Age Differences in Deactivation:
A Link to Cognitive Control?
Jonas Persson, Cindy Lustig, James K. Nelson,
and Patricia A. Reuter-Lorenz
Astratto
& The network of regions shown by functional imaging stud-
ies to be deactivated by experimental tasks relative to nomi-
nally more passive baselines (task < baseline) may reflect
processes engaged during the resting state or ‘‘default mode.’’
Deactivation may result when attention and resources are di-
verted from default-mode processes toward task processes.
Aging is associated with altered patterns of deactivation which
may be related to declining resources, difficulties with resource
allocation, or both. These possibilities predict that greater task
demand, which increases deactivation levels in younger adults,
should exacerbate age-related declines in allocating resources
away from the default mode. The present study investigated
the magnitude and temporal properties of deactivations in
young and older adults during tasks that varied in their de-
mand for cognitive control. Two versions of a verb generation
task that varied in their demand for selection among
competing alternatives were compared to word reading and
a fixation baseline condition. Consistent with our hypothesis,
greater deactivations were found with increasing demand.
Young and older adults showed equivalent deactivations in the
minimal selection condition. By contrast, age differences in
both the magnitude and time course of deactivation increased
with selection demand: Compared to young adults’, older
adults’ deactivation response showed less sensitivity to de-
mand. Demand-related changes in deactivation magnitude
correlated with performance changes, suggesting that indi-
vidual and group differences in deactivation have functional
significance. &
INTRODUCTION
Task-induced deactivations, or less activity during an
experimental task than during a passive baseline con-
dition, have become the target of much investigation
(Mazoyer et al., 2001; Binder et al., 1999; Shulman et al.,
1997). Such deactivations are generally thought to re-
flect a switch away from unconstrained ‘‘default-mode’’
processing in the passive condition (thus deactivating
regions supporting the default mode) to constrained
processing during the task (producing positive activation
[task > baseline] of regions that support task-related
processing) (Raichle et al., 2001). Older adults often have
difficulty with controlled processing, with some theories
emphasizing an age-related deficit in keeping thought
constrained to the relevant task (West, 1996; Hasher &
Zacks, 1988). Recent evidence suggests that both normal
aging (Grady, Springer, Hongwanishkul, McIntosh, &
Winocur, 2006; Lustig et al., 2003) and Alzheimer’s dis-
ease (Lustig et al., 2003) lead to reduced deactivations,
and that these responses may be delayed in pathological
aging (Rombouts, Goekoop, Stam, Barkhof, & Scheltens,
2005). The parameters of these effects, Tuttavia, are not
yet understood. Together, these possibilities suggest a
University of Michigan
potential relationship between age reductions in de-
activation magnitude and age-related declines in cogni-
tive control (per esempio., Braver & Barch, 2002). We investigate
this relationship by comparing the magnitude and time
course of deactivations in younger and older adults dur-
ing a verb generation task that varies cognitive control
by manipulating selection demands (Vedere, per esempio., Kan &
Thompson-Schill, 2004, for a review).
In young adults, deactivations are typically found in a
network of regions that includes the medial frontal,
medial and lateral parietal, and posterior cingulate cor-
tex (Mazoyer et al., 2001; Binder et al., 1999; Shulman
et al., 1997). The components of this network are
relatively stable over a wide range of tasks. This stability
suggests a coherent set of processes engaged during
baseline conditions (the default mode) that is not de-
pendent on the idiosyncracies of the particular experi-
mental task or procedure. Inoltre, spontaneous
fluctuations of activity (during relatively passive condi-
zioni) in the regions composing this network are tightly
correlated, and are anticorrelated with frontal (E
other) regions typically involved in active task process-
ing (Fox et al., 2005; Fransson, 2005; Greicius, Krasnow,
Reiss, & Menon, 2003; Raichle et al., 2001). Both the
stability of this network and its anticorrelation with
active-task regions buttress the idea that this network
D 2007 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 19:6, pag. 1021–1032
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
w
T
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
T
N
io
C
/
UN
e
–
R
P
T
D
io
C
1
l
9
e
6
–
1
P
0
D
2
F
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
C
/
N
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
C
2
N
1
.
P
2
D
0
0
B
7
sì
.
G
1
tu
9
e
.
S
6
T
.
o
1
N
0
0
2
8
1
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
.
.
F
.
.
.
o
N
1
8
M
UN
sì
2
0
2
1
supports default-mode processing from which partici-
pants switch away to focus on the active task. Further
supporting the idea that this network is involved in
default-mode processing, these regions, particularly the
medial parietal/posterior cingulate components, show
very high metabolism under rest conditions as measured
using fluorodeoxyglucose positron emission tomogra-
phy (FDG-PET; Phelps et al., 1981). Default-mode pro-
cesses have not been precisely characterized, but they
are thought to include attending to external environ-
mental stimuli, monitoring one’s own internal state and
emotion, and autobiographical/episodic memory pro-
cessazione (Gusnard, Akbudak, Shulman, & Raichle, 2001;
Raichle et al., 2001; Binder et al., 1999; Andreasen et al.,
1995). Deactivations may occur when the participant
switches from these unconstrained processes into the
specific processes demanded by the active task.
Although the components of the deactivation net-
work are stable across tasks, the magnitude of deactiva-
tion is responsive to task difficulty (McKiernan, Kaufman,
Kucera-Thompson, & Binder, 2003). Specifically, increas-
ing task difficulty by increasing factors such as stimulus
presentation rate or memory load results in greater de-
activation (greater distance from baseline) (McKiernan
et al., 2003). Inoltre, under equivalent difficulty
conditions, greater deactivations during learning are as-
sociated with successful encoding as demonstrated by
subsequent memory (Daselaar, Prince, & Cabeza, 2004).
These findings support the idea that deactivations re-
flect a reallocation of processing resources away from
default-mode processing and toward the demands of
the experimental task. Providing further support for this
idea, experimental manipulations that reduce the ability
to engage cognitive control result in reduced deactiva-
zioni (Choo, Lee, Venkatraman, Sheu, & Chee, 2005;
Chee & Choo, 2004).
Adult aging is thought to be associated with reduced
cognitive control (see Braver & Barch, 2002), which can
adversely affect the ability to constrain attention to a
relevant task (per esempio., Jennings & Jacoby, 1993; Hasher &
Zacks, 1988). Almost all functional neuroimaging studies
of aging and cognitive control have focused on age
differences in the positive activation of active-task re-
gions (see recent reviews by Rajah & D’Esposito, 2005;
Reuter-Lorenz & Lustig, 2005; Buckner, 2004; Hedden
& Gabrieli, 2004). Tuttavia, recent evidence suggests
that both healthy aging and dementia are also associ-
ated with altered deactivations of the default-mode net-
lavoro (Grady et al., 2006; Rombouts, Barkhof, Goekoop,
Stam, & Scheltens, 2005; Lustig et al., 2003). Are age
differences in deactivations linked to age differences in
cognitive control?
The present report addresses this question by ex-
amining the magnitude and time course of deactivations
during a task that manipulates the demand for control
by varying the requirement to select among competing
conceptual representations (Kan & Thompson-Schill,
2004). Participants completed a verb generation task,
during which they are instructed to generate an ap-
propriate verb in response to a visually presented noun
(Persson et al., 2004; Thompson-Schill, D’Esposito,
Aguirre, & Farah, 1997). Within the verb generation task,
two levels of selection demand were used. In the high-
selection condition, items were nouns with many appro-
priate associated responses (per esempio., BALL—THROW, KICK,
BOUNCE, eccetera.), but without a clear dominant response.
