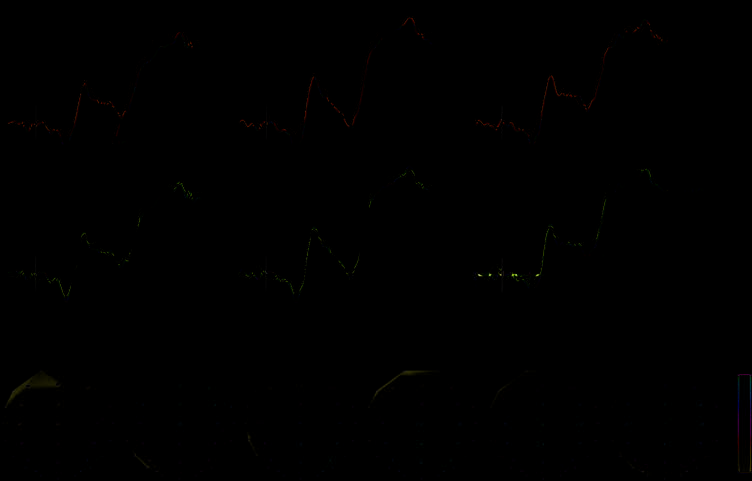Anticipating Conflict Facilitates Controlled
Stimulus-response Selection
A´ngel Correa1,2, Anling Rao2, and Anna C. Nobre2
Astratto
& Cognitive control can be triggered in reaction to previous
conflict, as suggested by the finding of sequential effects in
conflict tasks. Can control also be triggered proactively by
presenting cues predicting conflict (‘‘proactive control’’)?
We exploited the high temporal resolution of ERPs and con-
trolled for sequential effects to ask whether proactive con-
trol based on anticipating conflict modulates neural activity
related to cognitive control, as may be predicted from the
conflict-monitoring model. ERPs associated with conflict de-
tection (N2) were measured during a cued flanker task. Sym-
bolic cues were either informative or neutral with respect
to whether the target involved conflicting or congruent re-
sponsorizzato. Sequential effects were controlled by analyzing the
congruency of the previous trial. The results showed that cueing
conflict facilitated conflict resolution and reduced the N2 la-
tency. Other potentials (frontal N1 and P3) were also modu-
lated by cueing conflict. Cueing effects were most evident after
congruent than after incongruent trials. This interaction be-
tween cueing and sequential effects suggests neural overlap
between the control networks triggered by proactive and re-
active signals. This finding clarifies why previous neuroimaging
in which reactive sequential effects were not con-
studies,
trolled, have rarely found anticipatory effects upon conflict-
related activity. Finalmente, the high temporal resolution of ERPs
was critical to reveal a temporal modulation of conflict de-
tection by proactive control. This novel finding suggests that
anticipating conflict speeds up conflict detection and resolu-
zione. Recent research suggests that this anticipatory mecha-
nism may be mediated by preactivation of ACC during the
preparatory interval. &
INTRODUCTION
Most classic models of cognition posit a control system
that selects stimuli and responses according to task
goals (per esempio., Norman & Shallice, 1986; Kahneman, 1973;
Atkinson & Shiffrin, 1968). This system is engaged under
conflicting situations, such as when simultaneously oc-
curring events call for multiple and incompatible re-
sponsorizzato (Eriksen & Eriksen, 1974), and after negative
behavioral outcomes, such as errors (for a review, Vedere
Ridderinkhof, Ullsperger, Crone, & Nieuwenhuis, 2004).
After conflict or errors, behavioral measures reveal a
more deliberate mode of responding (‘‘sequential ef-
fects,’’ Gratton, Coles, & Donchin, 1992; ‘‘posterror slow-
ing,’’ Rabbitt, 1966).
Neural models of conflict monitoring (per esempio., Botvinick,
Braver, Barch, Carter, & Cohen, 2001) posit that the
ACC detects conflict and recruits DLPF cortex to bias
stimulus-response selection through top–down signals.
Neuroimaging research has confirmed increased ACC
activation by conflict, followed by decreased ACC activa-
tion on the subsequent conflicting trial, suggesting suc-
cessful conflict reduction by an increased attentional set
(Kerns et al., 2004; Botvinick, Nystrom, Fissell, Carter, &
Cohen, 1999).
1Universidad de Granada, Spain, 2University of Oxford, UK
Conflict monitoring is triggered in reaction to previous
conflict (‘‘reactive control’’), as suggested by the finding
of sequential effects in conflict tasks (Gratton et al., 1992).
Gratton et al. (1992) used a flanker task (Eriksen &
Eriksen, 1974) and found that the conflict effect (RT dif-
ference between incongruent and congruent conditions)
was reduced when the previous trial was incongruent
rather than congruent. Tuttavia, top–down control
È
not only triggered reactively but also proactively (Logan
& Zbrodoff, 1982). Logan and Zbrodoff (1982) induced
‘‘proactive control’’ by presenting cues predicting con-
flict in a Stroop (1935) task. The results showed that the
anticipation of an incongruent trial reduced the conflict
effect. Così, proactive control is analogous to attentional
orienting more generally, during which expectations
about a particular event attribute trigger top–down biases
of relevant stimulus/response representations (Kastner &
Ungerleider, 2000).
Although the conflict-monitoring model was primarily
based on reactive control (Botvinick et al., 2001), a clear
prediction can also be made for proactive control. Essere-
havioral facilitation of conflict resolution by proactive
control should be associated with modulations of conflict-
related neural activity (per esempio., decreased ACC activity). How-
ever, neuroimaging studies using conflict-predicting cues
have led to inconclusive results (Luks, Simpson, Dale, &
Hough, 2007; Sohn, Albert, Jung, Carter, & Anderson,
D 2008 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 21:8, pag. 1461–1472
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
8
e
–
1
P
4
D
6
F
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
C
1
N
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
9
e
.
S
T
2
o
1
N
1
3
0
6
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
/
T
.
F
.
o
N
1
8
M
UN
sì
2
0
2
1
2007; Fassbender, Foxe, & Garavan, 2006; Luks, Simpson,
Feiwell, & Mugnaio, 2002). The scarce evidence showing
that proactive control modulates conflict processing could
be explained by two factors: (1) the limited temporal res-
olution of hemodynamic measures or (2) the masking of
proactive control effects due to modulatory effects caused
by reactive control. None of these studies manipulating
proactive control considered the effect of previous trial
congruency. If proactive and reactive control share neu-
ral mechanisms, then the effects of proactive control
would be most evident when reactive control is low (cioè.,
after congruent rather than incongruent trials).
Here, we (1) exploited the high temporal resolution
of ERPs and (2) controlled for the effects of reactive
control in a cued flanker task to ask whether proactive
control modulates neural activity related to conflict.
Previous research using conflict tasks and ERPs has
identified two frontal potentials related to cognitive
controllo. A negative deflection is observed after stimuli
that carry incongruent versus congruent response ten-
dencies (N2; Kopp, Rist, & Mattler, 1996) as well as after
an incorrect response (error-related negativity—‘‘ERN’’
or ‘‘Ne’’; Gehring, Goss, Coles, Meyer, & Donchin, 1993;
Falkenstein, Hohnsbein, Hoormann, & Blanke, 1990).
ACC is thought to constitute an important source to
both the N2 and the ERN (Debener et al., 2005; van Veen
& Carter, 2002), Quale, rispettivamente, have been inter-
preted as indices of conflict detection (van Veen & Carter,
2002) and error detection (reviewed by Falkenstein,
Hoormann, Christ, & Hohnsbein, 2000).
