Brain Responses to Biological Relevance
Christine M. Tipper1, Todd C. Handy1, Barry Giesbrecht2,
and Alan Kingstone1
Astratto
& This study examines whether orienting attention to biolog-
ically based social cues engages neural mechanisms distinct
from those engaged by orienting to nonbiologically based non-
social cues. Participants viewed a perceptually ambiguous stim-
ulus presented centrally while performing a target detection
task. By having participants alternate between viewing this stim-
ulus as an eye in profile or an arrowhead, we were able to
directly compare the neural mechanisms of attentional ori-
enting to social and nonsocial cues while holding the physical
stimulus constant. The functional magnetic resonance imaging
results indicated that attentional orienting to both eye gaze
and arrow cues engaged extensive dorsal and ventral fronto-
parietal networks. Eye gaze cues, Tuttavia, more vigorously
engaged two regions in the ventral frontal cortex associated
with attentional reorienting to salient or meaningful stimuli,
as well as lateral occipital regions. An event-related potential
study demonstrated that this enhanced occipital response was
attributable to a higher-amplitude sensory gain effect for tar-
gets appearing at locations cued by eye gaze than for those
cued by an arrowhead. These results endorse the hypothesis
that differences in attention to social and nonsocial cues are
quantitative rather than qualitative, running counter to current
models that assume enhanced processing for social stimuli
reflects the involvement of a unique network of brain regions.
An intriguing implication of the present study is the possibility
that our ability to orient volitionally and reflexively to socially
irrelevant stimuli, including arrowheads, may have arisen as a
useful by-product of a system that developed first, and fore-
most, to promote social orienting to stimuli that are biolog-
ically relevant. &
INTRODUCTION
Most of us have had the experience of trying to carry on
a conversation with someone who looks away distract-
edly. When this happens, it is often difficult to continue
the conversation because your attention is diverted to
whatever your conversation partner is looking at. Far
from being anecdotal, this phenomenon—that one’s vi-
sual attention can be directed reflexively to locations
indicated by another’s eye gaze—is well documented
(Frischen & Tipper, 2004; Langton & Bruce, 1999;
Friesen & Kingstone, 1998). These studies have given
rise to the theoretical claim that eye gaze is a particular-
ly powerful,
‘‘special’’ cue for visuospatial attention.
Although there is evidence that contextual information
such as head orientation and body movements modu-
lates attention to gaze direction (Langton, Watt, & Bruce,
2000), the importance of the eyes themselves as a social
cue is hard to deny.
The special status of eye gaze as a cue for spatial at-
tention may owe, at least in part, to specialized neural
systems for processing eye gaze information. The supe-
rior temporal sulcus (STS) has been implicated in nu-
merous studies as a region specialized for processing eye
1University of British Columbia, 2University of California at Santa
Barbara
gaze (Kingstone, Tipper, Ristic, & Ngan, 2004; Allison,
Puce, & McCarthy, 2000; Hoffman & Haxby, 2000; Perrett
et al., 1985). More recent neuroimaging work suggests
that specific regions within the STS may be specialized not
only for the processing of eye gaze information but also
for the processing of several forms of biological motion,
including mouth, eye, and hand movements (Pelphrey
& Morris, 2006; Pelphrey, Morris, Michelich, Allison, &
McCarthy, 2005). Although there is ongoing debate re-
garding the specificity of STS functionality, there is grow-
ing consensus that the STS plays an integral role in the
perception of social cues in particular, rather than sim-
ply any directional stimuli (Hooker et al., 2003). Never-
theless, although the STS is an important player in the
perceptual processing of eye gaze, whether there are
neural systems specialized for orienting attention to this
biologically based social cue remains an open question.
The use of biologically based cues in shifting visual
Attenzione, what we will call social attention, provides in-
formation regarding one’s surroundings even in the ab-
sence of direct visual perception. The social extension
of one’s own attentional and perceptual reach would
have constituted a beneficial cognitive adaptation in an
ancestral environment rife with intergroup conflicts and
predators with far greater strength and speed. This evo-
lutionary argument for specialized mechanisms mediat-
ing social attention is supported by the finding that
D 2008 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 20:5, pag. 879–891
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
.
/
.
T
.
F
o
N
1
8
M
UN
sì
2
0
2
1
monkeys and humans may share a homologous neural
mechanism for social attention (Deaner & Platt, 2003).
The present study examines whether specialized neural
mechanisms facilitate the orienting of attention to social
cues in humans. Specifically, we asked whether visuo-
spatial attentional orienting to directional biological cues
(occhi) engages neural mechanisms distinct from those
engaged by orienting to directional nonbiological cues
(arrows). We will refer to the former as social cues, E
the latter as nonsocial cues.
One possibility is that orienting to social cues does
utilize specialized neural modules. In patients with visuo-
spatial neglect, Per esempio, gaze direction cues can in-
duce shifts in attention to regions of space to which these
patients cannot otherwise attend (Vuilleumier, 2002). In
aggiunta, although both hemispheres in a split-brain pa-
tient were recruited in orienting attention to arrow stim-
uli, only the predominant face-processing hemisphere
was engaged while orienting attention to eye gaze stimuli
(Kingstone, Friesen, & Gazzaniga, 2000). Consistent with
these patient studies, one recent functional magnetic
resonance imaging (fMRI) study reported that although
arrow cues engaged areas of the frontal and parietal
cortex typically involved in volitional orienting, gaze cues
did not (Hietanen, Nummenmaa, Nyman, Parkkola, &
Ha¨ma¨la¨inen, 2006). These results suggest that attentional
orienting to eye gaze may utilize neural mechanisms
distinct from those needed for orienting in response to
nonsocial stimuli, such as arrows. A second possibility,
Tuttavia, is that the same neural mechanisms subserve
attentional orienting to any meaningful or symbolic stim-
ulus. This possibility is consistent with behavioral studies
demonstrating equivalent orienting to centrally pre-
sented eyes and arrows (Quadflieg, Mason, & Macrae,
2004; Ristic, Friesen, & Kingstone, 2002; Tipples, 2002).
To test between these competing hypotheses, we asked
participants to view a perceptually ambiguous object that
could be interpreted either as an eye in profile (Figura 1,
looking to the right), or an arrowhead (Figura 1, pointing
to the left). By instructing participants to alternate be-
tween viewing this object as an eye or an arrow while they
underwent fMRI scanning, we were able to compare the
neural mechanisms of attentional orienting to social and
nonsocial cues while holding the physical cue stimulus
constant. If there are specialized neural modules for ori-
enting to social cues such as eye gaze, then we would
expect to find greater blood oxygenation level-dependent
(BOLD) activity in these regions when viewing the ambig-
uous object as an eye relative to when viewing that same
stimulus as an arrow. If, on the other hand, both eye gaze
and arrow cues utilize the same orienting network, we
would expect both percepts to equally engage a fronto-
parietal orienting network (Corbetta & Shulman, 2002).
EXPERIMENT 1
While undergoing fMRI scanning, participants viewed the
centrally presented object as either an eye or an arrow.
Periodically, we instructed participants to switch how they
perceived the object. The results of a pilot study ensured
that participants were able to maintain each percept with
equal ease, and switch percepts effectively when to do so
(and see Experiment 3 for direct behavioral evidence
supporting these claims). Participants responded with a
button press as quickly as possible when they saw an
asterisk appear in the left or right periphery either 100 O
600 msec following the onset of the cue. Importantly, Essi
could not predict the location of the target based on the
central object, a fact about which they were informed. On
half of the trials, the target appeared at the location to
which the eye gazed or the arrow pointed (cued). On the
other half of trials, the target appeared at the opposite
location (uncued). Because participants could not predict
the location of the target based on the central object,
there was no explicit reason for participants to attend to
one location or the other in response to the central cue
stimulus. Shorter response latencies to targets appearing
at the cued relative to the uncued location could therefore
be interpreted as indicating a reflexive, or automatic, shift
in visuospatial attention in the direction cued by the
central object (Posner, 1980).