In the low-selection condition, items were nouns with
one clear dominant response, or a few associated re-
sponsorizzato (per esempio., SCISSORS—CUT). The high-selection
condition is presumed to place greater demands on
processes involved in resolving competition between
multiple possible alternatives to determine which will
ultimately be given as a response. This results in an
‘‘interference effect,’’ or longer reaction times (RTs) In
the high-selection condition than in the low-selection
condition. In addition to the rest baseline, a read con-
dition was used as an active-task condition with minimal
selection demands. In a previous report based on a
subset of the same subjects performing the same gen-
eration task, we documented age-related differences
in positive (task > baseline) activation (Persson et al.,
2004). In particular, older adults showed less activation
in the left inferior frontal gyrus (IFG) coupled with more
activation in the right IFG compared to young adults,
possibly reflecting compensation.
In light of recent findings, we investigated deactiva-
tion within the same dataset (plus several additional
participants), and we hypothesized that deactivation
magnitudes should be larger in the high-selection de-
mand conditions than in the low-demand conditions (cf.
McKiernan et al., 2003). Also, if age differences in de-
activation reflect age differences in the ability to reallo-
cate resources, age differences in deactivation should be
most evident in the high-demand conditions. We further
tested the idea that deactivation is related to the reallo-
cation of processing resources by correlating deactiva-
tion magnitude with activation in frontal regions, E
with behavioral performance. Finalmente, exploratory analy-
ses characterized the time course of deactivations as a
function of region, age, and selection demand.
METHODS
Participants
Thirty-two young adults (15 men; age range: 18–30 years)
E 28 senior adults (14 men; age range: 60–81 years)
were recruited from local newspapers and posted adver-
tisements, and completed a self-report health screening
and neuropsychological tests. All participants were right-
handed native English speakers who reported no existing
neurological or psychiatric illness. Vision was normal or
corrected to near normal using MRI-compatible glasses
or contact lenses. None of the participants reported
1022
Journal of Cognitive Neuroscience
Volume 19, Numero 6
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
w
T
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
T
N
io
C
/
UN
e
–
R
P
T
D
io
C
1
l
9
e
6
–
1
P
0
D
2
F
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
C
/
N
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
C
2
N
1
.
P
2
D
0
0
B
7
sì
.
G
1
tu
9
e
.
S
6
T
.
o
1
N
0
0
2
8
1
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
.
.
.
F
.
.
o
N
1
8
M
UN
sì
2
0
2
1
medical conditions (per esempio., high blood pressure) or medi-
cations that could affect blood-oxygen levels. Seniors’
scores on the neuropsychological tests were in the
normal range for this age group. Senior participants were
excluded if they scored below 25 on the Mini-Mental State
Examination (MMSE; Folstein, Folstein, & McHugh,
1975). None of the participants that attended the pre-
scanning session were excluded based on this criterion.
Although one senior participant did score a 25 on the
MMSE, the scores were overall high (mean = 28.6, SD =
1.29). All participants were paid $10–20 per hour for the
experiment, and senior participants were additionally
paid for their travel time to campus.
Behavioral Methods
The study consisted of two separate sessions: neuro-
psychological pretesting (for senior adults only) E
fMRI scanning with neuropsychological posttesting. Neu-
ropsychological testing included standardized tests such
as the MMSE, the Wisconsin Card Sorting Test (WCST;
Heaton et al., 1993), and the California Verbal Learning
Test (CVLT; Delis, Kramer, Kaplan, & Ober, 1987) (Vedere
Tavolo 1). The tasks used in the scanning session required
participants either to generate a verb related to a visually
presented noun or to simply read the noun. For each of
the nouns, the participants responded by pressing a
button with their right hand after they had covertly
generated a single verb response in the ‘‘generate task,’’
or after they had read the noun in the ‘‘read task.’’ The
nouns in each of the conditions (MANY, FEW, READ)
Tavolo 1. Mean Scores and Standard Deviation (SD) for the
Demographic and Neuropsychological Data
Demographics
N
Age
Young
Senior
32
28
21.7 (2.5)
(18–30 years)
68.1 (5.8)
(60–81 years)
Gender (M:F)
Educationa (years)
15:17
12:16
3.0 (1.4)
3.1 (2.6)
Neuropsychological
Fluency
Semantic
23.3 (5.2)
18.0 (4.9)
First-letter cue
47.7 (10.9)
43.4 (12.2)
Vocabulary
53.5 (5.5)
Letter–number sequencing
14.4 (2.3)
MMSEb
–
54.9 (7.7)
11.2 (2.1)
28.6 (1.3)
aNumber of years after high school.
bMMSE collected as a screening criterion for senior (min = 25).
were presented only once. Due to the nature of the
task, the words used in the three conditions were differ-
ent, but were of similar length (3–8 letters) and frequency
(Kucera-Francis range from 0 A 591).
The high- and low-selection conditions were blocked,
and participants were not informed about this selection
manipulation. They also completed a low-level baseline
condition in which participants gazed at a central fixation
cross. Four nouns were presented in each 16-sec block
(except for REST), with each word presented for 4 sec.
The study was divided into two runs, each with eight al-
ternating sets of MANY, FEW, and READ blocks (24 blocks
total), as well as four baseline blocks. Before each task
block began, an instruction was placed on the screen for
2 sec (cioè., ‘‘GENERATE’’ or ‘‘READ’’). The order of the
blocks was counterbalanced. Before noun presentation, UN
small letter ‘‘G’’ or ‘‘R’’ was displayed for 500 msec in the
center of the screen to remind participants of the task at
hand. Further details of the behavioral methods have
been reported elsewhere (Persson et al., 2004).
In all behavioral analyses, an effect was considered
significant if it reached a threshold of p < .05.
fMRI Methods
Images were acquired using a 3-T whole-body MRI
scanner (General Electric) equipped with a standard
quadrature head coil. Functional T2* blood oxygenation
level-dependent (BOLD) images were acquired using
a spiral sequence with 25 contiguous axial 5-mm slices
(repetition time [TR] = 1500 msec, echo time [TE] =
25 msec, flip angle = 908, and a field of view [FOV] =
24 cm). A T1-weighted gradient-echo (GRE) anatomical
image was also acquired by using the same parameters
and slices as were used in the functional scans (TR =
275 msec, TE = 35 msec, and flip angle 908). In addition,
a 60-slice high-resolution set of anatomical images was
acquired by using spoiled gradient-recalled acquisition
in steady state (SPGR) imaging (TR = 35 msec, TE =
3 msec, flip angle = 358, and FOV = 24 cm, 2.5 mm slice
thickness, 3.75 (cid:1) 3.75-mm in-plane resolution). Experi-
mental tasks were presented using E-Prime (Psychology
Software Tools, Pittsburgh, PA) and the IFIS 9.0 system
(MRI Devices, Waukesha, WI), and responses were col-
lected using two 5-button glove-like response pads.
Subsequent preprocessing and analyses were done us-
ing SPM99 (Wellcome Department of Cognitive Neurol-
ogy, London, UK). Further details of the methods for
the fMRI session have been reported elsewhere (Persson
et al., 2004).
All conditions (high, low, and read) were modeled as a
fixed response (box-car) waveform convolved with the
hemodynamic response function. Statistical parametric
maps were generated using t statistics to identify re-
gions deactivated according to the model. Group data
were then analyzed using a random-effects model. All re-
ported overall deactivations passed a threshold corrected
Persson et al.
1023
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
9
e
6
-
1
p
0
d
2
f
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
c
/
n
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
c
2
n
1
.
p
2
d
0
0
b
7
y
.
g
1
u
9
e
.
s
6
t
.
o
1
n
0
0
2
8
1
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
f
.
.