The central hypothesis of this study was that the an-
ticipation of conflict should modulate control-related
potentials. Two ERP analyses were conducted on control-
related ERPs: stimulus locked and response locked. IL
stimulus-locked analysis tested whether conflict process-
ing, as indexed by the N2 potential, was attenuated by
cueing conflict. Additionally, this analysis allowed us to
explore whether proactive and reactive control operate
through a common attentional system, as suggested by
their analogous behavioral effects (Gratton et al., 1992;
Logan & Zbrodoff, 1982). We compared modulations
of stimulus-locked potentials by cueing versus by previous
conflict processing. If their neural mechanisms overlap,
proactive and reactive control should interact in their
modulation of conflict-related neural processing. Stron-
ger effects of proactive control should hence be ex-
pected under low versus high reactive control. On the
other hand, if proactive and reactive control involve in-
dependent mechanisms, we should observe dissociable
modulations of conflict processing.
The response-locked analysis tested whether error-
related processing indexed by the ERN and the ensu-
ing ‘‘error positivity’’ (Pe; Falkenstein et al., 1990, 2000)
was modulated by cueing conflict. High time pressure
for responding was imposed to ensure reliable numbers
of error trials for ERP analysis (per esempio., Nieuwenhuis et al.,
2006; Ullsperger, Bylsma, & Botvinick, 2005).
METHODS
Participants
Twenty participants from the University of Oxford (aged
19–49, nine women) took part voluntarily in the exper-
iment. Data from one participant were rejected due to
excessively low accuracy during task performance (58%
correct). The experimental methods were noninvasive
and had ethical approval from the University of Oxford.
Stimuli and Task
Participants completed a cued flanker task under high
time pressure. They were to respond according to the
direction pointed by a central arrow while ignoring the
direction of the flanking arrows. The target was pre-
ceded by a cue stimulus, which was either informative
or neutral with respect to the congruency between the
central and flanking arrows. Informative cues were 100%
predictive (‘‘cued’’ condition). A green checkmark indi-
cated that the central and the flanking arrows would be
congruent. A red cross indicated they would be incon-
gruent. A yellow question mark provided no prediction
about congruency (‘‘neutral’’ condition). A sufficiently
long interval was used between the cue and the flanker
array (1000–1500 msec) to enable the implementation
of endogenously generated expectations about the up-
coming stimulus congruency (per esempio., see Monsell, 2003;
Gratton et al., 1992; Mu¨ller & Rabbitt, 1989). The cue
stimulus subtended 1.28 (cid:1) 1.28. Target arrays consisted
of five white arrows (1.48 (cid:1) 18 each) appearing in a
row and centered on the fixation point (a gray dot with
0.38 diameter). The direction of the central arrow (left
or right) could be the same (congruent) or different
(incongruent) from that of the flankers. All flankers
pointed in the same direction.
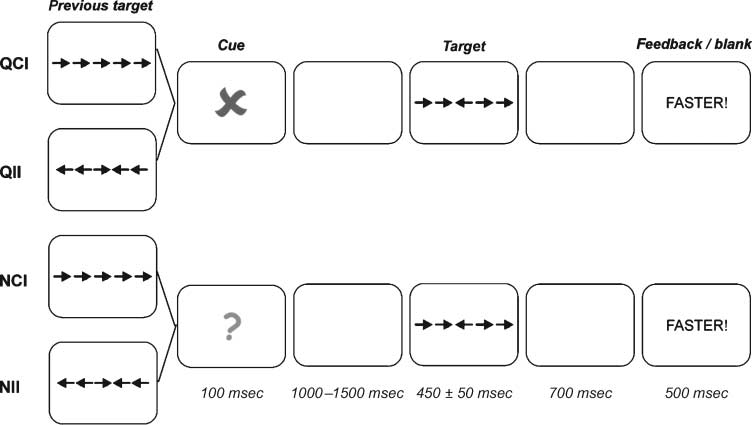
Figura 1 illustrates the main events of the task for the
four incongruent conditions used to compare proactive
cueing and reactive sequential effects. These included
cueing (cued and neutral) and congruency of the target
on the previous trial (‘‘previous congruency’’: congru-
ent and incongruent).
Each trial began with a fixation point presented cen-
trally for a random interval that ranged between 500
E 1000 msec. The cue was then presented centrally
for 100 msec and followed by a blank display for a ran-
dom interval of 1000–1500 msec. The target array then
appeared and remained until the participant responded
or a maximum deadline elapsed. The deadline was set
at 450 msec and was adjusted across blocks to main-
tain a good level of performance that was below ceiling
(around 70%). The deadline was increased by 50 msec
if the error rate was above 35% in the preceding block
or was decreased by 50 msec if it was below 25%. IL
participant had to press the ‘‘z’’ key with the left index
finger or the ‘‘m’’ key with the right index finger,
according to the direction pointed by the central arrow.
1462
Journal of Cognitive Neuroscience
Volume 21, Numero 8
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
8
e
–
1
P
4
D
6
F
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
C
1
N
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
9
e
.
S
T
2
o
1
N
1
3
0
6
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
F
/
.
T
.
o
N
1
8
M
UN
sì
2
0
2
1
Figura 1. The main events
of the task. The four
conditions included in the
stimulus-locked analysis are
displayed. This analysis
included current incongruent
targets with the factors of
cueing (cued and neutral)
and previous congruency
(congruent and incongruent).
QCI: cued, previous
congruent, and current
incongruent target; QII:
cued, previous incongruent,
and incongruent; NCI: neutro,
previous congruent, E
incongruent; NII: neutro,
previous incongruent, E
incongruent.
After a timely response or the deadline, there was a
blank display of 700 msec. If participants had failed to
respond within the deadline, a visual feedback ‘‘faster!’’
then appeared for 500 msec. Following a timely response
or the feedback, a blank display of 500 msec preceded the
next trial.
The experiment included three practice blocks plus
42 experimental blocks. Each block consisted of 16 trials,
including two trials of each type according to the type of
cueing (cued, neutro), flanker (congruent, incongruent),
and direction of the central arrow (left, right). The pro-
portion of congruent and incongruent targets was 0.5.
On average, half of current trials were preceded by con-
gruent or incongruent trials.
EEG Recording
The EEG recording was performed in an electrically
shielded room, using Ag/AgCl electrodes mounted on
an elastic cap and distributed along 34 scalp sites ac-
cording to the 10-20 International system (AEEGS, 1991)
using NuAmp amplifiers (Neuroscan, El Paso, TX). IL
montage included six midline sites (FZ, FCZ, CZ, CPZ,
PZ, and OZ) E 14 sites over each hemisphere (FP1/
FP2, F7/F8, F3/F4, FT7/FT8, FC3/FC4, T7/T8, C3/C4,
TP7/TP8, CP3/CP4, P7/P8, P3/P4, PO7/PO8, PO3/PO4,
and O1/O2). Additional electrodes were used as ground
and reference sites and for recording the EOG. All elec-
trodes were referenced to the right mastoid during the
recording and were algebraically rereferenced off-line
to calculate the average of the right and the left mas-
toids. Eye movements were monitored by horizontal and
vertical EOG bipolar recordings with electrodes placed
around the eyes. The EEG was amplified with a low-
pass filter of 300 Hz and digitized at a sampling rate of
1000 Hz.
ERP Analysis
The continuous EEG was filtered off-line with a 40-Hz
low-pass filter. Separate epochs were constructed for
targets (between (cid:2)100 E 600 msec relative to target
onset) and responses (between (cid:2)600 E 700 msec rel-
ative to response onset). Their respective baselines
consisted of a 100-ms epoch before target onset and a
50-ms epoch between (cid:2)100 E (cid:2)50 msec relative to
response onset. Epochs in which an eye blink or eye
movement occurred were rejected based on large de-
flections (±50 AV) in the HEOG or the VEOG elec-
trodes. Epochs with large signal drift were also removed
based on large deflections (±100 AV) in any channel.
