Adaptation to Binocular Anticorrelation Results
in Increased Neural Excitability
Reuben Rideaux, Elizabeth Michael, and Andrew E. Welchman
Astratto
■ Throughout the brain, information from individual sources
converges onto higher order neurons. Per esempio, informazione
from the two eyes first converges in binocular neurons in area V1.
Some neurons are tuned to similarities between sources of infor-
mazione, which makes intuitive sense in a system striving to match
multiple sensory signals to a single external cause—that is, estab-
lish causal inference. Tuttavia, there are also neurons that are
tuned to dissimilar information. In particular, some binocular neu-
rons respond maximally to a dark feature in one eye and a light
feature in the other. Despite compelling neurophysiological and
behavioral evidence supporting the existence of these neurons
[Katyal, S., Vergeer, M., Lui, S., Lui, B., & Engel, S. UN. Conflict-
sensitive neurons gate interocular suppression in human visual
cortex. Scientific Reports, 8, 1239, 2018; Kingdom, F. UN. A.,
Jennings, B. J., & Georgeson, M. UN. Adaptation to interocular
difference. Journal of Vision, 18, 9, 2018; Janssen, P., Vogels, R.,
Liu, Y., & Orban, G. UN. At least at the level of inferior temporal
cortex, the stereo correspondence problem is solved. Neuron, 37,
693–701, 2003; Tsao, D. Y., Conway, B. R., & Livingstone, M. S.
Receptive fields of disparity-tuned simple cells in macaque V1.
Neuron, 38, 103–114, 2003; Cumming, B. G., & Parker, UN. J.
Responses of primary visual cortical neurons to binocular dispar-
ity without depth perception. Nature, 389, 280–283, 1997], their
function has remained opaque. To determine how neural mech-
anisms tuned to dissimilarities support perception, here we use
electroencephalography to measure human observers’ steady-
state visually evoked potentials in response to change in depth
after prolonged viewing of anticorrelated and correlated
random-dot stereograms (RDS). We find that adaptation to an-
ticorrelated RDS results in larger steady-state visually evoked
potentials, whereas adaptation to correlated RDS has no effect.
These results are consistent with recent theoretical work sug-
gesting “what not” neurons play a suppressive role in supporting
stereopsis [Goncalves, N. R., & Welchman, UN. E. “What not”
detectors help the brain see in depth. Current Biology, 27,
1403–1412, 2017]; questo è, selective adaptation of neurons tuned
to binocular mismatches reduces suppression resulting in
increased neural excitability. ■
INTRODUCTION
It remains an important challenge in neuroscience to un-
derstand how the brain combines a pair of 2-D retinal im-
ages to support 3-D perception. Classically, this problem
has been framed as one of matching features between
the two eyes, questo è, solving the “stereoscopic correspon-
dence problem,” so that the depth of objects can be tri-
angulated (Julesz & Chang, 1976; Marr & Poggio, 1976).
This problem is nontrivial, as the number of “false matches”
(cioè., correspondences between features that do not origi-
nate from the same object) rapidly increases with the
number of to-be-matched elements.
Random-dot stereograms (RDSs) are frequently used to in-
vestigate binocular vision because of their ability to divorce
information about 2-D form from differences between the
two eyes. These stimuli are composed of many self-similar
caratteristiche, potentially posing a severe challenge to establishing
binocular correspondence. The classic framework for under-
standing stereopsis is to find correspondence by considering
a range of potential disparities and selecting the offset that
University of Cambridge
maximizes the image similarity between the two eyes
(Fleet, Wagner, & Heeger, 1996; Ohzawa, DeAngelis, &
Freeman, 1990). This makes intuitive sense; Tuttavia, some
disparity-selective neurons in V1 appear poorly optimized for
such a computation in that they respond maximally to differ-
ent images presented on the two retinae (Read & Cumming,
2007; Cumming & Parker, 1997). Inoltre, binocular neu-
rons can show tuning to images that are difficult to imagine
being produced in the real world. A prime example of this is
the test of neural function with anticorrelated RDSs (aRDSs)
in which the polarity of image features is reversed between
the two eyes. Unlike correlated RDSs (cRDSs), viewing
aRDS does not support reliable depth perception; neverthe-
less, some disparity-selective neurons in V1 respond strongly
to these stimuli. Despite empirical evidence supporting the
existence of these neurons in macaques (Janssen, Vogels,
Liu, & Orban, 2003; Tsao, Conway, & Livingstone, 2003;
Cumming & Parker, 1997) and humans (Katyal, Vergeer,
Lui, Lui, & Engel, 2018; Kingdom, Jennings, & Georgeson,
2018), their functional role remains opaque.
