Attentional Capacity Limits Gap Detection
during Concurrent Sound Segregation
Ada W. S. Leung1,2, Pierre Jolicoeur3,4,5,6, and Claude Alain2,7
Astratto
■ Detecting a brief silent interval (cioè., a gap) is more difficult
when listeners perceive two concurrent sounds rather than one
in a sound containing a mistuned harmonic in otherwise in-
tune harmonics. This impairment in gap detection may reflect
the interaction of low-level encoding or the division of attention
between two sound objects, both of which could interfere with
signal detection. To distinguish between these two alternatives,
we compared ERPs during active and passive listening with
complex harmonic tones that could include a gap, a mistuned
harmonic, both features, or neither. During active listening, par-
ticipants indicated whether they heard a gap irrespective of mis-
tuning. During passive listening, participants watched a
subtitled muted movie of their choice while the same sounds
were presented. Gap detection was impaired when the complex
sounds included a mistuned harmonic that popped out as a sep-
arate object. The ERP analysis revealed an early gap-related activ-
ity that was little affected by mistuning during the active or
passive listening condition. Tuttavia, during active listening,
there was a marked decrease in the late positive wave that was
thought to index attention and response-related processes.
These results suggest that the limitation in detecting the gap is
related to attentional processing, possibly divided attention in-
duced by the concurrent sound objects, rather than deficits in
preattentional sensory encoding. ■
INTRODUCTION
The auditory environment is often composed of a myriad
of sound sources, all of which compete for our attention.
According to the object-based account of auditory atten-
zione (Shinn-Cunningham, 2008; Alain & Arnott, 2000), UN
predominantly automatic segregation process results in
the formation of objects that are subsequently stored in
STM. The notion of “auditory object” refers to a grouping
of sounds into a coherent whole or stream such that it
seems to emanate from a single acoustic source (Alain,
2007). According to the auditory scene analysis account
(Bregman, 1990), incoming concurrent sounds are initially
analyzed and then perceptually grouped into distinct
sound objects based on the physical properties of the
sound, such as common onsets and offsets, harmonic
structure, and continuity of frequency over time (Bregman,
1990). These sound objects form the basic units for atten-
tional selection (Dyson & Ishfag, 2008; Shinn-Cunningham,
2008; Alain & Arnott, 2000). Evidence from behavioral and
EEG studies demonstrated that sequential (Snyder & Alain,
1University of Alberta, 2Rotman Research Institute, Baycrest
Centre for Geriatric Care, Toronto, Canada, 3Université de
Montréal, 4Centre de Recherche en Neuropsychologie et
Cognition (CERNEC), Montréal, Canada, 5BRAMS (Internazionale
Laboratory for Brain, Music, and Sound Research), Montréal,
Canada, 6Centre de Recherche de l’Institut Universitaire de
Gériatrie de Montréal (CRIUGM), 7University of Toronto
© 2015 Istituto di Tecnologia del Massachussetts
2007; Cusack, Carlyon, & Robertson, 2000) and concurrent
(Dyson, Alain, & Lui, 2005; Alain & Izenberg, 2003) sound
segregation can occur irrespective of a listener’s attention.
Once the auditory scene has been partitioned into distinct
sound objects, a selection process allows an individual to
focus or switch their attention from one sound object to
another (Backer & Alain, 2014). Although there is good ev-
idence that the perception of sound stimuli is influenced
by factors like stimulus salience or behavioral goals and that
these factors likely interact to determine which sound(S)
are perceived, attended, and remembered, how these pro-
cesses interact when multiple sound objects compete for
attention remains unknown.
Recentemente, we showed that the perception of a gap (cioè.,
a brief silence) inserted into a complex harmonic sound
was more difficult when it included a mistuned compo-
nent that “popped out” as a separate auditory object
(Leung, Jolicoeur, Vachon, & Alain, 2011). This effect
was observed even for gap durations well above the usual
perceptual threshold (for sounds that do not contain a
mistuned component). Leung et al. proposed that per-
ceiving the gap was more difficult because the attention
was divided between two competing sound objects,
thereby interfering with the detection of the gap. How-
ever, other reasons could explain why detecting the gap
was more difficult during the presence of an inharmonic
component (Heinrich, Alain, & Schneider, 2004). One
could imagine that the presence of a mistuned harmonic
in an otherwise in-tune harmonic complex could reduce
Journal of Cognitive Neuroscience 27:11, pag. 2186–2196
doi:10.1162/jocn_a_00849
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
o
D
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
C
3
N
5
_
8
UN
/
_
j
0
o
0
C
8
N
4
9
_
UN
P
_
D
0
0
B
8
sì
4
G
9
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
2
io
3
e
S
/
j
F
/
T
.
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
gap detection because of low-level peripheral interac-
tions such as beating, auditory induction, perceptual illu-
sion, or restoration, which are thought to result from the
reorganization of acoustic energy across multiple wave
bands and phases (Recanzone & Sutter, 2008). From be-
havioral data alone, it is difficult to determine whether
the gap detection impairment is because of impover-
ished sensory or perceptual processing of the sound
stimuli or a division of attention. Questo è, deficits in gap
detection when two sound objects are simultaneously
presented may be because of a failure in encoding or in-
sufficient processing at a higher cognitive level.
The recording of scalp ERPs provides a mean to assess
the automatic encoding of sound features such as harmo-
nicity and gap as well as higher cognitive processes such
as attention and memory. Prior research has revealed
neural correlates for both concurrent sound perception
and gap detection. For instance, the perception of con-
current sound objects is associated with an enhanced
negativity that overlaps with the N1 and P2 waves elicited
by sound onset (Dyson & Alain, 2004; Alain, Schuler, &
McDonald, 2002; Alain, Arnott, & Picton, 2001). This en-
hanced negativity, referred to as the object-related nega-
attività (ORN), is most prominent over the frontocentral
scalp region and is best isolated by a difference wave be-
tween the ERPs elicited by tuned and mistuned stimuli
(such as those including a component mistuned by 16%
that clearly pops out of the complex; Alain, 2007; Alain
et al., 2001). With respect to gap detection, short duration
sounds (per esempio., 200 msec) that include even shorter gaps in
the middle generate smaller sensory evoked responses
(cioè., N1 and/or P2 waves) relative to stimuli without a
gap (Ross, Schneider, Snyder, & Alain, 2010; Heinrich
et al., 2004; Hillyard & Picton, 1978). Gap-related neural ac-
tivity can be isolated by subtracting auditory ERPs elicited
by sounds with and without the gap. Both the ORN and the
gap-related activity can even be observed when listeners
are performing another task (per esempio., reading a book, watching
a movie; Alain, 2007; Alain et al., 2001).
È interessante notare, when participants are actively processing
a mistuned harmonic or gap, the perceptual decision re-
garding the stimuli elicits a late positive complex (LPC)
that peaks at about 600 msec over the parietal scalp re-
gion in addition to the ORN or gap-related activity. Questo
component has been associated with conscious identifi-
cation of an auditory event and is indicative of attentional
processes (Alain et al., 2002; Martin, Sigal, Kurtzberg, &
Stapells, 1997; Parasuraman, Richer, & Beatty, 1982).
The amplitude of the LPC is strongly modulated by atten-
tion and target detectability (per esempio., Dell’Acqua et al., 2015;
Picton, 1992). Many studies have reported that the mean
amplitude and mean area of the LPC are associated with
behavioral responses in attention and STM tasks (Xu,
Zhang, Ouyang, & Hong, 2013; Wolk et al., 2006; Curran,
Schacter, Johnson, & Spinks, 2001).
In this study, we measured auditory ERPs during both
passive and active listening conditions to examine whether
harmonicity and gap-related activities interact during the
encoding and attentional processes. By examining the N1
and P2 waves as well as the attention-dependent LPC, one
could determine whether gap detection impairments in
the previous study (Leung et al., 2011) were because of fail-
ure of low-level sensory encoding or the division of atten-
tion between concurrent sound objects. If the deficits in
gap detection were because of limitations during the en-
coding process, then the amplitude of sensory evoked re-
sponses elicited by the gap stimuli should be modulated by
mistuning. Conversely, if the deficits in gap detection were
because of a division of attention, then one would expect
greater LPC amplitudes for gap detection in tuned com-
pared with mistuned stimuli as well as correct compared
with incorrect responses. Inoltre, if the ORN indexes
the perception of concurrent sound objects, then one
would expect greater ORN amplitudes for incorrect than
correct responses (cioè., deficits in hearing the gap imply
greater likelihood of hearing the mistuned harmonic as a
separate object).