An initial analysis compared ERPs triggered by con-
gruent versus incongruent targets to replicate the well-
established effects of conflict on the N2 potential. IL
analysis of main interest followed, which tested the ef-
fects of proactive control upon conflict processing and
compared them with the effects of reactive control. Ac-
cordingly, only ERPs elicited by the conflict-carrying in-
congruent target conditions were included. Epochs were
averaged according to the four conditions defined by
cueing (cued and neutral) and previous congruency (con-
gruent and incongruent). Only incongruent targets with
correct responses were analyzed. Identifiable target-
related potentials were analyzed at electrode locations
and temporal windows where they were most evident
according to the segmentation analysis performed with
Cartool software (see Topographical analyses section).
Target-locked waveforms averaged across participants are
shown in Figure 3 (top). The first negative deflection
peaked at 110 msec over frontal electrodes and was re-
ferred to as ‘‘frontal N1’’ (N1f ). According to the segmen-
tation analysis, the N1f occurred around 90–130 msec and
extended from frontal to centroparietal electrodes with
maximal intensity over the left frontal scalp (Figura 3,
Correa, Rao, and Nobre
1463
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
8
e
–
1
P
4
D
6
F
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
C
1
N
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
9
e
.
S
T
2
o
1
N
1
3
0
6
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
T
.
F
/
.
o
N
1
8
M
UN
sì
2
0
2
1
bottom). The N1f was therefore analyzed during 90–
130 msec over frontal to centroparietal electrode posi-
zioni (F3/Z/4, FC3/Z/4, C3/Z/4, and CP3/Z/4). The P1
overlapped temporally with the N1f, peaking at 120 msec,
and was maximal over occipital electrodes (O1/2, PO3/4,
and PO7/8). N1 peaked at 170 msec and was also maximal
over occipital electrodes (O1/2, PO3/4, and PO7/8). P1 and
N1 were analyzed during 110–130 and 160–180 msec over
three occipital positions (O1/2, PO3/4, and PO7/8). IL
N2 peaked at about 305 msec over frontal electrodes and
was analyzed during 290–330 msec at F3/Z/4 and FC3/Z/4.
The P3 peaked at 425 msec and was broadly distributed
over the central scalp. The P3 was analyzed during 360–
460 msec at FC3/Z/4, C3/Z/4, CP3/Z/4, and P3/Z/4.
An analysis of response-related ERPs focused on
whether cueing conflict modulated error processing. IL
epochs were averaged separately for errors and correct
responses during incongruent trials, according to the
cueing conditions (cued and neutral). Responses to con-
gruent targets led to insufficient numbers of observations
on the error condition and hence were not analyzed.
Likewise, conditions with previous congruent and incon-
gruent targets were collapsed to gain statistical power.
Error analyses followed the same approach as for tar-
gets, using periods defined by topographical segmenta-
tion and electrodes showing the maximal distribution
of potentials. Around the time of response, ERPs elic-
ited by errors contained an ERN that peaked at about
20 msec after the response (Guarda la figura 4, top)1 and was
focally distributed over frontocentral electrodes (Fig-
ure 4, bottom, Map 2). This was followed by positive-
going potentials, similar to the Pe, which separated into
two different topographical states (Maps 4 E 5). ERPs
elicited by correct responses were characterized by a
positive potential, the ‘‘correct positivity’’ (Pc), at the
same time range as the ERN, which was distributed over
central electrodes (Figura 4, bottom,
‘‘Map 3’’). IL
Pc was then followed by continued positive potentials
subdivided into two topographical states. The ERN was
analyzed at 0–40 msec over frontal and frontocentral
electrodes (F3/Z/4 and FC3/Z/4). The Pe was analyzed
over frontal to central electrodes (F3/Z/4, FC3/Z/4, E
C3/Z/4) at two epochs: early Pe (50–110 msec, Map 4)
and late Pe (250–290 msec, Map 5) (van Veen & Carter,
2002). Analyses of correct responses included the Pc
at 0–40 msec over frontal to central electrodes: F3/Z/4,
FC3/Z/4, and C3/Z/4 (see Map 3).
Mean amplitude values were obtained for the tempo-
ral window of each potential and submitted to repeated
measures ANOVAs. Target-related analyses consisted of
one ANOVA, which tested the effects of cueing, previous
congruency, electrode position, and electrode side.
Response-related analyses included separate ANOVAs for
error and correct conditions with the factors of cueing,
electrode position, and electrode side. Significant ef-
fects of electrode position and electrode side were not
reported, unless they involved interactions with either
cueing or previous congruency. Peak latencies of the main
potentials of interest (N2 and ERN) and potentials show-
ing amplitude modulation (N1f and P3) were also sub-
jected to ANOVAs. The latency analyses included the same
temporal windows as the mean amplitude analyses and
focused on the electrode showing the highest activity: F3
electrode for the N2, N1f, and ERN potentials and Cz for
the P3. The criterion to include data from a participant
in either target or response-related analyses consisted of
20 artifact-free trials per condition. This criterion led to
the exclusion of two participants from the target-related
analyses and three participants from the response-related
analyses.
Topographical Analysis
The topographical analysis determined how the distri-
bution of voltage over the scalp (topographical maps)
evolved over time in each experimental condition
and compared the distributions across conditions, using
Cartool software (D. Brunet, Geneva, Svizzera; http://
brainmapping.unige.ch/Cartool.php). The segmentation
consisted of a spatiotemporal cluster analysis of the
ERP group-averaged data normalized for global field
power (with the constraints that each scalp topography
should remain stable for at least 20 msec and that the
correlation between different topographies should not
exceed 90%). The choice of the optimal number of to-
pographies that best explained the whole data was based
on a cross-validation criterion (Pascual-Marqui, Michel,
& Lehmann, 1995). The output of this segmentation
analysis consists of a set of topographical maps that rep-
resent stable periods of electrical field patterns, Quale
reflect dissociable functional states of the brain (Vedere
Figura 4, center and bottom). Different maps reflect dif-
ferent stages of information processing with different un-
derlying brain sources (Lehmann, 1987). This procedure
served to guide the selection of the optimal temporal
windows and electrodes for the ERP analysis. Inoltre,
the segmentation analysis served to compare the topo-
graphical maps and their time courses across different
experimental conditions. The comparison across condi-
zioni (per esempio., Figura 4, Maps 2 E 3) was performed
statistically at the single-subject level through a fitting
procedure. The topographical maps identified for the
group-average were fitted to the scalp topography of
each participant and for each time point of data. During
this procedure, data from each subject and condition
were compared with the maps identified at the group-
average level, using a spatial correlation. Each time point
of the data of each participant was then labeled with the
map with which it had the highest spatial correlation.
For each participant, the number of milliseconds during
which one specific map was present on each condition
was computed. These values would indicate whether one
map was dominant over another map for a given con-
dizione. These values were compared across conditions
1464
Journal of Cognitive Neuroscience
Volume 21, Numero 8
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
8
e
–
1
P
4
D
6
F
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
C
1
N
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
P
C
D
N
.
B
sì
2
0
G
0
tu
9
e
.
S
T
2
o
1
N
1
3
0
6
8
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
F
T
.
.
/
o
N
1
8
M
UN
sì
2
0
2
1
with ANOVAs with the maps and the experimental con-
ditions as factors. An interaction involving the factor
map would indicate that a particular experimental condi-
tion was better explained by one map than another, E
therefore that a different configuration of brain genera-
tors better explains the results.
RESULTS
Behavioral Results
Error rates and mean RTs from 19 participants were
submitted to separate repeated measures ANOVAs with
the factors of cueing (cued and neutral), previous con-
gruency (congruent and incongruent), and congruency
(congruent and incongruent). Error rates constituted
the main dependent variable of interest because RTs
were highly constrained by our time-pressure procedure.
Tavolo 1 shows detailed data for all conditions.