.
o
n
1
8
M
a
y
2
0
2
1
for multiple comparisons of p < .01, and deactivation in
the group comparisons (young vs. old) passed an uncor-
rected threshold of p < .005. Regions of interests were
functionally defined on the voxels that showed peak
deactivations in a comparison of the combined selection
conditions (HIGH and LOW) versus resting (fixation)
baseline in the current dataset, and that corresponded
to typical deactivation/default-mode regions in the litera-
ture. Each region was created by including activated
voxels ( p < .01, corrected) within a 10-mm sphere
around the peak voxel corresponding to default-mode
regions reported in the literature. Each ROI contained a
minimum of 40 contiguous voxels. Peak coordinates are
presented in Table 2 (coordinates in bold are those used
as seeds for the ROIs) and corresponding regions in
Figure 3. They included the medial and lateral parietal
cortex (LPC), the medial frontal cortex, and medial-
temporal regions. For each ROI, effect sizes (% signal
change) for the different conditions were then extracted
for each age group separately. The effect sizes represent
an average of the time points across the task blocks
versus the average across the fixation blocks.
RESULTS
Behavioral Data
Reaction time data were collected for the three behav-
ioral tasks (high, low, and read) during scanning (Fig-
ure 1). Four participants (2 seniors) were excluded from
the analysis due to technical problems. An analysis of
variance (ANOVA) of these data, including age group as
a between-subjects variable and task condition as a
within-subjects variable, indicated that the senior adults’
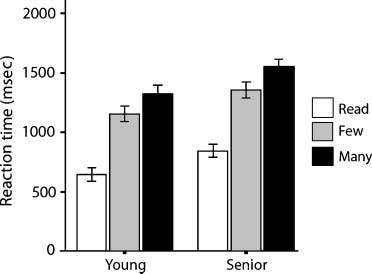
Figure 1. Mean RTs as a function of age group and task condition
based on median RTs for each participant. Error bars represent
standard error of the mean.
responses were slower than those of younger adults
[F(1, 54) = 6.20, p < .05]. The task condition main effect
was also significant [F(2, 54) = 328, p < .001]. Paired
comparisons of the task conditions indicated that re-
sponse times for high and low selection were signifi-
cantly different [t(55) = 14.7, p < .001], as were the
differences between low selection and read [t(55) =
16.7, p < .001], and high selection and read [t(55) =
20.4, p < .001]. The Age by Task interaction was not
significant [F(2, 54) = 0.16, p = .85].
For the neuropsychological tests, senior participants
performed significantly worse on the semantic fluency
test [t(56) = 3.99, p < .001] and letter–number se-
quencing [t(45) = 5.08, p < .001]. Performance was
equivalent for senior and young adults in WAIS-III Vo-
cabulary [t(55) = 0.81, p = .42] and first-letter fluency
[t(56) = 1.42, p = .16]. Education level was also equiv-
alent [t(51) = 0.58, p = .89].
Table 2. Talairach Coordinates for Areas that Show Maximal
Deactivation ( p < .01, Corrected for Multiple Comparisons)
fMRI Results
Anatomical Localization
BA
x
y
z
Z
R precuneus
R lateral parietal
L precuneus
L precuneus
R precuneus
L posterior cingulate
L superior temporal
7
39
30
31
31
31
22
11 (cid:2)57
30 14.70
49 (cid:2)61
17 14.30
(cid:2)15 (cid:2)68
26 13.31
(cid:2)11 (cid:2)52
11 (cid:2)52
(cid:2)15 (cid:2)35
49
49
39
12.10
12.07
11.49
(cid:2)41 (cid:2)22
0
10.95
R superior frontal gyrus
6./8
22
22
55
9.01
R medial temporal
30 (cid:2)45 (cid:2)5
7.85
L inferior parietal
40
(cid:2)59 (cid:2)28
R medial frontal
9./10
11
49
29
21
7.47
7.14
L = left; R = right; BA = Brodmann’s area; x, y, z = stereotactic
coordinates. The regions in bold were selected for ROI analyses.
To investigate deactivation related to task selection
based on the two age groups combined, we contrasted
the rest baseline with the verb generation conditions
(high and low selection combined). The results from
this contrast are presented in Figure 2 and Table 2.
Consistent with previous findings (e.g., McKiernan et al.,
2003; Mazoyer et al., 2001), task-induced deactivations
were found in multiple cortical regions including a me-
dial prefrontal region (PFC; Brodmann’s area [BA] 9/10),
bilateral medial parietal regions (posterior cingulate
cortex [PCC]; BA 30/31), the precuneus (BA 7), and a
right lateral temporo-parietal region (BA 39). Consistent
with some previous findings (e.g., Greicius, Srivastava,
Reiss, & Menon, 2004), we also found deactivation in
a region of the medial-temporal lobe (MTL). Based on
this whole-brain analysis (baseline > verb generate),
five regions were functionally determined using Marsbar
(http://marsbar.sourceforge.net). These regions have
typically been associated with task-induced deactivations
1024
Journal of Cognitive Neuroscience
Volume 19, Numero 6
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
w
T
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
T
N
io
C
/
UN
e
–
R
P
T
D
io
C
1
l
9
e
6
–
1
P
0
D
2
F
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
C
/
N
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
C
2
N
1
.
P
2
D
0
0
B
7
sì
.
G
1
tu
9
e
.
S
6
T
.
o
1
N
0
0
2
8
1
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
.
.
.
F
.
.
o
N
1
8
M
UN
sì
2
0
2
1
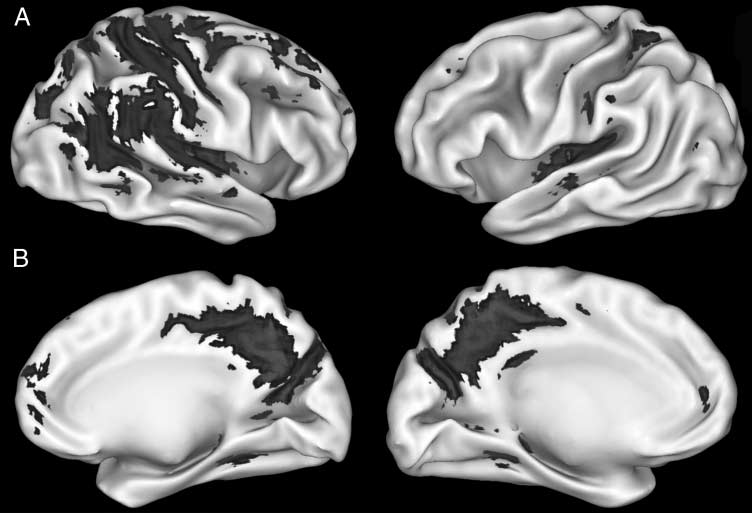
Figura 2. Statistical activation
map for deactivations
(baseline > verb generation;
corrected threshold at p < .01)
across all subjects.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
9
e
6
-
1
p
0
d
2
f
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
c
/
n
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
c
2
n
1
.
p
2
d
0
0
b
7
y
.
g
1
u
9
e
.
s
6
t
.
o
1
n
0
0
2
8
1
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
f
.
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
(Mazoyer et al., 2001; Binder et al., 1999; Shulman et al.,
1997), and were selected for additional ROI analyses.
For all subsequent ROI analyses, we focused on these
five regions (Figure 3).