Recent theoretical work suggested a potential explana-
tion for neurons tuned to mismatched binocular fea-
tures. In their binocular likelihood model of stereopsis,
© 2019 Istituto di Tecnologia del Massachussetts. Published under a
Creative Commons Attribution 4.0 Internazionale (CC BY 4.0) licenza.
Journal of Cognitive Neuroscience 32:1, pag. 100–110
https://doi.org/10.1162/jocn_a_01471
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
T
F
.
/
o
N
0
5
M
UN
sì
2
0
2
1
Goncalves and Welchman (2017) suggested a simple de-
coding rule for binocular neurons: Information about
depth can be read out from a population of binocular
neurons where the decoding scheme is based on the
cross-correlation between the encoding receptive fields.
Under this scheme, the activity of a binocular neuron
can lead to increased excitation for a particular depth
interpretation or drive suppression of a specific depth
estimate. By reading out a population of binocular neu-
rons, it is possible to derive a likelihood estimate of the
depth of the scene. This provides a plausible explanation
for why neurons should respond to binocular correspon-
dences that do not relate to a single physical object in the
ambiente. In particular, the “what not” responses of
binocular neurons can be used to drive suppression of
unlikely interpretations of the scene. Despite this theoret-
ical promise, there is little empirical evidence for the role
of “what not” responses in the human visual system.
The idea that binocular mismatches are used to drive
suppression in visual cortex yields a distinct prediction
concerning the balance of excitation and inhibition fol-
lowing a period of adaption. In particular, adapting the
responses of units that drive suppression should lead
to less inhibition, thereby increasing the net excitation
of the cortex. To investigate the role of “what not” re-
sponses within the visual cortex, here we use electro-
encephalography to measure human observers’ brain
activity during and after prolonged viewing of aRDS.
Specifically, we measure steady-state visually evoked po-
tentials (SSVEP) in response to cRDS and aRDS, following
adaptation to either aRDS or cRDS. We find that, follow-
ing adaptation to aRDS, SSVEP amplitude in response to
cRDS increases relative to a preadaptation baseline.
These results are consistent with the idea that “what
not” responses play a suppressive role in supporting
stereopsis; questo è, selective adaptation of “what not” re-
sponses reduce suppression, resulting in increased neu-
ral excitability.
METHODS
Participants
Observers were recruited from the University of Cambridge,
had normal or corrected-to-normal vision, and were
screened for stereo deficits. Thirty-three right-handed hu-
man adults (eight men, age = 25.2 ± 4.8 years) participated
in the main experiment; Tuttavia, two were not included in
the analysis: One was unable to see depth in the stimulus,
and a hardware fault stopped acquisition midway through
the experiment for the other. Of the 31 participants in-
cluded in the analysis, 22 completed all experimental
conditions; the remaining nine did not participate in the
baseline condition. Twenty-two right-handed human adults
(five men, age = 25.4 ± 4.6 years) participated in the
control experiment. Experiments were approved by the
University of Cambridge Ethics Committee; all observers
provided written informed consent.
The sample sizes used in the experiments were se-
lected based on previous studies using similar tech-
niques to study stereopsis (Cottereau, McKee, Ales, &
Norcia, 2012; Cottereau, McKee, & Norcia, 2012).
Apparatus and Stimuli
Stimuli were generated in MATLAB (The MathWorks, Inc.)
using Psychophysics Toolbox and Eyelink Toolbox exten-
sions (Cornelissen, Peters, & Palmer, 2002; Brainard,
1997; Pelli, 1997; see psychtoolbox.org/). Binocular pre-
sentation was achieved using a pair of Samsung 2233RZ
LCD monitors (120 Hz, 1680 × 1050) viewed through
mirrors in a Wheatstone stereoscope configuration. IL
viewing distance was 50 cm, and participant head position
was stabilized using an eye mask, headrest, and chin rest.
Eye movement was recorded binocularly at 1 kHz using an
EyeLink 1000 (SR Research Ltd.).