The analysis of the error rates only considered re-
sponses given within the deadline (18% rejected). IL
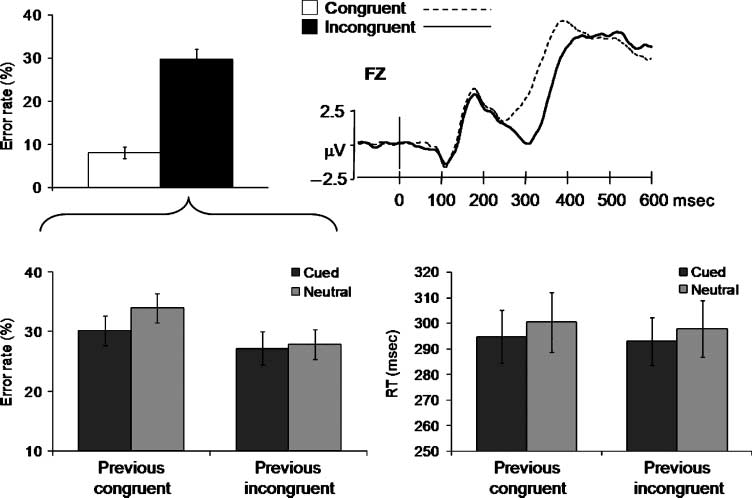
ANOVA showed a main effect of congruency (cioè., IL
conflict effect), F(1,18) = 80.07, P < .001, so that in-
congruent targets induced higher error rates than con-
gruent targets (Figure 2, top left).
A significant main effect of previous congruency,
F(1,18) = 6.19, p = .023, revealed that fewer errors
were committed after an incongruent trial (18%) than
after a congruent trial (20%). These two effects also
interacted significantly leading to sequential effects2
(previous congruency by congruency), F(1,18) = 8.25,
p = .01. Specifically, the conflict effect was smaller
when the previous trial was incongruent (20%) rather
than congruent (24%). Cueing also modulated the con-
flict effect (cueing by congruency), F(1,18) = 12.75, p =
.002, which was smaller on cued (19%) compared with
neutral (25%) conditions. Follow-up analysis of incon-
gruent targets showed that cueing conflict tended to re-
duce error rates, especially when the previous trial
was congruent, F(1,18) = 3.49, p = .078 (see Figure 2,
bottom left).
The RT analysis only considered responses that were
slower than 100 msec and within the deadline (14% re-
jected). The main effect of congruency was significant,
F(1,18) = 53.1, p < .001, revealing slower responses
for incongruent (296 msec) compared with congruent
(261 msec) trials. The main effect of cueing was also
significant, F(1,18) = 26.41, p < .001; responses were
faster in valid versus neutral targets. The interaction be-
tween previous congruency and congruency (sequen-
tial effects) was only marginally significant, F(1,18) =
2.92, p = .1, which may be due to the strong constraints
placed upon speeded responses. The interaction be-
tween cueing and congruency was significant, F(1,18) =
15.1, p = .001, showing larger cueing effects on congru-
ent versus incongruent trials (see also Gratton et al.,
1992, Experiment 3). Most relevant, follow-up analysis
of RTs in the incongruent condition revealed a signifi-
cant effect of cueing, with faster RTs in cued (294 msec)
versus neutral (299 msec) conflicting targets, F(1,18) =
5.21, p = .035.
Combined, the analyses of error rates and RTs show
that that the anticipation of conflict can speed up and
improve the performance during conflict resolution. Fig-
ure 2 (bottom) shows cueing benefits on both errors and
RT to incongruent targets. The neural correlates of this
attentional improvement in conflict resolution were stud-
ied by ERPs to the incongruent condition.
Electrophysiological Results
Target-locked ERPs
Figure 2 (top right) shows that congruent and incon-
gruent targets were processed differently. Consistent
with previous research (Kopp et al., 1996), the fronto-
central N2 was sensitive to different degrees of conflict
between congruent and incongruent conditions despite
being matched for stimulus frequency (50%). The con-
flict effect was evident as a larger and later N2 for in-
congruent compared with congruent targets, F(1,16) =
7.93, p = .01.3 This electrophysiological correlate of
conflict processing was consistent with the conflict ef-
fect observed on behavior. Once the conflict effect was
validated, the ERP analyses focused on incongruent tar-
gets yielding correct responses. These analyses tested
whether processing of a conflicting stimulus was mod-
ulated by cueing when sequential effects were controlled.
Table 1. Mean RTs and Error Rates from 19 Participants Broken Down by Cueing (Cued and Neutral), Previous Congruency
(Congruent and Incongruent), and Congruency (Congruent and Incongruent)
Cued
Neutral
Previous Congruent
Previous Incongruent
Previous Congruent
Previous Incongruent
Congruent
Incongruent
Congruent
Incongruent
Congruent
Incongruent
Congruent
Incongruent
252 (6.8)
295 (10.5)
253 (6.8)
293 (9.4)
267 (7.6)
300 (11.8)
271 (8.2)
298 (11)
11 (1.8)
30 (2.5)
10 (1.7)
27 (2.8)
6 (1.1)
34 (2.4)
6 (1.1)
28 (2.5)
Standard errors are shown in parentheses.
Correa, Rao, and Nobre
1465
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
8
e
-
1
p
4
d
6
f
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
c
1
n
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
t
/
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Top: Behavioral
(left) and electrophysiological
(right) correlates of the
conflict effect. Target-locked
ERP waveforms averaged
across 17 participants for
congruent and incongruent
trials. The frontal N2
potential was larger and
peaked later for incongruent
versus congruent conditions.
Bottom: Mean error rates
(left) and RTs (right) from
the incongruent condition
as a function of cueing (cued
and neutral) and previous
congruency (congruent and
incongruent). Vertical bars
represent standard errors.
The ANOVA performed on the mean amplitudes of the
N2 showed an interaction between cueing and electrode
side, F(2,32) = 3.57, p = .04, and a further three-way in-
teraction between cueing, previous congruency, and elec-
trode side, F(2,32) = 3.30, p = .05. Together, these effects
demonstrated that previous incongruent targets reliably
attenuated the N2 over right frontal electrodes (F4 and
FC4), F(1,16) = 4.32, p = .05, but only in the absence of
predictive cueing (i.e., on neutral conditions; see Figure 3,
NII vs. NCI). The N2 appeared to be attenuated by cueing
over left frontal electrodes, but this did not reach statis-
tical significance (F3 and FC3), F(1,16) = 1.37, p = .26.
Figure 3. Top: Stimulus-
locked ERP waveforms
averaged across 17 participants
showing the effects of cueing
(top row) and sequential
effects (bottom row) over
the frontal electrode sites
[F3 (left), FZ (midline), and
F4 (right)] where the N2
potential was largest. To
illustrate the proactive,
cueing effects, waveforms
elicited by cued incongruent
targets preceded by congruent
targets (QCI, thick blue line)
are compared with neutral
incongruent targets preceded
by congruent targets (NCI,
thin black line). To illustrate
the reactive, sequential
effects, waveforms elicited
by neutral incongruent targets
preceded by incongruent
targets (NII, thick purple line)
are compared with neutral
incongruent targets preceded by congruent targets (NCI, thin black line). The significant effects on the N1f and N2 potentials are labeled
where they were strongest. The P3 effects occurred more posteriorly and are not shown in the figure. Bottom: Topographical representation
of the voltage distribution over the scalp averaged during each segmentation map. Positive voltage is plotted in red, and negative voltage
is plotted in blue. Voltage was normalized during each period shown, according to the scale provided on the right. Yellow dots show the
electrode montage.