Methods
Participants
Eight neurologically healthy, right-handed participants
(mean age = 23.75 years, 3 women) from the University
of British Columbia took part in the study with written
consent. All participants had normal or corrected-to-normal
vision. Experimental procedures were approved by the Uni-
versity of British Columbia Clinical Research Ethics Board.
Participants were remunerated with structural images of
their brains on CD.
Figura 1. Ambiguous cue stimulus. Participants were instructed to
perceive this stimulus as either an arrow (per esempio., pointing the left) O
as an eye in profile (per esempio., looking to the right). How the participants
viewed this ambiguous shape was alternated via instructions. This cue
stimulus was presented at fixation at the beginning of each trial and
varied in its gazing/pointing direction from trial-to-trial.
Stimuli and Task
Visual stimuli were presented to participants via rear pro-
jection through the scanner bore onto a mirror, Quale
reflected the image to the participant. Experimental
880
Journal of Cognitive Neuroscience
Volume 20, Numero 5
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
.
/
.
T
F
.
o
N
1
8
M
UN
sì
2
0
2
1
stimuli consisted of a central fixation point, centrally
presented cues and peripherally presented response
targets. The cue was always presented at fixation. IL
response target was simply an asterisk presented to the
left or the right of center. All stimuli were black shapes
presented on a white background.
The task was to fixate centrally on a small point, and to
actively perceive the cue stimulus according to instruction
(either as an eye or as an arrow). Inoltre, participants
were told to press a response button as quickly and as
accurately as possible when they saw the target appear.
Importantly, the direction of the reflexive attentional shift
induced by the cue should vary depending on whether it
is being viewed as an eye or as an arrow.
Procedure
At the beginning of the testing session, participants were
instructed to see the cue stimulus as either an eye or
as an arrow. They were not given any indication at the
outset that the cue stimulus could be viewed any other
modo. Halfway through the testing session, Tuttavia, par-
ticipants were instructed to switch their perceptual set
in order to see the cue as the alternative object; suc-
cessful perceptual switching could be confirmed via the
pattern of response times (RTs) to the targets as a func-
tion of their visual field and the orienting direction of
the percept (see Experiment 3). The testing session was
divided into four functional scanning runs, including two
consecutive runs viewing the cue as an eye, e due
consecutive runs viewing the cue as an arrow. Whether
the cue was first viewed as an eye or as an arrow was
counterbalanced across participants. In all cases, partic-
ipants were informed that cue direction did not reliably
predict the target location.
Each scanning run consisted of 93 trials, which includ-
ed 64 cue–target trials (in which both a cue and a target
occurred), 20 catch trials (in which a cue was not fol-
lowed by a target), E 9 fixation-only trials (in which nei-
ther a cue nor a target occurred) that lasted a duration
of either one, two, or three TRs.
Each cue–target and catch trial began with a 750-msec
blank screen. A small fixation point would then appear
for 750 msec, at which time it would be replaced by the
cue stimulus. For cue–target trials, either 100 O 600 msec
following the onset of the cue, the target would appear
on the left or the right of the cue. The target stayed on
the screen for the remainder of the trial, which lasted for
2250 msec, irrespective of when the response was made.
The screen then blanked for a 750 msec intertrial interval
(ITI). Half of all cue–target trials were cued trials, in which
the target would appear at the gazed-at or pointed-at
location, and the other half of trials were uncued trials, In
which the target would appear at the opposite location.
An equal number of left gazing/pointing and right gazing/
pointing cues were presented randomly in each experi-
mental condition.
fMRI Acquisition and Image Processing
fMRI data were collected on a research-dedicated Phillips
3-T system fitted with a SENSE head coil. Conventional
spin-echo, T1-weighted sagittal localizers were used to
view the positioning of the participant’s head and to set
the functional image volumes to be acquired in line with
the AC–PC plane. Functional image volumes were col-
lected with an echo-planar imaging gradient-echo pulse
sequence (TR = 2000 msec, TE = 30 msec, 908 flip angle,
FOV = 240 (cid:1) 240 mm, 128 (cid:1) 128 matrix, 62.5 kHz band-
width, 3.00 (cid:1) 3.00 mm final measured in-plane resolution,
3.00 mm slice thickness, 1.00 mm slice gap, 36 interleaved
transverse slices), which is sensitive to BOLD contrast.
A transistor-transistor logic (TTL) pulse from the scan-
ner started the scanning run such that the scanner and
the visual presentation of stimuli were synched for
timing. The run consisted of four initial dummy scans
(to allow time to achieve steady-state magnetization) E
166 functional scans. After participants completed four
functional runs, a high-resolution T1-weighted structural
image was acquired with a 3-D gradient-echo pulse
sequence (TR = shortest, TE = shortest, 88 flip angle,
FOV 256 (cid:1) 256 mm, 256 (cid:1) 256 matrix, 1.00 (cid:1) 1.00 mm in-
plane resolution, 1.00 mm slice thickness, 170 slices).
The task was programmed and presented on a per-
sonal computer running Presentation software (Neuro-
behavioral Systems, San Francisco, CA). An event-related
fMRI design was used, which allowed the pseudoran-
dom intermixed presentation of cued, uncued, and catch
trials, as well as fixation-only trials. Responses were col-
lected on a fiber-optic MR-compatible response device
(Lightwave Medical, Vancouver, BC). Stimulus and re-
sponse timing was recorded in a log file that was later
analyzed to generate behavioral RTs.
Functional images were reconstructed off-line. Statis-
tical Parametric Mapping software (SPM2, Wellcome
Institute of Cognitive Neurology, London, UK) was used
for image orientation, motion correction, and spatial nor-
malization into modified Talairach anatomical space. UN
low-pass filter (high frequency cutoff = 6.25 sec) imple-
mented in MATLAB (The Mathworks, Natick, MA) era
applied to the data prior to statistical interrogation in
order to eliminate high-frequency noise confounds not
associated with the BOLD effect.
Statistical Analyses
SPM2 was used to construct a 2 (cid:1) 2 (cid:1) 3 fixed-effects
general linear model for analyzing the group-wise data,
with percept (eye vs. arrow), cue direction (left vs.
right), and cueing (cued vs. uncued vs. catch) as factors.
Each condition was modeled as a set of events time-
locked to the onset of the cue and convolved with a
synthetic hemodynamic response function. Temporal
derivative regressors were also included in the model.
Specific BOLD effects of interest were examined by
Tipper et al.
881
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
.
/
.
F
T
.
o
N
1
8
M
UN
sì
2
0
2
1
creating linear contrasts of the parameter estimates for
each condition. The linear contrasts resulted in a t
statistic for each voxel (unit of fMRI spatial resolution),
which could then be assessed for statistical significance
across the whole brain, thresholded at p < .05, cor-
rected for multiple comparisons.
In order to identify the cortical networks associated
with reflexive attentional orienting to centrally present-
ed spatially nonpredictive cues, we looked at the BOLD
response to eye cues and arrow cues independently. In
order to ascertain whether there are any differences in
attentional networks for orienting to eyes as opposed to
arrows, we looked at the relative BOLD effects (i.e., Eye
cues > Arrow cues).
Results
Behavior
Tavolo 1 shows mean RTs. Regardless of whether the cue
was perceived as an eye or an arrow, responses to cued
targets were faster than responses to uncued targets.
