Age- and Episodic Memory-related Differences in
Task-based Functional Connectivity in Women and Men
Sivaniya Subramaniapillai1,2*, Sricharana Rajagopal2, Elizabeth Ankudowich1,2,
Stamatoula Pasvanis2, Bratislav Misic2, and M. Natasha Rajah1,2*
Astratto
■ Aging is associated with episodic memory decline and
changes in functional brain connectivity. Understanding
whether and how biological sex influences age- and memory
performance-related functional connectivity has important the-
oretical implications for the cognitive neuroscience of memory
and aging. Here, we scanned 161 healthy adults between 19 E
76 years of age in an event-related fMRI study of face–location
spatial context memory. Adults were scanned while performing
easy and difficult versions of the task at both encoding and
retrieval. We used multivariate whole-brain partial least squares
connectivity to test the hypothesis that there are sex differences
in age- and episodic memory performance-related functional
connectivity. We examined how individual differences in age
and retrieval accuracy correlated with task-related connectivity.
We then repeated this analysis after disaggregating the data by
self-reported sex. We found that increased encoding and
retrieval-related connectivity within the dorsal attention net-
lavoro (DAN), and between DAN and frontoparietal network
and visual networks, were positively correlated to retrieval accu-
racy and negatively correlated with age in both sexes. Anche noi
observed sex differences in age- and performance-related func-
tional connectivity: (UN) Greater between-networks integration
was apparent at both levels of task difficulty in women only,
E (B) increased DAN–default mode network connectivity
with age was observed in men and was correlated with poorer
memory performance. Perciò, the neural correlates of
age-related episodic memory decline differ in women and
men and have important theoretical and clinical implications
for the cognitive neuroscience of memory, aging, and dementia
prevention. ■
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
INTRODUCTION
Healthy aging is associated with episodic memory decline,
a reduced ability to encode, store, and retrieve past expe-
riences in rich spatiotemporal contextual detail (Grady &
Craik, 2000; Tulving, 1972). Age-associated episodic
memory decline impairs older adults’ quality of life and
can be an early sign of sporadic Alzheimer disease (AD;
Mol et al., 2007; Mol, van Boxtel, Willems, & Jolles, 2006).
Given that the proportion of older adults is increasing
worldwide, and age is the strongest predictor of AD, there
is an urgent need to understand how normative aging
influences memory and related brain function.
To this aim, there is a large body of research that
has investigated how normative aging affects episodic
memory and related brain activity using task fMRI (Maillet
& Rajah, 2014; Nyberg, Lövdén, Riklund, Lindenberger, &
Bäckman, 2012; Spaniol et al., 2009; Grady, 2008; Sperling,
2007; Rajah & McIntosh, 2005; Naveh-Benjamin, Hussain,
Guez, & Bar-On, 2003). This research has shown that
age-related reductions in episodic memory, as measured
by associative memory tasks (per esempio., spatial context memory
1McGill University, Montréal, Quebéc, Canada, 2Douglas Mental
Health University Institute, Montréal, Quebéc, Canada
*Both authors contributed equally to writing the article.
© 2022 Istituto di Tecnologia del Massachussetts
compiti), are present at midlife and increase with advanced
age (Ankudowich, Pasvanis, & Rajah, 2016; Kwon et al.,
2016; Cansino, 2009), and that these behavioral reduc-
tions are associated with altered activation in occipito-
temporal, pFC, inferior parietal cortex, and medial
temporal lobe with age (Ankudowich, Pasvanis, & Rajah,
2017, 2019; Ankudowich et al., 2016). Inoltre, con
the growing consensus that human cognition and behavior
depends on the dynamic interactions of large-scale neural
reti (Sporns & Betzel, 2016; McIntosh, 2000;
Strother, Kanno, Rottenberg, Friston, & Ford, 1995;
Friston, 1994; Mesulam, 1990), several cognitive neurosci-
ence studies of aging have focused on how age differences
in interregional or internetwork correlations in brain
activity (functional connectivity) during resting state fMRI
(rsfMRI) relate to cognitive task performance assessed
outside of the scanner (Uddin, Sì, & Spreng, 2019;
Power et al., 2011; Yeo et al., 2011; Biswal, Zerrin Yetkin,
Haughton, & Hyde, 1995).
Studies of rsfMRI connectivity have found that
age-related decreases in cognitive task performance were
associated with reduced anticorrelation between the
dorsal attention network (DAN) and default mode net-
lavoro (DMN), possibly as a consequence of disrupted
frontoparietal network (FPN) engagement (Esposito
Journal of Cognitive Neuroscience 34:8, pag. 1500–1520
https://doi.org/10.1162/jocn_a_01868
et al., 2018; Avelar-Pereira, Bäckman, Wåhlin, Nyberg, &
Salami, 2017; Dixon et al., 2017; Amer, Campbell, &
Hasher, 2016; Grady, Sarraf, Saverino, & Campbell, 2016;
Spreng, Stevens, Viviano, & Schacter, 2016; Prakash, Heo,
Voss, Patterson, & Kramer, 2012; Sala-Llonch et al., 2012;
Fox et al., 2005). More generally, aging has also been cor-
related with increased connectivity between networks
(cioè., network integration) and decreased connectivity
within networks (cioè., network segregation; Damoiseaux,
2017; Chan, Park, Savalia, Petersen, & Wig, 2014). How-
ever, only a few rsfMRI studies have directly explored
whether age-related differences in connectivity correlated
with pre/post-scan performance on episodic memory
compiti (Nordin et al., 2021; Edde et al., 2020; Zhang,
Andreano, Dickerson, Touroutoglou, & Barrett, 2020;
King, de Chastelaine, & Rugg, 2018; Nyberg, 2017; Grady
et al., 2016; Kukolja, Goreci, Onur, Riedl, & Fink, 2016;
Fjell et al., 2015; Wang et al., 2010). Most of these studies
focused on specific a priori defined networks of interest
(but see the work of Fjell et al., 2015). Perciò, there
remains a paucity of knowledge about how age-related dif-
ferences in whole-brain functional connectivity contribute
to decreases in episodic memory with age. Inoltre,
most of what we know about the correlation between
age-related differences in functional connectivity and epi-
sodic memory is based on rsfMRI paradigms. Although
resting-state research has provided a greater understanding
of functional architecture, solely relying on resting state
scans as an indirect proxy for cognitive processes is not
sufficient to understand brain-cognitive processes (Vedere
reviews by Finn, 2021; Campbell & Schacter, 2016).
A nostra conoscenza, no prior work has specifically inves-
tigated how age and performance correlates with whole-
brain, task-based functional connectivity during episodic
encoding and retrieval, across the adult lifespan. One
recent study investigated age-related differences in
whole-brain connectivity during encoding of an associa-
tive memory task across the adult lifespan (Capogna
et al., 2022). Using a whole-brain psychophysiological
interaction analysis to investigate direct brain-cognitive
processes, the authors found that in older age, greater
connectivity between medial temporal and posterior pari-
etal regions during encoding was associated with better
performance, whereas increased connectivity between
frontal, parietal, and visual regions was associated with
worse performance. The functional connectivity patterns
associated with successful memory performance in older
adults are associated with cognitive processes that involve
integrative and multisensory strategies and mental imag-
ery. Tuttavia, this study controlled for sex in their analyses
hindering any further interpretations of how these find-
ings may separately relate to women and men.
Infatti, most fMRI connectivity studies of aging have
assumed that age-related differences in functional connec-
tivity were the same in women and men, because data
were not disaggregated by sex and/or gender at analysis.
Tuttavia, depending on the task stimuli and design,
studies have repeatedly demonstrated behavioral sex dif-
ferences on episodic memory performance. Women typi-
cally perform better than men on episodic memory tasks
of verbal stimuli (Gur & Gur, 2002; Ragland, Coleman,
Gur, Glahn, & Gur, 2000; Herlitz, Nilsson, & Bäckman,
1997), whereas men tend to perform better than women
on visuospatial memory tasks (De Frias, Nilsson, & Herlitz,
2006; Weiss, Kemmler, Deisenhammer, Fleischhacker, &
Delazer, 2003). Tuttavia, these sex differences have small
to medium effect sizes and are stable across the adult
lifespan (Asperholm, Van Leuven, & Herlitz, 2020; Jack
et al., 2015; Voyer, Postma, Brake, & Imperato-McGinley,
2007; De Frias et al., 2006). This may account for the few
studies investigating sex differences in age effects on
memory and associated brain activity and connectivity.
Tuttavia, even if there are no significant Sex main effects
and/or Sex × Age interactions in behavioral outcomes, sex
differences in the underlying neural systems supporting
episodic memory across the adult lifespan may still exist
(Becker & Koob, 2016; McCarthy, Arnold, Ball, Blaustein,
& de Vries, 2012). Consistent with the view that there may
be sexual divergence in the brain systems supporting epi-
sodic memory function in older women and men, recente
studies have found that age-related memory decline was
correlated with different patterns of activations in women
compared with men (Rabipour, Rajagopal, Pasvanis, &
Rajah, 2021; Subramaniapillai et al., 2019). Yet, it remains
unclear if there are sex differences in how age and memory
performance correlate with task-based functional connec-
tivity during episodic memory encoding and retrieval. Questo
information is important to know because, historically, Esso
has been assumed that the neural basis of age-associated
memory decline is the same in both sexes, but this may not
be the case (Subramaniapillai, Almey, Natasha Rajah, &
Einstein, 2021; Rahman et al., 2020; Ferretti et al., 2018;
Nebel et al., 2018; Snyder et al., 2016). Investigating sex
and gender differences in functional brain connectivity
in a normative adult lifespan sample can help determine
if there are sex and/or gender-specific markers of memory
decline in the aging brain. Such knowledge informs us if
the underlying neurocognitive mechanisms linked to
age-related episodic memory decline is the same in
women and men, and if interventions aimed at support-
ing memory into late life should be the same for women
and men.
Here, we present whole-brain functional connectivity
results from an episodic memory task fMRI study of 161
healthy adults aged 19–76 years of age who were scanned
while performing both encoding and retrieval phases of a
face–location spatial context memory paradigm. We par-
cellated task fMRI data into canonical brain networks
defined by Power et al. (2011) and used whole-brain
behavior partial least squares (B-PLS) connectivity analysis
to examine the orthogonalized contributions of age and
memory performance on task-based functional connec-
attività. We then repeated this analysis after disaggregating
the data by self-reported sex to investigate whether both
Subramaniapillai et al.
1501
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
sexes exhibited similar age- and performance-related pat-
terns of connectivity. We hypothesized that age would be
correlated with decreased connectivity between DAN and
FPN and increased connectivity between DAN and DMN,
and memory performance would exhibit the opposite
patterns of network associations (Esposito et al., 2018;
Avelar-Pereira et al., 2017; Dixon et al., 2017; Amer
et al., 2016; Grady et al., 2016; Spreng et al., 2016; Prakash
et al., 2012; Sala-Llonch et al., 2012; Turner & Spreng,
2012; Fox et al., 2005). Based on prior activation analyses
of sex differences in the effect of age and memory accuracy
on task-related brain activity across the adult lifespan
(Subramaniapillai et al., 2019), we hypothesized that both
sexes will exhibit similar patterns of performance-related
functional connectivity at encoding, but not retrieval. Noi
also hypothesized that there would be sex differences
in age-related functional connectivity at both encoding
and retrieval.
METHODS
Participants
Volunteer research participants were recruited from the
Montreal and surrounding area using on-line and print
advertisements and community outreach. Research volun-
teers were told they would first be asked to participate in a
behavioral and neuropsychological testing session ( Visit
1), and if they met our inclusion criteria, they would be
invited back for an fMRI session ( Visit 2). Two hundred
seventy-five participants (102 self-identified as men, 173
self-identified as women) were tested in Visit 1. Of these,
49 were excluded for not meeting our neuropsychologi-
cal inclusion criteria (listed below), 26 were excluded for
having medical/psychiatric exclusionary criteria (listed
below), E 15 participants could not be reached for
scheduling a Visit 2. Perciò, 185 participants were
invited back for Visit 2 and participated in the fMRI portion
of this study. Of these participants, we identified incidental
findings in 9 participants, 5 participants’ fMRI data did not
meet our quality control criteria (listed below), E 10 par-
ticipants did not perform the fMRI task as instructed,
resulting in a sample of 161 participants (49 men, 112
women) who reported no history of neurological or psy-
chological illness, or serious cardiovascular disease. Tutto
participants were right-handed, as confirmed by the Edin-
burgh Inventory for Handedness. Of the 53 middle-aged
women, we had self-reported menopause status for 41
women, 18 of these self-reported having irregular periods,
symptoms of the menopausal transition, and/or had
undergone hormone replacement therapy (HRT). Two
older adult women had also undergone HRT. Così, we
excluded these 20 women from further analyses because
menopause transition and HRT influences memory-
related brain activity (Rentz et al., 2017; Li, Cui, & Shen,
2014; Henderson, 2010; Yonker et al., 2006). Our final
cohort consisted of 141 participants (49 men, 92 women;
65% women) between the ages of 19 E 76 years (mean
age = 47.11 years, SE = 1.41 years; mean education =
15.73 years, SE = 0.18 years). Of the 35 middle-aged
women, we had a self-reported premenopausal status for
23 women, with unknown status for 12 women. As we did
not have hormonal data to verify self-reported meno-
pausal status, we focus here on age and sex effects and
note in our Caveats section the need to consider repro-
ductive age and health in future studies examining sex
differences in brain aging.
Behavioral Methods
Visit 1: Behavioral and Neuropsychological Session
During an initial session, participants provided informed
consent and then were administered a medical screening
questionnaire to assess neurological, psychological, E
physical health. Medical health exclusion criteria for this
study included having a current diagnosis of diabetes,
untreated cataracts and glaucoma, and a current diagnosis
of high cholesterol levels and/or high blood pressure left
untreated in the past 6 months. Inoltre, participants
were excluded if they had a history of a major psychiatric
illness or neurological insult. Participants then underwent
neuropsychological assessment (Mini-International Neu-
ropsychiatric Interview, inclusion cutoff ≤ 2; the Folstein
Mini Mental State Examination, exclusion cutoff < 27; the
Beck Depression Inventory [BDI-II], exclusion cutoff <
15; California Verbal Learning Task [CVLT-I English,
CVLT-II French], exclusion cutoff based on recommenda-
tions by Norman, Evans, Miller, & Heaton, 2000). Only par-
ticipants who met the above neuropsychological criteria
and performed above chance on the practice context
memory task presented in a mock fMRI scanner were
invited to return for a second visit and participate in the
fMRI scanning portion of the study. All participants were
paid for their participation, and the research ethics board
of the Faculty of Medicine at McGill University approved
the study protocol.