METHODS
Participants
Twenty right-handed young adults (mean age = 24.30 ±
4.27 years, 10 women) gave informed consent and partic-
ipated in the study. All participants had pure tone thresh-
olds below 30 dB HL for frequencies ranging from 250 A
8000 Hz. None of them had neurological or psychological
illnesses or were taking medication at the time of the ex-
periment. The study was approved by the research ethics
board of the Toronto Academic Health Sciences Network
and the University of Toronto Human Subject Review
Committee. Participants received $25 in compensation
for their participation in the study.
Stimuli and Task
Stimuli consisted of four different complex sounds
(200 msec in duration, 2.5-msec rise and fall time) gener-
ated by adding 10 pure tones of equal intensity. All stimuli
had a fundamental frequency ( f0) Di 200 Hz. For half of the
stimuli, which were referred to as “tuned,” all tonal ele-
ments were an exact integer multiple of f0 (cioè., 200, 400,
600, 800, 1000, 1200, 1400, 1600, 1800, E 2000 Hz). For
the other half of the stimuli, which were referred to as
“mistuned,” the fourth tonal element (4 × f0) was shifted
upward by 16% of the original frequency (cioè., 928 Hz in-
stead of 800 Hz). For each of the tuned and mistuned
harmonics, half of the stimuli included a 15-msec gap
that had a 2.5-msec rise and fall time and a 10-msec
zero-amplitude plateau inserted in the middle of the third
tonal element (600 Hz). The sounds were generated digi-
tally at a sampling rate of 48.8 kHz using the System 3
Real-Time Processor from Tucker Davis Technologies RP-
2 real-time processor (Alachua, FL). They were presented
Leung, Jolicoeur, and Alain
2187
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
o
D
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
C
3
N
5
_
8
UN
/
_
j
0
o
0
C
8
N
4
9
_
UN
P
_
D
0
0
B
8
sì
4
G
9
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
2
io
3
e
S
/
j
/
F
.
T
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
binaurally through insert earphones (ER-3A; Etymotic
Research, Elk Grove, CA) at 68-dB sound pressure level
as measured with a sound pressure level meter using an
artificial ear (Dalimar Instrument Inc., Quebec, Canada).
All participants took part in a passive listening condi-
tion and an active listening condition. A total of 1200
stimuli were presented in each condition. The passive
condition consisted of two blocks of 600 stimuli pre-
sented in random order (150 stimulus presentations ×
4 stimulus types). The ISI varied randomly between
1800 E 2200 msec (100-msec steps, rectangular distri-
bution). The active condition consisted of six blocks of
200 stimuli presented in random order (50 stimulus pre-
sentations × four stimulus types). The ISI varied between
2300 E 2700 msec (100-msec steps, rectangular dis-
tribution) to accommodate the participant’s response
on each trial. We used a custom Matlab program with
Psychophysical Toolbox (version 11.0; The MathWorks,
Natick, MA) on a Dell Pentium 4 PC with a SoundBlaster
Live sound card (Creative Technology, Ltd.) to run the
experiment.
All participants completed the passive listening condi-
tion before completing the active listening condition.
During passive listening, participants watched a silent
subtitled movie of their choice while the auditory stimuli
were presented. Before participants started the active lis-
tening condition, they were given a chance to familiarize
themselves with the stimuli by listening to each stimulus
played six times consecutively in the following order:
tuned, tuned gap, mistuned, and mistuned gap. During
active listening, the participants indicated the presence
or absence of the gap for each stimulus by pressing 1
O 3 on a computer keypad, rispettivamente. Participants
were given rest breaks between blocks. The experiment
took place in a sound-attenuating chamber.
Recording of Neuroelectric Brain Activity
The EEG was digitized continuously (sampling rate =
500 Hz) from an array of 64 electrodes with a bandpass
filter of 0.05–100 Hz using NeuroScan Synamps2 (Com-
pumedics, El Paso, TX). Eye movements were monitored
with electrodes placed at the outer canthi and at the in-
ferior orbits. During recording, all electrodes were refer-
enced to the midline central electrode (cioè., Cz). For
offline data analysis, they were rereferenced to an aver-
age reference. The analysis epoch consisted of 200 msec
of prestimulus activity and 800 msec of poststimulus ac-
attività. For each participant, a set of ocular movements
was obtained before and after the experiment (Picton
et al., 2000). From this set, averaged eye movements
were calculated for both lateral and vertical eye move-
ments as well as for eye blinks. A PCA of these averaged
recordings provided a set of components that best ex-
plained the eye movements. The scalp projections of
these components were then removed from the experi-
mental ERPs to minimize ocular contamination, using
BESA 5.2.0. Epochs contaminated by excessive deflec-
zioni (greater than ±100 μV anywhere in the epoch) after
correcting for ocular contaminations were excluded from
the averages. For each participant, the remaining epochs
were averaged according to electrode position, stimulus
type (cioè., tuned, no gap; tuned, with gap; mistuned, NO
gap; and mistuned, with gap), experimental condition
(cioè., active and passive listening conditions), and gap de-
tection performance (cioè., correct rejection, hit, false
alarm, and miss responses) using BESA. The ERPs were
digitally filtered to attenuate frequencies above 30 Hz
(12 dB/Oct, zero phase).
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
H
T
T
P
:
/
D
/
o
M
w
io
N
T
o
P
UN
R
D
C
e
.
D
S
F
io
R
o
l
M
v
e
H
R
C
P
H
UN
D
io
io
R
R
e
.
C
C
T
.
o
M
M
/
j
e
o
D
tu
C
N
o
/
C
UN
N
R
UN
T
R
io
T
io
C
C
l
e
e
–
P
–
D
P
D
2
F
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
C
3
N
5
_
8
UN
/
_
j
0
o
0
C
8
N
4
9
_
UN
P
_
D
0
0
B
8
sì
4
G
9
tu
.
e
P
S
T
D
o
F
N
B
0
sì
8
S
M
e
IO
P
T
e
M
l
io
B
B
e
R
R
UN
2
R
0
2
io
3
e
S
/
j
.
/
T
F
tu
S
e
R
o
N
1
7
M
UN
sì
2
0
2
1
Data Analysis
Behavioral Data
0) and response bias (β) were calcu-
A sensitivity index (D
lated from hit and false alarm rates of gap detection in
each of the two conditions (tuned and mistuned har-
monics) during the active listening condition (Macmillan
& Creelman, 1991). Paired t tests were performed on the
0 and β values as well as the hit and false alarm rates, E
D
RT, to compare gap detection performance as a function
of harmonicity (tuned vs. mistuned).
Analyzing sensory encoding from 0 A 300 msec. IL
analysis of sensory encoding focused on comparing the
magnitude of the evoked potential for the mistuning and
gap-related component. A four-way repeated-measures
ANOVA was computed and included the following within-
subject factors: condition (active vs. passive), harmonicity
(tuned vs. mistuned), gap (presence vs. absence), and site
(cioè., frontocentral electrodes: FC1, FCz, FC2, C1, Cz, C2,
CP1, CPz, and CP2). These nine electrodes were selected
because they best captured the evoked responses corre-
sponding to the mistuning and the gap-related compo-
nents (Heinrich et al., 2004; Alain et al., 2001). The ERPs
for the mistuning and gap conditions were quantified as
the mean amplitude between 100 E 200 msec and be-
tween 175 E 275 msec, rispettivamente. These time windows
were chosen based on prior research using comparable
stimuli. The effect of mistuning typically peaks at about
150 msec after sound onset (Alain et al., 2001, 2002). IL
gap-related activity peaks at about 125 msec after gap onset
(Campbell & Macdonald, 2011; Heinrich et al., 2004),
which would correspond to about 225 msec after sound
onset as the gap onset was at about 100 msec in this study.
This 100-msec adjustment is not applicable to the mistun-
ing component as the onset of the mistuning is the same as
the onset of the sound.