This behavioral cueing effect did not vary as a function of
stimulus-onset asynchrony (SOA) or percept order. An
ANOVA with percept (eye vs. arrow), cueing (cued vs.
uncued), and SOA (100 msec vs. 600 msec) as within-
subject factors, and percept order (eye first vs. arrow
first) as a between-subject factor established the statis-
tical significance of these findings. The main effect of
percept was not significant [F(1, 7) = 1.5, p > .05], In-
dicating no overall difference in RTs between eye and
arrow percepts. The main effect of cueing was significant
[F(1, 7) = 29.5, P < .05], as was the main effect of SOA
[F(1, 7) = 10.0, p < .05]. Neither the cueing factor nor
the SOA factor interacted with any other factors (all ps >
.05). Given that both percepts gave rise to equivalent
Tavolo 1. Response Time Data for Experiment 1 (fMRI)
SOA
100 msec
600 msec
Cue Type
Mean
SE
Mean
SE
Eye Gaze
Cued
Uncued
Arrow
Cued
Uncued
428.78
460.97
417.37
427.91
28.79
34.62
31.62
38.67
363.55
381.85
348.65
374.78
15.75
15.96
18.84
19.38
The results indicate faster responses for cued relative to uncued targets
at both short and long cue–target intervals, regardless of whether the
cue was perceived as an arrow or as an eye. These equivalent atten-
tional orienting effects occurred despite the fact that participants were
assured that the cue did not reliably predict the location of the im-
pending target.
reflexive attentional orienting, the question then was
whether these two types of attentional cues would en-
gage distinct cortical mechanisms.
Functional Magnetic Resonance Imaging
We conducted a two-part analysis of the fMRI data. Primo,
we looked at BOLD responses to the eye gaze and ar-
row cues independently. This allowed us to identify the
cortical regions subserving attentional orienting to each
type of cue. Secondo, we directly compared BOLD re-
sponses to eye and arrow cues in order to identify re-
gions having a differential response to the two types of
cue. Both analyses were time-locked to the onset of the
cue stimulus, and included cued trials only.
The cortical regions in which the BOLD response in-
creased significantly ( P < .05, corrected) with the pre-
sentation of the central object are shown in Figure 2.
The BOLD responses to eye and arrow cues were
similar. For both cue types, posterior regions of activity
included large clusters in the bilateral intraparietal sul-
cus, superior parietal lobule, and the temporal–parietal
junction, including the inferior parietal lobule, and the
superior temporal gyrus (STG). In more anterior brain
regions, there were significant clusters in the bilateral
dorsal frontal cortex, including the middle frontal and
superior frontal gyri. Bilateral anterior clusters were also
found more ventrally in the STG. Prominent BOLD re-
sponses in the occipital cortex, extending into posterior
ventral temporal regions, were also observed. Table 2
provides coordinates and t values for some of the local
maxima in each of these regions.
Although orienting to eye and arrow cues activated
largely similar cortical regions, we were interested in
directly probing the question of whether any cortical re-
gions were uniquely associated with attentional orient-
ing to eye gaze cues. The results of the relative BOLD
contrast ( p < .05, corrected) showing regions respond-
ing more vigorously to eye cues than to arrow cues are
depicted in Figure 3. Four clusters were identified,
including two clusters in the bilateral middle occipital
gyri, a cluster on the ventral surface of the right medial
frontal gyrus, and a cluster in the right inferior precen-
tral gyrus. Table 3 lists coordinates and t values for the
most highly activated voxel in each of these clusters.
When we conducted the reverse contrast, we found no
regions with a significantly greater BOLD response for
arrows than for eyes.
EXPERIMENT 2
The finding that the lateral occipital cortex showed a
larger BOLD response for eye gaze than for arrow cues
was interesting given that the physical stimulus for each
type of cue was identical. One possible explanation for
the enhanced occipital activation is that an eye gaze cue
may be particularly effective for enhancing visual sensory
882
Journal of Cognitive Neuroscience
Volume 20, Number 5
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
5
e
-
8
p
7
d
9
f
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
c
9
n
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
5
1
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
f
t
/
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Attentional
orienting network.
Independent analyses of the
BOLD response to eye gaze
and arrow cues are shown
here. A general linear model
approach (see Experiment 1
Methods) was used to identify
BOLD activity specific to
the processing of the cue.
Activation maps, representing
voxels with a t statistic
greater than 4.63 ( p < .05,
corrected), are overlaid
on the SPM2 single-subject
rendered brain template
(MNI). Largely the same
network was activated while
orienting attention to eye gaze
and arrow stimuli. R = right
hemisphere, L = left
hemisphere; r = rostral; c =
caudal; a = anterior; p =
posterior; IFG = inferior
frontal gyrus; MFG = middle frontal gyrus; SFG = superior frontal gyrus; PreCG = precentral gyrus; Ins = insula; IPL = inferior parietal
lobule; PoCG = postcentral gyrus; SPL = superior parietal lobule; STG = superior temporal gyrus; MTG = middle temporal gyrus; MOG =
middle occipital gyrus; IOG = inferior occipital gyrus.
processing for stimuli appearing at gazed-at locations
(Hopfinger & Ries, 2005). That is, although both eye
and arrow cues induce reflexive shifts in spatial atten-
tion, and utilize largely the same cortical regions to do
so, eye gaze cues may be associated with a larger visual
sensory gain effect in the lateral occipital cortex than ar-
row cues. In order to test this possibility, a second ex-
periment was conducted with an independent group of
participants using event-related potentials (ERPs).
An attention-related sensory gain effect is characterized
by a larger-amplitude P1 ERP component in response
to the onset of a visual stimulus when that stimulus is
presented at an attended (cued) location than when it
is presented at an unattended (uncued) location (cf.
Mangun & Hillyard, 1991). If indeed the more robust
BOLD response in the occipital cortex was caused by a
larger sensory gain effect, then we would expect to find a
larger difference in P1 amplitudes for cued and uncued
targets when the attention-orienting stimulus is perceived
as an eye than when it is perceived as an arrow. The
stimuli and task used in Experiment 2 were identical to
those employed in Experiment 1, with the exception of
some changes made to the timing of stimulus presenta-
tion in order to facilitate ERP data collection.
Methods
Participants
(EEG), and a failure to evoke a distinguishable P1 ERP
component. Of the remaining 11 participants, 5 were
women, and the mean age was 20.55 years. All partic-
ipants had normal or corrected-to-normal vision. Exper-
imental procedures were approved by the University
of British Columbia Behavioral Research Ethics Board.
Participants were remunerated with 10 dollars per hour
of their time.
Stimuli and Task
The stimuli and task employed were nearly identical to
those used in Experiment 1. On most trials, a cue stim-
ulus was presented, followed shortly by an asterisk tar-
get. The directionality of the cue did not predict the
location of the target. There were a few changes made
in order to accommodate the requirements of an ERP
study. The display was presented on a 17-inch CRT mon-
itor at a viewing distance of approximately 100 cm. In
addition, the timing of the stimulus presentation dif-
fered from that used in Experiment 1. The interval
between the cue and the target was randomly selected
on each trial from a rectangular distribution between
500 and 700 msec. A long intertrial interval, randomly
varied between 2400 and 2600 msec, was added for the
purpose of obtaining accurate baseline measures for ERPs.
Thirteen neurologically healthy, right-handed partici-
pants from the University of British Columbia took part
in the study with written consent. The data from two
participants were discarded due to technical problems
leading to excessive noise in the electroencephalogram
Procedure
Participants were fitted with an elastic cap containing an
array of 31 tin electrodes (Electro-Cap International,
Eaton, OH). Half of the participants were instructed
to perceive the ambiguous cue shape as an arrow; the
Tipper et al.