Visit 2: Task fMRI Session
Stimuli and procedure. The task fMRI stimulus set has
been used in previous studies and has been independently
rated for pleasantness (Kwon et al., 2016; Rajah, Languay,
& Valiquette, 2010). Stimuli consisted of black-and-white
photographs of faces that were varied in age and balanced
for age and sex across experimental conditions. Each
face presented during initial encoding was tested during
subsequent retrieval, and participants were scanned
during both encoding and retrieval memory phases (see
Figure 1 for schematic representation of the task). A
detailed description of the task paradigm used in the
current study can be found in previous studies from our
laboratory (Ankudowich et al., 2016, 2017).
Using a mixed rapid event-related design, participants
were scanned across 12 experimental runs while they
1502
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
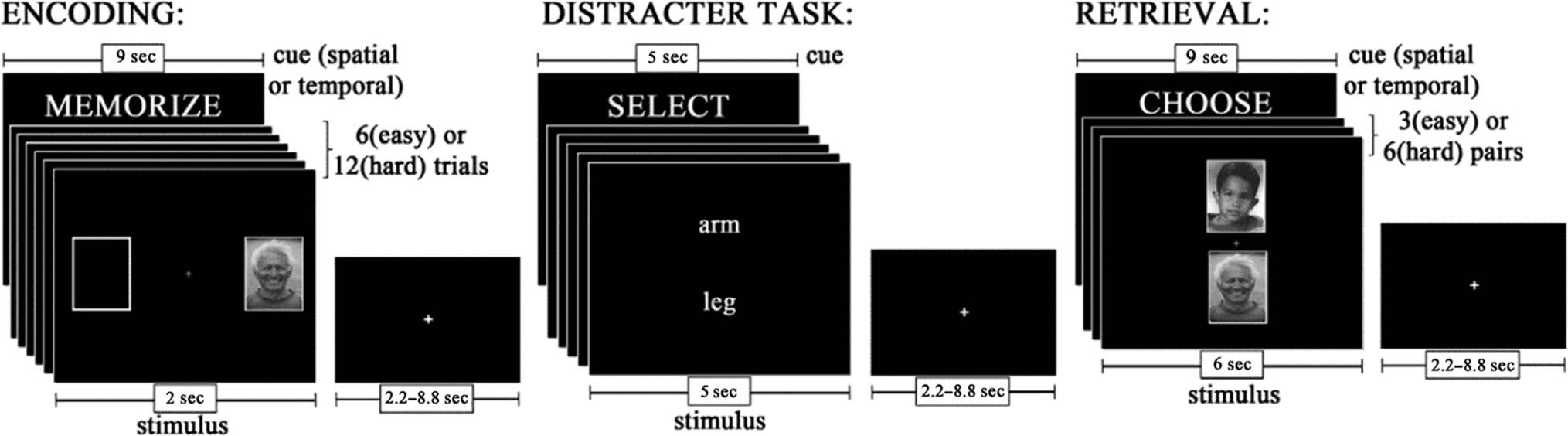
Figure 1. Task fMRI procedure.
encoded and retrieved the spatial and temporal details of
faces. Each run consisted of an “easy” temporal context
memory task (TE) and an “easy” spatial context memory
task (SE), and either a “hard” temporal context memory
task (TH) or a “hard” spatial context memory task (SH).
Easy and hard tasks differed in the number of stimuli to
be encoded: six encoding stimuli for “easy” tasks and 12
encoding stimuli for “hard” tasks. In total, there were 72
trials presented for each encoding event type (i.e., 288
trials total) and 36 trials presented for each retrieval event
type (i.e., 144 trials total). The current study focused on
the behavioral and fMRI data collected during the spatial
context memory tasks to compare our study findings with
our previous activation analyses using the same paradigm
(Subramaniapillai et al., 2019), and to further contextualize
our work with the substantial psychological literature
investigating sex differences in spatial episodic memory
(Sommer, Hildebrandt, Kunina-Habenicht, Schacht, &
Wilhelm, 2013; Young, Bellgowan, Bodurka, & Drevets,
2013; Bender, Naveh-Benjamin, & Raz, 2010; De Frias
et al., 2006; Weiss et al., 2003; Yonker, Eriksson, Nilsson,
& Herlitz, 2003; Gur & Gur, 2002; Herlitz et al., 1997). Our
choice to only focus on the spatial context memory task
further allows us to comprehensively address our aim of
investigating sex differences in performance-related func-
tional connectivity by comparing findings across several
sex-aggregated and -disaggregated B-PLS analyses. Please
refer to the work of Ankudowich et al. (2016, 2017) for
details regarding the temporal context memory tasks.
Herein, we present the details of the spatial context
memory tasks.
Encoding was intentional, and at the start of each
encoding phase, participants were cued (9 sec) to memo-
rize the spatial location (whether a face appeared on the
LEFT or the RIGHT during encoding) of the faces and to
the level of task difficulty. At encoding, each face was pre-
sented (2 sec) on either the left or the right of a central
fixation cross. There was a variable intertrial interval
(ITI) of 2.2–8.8 sec. During encoding, participants were
instructed to rate the pleasantness of each face. Partici-
pants pressed a button with their right thumb to indicate
a pleasant response and a button with their left thumb to
indicate a neutral response using an MRI-compatible fiber
optic response box. Between encoding and retrieval mem-
ory phases, participants performed a 1-min distractor task
in which they were required to reverse alphabetize two
words presented centrally on the computer screen. The
distractor task was used to deter participants from actively
rehearsing the encoding stimuli.
Following the distractor task, participants were pre-
sented with task instructions for retrieval (9 sec) to remind
them of the spatial context task demands. During retrieval,
participants were presented with pairs of previously
encoded faces for 6 sec. One of the faces was presented
above a central fixation cross, and the other was presented
below. During the easy versions of the retrieval task, par-
ticipants viewed three pairs of faces, and during the hard
versions of the retrieval task, they viewed six pairs of faces.
There was a variable ITI of 2.2–8.8 sec between retrieval
events. For the spatial task, participants were asked to indi-
cate which of the two faces was originally presented on the
LEFT/RIGHT. Participants pressed a button under their
right thumb to indicate a face at the top of the screen,
and they pressed a button under their left thumb to indi-
cate a face at the bottom of the screen. Therefore, fMRI
task-related activation for the spatial context memory par-
adigm was collected for four different event types in this
experiment: encoding spatial easy (eSE), encoding spatial
hard (eSH), retrieval spatial easy (rSE), retrieval spatial
hard (rSH).
Task fMRI Imaging Methods
Structural and fMRI data were collected at the Douglas
Institute Brain Imaging Centre. Participants lied supine
in a 3-T Siemens Magnetom Trio scanner and wore a
standard 12-channel head coil. T1-weighted anatomical
images were first acquired for each participant at the start
of the scanning session using a 3-D magnetization pre-
pared rapid gradient echo sequence (repetition time =
2300 msec, echo time = 2.98 msec, flip angle = 9°, field
of view = 256, one hundred seventy-six 1-mm sagittal
Subramaniapillai et al.
1503
slices, 1 × 1 × 1 mm voxels). BOLD images were acquired
with a single-shot T2*-weighted gradient EPI pulse
sequence (repetition time = 2000 msec, echo time =
30 msec, field of view = 256, matrix size = 64 × 64,
in-plane resolution 4 × 4 mm, 32 oblique slices per
whole-brain volume) while participants performed the
context memory tasks. Visual task stimuli were back-
projected onto a screen in the scanner bore using E-Prime
software, and participants requiring correction for visual
acuity wore plastic corrective lenses. A variable ITI (2.2–
8.8 sec) was introduced to add jitter to event-related
acquisitions.
fMRI Basic Preprocessing
Reconstructed images were preprocessed in SPM Version
8 software. For each participant, the origin of functional
images was reoriented to the anterior commissure of that
individual’s acquired T1-weighted structural image. All
functional images were then realigned to the first image,
and motion artifacts were corrected using a 6° rigid-body
transformation (three translation and three rotational
parameters). Any experimental run in which within-run
motion exceeded 1.5 mm was excluded from analysis. In
total, 22 runs (1.2%) were excluded: 12 runs because of
task noncompliance (e.g., failure to record participant
responses, issues with the response box), 6 runs because
of frontal/medial BOLD signal loss after fMRI preprocess-
ing, 2 runs because of poor volumes, 2 runs because of
scanner failure, and none because of excessive motion.
Functional images were then normalized to an Montreal
Neurological Institute EPI template and resliced at 4 ×
4 × 4 mm voxel resolution and smoothed with an 8-mm
FWHM isotropic Gaussian kernel. ArtRepair toolbox for
SPM8 (cibsr.stanford.edu/tools/ human-brain-project
/artrepair-software.html) was used to correct slice artifacts
before realignment and volume artifacts after normaliza-
tion and smoothing (< 5% interpolated data). Any run
in which interpolated data exceeded 5% was excluded
from analysis.
Analysis
Behavioral Data Analysis
Spatial context retrieval accuracy and RT. Using R (R
Core Team, 2013), we conducted robust linear mixed-
effects regression (rlmer) models (using the robustlmm
package; Koller, 2016) in the full cohort to test the
three-way interaction between age, sex (2: men, women),
and task difficulty (2: easy, hard) on retrieval accuracy (%
correct) and RT (msec), respectively. The rlmer model is
similar to the lmer model (see the work of Bates, Mächler,
Bolker, & Walker, 2015, for the lme4 package details), but
additionally, it is robust to outliers by down-weighting the
impact of extreme measures on the model performance
(Koller, 2016). The models contained the random effect
of participants to account for the variability of participants’
performance between the easy and hard versions of the
spatial context task. The models used in terms of R syntax
for spatial retrieval accuracy and RT, respectively, were
as follows:
Spatial Retrieval Accuracy ∼ Age × Sex × Task
Difficulty + (1|Participant).
Spatial Retrieval Reaction Time ∼ Age × Sex × Task
Difficulty + (1|Participant).
The continuous variable of age was standardized using a
z-score transformation, whereas the variables of sex and
task difficulty were treated as categorical variables through
deviation coding (−1, 1).
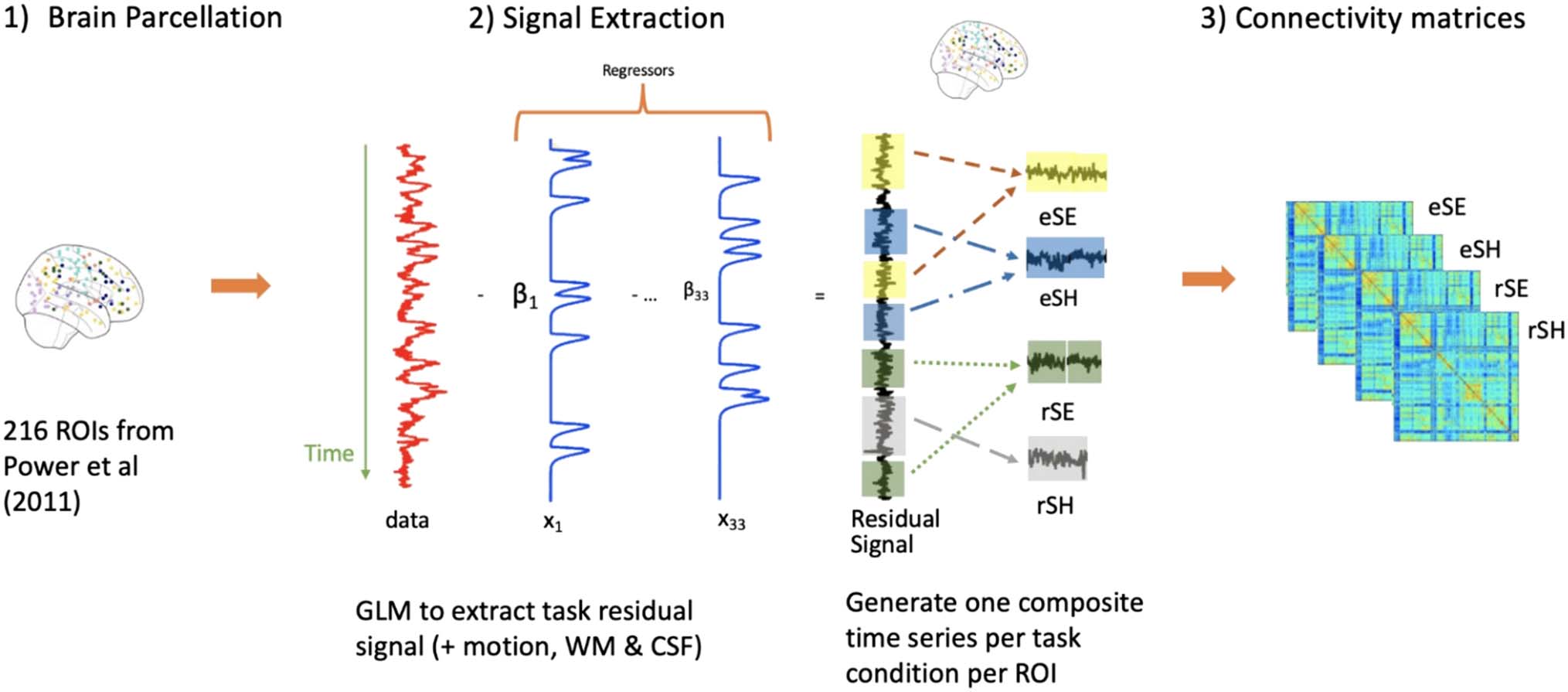
fMRI preprocessing for PLS connectivity analysis brain
parcellation. Figure 2 illustrates the preprocessing
steps used to generate the connectivity matrices for partic-
ipants across the four task conditions, which were subse-
quently submitted to the PLS analysis. Using SPM’s
MarsBaR toolbox, the average time series for 264 ROIs
defined by the Power et al. (2011) functional parcellation
atlas were extracted for each participant for all task-related
event types across the full experiment. Each ROI was reg-
istered from the 2 × 2 × 2 mm3 Power et al. atlas to the 4 ×
4 × 4 mm3 voxel resolution of our functional scans. To do
this, we took each ROI’s central coordinates from the
Power et al. (2011) ROIs and identified a 7-voxel sphere
surrounding the central coordinates. During this process
of scaling down to the 4 × 4 × 4 mm3 voxel resolution, we
eliminated ROIs with voxels that were not common to all
participants and/or overlapped with other ROIs. We also
excluded cerebellar ROIs because our fMRI acquisition
did not completely acquire these regions, and the uncer-
tain network ROIs because they did not belong to a major
functional system in the brain. We additionally combined
the memory retrieval network with the DMN because the
few nodes belonging to the memory retrieval network are
activated in cognitive functions (e.g., memory, imagina-
tion) commonly attributed to the DMN (Huo, Li, Wang,
Zheng, & Li, 2018). Thus, we identified a total of 216
unique ROIs assigned to nine brain networks: auditory,
cingulo-opercular task control network (CON), DMN,
DAN, fronto-parietal task control network (FPN), salience,
sensory/somatomotor network (SSM), visual attention
network ( VAN), visual (the list of Montreal Neurological
Institute coordinates and network affiliation can be
located through the Data Availability Statement below).
fMRI signal extraction. To examine task-related func-
tional connectivity, it is recommended that first the mean
task/event-related activity across the full experiment be
regressed out of the fMRI signal. This accounts for the con-
found of task-timing-driven statistical associations (Cole
et al., 2019). To this aim, event-related task activation for
all 216 ROIs was estimated using SPM’s general linear
model (GLM) with an ordinary least squares approach
(i.e., with AR(1) off ), using a high-pass filter set at
1504
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. The fMRI preprocessing steps involved (1) functional parcellation of each participant across the 216 unique ROIs from the Power et al.
atlas; (2) applying a GLM to extract the task residual signal after regressing 33 regressors to generate one composite time series per task condition for
each ROI; (3) generating four connectivity matrices for each task condition for every participant. GLM = general linear model; WM = white matter;
CSF = cerebrospinal fluid; eSE = encoding spatial easy; eSH = encoding spatial hard; rSE = retrieval spatial easy; rSH = retrieval spatial hard.