1466
Journal of Cognitive Neuroscience
Volume 21, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
8
e
-
1
p
4
d
6
f
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
c
1
n
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
/
.
t
o
n
1
8
M
a
y
2
0
2
1
Crucially, the analysis of the N2 latency revealed a re-
liable main effect of cueing, F(1,16) = 5.15, p = .04, such
that the N2 peaked earlier for cued (303 msec) com-
pared with neutral conditions (310 msec). Figure 3 (top)
shows that the N2 peak and the subsequent rising oc-
curred earlier for cued (QCI) versus neutral (NCI) con-
ditions.4 In contrast to cueing, previous congruency did
not modulate the N2 latency (F < 1).
The analysis of the early N1f revealed a significant at-
tenuation by cueing, F(1,16) = 4.42, p = .05. This main
effect was modulated by the congruency of the previous
trial (cueing by previous congruency), F(1,16) = 4.98, p =
.04. When the previous trial was congruent, the N1f was
clearly attenuated by the cued condition (QCI) as com-
pared with the neutral condition (NCI), F(1,16) = 8.27,
p = .01 (Figure 3). When the previous trial was incon-
gruent, the cueing effect was far from significant (F < 1).
In contrast to cueing, previous congruency did not exert
a main effect upon the N1f, F(1,16) < 1, in either cued
(QII vs. NCI), F(1,16) = 1.66, p = .2, or neutral con-
ditions (NII vs. NCI), F(1,16) = 1.08, p = .3. No sig-
nificant main effects of cueing, previous congruency, or
interactions were observed for the latency of the N1f
potential (all Fs < 1).
The P1 potential showed a significant interaction be-
tween cueing, previous congruency, and electrode po-
sition, F(2,32) = 3.81, p = .03. When effects were tested
at the separate electrode locations, however, the inter-
action only approached a trend over electrodes PO3/4,
F(1,16) = 2.83 p = .11, but remained clearly nonsignif-
icant over O1/2 and PO7/8 ( ps > .2). The N1 analysis
showed no significant effects of cueing or previous
congruency (Fs < 1) or interactions ( ps > .1).
The P3 analysis also showed an interaction between
cueing and previous congruency, F(1,16) = 9.20, p = .01.
When the previous trial was congruent, the P3 was en-
hanced by cued rather than neutral conditions, F(1,16) =
8.01, p = .01. When the previous trial was incongruent,
the cueing effect was far from significant (F < 1). Analo-
gously, previous congruency was only significant on neu-
tral conditions, F(1,16) = 7.53, p = .01, rather than on
cued conditions (F < 1). Both cueing and previous con-
gruency effects consisted of an enhancement on the P3
amplitude. Cueing also interacted with electrode position
and side, F(6,96) = 3.02, p = .01, revealing the largest
cueing effect over the central left electrode (C3), F(1,16) =
4.83, p = .04. The latency of the P3 potential was not
modulated by cueing, F(1,16) = 1.37, p = .26, or previous
congruency (F < 1).
Response-locked ERPs
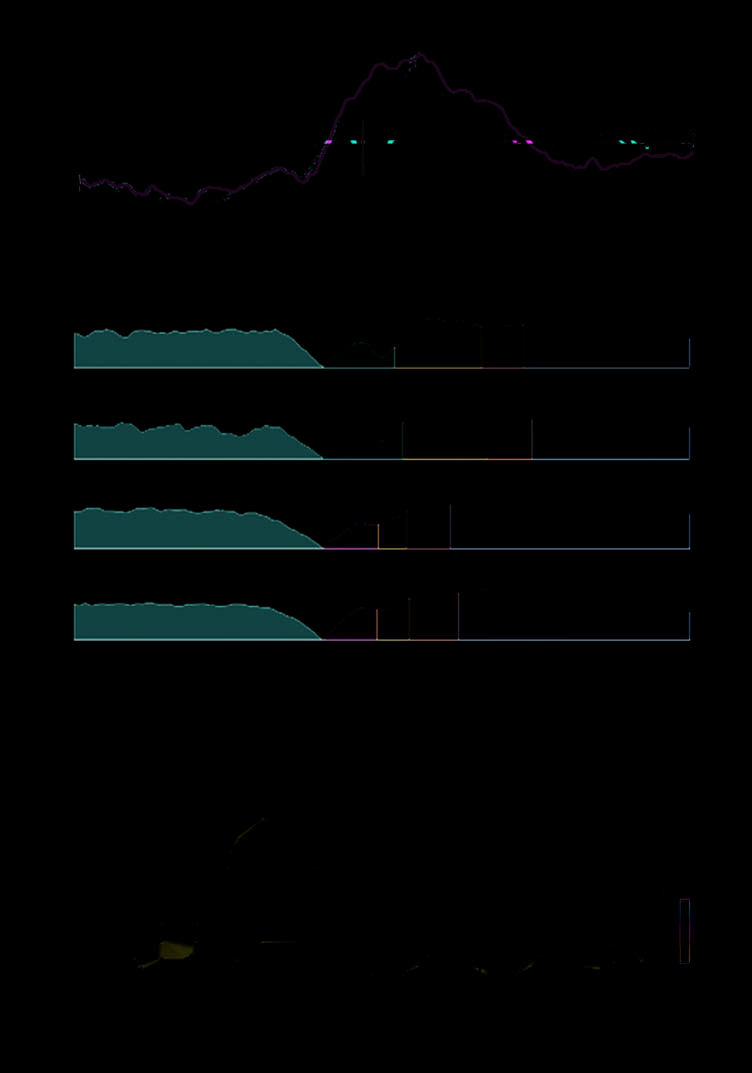
Figure 4 (top) shows that errors elicited an ERN peak-
ing shortly after the response (Map 2). The ANOVA
showed no effect of cueing on either ERN amplitude,
F(1,15) < 1, or latency, F(1,15) < 1. The ERN was fol-
lowed by two positive peaks (early and late Pe, Maps 4
and 5). The ANOVAs showed no effects of cueing on
the amplitudes of either early or late Pe, F(1,15) = 1.39,
p = .26 and F(1,15) < 1, respectively.
Correct responses were associated with the Pc po-
tential (Pc, Map 3), which was observed in the same
time range as the ERN on error conditions. The ampli-
tude of the Pc was enhanced by cueing, F(1,15) = 14.28,
p = .002. Following the Pc, the topographical seg-
mentation (Figure 4, center) suggested that waveforms
elicited by correct responses contained the same two pos-
itive potentials observed on error conditions but greatly
anticipated in time (100–150 msec earlier than in error
conditions).
Topographical Analysis of Response-locked ERPs
Topographical analyses were performed to test the fol-
lowing two observations based on the morphology of
the waveforms: (a) whether the ERN and the Pc had
different topographies associated to errors and correct
responses, respectively, and (b) whether the similari-
ties in the two positive peaks after the ERN and the
Pc for errors and correct responses, respectively, corre-
sponded to similar topographical distributions that were
offset in time.
Figure 4 (center) shows the time course of the seg-
mentation maps corresponding to stable voltage top-
ographies, colored on the global field power waveform,
for each of the four conditions. The progression of maps
was equivalent for all the conditions except during the
period of the ERN and the Pc (Maps 2 and 3). Map 2 was
only present on error conditions, whereas Map 3 was
only present on correct responses. The topographical
representation of these segments (Figure 4, bottom)
showed a frontal negativity that was specific to Map 2
but not Map 3, which was slightly lateralized to the left
hemisphere. In the group-averaged waveforms (Figure 4,
top), this negativity corresponds to the ERN potential
that was specifically observed on error conditions.
An ANOVA testing the preponderance of Maps 2 and
3 across conditions between (cid:2)100 and +60 msec con-
firmed the difference in distribution for error versus
correct trials. The interaction between response and to-
pographical map was significant, F(1,15) = 26.12, p <
.001. For errors, the ERN topography (Map 2) was signif-
icantly dominant over Map 3 (65% of the entire 160-ms
temporal window vs. 35%), t(15) = 2.1, p = .027, one-
tailed. In contrast, for correct responses, Map 3 was sig-
nificantly dominant over Map 2 (63% vs. 37%), t(15) =
1.77, p = .048, one-tailed.