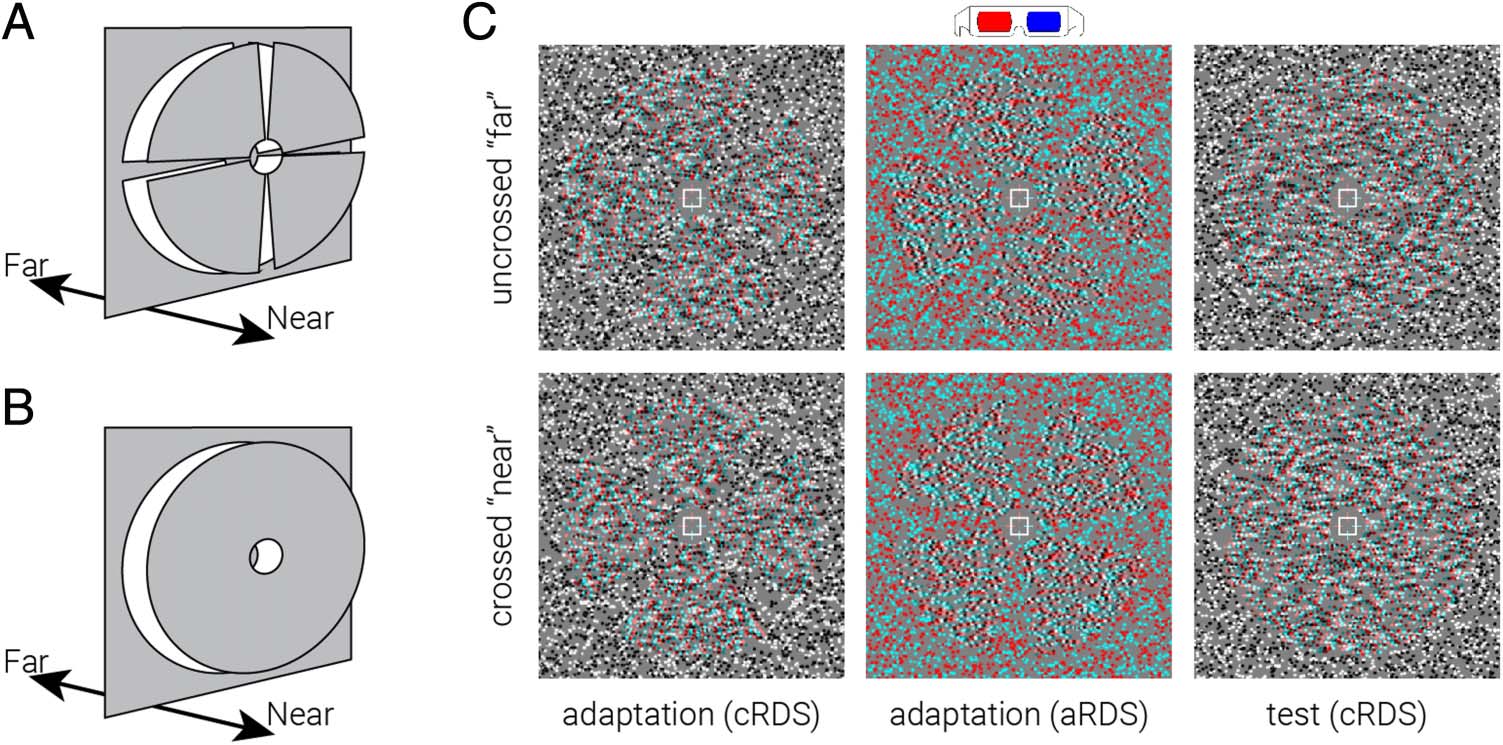
Adaptation stimuli consisted of RDS (12° × 12°) on a
mid-gray background surrounded by a static grid of black
and white squares intended to facilitate stable vergence.
Dots in the stereogram followed a black or white
Gaussian luminance profile, subtending 0.07° at half
maximum. There were 108 dots/deg2, resulting in ∼38%
coverage of the background. In the center of the stereo-
gram, four wedges were equally distributed around a cir-
cular aperture (1.2°), each subtending 10° in the radial
direction and 70° in polar angle, with a 20° gap between
wedges (Figure 1A). The wedge formation was used to
perceptually accentuate the near/far regions from the
surrounding zero disparity surface. Dots constituting
the wedges were offset by 10 arcmin between the left
and right eyes, and the remaining dots had zero offset.