200 sec. This GLM consisted of 12 task-related regressors:
correct subsequent memory events for all experimental
tasks at encoding and retrieval, incorrect subsequent
memory responses for all encoding tasks, incorrect con-
text retrieval responses for all retrieval tasks, encoding
and retrieval task instructions, and distraction task. In addi-
tion, the six movement regressors generated by SPM dur-
ing motion correction, the mean white matter, and the
cerebrospinal fluid signals were also included as regres-
sors in the GLM to correct for physiological noise (Birn
et al., 2014). Finally, the temporal derivatives of the hemo-
dynamic response function for each of the task-related
regressors and the constant (i.e., intercept) resulted in a
total of 33 regressors used in the GLM. Thus, this one
GLM model was used to extract the mean residual time
series for each ROI per event type using the MarsBaR tool-
box in SPM (marsbar.sourceforge.net/).
Generating functional connectivity matrices. Because
the focus of our current analysis is the spatial version of the
task, we only generated functional connectivity matrices for
each event type of the spatial task. Each participant’s
residual time series were concatenated across similar event
types to generate composite time series for each event
type. The minimum length of time for a concatenated
event was 186 sec in the current study. Previous work has
established that a minimum length of 30 sec is sufficient for
reliable task-based connectivity analyses (e.g., Mohr et al.,
2016). As a measure of functional connectivity, we com-
puted Pearson correlations for each ROI with every other
ROI across the time series. Connectivity matrices were
created for each participant and event type from the
correlation coefficients, which then underwent Fisher
z-transformation. Thus, in total, each participant had four
connectivity matrices, one for each of the four event types
(i.e., eSE, eSH, rSE, and rSH) of size 216 × 216. There were
a total of 23,220 unique connections, after accounting for
the symmetry around the diagonal of the matrix.
PLS functional connectivity analysis. Behavioral multi-
variate partial least squares (B-PLS) connectivity analysis
was used to identify patterns of task-based functional
connectivity (McIntosh & Mišić, 2013), because of its
ability to simultaneously detect distributed patterns of
whole-brain connectivity that differ based on participants’
age, sex, and memory performance. We conducted two B-
PLS connectivity analyses. The first was a full group analy-
sis (B-PLS1), in which we examined how age and memory
performance in the full sample of adults (i.e., without
disaggregating by sex) related to task-based connectivity
during encoding and retrieval of SE and SH tasks. The
second was a between-sex (women, men) group B-PLS
analysis (B-PLS2), in which we explored sex differences in
age- and performance-related patterns of brain connectivity.
In the first analysis, connectivity matrices for each indi-
vidual were organized by task event type and then stored
in a single group-level fMRI connectivity matrix. In the
second analysis, the between-groups factor of sex was
included in the group-level fMRI connectivity matrices.
In both B-PLS analyses, normalized measures of partici-
pants’ age and retrieval accuracy were the behavioral
measures of interest. We orthogonalized our behavioral
vectors of age and accuracy to assess independent effects
of age and performance (consistent with Subramaniapillai
Subramaniapillai et al.
1505
et al., 2019; see also the work of Ankudowich et al., 2017).
That is, before the PLS analyses, we conducted a regres-
sion analysis where task-specific retrieval accuracy was
used to predict age to obtain an age-residual vector that
would be uncorrelated with retrieval accuracy. These
age-residual and retrieval accuracy vectors were then
stacked in the same manner as the fMRI data matrix for
each analysis, respectively (e.g., participant sex and by
event type for the between-sex group B-PLS). Given that
the retrieval accuracy behavioral vector did not have age
regressed from it, it allowed us to assess connectivity asso-
ciated with age-related performance effects, whereas the
age residual allowed us to assess age effects orthogonal
to performance effects. The following steps would be
identical for both analyses, so they are presented once.
The stacked fMRI data matrix was then cross-correlated
with the similarly stacked behavioral vectors. The result-
ing cross-correlation matrix was submitted to singular
value decomposition. Singular value decomposition
re-expresses the matrix as a set of orthogonal singular
vectors or latent variables (LVs). Each LV consists of a
singular value that reflects the proportion of matrix
accounted for by that LV, and a pair of vectors (a left
singular vector consisting of the behavioral weights and
a right singular vector consisting of the connectivity
weights) that reflect a symmetrical relationship between
the pattern of whole-brain connectivity and the experi-
mental design/behavior measures. The profile of behav-
ioral weights shows how the behavioral vectors of age
and retrieval accuracy are correlated to the pattern of
whole-brain connectivity identified in the singular vector
of connectivity weights. The connectivity weights identify
the collection of edges that, as a group, are maximally
related to the behavioral weights.
Significance testing for the LVs was done using 500 per-
mutations ( p < .05). The permutation test assesses
whether the functional networks and behavioral profiles
are more strongly associated with one another than
expected by chance. Bootstrap resampling was performed
to assess the reliability of each of the edges (500 boot-
straps, bootstrap ratio threshold was set at 95th percentile,
p < .001). Connectivity edge contribution was estimated
with edge loadings, which is calculated as the correlation
of the participants’ PLS-derived brain score pattern with
their stacked connectivity matrices. The pattern of edge
loadings (i.e., correlations) is referred to as the loading
matrix and reflects whether edges are more positively or
negatively associated with the behavioral weights. A posi-
tive correlation coefficient in the loading matrix indicates a
positive association with positive behavioral weights. Con-
versely, a negative correlation coefficient in the loading
matrix is positively associated with the negative behavioral
weights. Because the relationship between the behavioral
weights and the loading matrix (i.e., connectivity weights)
is symmetric, the inverse is also true. That is, a positive
correlation coefficient indicates a negative association
with negative behavioral weights and vice versa.
RESULTS
Behavioral Results
Table 1 summarizes the participant demographic and neu-
ropsychological information across the age groups for the
full (n = 141) and sex-disaggregated sample (n = 49 men,
92 women). Behaviorally, the rlmer model investigating
the effects of age, sex, and task difficulty on memory
accuracy showed a main effect of Age (β = −0.03 [SE,
0.01]; t = −2.35, p < .05) and Task Difficulty (β =
−0.04 [SE, 0.01]; t = −3.00, p < .05). Younger adults
had greater accuracy than older adults on the tasks, and
generally, participants performed worse on the SH task
compared with the SE task. No other main effects or inter-
actions were significant.
There were also significant main effects of Age (β =
145.60 [SE = 68.71]; t = 2.12, p < .05) and Task Difficulty
(β = 130.23, [SE = 36.71]; t = 3.55, p < .05) on RT. Young
adults were faster than older adults across SE and SH tasks,
and participants took longer to respond to the SH task
than the SE task. No other main effects or interactions
were significant. Therefore, there were no sex differences,
nor Sex × Age interactions in task performance.
Functional Connectivity Results
Four participants’ fMRI images failed preprocessing and
were excluded from the PLS analyses (two women and
two men). Therefore, the sample size for the PLS analyses
was 137 (47 men and 90 women). Figures 3 through 6
depict the relevant information for the significant LVs in
both the full group B-PLS1 and the between-sex group
B-PLS2 analyses, respectively. The subplots include the
(1) thresholded loading matrix, (2) behavioral correlation
weights, (3) network density matrix, and (4) brain figure
representing the highly involved nodes. The thresholded
connectivity matrix (1) represents the 95th percentile of
the z-score values of correlation coefficients. The behav-
ioral weights (2) indicate how the loading matrix relates
to the behavioral vectors of age and accuracy in women
and men. The network density matrix (3) represents the
sum of the unthresholded significant edges divided by
the total number of possible edges between any two net-
works (or within a network). Each LV generated two den-
sity plots because calculations were done separately on the
positive and negative correlation coefficients. Density
matrices that produced sparse significant edges (< 5%)
were not included. Finally, the brain figures (4) identify
the most highly contributing nodes from the thresholded
loading matrix, as determined by the ranked sum of the
correlation values from most to least involved. Below,
we report the detailed findings of each B-PLS analysis.
Full Group B-PLS1 Results
The full group B-PLS1 analysis examining age and perfor-
mance effects in connectivity identified two significant
1506
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 1. Mean Demographic and Behavioral Measures (and Standard Errors)
Sample size (n)
141
137
49 – Total behavioral;
47 – fMRI Sample
92 – Total behavioral;
90 fMRI sample
Total Behavioral Sample
Total fMRI Sample
Men
Women
p
Age (years)
Educations (years)
Predicted full-scale IQ
BDI
CVLT-LFR
CVLT-LCR
CVLT-RG
BMI (kg/m2)
SE retrieval accuracy (% correct)
SH retrieval accuracy (% correct)
SE retrieval RT (msec)
SH retrieval RT (msec)
47.11 (1.41)
15.73 (0.18)
119.51 (0.44)
3.90 (0.32)a
13.17 (0.18)
13.43 (0.17)
15.33 (0.69)
24.26 (0.31)a
0.86 (0.01)
0.83 (0.01)
2474.95 (47.27)
2570.99 (43.85)
47.26 (1.44)
15.72 (0.18)
119.60 (0.44)
3.96 (0.32)a
13.19 (0.19)
13.46 (0.17)
15.36 (0.68)
24.25 (0.31)a
0.86 (0.01)
0.83 (0.01)
2488.80 (47.43)
2582.56 (43.94)
46.96 (2.44)
16.06 (0.27)
119.66 (0.73)
3.84 (0.53)
12.35 (0.36)
12.76 (0.30)
15.29 (0.11)
24.49 (0.39)
0.85 (0.01)
0.80 (0.02)
2417.32 (72.44)
2550.29 (72.25)
47.20 (1.73)
15.55 (0.23)
119.43 (0.56)
3.93 (0.40)a
13.61 (0.19)
13.78 (0.20)
15.36 (0.09)
24.14 (0.43)a
0.86 (0.01)
0.84 (0.01)
2505.31 (61.36)
2582.92 (55.35)
ns
ns
ns
p < .05b,d
p < .05b,d
ns
p < .001c
p < .001d
p < .001d
p < .001d
p < .001d
The breakdown of demographics between sexes relates to the full behavioral sample (n = 141). BDI = Beck Depression Inventory; CVLT = California Verbal Learning Test; LFR = long-form free recall;
LCR = long-form cued recall; ns = not significant; RG = recognition; BMI = body mass index; SE = Spatial Easy, SH = Spatial Hard.
a One participant had missing information. Values in brackets represent the standard error. A linear regression of Age × Sex was performed on each of the measures (significance of p < .05 used) on the
total sample (n = 141).
b The linear regression produced a significant effect of sex, such that women outperformed men on this score.
c Age × Sex interaction of BMI: Age-related increase in BMI; younger and middle-aged adult men had higher BMI than their female counterparts; and older men had higher BMI than older women.
d The linear regression produced a significant main effect of Age. The fMRI behavioral measures revealed that older adult participants performed significantly worse than younger and middle-aged par-
ticipants and with significantly greater RT to complete the spatial tasks.
S
u
b
r
a
m
a
n
i
a
p
i
l
l
a
i
e
t
a
l
.