The maps after the ERN map (Maps 4–6) followed iden-
tical sequences for both error and correct responses.
The main difference consisted of Map 4 (early Pe) being
present for a longer duration on errors than on correct
responses. This visual impression was tested by ANOVAs
Correa, Rao, and Nobre
1467
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
8
e
-
1
p
4
d
6
f
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
c
1
n
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
/
.
.
f
.
o
n
1
8
M
a
y
2
0
2
1
Figure 4. Top: Response-
locked ERP waveforms
averaged across 16 participants
for errors (red) and correct
responses (green) as a
function of cueing (cued, solid
line; neutral, dashed line).
ERPs recorded at frontocentral
electrodes are shown (left,
center, and right electrode
sides are collapsed in this
figure). The main error-related
potentials are labeled (PEe/PEl,
early/late Pe). On errors,
the ERN and the Pe did not
differ for cued versus neutral
conditions. On correct
responses, the first positive
potential was enhanced by
cueing. Center: Segmentation
maps of the group-averaged
data for the four conditions
during the interval between
(cid:2)600 and +700 msec (zero
indicates response execution,
keypressing). Map 2 was
present on errors, whereas
Map 3 was present on correct
responses. Maps 4, 5, and 6
followed the same sequence
for both errors and correct
responses. Map 4 was longer
after errors compared with
correct responses. Bottom:
Topographical representation
of the voltage distribution
over the scalp averaged across
each segmentation map. Map 2
shows the ERN distribution as
a negative voltage (blue) over
the frontal part of the head.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
8
e
-
1
p
4
d
6
f
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
c
1
n
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
t
/
.
f
o
n
1
8
M
a
y
2
0
2
1
testing the preponderance of Maps 4 and 5 for errors
and correct responses over three consecutive 80-ms
bins from 0 to 240 msec. The interaction between re-
sponse, topographical map, and time bin was significant,
F(2,30) = 7.58, p = .002. For errors, Map 4 was signifi-
cantly dominant over Map 5 during the first time bin
(77% vs. 23%), t(15) = 3.05, p = .004, one-tailed, and
the second bin (68% vs. 32%), t(15) = 2.04, p = .029,
one-tailed, but not at the third bin (56% vs. 44%), t(15) =
0.59, p = .28, one-tailed. In contrast, for correct re-
sponses, Map 4 was dominant over Map 5 during the first
time bin only (80% vs. 20%), t(15) = 3.91, p < .001, one-
tailed, but not at the second bin (60% vs. 40%), t(15) = 1,
p = .18, one-tailed. At the third bin, the pattern reversed
significantly, so that Map 5 was now dominant over Map 4
(75% vs. 25%), t(15) = 2.24, p = .02, one-tailed, suggest-
ing that the processing stage indexed by topographical
Map 4 was resolved at that time for correct responses. In
brief, Map 4 remained for longer after errors (0–160 msec)
than after correct responses (0–80 msec).
1468
Journal of Cognitive Neuroscience
Volume 21, Number 8
DISCUSSION
Modulation of Conflict Processing by Proactive
and Reactive Control
The analysis of behavioral performance confirmed that
both proactive and reactive control effectively facilitated
conflict resolution. This facilitation was also reflected in
ERP measures. The main finding showed that proactive
cueing altered the time course of conflict processing.
We expected proactive anticipation of conflict to en-
gage increased top–down control, hence to facilitate
conflict detection, as indexed by the N2 potential. The
high temporal resolution of ERPs was critical to un-
cover that proactive control enhanced on-line conflict
processing by reducing the latency of the N2. This
novel finding reveals the ability to prepare for conflict
based on predictive information. More generally, this
finding confirms that top–down attention is not only
constrained to cued physical attributes of forthcoming
stimuli and responses (for a review, see Nobre, 2004).
Rather, in the present study, cueing prompted the an-
ticipation of conflict and the need to engage the con-
trol system for cautious stimulus-response selection.
As a result, conflict was detected and resolved more
rapidly. The rapid detection of conflict was particularly
useful under the task parameters of the current exper-
iment, which imposed very high temporal pressure for
responding. When more deliberate processing is possi-
ble under normal conditions of time pressure (being
all the remaining task parameters identical to the cur-
rent experiment), the effects of proactive cueing of con-
flict may become less conspicuous (A. Correa, A. Rao,
J. Lupia´n˜ez, and A. C. Nobre, unpublished observations).
The temporal modulation of conflict detection by pro-
active control had remained unnoticed so far. Hemody-
namic neuroimaging studies have provided unclear results,
probably because the technique is highly insensitive to
short-lived or latency effects. During the anticipatory in-
terval, increments in ACC activation have been observed
in some studies (Sohn et al., 2007; Luks et al., 2002)
but not others (Luks et al., 2007; Fassbender et al., 2006;
MacDonald, Cohen, Stenger, & Carter, 2000). During
conflict processing, decrements in ACC activation selec-
tively related to cued conditions have been observed in
some studies (Luks et al., 2007; Sohn et al., 2007) but
not others (Fassbender et al., 2006; Luks et al., 2002).
The inconsistency of the fMRI results has then precluded
establishing strong conclusions about the role of ACC
in anticipatory conflict monitoring. Importantly, however,
the present ERP results strongly suggest that conflict pro-
cessing can be anticipated. As recently suggested, this
anticipatory mechanism may be mediated by preactiva-
tion of ACC during the preparatory interval (Sohn et al.,
2007).
An additional explanation for the absence of proac-
tive modulations upon ACC activity considers that the
previous studies cueing conflict did not control for se-
quential effects. It is possible that proactive cueing could
not attenuate ACC activity further when it was already
attenuated by reactive control. Therefore, this possibil-
ity assumes that proactive and reactive control involve
common or interacting neural mechanisms. To our knowl-
edge, the current study is the first to test this assumption
by comparing neural modulation resulting from both
types of control directly. The temporally rich dependent
variables provided by ERPs were able to identify particu-
lar stages during which proactive and reactive control
interact. From early through late neural target process-
ing, proactive control was most effective under low reac-
tive control. Specifically, the attenuation of the early N1f
by cueing was stronger after congruent versus incongruent
trials and the amplification of the late P3 was only signifi-
cant after congruent trials. Analogously, reactive control
was only effective in the absence of proactive control: N2
attenuation and P3 amplification by previous conflict were
only significant on neutral versus cued conditions. These
interactions suggest neural overlap or crosstalk between
the control networks triggered by proactive and reactive
signals.
The overlap between proactive and reactive control
was only partial, however: Each modulated conflict pro-
cessing with distinct time courses and spatial distribu-
tions. The anticipatory effects of proactive control upon
conflict processing started early (frontal N1 at 110 msec)5
and later became lateralized to the left hemisphere (P3).
In contrast, the effects of reactive control started later
(N2) and were lateralized to the right. Moreover, the N2
latency was selectively reduced by cueing, thus illustrating
the proactive nature of this type of control over more
reactive sequential effects. Overall, these findings would
suggest multiple rather than a single control mechanism,
although such differences could be specific to our exper-
imental procedure. For example, signals for reactive ver-
sus proactive control took place at different times, which
could have led to differential modulations of processing.
Further research will be useful to clarify whether neural
mechanisms subserving reactive and proactive control can
be more clearly dissociated.
