. R., & Jaeger, T. F. (2016). What do we mean by
prediction in language comprehension? Language, Cognition
and Neuroscience, 31, 32–59. DOI: https://doi.org/10.1080
/23273798.2015.1102299, PMID: 27135040, PMCID:
PMC4850025
Kutas, M., & Federmeier, K. D. (2011). Thirty years and
counting: Finding meaning in the N400 component of the
event-related brain potential (ERP). Annual Review of
Psychology, 62, 621–647. DOI: https://doi.org/10.1146
/annurev.psych.093008.131123, PMID: 20809790, PMCID:
PMC4052444
Kutas, M., Van Petten, C. K., & Kluender, R. (2006).
Psycholinguistics electrified II (1994–2005). In M. J. Traxler &
M. UN. Gernsbacher (Eds.), Handbook of psycholinguistics
(pag. 659–724). New York: Academic Press. DOI: https://doi
.org/10.1016/B978-012369374-7/50018-3
Lakatos, P., Karmos, G., Mehta, UN. D., Ulbert, I., & Schroeder,
C. E. (2008). Entrainment of neuronal oscillations as a
mechanism of attentional selection. Scienza, 320, 110–113.
DOI: https://doi.org/10.1126/science.1154735, PMID:
18388295
Lakatos, P., Shah, UN. S., Knuth, K. H., Ulbert, I., Karmos, G., &
Schroeder, C. E. (2005). An oscillatory hierarchy controlling
neuronal excitability and stimulus processing in the auditory
cortex. Journal of Neurophysiology, 94, 1904–1911. DOI:
https://doi.org/10.1152/jn.00263.2005, PMID: 15901760
Large, E. W., & Jones, M. R. (1999). The dynamics of attending:
How people track time-varying events. Psychological Review,
106, 119–159. DOI: https://doi.org/10.1037/0033-295X.106
.1.119
Large, E. W., & Kolen, J. F. (1994). Resonance and the
perception of musical meter. Connection Science, 6,
177–208. DOI: https://doi.org/10.1080/09540099408915723
Ljung, L., Chen, T., & Mu, B. (2020). A shift in paradigm for
system identification. International Journal of Control, 93,
173–180. DOI: https://doi.org/10.1080/00207179.2019
.1578407
Luo, H., Liu, Z., & Poeppel, D. (2010). Auditory cortex tracks
both auditory and visual stimulus dynamics using low-
frequency neuronal phase modulation. PLoS Biology, 8,
e1000445. DOI: https://doi.org/10.1371/journal.
pbio.1000445, PMID: 20711473, PMCID: PMC2919416
Luo, H., & Poeppel, D. (2007). Phase patterns of neuronal
responses reliably discriminate speech in human auditory
cortex. Neuron, 54, 1001–1010. DOI: https://doi.org/10.1016
/j.neuron.2007.06.004, PMID: 17582338, PMCID:
PMC2703451
MacDonald, M. C. (1993). The interaction of lexical and
syntactic ambiguity. Journal of Memory and Language, 32,
692–715. DOI: https://doi.org/10.1006/jmla.1993.1035
MacDonald, M. C., Pearlmutter, N. J., & Seidenberg, M. S.
(1994). The lexical nature of syntactic ambiguity resolution.
Psychological Review, 101, 676–703. DOI: https://doi.org
/10.1037/0033-295x.101.4.676, PMID: 7984711
Martin, UN. E., & Doumas, l. UN. UN. (2017). A mechanism for
the cortical computation of hierarchical linguistic structure.