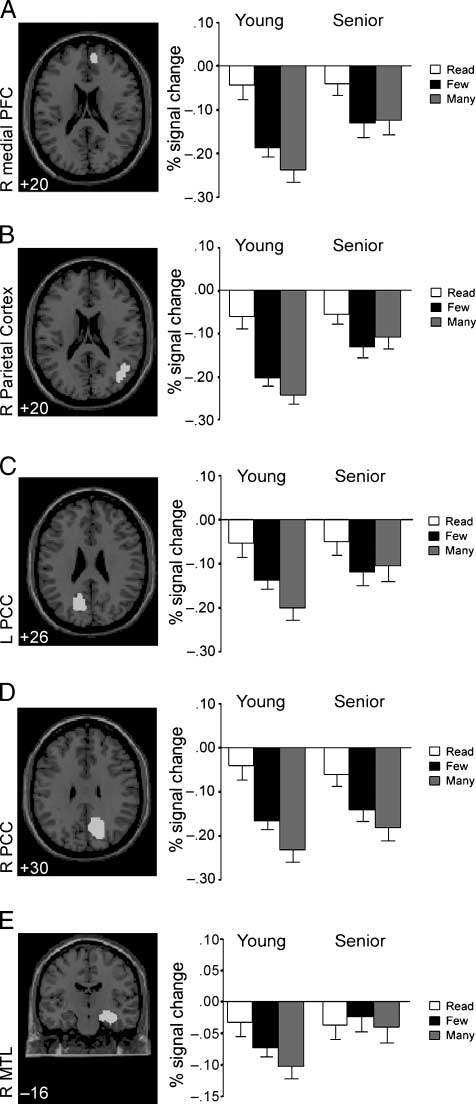
ROI Analyses
The primary objective for the ROI analyses was to
investigate age-related differences in magnitude and
temporal properties in regions associated with the
default-mode network. A critical feature of these analyses
was to investigate whether age-related differences in
deactivation varied as a function of task demand. Sepa-
rate Group (young, senior) by Condition (read, few,
many) ANOVAs were performed for each of the ROIs.
In the analysis of deactivation magnitude for the me-
dial frontal region (Figure 3A), there was a main effect
of condition [F(2, 54) = 29.6, p < .001)]. Deactivation
became greater (task (cid:2) baseline values became more
negative) as a function of task demand. The Condition (cid:1)
Age interaction was also significant [F(2, 54) = 4.33,
p < .05)], indicating that age differences in deactivation
magnitude changed as a function of demand. Follow-up
analyses showed that young participants had greater
deactivation than seniors in the MANY condition [high-
selection demand: t(54) = 2.40, p < .05], whereas deac-
tivations for the FEW condition [low selection demand:
t(54) = 0.776, p = .441] and the read condition [t(54) =
0.769, p = .445] were nonsignificant. These results sug-
gest that as processing demand increases, the magnitude
of medial prefrontal deactivation in this region increases
accordingly. Moreover, age differences emerge when
higher levels of selection are required.
The same pattern was evident for the right LPC (Fig-
ure 3B). The main effect for condition [F(2, 54) = 44.97,
p < .001], and the Task (cid:1) Age interaction [F(2, 54) =
3.76, p < .05] were both significant. The deactivation
was greater during more demanding task conditions;
older adults showed less deactivation than young adults
in the high-selection condition [t(54) = 2.85, p < .01],
but not in the low-selection or read conditions [t(54) =
.487, p = .628; t(54) = .003, p = .998].
Data for the left PCC (Figure 3C) yielded a significant
Condition (cid:1) Age interaction [F(2, 54) = 4.03, p < .05].
The main effects for condition and age were not signifi-
cant. In contrast to the observation of greater deactivation
for more demanding conditions in young participants,
senior participants did not show variations due to task
demand. In line with the findings for the medial PFC re-
gion and the lateral parietal region, age differences were
found in the most demanding condition [t(54) = 2.26,
p < .05], but not in the condition with low-selection
demands [t(54) = 1.28, p = .203] or the read condition
[t(54) = 0.017, p = .986].
In the analysis of the magnitude of deactivation in
the right PCC (Figure 3D), the main effect of condition
was significant [F(2, 54) = 15.9, p < .001]. The main
effect of age was not significant, and the Condition (cid:1)
Age interaction showed a trend [F(2, 54) = 2.71, p <
.071] indicating that the age differences were exacer-
bated with increasing selection demand. Once again, a
significant group difference was present for the high-
selection condition [t(54) = 1.73, p < .05], whereas
neither the low-selection [t(54) = 0.318, p = .751] nor
the read condition [t(54) = 0.048, p = .962] differed be-
tween groups.
Persson et al.
1025
were significant. The Condition by Age interaction sug-
gests that although younger individuals show more deac-
tivation in more demanding selection conditions, older
individuals show similar levels of deactivation across se-
lection demands.
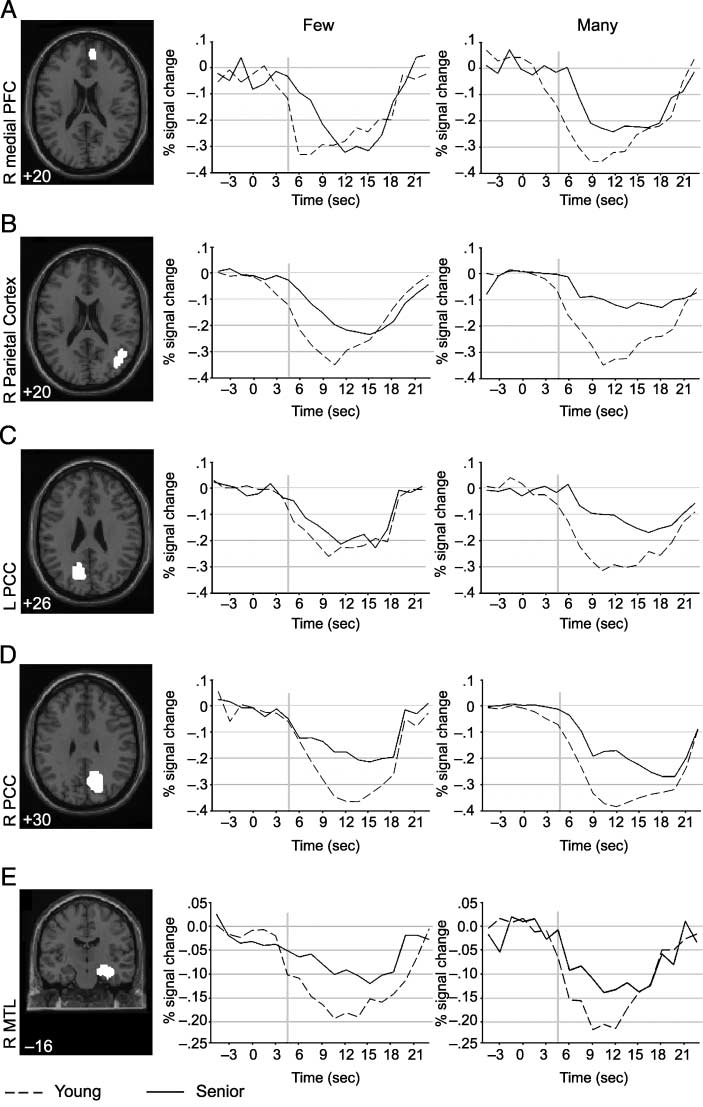
Time Course Analyses
Although time course analyses are most frequently per-
formed for event-related designs, they can also be in-
formative about temporal properties in block designs
(Fox, Snyder, Barch, Gusnard, & Raichle, 2005; Konishi,
Donaldson, & Buckner, 2001; Friston, Frith, Turner, &
Frackowiak, 1995). Konishi et al. suggested that effects
linked to the transition between baseline and task blocks
could reflect updating or changing the behavioral mode
(i.e., ‘‘task set’’), or alternatively might reflect competi-
tion between brain regions (and their respective cogni-
tive processes) to control performance during the task.