Our results regarding reactive control are consistent
with neuroimaging research showing that increments
in ACC activity on incongruent conditions correlate with
subsequent reductions of ACC activity and behavioral
conflict (for a review, see Botvinick, Cohen, & Carter,
2004; Durston et al., 2003), even when repetition priming
effects are controlled (Notebaert, Gevers, Verbruggen, &
Liefooghe, 2006; Kerns et al., 2004; see also Ullsperger
et al., 2005; but see Nieuwenhuis et al., 2006; Mayr,
Awh, & Laurey, 2003). Previous ERP studies control-
ling for priming effects have provided mixed results. In
one case, the sequential modulation of conflict-related
processing survived (Scerif, Worden, Davidson, Seiger,
& Casey, 2006), but in another case it was abolished
(Wendt, Heldmann, Munte, & Kluwe, 2007). Our behav-
ioral analysis showed that priming did not account for
Correa, Rao, and Nobre
1469
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
8
e
-
1
p
4
d
6
f
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
c
1
n
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
.
.
t
.
f
o
n
1
8
M
a
y
2
0
2
1
the reductions in conflict effect exerted by either proac-
tive or reactive control. Nevertheless, because we had in-
sufficient trials to confirm our ERP results after controlling
for stimulus-response repetitions, these must be taken
with caution.
Electrophysiological Correlates of
Error Processing
Assuming that the ERN reflects ACC activity related to
response-related conflict (Botvinick et al., 2004) or error
detection (Debener et al., 2005), we hypothesized that
the anticipation of error in conflict trials would modu-
late the ERN. Unexpectedly, we observed no modulation
of the ERN. In light of the strong interactions between
cueing and sequential effects we observed in the analysis
of target-related processing, it is possible that modula-
tion of the ERN would be detectable in conditions of low
reactive control. However, due to an insufficient num-
ber of error trials in the different conditions of previous
congruency, this possibility could not be tested here.
The analysis of map topographies revealed that the
ERN was specific to trials with incorrect responses, rep-
licating earlier studies (Gehring et al., 1993; Falkenstein
et al., 1990). The absence of the typical ERN topogra-
phy on ‘‘correct’’ conditions replicates previous research
(e.g., Ullsperger, von Cramon, & Mu¨ller, 2002) but con-
trasts with reports of the so-called ‘‘correct response
negativity’’ (Bartholow et al., 2005; Falkenstein et al.,
2000; Vidal, Hasbroucq, Grapperon, & Bonnet, 2000).
The current topographical analysis showed that the
ERPs associated with errors and correct responses were
generated by differing configuration of neural sources
rather than differing only in the overall strength of their
activation.
Rather surprisingly, the topographical analysis further
revealed that, following the ERN, the stages of neural
processing in error trials were equivalent to those in
correct trials, although protracted in time. This find-
ing questions the specificity of the Pe potential to error
processing (Falkenstein et al., 1990, 2000). Our findings
suggest instead that a common late stage of postre-
sponse processing is achieved in both correct and error
trials (e.g., contextual updating), but that the conflict
and/or error detection delays the onset of this stage in
error trials.
In summary, proactive control based on the anticipa-
tion of conflict significantly modulated subsequent tar-
get processing and did so through neural mechanisms
that interacted with reactive control based on sequential
effects. The effects of proactive control were maximal
when controlled stimulus-response selection was not al-
ready engaged by reactive signals coming from conflict
in the previous trial. The present findings suggest that
anticipating conflict enhances cognitive control by speed-
ing up conflict detection and resolution.
Acknowledgments
This research was supported by the Spanish Ministerio de
Educacio´n y Cultura with postdoctoral grants (EX-2005-1028
and RYC-2007-00296) to A. C. and by an award from the James
S. McDonnell Foundation to A. C. N.
Reprint requests should be sent to A´ngel Correa, Departa-
mento de Psicologı´a Experimental y Fisiologı´a del Comporta-
miento, Campus Universitario de Cartuja s/n, 18011, Granada,
Spain, or via e-mail: act@ugr.es.
Notes
1. The use of a standard keyboard as response device may
have delayed (and jittered) the registration of the response
event, which might explain the unusually short apparent laten-
cies (and amplitudes) observed for the ERN.
2. Given the controversy on whether sequential effects re-
flect top–down conflict monitoring (Notebaert et al., 2006;
Ullsperger et al., 2005; Kerns et al., 2004) or bottom–up prim-
ing effects due to stimulus-response repetitions (Nieuwenhuis
et al., 2006; Mayr et al., 2003), the influence of priming was
tested by including ‘‘repetition’’ as a within-subjects factor
(e.g., Mayr et al., 2003). Repetition exerted no influence on
either the Cueing (cid:1) Congruency interaction (F < 1) or the
Previous congruency (cid:1) Congruency interaction, F(1,18) = 1.27,
p = .27. In light of this null effect and because of restricted trial
numbers, this variable was not analyzed further in the ERP
analyses.
3. Note that Figure 2 shows the N2 peaking at about 250 msec
for congruent targets, thus falling outside the time window
used in the analysis. In any case, the conflict effect was clearly
reliable for a broader time window of 240–300 msec including
both incongruent and congruent N2 peaks [mean amplitude
effect: F(1,16) = 11.05, p = .004].
It could be argued that this latency difference may have
4.
determined the mean amplitude results. A peak amplitude
analysis was performed on the N2, which replicated the mean
amplitude data. This analysis replicated the significant inter-
action between cueing and electrode side, F(2,32) = 4.06, p =
.027, and the three-way interaction between cueing, previous
congruency, and electrode side, F(2,32) = 3.19, p = .055,
which showed that sequential effects attenuated the N2 peak
amplitude over right frontal electrodes under neutral cueing
conditions (mean peak amplitudes: 1.33 AV for previous con-
gruent vs. 2.61 AV for previous incongruent), F(1,16) = 5.10,
p = .038.
5. The early N1f potential has not been specifically associated
with conflict processing. However, the attentional modula-
tion observed here is reminiscent of ERP studies on attention
and emotion, which reported attenuation of the anterior N1
when fearful faces were attended (e.g., Holmes, Vuilleumier,
& Eimer, 2003). Hence, it might be that our N1f was reflect-
ing emotional processing in reaction to conflict and/or the
prediction of error.
REFERENCES
AEEGS. (1991). American Electroencephalographic Society
(AEEGS) guidelines for standard electrode position
nomenclature. Journal of Clinical Neurophysiology,
8, 200–202.
Atkinson, R., & Shiffrin, R. (1968). Human memory: A
proposed system and its control processes. In K. Spence
& J. Spence (Eds.), The psychology of learning and
1470
Journal of Cognitive Neuroscience
Volume 21, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
8
e
-
1
p
4
d
6
f
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
c
1
n
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
f
t
/
.
o
n
1
8
M
a
y
2
0
2
1
motivation: Advances in research and theory (Vol. 2,
pp. 742–775). New York: Academic Press.
Bartholow, B. D., Pearson, M. A., Dickter, C. L., Sher, K. J.,
Fabiani, M., & Gratton, G. (2005). Strategic control and
medial frontal negativity: Beyond errors and response
conflict. Psychophysiology, 42, 33–42.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S.,
& Cohen, J. D. (2001). Conflict monitoring and cognitive
control. Psychological Review, 108, 624–652.
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004).
Conflict monitoring and anterior cingulate cortex:
An update. Trends in Cognitive Sciences, 8, 539–546.
Botvinick, M. M., Nystrom, L. E., Fissell, K., Carter, C. S.,
& Cohen, J. D. (1999). Conflict monitoring versus
selection-for-action in anterior cingulate cortex. Nature,
402, 179–181.
Debener, S., Ullsperger, M., Siegel, M., Fiehler, K., von
Cramon, D. Y., & Engel, A. K. (2005). Trial-by-trial
coupling of concurrent electroencephalogram and
functional magnetic resonance imaging identifies the
dynamics of performance monitoring. Journal of
Neuroscience, 25, 11730–11737.
Durston, S., Davidson, M. C., Thomas, K. M., Worden, M. S.,
Tottenham, N., Martinez, A., et al. (2003). Parametric
manipulation of conflict and response competition using
rapid mixed-trial event-related fMRI. Neuroimage, 20,
2135–2141.