PLoS Biology, 15, e2000663. DOI: https://doi.org/10
.1371/journal.pbio.2000663, PMID: 28253256, PMCID:
PMC5333798
Matchin, W., & Hickok, G. (2020). The cortical organization
of syntax. Cerebral Cortex, 30, 1481–1498. DOI: https://
doi.org/10.1093/cercor/bhz180, PMID: 31670779, PMCID:
PMC7132936
Meyer, L.. (2018). The neural oscillations of speech processing
and language comprehension: State of the art and emerging
mechanisms. European Journal of Neuroscience, 48,
2609–2621. DOI: https://doi.org/10.1111/ejn.13748, PMID:
29055058
Meyer, L., & Gumbert, M. (2018). Synchronization of
electrophysiological responses with speech benefits syntactic
information processing. Journal of Cognitive Neuroscience,
30, 1066–1074. DOI: https://doi.org/10.1162/jocn_a_01236,
PMID: 29324074
590
Journal of Cognitive Neuroscience
Volume 33, Numero 4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Meyer, L., Henry, M. J., Gaston, P., Schmuck, N., & Friederici,
UN. D. (2017). Linguistic bias modulates interpretation of
speech via neural delta-band oscillations. Cerebral Cortex,
27, 4293–4302. DOI: https://doi.org/10.1093/cercor/bhw228,
PMID: 27566979
Meyer, L., Sun, Y., & Martin, UN. E. (2020). Synchronous, but not
entrained: Exogenous and endogenous cortical rhythms of
speech and language processing. Language, Cognition and
Neuroscience, 35, 1089–1099. DOI: https://doi.org/10.1080
/23273798.2019.1693050
Millman, R. E., Johnson, S. R., & Prendergast, G. (2015). IL
role of phase-locking to the temporal envelope of speech in
auditory perception and speech intelligibility. Journal of
Cognitive Neuroscience, 27, 533–545. DOI: https://doi.org
/10.1162/jocn_a_00719, PMID: 25244119
Morillon, B., & Schroeder, C. E. (2015). Neuronal oscillations as
a mechanistic substrate of auditory temporal prediction.
Annals of the New York Academy of Sciences, 1337, 26–31.
DOI: https://doi.org/10.1111/nyas.12629, PMID: 25773613,
PMCID: PMC4363099
Moseley, R. L., & Pulvermüller, F. (2014). Nouns, verbs, objects,
actions, and abstractions: Local fMRI activity indexes
semantics, not lexical categories. Brain and Language, 132,
28–42. DOI: https://doi.org/10.1016/j.bandl.2014.03.001,
PMID: 24727103, PMCID: PMC4029073
Myers, J. (2009). Syntactic judgment experiments. Language
and Linguistics Compass, 3, 406–423. DOI: https://doi.org
/10.1111/j.1749-818X.2008.00113.x
Nakajima, S., & Allen, J. F. (1993). A study on prosody and
discourse structure in cooperative dialogues. Phonetica,
50, 197–210. DOI: https://doi.org/10.1159/000261940
Nobre, UN. C., & van Ede, F. (2018). Anticipated moments: Temporal
structure in attention. Nature Reviews Neuroscience, 19, 34–48.
DOI: https://doi.org/10.1038/nrn.2017.141, PMID: 29213134
Nolan, F., & Jeon, H.-S. (2014). Speech rhythm: A metaphor?
Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences, 369, 20130396. DOI: https://
doi.org/10.1098/rstb.2013.0396, PMID: 25385774, PMCID:
PMC4240963
Nora, A., Faisal, A., Seol, J., Renvall, H., Formisano, E., &
Salmelin, R. (2020). Dynamic time-locking mechanism in
the cortical representation of spoken words. eNeuro, 7,
ENEURO.0475-19.2020. DOI: https://doi.org/10.1523
/ENEURO.0475-19.2020, PMID: 32513662, PMCID:
PMC7470935
Nozaradan, S., Peretz, I., Missal, M., & Mouraux, UN. (2011).
Tagging the neuronal entrainment to beat and meter.
Journal of Neuroscience, 31, 10234–10240. DOI: https://doi
.org/10.1523/JNEUROSCI.0411-11.2011, PMID: 21753000,
PMCID: PMC6623069
Obleser, J., & Kotz, S. UN. (2011). Multiple brain signatures of
integration in the comprehension of degraded speech.