Analyzing attention during gap detection from 0 A
300 msec. The first analysis was performed to examine
the perception of the mistuned harmonic in the complex
sound during the active listening condition by extracting
the ORN from the ERP waveforms. This analysis allows us
2188
Journal of Cognitive Neuroscience
Volume 27, Numero 11
to determine whether mistuning has an impact on the
detection of a gap in the sound stimulus, questo è, we would
expect a significantly larger ORN for miss responses if the
impairment of detecting the gap is because of the inter-
ference of the mistuning. The ORN was obtained by
computing a difference wave between the tuned and mis-
tuned stimuli. Mean ORN amplitude was extracted be-
tween 100 E 200 msec (Alain et al., 2001, 2002). For
the ORN analysis, a three-way repeated-measures ANOVA
was computed, which included the within-subject factors
Gap, Response type, and Site.
A second analysis was performed to determine wheth-
er there was an interaction between gap detection accu-
racy and harmonicity. A three-way repeated-measures
ANOVA on gap-related activity (mean amplitude between
175 E 275 msec) was computed, which included the
factors Gap detection response (cioè., hit vs. miss), Harmo-
nicity (cioè., tuned vs. mistuned), and Site.
Analyzing attention during gap detection from 400 A
800 msec. The LPC was quantified using the total posi-
tive area between 400 E 800 msec poststimulus onset.
The mean area-under-the-curve provides a better estimate
of the LPC than peak amplitude at the individual level
(Luck, 2014), and the use of this measure, or the mean
amplitude in a given window, is common (per esempio., Dell’Acqua
et al., 2015; Hasko, Groth, Bruder, Bartling, & Schulte-
Korne, 2013). A four-way repeated-measures ANOVA was
calculated using the factors Harmonicity (tuned vs. mis-
0 and β (UN)
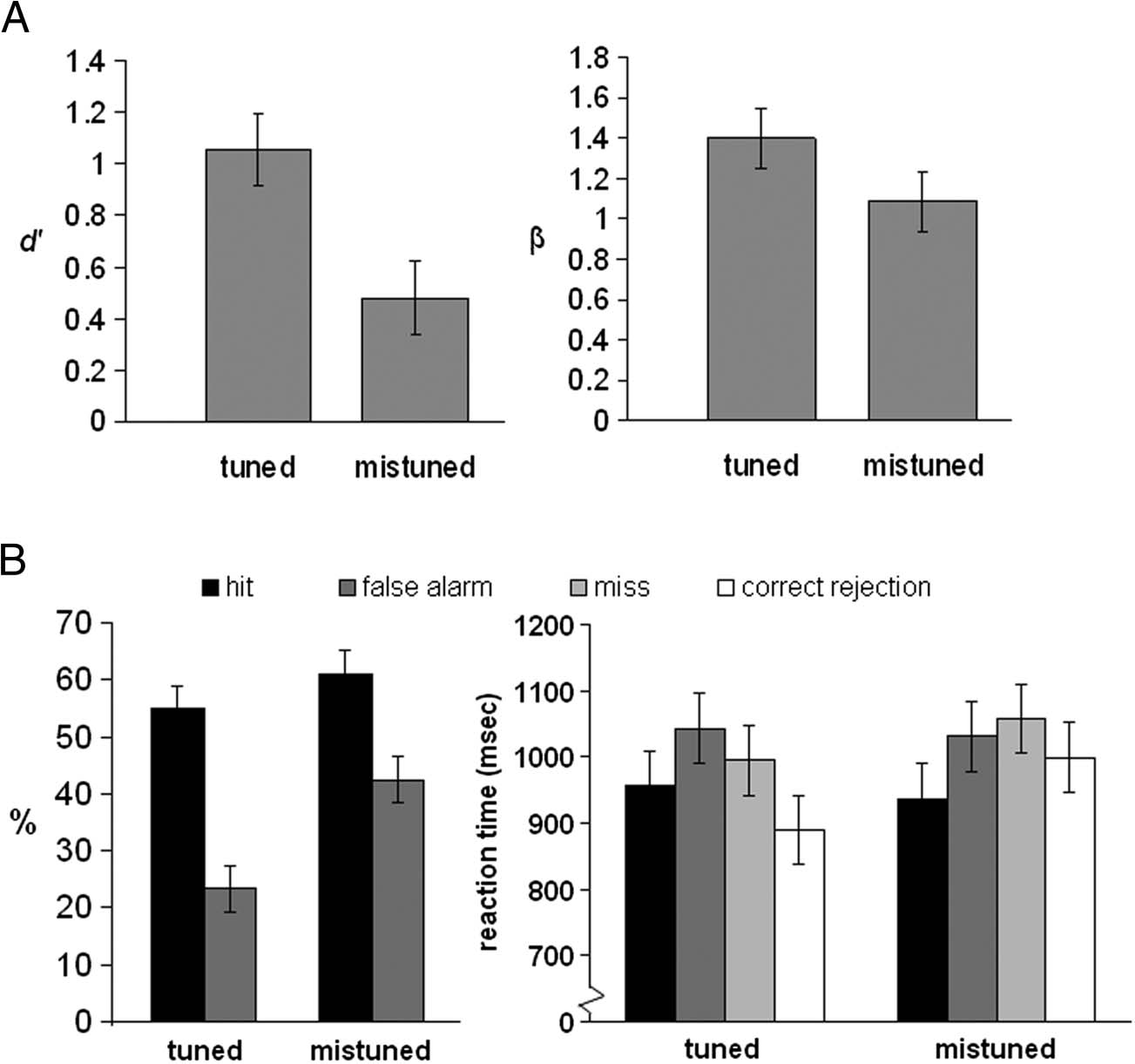
Figura 1. Plot of d
and hit rate and RT (B) for the
gap detection task. Error bars
indicate the SEM.
tuned), Gap (presence vs. absence), Response type (cor-
rect vs. incorrect), and Site (parieto-occipital electrode
sites, namely, P1, Pz, P2, PO3, POz, PO4, O1, Oz, E
O2). These nine electrodes best captured the LPC
elicited by auditory stimuli (McDonald & Alain, 2005;
Alain et al., 2001). Significant interactions between har-
monicity, gap, and/or response type were examined
using pairwise comparisons with Bonferroni adjust-
ment. For all repeated-measures ANOVAs, we used the
Greenhouse–Geisser to correct for any violations of as-
sumption of sphericity (homogeneity of the covariance
matrix).