1
5
0
7
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
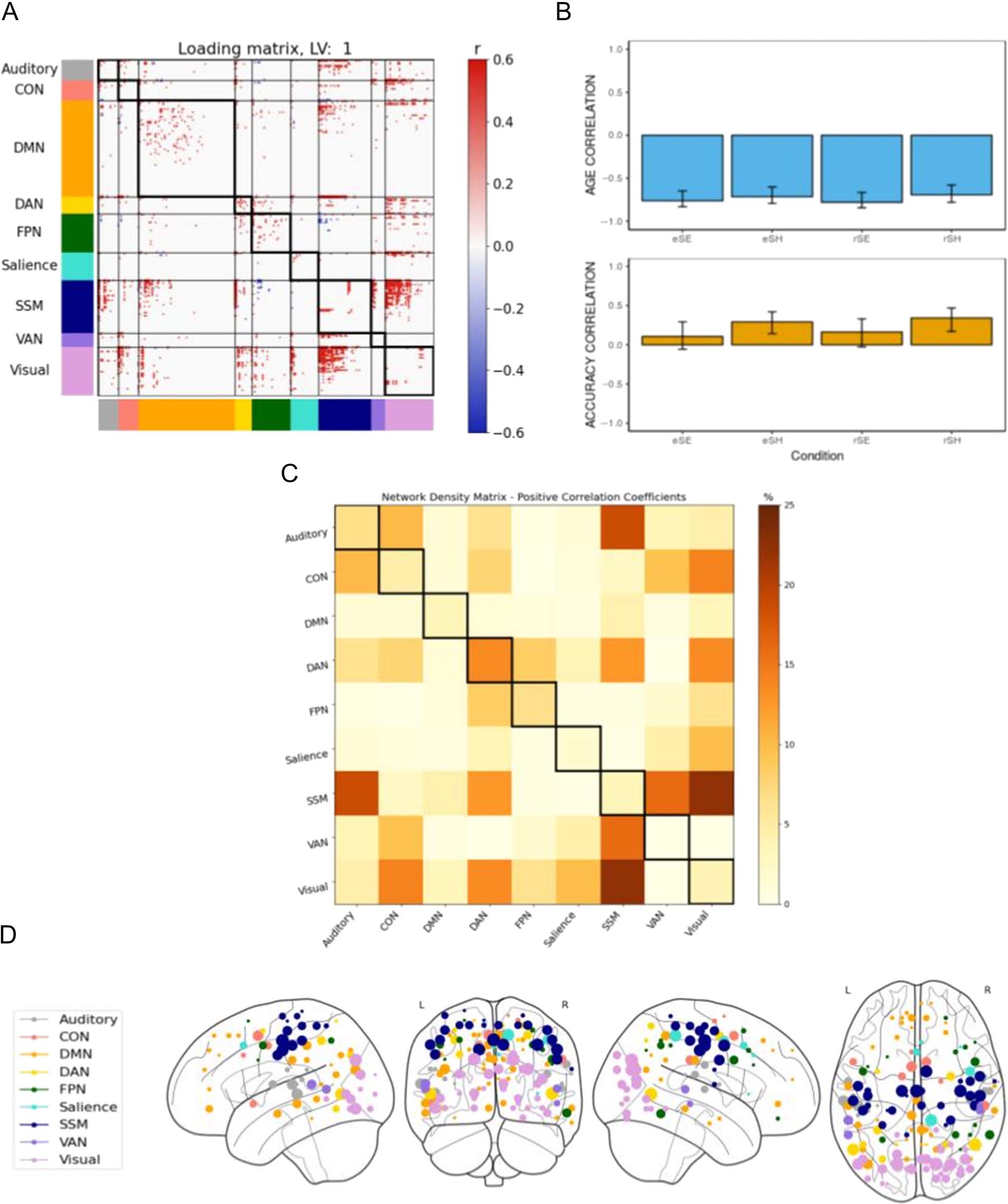
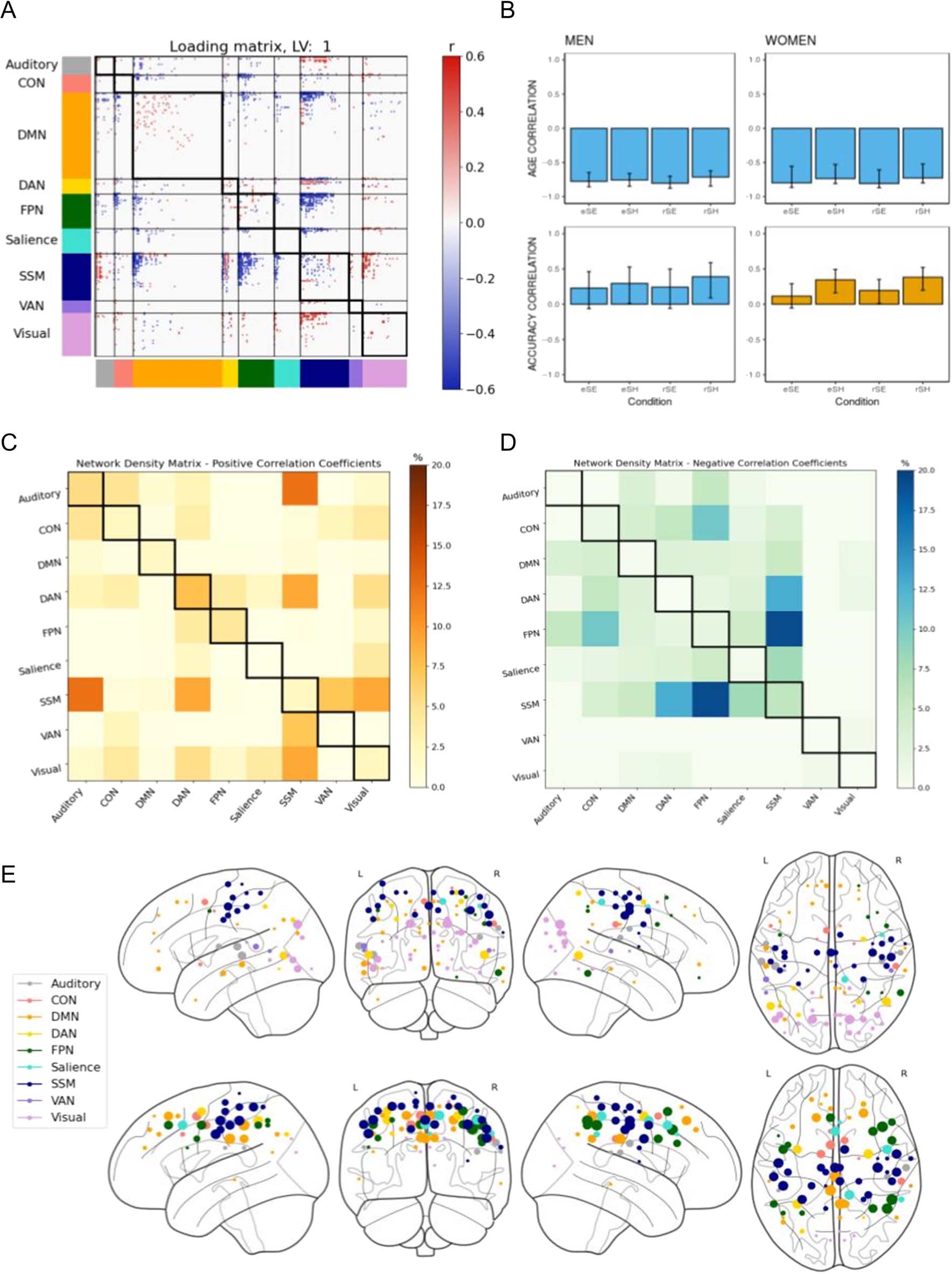
Figure 3. B-PLS1, LV1: Differential effects of age and accuracy on task-related brain connectivity. B-PLS1, LV1 reflects differences in how age and
accuracy on the task influence task-related brain connectivity. (A) Thresholded 95th percentile of correlations between participants’ task fMRI data
and behavioral profile indicated in (B). (B) Correlation between the behavioral vectors of age and accuracy with the task fMRI connectivity of
participants (behavior correlation weights). Error bars represent bootstrapped standard deviations. (C) The density plot for the positive correlation
coefficients (i.e., sum of the significant correlation coefficients after thresholding, divided by the total number of edges between any two networks).
The density matrix for the negative correlation coefficients is not presented because there were no significant edges. (D) Most densely connected
nodes from the positive salience loading matrix as represented by the rank sum of the correlation coefficients of the thresholded matrix. Greater
node size represents greater node involvement. eSE = encoding spatial easy; eSH = encoding spatial hard; rSE = retrieval spatial easy; rSH =
retrieval spatial hard; CON = cingulo-opercular network; DMN = default mode network; DAN = dorsal attention network; FPN = frontoparietal
network; SSM = somatomotor network; VAN = ventral attention network.
1508
Journal of Cognitive Neuroscience
Volume 34, Number 8
LVs at p < .05. The first LV (LV1, accounting for 70.15%
cross-block covariance) identified significant positive con-
nectivity weights (in red) between several networks
(Figure 3A).
The loading matrix and density matrix for LV1 (Figure 3A
and C) indicates that there were three dominant patterns
of positive connectivity involving the DAN, visual network,
and SSM network. First, LV1 identified positive within-
network connectivity weights in the DAN and FPN, and
between the DAN and FPN, SSM, and visual network. Sec-
ond, there was positive network connectivity between the
(i) visual network and CON, and (ii) SSM and the auditory
network and VAN. The matrices and behavioral correlation
weights (Figure 3B) together indicates that this pattern of
positive brain connectivity was negatively correlated with
age across all encoding and retrieval conditions and was
positively correlated with memory performance during
the hard spatial context memory task. Specifically, greater
positive functional connectivity among these networks
during the encoding and retrieval phases of the hard,
but not easy, spatial context memory task was positively
correlated with memory accuracy but negatively corre-
lated with age. Therefore, LV1 identified patterns of task-
related functional connectivity that differentiated age and
memory performance effects for the hard spatial context
memory tasks.
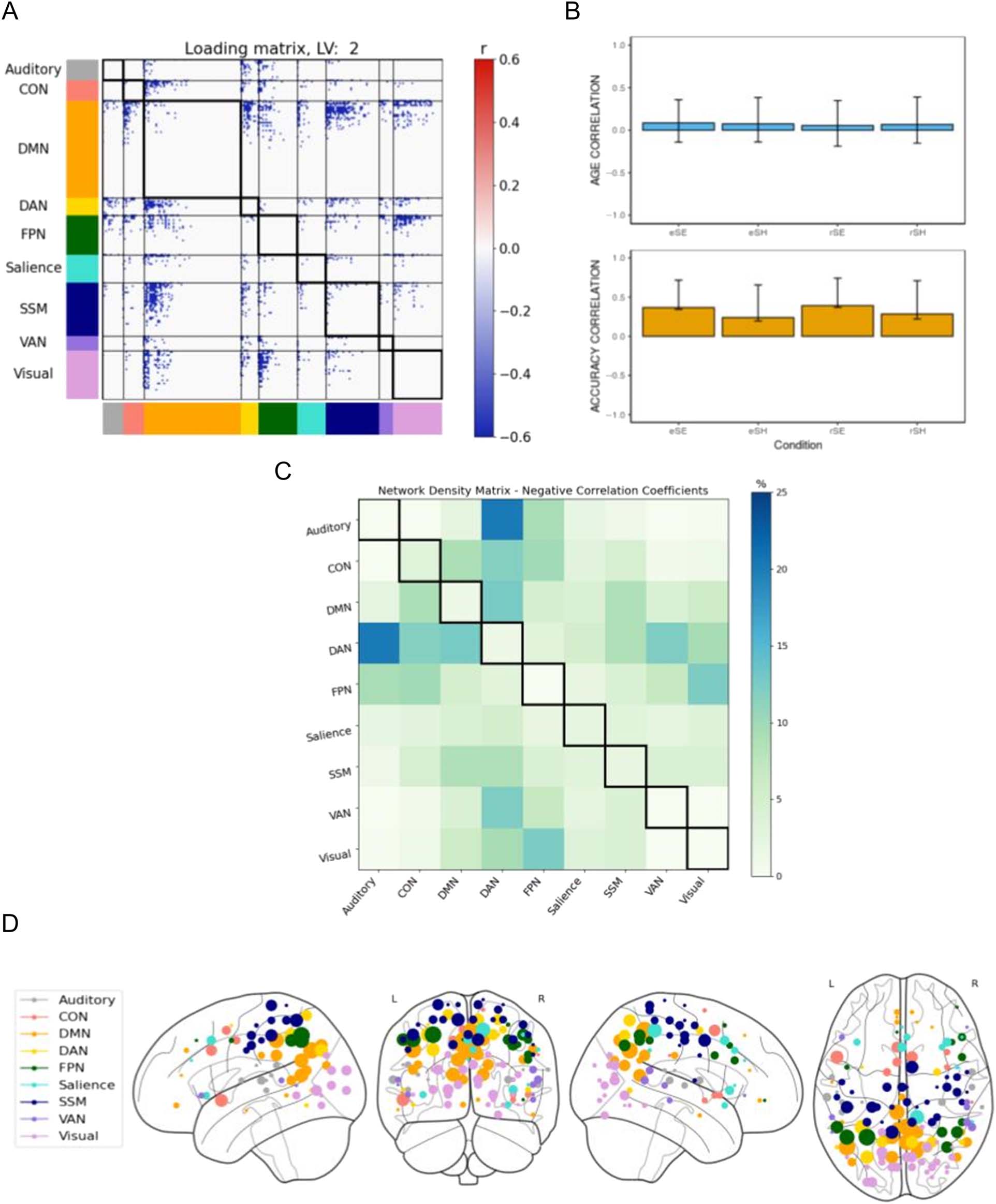
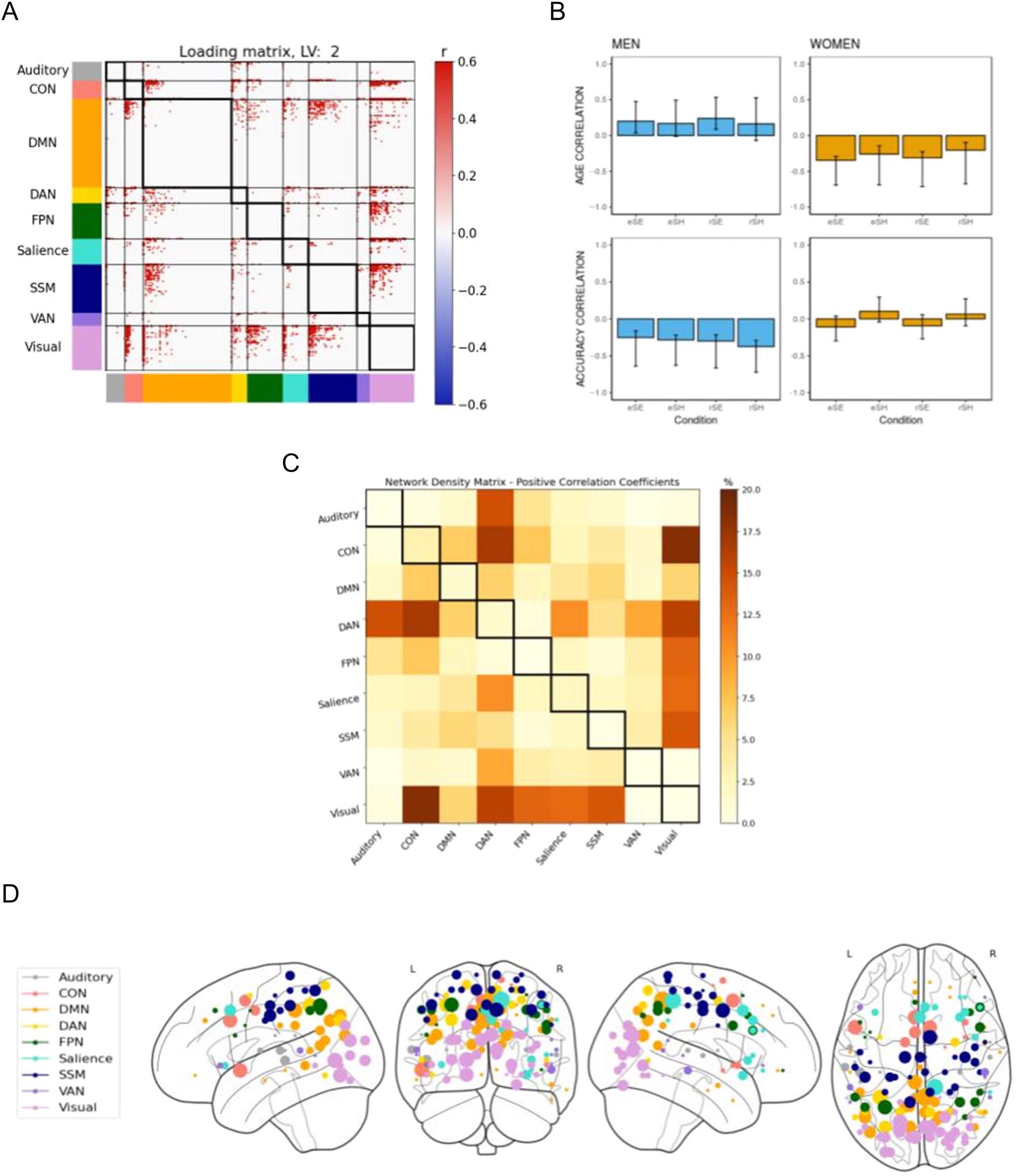
The second LV accounted for 17.47% cross-block covari-
ance and identified only significant negative connectivity
weights (in blue) as seen in the loading matrix (Figure 4A).
The density matrix (Figure 4C) identified dense patterns
of connectivity between DAN and auditory, CON, DMN,
and VAN. Taken together with the behavior correlation
weights (Figure 4B), these networks showed a negative
correlation with retrieval accuracy. That is, greater con-
nectivity between these networks during encoding and
retrieval was related to poorer performance for all
memory tasks.