883
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
5
e
-
8
p
7
d
9
f
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
c
9
n
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
5
1
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
f
/
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Table 2. BOLD Cluster Maxima for Eye Gaze and Arrow
Cues, p < .05, Corrected for Multiple Comparisons
Eye Gaze Cue
Arrow Cue
x, y, z
Coordinates
(mm)
t-Score
x, y, z
Coordinates
(mm)
t-Score
52
48
36
52
36
20
(cid:2)56
(cid:2)56
12
32
16 (cid:2)4
56
36
0
16
28
64
6.42
5.32
6.86
7.34
10.03
52
48
48
52
36
12
32
7.55
16 (cid:2)4
8.05
48
36
0
16
28
5.76
4.78
64 10.97
56 (cid:2)16
7.41
–
–
–
p > .05
12
28
8.21 (cid:2)56
12
28 11.01
12 (cid:2)4
4.68 (cid:2)48
12 (cid:2)4
5.05
Region
Frontal
RIFGr
RIFGc
RMFGa
RMFG
RMFGp
RSFG
LIFGr
LIFGc
LMFGa (cid:2)36
52
LMFGp (cid:2)32 (cid:2)4
LPreCG (cid:2)32 (cid:2)20
LIns
(cid:2)40
8
Parietal
RIPL
24 (cid:2)76
LPoCG (cid:2)60 (cid:2)16
LSPL
LIPL
(cid:2)24 (cid:2)64
(cid:2)28 (cid:2)40
Temporal
24
48
68
0
56
24
56
44
6.25 (cid:2)40
44
20
8.52
10.89 (cid:2)36 (cid:2)4
48 12.31
8.30 (cid:2)32 (cid:2)20
68 12.08
6.46 (cid:2)40
8
0
8.41
11.43
40 (cid:2)44
44 10.89
6.41 (cid:2)60
16
12.87 (cid:2)24 (cid:2)64
24
56
8.73
7.88
12.06 (cid:2)44 (cid:2)32
40 12.51
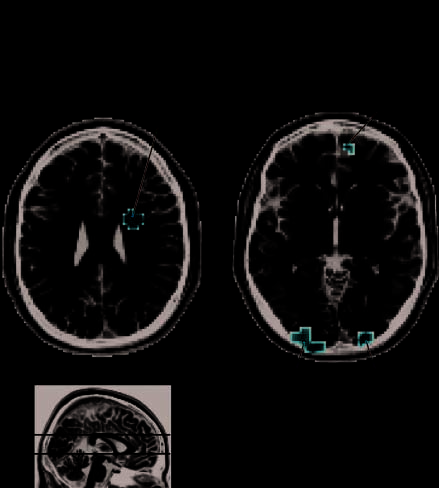
Figura 3. Regions preferentially engaged by the eye gaze cue.
Regions exhibiting a large BOLD response (t > 4.63, P < .05,
corrected) for the eye gaze percept than the arrow percept are
shown here. Activation maps are overlaid on two slices of the SPM
single-subject T1 image template (MNI). Four clusters responded
more vigorously while attending to the eye gaze cue than while
attending to the arrow cue. Panel A shows a cluster in the right
inferior precentral gyrus. Panel B shows three clusters, one on the
ventral surface of the right medial frontal gyrus, and two located in
bilateral middle occipital gyri. R = right hemisphere; L = left
hemisphere; MFG = middle frontal gyrus; PreCG = precentral
gyrus; MOG = middle occipital gyrus.
RSTG
48 (cid:2)40
8
RMTG
52
16 (cid:2)4
7.75
5.92
48 (cid:2)40
8
4.88
52
16 (cid:2)4
6.53
Table 3. BOLD Cluster Maxima for Eye Gaze Cue > Arrow
Cue Statistical Contrast
Occipital
RMOGp
32 (cid:2)92
12
11.29
32 (cid:2)92
12 10.06
RMOGa
48 (cid:2)72 (cid:2)8
14.72
48 (cid:2)72 (cid:2)8 14.63
RIOG
44 (cid:2)76
LMOGp (cid:2)32 (cid:2)92
8
8
15.88
44 (cid:2)76
8
6.41
14.94 (cid:2)32 (cid:2)92
8 11.74
LMOGa (cid:2)44 (cid:2)76 (cid:2)12
14.10 (cid:2)44 (cid:2)76 (cid:2)12 13.77
LIOG
(cid:2)44 (cid:2)84 (cid:2)8
16.14 (cid:2)44 (cid:2)84 (cid:2)8 10.69
Regions listed are represented in Figure 2. R = right hemisphere; L =
left hemisphere; r = rostral; c = caudal; a = anterior; p = posterior;
IFG = inferior frontal gyrus; MFG = middle frontal gyrus; SFG =
superior frontal gyrus; PreCG = precentral gyrus; Ins = insula; IPL =
inferior parietal lobule; PoCG = postcentral gyrus; SPL = superior pa-
rietal lobule; STG = superior temporal gyrus; MTG = middle temporal
gyrus; MOG = middle occipital gyrus; IOG = inferior occipital gyrus.
Region
Frontal
RMFG
RPreCG
Occipital
RMOG
LMOG
Eye Gaze > Arrow Cue
X, sì, z Coordinates (mm)
t-Score
16
36
60
0
28
(cid:2)24
(cid:2)100
(cid:2)100
(cid:2)4
28
(cid:2)4
4
5.57
7.18
6.27
5.14
Data reported here surpass the statistical threshold of p < .05, corrected
for multiple comparisons. Regions listed are represented in Figure 3. R =
right hemisphere; L = left hemisphere; MFG = middle frontal gyrus;
PreCG = precentral gyrus; MOG = middle occipital gyrus.
884
Journal of Cognitive Neuroscience
Volume 20, Number 5
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
5
e
-
8
p
7
d
9
f
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
c
9
n
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
p
c
d
n
.
b
2
y
0
g
0
u
8
e
.
s
t
2
o
0
n
5
1
0
0
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
/
.
.
f
t
.
o
n
1
8
M
a
y
2
0
2
1
other half were instructed to see it as an eye. After 10
trial blocks, participants were then instructed to see the
cue stimulus as the other possible shape, and another
10 blocks were run. Each block lasted approximately
4 min, and consisted of 30 trials, including 28 cue–target
trials, and 2 catch trials, in which no target occurred.
EEG data were collected from 24 scalp electrodes sites
(FP1, FP2, Fz, F7, F8, Cz, C3, C4, T3, T4, P1, P2, Pz, P5,
P6, PO1, PO2, Oz, OL, OR, P3, P4, T5, and T6) using a
Grass Instruments Model 12 amplifier, referenced to the
left mastoid. Three additional channels were recorded,
one from the right mastoid (for off-line referencing of
the data to the average of the two mastoid signals), one
from a pair of electrodes mounted on the outer canthi of
each eye (to record horizontal eye movements), and one
from below the right eye (to record vertical eye move-
ments and blink artifacts). EEG was amplified with a gain
of 50,000 and a half-amplitude band pass of 0.1 to 30 Hz.
Data were digitized at 256 Hz. Off-line, trials with eye
movement artifacts were flagged and not included in any
subsequent analysis. ERP waveforms were digitally re-
referenced to the average of the left and right mastoids,
and low-pass filtered (25.6 half-amplitude cutoff ) prior
to analysis. Peak amplitude measures for the P1 wave-
form were obtained by identifying the latency of the
P1 peak for each condition of interest in the grand-
averaged waveforms, and obtaining the voltage measure
at that latency within each participant. All statistical anal-
yses and waveform displays were conducted with a (cid:2)100
to 0 msec prestimulus baseline.
Results
Behavior
Table 4 shows mean RTs and standard errors. The re-
sults indicate faster responses to cued relative to uncued
targets, regardless of whether the cue was perceived as
an eye or an arrow. Response time data were analyzed
Table 4. Response Time Data for Experiment 2 (ERP)
SOA
500–700 msec
Cue Type
Eye Gaze
Cued
Uncued
Arrow
Cued
Uncued
Mean
276.31
281.08
264.46
276.44
SE
14.67
16.41
14.03
14.99
The results show statistically equivalent attentional orienting effects for
arrow and eye gaze percepts.
with an ANOVA, with percept (eye vs. arrow) and cueing
(cued vs. uncued) as within-subject factors. The main
effect of percept was not significant, [F(1, 10) = 0.5,
p > .05], indicating no overall difference in RTs between
eye and arrow percepts. There was a significant main
effect of cueing [F(1, 10) = 9.7, P < .05]. The interac-
tion of percept and cueing was not significant ( p >
.05), replicating the finding from Experiment 1 that eye
and arrow cues induce equivalent attentional orienting
effects.