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
RESULTS
Behavioral Response and Trial Types for
ERP Analysis
0 and β values
Figura 1 shows the mean accuracy (mean d
as well as hit and false alarm rates) and mean RT for the
gap detection task. The hit rate was comparable across
tuned and mistuned stimuli, T(19) = 1.01, p = .325, Ma
the false alarm rate was higher for trials with mistuned
stimuli, T(19) = 3.96, P < .001. Using the signal detection
0 was significantly lower for mistuned
index, the mean d
stimuli, t(19) = 3.98, p < .001. There was also a marginal
effect of mistuning for the mean response bias, β, t(19) =
2.01, p = .06. Participants were more prone to respond
“gap” during trials with mistuned stimuli. For the tuned
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
c
3
n
5
_
8
a
/
_
j
0
o
0
c
8
n
4
9
_
a
p
_
d
0
0
b
8
y
4
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Leung, Jolicoeur, and Alain
2189
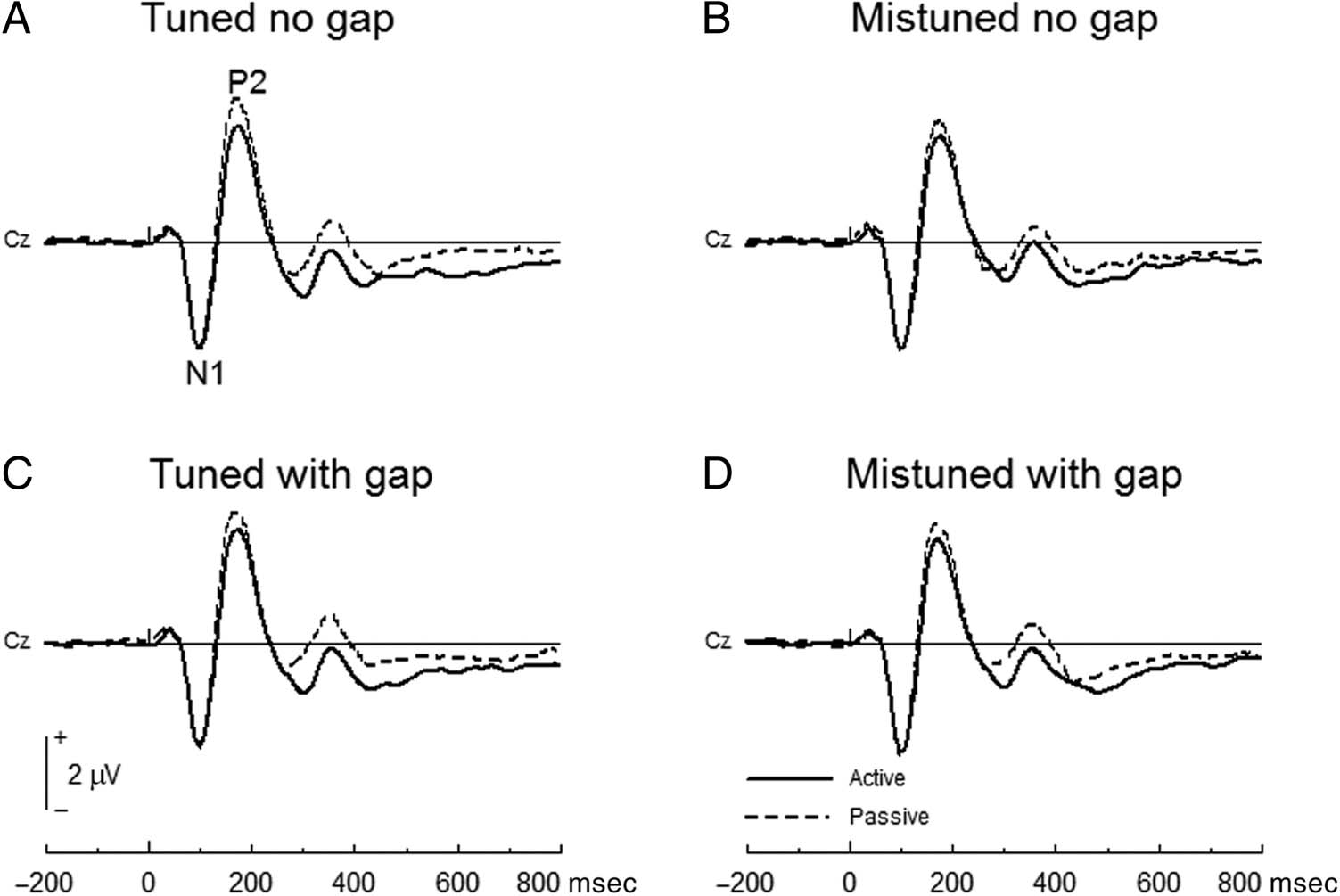
Figure 2. Plot of N1 and P2
waves of the active and passive
listening conditions.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
c
3
n
5
_
8
a
/
_
j
0
o
0
c
8
n
4
9
_
a
p
_
d
0
0
b
8
y
4
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
/
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
stimuli, the participants were more accurate in detecting
gaps during trials with gap stimuli (Figure 1B).
RT was significantly shorter for correct than incorrect
responses, F(1, 19) = 11.29, p = .0035, and shorter for
tuned than mistuned stimuli, F(1, 19) = 4.81, p = .041.
This pattern of results replicated the behavioral findings
in Leung et al.’s (2011) study.
Overall, the number of trials available for ERP averag-
ing was comparable among the eight trial types, with an
average of 136 ± 45 trials, F(1, 19) = 1.18–3.16, ps =
.091–.291. However, there were slightly more trials
where the participants correctly rejected the tuned stim-
uli (211 ± 50 trials) rather than the false alarms for tuned
stimuli response (65 ± 42 trials).
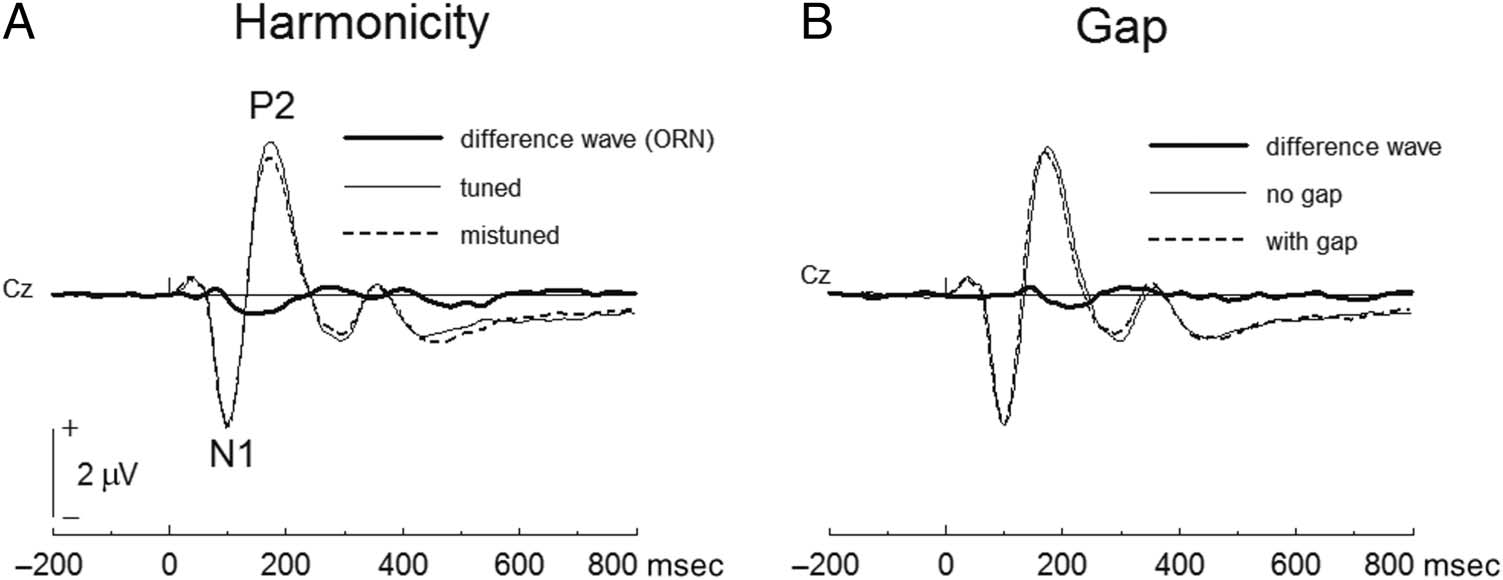
Assessing the Encoding of Mistuning and Gap
Figure 2 shows the ERP waveforms for the active and pas-
sive listening conditions. For the mistuning compo-
nent (i.e., the ORN: mean amplitude between 100 and
200 msec), repeated-measures ANOVA (Condition × Har-
monicity × Gap × Site) revealed significant main effects of
Condition, F(1, 19) = 8.79, p = .008 (larger mean ampli-
tude in passive than active conditions), and Harmonicity,
F(1, 19) = 27.22, p < .001 (larger mean amplitude in tuned
than mistuned). Interaction effects among Condition, Har-
monicity, and Gap were not significant, F(1, 19) = 0.16, p =
.690, and interaction effects of these factors with Site were
also not significant, F(8, 152) = 0.39–3.17, ps = .091–.927.
For the gap-related component (mean amplitude be-
tween 175 and 275 msec), repeated-measures ANOVA re-
vealed significant main effects of Condition, F(1, 19) =
5.01, p = .037 (larger mean amplitude in passive than
active conditions), and Gap, F(1, 19) = 8.80, p = .008
(larger mean amplitude in stimuli without a gap than
stimuli with a gap). Interaction effects among Condition,
Harmonicity, and Gap were not significant for both
the mistuning and gap responses, F(1, 19) = 2.641,
Figure 3. ERP waveforms at Cz
for comparing between tuned
and mistuned stimuli (A) and
gap and no-gap stimuli (B). The
waveforms are average of active
and passive listening conditions.
Difference waves show the ORN
(A) and gap-related activity (B).
2190
Journal of Cognitive Neuroscience
Volume 27, Number 11
p = .121, and interaction effects of these factors with Site
were also not significant, F(8, 152) = 0.55–2.06, ps =
.168–.466. The results indicate that the mistuning and
gap were independently encoded and the encoding pro-
cess did not differ between the active and passive listen-
ing conditions. Figure 3 shows the ERP waveforms of the
mistuning (the ORN component) and gap-related
response.