Either of these possibilities would require cognitive con-
trol in order to activate the appropriate task processes
or to inhibit inappropriate ones. Lustig et al. (2003)
noted qualitative variations in the temporal profile of
deactivations as a function of region (medial prefrontal,
lateral parietal, or posterior cingulate/medial parietal)
and group (young, old, or Alzheimer’s), but did not
conduct formal analyses (see also Rombouts, Goekoop,
et al., 2005, for a comparison of early vs. late phases of
the BOLD response in an event-related analysis of data
from healthy older adults, mildly cognitively impaired
older adults, and Alzheimer’s patients).
We investigated age differences in the magnitude
and shape of the time courses for each of the ROIs
(Figure 4). An analysis of the time courses for the low-
selection condition (FEW) revealed a significant Time (cid:1)
Group interaction for the medial PFC [Figure 4A; F(18,
54) = 2.25, p < .05]. For the high-selection condition
(MANY), a similar pattern was evident for the medial
PFC [Figure 4A; F(18, 54) = 2.49, p < .01], the right LPC
[Figure 4B; F(18, 54) = 7.29, p < .001], and the left
PCC [Figure 4C; F(18, 54) = 2.72, p < .001]. No Time (cid:1)
Group difference was found for the visual cortex (BA 17;
data not shown) in either the high- or low-selection con-
dition, suggesting that the differences in time courses
for the default-mode regions are not related to general
aspects of changes in the hemodynamic response that
occur with age. The significant interaction between time
and age indicates that the time course of the response in
that region had a different shape for young compared
to senior participants. In addition to the findings of less
deactivation for senior adults,
inspection of Figure 4
suggests that there is a slight delay in deactivation for
senior adults compared to young adults.
In order to investigate possible age and condition dif-
ferences in positive activation–deactivation dynamics, we
also plotted activation time courses from the right IFG
(BA 45/46; x, y, z = 41, 15, 5) region that was associated
Figure 3. Transverse sections depict the location of the areas used
for the ROI analyses. Bar graphs represent the average percent
signal change for young and senior participants for each of the
conditions (READ, FEW, MANY) compared to a rest baseline. Error
bars represent standard error of the mean.
The results for the right MTL region (Figure 3E)
showed a significant Condition (cid:1) Age interaction [F(2,
54) = 5.93, p < .01]. The main effects for condition and
age were not significant. Also, the group difference for
neither the selection condition nor the read condition
1026
Journal of Cognitive Neuroscience
Volume 19, Number 6
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
9
e
6
-
1
p
0
d
2
f
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
c
/
n
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
c
2
n
1
.
p
2
d
0
0
b
7
y
.
g
1
u
9
e
.
s
6
t
.
o
1
n
0
0
2
8
1
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
f
.
o
n
1
8
M
a
y
2
0
2
1
Figure 4. Transverse sections
depict the location of the areas
used for the ROI analyses. The
graphs represent time courses
for each of the verb generate
conditions (FEW, MANY).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
9
e
6
-
1
p
0
d
2
f
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
c
/
n
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
c
2
n
1
.
p
2
d
0
0
b
7
y
.
g
1
u
9
e
.
s
6
t
.
o
1
n
0
0
2
8
1
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
.
.
.
.
.
f
o
n
1
8
M
a
y
2
0
2
1
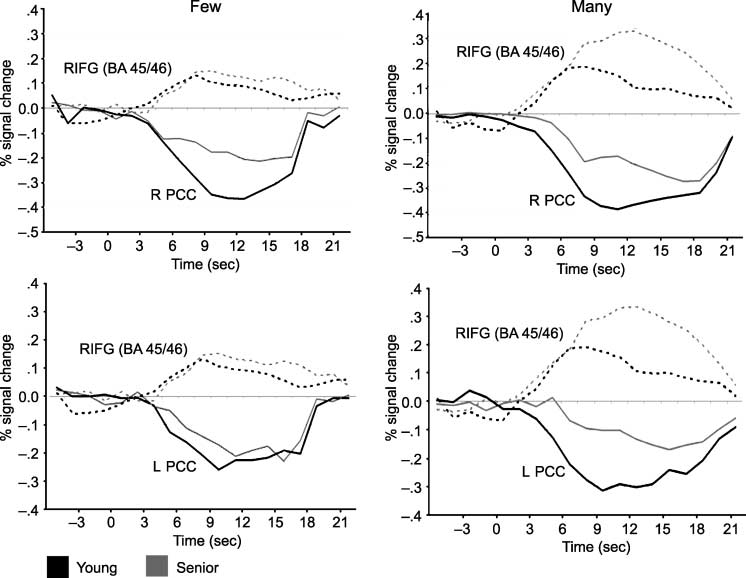
with age differences in activation in our previous analy-
ses of the positive activations on this dataset (Persson
et al., 2004), together with deactivation time courses for
the two medial PCC regions (Figure 5). Visual inspection
of these time courses revealed only small age differ-
ences in the low-selection condition, with the peak of
activation occurring before the peak of deactivation. Al-
though young adults maintain this pattern in the high-
selection demand condition, older adults show a more
extended frontal activation with a later peak. Power at
this sample size does not allow for strong statements
about whether this age difference interacts with selec-
tion difficulty. However, the more consistent findings
of age differences in deactivation in the high-selection
condition than in the low-selection condition are sug-
gestive in this regard.
We also asked whether deactivations in our ROIs
might be related to positive activations in regions that
Persson et al.
1027
Figure 5. The graphs
represent time courses
for each of the high- and
low-selection conditions for
the right IFG region together
with the medial PCC regions
for each age group.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
t
o
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
p
r
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
a
m
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
a
e
-
r
p
t
d
i
c
1
l
9
e
6
-
1
p
0
d
2
f
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
c
/
n
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
c
2
n
1
.
p
2
d
0
0
b
7
y
.
g
1
u
9
e
.
s
6
t
.
o
1
n
0
0
2
8
1
S
.
e
p
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
f
.
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
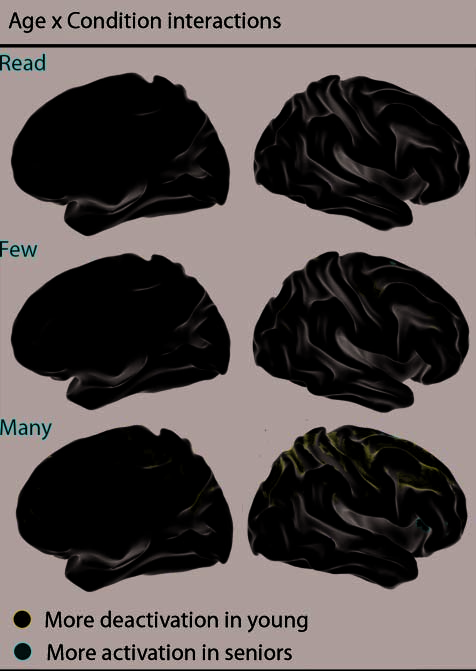
risk of missing effects elsewhere in the brain. We there-
fore conducted exploratory whole-brain analyses to
confirm and extend the a priori ROI analyses. To dis-
play the effects of age at each level of task difficulty,
we conducted three separate Group (young vs. old) (cid:1)
Condition (active task vs. fixation baseline) voxelwise
ANOVAs, one for each level of demand (Figure 7).
For each of these analyses, we distinguished between
regions showing more deactivation by young adults ver-
sus those showing more positive activation by older
adults. Consistent with the ROI analyses, age differences
in deactivation magnitude were most apparent in the
are typically associated with cognitive control. To test
this hypothesis, we examined potential correlations be-
tween the deactivation ROIs used here and a region in
the left IFG (BA 45/46; x, y, z = (cid:2)49, 26, 15) that showed
positive activation in the analysis by Persson et al. (2004),
and a region in the right IFG (BA 45/46; x, y, z = 41, 15, 5)
that showed an age-related increase in that analysis.
Neither of these regions correlated with any of the de-
activation ROIs.