Neuroimage, 55, 713–723. DOI: https://doi.org/10.1016
/j.neuroimage.2010.12.020, PMID: 21172443
Obleser, J., & Kayser, C. (2019). Neural entrainment and
attentional selection in the listening brain. Trends in
Cognitive Sciences, 23, 913–926. DOI: https://doi.org
/10.1016/j.tics.2019.08.004, PMID: 31606386
Payton, K. L., Uchanski, R. M., & Braida, l. D. (1994).
Intelligibility of conversational and clear speech in noise
and reverberation for listeners with normal and impaired
hearing. Journal of the Acoustical Society of America, 95,
1581–1592. DOI: https://doi.org/10.1121/1.408545, PMID:
8176061
Peelle, J. E., & Davis, M. H. (2012). Neural oscillations carry
speech rhythm through to comprehension. Frontiers
in Psychology, 3, 320. DOI: https://doi.org/10.3389/fpsyg
.2012.00320, PMID: 22973251, PMCID: PMC3434440
Peelle, J. E., Gross, J., & Davis, M. H. (2013). Phase-locked
responses to speech in human auditory cortex are enhanced
during comprehension. Cerebral Cortex, 23, 1378–1387.
DOI: https://doi.org/10.1093/cercor/bhs118, PMID:
22610394, PMCID: PMC3643716
Peelle, J. E., Mugnaio, R. L., Rogers, C. S., Spehar, B., Sommers,
M. S., & Van Engen, K. J. (2020). Completion norms for
3085 English sentence contexts. Behavior Research
Methods, 52, 1795–1799. DOI: https://doi.org/10.3758
/s13428-020-01351-1, PMID: 31993960, PMCID:
PMC7406521
Picheny, M. A., Durlach, N. I., & Braida, l. D. (1985). Speaking
clearly for the hard of hearing I: Intelligibility differences
between clear and conversational speech. Journal of Speech,
Language, and Hearing Research, 28, 96–103. DOI: https://
doi.org/10.1044/jshr.2801.96, PMID: 3982003
Pickering, M. J., & Ferreira, V. S. (2008). Structural priming: UN
critical review. Psychological Bulletin, 134, 427–459. DOI:
https://doi.org/10.1037/0033-2909.134.3.427, PMID:
18444704, PMCID: PMC2657366
Price, C. J. (2012). A review and synthesis of the first 20 years
of PET and fMRI studies of heard speech, spoken language
and reading. Neuroimage, 62, 816–847. DOI: https://doi.org
/10.1016/j.neuroimage.2012.04.062, PMID: 22584224,
PMCID: PMC3398395
Prystauka, Y., & Lewis, UN. G. (2019). The power of neural
oscillations to inform sentence comprehension: A linguistic
perspective. Language and Linguistics Compass, 13,
e12347. DOI: https://doi.org/10.1111/lnc3.12347, PMID:
33042211, PMCID: PMC7546279
Rimmele, J. M., Golumbic, E. Z., Schröger, E., & Poeppel, D.
(2015). The effects of selective attention and speech
acoustics on neural speech-tracking in a multi-talker scene.
Cortex, 68, 144–154. DOI: https://doi.org/10.1016/j.cortex
.2014.12.014, PMID: 25650107, PMCID: PMC4475476
Rimmele, J. M., Morillon, B., Poeppel, D., & Arnal, l. H. (2018).
Proactive sensing of periodic and aperiodic auditory patterns.
Trends in Cognitive Sciences, 22, 870–882. DOI: https://doi
.org/10.1016/j.tics.2018.08.003, PMID: 30266147
Rodd, J. M., Vitello, S., Woollams, UN. M., & Adank, P. (2015).
Localising semantic and syntactic processing in spoken and
written language comprehension: An activation likelihood
estimation meta-analysis. Brain and Language, 141, 89–102.