Assessing Attention during Gap Detection
ANOVA Results for the ORN
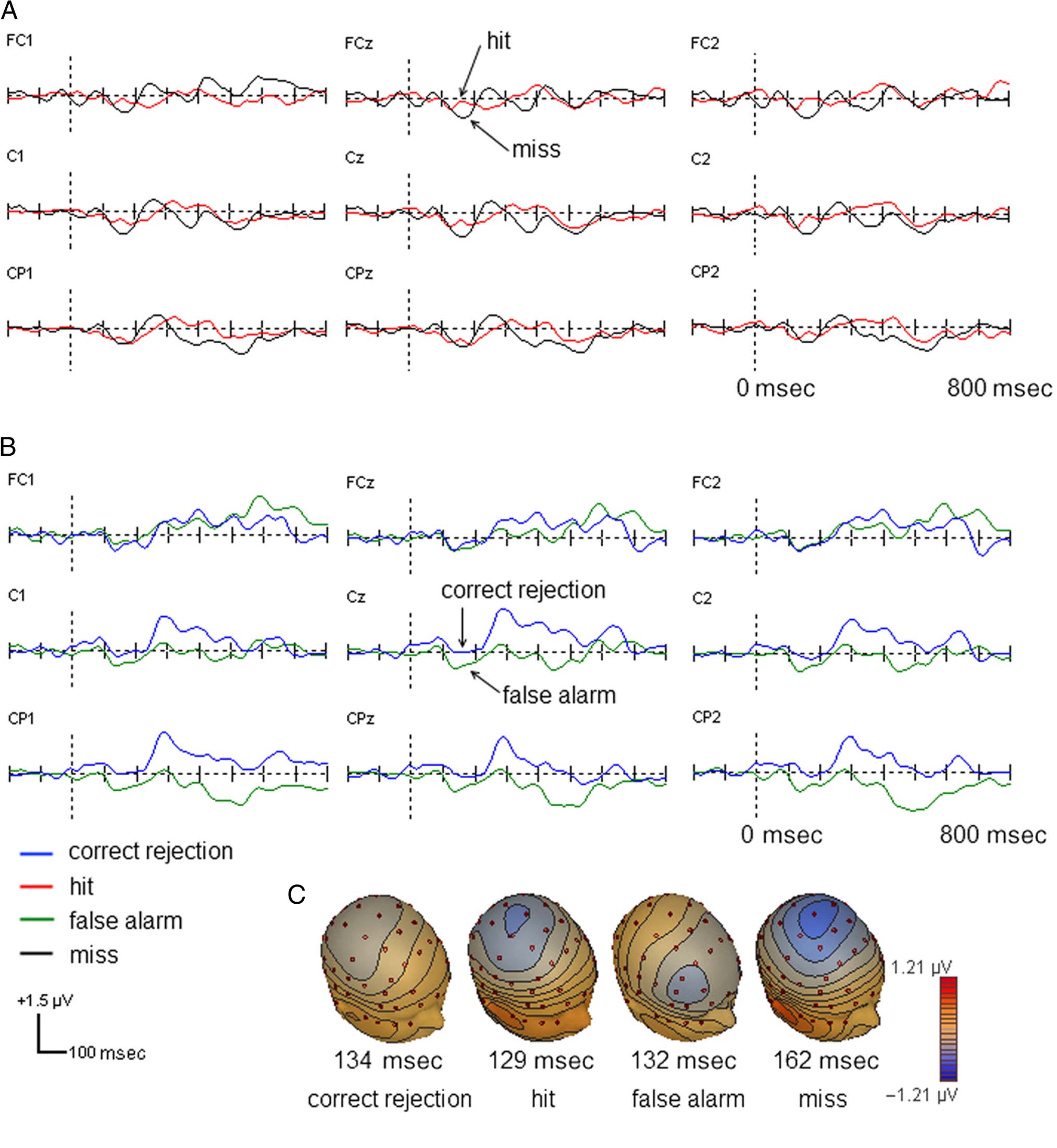
Figure 4 shows the group mean ORN waveforms. Using
the ORN (mean amplitude between 100 and 200 msec
from the difference waves between the tuned and mistuned
stimuli), repeated-measures ANOVA (Gap × Response
type) revealed a significant main effect of Response type,
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
c
3
n
5
_
8
a
/
_
j
0
o
0
c
8
n
4
9
_
a
p
_
d
0
0
b
8
y
4
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 4. Plots of the ORN for hit and miss responses (A) and correct rejection and false alarm (B) and their topography as a function of response (C).
Leung, Jolicoeur, and Alain
2191
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
c
3
n
5
_
8
a
/
_
j
0
o
0
c
8
n
4
9
_
a
p
_
d
0
0
b
8
y
4
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
/
.
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
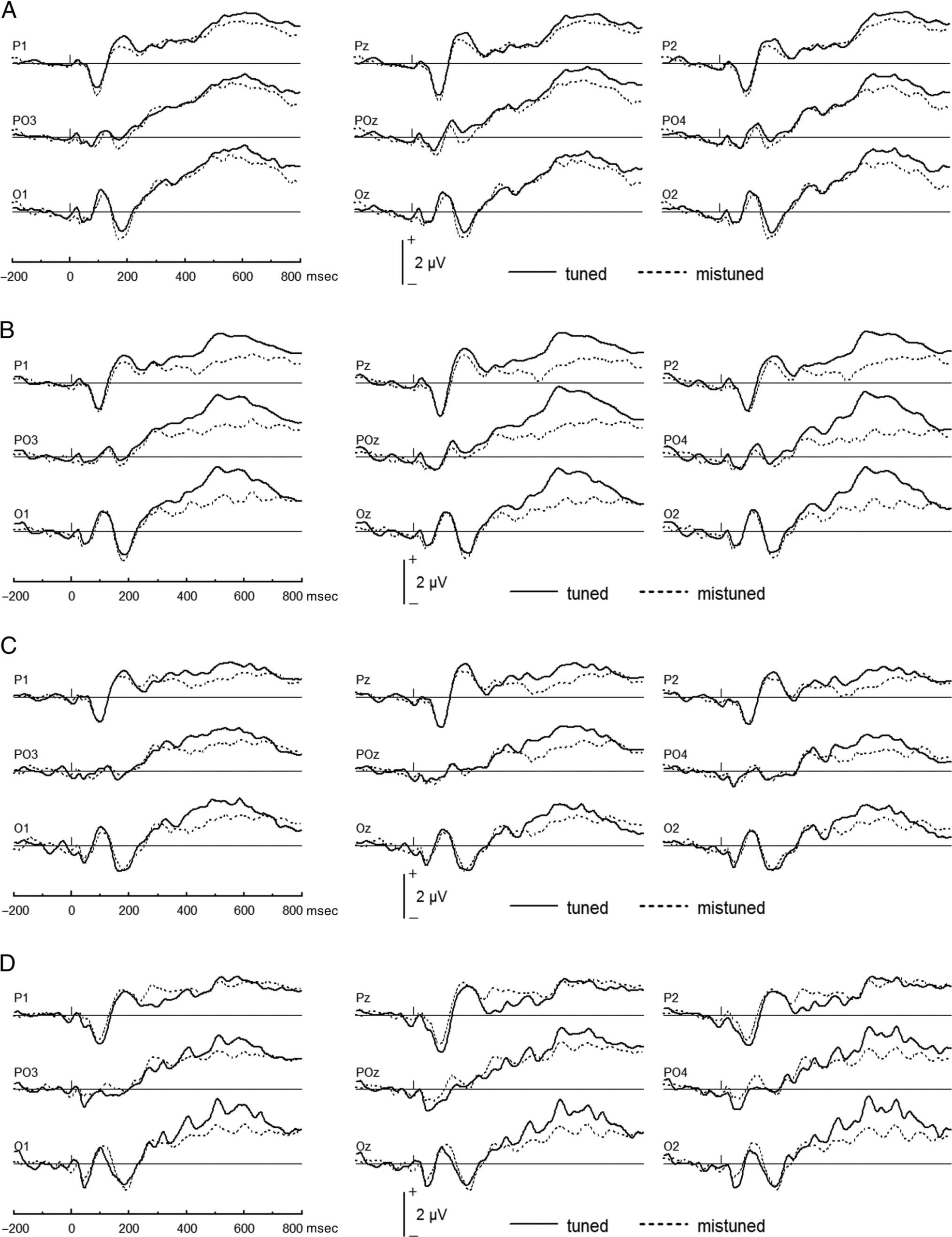
Figure 5. Plots of LPC at parietal-occipital scalp locations, which compare tuned with mistuned stimuli for hit (A), correct rejection (B), miss (C), and
false alarm (D).