Correlation Analyses of Behavioral Data
Do individual differences in deactivation correspond
to individual differences in behavior? The right PCC
showed a significant correlation (r = .40, p < .01) be-
tween changes in deactivation magnitude and changes
in response time between the high- and low-selection
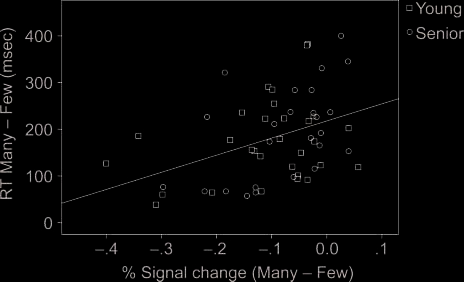
conditions (Figure 6). Those participants who showed
the smallest difference in deactivation magnitude be-
tween the high- and low-selection conditions were those
who showed the largest interference effects, as reflected
by longer response time. This pattern is consistent with
the idea that deactivation magnitudes correspond with
an individual’s response to demands for selection or
resolving interference.
Whole-brain Analyses
Although ROI analyses allow the rigor of a priori hy-
pothesis testing and increased sensitivity, they run the
Figure 6. Correlation between the behavioral interference effect
(MANY vs. FEW) and the magnitude of interference in deactivation
(MANY vs. FEW) in the right PCC.
1028
Journal of Cognitive Neuroscience
Volume 19, Number 6
between competing alternatives). Additionally, inspec-
tion of the time courses suggested that deactivations are
not only reduced in magnitude but also slower for older
adults than for young adults under high control demand
conditions, suggesting a slower reallocation of attention
or resources. Furthermore, control-related changes in
deactivation in the right PCC correlated with control-
related changes in performance. To our knowledge, this
is the first report of individual-level correlations between
deactivation change and performance change, and it pro-
vides support for the idea that deactivations have func-
tional significance.
The most likely explanation for deactivation-perform-
ance links is that deactivations reflect a diversion of at-
tention away from so-called default mode processes and
toward the experimental task. Spontaneous (non-task-
related) activity in regions associated with task-induced
deactivations and default-mode processing correlates
negatively with spontaneous activity in regions associ-
ated with task-induced positive activations, suggesting
the existence of two competing networks (Fox et al.,
2005). Other manipulations of difficulty (target discrim-
inability, stimulus presentation rate, and short-term
memory load) also lead to changes in deactivation mag-
nitude similar to those seen here (McKiernan et al.,
2003). Greater deactivations at encoding are related to
subsequent memory, again suggesting a reallocation of
attentional resources away from default mode processes
and toward learning and memory (Daselaar et al., 2004).
The group- and individual-differences level data re-
ported here converge with the network-level, task-level,
and item-level findings to suggest that deactivations re-
flect important processes related to cognitive control.
Difficulties with cognitive control are a major feature
of cognitive aging, but only a few prior studies have spe-
cifically examined the effects of aging on default mode
functioning (Rombouts, Barkhof, et al., 2005; Lustig et al.,
2003). Although the specifics differ between these stud-
ies and ours (e.g., by including Alzheimer’s disease pa-
tients), the overall pattern of age-related changes in
deactivation magnitude is generally consistent with the
current findings. For example, the observation of re-
duced medial parietal and frontal deactivations in older
adults found here during a verb generation task repli-
cates and extends previous findings of age differences
using a semantic classification task (Lustig et al., 2003).
These regions also show alterations in individuals with
mild cognitive impairment (Rombouts, Barkhof, et al.,
2005) and Alzheimer’s disease (Rombouts, Barkhof,
et al., 2005; Lustig et al., 2003), suggesting a relation be-
tween disruption of the default-mode network and de-
mentia severity. However, reduced deactivations are
likely a characteristic of normal aging as well as demen-
tia, as recent work suggests that a reduction in deacti-
vation magnitude is apparent even in middle age (Grady
et al., 2006). An important question for future work is
whether age differences in deactivation interact with the
Persson et al.
1029
Figure 7. Map of statistically significant group differences across
the brain for each of the conditions (i.e., baseline > read/few/many).
Each interaction was masked by an overall contrast to indicate the
direction of effects.
high-selection condition. Regions showing age differ-
ences in deactivation included the postcentral sulcus,
the medial PFC, the LPC, and the PCC (Figura 7). Age
differences in deactivation magnitude are unlikely to re-
flect a simple lack of responsiveness by older adult
brains: Older adults show greater positive activations
than do young adults in a medial dorsal frontal region
(SMA; BA 6) and right IFG (BA 45/46). The age differ-
ence in the right IFG was specific to the high-selection
condition (see also Figure 5), also discounting a generic
‘‘positive bias’’ account of age differences in activation
and deactivation.
DISCUSSION
Several significant findings emerged from these results.
Primo, age differences in deactivation were not present
in an active task with minimal selection demands, E
overall deactivations were small in this condition. Both
overall deactivation magnitude and the size of age dif-
ferences in deactivation became greater as a function of
the demand for cognitive control (specifically, selection
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
w
T
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
T
N
io
C
/
UN
e
–
R
P
T
D
io
C
1
l
9
e
6
–
1
P
0
D
2
F
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
C
/
N
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
C
2
N
1
.
P
2
D
0
0
B
7
sì
.
G
1
tu
9
e
.
S
6
T
.
o
1
N
0
0
2
8
1
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
.
.
.
F
.
.
o
N
1
8
M
UN
sì
2
0
2
1
type as well as the level of task demands; the selection
demands of the verb generation task revealed age differ-
ences in lateral parietal deactivation that were not found
in previous studies focusing on memory encoding and
retrieval (Grady et al., 2006; Lustig et al., 2003).
The Age by Condition interactions represent an ad-
vance over previous studies because they links age dif-
ferences in deactivation magnitude to increased demand
for cognitive control, rather than to more general age
differences in the hemodynamic response implied by
main effects for age (Vedere, per esempio., Johnson, Mitchell, Raye,
& Greene, 2004; Buckner, Snyder, Sanders, Raichle, &
Morris, 2000 for other examples of Age by Condition
interaction logic). Young and old adults were remarkably
similar during low-demand conditions. Age differences
were only strongly revealed when demand was relatively
high. The control demand specificity of age differences
in deactivation magnitude supports the idea that they
reflect age differences in the cognitive control required
to divert attention away from task-irrelevant default-
mode processing.
The failure to find Age by Condition interactions in
performance might at first seem to argue against this
conclusion. Infatti, the possibility exists that reduced
deactivations by older adults in the high demand con-
ditions may even reflect a form of compensation, COME
age differences in positive activation in the absence
of behavioral age differences are often interpreted as
compensatory (per esempio., Cabeza, Anderson, Locantore, &
McIntosh, 2002; Reuter-Lorenz et al., 2000; Grady et al.,
1994). Tuttavia, correlations with performance provide
evidence against the compensation explanation of re-
duced deactivations. The inverse relationship between
deactivation in the right PCC and interference scores
(many (cid:2) few difference) indicates that, at a particular
level of task complexity, greater deactivation may be
related to more efficient task performance.
Given that only a few studies have examined age
differences in deactivation, it is too early to rule out a
possible compensatory role. Tuttavia, we speculatively
propose the opposite hypothesis: Failures to divert
attention or resources away from default-mode process-
ing (as reflected by slower and reduced deactivations)
may, Infatti, be part of what older adults are compensat-
ing for (as reflected by increased or additional frontal
activations). Spared performance may result when older
adults recruit additional (prefrontal) regions to compen-
sate for lingering default-mode processes during the
experimental task (see Rombouts, Barkhof, et al., 2005
for a more extreme version of this pattern in Alzheimer’s
disease patients).