Between-Sex Group B-PLS2 Results
The between-sex group B-PLS2 analysis examining age
and performance effects separately in women and men
identified four significant LVs at p < .05. Because LV1
and LV2 accounted for most of the original variance in data
(87.62%), we present and discuss the findings for LV1 and
LV2 as they would represent the most valuable information
with regard to sex differences in age and memory accuracy
on task-related functional connectivity (Zeng & Wang,
2010). The results and figures for LV3 and LV4 can be
located through the Data Availability Statement below.
LV1 accounted for 44.58% of cross-block covariance and
showed both significant positive and negative connectivity
weights. The behavior correlation plot indicates that the
patterns of connectivity identified by LV1 was differentially
correlated with age and memory performance during hard
spatial context memory tasks in men and women, recapit-
ulating the LV1 effect of the full group B-PLS1. The loading
and density matrices (Figure 5A, C, D) showed dense pos-
itive connections involving DAN, SSM, and visual net-
works, consistent with LV1 from the B-PLS1. However,
by disaggregating our connectivity analysis by sex, we
observed that the positive functional connectivity patterns
also support retrieval performance during easy spatial con-
text memory tasks in women only (i.e., the confidence
interval does not contain zero). Furthermore, a unique
pattern of negative weighted connectivity involving
CON, DAN, FPN, and SSM was also identified. In both
sexes, age was positively correlated with increased con-
nectivity between SSM and DAN, FPN, and between
CON and FPN, whereas memory performance during hard
spatial context memory tasks was negatively correlated
with this pattern of connectivity in both sexes, and during
easy spatial context retrieval in women only.
LV2 accounted for 21.66% of the cross-block covariance
and identified significant positive between-networks con-
nections involving DAN, SSM, and the visual network
(Figure 6A and C). The behavior correlation weights
(Figure 6B) indicates there were sex differences in how
age and memory performance correlated with this pattern
of task-related brain connectivity. In men, positive connec-
tivity among these networks was negatively correlated
with memory performance across all tasks, and age was
related to increased connectivity among these networks
only during easy spatial context memory tasks. In contrast,
in women, memory performance was not related to con-
nectivity among these networks, but age was negatively
correlated with connectivity in these networks across all
tasks. Therefore, LV2 identified sex differences in how
both age and memory performance correlated with task-
based brain connectivity.
Supplementary analyses. We performed several
post hoc analyses to account for confounding factors that
may have influenced the findings and subsequent inter-
pretation of our primary analyses. Information regarding
the supplementary material can be retrieved through the
Data Availability Statement below. First, sex differences in
education and intracranial volume (ICV ) may have
impacted our study findings. Men typically have larger
ICV than women (Ruigrok et al., 2014) and education level
may have a strong involvement as a gendered reserve con-
tributor (Subramaniapillai et al., 2021). Thus, we ran a sup-
plementary analysis using a subcohort (n = 48) of women
and men selected from our full sample matched according
to age, education, and ICV to determine whether the LV
patterns identified in our primary analyses were similar
after controlling for these factors. This supplementary anal-
ysis revealed similar findings as those presented in our pri-
mary analyses (this supplementary analysis revealed similar
findings as those presented in our primary analyses).
Second, whereas the choice to regress mean task-
related activity is grounded in previous literature (Cole
et al., 2019), we conducted supplementary B-PLS analyses
without regressing mean task-related activity to enable
Subramaniapillai et al.
1509
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. B-PLS1, LV2: Accuracy but not age-related effects on task-related brain connectivity. B-PLS1, LV2 reflects how accuracy was related to task-
related brain connectivity but not age. (A) Thresholded 95th percentile of correlations between participants’ task fMRI data and behavioral profile
indicated in (B). (B) Correlation between the behavioral vectors of age and accuracy with the task fMRI connectivity of participants (behavioral
correlation weights). Error bars represent bootstrapped standard deviations. (C) The density plot for the negative correlation coefficients (i.e., sum of
the significant correlation coefficients after thresholding, divided by the total number of edges between any two networks). The density matrix for the
positive correlation coefficients is not presented because there were no significant edges. (D) Most densely connected nodes from the negative
salience loading matrix as represented by the rank sum of the correlation coefficients of the thresholded matrix. Greater node size represents greater
node involvement. eSE = encoding spatial easy; eSH = encoding spatial hard; rSE = retrieval spatial easy; rSH = retrieval spatial hard; CON =
cingulo-opercular network; DMN = default mode network; DAN = dorsal attention network; FPN = frontoparietal network; SSM = somatomotor
network; VAN = ventral attention network.
1510
Journal of Cognitive Neuroscience
Volume 34, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. B-PLS2, LV1: Sex similarities in age and accuracy effects on task-related brain connectivity. B-PLS2, LV1 sex similarities in age and performance
on task-related brain connectivity. (A) Thresholded 95th percentile of correlations between participants’ task fMRI data and behavioral profile indicated in
(B). (B) Correlation between the behavioral vectors of age and accuracy with the task fMRI connectivity of participants (behavioral correlation weights).
Error bars represent bootstrapped standard deviations. (C) The density plot for the positive correlation coefficients (i.e., sum of the significant correlation
coefficients after thresholding, divided by the total number of edges between any two networks). (D) The density plot for the negative correlation
coefficients. (E) Most densely connected nodes from the positive (top) and the negative (bottom) salience loading matrix as represented by the rank sum
of the correlation coefficients of the thresholded matrix. Greater node size represents greater node involvement. eSE = encoding spatial easy; eSH =
encoding spatial hard; rSE = retrieval spatial easy; rSH = retrieval spatial hard; CON = cingulo-opercular network; DMN = default mode network; DAN =
dorsal attention network; FPN = frontoparietal network; SSM = somatomotor network; VAN = ventral attention network.
Subramaniapillai et al.
1511
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
c
n
_
a
_
0
1
8
6
8
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. B-PLS2, LV2: Sex differences in age and accuracy effects on task-related brain connectivity. B-PLS2, LV2 sex differences in age and
performance on task-related brain connectivity. (A) Thresholded 95th percentile of correlations between participants’ task fMRI data and behavioral
profile indicated in B. (B) Correlation between the behavioral vectors of age and accuracy with the task fMRI connectivity of participants (behavioral
correlation weights). Error bars represent bootstrapped standard deviations. (C) The density plot for the positive correlation coefficients (i.e., sum of
the significant correlation coefficients after thresholding, divided by the total number of edges between any two networks). (D) Most densely
connected nodes from the positive salience loading matrix as represented by the rank sum of the correlation coefficients of the thresholded matrix.
Greater node size represents greater node involvement. eSE = encoding spatial easy; eSH = encoding spatial hard; rSE = retrieval spatial easy;
rSH = retrieval spatial hard; CON = cingulo-opercular network; DMN = default mode network; DAN = dorsal attention network; FPN =
frontoparietal network; SSM = somatomotor network; VAN = ventral attention network.
1512
Journal of Cognitive Neuroscience
Volume 34, Number 8
readers to compare findings across differences in this
preprocessing methodology. The LV effects from this
supplementary analysis were consistent with our primary
analysis.
DISCUSSION
The goals of the current study were twofold. First, we used
B-PLS connectivity analysis to test the hypothesis that age
and memory performance (retrieval accuracy) would be
inversely associated with task-based connectivity between
the DAN, DMN, and FPN during successful encoding and
retrieval of face–location associations (spatial context
memory). We then disaggregated our analyses by self-
reported sex and tested the hypothesis that there would
largely be similarities in performance-related connectivity
in both sexes and sex differences in the effect of age on
memory performance-related brain connectivity, consis-
tent with our prior task-based activation analyses of sex dif-
ferences during spatial context memory (Subramaniapillai
et al., 2019). The behavioral data from the current study
replicated our prior work based on smaller sample sizes:
There was no significant effect of Sex on accuracy and
RT, nor any significant interactions of Age and Sex. There
were significant main effects of Age and Task Difficulty on
spatial context memory accuracy and RT, as reported
previously (Subramaniapillai et al., 2019; Ankudowich
et al., 2017).
The multivariate behavior PLS results from the full
group B-PLS1 and between-sex group (women, men)
B-PLS2 results generally corroborated our age-related
hypotheses. Age and memory performance were inversely
correlated to connectivity between DAN, FPN, and visual
networks in both sexes. Aging was also related to greater
between-networks integration among nonsensory net-
works, which was related to lower performance on hard
spatial context memory tasks in both sexes, and lower per-
formance during easy spatial context retrieval in women
only. However, our sex-related hypotheses were not sup-
ported. We observed both similarities and differences in
age-related and performance-related patterns of task-
based functional connectivity, which did not differ by
memory phase (encoding and retrieval). We discuss the
details of our connectivity results below and highlight
the importance of disaggregating task-based connectivity
results by sex and gender in computational and clinical
neuroscience studies of normative aging and episodic
memory function.
Sex Similarities in Age- versus Performance-related
Patterns of Task-based Connectivity during Spatial
Context Memory Encoding and Retrieval
In both B-PLS analyses, LV1 indicated that in both women
and men, better memory performance during hard spatial
context memory tasks was related to increased positive
connectivity: (i) between DAN and the FPN, SSM, and
visual networks; (ii) between SSM and the VAN, auditory,
and visual networks; and (iii) within the DAN and FPN dur-
ing encoding and retrieval phases of the hard spatial con-
text memory tasks. In contrast, age was associated with
decreased connectivity among these networks across all
task conditions in both sexes (B-PLS1, LV1 and B-PLS2,
LV1). This pattern of connectivity was correlated only with
memory performance during hard but not easy tasks,
which suggests increasing encoding load and retrieval
demands during the spatial context hard > easy tasks,
resulting in the engagement of several domain-general
cognitive control and attention-related brain networks
(cioè., DAN, FPN) to support memory performance. Questo
observation is consistent with prior brain activation stud-
ies that have highlighted the importance of attention and
cognitive control processes for successful episodic encod-
ing and retrieval (Smallwood et al., 2021; Ciaramelli &
Moscovitch, 2020), particularly for the memory of source
and/or contextual details (Thakral, Wang, & Rugg, 2015;
Dulas & Duarte, 2014; Rajah et al., 2010; Rajah, Ames, &
D’Esposito, 2008). Inoltre, we observed that across
encoding and retrieval, men and women exhibited similar-
ities in performance-related functional connectivity. Questo
indicates that successful memory performance during
the hard spatial context tasks relied on the reinstatement
of functional connections present at encoding, during the
later retrieval phase. This finding is consistent with current
theories emphasizing the importance of recapitulation of
cognitive/ brain states and episodic replay to support
retrieval success (Hill, King, & Rugg, 2021; Stawarczyk,
Wahlheim, Etzel, Snyder, & Zacks, 2020; Wimmer, Liu,
Vehar, Behrens, & Dolan, 2020; Morcom, 2014). More-
over, our current findings indicate this reinstatement
occurs at a broad network level and is associated with indi-
vidual differences in retrieval success. The finding that
greater DAN–FPN connectivity during encoding and
retrieval was correlated with better performance during
harder spatial context memory tasks and younger age is
consistent with prior studies that reported that FPN con-
nectivity with DAN supports episodic memory, and with
our hypothesis that age-related declines in episodic mem-
ory are related to reduced DAN–FPN connectivity (Avelar-
Pereira et al., 2017; Spreng et al., 2016; Benoit & Schacter,
2015; Habeck et al., 2012; Kim, 2012; Cabeza & St Jacques,
2007). Beyond these predicted results, our task fMRI con-
nectivity results highlight that the distinct pattern of con-
nectivity among the visual network, SSM, and higher-order
CON and DAN networks supported successful encoding
and retrieval during hard spatial context memory in both
women and men, and easy spatial context retrieval in
women. Greater sensory and SSM connectivity in both
sexes likely reflected the complex sensory-motor remap-
ping demands of the task. At encoding, stimuli were pre-
sented left/right; at retrieval, two old faces were oriented
top/bottom, but retrieval was based on a left/right decision
and response data were collected from a horizontally
Subramaniapillai et al.
1513
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
oriented response box. The face stimuli were vertically
presented at retrieval to avoid stimulus masking effects.
Tuttavia, this task design choice likely increased the stimulus-
response mapping demands of the spatial context memory
compito (Power et al., 2011). Così, age-related decreases in
these connectivity patterns may reflect reductions in the
ability to attend and integrate visual and sensorimotor
information with goal-directed cognitive control processes.
This may in turn have contributed to poorer memory func-
tion in both women and men. The observation that this
pattern of connectivity was only correlated with better per-
formance on hard tasks in both sexes is consistent with
prior studies showing modulation of frontoparietal cogni-
tive control processes as a function of task difficulty across
cognitive tasks, including episodic memory tasks (Rajah,
Crane, Maillet, & Floden, 2011; Kim, 2010; Rajah et al.,
2008; Vincent, Kahn, Snyder, Raichle, & Buckner, 2008; Cole
& Schneider, 2007; Dobbins & Han, 2006). È interessante notare, In
women, the correlation between connectivity and mem-
ory performance was also observed for easy spatial context
retrieval and points to a sex difference in task-related func-
tional connectivity that is discussed in greater detail below.
Sex Differences in the Performance-related
Task-based Connectivity during Easy Spatial
Context Retrieval
The full group and between-sex group PLS LV1 results
supported the hypothesis that aging in women and men
was related to declines in within-network segregation in
DAN and FPN. Tuttavia, only after disaggregating our
analysis by sex did we observe the predicted age-related
increase in between-networks connectivity (integration)
among nonsensory networks, questo è, CON, DMN, DAN,
FPN, salience, and SSM, across all task conditions in both
women and men (B-PLS2, LV1, negative connectivity
matrix). This pattern of connectivity was negatively corre-
lated with memory performance during hard spatial
context memory tasks in both sexes, and with memory
performance during easy spatial context memory tasks in
women only. Perciò, by disaggregating our analyses by
sex, we were able to identify sex differences in perfor-
mance effects related to easy spatial context retrieval.
This result indicates that the age effects identified in LV1
had a more general effect on memory performance in
older compared with younger women, but only affected
memory performance on hard spatial context memory
tasks in older compared with younger men. Inoltre,
it is possible that the between-networks integration
observed in the sex disaggregated, but not the full group,
analyses may have been driven by performance effects in
older women during the easy spatial context retrieval con-
ditions. We have previously observed greater generaliza-
tion in activation patterns across women, compared with
men, in the activation analysis of a smaller sample of adults
who participated in the current study (Subramaniapillai
et al., 2019) and in a sample of older adults with a family
history of late-onset AD (Rabipour et al., 2021). IL
current results shows that greater between-networks
integration was apparent at both levels of task difficulty
in women only and may reflect increased generalization
(or dedifferentiation) of function as women age (Chan
et al., 2014).
Sex Differences in Age- and Performance-related
Patterns of Task Connectivity
Based on prior rsfMRI connectivity studies (Zonneveld
et al., 2019; Avelar-Pereira et al., 2017; Jockwitz et al.,
2017; Klaassens et al., 2017; Ferreira et al., 2016; Spreng
& Schacter, 2012), we hypothesized that there would be
age-related increases in DAN–DMN task-based connectiv-
ity during encoding and retrieval, which would be
inversely correlated with memory performance. Entrambi
our full group B-PLS1, LV2, and between-sex group
B-PLS2, LV2 indicated that increased connectivity between
DAN and DMN during spatial context encoding and
retrieval was related to poorer memory performance.