2192
Journal of Cognitive Neuroscience
Volume 27, Number 11
F(1, 19) = 11.60, p = .003, which indicated that the
ORN was significantly smaller in correct than incorrect
responses.
ANOVA Results for the Encoding of Gap during
Gap Detection
To examine the interaction between successful gap de-
tection and harmonicity, a repeated-measures ANOVA
(Gap detection response [i.e., hit vs. miss] × Harmoni-
city [i.e., tuned vs. mistuned]) on gap-related activity
was performed. The interaction effect between Gap de-
tection response and Harmonicity was not significant,
F(1, 19) = .03, p = .871, which indicated that the mis-
tuning did not cause any significant changes in low-level
processing of the gap.
ANOVA Results for the LPC
The g roup mean LPC waveforms are shown in
Figures 5 and 6. Overall, the LPC was larger for tuned
than mistuned stimuli, F(1, 19) = 11.22, p < .0035, and
for correct than incorrect responses, F(1, 19) = 9.22, p =
.007. A three-way repeated-measures ANOVA (Harmoni-
city, Gap, and Response type) yielded significant inter-
actions between Gap and Response type, F(1, 19) = 7.31,
p = .015, and between Harmonicity, Gap, and Response
type, F(1, 19) = 4.46, p = .049. Further pairwise compari-
sons were performed between experimental conditions.
To assess the impact of concurrent sound perception
on attention, the LPC was compared between tuned and
mistuned stimuli for different gap detection responses.
For correct rejections, the LPC was greater for tuned than
mistuned stimuli, F(1, 19) = 35.30, p < .0001, whereas
for correct hits, the LPC was comparable for tuned
and mistuned stimuli, F(1, 19) = 1.10, p = .307
(Figure 5A and B). For misses, the LPC was greater for
tuned than mistuned stimuli, F(1, 19) = 4.95, p =
.0038, whereas for false alarms, the LPC amplitude was
comparable for tuned and mistuned stimuli, F(1, 19) =
.26, p = .616 (Figure 5C and D).
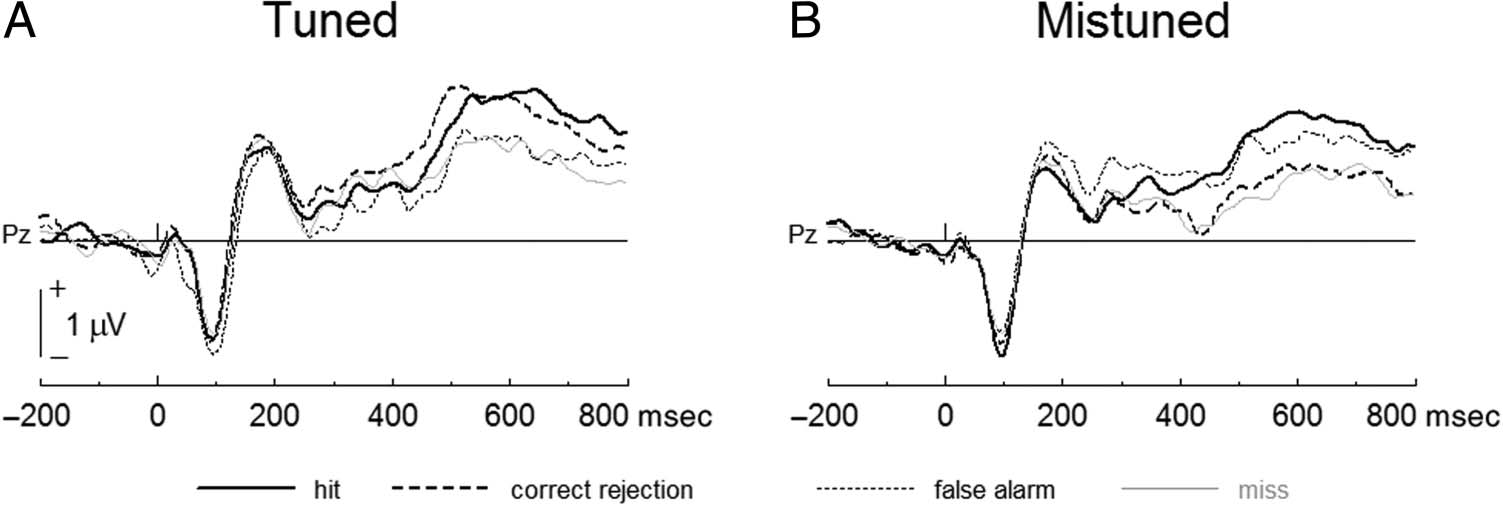
To address whether attention was related to successful
gap detection, the LPC was compared between gap
detection responses (i.e., hits vs. misses and correct re-
jections vs. false alarm) for tuned and mistuned stimuli.
For tuned stimuli, the LPC was greater for hits than mis-
ses, F(1, 19) = 6.99, p = .016, and comparable for false
alarms and correct rejections, F(1, 19) = 3.38, p = .082
(Figure 6A). For mistuned stimuli, the LPC was greater
for hits than misses, F(1, 19) = 8.39, p = .009, and great-
er for false alarms than correct rejections, F(1, 19) = 6.90,
p = .017 (Figure 6B). Additional significant results
showed that the LPC was greater for correct rejections
than misses for tuned stimuli, F(1, 19) = 31.20, p <
.0001, for hits than correct rejections, F(1, 19) = 9.67,
p = .006, and for false alarms than misses for mistuned
stimuli, F(1, 19) = 14.75, p < .001.
DISCUSSION
This study used auditory ERPs to determine whether gap
detection impairments were because of the failure of
low-level sensory encoding or the limitation of atten-
tional capacity. The behavioral results of the current
study are consistent with findings from a prior study by
Leung et al. (2011) in which gap detection performance
declined when mistuned harmonic stimuli were used.
The present results delineate whether the failure to de-
tect the gap is related to poor sensory encoding or taxed
attentional processing. Here, the analyses of sensory
evoked responses revealed comparable gap-related activ-
ity between tuned and mistuned harmonics during both
active and passive listening conditions. In contrast, the
analysis of attentional processes and response-related ac-
tivity revealed main effects of both harmonicity and re-
sponse accuracy as well as interaction effects among
harmonicity, gap, and response accuracy. We found a
marked decrease in the LPC amplitude when comparing
incorrect with correct responses. In addition, the ORN,
which indexes the perception of mistuned harmonics,
was greater for incorrect than correct responses. The
ORN has been proposed as a marker for the perception
of concurrent sound objects, that is, of mistuned har-
monics (e.g., Alain, 2007). A greater ORN for miss re-
sponses suggests that the perception of mistuned
harmonic interfered with the perception of the gap in
the sound complex. The results suggest that the presence
Figure 6. Comparing LPC
between the trial types, namely,
hit, correct rejection, miss, and
false alarm, in each of the tuned
(A) and mistuned (B) stimuli.
Only Pz was illustrated for
comparison as the waveform of
each trial type was the same as
Figure 5.
Leung, Jolicoeur, and Alain
2193
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
c
3
n
5
_
8
a
/
_
j
0
o
0
c
8
n
4
9
_
a
p
_
d
0
0
b
8
y
4
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
t
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
of a mistuned harmonic divided attention to more audi-
tory objects, which in turn reduced successful gap
detection.
In this study, the amplitude of the P2 wave was smaller
for gap than no-gap stimuli. The reduced P2 amplitude
could be accounted for by an N1 wave elicited by the on-
set of the gap that superimposed the P2 deflection gen-
erated by the earlier onset of the complex sound. A
similar decrease in P2 amplitude has been reported in an-
other study using similar gap durations (Heinrich et al.,
2004). Importantly, this gap-related activity is distinct
from the effects of mistuning as we found no interactions
between the two until response-related evoked poten-
tials. The fact that mistuning and gap effects do no inter-
act statistically in the ERP components observed during
the first 300 msec suggests that the lower detection rate
for gaps presented in mistuned stimuli results from post-
sensory-processing. We argue, therefore, that the reduced
gap detection accuracy reflects limitations of attentional pro-
cessing. Our findings could be explained by the object-
based account of auditory scene analysis, which posits
that attention operates on perceptual objects (Alain &
Arnott, 2000).