The hypothesis that increased frontal activations may
reflect compensation for failures to divert attention from
or inhibit default-mode processes receives tentative but
convergent support from several recent findings. Mir-
roring the control-demand specificity of age differences
in deactivation magnitude and time course found here,
Velanova et al. (2007) recently reported that extend-
ed positive activation in the right IFG for older adults
was specific to a high control-demand retrieval condi-
zione, and did not occur under low-demand conditions.
Within the current dataset, greater right IFG activation
was found for seniors compared to young adults in
the high-selection condition (Persson et al., 2004). Com-
parison of time courses for right IFG and medial PCC
regions across conditions (Figura 5) reveals a similar
time course of frontal activation for young and old
adults in the low-selection demand condition, replicat-
ing Velanova et al., and for both age groups, the peak of
frontal activation occurs before the point of greatest
deactivation. Young adults maintain this pattern in the
high-selection demand condition, but older adults show
a more extended frontal activation with a later peak,
which may be related to their slower deactivation time
course. Of interest, aging is not the only condition that is
associated with disruptions in cognitive control and its
neuroimaging correlates, including deactivations: Some
parallels exist between the present results and those
found due to sleep deprivation (Chee & Choo, 2004). Al-
though this area of research is relatively new, the find-
ings seem to converge on the hypothesis that successful
task execution results from a coordinated pattern of ac-
tivation in task-relevant areas and deactivation of task-
irrelevant ones.
Given the increased RT with task difficulty, one alter-
native explanation for the findings of greater deacti-
vation in conditions with high demands for cognitive
is increased time on task. RTs and demands
controllo
for cognitive control are typically tightly linked, making
it difficult to rule out this explanation at all levels of
analysis. Tuttavia, it is unlikely to apply as a general
principle. Primo, a simple time-on-task account of deacti-
vations would have difficulty explaining why older adults
showed similar changes in RT across conditions as did
young adults, but not similar changes in deactivation
magnitude. Secondo, overall older adults had longer RTs
than did young adults (spent more time on task), Ma
had smaller deactivations. Inoltre, a time-on-task
account would predict that even within an age group,
the fastest subjects should show the smallest deacti-
vations. Our results trended in the opposite direction
(data not shown). Finalmente, a comparison of the condi-
tions in which young and older adults were matched
on response time (young (cid:2) many versus old (cid:2) few) Rif-
sults in larger age differences in deactivation magnitude.
Taken together, these patterns make it very difficult to
support an account whereby greater deactivations rep-
resent generic effects of greater difficulty or greater time
on task. Invece, we suggest that greater deactivations
represent greater responsiveness to cognitive control
demands—here instantiated as a demand on selection
processes.
In conclusion, these findings suggest that advanced
age alters the functional properties of task-induced
1030
Journal of Cognitive Neuroscience
Volume 19, Numero 6
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
w
T
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
T
N
io
C
/
UN
e
–
R
P
T
D
io
C
1
l
9
e
6
–
1
P
0
D
2
F
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
C
/
N
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
C
2
N
1
.
P
2
D
0
0
B
7
sì
.
G
1
tu
9
e
.
S
6
T
.
o
1
N
0
0
2
8
1
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
.
.
.
F
.
.
o
N
1
8
M
UN
sì
2
0
2
1
deactivations, and that age-related differences are most
pronounced in conditions with greater demand for
cognitive control. Observations of an inverse relation-
ship between deactivation in specific regions and inter-
ference suggest that deactivations may be related to the
degree of cognitive efficiency. Reduced deactivation for
older adults in high-control conditions may indicate a
reduction in cognitive efficiency stemming from difficul-
ty disengaging from or inhibiting internal processes in
order to reallocate attention to the task at hand.
Ringraziamenti
We thank Ching-Yune Sylvester for her contributions to the
verb generation experiment and Sharon Thompson-Schill and
Deanna Barch for providing us with stimulus material for this
task. We are also grateful to Abraham Z. Snyder for technical
advice. This work was supported by the National Institutes of
Health Grant AG18286.
Reprint requests should be sent to Patricia A. Reuter-Lorenz,
Department of Psychology, University of Michigan, East Hall,
530 Church Street, Ann Arbor, MI 48109-1109, or via e-mail:
parl@umich.edu.
REFERENCES
Andreasen, N. C., O’Leary, D. S., Cizadlo, T., Arndt, S., Rezai, K.,
Watkins, L., et al. (1995). Remembering the past—2 facets
of episodic memory explored with Positron Emission
Tomography. American Journal of Psychiatry, 152,
1576–1585.
Binder, J. R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S.,
Rao, S. M., & Cox, R. W. (1999). Conceptual processing
during the conscious resting state. A functional MRI
study. Journal of Cognitive Neuroscience, 11, 80–95.
Braver, T. S., & Barch, D. M. (2002). A theory of cognitive
controllo, aging cognition, and neuromodulation.
Neuroscience and Biobehavioral Reviews, 26, 809–817.
Buckner, R. l. (2004). Memory and executive function in
aging and AD: Multiple factors that cause decline and
reserve factors that compensate. Neuron, 44, 195–208.
Buckner, R. L., Snyder, UN. Z., Sanders, UN. L., Raichle, M. E.,
& Morris, J. C. (2000). Functional brain imaging of
young, nondemented, and demented older adults.
Journal of Cognitive Neuroscience, 12(Suppl. 2),
24–34.
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975).
Mini-Mental State: A practical method for grading the
cognitive state of patients for the clinician. Journal of
Psychiatric Research, 12, 189–198.
Fox, M. D., Snyder, UN. Z., Barch, D. M., Gusnard, D. A.,
& Raichle, M. E. (2005). Transient BOLD responses at
block transitions. Neuroimage, 28, 956–966.
Fox, M. D., Snyder, UN. Z., Vincent, J. L., Corbetta, M.,
Van Essen, D. C., & Raichle, M. E. (2005). The human
brain is intrinsically organized into dynamic, anticorrelated
functional networks. Proceedings of the National
Academy of Sciences, U.S.A., 102, 9673–9678.
Fransson, P. (2005). Spontaneous low-frequency BOLD signal
fluctuations: An fMRI investigation of the resting-state
default mode of brain function hypothesis. Human Brain
Mapping, 26, 15–29.
Friston, K. J., Frith, C. D., Turner, R., & Frackowiak, R. S.
(1995). Characterizing evoked hemodynamics with fMRI.
Neuroimage, 2, 157–165.
Grady, C. L., Maisog, J. M., Horwitz, B., Ungerleider, l. G.,
Mentis, M. J., Salerno, J. A., et al. (1994). Age-related
changes in cortical blood flow activation during visual
processing of faces and location. Journal of Neuroscience,
14, 1450–1462.
Grady, C. L., Springer, M. V., Hongwanishkul, D., McIntosh,
UN. R., & Winocur, G. (2006). Age-related changes in brain
activity across the adult lifespan. Journal of Cognitive
Neuroscience, 18, 227–241.
Greicius, M. D., Krasnow, B., Reiss, UN. L., & Menon, V.
(2003). Functional connectivity in the resting brain:
A network analysis of the default mode hypothesis.
Proceedings of the National Academy of Sciences,
U.S.A., 100, 253–258.
Greicius, M. D., Srivastava, G., Reiss, UN. L., & Menon, V. (2004).
Default-mode network activity distinguishes Alzheimer’s
disease from healthy aging: Evidence from functional
MRI. Proceedings of the National Academy of Sciences,
U.S.A., 101, 4637–4642.
Gusnard, D. A., Akbudak, E., Shulman, G. L., & Raichle,
M. E. (2001). Medial prefrontal cortex and self-referential
mental activity: Relation to a default mode of brain function.
Proceedings of the National Academy of Sciences, U.S.A.,
98, 4259–4264.