Tuttavia, it was only after we disaggregated our analysis
by sex, we observed the predicted age effect—and only
in men. Specifically, men showed age-related increases
in DAN–DMN connectivity during easy spatial context
memory encoding and retrieval tasks, which was nega-
tively correlated to their memory performance. Men also
exhibited weak connectivity between DAN–FPN and an
increased connectivity pattern between DMN and the
auditory, CON, and visual networks. This suggests that
decoupling of DAN–FPN, greater DAN–DMN connectivity,
and greater connectivity between DAN and FPN with sen-
sory networks was correlated with men’s poorer episodic
encoding and retrieval. This result is consistent with the
hypothesis that suppression of DAN–DMN connectivity
and increased DAN–FPN connectivity during externally
oriented tasks, questo è, episodic memory tasks, supports
successful task performance (Smallwood et al., 2021;
Spreng & Turner, 2019), but highlights that this age-
related deficit in the suppression of DAN–DMN connectiv-
ity was specific to men in the current study. Inoltre,
these age- and performance-related differences in connec-
tivity in men suggests they may exhibit decreases in top–
down attentional control of visual processing with age that
was detrimental to performance (Esposito et al., 2018;
Grady et al., 2016; Vogel, Miezin, Petersen, & Schlaggar,
2012). This is also consistent with our prior activation anal-
ysis demonstrating that with advanced age, men engaged
visual sensory processing areas for successful memory per-
formanza, possibly relying on task strategies related to
semantic processing (Subramaniapillai et al., 2019).
Women, in contrast, exhibited an age-related decrease
in DAN–DMN connectivity and in DAN connectivity with
other networks. Inoltre, this age-related difference in
connectivity was not related to memory performance in
women. Così, age-related memory decline in women
in the current study was not associated with altered
1514
Journal of Cognitive Neuroscience
Volume 34, Numero 8
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
DAN–DMN connectivity. This was contrary to our hypoth-
esis that similar age effects would be observed in women
and men, and indicates that, in women, age-related spatial
context memory decline was primarily represented by the
effects observed in B-PLS2 LV1 (discussed above). More
broadly, our findings indicate there were sex differences
in DMN and DAN connectivity with age. This may be indic-
ative of different task orientations in older women, com-
pared with men (Ankudowich et al., 2017), or reflect sex
differences in the rate at which age effects functional
connectivity (Scheinost et al., 2015). Infatti, using
resting-state functional connectivity, Scheinost et al.
(2015) reported that between the ages of 18 E 65 years,
men exhibited steeper differences in DMN connectivity by
decade, compared with women. Given the fact that
age-related cognitive decline and neurodegenerative
diseases, questo è, AD has been linked to altered connectivity
involving the DMN (Hafkemeijer, van der Grond, &
Rombouts, 2012), future work should further explore if
there are sex differences in task-based DMN connectivity
in other memory paradigms, and at rest.
Caveats
This study examined sex similarities and differences in spa-
tial context memory across the lifespan using a novel func-
tional connectivity methodological approach. Tuttavia,
our study has several limitations that future work should
address. Primo, our findings are specific to the tasks
analyzed and future studies aimed at replicating results
in different episodic memory paradigms is essential to val-
idating the generalizability of our current finding. Secondo,
a comprehensive data collection approach was not used
when collecting participants’ biological sex or menopause
status. Our current study acquired participants’ biological
sex through self-report, although it could also be ascer-
tained through other means, including participants’ sex
hormone measurements. Hormone collection is espe-
cially relevant when investigating major life transitions,
such as menopause, which is associated with age-related
differences in women’s hormonal profiles. As a conse-
quence of women’s greater menopause-related hormonal
changes and the established literature of memory effects
during this transition (Rentz et al., 2017; Li et al., 2014;
Henderson, 2010; Yonker et al., 2006), we decided to omit
our cohort of women transitioning through menopause
and those who underwent HRT. Although our small
cohort size of women in the menopause transition pre-
vented us from including them in our primary analysis, Esso
is essential that future research integrate important life
transitions to better inform our understanding of healthy
aging models in women and men. Lastly, given that we did
not collect information about participants’ sociocultural
genere, it is further challenging to disentangle the effects
of biological sex and sociocultural gender on age- E
performance-related connectivity differences.
Inoltre, our relatively small cohort size constitutes
another limitation of the current study. Despite the small
cohort, our findings complement our previous activation
studies, both at the behavioral and functional level, using
the same lifespan cohort (Subramaniapillai et al., 2019;
Ankudowich et al., 2016, 2017). Inoltre, we found that
our PLS connectivity findings were robust to several meth-
odological confounds. Primo, one challenge that we foresaw
was that sex differences in ICV, with men typically having
greater ICV than women, may be driving our functional
connectivity results. Tuttavia, when we ran our analysis
on a smaller cohort of participants matched on ICV (E
age and education), our findings corroborate our primary
analysis.
Finalmente, although we have theoretical justification for
regressing task mean activity from the fMRI signal, uno
might rightfully ask what the error term actually means,
in terms of functional relevance. When we ran the PLS con-
nectivity analysis without regressing mean task-related
activity, the analysis generated the same exact LV results
and functional network connectivity with minimal differ-
ences observed in connectivity at the nodal rather than
network level. This enabled us to conclude that the level
of interpretation we used for the current study (cioè., at the
network level) would have resulted in the same interpre-
tations of findings, whether we chose to regress mean
task-related activity. Future work should endeavor to
understand what these minute differences mean at the
node level, both theoretically and conceptually. Così,
although there was the possibility of several confounds,
our supplementary analyses findings demonstrate our
primary analysis was robust to different preprocessing
strategies and methodological confounds.
Conclusions
The current study is the first to examine age- E
performance-related differences in task-based connectiv-
ity during episodic encoding and retrieval in a normative
adult lifespan sample and to explore how self-reported sex
affects these patterns of connectivity. In both sexes, age
and memory performance were inversely correlated with
DAN–FPN connectivity. Inoltre, we observed the pre-
dicted age-related increase in DAN–DMN connectivity but
only in men, whereas women showed more between-
networks integration and generalization of function with
advanced age. Così, different neurocognitive mechanisms
contribute to normative age-related differences in epi-
sodic memory in women and men. These sex and gender
differences should be considered when interpreting
task-related and resting-state fMRI studies of AD, E
other age-related neurological and psychiatric diseases
that have sex differences in prevalence rates and are
known to affect individuals’ episodic memory function
(cioè., Parkinson disease). Overall, our results highlight
the importance of considering sex and gender in study
Subramaniapillai et al.
1515
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
progetto, analysis, and interpretation in cognitive neurosci-
ence studies of aging and memory.
. o r g / 1 0 . 1 3 0 3 9 / 5 0 1 1 0 0 0 0 0 0 2 4 ) , g r a n t n u m b e r s :
201610PJT- 374992, GS9–171369.
Ringraziamenti
We thank all the research participants who made this work pos-
sible. This work was supported by CIHR Operating Grants
(GS9–171369 and 201610PJT- 374992) and NSERC Discovery
Grant (RGPIN-2018-05761) awarded to M.N. Rajah; Canada
Research Chair II to B. Misic; the Natural Science and Engi-
neering Research Council Graham Bell Canada Graduate
Scholarship-Doctoral and the Healthy Brains Healthy Lives
Doctoral Fellowship awarded to S. Subramaniapillai.
Reprint requests should be sent to M. Natasha Rajah, Room
2114 CIC Pavilion, Douglas Mental Health University Institute,
6875 LaSalle Blvd, Montreal, QC, Canada H4H 1R3, or via e-mail:
maria.rajah@mcgill.ca.
Author Contributions
M. N. Rajah (M. N. R.) designed the study. S. Subramaniapillai
(S. S.), S. Rajagopal (S. R.) and E. Ankudowich (E. A.) con-
tributed to data processing and analysis. S. Pasvanis (S. P.)
and E. UN. led data collection and quality control. S. S. E
S. R. created figures and tables. Bratislav Misic (B. M.) pro-
vided the PLS connectivity code, S. R. edited and created
the GitHub code used in the current publication. S. S.,
S. R., E. A., B. M., and M. N. R. provided analytic, theoretical
input and editorial feedback on drafts of this paper. E. UN.
wrote and earlier version of this manuscript focused on
the age effects; S. S. and M. N.R. co-wrote the current ver-
sion of the manuscript.
Sivaniya Subramaniapillai: Conceptualization; Data
curation; Formal analysis; Investigation; Methodology;
Validation; Visualization; Writing—Original draft;
Writing—Review & editing. Sricharana Rajagopal: Concep-
tualization; Formal analysis; Investigation; Methodology;
Software; Supervision; Validation; Visualization;
Writing—Original draft; Writing—Review & editing.
Elizabeth Ankudowich: Conceptualization; Formal analy-
sis; Investigation; Methodology; Validation; Visualization;
Writing—Original draft. Stamatoula Pasvanis: Data cura-
zione; Methodology; Project administration; Validation;
Visualization. Bratislav Misic: Formal analysis; Methodol-
ogy; Software; Validation; Visualization. M. Natasha Rajah:
Conceptualization; Data curation; Formal analysis;
Funding acquisition; Investigation; Methodology; Project
administration; Resources; Software; Supervision; Valida-
zione; Visualization; Writing—Original draft; Writing—
Review & editing.
Funding Information
M. Natasha Rajah, Natural Sciences and Engineering
Research Council of Canada (https://dx.doi.org/10.13039
/501100000038), grant number: RGPIN-2018-05761;
Canadian Institutes of Health Research (https://dx.doi
Data Availability Statement
The code used to run the functional connectivity analysis
can be retrieved from: https://tinyurl.com/tfcon. The sup-
plementary material for this paper can be retrieved from
https://tinyurl.com/fcsuppl.
Diversity in Citation Practices
Retrospective analysis of the citations in every article pub-
lished in this journal from 2010 A 2021 reveals a persistent
pattern of gender imbalance: Although the proportions of
authorship teams (categorized by estimated gender
identification of first author/ last author) publishing in
the Journal of Cognitive Neuroscience ( JoCN ) during this
period were M(an)/M = .407, W(oman)/M = .32, M/ W =
.115, and W/ W = .159, the comparable proportions for the
articles that these authorship teams cited were M/M =
.549, W/M = .257, M/ W = .109, and W/ W = .085 (Postle
and Fulvio, JoCN, 34:1, pag. 1–3). Consequently, JoCN
encourages all authors to consider gender balance
explicitly when selecting which articles to cite and gives
them the opportunity to report their article’s gender cita-
tion balance.
REFERENCES
Amer, T., Campbell, K. L., & Hasher, l. (2016). Cognitive
control as a double-edged sword. Trends in Cognitive
Scienze, 20, 905–915. https://doi.org/10.1016/j.tics.2016.10
.002, PubMed: 27863886
Ankudowich, E., Pasvanis, S., & Rajah, M. N. (2016). Changes in
the modulation of brain activity during context encoding vs.
context retrieval across the adult lifespan. Neuroimage, 139,
103–113. https://doi.org/10.1016/j.neuroimage.2016.06.022,
PubMed: 27311641
Ankudowich, E., Pasvanis, S., & Rajah, M. N. (2017). Changes in
the correlation between spatial and temporal source memory
performance and BOLD activity across the adult lifespan.
Cortex, 91, 234–249. https://doi.org/10.1016/j.cortex.2017.01
.006, PubMed: 28190516
Ankudowich, E., Pasvanis, S., & Rajah, M. N. (2019). Age-related
differences in prefrontal-hippocampal connectivity are
associated with reduced spatial context memory. Psychology
and Aging, 34, 251–261. https://doi.org/10.1037/pag0000310,
PubMed: 30407034
Asperholm, M., Van Leuven, L., & Herlitz, UN. (2020). Sex
differences in episodic memory variance. Frontiers in
Psychology, 11, 52–56. https://doi.org/10.3389/fpsyg.2020
.00613, PubMed: 32362856
Avelar-Pereira, B., Bäckman, L., Wåhlin, A., Nyberg, L., & Salami,
UN. (2017). Age-related differences in dynamic interactions
among default mode, frontoparietal control, and dorsal
attention networks during resting-state and interference
resolution. Frontiers in Aging Neuroscience, 9, 1–15. https://
doi.org/10.3389/fnagi.2017.00152, PubMed: 28588476
Bates, D., Mächler, M., Bolker, B. M., & Walker, S. C. (2015).
Fitting linear mixed-effects models using lme4. Diario di
1516
Journal of Cognitive Neuroscience
Volume 34, Numero 8
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Statistical Software, 67, 1–48. https://doi.org/10.18637/jss
.v067.i01
Becker, J. B., & Koob, G. F. (2016). Sex differences in animal
Modelli: Focus on addiction. Pharmacological Reviews, 68,
242–263. https://doi.org/10.1124/pr.115.011163, PubMed:
26772794
Bender, UN. R., Naveh-Benjamin, M., & Raz, N. (2010). Associative
deficit in recognition memory in a lifespan sample of healthy
adults. Psychology and Aging, 25, 940–948. https://doi.org/10
.1037/a0020595, PubMed: 20822256
Benoit, R., & Schacter, D. (2015). Specifying the core network
supporting episodic simulation and episodic memory by
activation likelihood estimation Roland. Neuropsychologia,
75, 450–457. https://doi.org/10.1016/j.neuropsychologia.2015
.06.034, PubMed: 26142352
Birn, R. M., Cornejo, M. D., Molloy, E. K., Patriat, R., Meier, T. B.,
Kirk, G. R., et al. (2014). The influence of physiological noise
correction on test–retest reliability of resting-state functional
connectivity. Brain Connectivity, 4, 511–522. https://doi.org
/10.1089/brain.2014.0284, PubMed: 25112809
Biswal, B., Zerrin Yetkin, F., Haughton, V. M., & Hyde, J. S.