DOI: https://doi.org/10.1016/j.bandl.2014.11.012, PMID:
25576690
Russo, UN. G., De Martino, M., Mancuso, A., Iaconetta, G.,
Manara, R., Elia, A., et al. (2020). Semantics-weighted lexical
surprisal modeling of naturalistic functional MRI time-series
during spoken narrative listening. Neuroimage, 222,
117281. DOI: https://doi.org/10.1016/j.neuroimage.2020
.117281, PMID: 32828929
Salverda, UN. P., Brown, M., & Tanenhaus, M. K. (2011). UN
goal-based perspective on eye movements in visual world
studies. Acta Psychologica, 137, 172–180. DOI: https://doi
.org/10.1016/j.actpsy.2010.09.010, PMID: 21067708, PMCID:
PMC3109199
Sanders, l. D., & Neville, H. J. (2003). An ERP study of
continuous speech processing: IO. Segmentation, semantics,
and syntax in native speakers. Cognitive Brain Research, 15,
228–240. DOI: https://doi.org/10.1016/S0926-6410(02)00195-7,
PMID: 12527097
Sanford, UN. J., & Sturt, P. (2002). Depth of processing in
language comprehension: Not noticing the evidence. Trends
in Cognitive Sciences, 6, 382–386. DOI: https://doi.org
/10.1016/S1364-6613(02)01958-7, PMID: 12200180
Sassenhagen, J. (2019). How to analyse electrophysiological
responses to naturalistic language with time-resolved
multiple regression. Language, Cognition and
Beier et al.
591
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Neuroscience, 34, 474–490. DOI: https://doi.org/10.1080
/23273798.2018.1502458
Schacht, A., Sommer, W., Shmuilovich, O., Martíenz, P. C., &
Martín-Loeches, M. (2014). Differential task effects on N400
and P600 elicited by semantic and syntactic violations. PLoS
One, 9, e91226. DOI: https://doi.org/10.1371/journal.pone
.0091226, PMID: 24614675, PMCID: PMC3948820
Schmidt-Kassow, M., & Kotz, S. UN. (2009UN). Event-related brain
potentials suggest a late interaction of meter and syntax in
the P600. Journal of Cognitive Neuroscience, 21, 1693–1708.
DOI: https://doi.org/10.1162/jocn.2008.21153, PMID:
18855546
Schmidt-Kassow, M., & Kotz, S. UN. (2009B). Attention and
perceptual regularity in speech. NeuroReport, 20, 1643–1647.
DOI: https://doi.org/10.1097/ WNR.0b013e328333b0c6,
PMID: 19907350
Schwartze, M., & Kotz, S. UN. (2013). A dual-pathway neural
architecture for specific temporal prediction. Neuroscience &
Biobehavioral Reviews, 37, 2587–2596. DOI: https://doi.org
/10.1016/j.neubiorev.2013.08.005, PMID: 23994272
Shattuck-Hufnagel, S., & Turk, UN. E. (1996). A prosody tutorial
for investigators of auditory sentence processing. Journal of
Psycholinguistic Research, 25, 193–247. DOI: https://doi.org
/10.1007/BF01708572, PMID: 8667297
Song, J., & Iverson, P. (2018). Listening effort during speech
perception enhances auditory and lexical processing for non-
native listeners and accents. Cognition, 179, 163–170. DOI:
https://doi.org/10.1016/j.cognition.2018.06.001, PMID:
29957515
Staub, A., Rayner, K., Pollatsek, A., Hyönä, J., & Majewski, H.
(2007). The time course of plausibility effects on eye
movements in reading: Evidence from noun–noun
compounds. Journal of Experimental Psychology: Apprendimento,
Memory, and Cognition, 33, 1162–1169. DOI: https://doi.org
/10.1037/0278-7393.33.6.1162, PMID: 17983320
Stefanics, G., Hangya, B., Hernádi, I., Winkler, I., Lakatos, P., &
Ulbert, IO. (2010). Phase entrainment of human delta
oscillations can mediate the effects of expectation on
reaction speed. Journal of Neuroscience, 30, 13578–13585.