Electrophysiology
ERP waveforms time-locked to target onset and averaged
across participants are plotted in Figure 4. These data
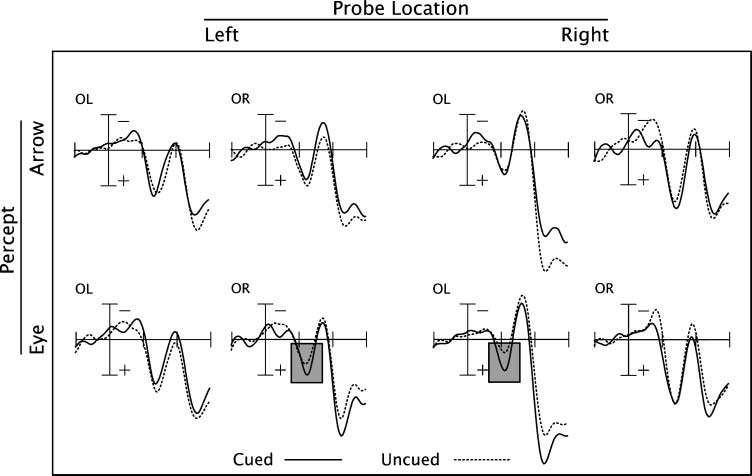
indicate that although both eye gaze and arrow cues
produced behavioral cueing effects, an attention-related
sensory gain effect was present only for eye gaze cues.
Peak amplitude values in the P1 time window (Tavolo 5)
were submitted as the dependent variable in a 2 (cid:1) 2 (cid:1)
2 (cid:1) 2 ANOVA with percept (eye vs. arrow), cueing (cued
vs. uncued), visual Field of the target (left vs. right), E
electrode (ipsilateral vs. contralateral) as within-subject
factors. A significant three-way interaction between per-
cept, cueing, and electrode [F(1, 10) = 4.9, p =.05] era
followed up with simple effects analyses. These analyses
indicated that the three-way interaction was driven by
the presence of a significant sensory gain effect (greater
P1 amplitudes for cued relative to uncued targets) for
the eye gaze percept at electrode sites contralateral [F(1,
50) = 4.6, P < .05], but not ipsilateral [F(1, 50) = 0.4,
p > .05], to the visual field of target presentation. There
was no statistically significant sensory gain effect for the
arrow percept at either contralateral [F(1, 50) = 0.03,
p > .05] or ipsilateral [F(1, 50) = 0.6, p > .05] electrode
sites. The results are consistent with the hypothesis that
attentional orienting in response to an eye gaze cue is
particularly effective at highlighting sensory information
being processed at the gazed-at location.
EXPERIMENT 3
To date, all studies comparing the neural mechanisms of
attentional orienting in response to eye gaze and arrow
stimuli have used physically distinct stimuli. Although
this is an obvious and unavoidable fact of comparing
the orienting response to realistic depictions of distinct
objects, it is important to note that it represents an inher-
ent methodological confound for controlled investi-
gations. Specifically, when comparing the orienting
response to eye gaze and arrow cues with a study that
utilizes physically distinct cues (per esempio., Hietanen et al.,
2006), one can never be certain of whether observed
differences arise because of differences in the physical
stimulus parameters, or due to differences in the mean-
ingful (semantic) representations of those stimuli.
Tipper et al.
885
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
F
.
T
.
.
/
o
N
1
8
M
UN
sì
2
0
2
1
Figura 4. Grand-averaged ERP
waveforms from Experiment 2.
Event-related potentials
recorded at left and right
lateral occipital electrode sites
(OL and OR, rispettivamente),
time-locked to the onset of
the peripheral target stimulus,
were averaged across subjects.
The time window shown spans
from 100 msec pretarget to
300 msec posttarget. The first
large positive deflection in
the waveform represents the
P1 ERP component, peaking
approximately 120 msec
posttarget. Shaded boxes
represent the conditions in
which a significant sensory gain
effect (greater P1 amplitudes for
cued relative to uncued targets)
was observed. The results
indicate the presence of a
sensory gain effect in response
to eye gaze, but not arrow cues
at contralateral electrode sites.
In the present study, having participants perceive the
same physical stimulus as either an eye or an arrow en-
abled a direct comparison of attentional orienting to eye
gaze and arrow cues without confounding different cues
with different stimulus attributes. Tuttavia, the use of
a perceptually ambiguous cue stimulus poses its own
procedural and inferential challenges, where it is of pri-
mary importance to design a stimulus that (1) could be
viewed either as an eye or an arrow with equal ease, (2)
would allow perceptual switches without any negative
transfer, E (3) would produce equivalent attentional
orienting. Although the data from Experiments 1 E 2
support the conclusion that we have met these criteria,
one might argue that the relatively small sample sizes
used in the present study undermined the ability to de-
tect significant differences in the effects of eye gaze
and arrow percepts in Experiments 1 E 2. To address
this concern, we conducted Experiment 3; a behavioral
study designed to (1) ensure that switching percepts
without negative carryover effects is, Infatti, possible,
E (2) replicate the behavioral findings of Experiments
1 E 2 with a larger group of participants enabling more
power.
Experiment 3 specifically addressed the issue of whether
participants are able to switch their perception of the
ambiguous cue-stimulus without negative transfer ef-
fects. We tested two groups of participants: one group
experienced an exact replication of the experimental
design used in Experiment 1, in which the cue percept
was switched once halfway through testing and one group
experienced a different design in which the percept was
switched several times during testing. The logic here is
that if switching percepts interferes with the orienting
effect to eye gaze and/or arrow percepts, these negative
transfer effects will be more pronounced when partici-
pants switch percepts more frequently, leading to differ-
ences in the pattern of results between the single-switch
and multi-switch groups.
Methods
Participants
Seventeen neurologically healthy, right-handed partici-
pants from the University of British Columbia took part
in the study with written consent. The data from one
participant were discarded due to a technical failure
leading to the loss of a large proportion of RT data. Of
Tavolo 5. Mean Peak P1 ERP Voltage Amplitudes (AV) for
Experiment 2
Electrode
Contralateral
Ipsilateral
Cue Type
Mean
SE
Mean
SE
Eye Gaze
Cued
Uncued
Arrow
Cued
Uncued
1.84
1.03
1.50
1.57
0.59
0.53
0.53
0.64
3.02
3.26
2.95
2.66
0.43
0.45
0.44
0.43
Results are averaged over target location. A significant Percept (cid:1) Cueing (cid:1)
Electrode interaction indicates a larger sensory gain effect at contralateral
electrode sites for eye gaze cues than for arrow cues.
886
Journal of Cognitive Neuroscience
Volume 20, Numero 5
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
F
.
T
/
.
.
o
N
1
8
M
UN
sì
2
0
2
1
the remaining 16 participants, 9 were women, and the
mean age was 21.32 years. Participants were assigned to
one of two groups. The single-switch group was in-
structed to switch percepts only once, midway through
testing, as in Experiments 1 E 2. The multi-switch group
was instructed to switch percepts repeatedly throughout
the experiment.
All participants had normal or corrected-to-normal
vision. Experimental procedures were approved by the
University of British Columbia Behavioral Research Ethics
Board. Participants were remunerated with 10 dollars per
hour of their time.
Stimuli, Task, and Procedure
The stimulus design and task were identical to that of Ex-
periment 1, with the exception that half the participants
were requested via an on-screen instruction to switch per-
cepts six times throughout the experiment.
At the beginning of the testing session, participants
in the single-switch group were instructed to perceive the
cue stimulus as either an eye or as an arrow. They were
not given any indication at the outset that the cue stim-
ulus could be viewed any other way. Halfway through the
testing session, Tuttavia, these participants were in-
structed to switch their perceptual set in order to perceive
the cue as the alternative object. Participants in the multi-
switch group, Tuttavia, were instructed from the outset
that they would have to make perceptual switches, E
to do so by following instructions presented on the screen
at regular intervals throughout the experiment.