(1995). Functional connectivity in the motor cortex of resting
human brain using echo-planar MRI. Magnetic Resonance in
Medicine, 34, 537–541. https://doi.org/10.1002/mrm
.1910340409, PubMed: 8524021
Cabeza, R., & St Jacques, P. (2007). Functional neuroimaging of
autobiographical memory. Trends in Cognitive Sciences, 11,
219–227. https://doi.org/10.1016/j.tics.2007.02.005, PubMed:
17382578
Campbell, K. L., & Schacter, D. l. (2016). Aging and the resting
state: Cognition is not obsolete. Language, Cognition and
Neuroscienza, 32, 692–694. https://doi.org/10.1080/23273798
.2016.1265658, PubMed: 28603744
Cansino, S. (2009). Episodic memory decay along the adult
lifespan: A review of behavioral and neurophysiological
evidence. International Journal of Psychophysiology, 71,
64–69. https://doi.org/10.1016/j.ijpsycho.2008.07.005,
PubMed: 18725253
Capogna, E., Sneve, M. H., Raud, L., Folvik, L., Ness, H. T.,
Walhovd, K. B., et al. (2022). Whole-brain connectivity during
encoding: Age-related differences and associations with
cognitive and brain structural decline. Cerebral Cortex,
bhac053. https://doi.org/10.1093/cercor/bhac053, PubMed:
35193146
Chan, M. Y., Park, D. C., Savalia, N. K., Petersen, S. E., & Wig,
G. S. (2014). Decreased segregation of brain systems
across the healthy adult lifespan. Atti del
National Academy of Sciences, U.S.A., 111, E4997–E5006.
https://doi.org/10.1073/pnas.1415122111, PubMed:
25368199
Ciaramelli, E., & Moscovitch, M. (2020). The space for memory
in posterior parietal cortex: Re-analyses of bottom–up
attention data. Neuropsychologia, 146, 107551. https://doi
.org/10.1016/j.neuropsychologia.2020.107551, PubMed:
32623010
Cole, M. W., Ito, T., Schultz, D., Mill, R., Chen, R., & Cocuzza, C.
(2019). Task activations produce spurious but systematic
inflation of task functional connectivity estimates.
Neuroimage, 189, 1–18. https://doi.org/10.1016/j.neuroimage
.2018.12.054, PubMed: 30597260
Cole, M. W., & Schneider, W. (2007). The cognitive control
rete: Integrated cortical regions with dissociable
functions. Neuroimage, 37, 343–360. https://doi.org/10.1016
/j.neuroimage.2007.03.071, PubMed: 17553704
Damoiseaux, J. S. (2017). Effects of aging on functional and
structural brain connectivity. Neuroimage, 160, 32–40.
https://doi.org/10.1016/j.neuroimage.2017.01.077, PubMed:
28159687
De Frias, C., Nilsson, l. G., & Herlitz, UN. (2006). Sex differences
in cognition are stable over a 10-year period in adulthood
and old age. Aging, Neuropsychology, and Cognition, 13,
574–587. https://doi.org/10.1080/13825580600678418,
PubMed: 16887790
Dixon, M. L., Andrews-Hanna, J. R., Spreng, R. N., Irving, Z. C.,
Mills, C., Girn, M., et al. (2017). Interactions between the
default network and dorsal attention network vary across
default subsystems, time, and cognitive states. Neuroimage,
147, 632–649. https://doi.org/10.1016/j.neuroimage.2016.12
.073, PubMed: 28040543
Dobbins, IO. G., & Han, S. (2006). Cue- versus probe-dependent
prefrontal cortex activity during contextual remembering.
Journal of Cognitive Neuroscience, 18, 1439–1452. https://
doi.org/10.1162/jocn.2006.18.9.1439, PubMed: 16989546
Dulas, M. R., & Duarte, UN. (2014). Aging affects the interaction
between attentional control and source memory: An fMRI
study. Journal of Cognitive Neuroscience, 26, 2653–2669.
https://doi.org/10.1162/jocn_a_00663, PubMed: 24800631
Edde, M., Dilharreguy, B., Theaud, G., Chanraud, S., Helmer, C.,
Dartigues, J. F., et al. (2020). Age-related change in episodic
memory: Role of functional and structural connectivity
between the ventral posterior cingulate and the parietal
cortex. Brain Structure and Function, 225, 2203–2218.
https://doi.org/10.1007/s00429-020-02121-7, PubMed:
32728934
Esposito, R., Cieri, F., Chiacchiaretta, P., Cera, N., Lauriola, M.,
Di Giannantonio, M., et al. (2018). Modifications in resting
state functional anticorrelation between default mode
network and dorsal attention network: Comparison among
young adults, healthy elders and mild cognitive impairment
patients. Brain Imaging and Behavior, 12, 127–141. https://
doi.org/10.1007/s11682-017-9686-y, PubMed: 28176262
Ferreira, l. K., Regina, UN. C. B., Kovacevic, N., Martin, M. D. G. M.,
Santos, P. P., Carneiro, C. D. G., et al. (2016). Aging effects
on whole-brain functional connectivity in adults free of
cognitive and psychiatric disorders. Cerebral Cortex, 26,
3851–3865. https://doi.org/10.1093/cercor/bhv190, PubMed:
26315689
Ferretti, M. T., Iulita, M. F., Cavedo, E., Chiesa, P. A., Dimech,
UN. S., Chadha, UN. S., et al. (2018). Sex differences in
Alzheimer disease—The gateway to precision medicine.
Nature Reviews Neurology, 14, 457–469. https://doi.org/10
.1038/s41582-018-0032-9, PubMed: 29985474
Finn, E. S. (2021). Is it time to put rest to rest? Trends in
Cognitive Sciences, 25, 1021–1032. https://doi.org/10.1016/j
.tics.2021.09.005, PubMed: 34625348
Fjell, UN. M., Sneve, M. H., Grydeland, H., Storsve, UN. B., Di
Lange, A.-M. G., Amlien, IO. K., et al. (2015). Functional
connectivity change across multiple cortical networks relates
to episodic memory changes in aging. Neurobiology of
Aging, 36, 3255–3268. https://doi.org/10.1016/j
.neurobiolaging.2015.08.020, PubMed: 26363813
Fox, M. D., Snyder, UN. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks.
Proceedings of the National Academy of Sciences, U.S.A.,
102, 9673–9678. https://doi.org/10.1073/pnas.0504136102,
PubMed: 15976020
Friston, K. J. (1994). Functional and effective connectivity
in neuroimaging: A synthesis. Human Brain Mapping, 2,
56–78. https://doi.org/10.1002/hbm.460020107
Grady, C. (2008). Cognitive neuroscience of aging. Annals of
the New York Academy of Sciences, 1124, 127–144. https://
doi.org/10.1196/annals.1440.009, PubMed: 18400928
Grady, C. L., & Craik, F. IO. (2000). Changes in memory
processing with age. Current Opinion in Neurobiology, 10,
224–231. https://doi.org/10.1016/S0959-4388(00)00073-8
Subramaniapillai et al.
1517
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Grady, C., Sarraf, S., Saverino, C., & Campbell, K. (2016). Age
differences in the functional interactions among the default,
frontoparietal control, and dorsal attention networks.
Neurobiology of Aging, 41, 159–172. https://doi.org/10.1016/j
.neurobiolaging.2016.02.020, PubMed: 27103529
Gur, R. E., & Gur, R. C. (2002). Gender differences in aging:
Cognition, emozioni, and neuroimaging studies. Dialogues
in Clinical Neuroscience, 4, 197–210. https://doi.org/10
.31887/DCNS.2002.4.2/rgur, PubMed: 22033483
Habeck, C., Risacher, S., Lee, G. J., Glymour, M. M., Mormino,
E., Mukherjee, S., et al. (2012). Relationship between baseline
brain metabolism measured using F-18 FDG PET and
memory and executive function in prodromal and early
Alzheimer’s disease. Brain Imaging and Behavior, 6,
568–583. https://doi.org/10.1007/s11682-012-9208-x, PubMed:
23179062
Hafkemeijer, A., van der Grond, J., & Rombouts, S. UN. R. B.
(2012). Imaging the default mode network in aging and
dementia. Biochimica et Biophysica Acta – Molecular Basis
of Disease, 1822, 431–441. https://doi.org/10.1016/j.bbadis
.2011.07.008, PubMed: 21807094
Henderson, V. W. (2010). Action of estrogens in the aging
brain: Dementia and cognitive aging. Biochimica et
Biophysica Acta – General Subjects, 1800, 1077–1083.
https://doi.org/10.1016/j.bbagen.2009.11.005, PubMed:
19913598
Herlitz, A., Nilsson, l. G., & Bäckman, l. (1997). Gender
differences in episodic memory. Memory & Cognition, 25,
801–811. https://doi.org/10.3758/BF03211324, PubMed:
9421566
Hill, P. F., King, D. R., & Rugg, M. D. (2021). Age differences
in retrieval-related reinstatement reflect age-related
dedifferentiation at encoding. Cerebral Cortex, 31, 106–122.
https://doi.org/10.1093/cercor/bhaa210, PubMed: 32829396
Huo, L., Li, R., Wang, P., Zheng, Z., & Li, J. (2018). The default
mode network supports episodic memory in cognitively
unimpaired elderly individuals: Different contributions to
immediate recall and delayed recall. Frontiers in Aging
Neuroscienza, 10, 6. https://doi.org/10.3389/fnagi.2018
.00006, PubMed: 29416508
Jack, C. R., Wiste, H. J., Weigand, S. D., Knopman, D. S., Vemuri,
P., Mielke, M. M., et al. (2015). Age, sex, and APOE (cid:1)4 effects
on memory, brain structure, and β-amyloid across the adult
life span. JAMA Neurology, 72, 511–519. https://doi.org/10
.1001/jamaneurol.2014.4821, PubMed: 25775353
Jockwitz, C., Caspers, S., Lux, S., Jütten, K., Schleicher, A.,
Eickhoff, S. B., et al. (2017). Age- and function-related
regional changes in cortical folding of the default mode
network in older adults. Brain Structure and Function, 222,
83–99. https://doi.org/10.1007/s00429-016-1202-4, PubMed:
26943919
Kim, H. (2010). Dissociating the roles of the default-mode,
dorsal, and ventral networks in episodic memory retrieval.
Neuroimage, 50, 1648–1657. https://doi.org/10.1016/j
.neuroimage.2010.01.051, PubMed: 20097295
Kim, H. (2012). A dual-subsystem model of the brain’s default
rete: Self-referential processing, memory retrieval
processes, and autobiographical memory retrieval.
Neuroimage, 61, 966–977. https://doi.org/10.1016/j
.neuroimage.2012.03.025, PubMed: 22446489
King, D. R., de Chastelaine, M., & Rugg, M. D. (2018).
Recollection-related increases in functional connectivity
across the healthy adult lifespan. Neurobiology of Aging, 62,
1–19. https://doi.org/10.1016/j.neurobiolaging.2017.09.026,
PubMed: 29101898
Klaassens, B. L., van Gerven, J. M. A., van der Grond, J., de Vos,
F., Möller, C., & Rombouts, S. UN. R. B. (2017). Diminished
posterior precuneus connectivity with the default mode
network differentiates normal aging from Alzheimer’s
disease. Frontiers in Aging Neuroscience, 9, 97. https://doi
.org/10.3389/fnagi.2017.00097, PubMed: 28469571
Koller, M. (2016). Robustlmm: An R package for robust
estimation of linear mixed-effects models. Diario di
Statistical Software, 75. https://doi.org/10.18637/jss.v075.i06
Kukolja, J., Goreci, D. Y., Onur, O. A., Riedl, V., & Fink, G. R.
(2016). Resting-state fMRI evidence for early episodic
memory consolidation: Effects of age. Neurobiology of
Aging, 45, 197–211. https://doi.org/10.1016/j.neurobiolaging
.2016.06.004, PubMed: 27459940
Kwon, D., Maillet, D., Pasvanis, S., Ankudowich, E., Grady, C. L.,
& Rajah, M. N. (2016). Context memory decline in middle
aged adults is related to changes in prefrontal cortex
function. Cerebral Cortex, 26, 2440–2460. https://doi.org/10
.1093/cercor/bhv068, PubMed: 25882039
Li, R., Cui, J., & Shen, Y. (2014). Brain sex matters: Estrogen in
cognition and Alzheimer’s disease. Molecular and Cellular
Endocrinology, 389, 13–21. https://doi.org/10.1016/j.mce
.2013.12.018, PubMed: 24418360
Maillet, D., & Rajah, M. N. (2014). Age-related differences in
brain activity in the subsequent memory paradigm: UN
meta-analysis. Neuroscience and Biobehavioral Reviews, 45,
246–257. https://doi.org/10.1016/j.neubiorev.2014.06.006,
PubMed: 24973756
McCarthy, M. M., Arnold, UN. P., Ball, G. F., Blaustein, J. D., & Di
Vries, G. J. (2012). Sex differences in the brain: The not so
inconvenient truth. Journal of Neuroscience, 32, 2241–2247.
https://doi.org/10.1523/JNEUROSCI.5372-11.2012, PubMed:
22396398
McIntosh, UN. R. (2000). From location to integration: How neural
interactions form the basis for human cognition. Memory,
Consciousness and the Brain: The Tallin Conference, 346–362.
McIntosh, UN. R., & Mišić, B. (2013). Multivariate statistical
analyses for neuroimaging data. Annual Review of
Psychology, 64, 499–525. https://doi.org/10.1146/annurev
-psych-113011-143804, PubMed: 22804773
Mesulam, M. (1990). Large-scale neurocognitive networks and
distributed processing for attention, lingua, and memory.
Neurological Progress, 28, 597–613. https://doi.org/10.1002
/ana.410280502, PubMed: 2260847
Mohr, H., Wolfensteller, U., Betzel, R. F., Mišić, B., Sporns, O.,
Richiardi, J., et al. (2016). Integration and segregation
of large-scale brain networks during short-term task
automatization. Nature Communications, 7. https://doi.org
/10.1038/ncomms13217, PubMed: 27808095
Mol, M., Carpay, M., Ramakers, I., Rozendaal, N., Verhey, F., &
Jolles, J. (2007). The effect of perceived forgetfulness
on quality of life in older adults; A qualitative review.
International Journal of Geriatric Psychiatry, 22, 393–400.
https://doi.org/10.1002/gps.1686, PubMed: 17044138
Mol, M. E. M., van Boxtel, M. P. J., Willems, D., & Jolles, J.
(2006). Do subjective memory complaints predict cognitive
dysfunction over time? A six-year follow-up of the Maastricht
aging study. International Journal of Geriatric Psychiatry,
21, 432–441. https://doi.org/10.1002/gps.1487, PubMed:
16676287
Morcom, UN. M. (2014). Re-engaging with the past: Recapitulation
of encoding operations during episodic retrieval. Frontiers
in Human Neuroscience, 8, 351. https://doi.org/10.3389
/fnhum.2014.00351, PubMed: 24904386
Naveh-Benjamin, M., Hussain, Z., Guez, J., & Bar-On, M. (2003).
Adult age differences in episodic memory: Further support
for an associative-deficit hypothesis. Journal of Experimental
Psychology: Apprendimento, Memory, and Cognition, 29, 826–837.
https://doi.org/10.1037/0278-7393.29.5.826, PubMed: 14516216
Nebel, R. A., Aggarwal, N. T., Barnes, l. L., Gallagher, A.,
Goldstein, J. M., Kantarci, K., et al. (2018). Understanding the
1518
Journal of Cognitive Neuroscience
Volume 34, Numero 8
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
impact of sex and gender in Alzheimer’s disease: A call to
action. Alzheimer’s and Dementia, 14, 1171–1183. https://
doi.org/10.1016/j.jalz.2018.04.008, PubMed: 29907423
Nordin, K., Nyberg, L., Andersson, M., Karalija, N., Riklund, K.,
Bäckman, L., et al. (2021). Distinct and common large-scale
networks of the hippocampal long axis in older age: Links
to episodic memory and dopamine D2 receptor availability.