Eriksen, B. A., & Eriksen, C. W. (1974). Effects of noise
letters upon the identification of a target letter in a
nonsearch task. Perception and Psychophysics, 16,
143–149.
Falkenstein, M., Hohnsbein, J., Hoormann, J., & Blanke, L.
(1990). Effects of errors in choice reaction tasks on the
ERP under focused and divided attention. In C. H. M. Brunia,
A. W. K. Gaillard, & A. Kok (Eds.), Psychophysiological
brain research 1990 (pp. 192–195). Tilburg: Tilburg
University Press.
Falkenstein, M., Hoormann, J., Christ, S., & Hohnsbein, J.
(2000). ERP components on reaction errors and their
functional significance: A tutorial. Biological Psychology,
51, 87–107.
Fassbender, C., Foxe, J. J., & Garavan, H. (2006). Mapping
the functional anatomy of task preparation: Priming
task-appropriate brain networks. Human Brain Mapping,
27, 819–827.
Gehring, W. J., Goss, B., Coles, M. G. H., Meyer, D. E., &
Donchin, E. (1993). A neural system for error detection
and compensation. Psychological Science, 4, 385–390.
Gratton, G., Coles, M. G., & Donchin, E. (1992). Optimizing
the use of information: Strategic control of activation of
responses. Journal of Experimental Psychology: General,
121, 480–506.
Holmes, A., Vuilleumier, P., & Eimer, M. (2003). The processing
of emotional facial expression is gated by spatial attention:
Evidence from event-related brain potentials. Cognitive
Brain Research, 16, 174–184.
Lehmann, D. (1987). Principles of spatial analyses. In
A. S. Gevins & A. Remond (Eds.), Handbook of
electroencephalography and clinical neurophysiology:
Methods of analysis of brain electrical and magnetic
signals (Vol. 1, pp. 309–354). Amsterdam: Elsevier.
Logan, G. D., & Zbrodoff, N. J. (1982). Constraints on
strategy construction in a speeded discrimination
task. Journal of Experimental Psychology: Human
Perception and Performance, 8, 502–520.
Luks, T. L., Simpson, G. V., Dale, C. L., & Hough, M. G.
(2007). Preparatory allocation of attention and
adjustments in conflict processing. Neuroimage, 35,
949–958.
Luks, T. L., Simpson, G. V., Feiwell, R. J., & Miller, W. L.
(2002). Evidence for anterior cingulate cortex involvement
in monitoring preparatory attentional set. Neuroimage,
17, 792–802.
MacDonald, A. W., Cohen, J. D., Stenger, V. A., & Carter, C. S.
(2000). Dissociating the role of the dorsolateral prefrontal
and anterior cingulate cortex in cognitive control. Science,
288, 1835–1838.
Mayr, U., Awh, E., & Laurey, P. (2003). Conflict adaptation
effects in the absence of executive control. Nature
Neuroscience, 6, 450–452.
Monsell, S. (2003). Task switching. Trends in Cognitive
Sciences, 7, 134–140.
Mu¨ller, H. J., & Rabbitt, P. M. (1989). Reflexive and voluntary
orienting of visual attention: Time course of activation
and resistance to interruption. Journal of Experimental
Psychology: Human Perception and Performance, 15,
315–330.
Nieuwenhuis, S., Stins, J. F., Posthuma, D., Polderman, T. J.,
Boomsma, D. I., & de Geus, E. J. (2006). Accounting
for sequential trial effects in the flanker task: Conflict
adaptation or associative priming? Memory and
Cognition, 34, 1260–1272.
Nobre, A. C. (2004). Probing the flexibility of attentional
orienting in the human brain. In M. I. Posner (Ed.),
Cognitive neuroscience of attention (pp. 157–179).
New York: Guilford Press.
Norman, D. A., & Shallice, T. (1986). Attention to action:
Willed and automatic control of behaviour. In R. J. Davidson,
G. E. Schwartz, & D. Shapiro (Eds.), Consciousness
and self-regulation: Advances in research and theory
(pp. 1–18). New York: Plenum Press.
Notebaert, W., Gevers, W., Verbruggen, F., & Liefooghe, B.
(2006). Top–down and bottom–up sequential modulations
of congruency effects. Psychonomic Bulletin and Review,
13, 112–117.
Pascual-Marqui, R. D., Michel, C. M., & Lehmann, D. (1995).
Segmentation of brain electrical activity into microstates:
Model estimation and validation. IEEE Transactions on
Biomedical Engineering, 42, 658–665.
Rabbitt, P. M. A. (1966). Errors and error correction in choice
reaction tasks. Journal of Experimental Psychology, 71,
264–272.
Kahneman, D. (1973). Attention and effort. London:
Ridderinkhof, K. R., Ullsperger, M., Crone, E. A., &
Prentice Hall.
Kastner, S., & Ungerleider, L. G. (2000). Mechanisms of
visual attention in the human cortex. Annual Review
of Neuroscience, 23, 315–341.
Kerns, J. G., Cohen, J. D., MacDonald, A. W., III, Cho, R. Y.,
Stenger, V. A., & Carter, C. S. (2004). Anterior cingulate
conflict monitoring and adjustments in control. Science,
303, 1023–1026.
Kopp, B., Rist, F., & Mattler, U. (1996). N200 in the flanker
task as a neurobehavioral tool for investigating executive
control. Psychophysiology, 33, 282–294.
Nieuwenhuis, S. (2004). The role of the medial frontal
cortex in cognitive control. Science, 306, 443–447.
Scerif, G., Worden, M. S., Davidson, M., Seiger, L., & Casey,
B. J. (2006). Context modulates early stimulus processing
when resolving stimulus-response conflict. Journal of
Cognitive Neuroscience, 18, 781–792.
Sohn, M. H., Albert, M. V., Jung, K., Carter, C. S., &
Anderson, J. R. (2007). Anticipation of conflict monitoring
in the anterior cingulate cortex and the prefrontal cortex.
Proceedings of the National Academy of Sciences, U.S.A.,
104, 10330–10334.
Correa, Rao, and Nobre
1471
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
8
e
-
1
p
4
d
6
f
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
c
1
n
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
f
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Stroop, J. R. (1935). Studies of interference in serial verbal
van Veen, V., & Carter, C. S. (2002). The timing of
reactions. Journal of Experimental Psychology, 18,
643–662.
Ullsperger, M., Bylsma, L. M., & Botvinick, M. M. (2005).
The conflict adaptation effect: It’s not just priming.
Cognitive, Affective, and Behavioral Neuroscience, 5,
467–472.
Ullsperger, M., von Cramon, D. Y., & Mu¨ller, N. G.
(2002). Interactions of focal cortical lesions with error
processing: Evidence from event-related brain potentials.
Neuropsychology, 16, 548–561.
action-monitoring processes in the anterior cingulate
cortex. Journal of Cognitive Neuroscience, 14, 593–602.
Vidal, F., Hasbroucq, T., Grapperon, J., & Bonnet, M. (2000).
Is the ‘‘error negativity’’ specific to errors? Biological
Psychology, 51, 109–128.
Wendt, M., Heldmann, M., Munte, T. F., & Kluwe, R. H.
(2007). Disentangling sequential effects of stimulus- and
response-related conflict and stimulus-response repetition
using brain potentials. Journal of Cognitive Neuroscience,
19, 1104–1112.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
8
e
-
1
p
4
d
6
f
1
/
1
2
9
1
3
/
7
8
8
/
2
1
5
4
o
6
c
1
n
/
1
2
0
7
0
6
9
0
2
2
5
1
2
1
3
/
6
j
o
p
c
d
n
.
b
y
2
0
g
0
u
9
e
.
s
t
2
o
1
n
1
3
0
6
8
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
.
t
.
/
o
n
1
8
M
a
y
2
0
2
1
1472
Journal of Cognitive Neuroscience
Volume 21, Number 8