As in Experiment 1, the testing session was divided into
four blocks of trials, each separated with a short break.
For the single-switch group, there were two consecutive
blocks viewing the cue as an eye, and two consecutive
blocks viewing the cue as an arrow. For the multi-switch
group, participants were instructed to switch percepts
midway through each of the four blocks, as well as be-
tween most blocks. There were two consecutive eye
gaze then arrow blocks, and two consecutive arrow then
eye gaze blocks. Whether the cue was first viewed as an
eye or as an arrow was counterbalanced across partic-
ipants. This design resulted in six perceptual switches
throughout the experiment with a switch order of either
eye/arrow–eye/arrow–arrow/eye–arrow/eye or arrow/eye–
arrow/eye–eye/arrow–eye/arrow. In all cases, participants
were informed that cue direction did not reliably predict
the target location.
Results
Mean RTs are presented in Table 6. The data were ana-
lyzed by a 2 (cid:1) 2 (cid:1) 2 (cid:1) 2 ANOVA, with percept (eye vs.
arrow), cueing (cued vs. uncued), and SOA (100 msec
vs. 600 msec) as within-subject factors, and switch group
(single-switch vs. multi-switch) as a between-subject fac-
tor. The results indicate statistically equivalent patterns
Tavolo 6. Mean Response Times for Experiment 3
SOA
100 msec
600 msec
Group
Cue Type
Mean
SE Mean
SE
Single-Switch Eye Gaze Cued
422.97 24.39 337.91 17.86
Uncued 441.18 28.96 354.09 16.33
Arrow
Cued
401.74 25.34 340.66 20.05
Uncued 449.30 32.00 361.34 18.28
Multi-Switch Eye Gaze Cued
408.31 29.23 362.52 19.70
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
F
.
.
/
T
.
o
N
1
8
M
UN
sì
2
0
2
1
Uncued 418.46 28.86 366.68 20.90
Arrow
Cued
395.29 28.56 363.22 22.98
Uncued 410.40 31.47 376.79 21.19
The results indicate no significant differences between groups, sug-
gesting effective perceptual switching that does not produce any nega-
tive transfer effects. For both groups, responses were faster for cued
relative to uncued targets at both short and long cue–target intervals,
regardless of whether the cue was perceived as an arrow or as an eye.
of RTs associated with eye gaze and arrow percepts
across both groups. There was no significant main effect
of switch group [F(1, 14) = 0.08, p > .05], and no inter-
action of switch group with any other factor (all ps >
.05). Overall, there was no main effect of percept [F(1,
14) = 0.05, p > .05]. There was, Tuttavia, a significant
main effect of cueing [F(1, 14) = 21.4, P < .05], as well
as a main effect of SOA [F(1, 14) = 44.0, p < .05], but no
Percept (cid:1) Cueing interaction [F(1, 14) = 3.6, p > .05].
Così, in both the single-switch and multi-switch groups,
participants were switching percepts appropriately, E
the percepts triggered equivalent attentional orienting
to the cued locations. These results support the conclu-
sion that switching between interpreting the ambiguous
cue stimulus as an eye or an arrow produces no negative
carryover effects.
META-ANALYSIS: EXPERIMENTS 1 TO 3
Although the same pattern of results have been demon-
strated in Experiments 1 A 3, one still might wish to
argue that the relatively small sample sizes within each
of these studies precluded the detection of differences
between either the overall RTs within each percept
condition, or the orienting effects generated by each
percept. To maximize statistical power, we combined
the data from Experiments 1 A 3 in a meta-analysis with
a total of 35 participants.
Methods
As Experiment 2 included only one 600-msec SOA
condition, the RTs from the 100-msec and 600-msec
Tipper et al.
887
SOA conditions within both Experiment 1 and Experi-
ment 3 were averaged. UN 2 (cid:1) 2 (cid:1) 4 ANOVA was
performed, with percept (eye vs. arrow) and cueing
(cued vs. uncued) as within-subject factors, and exper-
iment (Experiments 1, 2, 3) as a between-subject factor.
Results
Although the meta-analysis revealed a difference in over-
all RTs between experiments with a significant main
effect of experiment [F(3, 31) = 8.5, P < .05], experi-
ment did not interact with any other factor (all ps > .05).
There was a significant cueing effect [F(1, 31) = 55.5,
P < .05], indicating faster RTs for cued than for uncued
targets. This cueing effect was equivalent for both eye
gaze and arrow percepts, as indicated by the absence of
a Percept (cid:1) Cueing interaction [F(1, 31) = 2.2, p > .05].
Inoltre, there was no main effect of percept [F(1,
31) = 2.3, p > .05], indicating equivalent RTs in both eye
gaze and arrow percept conditions. This more powerful
meta-analysis converges with, and reinforces, the con-
clusion that participants resolve and maintain eye and
arrow cue percepts with equal ease and efficiency, E
that these percepts induce behaviorally equivalent re-
flexive attentional orienting effects.
GENERAL DISCUSSION
The present study was conducted to test the hypothesis
that humans may have evolved neural mechanisms spe-
cialized for orienting attention to biologically based so-
cial cues. To answer this question, we used fMRI to test
whether different regions of the brain were engaged
when orienting to biologically based eye gaze cues and
nonbiologically based arrow cues. Experiment 1 revealed
an extensive and highly similar BOLD response for both
types of cue, suggesting that, at least in the context of
the present study, the same cortical network subserved
attentional orienting to social and nonsocial cues. Al-
though we did not identify distinct neural modules that
responded only to eye gaze cues, there were specific
regions that responded more vigorously to eye gaze rel-
ative to arrow cues, including the lateral occipital cortex.
The results of Experiment 2, a follow-up ERP study, Rif-
vealed that this larger BOLD response in the occipital
cortex may well have been attributable to the eye gaze
cues evoking a larger sensory gain effect for targets pre-
sented at attended locations.
Before discussing the broader implications of the pres-
ent study, it is important to note that the present results
contrast that of a previous fMRI study that reported that
orienting to arrows recruited areas of the frontal and pa-
rietal attention network, but gaze cues did not (Hietanen
et al., 2006). Hietanen et al. (2006) reported the non-
overlapping networks for directional arrow and diverted
gaze cues within the context of a blocked fMRI design
that required an initial subtraction of activity observed in
a baseline control conditions that consisted of a nondi-
rectional line stimulus and an undiverted gaze stimulus,
rispettivamente. Given the blocked design of the Hietanen
et al., it is possible that either the target stimuli and/or the
undiverted gaze stimulus in the control condition evoked
activity in the frontal and parietal cortex, thus reducing
the likelihood of revealing activations in those areas in
the diverted gaze condition (the authors themselves note
this possibility on p. 411). In contrasto, the present event-
related design permitted the deconvolution of activity
evoked by gaze cues which were physically identical to
the arrow cues without relying on nonequivalent baseline
control conditions, thereby making the present design
much more sensitive to detecting activity in the frontal
and parietal cortex evoked by gaze cues.
A Cortical Network for Reflexive Attention to
Meaningful Stimuli
In addition to our primary research goal—the question
of whether there are distinct neural systems for atten-
tional orienting to social and nonsocial cues—the pres-
ent study addresses more generally the neural systems
involved in reflexive attentional orienting to centrally
presented cues. There has been some debate as to
whether reflexive and volitional orienting are underlain
by distinct neural mechanisms. Early work by Posner
(1980), as well as more recent studies (Sapir, Soroker,
Berger, & Henik, 1999) provided evidence of a role for
the superior colliculus in reflexive attentional orienting.
This role for the superior colliculus, Tuttavia, is largely
associated with abrupt peripheral events that activate
the eye movement system (Rafal & Smith, 1990; Rafal,
Calabresi, Brennan, & Sciolto, 1989). As the present
study used centrally presented cues, and prohibited eye
movements, it is not surprising that there was no BOLD
activation found for the superior colliculus.