Neisser (1967) proposed an object-based account of vi-
sual attention in which perceptual analysis takes place in
two successive stages, a preattentive and a focal attention
stage. The former is based on Gestalt properties to sep-
arate objects within a complex scene (Duncan, 1984).
From this theory, the gap in this experiment was per-
ceived as an entity having shared properties with the
tuned harmonic sound, and the mistuned harmonic
formed a separate auditory object that likely interfered
with the perception of the tuned harmonic and gap. In
the focal attention stage, attention is devoted to analyze
a particular object in more detail (Neisser, 1967). How-
ever, previous studies have found that, even when a listen-
er is sure which object is the target, object selection can fail
when a competing object is inherently more salient, such
as a much louder sound, than the target (Conway, Cowan,
& Bunting, 2001). In our study, the most plausible explana-
tion for this impairment of gap detection lies on the focal
attention stage of the object-based theory. The mistuning
appeared to be perceptually more salient than the gap be-
cause it occurs throughout the entire duration of the
sound, whereas the gap only lasts a few milliseconds in
the middle of the sound and is embedded in only 1 of
the 10 tonal elements. Alain et al.’s (2001) study showed
that, by mistuning a tone by 16%, like the one applied in
this study, participants were very likely (over 95%) to per-
ceive two sounds. However, the saliency of a gap could be
affected by the frequency of the sound complex (Shailer &
Moore, 1983, 1987). Some studies have found that the sa-
liency of a gap in a tonal element could decrease because
of the frequency difference between tonal elements of a
complex sound (e.g., Oxenham, 2000). Therefore, the
mistuned harmonic could compete more successfully
for attention and prevent the gap from being detected.
One aspect of our ERP results supports this interpreta-
tion. Recall that we found a larger ORN on miss trials
compared with hit trials. The suggestion is that, for mis-
ses, attention was more likely to be biased toward the more
salient mistuned harmonic object, producing a larger ORN
and drawing attention away from the gap. This argument is
in line with the notion of biased competition, which
proposes that the competition among representations
happens before the volitional selection of objects as at-
tention is involuntarily drawn through the salience of ob-
jects (Desimone & Duncan, 1995). In this study, it appears
that the salience associated with the mistuned harmonic
often wins the competition for representation presumably
because the selective attention bias toward the gap is insuf-
ficient to override the interference from mistuning.
This study revealed differences in the LPC as a function
of mistuning and response accuracy. Previous studies
have shown that an increase in LPC amplitude typically
reflects greater attentional resource allocation to a stim-
ulus (Dien, Spencer, & Donchin, 2004; Solbakk, Reinvang,
& Nielsen, 2000). Our data are consistent with this inter-
pretation given that the LPC was larger during tuned than
mistuned trials. For tuned stimuli, attention was initially
allocated to the single tuned harmonic complex resulting
in a large LPC waveform when the gap was correctly de-
tected. In contrast, the presence of mistuning induces
concurrent sound perception, which in turn increases
the number of sound objects that compete for the limited
attentional resource. Such a competition for attention
among multiple auditory objects could account for the re-
duced LPC amplitude. This interpretation is consistent
with prior studies showing a reduced LPC under the influ-
ence of competitive interference between concurrent
stimuli (Martin, Jerger, & Mehta, 2007; Fischler & Bradley,
2006). This phenomenon is also consistent with Wicken’s
attentional resource model in which attention is divided
among mental operations, which impairs performance
task demands (Wickens, 1976, 2008).
The LPC is thought to reflect demands of attentional
resources for the successful performance of the task
(Starr & Don, 1988). Similarly, we found that the correct-
ly responded trials elicited larger LPC than incorrectly
responded trials for both the tuned and mistuned stimuli.
This is consistent with previous studies that have re-
ported positive relationships between behavioral
response and LPC amplitude in auditory discrimination
tasks (Xu et al., 2013).
Another consideration is that the gap is not only any
stimulus but also the most critical stimulus for the task.
Therefore, it is likely that attention was intentionally de-
ployed so as to extract the gap of the sound in an optimal
way. Prior studies have suggested that the LPC amplitude
represents the amount of information extracted from the
stimulus by the participants ( Johnson, 1986). This might
also be the case in this study as the LPC for hits was com-
parable between the tuned and mistuned stimuli, sug-
gesting that attention might have been deployed to
2194
Journal of Cognitive Neuroscience
Volume 27, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
c
3
n
5
_
8
a
/
_
j
0
o
0
c
8
n
4
9
_
a
p
_
d
0
0
b
8
y
4
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
extract the gap regardless of whether the distracting fea-
ture (i.e., the mistuning) was present.
situations in which attention is shared among fewer
objects.
One thing worth noting is that the gap was always em-
bedded in the tuned component of the harmonic complex
and never in the mistuned harmonic. The participants
might have adopted the strategy to keep their attention
focused on the tuned harmonic. Previous findings have
shown that the LPC amplitude was greater in conditions
where participants were asked to focus their attention to
a particular stimulus than in conditions where focused at-
tention to that particular stimulus was not required in ac-
tive listening tasks (Martin et al., 2007). To verify this
possibility, future studies may consider comparing LPC
amplitudes during gap detection, with the gap having
equal probability of being inserted in the tuned and mis-
tuned components of a sound complex.
Last but not the least, it is noted that the ORN occurs at
about the same latencies as the MMN component, which
peaks between 150 and 250 msec after deviant onset
(Näätänen, Paavilainen, Rinne, & Alho, 2007). The MMN
has been used as an index of automatic change detection
in the brain and is typically elicited by task-irrelevant
sounds that occur infrequently in an otherwise regular
stream of sounds. For example, it occurs when an incom-
ing stimulus differs from the memory representation
formed by the preceding stimulus sequence (Campbell,
2015; Näätänen et al., 2007). However, it is unlikely that
the ORN was affected by the MMN in this experiment.
One could imagine that a small MMN could occur after
the occurrence of an inharmonic sound that would have
followed a short series of repeated harmonic sounds.
However, the opposite would be equally likely, given that
the probability of presenting each type of sound was the
same. As such, we would expect no net overall MMN ef-
fect for either type of stimuli.
Conclusion
This study investigated whether the failure to detect gaps
during concurrent sound perception is related to sensory
encoding or attentional processing. During the first
300 msec, the ERP data showed comparable gap-related
activity across the tuned and mistuned groups of har-
monics. This suggests that the presence of a mistuned
harmonic does not significantly alter early sensory re-
sponses of gap detection. Moreover, it suggests that
the impaired gap detection is not likely because of the
failure of sensory encoding of the gap. In contrast, the
analysis of the late positive waves (LPC) revealed marked
differences as a function of mistuning and response.
These later effects suggest that the failure and success of
detecting a gap are influenced by attentional resources.
The results are consistent with the object-based account
of auditory scene analysis in which a listener’s attention
is shared among multiple objects, which can lead to the
reduction of attentional processing when compared with
Acknowledgments
This research was supported by grants from the Canadian Insti-
tutes of Health Research (MOP 106619) and the Natural Sci-
ences and Engineering Research Council of Canada.
Reprint requests should be sent to Claude Alain, Rotman Re-
search Institute, Baycrest Centre, 3560 Bathurst Street, Toronto,
Ontario, Canada M6A 2E1, or via e-mail: calain@research.
baycrest.org.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
c
3
n
5
_
8
a
/
_
j
0
o
0
c
8
n
4
9
_
a
p
_
d
0
0
b
8
y
4
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
t
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
REFERENCES
Alain, C. (2007). Breaking the wave: Effects of attention and
learning on concurrent sound perception. Hearing Research,
229, 225–236.
Alain, C., & Arnott, S. R. (2000). Selectively attending to auditory
objects. Frontiers in Bioscience, 5, D202–D212.
Alain, C., Arnott, S. R., & Picton, T. W. (2001). Bottom–up and
top–down influences on auditory scene analysis: Evidence
from event-related brain potentials. Journal of Experimental
Psychology: Human Perception and Performance, 27,
1072–1089.
Alain, C., & Izenberg, A. (2003). Effects of attentional load on
auditory scene analysis. Journal of Cognitive Neuroscience,
15, 1063–1073.