DOI: https://doi.org/10.1523/JNEUROSCI.0703-10.2010,
PMID: 20943899, PMCID: PMC4427664
Strijkers, K., Costa, A., & Thierry, G. (2010). Tracking lexical
access in speech production: Electrophysiological correlates
of word frequency and cognate effects. Cerebral Cortex, 20,
912–928. DOI: https://doi.org/10.1093/cercor/bhp153,
PMID: 19679542
Swaab, T. Y., Ledoux, K., Camblin, C. C., & Boudewyn, M. UN.
(2012). Language-related ERP components. In S. J. Luck &
E. S. Kappenman (Eds.), Oxford handbook of event-related
potential components (pag. 397–440). Oxford: Oxford
Stampa universitaria. DOI: https://doi.org/10.1093/oxfordhb
/9780195374148.013.0197
Swets, B., Desmet, T., Hambrick, D. Z., & Ferreira, F. (2007).
The role of working memory in syntactic ambiguity
resolution: A psychometric approach. Journal of
Experimental Psychology: General, 136, 64–81. DOI: https://
doi.org/10.1037/0096-3445.136.1.64, PMID: 17324085
Taylor, W. l. (1953). “Cloze procedure”: A new tool for
measuring readability. Giornalismo & Comunicazione di massa
Trimestrale, 30, 415–433. DOI: https://doi.org/10.1177
/107769905303000401
Teinonen, T., & Huotilainen, M. (2012). Implicit segmentation
of a stream of syllables based on transitional probabilities: An
MEG study. Journal of Psycholinguistic Research, 41, 71–82.
DOI: https://doi.org/10.1007/s10936-011-9182-2, PMID:
21993901
Tooley, K. M., Traxler, M. J., & Swaab, T. Y. (2009).
Electrophysiological and behavioral evidence of syntactic
priming in sentence comprehension. Journal of
Experimental Psychology: Apprendimento, Memory, E
Cognition, 35, 19–45. DOI: https://doi.org/10.1037/a0013984,
PMID: 19210079
Traxler, M. J. (2014). Trends in syntactic parsing: Anticipation,
Bayesian estimation, and good-enough parsing. Trends in
Cognitive Sciences, 18, 605–611. DOI: https://doi.org/10.1016
/j.tics.2014.08.001, PMID: 25200381, PMCID: PMC6814003
Trueswell, J. C., & Tanenhaus, M. K. (1994). Toward a lexicalist
framework of constraint-based syntactic ambiguity resolution.
In C. Clifton Jr., l. Frazier, & K. Rayner (Eds.), Perspectives on
sentence processing (pag. 155–179). Hillsdale, NJ: Erlbaum.
Tyler, l. K. (Ed.). (2020). Meyer forum [special issue].
Language, Cognition and Neuroscience, 35, 1089–1222.
DOI: https://doi.org/10.1080/23273798.2019.1693050
Uchanski, R. M., Choi, S. S., Braida, l. D., Reed, C. M., &
Durlach, N. IO. (1996). Speaking clearly for the hard of
hearing IV: Further studies of the role of speaking rate.
Journal of Speech, Language, and Hearing Research, 39,
494–509. DOI: https://doi.org/10.1044/jshr.3903.494,
PMID: 8783129
van Berkum, J. J. UN. (2004). Sentence comprehension in a wider
discourse: Can we use ERPs to keep track of things? In M.
Carreiras & C. Clifton Jr. (Eds.), The on-line study of sentence
comprehension: Eyetracking, ERPs and beyond (pag. 229–270).