To date, there have been very few neuroimaging stud-
ies of reflexive attention in and of itself. The few that
have been conducted have manipulated orienting via
abrupt peripheral events. These results are difficult to
compare with the vast majority of neuroimaging studies
of attention, which typically use centrally presented pre-
dictive arrow cues to induce volitional attentional shifts
(cf. Ristic & Kingstone, 2006 for a detailed consideration
if this issue). By using a nonpredictive centrally pre-
sented directional cue in the present study, the results
can be directly compared to the existing literature re-
garding the cortical networks associated with volitional
attention without confounding reflexive attention with
eye movement preparation.
The large fronto-parietal networks that showed a
significant BOLD response when orienting to both eye
and arrow cues map very well onto the dorsal and ven-
tral fronto-parietal networks identified by Corbetta and
Shulman (2002). Their meta-analysis revealed that the
dorsal fronto-parietal network, consisting of regions of
888
Journal of Cognitive Neuroscience
Volume 20, Numero 5
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
T
/
.
F
.
.
o
N
1
8
M
UN
sì
2
0
2
1
the superior parietal lobule, intraparietal sulcus, middle
and superior frontal gyri, has been, in most studies, asso-
ciated with volitional or willful processes for directing
spatial attention. The ventral fronto-parietal network, con-
versely, consisting of regions in the temporal–parietal
junction (inferior parietal lobule and STG), as well as
the ventral frontal cortex, has been associated with re-
flexive orienting to particularly salient stimuli or infre-
quent events. This pattern of results seems to support
the idea that, Infatti, distinct neural systems underlie
volitional and reflexive attentional orienting. Tuttavia,
it is grounded largely on a confound of central and pe-
ripheral cueing.
The present results demonstrate that when this con-
found is removed, both dorsal and ventral fronto-parietal
networks are involved in reflexive orienting to a direc-
tional cue presented at fixation. This finding suggests an
interaction between dorsal and ventral fronto-parietal net-
works in reflexive attentional orienting to meaningful
stimuli. This is consistent with Corbetta and Shulman’s
(2002) alternative proposal that effective reflexive atten-
tional orienting to a spatial location may require the co-
activation of both these networks—the ventral network
subserving a circuit-breaking or attentional reorienting
function in response to a salient or meaningful stimulus,
and the dorsal network contributing a spatial selectivity
function. An additional implication of the present results
is that dorsal and ventral fronto-parietal networks should
not simply be functionally mapped to volitional (endog-
enous) and reflexive (exogenous) orienting processes.
Nor for that matter should central and peripheral cueing
be mapped to volitional and reflexive orienting, as is mis-
takenly done on occasion (Vecera & Rizzo, 2006).
Biologically Relevant Cues as Inherently
Meaningful Stimuli
A direct comparison of the cortical networks associated
with attentional orienting to eye gaze and arrow cues re-
vealed two clusters in the frontal cortex (in addition to
those found in the lateral occipital cortex) that were
more vigorously engaged by eye gaze than by arrow cues.
Although these regions showed a BOLD response for
both eye gaze and arrow cues, eye gaze cues evoked a
significantly larger BOLD response in these areas. Questo
finding is consistent with previous work suggesting that
eye gaze cues do not engage distinct neural mechanisms
for orienting attention, but recruit the same neural re-
sources more efficiently (Quadflieg et al., 2004). Both of
the ventral frontal regions showing a larger BOLD re-
sponse for eye gaze cues are part of the ventral frontal–
parietal network, which is associated with attentional
reorienting to particularly salient or meaningful stimuli.
Enhanced BOLD activity in these ventral frontal regions is
consistent with the idea that eye gaze cues are particularly
meaningful and are recognized by the attentional system
as such.
This result lends itself to the provocative possibility
that other types of inherently meaningful stimuli may also
induce reflexive attentional orienting, and bring about
enhanced activity in these ventral frontal regions. Our re-
sults indicate that eye gaze is one such meaningful cue.
Other biologically relevant social stimuli, such as finger
pointing or head turning, may also serve as particularly
effective cues for ventral fronto-parietal activation and
reflexive orienting. Inoltre, nonsocial stimuli that are
inherently meaningful in their provision of information
directly useful for planning and coordinating adaptive
behavior, such as looming stimuli (Franconeri & Simons,
2003), or one’s heading point or obstacles on one’s path
during self-motion, may serve as particularly effective
reflexive cues.
Critical Considerations
Does the STS Specialize in Biological Relevance?
È interessante notare, the STS, a structure implicated in atten-
tional orienting to eye gaze stimuli
in both lesion
(Akiyama et al., 2006) and fMRI (Hoffman & Haxby,
2000) studies, did not show greater activation for eye
gaze than for arrow cues. Akiyama et al. (2006) showed
that a patient with a circumscribed lesion to the right
STG was not able to orient attention to eye gaze stimuli,
but orienting to arrow cues was left intact. The authors
concluded that the STS specializes in processing eye
gaze.
Although this result may seem to provide compelling
evidence for the specialization of the STS for processing
eye gaze, it is important to note that the eye gaze and
arrow cues employed in the Akiyama study consisted of
physically different stimuli. Così, it is quite possible that
the physical construction of the stimuli, rather than the
meaning of the stimuli, produced the observed differ-
ences in orienting. In any case, it is also true that al-
though Akiyama et al.’s results speak to the necessity of
the STS in processing eye gaze, they do not preclude the
engagement of the STS during orienting to arrow cues if
the STS is functionally intact. Our data show that when
cue-type and stimulus-construction are not confounded
and the STS is functionally intact, there is a significant
BOLD response in the right STS for both eye gaze and
the arrow cue conditions (Figura 2).
Are the Present Orienting Effects Truly Reflexive?
One might question whether reflexive orienting to gaze
and arrows cues should be considered spatially reflexive,
for instance, in light of the fact that volitional top–down
processes are critical to whether the stimulus cue is
perceived as an eye or an arrow. This concern, how-
ever, confuses volitional acquisition of a percept with
volitional spatial orienting itself. In all cases, and regard-
less of whether an eye or arrow percept is acquired,
Tipper et al.
889
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
F
/
.
.
.
T
o
N
1
8
M
UN
sì
2
0
2
1
participants have no incentive to attend volitionally to
the cued location, questo è, they are informed repeatedly
that the direction of the cue never predicts reliably the
location of a target. Nevertheless, and for both percepts,
attention is shifted rapidly and consistently to the cued
location, thereby satisfying the key criteria for reflexive
spatial orienting (Ristic, Wright, & Kingstone, 2007; Gibson
& Kingstone, 2006; Ristic et al., 2002; Tipples, 2002;
Friesen & Kingstone, 1998). With this point in place, it is
also important to note that although both cues engage
reflexive orienting, and their behavioral effects in the
present study are equivalent, it does not follow that their
attention effects must be identical on all fronts (Ristic,
Wright, & Kingstone, 2007). Infatti, as we found in the
present study, the sensory gain effect from eye gaze cues is
significantly greater than for arrow cues.
Conclusione
The present study demonstrates that reflexive social
Attenzione, at least in the context of perceiving eye gaze
cues, does not require the involvement of a specialized
attentional module network per se. Piuttosto, eye gaze
cues more vigorously engage ventral frontal regions
within a common attentional network and bring about
a larger sensory gain effect at the attended location
than do arrow cues. Ventral frontal regions associated
with the detection of stimulus salience were more highly
activated by eye gaze than arrow cues, even though
these cues consisted of identical physical stimulation
on the retina. Così, the salience of eye gaze cues can-
not be attributed solely to low-level visual properties
such as high contrast or spatial frequency. Piuttosto, any
benefit for eye gaze cues had to have been brought
about by how that cue was being represented. In other
parole, the enhanced sensory processing at the gazed-at
location occurred because the eye gaze cue was more
salient to the attentional system, presumably because
eyes are socially and biologically meaningful (Birmingham,
in press-b). A final,
in press-a,
Bischof, & Kingstone,
intriguing implication of these data, is that our ability to
orient volitionally and reflexively to socially irrelevant
stimuli, including arrowheads, may have arisen as a useful
by-product of a system that developed first, and foremost,
to promote social orienting to stimuli that are biologically
relevant.