Hasher, L., & Zacks, R. T. (1988). Working memory,
comprehension, and aging: A review and a new view.
Psychology of Learning and Motivation, 22, 193–225.
Heaton, R. K., Chelune, C. J., Talley, J. L., Kay, G. G., &
Curtis, G. (1993). Wisconsin card sorting test manual,
revised and expanded. Odessa, FL: Psicologico
Assessment Resources.
Cabeza, R., Anderson, N. D., Locantore, J. K., & McIntosh,
Hedden, T., & Gabrieli, J. D. (2004). Insights into the
UN. R. (2002). Aging gracefully: Compensatory brain
activity in high-performing older adults. Neuroimage,
17, 1394–1402.
Chee, M. W., & Choo, W. C. (2004). Functional imaging of
working memory after 24 hr of total sleep deprivation.
Journal of Neuroscience, 24, 4560–4567.
Choo, W. C., Lee, W. W., Venkatraman, V., Sheu, F. S., &
Chee, M. W. (2005). Dissociation of cortical regions
modulated by both working memory load and sleep
deprivation and by sleep deprivation alone. Neuroimage,
25, 579–587.
Daselaar, S. M., Prince, S. E., & Cabeza, R. (2004). When less
means more: Deactivations during encoding that predict
subsequent memory. Neuroimage, 23, 921–927.
Delis, D. C., Kramer, J. H., Kaplan, E., & Ober, B. UN. (1987).
The California Verbal Learning Test. New York: IL
Psychological Corporation.
ageing mind: A view from cognitive neuroscience. Nature
Reviews Neuroscience, 5, 87–96.
Jennings, J. M., & Jacoby, l. l. (1993). Automatic versus
intentional uses of memory: Aging, Attenzione, and control.
Psychology and Aging, 8, 283–293.
Johnson, M. K., Mitchell, K. J., Raye, C. L., & Greene, E. J.
(2004). An age-related deficit in prefrontal cortical
function associated with refreshing information.
Psychological Science, 15, 127–132.
Kan, IO. P., & Thompson-Schill, S. l. (2004). Selection from
perceptual and conceptual representations. Cognitive,
Affective, and Behavioral Neuroscience, 4, 466–482.
Konishi, S., Donaldson, D. I., & Buckner, R. l. (2001).
Transient activation during block transition. Neuroimage,
13, 364–374.
Lustig, C., Snyder, UN. Z., Bhakta, M., O’Brien, K. C., McAvoy, M.,
Raichle, M. E., et al. (2003). Functional deactivations:
Persson et al.
1031
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
w
T
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
T
N
io
C
/
UN
e
–
R
P
T
D
io
C
1
l
9
e
6
–
1
P
0
D
2
F
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
C
/
N
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
C
2
N
1
.
P
2
D
0
0
B
7
sì
.
G
1
tu
9
e
.
S
6
T
.
o
1
N
0
0
2
8
1
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
F
.
.
.
.
.
o
N
1
8
M
UN
sì
2
0
2
1
Change with age and dementia of the Alzheimer type.
Proceedings of the National Academy of Sciences, U.S.A.,
100, 14504–14509.
in the frontal lateralization of verbal and spatial working
memory revealed by PET. Journal of Cognitive
Neuroscience, 12, 174–187.
Mazoyer, B., Zago, L., Mellet, E., Bricogne, S., Etard, O.,
Reuter-Lorenz, P. A., & Lustig, C. (2005). Brain aging:
Houde´, O., et al. (2001). Cortical networks for working
memory and executive functions sustain the conscious
resting state in man. Brain Research Bulletin, 54,
287–298.
McKiernan, K. A., Kaufman, J. N., Kucera-Thompson, J.,
& Binder, J. R. (2003). A parametric modulation of
factors affecting task-induced deactivation in functional
neuroimaging. Journal of Cognitive Neuroscience, 15,
394–408.
Persson, J., Sylvester, C.-Y. C., Nelson, J. K., Welsh, K. M.,
Jonides, J., & Reuter-Lorenz, P. UN. (2004). Selection
requirements during verb generation: Differential
recruitment in older and younger adults. Neuroimage,
23, 1382–1390.
Phelps, M. E., Mazziotta, J. C., Kuhl, D. E., Nuwer, M.,
Packwood, J., Metter, J., et al. (1981). Tomographic mapping
of human cerebral metabolism visual stimulation and
deprivation. Neurology, 31, 517–529.
Raichle, M. E., MacLeod, UN. M., Snyder, UN. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. l. (2001). A default mode
of brain function. Proceedings of the National Academy
of Sciences, U.S.A., 98, 676–682.
Rajah, M. N., & D’Esposito, M. (2005). Region-specific changes
in prefrontal function with age: A review of PET and
fMRI studies on working and episodic memory. Brain,
128, 1964–1983.
Reuter-Lorenz, P. A., Jonides, J., Smith, E. E., Hartley, A.,
Mugnaio, A., Marshuetz, C., et al. (2000). Age differences
Reorganizing discoveries about the aging mind. Current
Opinion in Neurobiology, 15, 245–251.
Rombouts, S. A., Barkhof, F., Goekoop, R., Stam, C. J.,
& Scheltens, P. (2005). Altered resting state networks
in mild cognitive impairment and mild Alzheimer’s
disease: An fMRI study. Human Brain Mapping, 26,
231–239.
Rombouts, S. A., Goekoop, R., Stam, C. J., Barkhof, F.,
& Scheltens, P. (2005). Delayed rather than decreased
BOLD response as a marker for early Alzheimer’s disease.
Neuroimage, 26, 1078–1085.
Shulman, G. L., Fiez, J. A., Corbetta, M., Buckner, R. L.,
Miezin, F. M., Raichle, M. E., et al. (1997). Common
blood flow changes across visual tasks: II. Decreases in
cerebral cortex. Journal of Cognitive Neuroscience, 9,
648–663.
Thompson-Schill, S. L., D’Esposito, M., Aguirre, G. K., &
Farah, M. J. (1997). Role of left inferior prefrontal cortex
in retrieval of semantic knowledge: A reevaluation.
Proceedings of the National Academy of Sciences, U.S.A.,
94, 14792–14797.
Velanova, K., Lustig, C., Jacoby, l. L., & Buckner, R. l.
(2007). Evidence for frontally-mediated controlled
processing differences in older adults. Cerebral Cortex,
17, 1033–1046.
West, R. l. (1996). An application of prefrontal cortex
function theory to cognitive aging. Psychological Bulletin,
120, 272–292.
D
o
w
N
l
o
UN
D
e
D
F
R
o
M
l
l
/
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
D
H
T
o
w
T
P
N
:
o
/
UN
/
D
M
e
io
D
T
F
P
R
o
R
M
C
.
H
S
io
P
l
v
D
e
io
R
R
e
C
C
H
T
.
UN
M
io
R
e
.
D
C
tu
o
M
o
/
C
j
N
o
UN
C
R
T
N
io
C
/
UN
e
–
R
P
T
D
io
C
1
l
9
e
6
–
1
P
0
D
2
F
1
/
1
1
9
9
/
3
6
6
3
/
9
1
4
0
2
o
1
C
/
N
1
2
7
0
5
0
6
7
7
1
2
9
1
/
6
j
1
o
0
C
2
N
1
.
P
2
D
0
0
B
7
sì
.
G
1
tu
9
e
.
S
6
T
.
o
1
N
0
0
2
8
1
S
.
e
P
P
D
F
e
M
B
B
sì
e
R
G
2
tu
0
e
2
S
3
T
/
j
T
.
.
.
.
F
.
o
N
1
8
M
UN
sì
2
0
2
1
1032
Journal of Cognitive Neuroscience
Volume 19, Numero 6