Alain, C., Schuler, B. M., & McDonald, K. L. (2002). Neural
activity associated with distinguishing concurrent auditory
objects. Journal of the Acoustical Society of America,
111, 990–995.
Backer, K. C., & Alain, C. (2014). Attention to memory:
Orienting attention to sound object representation.
Psychological Research, 78, 439–452.
Bregman, A. S. (1990). Auditory scene analysis: The perceptual
organization of sound. Cambridge, MA: MIT Press.
Campbell, K., & Macdonald, M. (2011). The effects of attention
and conscious state on the detection of gaps in long duration
auditory stimuli. Clinical Neurophysiology, 122, 738–747.
Campbell, T. A. (2015). A theory of attentional modulations
of the supratemporal generation of the auditory mismatch
negativity (MMN). Frontiers in Human Neuroscience,
9, e1065.
Conway, A. R., Cowan, N., & Bunting, M. F. (2001). The cocktail
party phenomenon revisited: The importance of working
memory capacity. Psychonomic Bulletin and Review,
8, 331–335.
Curran, T., Schacter, D. L., Johnson, M. K., & Spinks, R. (2001).
Brain potentials reflect behavioral differences in true and
false recognition. Journal of Cognitive Neuroscience,
13, 201–216.
Cusack, R., Carlyon, R. P., & Robertson, I. H. (2000). Neglect
between but not within auditory objects. Journal of
Cognitive Neuroscience, 12, 1056–1065.
Dell’Acqua, R., Dux, P. E., Wyble, B., Doro, M., Sessa, P.,
Meconi, F., et al. (2015). The attentional blink impairs
detection and delays encoding of visual information:
Evidence from human electrophysiology. Journal of
Cognitive Neuroscience, 27, 720–735.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of
selective visual attention. Annual Review of Neuroscience,
18, 193–222.
Dien, J., Spencer, K. M., & Donchin, E. (2004). Parsing the
late positive complex: Mental chronometry and the ERP
Leung, Jolicoeur, and Alain
2195
components that inhabit the neighborhood of the P300.
Psychophysiology, 41, 665–678.
Duncan, J. (1984). Selective attention and the organization of
visual information. Journal of Experimental Psychology:
General, 113, 501–517.
Dyson, B. J., & Alain, C. (2004). Representation of concurrent
acoustic objects in primary auditory cortex. Journal of the
Acoustical Society of America, 115, 280–288.
Dyson, B. J., Alain, C., & He, Y. (2005). Effects of visual
attentional load on low-level auditory scene analysis.
Cognitive, Affective & Behavioral Neuroscience,
5, 319–338.
Dyson, B. J., & Ishfag, F. (2008). Auditory memory can be object
based. Psychonomic Bulletin & Review, 15, 409–412.
Fischler, I., & Bradley, M. (2006). Event-related potential studies
of language and emotion: Words, phrases, and task effects.
Progress in Brain Research, 156, 185–203.
Hasko, S., Groth, K., Bruder, J., Bartling, J., & Schulte-Korne, G.
(2013). The time course of reading processes in children with
and without dyslexia: An ERP study. Frontiers in Human
Neuroscience, 7, 1–19.
Heinrich, A., Alain, C., & Schneider, B. A. (2004). Within- and
between-channel gap detection in the human auditory
cortex. NeuroReport, 15, 2051–2056.
auditory processing: A review. Clinical Neurophysiology,
118, 2544–2590.
Neisser, U. (1967). Cognitive psychology. New York: Appleton-
Century-Crofts.
Oxenham, A. J. (2000). Influence of spatial and temporal coding
on auditory gap detection. Journal of the Acoustical Society
of America, 107, 2215–2223.
Parasuraman, R., Richer, F., & Beatty, J. (1982). Detection and
recognition: Concurrent processes in perception. Perception
& Psychophysics, 31, 1–12.
Picton, T. W. (1992). The P300 wave of the human event-related
potential. Journal of Clinical Neurophysiology, 9, 456–479.
Picton, T. W., van Roon, P., Armilio, M. L., Berg, P., Ille, N.,
& Scherg, M. (2000). The correction of ocular artifacts: A
topographic perspective. Clinical Neurophysiology, 111,
53–65.
Recanzone, G. H., & Sutter, M. L. (2008). The biological basis
of audition. Annual Review of Psychology, 59, 119–142.
Ross, B., Schneider, B., Snyder, J. S., & Alain, C. (2010).
Biological markers of auditory gap detection in young,
middle-aged, and older adults. PLoS One, 5, e10101.
Shailer, M. J., & Moore, B. C. (1983). Gap detection as a
function of frequency, bandwidth, and level. Journal of
the Acoustical Society of America, 74, 467–473.
Hillyard, S. A., & Picton, T. W. (1978). On and off components
Shailer, M. J., & Moore, B. C. (1987). Gap detection and the
in the auditory evoked potential. Perception &
Psychophysics, 24, 391–398.
Johnson, R. (1986). A triarchic model of P300 amplitude.
Psychophysiology, 23, 367–384.
auditory filter: Phase effects using sinusoidal stimuli. Journal
of the Acoustical Society of America, 81, 1110–1117.
Shinn-Cunningham, B. G. (2008). Object-based auditory and
visual attention. Trends in Cognitive Sciences, 12, 182–186.
Leung, A. W., Jolicoeur, P., Vachon, F., & Alain, C. (2011).
Snyder, J. S., & Alain, C. (2007). Sequential auditory scene
The perception of concurrent sound objects in harmonic
complexes impairs gap detection. Journal of Experimental
Psychology: Human Perception and Performance, 37,
727–736.
Luck, S. J. (2014). An introduction to the event-related
potential technique. Cambridge, MA: MIT Press.
Macmillan, N. A., & Creelman, C. D. (1991). Detection theory:
A user’s guide. New York: Cambridge University Press.
Martin, B. A., Sigal, A., Kurtzberg, D., & Stapells, D. R. (1997).
The effects of decreased audibility produced by high-pass
noise masking on cortical event-related potentials to speech
sounds /ba/ and /da/. Journal of the Acoustical Society of
America, 101, 1585–1599.
Martin, J., Jerger, J., & Mehta, J. (2007). Divided-attention
and directed-attention listening modes in children with
dichotic deficits: An event-related potential study.
Journal of the American Academy of Audiology, 18,
34–53.
McDonald, K. L., & Alain, C. (2005). Contribution of harmonicity
and location to auditory object formation in free field:
Evidence from event-related brain potentials. Journal of the
Acoustical Society of America, 118, 1593–1604.
Näätänen, R., Paavilainen, P., Rinne, T., & Alho, K. (2007).
The mismatch negativity (MMN) in basic research of central
analysis is preserved in normal aging adults. Cerebral Cortex,
17, 501–512.
Solbakk, A. K., Reinvang, I., & Nielsen, C. S. (2000). ERP indices
of resource allocation difficulties in mild head injury.
Journal of Clinical and Experimental Neuropsychology,
22, 743–760.
Starr, A., & Don, M. (1988). Brain potentials evoked by
acoustic stimuli. In T. W. Picton (Ed.), Human
event-related potentials: EEG handbook (Vol. 3,
pp. 97–157). Amsterdam: Elsevier.
Wickens, C. D. (1976). The effects of divided attention on
information processing in manual tracking. Journal of
Experimental Psychology: Human Perception and
Performance, 2, 1–13.
Wickens, C. D. (2008). Multiple resources and mental workload.
Human Factors, 50, 449–455.
Wolk, D. A., Schacter, D. L., Lygizos, M., Sen, N. M., Holcomb,
P. J., Daffner, K. R., et al. (2006). ERP correlates of recognition
memory: Effects of retention interval and false alarms.
Brain Research, 1096, 148–162.
Xu, H., Zhang, D., Ouyang, M., & Hong, B. (2013). Employing
an active mental task to enhance the performance of
auditory attention-based brain-computer interfaces.
Clinical Neurophysiology, 124, 83–90.
2196
Journal of Cognitive Neuroscience
Volume 27, Number 11
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
1
2
1
7
/
2
1
1
1
8
/
6
2
1
1
9
8
4
6
9
/
6
1
3
3
8
1
o
3
c
3
n
5
_
8
a
/
_
j
0
o
0
c
8
n
4
9
_
a
p
_
d
0
0
b
8
y
4
g
9
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1