New York: Psychology Press.
van Berkum, J. J. A., Brown, C. M., Zwitserlood, P., Kooijman, V.,
& Hagoort, P. (2005). Anticipating upcoming words in
discourse: Evidence from ERPs and reading times. Journal
of Experimental Psychology: Apprendimento, Memory, E
Cognition, 31, 443–467. DOI: https://doi.org/10.1037/0278
-7393.31.3.443, PMID: 15910130
Van Petten, C., & Kutas, M. (1990). Interactions between
sentence context and word frequency in event-related brain
potentials. Memory & Cognition, 18, 380–393. DOI: https://
doi.org/10.3758/bf03197127, PMID: 2381317
Wang, L., Bastiaansen, M., Yang, Y., & Hagoort, P. (2011). IL
influence of information structure on the depth of semantic
processing: How focus and pitch accent determine the size of
the N400 effect. Neuropsychologia, 49, 813–820. DOI:
https://doi.org/10.1016/j.neuropsychologia.2010.12.035,
PMID: 21195102
Wang, L., Bastiaansen, M., Yang, Y., & Hagoort, P. (2012).
Information structure influences depth of syntactic processing:
Event-related potential evidence for the Chomsky illusion. PLoS
One, 7, e47917. DOI: https://doi.org/10.1371/journal.pone
.0047917, PMID: 23110131, PMCID: PMC3480462
Wang, L., Hagoort, P., & Jensen, O. (2018). Language prediction
is reflected by coupling between frontal gamma and posterior
alpha oscillations. Journal of Cognitive Neuroscience, 30,
432–447. DOI: https://doi.org/10.1162/jocn_a_01190, PMID:
28949823
Weissbart, H., Kandylaki, K. D., & Reichenbach, T. (2020).
Cortical tracking of surprisal during continuous speech
comprehension. Journal of Cognitive Neuroscience, 32,
155–166. DOI: https://doi.org/10.1162/jocn_a_01467, PMID:
31479349
Willems, R. M., Frank, S. L., Nijhof, UN. D., Hagoort, P., & van den
Bosch, UN. (2016). Prediction during natural language
comprehension. Cerebral Cortex, 26, 2506–2516. DOI:
https://doi.org/10.1093/cercor/bhv075, PMID: 25903464
Winsler, K., Midgley, K. J., Grainger, J., & Holcomb, P. J. (2018).
An elecrophysiological megastudy of spoken word
recognition. Language, Cognition and Neuroscience, 33,
1063–1082. DOI: https://doi.org/10.1080/23273798.2018
.1455985
Zaccarella, E., Schell, M., & Friederici, UN. D. (2017). Reviewing
the functional basis of the syntactic Merge mechanism for
592
Journal of Cognitive Neuroscience
Volume 33, Numero 4
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
lingua: A coordinate-based activation likelihood estimation
meta-analysis. Neuroscience & Biobehavioral Reviews, 80,
646–656. DOI: https://doi.org/10.1016/j.neubiorev.2017.06.011,
PMID: 28743620
Zoefel, B., Archer-Boyd, A., & Davis, M. H. (2018). Phase
entrainment of brain oscillations causally modulates neural
responses to intelligible speech. Current Biology, 28,
401–408. DOI: https://doi.org/10.1016/j.cub.2017.11.071,
PMID: 29358073, PMCID: PMC5807089
Zoefel, B., & VanRullen, R. (2015). The role of high-level
processes for oscillatory phase entrainment to speech sound.
Frontiers in Human Neuroscience, 9, 651. DOI: https://doi.org
/10.3389/fnhum.2015.00651, PMID: 26696863, PMCID:
PMC4667100
Zoefel, B., & VanRullen, R. (2016). EEG oscillations entrain their
phase to high-level features of speech sound. Neuroimage,
124, 16–23. DOI: https://doi.org/10.1016/j.neuroimage
.2015.08.054, PMID: 26341026
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
/
3
3
4
5
7
4
2
0
3
2
7
3
8
/
j
o
C
N
_
UN
_
0
1
6
7
6
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
7
S
e
P
e
M
B
e
R
2
0
2
3
Beier et al.
593