Ringraziamenti
This work was supported by graduate student awards to C. T.
by NSERC and MSFHR; grants to T. H. by NIH, NSERC, E
MSFHR; and grants to A. K. by NSERC and MSFHR. We thank
the comments by Todd Heatherton and two anonymous re-
viewers on a previous version of this manuscript.
REFERENCES
Akiyama, T., Kato, M., Muramatsu, T., Saito, F., Umeda, S., &
Kahima, H. (2006). Gaze but not arrows: A dissociative
impairment after right superior temporal gyrus damage.
Neuropsychologia, 44, 1804–1810.
Allison, T., Puce, A., & McCarthy, G. (2000). Sociale
perception from visual cues: Role of the STS region.
Trends in Cognitive Sciences, 4, 267–278.
Birmingham, E., Bischof, W. F., & Kingstone, UN. (in press-a).
Social attention and real world scenes: The roles of action,
competition and social content. Quarterly Journal of
Experimental Psychology.
Birmingham, E., Bischof, W. F., & Kingstone, UN. (in press-b).
Gaze selection in complex social scenes. Visual Cognition.
Corbetta, M., & Shulman, G. l. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neuroscience, 3, 201–215.
Deaner, R. O., & Platt, M. l. (2003). Reflexive social attention
in monkeys and humans. Current Biology, 13, 1609–1613.
Franconeri, S. L., & Simons, D. J. (2003). Moving and
looming stimuli capture attention. Perception &
Psychophysics, 65, 999–1010.
Friesen, C. K., & Kingstone, UN. (1998). The eyes have it!:
Reflexive orienting is triggered by nonpredictive gaze.
Psychonomic Bulletin & Review, 5, 490–495.
Frischen, A., & Tipper, S. P. (2004). Orienting attention via
observed gaze shift evokes longer term inhibitory effects:
Implications for social interactions, Attenzione, and memory.
Journal of Experimental Psychology: General, 133, 516–533.
Gibson, B. S., & Kingstone, UN. (2006). Visual attention and the
semantics of space: Beyond central and peripheral cues.
Psychological Science, 17, 622–627.
Hietanen, J. K., Nummenmaa, L., Nyman, M. J., Parkkola, R.,
& Ha¨ma¨la¨inen, H. (2006). Automatic attention orienting by
social and symbolic cues activates different neural networks:
An fMRI study. Neuroimage, 33, 406–413.
Hoffman, E. A., & Haxby, J. V. (2000). Distinct representations
of eye gaze and identity in the distributed human neural
system for face perception. Nature Neuroscience, 3, 80–84.
Hooker, C. I., Paller, K. A., Gitelman, D. R., Parrish, T. B.,
Mesulam, M. M., & Reber, P. J. (2003). Brain networks for
analyzing eye gaze. Cognitive Brain Research, 17, 406–418.
Hopfinger, J. B., & Ries, UN. J. (2005). Automatic versus
contingent mechanisms of sensory-driven neural biasing and
reflexive attention. Journal of Cognitive Neuroscience, 17,
1341–1352.
Kingstone, A., Friesen, C. K., & Gazzaniga, M. (2000). Reflexive
joint attention depends on lateralized cortical connections.
Psychological Science, 11, 159–166.
Kingstone, A., Tipper, C., Ristic, J., & Ngan, E. (2004). The eyes
have it!: An fMRI investigation. Brain and Cognition, 55,
269–271.
Langton, S. R. H., & Bruce, V. (1999). Reflexive visual orienting
in response to the social attention of others. Visual
Cognition, 6, 541–568.
Langton, S. R. H., Watt, R. J., & Bruce, V. (2000). Do the eyes
have it? Cues to the direction of social attention. Trends in
Cognitive Sciences, 4, 50–58.
Mangun, G. R., & Hillyard, S. UN. (1991). Modulation of
sensory-evoked brain potentials provide evidence for
changes in perceptual processing during visual–spatial
priming. Journal of Experimental Psychology: Umano
Perception and Performance, 17, 1057–1074.
Pelphrey, K. A., & Morris, J. P. (2006). Brain mechanisms
Reprint requests should be sent to Christine M. Tipper, 2136
West Mall, Vancouver, BC, Canada, V6T 1Z4, 604-822-3120, O
via e-mail: tipper@interchange.ubc.ca.
for interpreting the actions of others from biological-motion
cues. Current Directions in Psychological Science, 15,
136–140.
890
Journal of Cognitive Neuroscience
Volume 20, Numero 5
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
.
.
/
T
.
F
o
N
1
8
M
UN
sì
2
0
2
1
Pelphrey, K. A., Morris, J. P., Michelich, C. R., Allison, T., &
McCarthy, G. (2005). Functional anatomy of biological
motion perception in posterior temporal cortex: An fMRI
study of eye, mouth, and hand movements. Cerebral Cortex,
15, 1866–1876.
Perrett, D. I., Smith, P. UN. J., Potter, D. D., Mistlin, UN. J., Head,
UN. S., Milner, UN. D., et al. (1985). Visual cells in the temporal
cortex sensitive to face view and gaze direction. Proceedings
of the Royal Society of London, Series B, 223, 293–317.
Posner, M. IO. (1980). Orienting of attention. Trimestrale
Journal of Experimental Psychology, 32, 3–25.
Quadflieg, S., Mason, M. F., & Macrae, C. N. (2004). The owl
and the pussycat: Gaze cues and visuospatial orienting.
Psychonomic Bulletin & Review, 11, 826–831.
Rafal, R., Calabresi, P., Brennan, C., & Sciolto, T. (1989).
Saccade preparation inhibits reorienting to recently
attended locations. Journal of Experimental Psychology:
Human Perception and Performance, 15, 673–685.
Rafal, R., & Smith, J. (1990). Extrageniculate vision in
hemianopic humans: Saccade inhibition by signals in the
blind field. Scienza, 250, 118–121.
Ristic, J., Friesen, C. K., & Kingstone, UN. (2002). Are eyes
special? It depends on how you look at it. Psychonomic
Bulletin & Review, 9, 507–513.
Ristic, J., & Kingstone, UN. (2006). Attention to arrows:
Pointing to a new direction. Quarterly Journal of
Experimental Psychology, 59, 1921–1930.
Ristic, J., Wright, A., & Kingstone, UN. (2007). Attentional
control and reflexive orienting to gaze and arrow cues.
Psychonomic Bulletin & Review, 14, 964–969.
Sapir, A., Soroker, N., Berger, A., & Henik, UN. (1999). Inhibition
of return in spatial attention: Direct evidence for collicular
generation. Nature Neuroscience, 2, 1053–1054.
Tipples, J. (2002). Eye gaze is not unique: Automatico
orienting in response to uninformative arrows.
Psychonomic Bulletin & Review, 9, 314–318.
Vecera, S. P., & Rizzo, M. (2006). Eye gaze does not
produce reflexive shifts of attention: Evidence from
frontal lobe damage. Neuropsychologia, 44, 150–159.
Vuilleumier, P. (2002). Facial expression and selective
Attenzione. Current Opinion in Psychiatry, 15,
291–300.
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
0
l
5
e
–
8
P
7
D
9
F
1
/
9
2
3
0
6
/
7
5
9
/
8
8
o
7
C
9
N
/
1
2
0
7
0
5
8
9
5
2
1
0
6
5
1
/
0
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
8
e
.
S
T
2
o
0
N
5
1
0
0
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
.
F
/
.
T
.
o
N
1
8
M
UN
sì
2
0
2
1
Tipper et al.
891