Cerebral Cortex, 31, 3435–3450. https://doi.org/10.1093
/cercor/bhab023, PubMed: 33676372
Norman, M. A., Evans, J. D., Mugnaio, S. W., & Heaton, R. K.
(2000). Demographically corrected norms for the California
verbal learning test. Journal of Clinical and Experimental
Neuropsychology, 22, 80–94. https://doi.org/10.1076/1380
-3395(200002)22:1;1-8;FT080, PubMed: 10649547
Nyberg, l. (2017). Functional brain imaging of episodic memory
decline in ageing. Journal of Internal Medicine, 281, 65–74.
https://doi.org/10.1111/joim.12533, PubMed: 27453565
Nyberg, L., Lövdén, M., Riklund, K., Lindenberger, U., &
Bäckman, l. (2012). Memory aging and brain maintenance.
Trends in Cognitive Sciences, 16, 292–305. https://doi.org/10
.1016/j.tics.2012.04.005, PubMed: 22542563
Energia, J. D., Cohen, UN. L., Nelson, S. M., Wig, G. S., Barnes, K. A.,
Church, J. A., et al. (2011). Functional network organization of
the human brain. Neuron, 72, 665–678. https://doi.org/10
.1016/j.neuron.2011.09.006, PubMed: 22099467
Prakash, R. S., Heo, S., Voss, M. W., Patterson, B., & Kramer,
UN. F. (2012). Age-related differences in cortical recruitment
and suppression: Implications for cognitive performance.
Behavioural Brain Research, 230, 192–200. https://doi.org
/10.1016/j.bbr.2012.01.058, PubMed: 22348896
Rabipour, S., Rajagopal, S., Pasvanis, S., & Rajah, M. N. (2021).
Generalization of memory-related brain function in
asymptomatic older women with a family history of late onset
Alzheimer’s disease: Results from the PREVENT-AD cohort.
Neurobiology of Aging, 104, 42–56. https://doi.org/10.1016/j
.neurobiolaging.2021.03.009, PubMed: 33964608
Ragland, J. D., Coleman, UN. R., Gur, R. C., Glahn, D. C., &
Gur, R. E. (2000). Sex differences in brain–behavior
relationships between verbal episodic memory and resting
regional cerebral blood flow. Neuropsychologia, 38,
451–461. https://doi.org/10.1016/S0028-3932(99)00086-X,
PubMed: 10683395
Rahman, A., Schelbaum, E., Hoffman, K., Diaz, I., Hristov, H.,
Andrews, R., et al. (2020). Sex-driven modifiers of Alzheimer
risk: A multimodality brain imaging study. Neurology, 95,
E166–E178. https://doi.org/10.1212/ WNL.0000000000009781,
PubMed: 32580974
Rajah, M. N., Ames, B., & D’Esposito, M. (2008). Prefrontal
contributions to domain-general executive control processes
during temporal context retrieval. Neuropsychologia, 46,
1088–1103. https://doi.org/10.1016/j.neuropsychologia.2007
.10.023, PubMed: 18155254
Rajah, M. N., Crane, D., Maillet, D., & Floden, D. (2011).
Similarities in the patterns of prefrontal cortex activity during
spatial and temporal context memory retrieval after equating
for task structure and performance. Neuroimage, 54,
1549–1564. https://doi.org/10.1016/j.neuroimage.2010.09
.001, PubMed: 20837150
Rajah, M. N., Languay, R., & Valiquette, l. (2010). Age-related
changes in prefrontal cortex activity are associated with
behavioural deficits in both temporal and spatial context
memory retrieval in older adults. Cortex, 46, 535–549.
https://doi.org/10.1016/j.cortex.2009.07.006, PubMed:
19674742
Rajah, M. N., & McIntosh, UN. R. (2005). Overlap in the functional
neural systems involved in semantic and episodic memory
retrieval. Journal of Cognitive Neuroscience, 17, 470–482.
https://doi.org/10.1162/0898929053279478, PubMed: 15814006
R Core Team. (2013). A language and environment for
statistical computing. Vienna, Austria: R Foundation for
Statistical Computing.
Rentz, D. M., Weiss, B. K., Jacobs, E. G., Cherkerzian, S.,
Klibanski, A., Remington, A., et al. (2017). Sex differences in
episodic memory in early midlife: Impact of reproductive
aging. Menopause, 24, 400–408. https://doi.org/10.1097/GME
.0000000000000771, PubMed: 27824681
Ruigrok, UN. N. V., Salimi-Khorshidi, G., Lai, M. C., Baron-Cohen,
S., Lombardo, M. V., Tait, R. J., et al. (2014). A meta-analysis of
sex differences in human brain structure. Neuroscience and
Biobehavioral Reviews, 39, 34–50. https://doi.org/10.1016/j
.neubiorev.2013.12.004, PubMed: 24374381
Sala-Llonch, R., Peña-Gómez, C., Arenaza-Urquijo, E. M., Vidal-
Piñeiro, D., Bargalló, N., Junqué, C., et al. (2012). Brain
connectivity during resting state and subsequent working
memory task predicts behavioural performance. Cortex, 48,
1187–1196. https://doi.org/10.1016/j.cortex.2011.07.006,
PubMed: 21872853
Scheinost, D., Finn, E. S., Tokoglu, F., Shen, X., Papademetris,
X., Hampson, M., et al. (2015). Sex differences in normal age
trajectories of functional brain networks. Human Brain
Mapping, 36, 1524–1535. https://doi.org/10.1002/hbm.22720,
PubMed: 25523617
Smallwood, J., Bernhardt, B. C., Leech, R., Bzdok, D., Jefferies,
E., & Margulies, D. S. (2021). The default mode network in
cognition: A topographical perspective. Nature Reviews
Neuroscienza, 22, 503–513. https://doi.org/10.1038/s41583
-021-00474-4, PubMed: 34226715
Snyder, H. M., Asthana, S., Bain, L., Brinton, R., Craft, S., Dubal,
D. B., et al. (2016). Sex biology contributions to vulnerability
to Alzheimer’s disease: A think tank convened by the
Women’s Alzheimer’s research initiative. Alzheimer’s and
Dementia, 12, 1186–1196. https://doi.org/10.1016/j.jalz.2016
.08.004, PubMed: 27692800
Sommer, W., Hildebrandt, A., Kunina-Habenicht, O., Schacht,
A., & Wilhelm, O. (2013). Sex differences in face cognition.
Acta Psychologica, 142, 62–73. https://doi.org/10.1016/j
.actpsy.2012.11.001, PubMed: 23232336
Spaniol, J., Davidson, P. S. R., Kim, UN. S. N., Han, H.,
Moscovitch, M., & Grady, C. l. (2009). Neuropsychologia
event-related fMRI studies of episodic encoding and retrieval:
Meta-analyses using activation likelihood estimation.
Neuropsychologia, 47, 1765–1779. https://doi.org/10.1016/j
.neuropsychologia.2009.02.028, PubMed: 19428409
Sperling, R. (2007). Functional MRI studies of associative
encoding in normal aging, mild cognitive impairment, E
Alzheimer’s disease. Annals of the New York Academy of
Scienze, 1097, 146–155. https://doi.org/10.1196/annals.1379
.009, PubMed: 17413017
Sporns, O., & Betzel, R. F. (2016). Modular brain networks.
Annual Review of Psychology, 67, 613–640. https://doi.org/10
.1146/annurev-psych-122414-033634, PubMed: 26393868
Spreng, R. N., & Schacter, D. l. (2012). Default network
modulation and large-scale network interactivity in
healthy young and old adults. Cerebral Cortex, 22,
2610–2621. https://doi.org/10.1093/cercor/bhr339, PubMed:
22128194
Spreng, R. N., Stevens, W. D., Viviano, J. D., & Schacter, D. l.
(2016). Attenuated anticorrelation between the default
and dorsal attention networks with aging: Evidence from
task and rest. Neurobiology of Aging, 45, 149–160. https://
doi.org/10.1016/j.neurobiolaging.2016.05.020, PubMed:
27459935
Spreng, R. N., & Turner, G. R. (2019). The shifting architecture
of cognition and brain function in older adulthood.
Perspectives on Psychological Science, 14, 523–542. https://
doi.org/10.1177/1745691619827511, PubMed: 31013206
Subramaniapillai et al.
1519
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Stawarczyk, D., Wahlheim, C. N., Etzel, J. A., Snyder, UN. Z., &
Zacks, J. M. (2020). Aging and the encoding of changes in
events: The role of neural activity pattern reinstatement.
Proceedings of the National Academy of Sciences, U.S.A.,
117, 29346–29353. https://doi.org/10.1073/pnas.1918063117,
PubMed: 33229530
Strother, S. C., Kanno, I., Rottenberg, D. A., Friston, K. J., &
Ford, IO. (1995). Commentary and opinion: IO. Principal
component analysis, variance partitioning, and “functional
connectivity”. Journal of Cerebral Blood Flow and
Metabolism, 15, 353–377. https://doi.org/10.1038/jcbfm.1995
.44, PubMed: 7713992
Subramaniapillai, S., Almey, A., Natasha Rajah, M., & Einstein, G.
(2021). Sex and gender differences in cognitive and brain
reserve: Implications for Alzheimer’s disease in women.
Frontiers in Neuroendocrinology, 60. https://doi.org/10.1016
/j.yfrne.2020.100879, PubMed: 33137359
meta-analysis. Psychonomic Bulletin and Review, 14, 23–38.
https://doi.org/10.3758/BF03194024, PubMed: 17546728
Wang, L., LaViolette, P., O’Keefe, K., Putcha, D., Bakkour, A.,
Van Dijk, K. R. A., et al. (2010). Intrinsic connectivity between
the hippocampus and posteromedial cortex predicts memory
performance in cognitively intact older individuals.
Neuroimage, 51, 910–917. https://doi.org/10.1016/j
.neuroimage.2010.02.046, PubMed: 20188183
Weiss, E. M., Kemmler, G., Deisenhammer, E. A., Fleischhacker,
W. W., & Delazer, M. (2003). Sex differences in cognitive
functions. Personality & Individual Differences, 35, 863.
https://doi.org/10.1016/s0191-8869(02)00288-X
Wimmer, G. E., Liu, Y., Vehar, N., Behrens, T. E. J., & Dolan, R. J.
(2020). Episodic memory retrieval success is associated
with rapid replay of episode content. Nature Neuroscience,
23, 1025–1033. https://doi.org/10.1038/s41593-020-0649-z,
PubMed: 32514135
Subramaniapillai, S., Rajagopal, S., Elshiekh, A., Pasvanis, S.,
Sì, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R.,
Ankudowich, E., & Rajah, M. N. (2019). Sex differences in the
neural correlates of spatial context memory decline in healthy
aging. Journal of Cognitive Neuroscience, 31, 1895–1916.
https://doi.org/10.1162/jocn_a_01455, PubMed: 31393233
Thakral, P. P., Wang, T. H., & Rugg, M. D. (2015). Cortical
reinstatement and the confidence and accuracy of source
memory. Neuroimage, 109, 118–129. https://doi.org/10.1016
/j.neuroimage.2015.01.003, PubMed: 25583615
Tulving, E. (1972). Episodic and semantic memory. In E.
Tulving & W. Donaldson (Eds.), The curated reference
collection in neuroscience and biobehavioral psychology.
New York: Academic Press. https://doi.org/10.1016/B978-0-12
-809324-5.21037-7
Turner, G. R., & Spreng, R. N. (2012). Executive functions and
neurocognitive aging: Dissociable patterns of brain activity.
Neurobiology of Aging, 33, 326–e1. https://doi.org/10.1016/j
.neurobiolaging.2011.06.005, PubMed: 21791362
Uddin, l. Q., Sì, B. T. T., & Spreng, R. N. (2019). Towards a
universal taxonomy of macro-scale functional human brain
reti. Brain Topography, 32, 926–942. https://doi.org/10
.1007/s10548-019-00744-6, PubMed: 31707621
Vincent, J. L., Kahn, I., Snyder, UN. Z., Raichle, M. E., & Buckner,
R. l. (2008). Evidence for a frontoparietal control system
revealed by intrinsic functional connectivity. Diario di
Neurophysiology, 100, 3328–3342. https://doi.org/10.1152/jn
.90355.2008, PubMed: 18799601
Vogel, UN. C., Miezin, F. M., Petersen, S. E., & Schlaggar, B. l.
(2012). The putative visual word form area is functionally
connected to the dorsal attention network. Cerebral Cortex,
22, 537–549. https://doi.org/10.1093/cercor/bhr100, PubMed:
21690259
Voyer, D., Postma, A., Brake, B., & Imperato-McGinley, J.
(2007). Gender differences in object location memory: UN
Lashkari, D., Hollinshead, M., et al. (2011). The organization
of the human cerebral cortex estimated by intrinsic functional
connectivity. Journal of Neurophysiology, 106, 1125–1165.
https://doi.org/10.1152/jn.00338.2011, PubMed: 21653723
Yonker, J., Adolfsson, R., Eriksson, E., Hellstrand, M., Nilsson, l. G.,
& Herlitz, UN. (2006). Verified hormone therapy improves
episodic memory performance in healthy postmenopausal
women. Aging, Neuropsychology, and Cognition, 13,
291–307. https://doi.org/10.1080/138255890968655, PubMed:
16887775
Yonker, J. E., Eriksson, E., Nilsson, l. G., & Herlitz, UN. (2003).
Sex differences in episodic memory: Minimal influence of
estradiol. Brain and Cognition, 52, 231–238. https://doi.org
/10.1016/S0278-2626(03)00074-5, PubMed: 12821106
Young, K. D., Bellgowan, P. S. F., Bodurka, J., & Drevets, W. C.
(2013). Functional neuroimaging of sex differences in
autobiographical memory recall. Human Brain Mapping,
34, 3320–3332. https://doi.org/10.1002/hbm.22144, PubMed:
22807028
Zeng, Z., & Wang, J. (2010). Advances in neural network
research and applications (Vol. 67). Berlin: Springer Science
& Business Media.
Zhang, J., Andreano, J. M., Dickerson, B. C., Touroutoglou,
A., & Barrett, l. F. (2020). Stronger functional connectivity
in the default mode and salience networks is associated
with youthful memory in superaging. Cerebral Cortex,
30, 72–84. https://doi.org/10.1093/cercor/bhz071, PubMed:
31058917
Zonneveld, H. I., Pruim, R. H., Bos, D., Vrooman, H. A., Muetzel,
R. L., Hofman, A., et al. (2019). Patterns of functional
connectivity in an aging population: The Rotterdam Study.
Neuroimage, 189, 432–444. https://doi.org/10.1016/j
.neuroimage.2019.01.041, PubMed: 30659958
1520
Journal of Cognitive Neuroscience
Volume 34, Numero 8
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
j
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
4
8
1
5
0
0
2
0
5
7
1
3
2
/
/
j
o
C
N
_
UN
_
0
1
8
6
8
P
D
.
F
B
sì
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3