This disparity was large enough to clearly distinguish
the near/far surface from the surrounding region while
still being comfortable to stereoscopically fuse for a pro-
longed period. Stimuli were presented for 1 sec and sep-
arated by 1-sec interstimulus intervals consisting of only
the background and fixation cross. On each presentation,
we applied a random polar rotation to the set of wedges
such that the disparity edges of the stimuli were in differ-
ent locations for each stimulus presentation (cioè., a rigid
body rotation of the four depth wedges together around
the fixation point). In every eight presentations, we re-
versed the sign of the disparity of the wedges (crossed
and uncrossed; Figure 1C). We balanced the presentation
of near and far disparities to ensure refixation on the zero
disparity reference plane, rather than promoting fixation
away from the reference plane. At a given time point, Tutto
wedges were presented the same disparity. In the center
of the wedge field, we presented a fixation square (side
length = 1°) paired with horizontal and vertical nonius
lines.
Test stimuli were similar to adaptation stimuli, except
Quello, instead of rotating wedges, an annulus was used
Rideaux, Michael, and Welchman
101
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
.
/
F
T
o
N
0
5
M
UN
sì
2
0
2
1
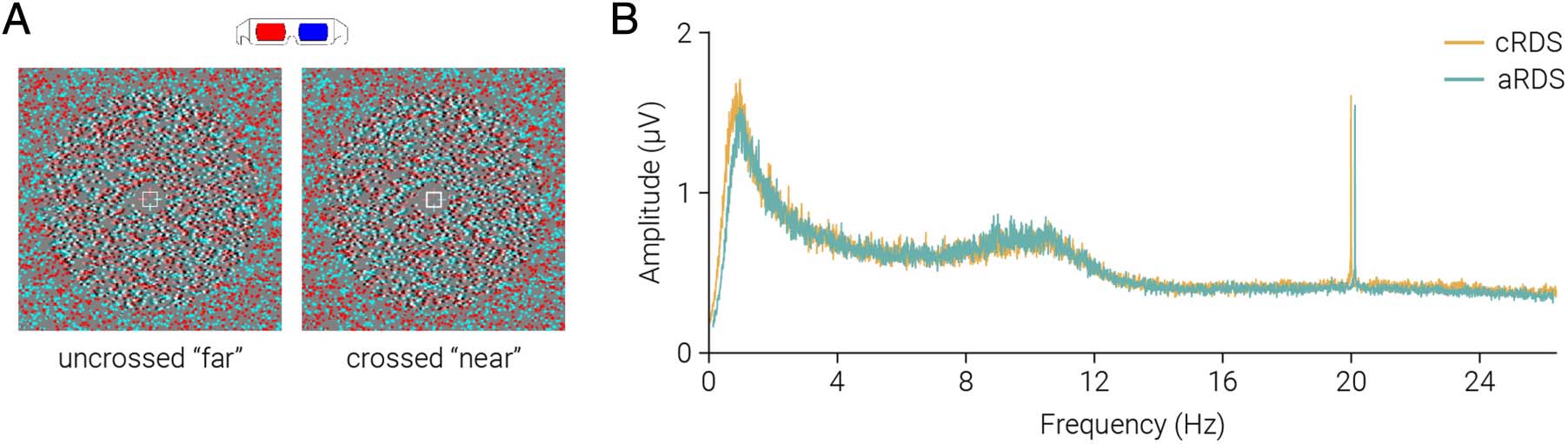
Figura 1. Adaptation and test
stimuli used in the experiment.
Diagram of the depth
arrangement of the (UN)
adaptation and (B) test stimuli.
(C) Example stimuli used in the
experiment, designed for red-
cyan anaglyph viewing.
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
F
.
T
o
N
0
5
M
UN
sì
2
0
2
1
(Figure 1B, C). The depth sign of the annulus was re-
versed at 4-Hz frequency, whereas all dots were regener-
ated at 20-Hz frequency.
Procedure
Participants underwent three different runs: an initial base-
line run, followed by correlated and anticorrelated runs
(counterbalanced across participants). The baseline run
consisted of five blocks of test stimulus presentations,
each lasting 25 sec, separated by 20-sec blank interblock
intervals. Correlated runs consisted of 12 adaptation
blocks, each followed by a 2-sec blank interblock interval
before a test block. Adaptation blocks consisted of 64 sec
of adaptation stimuli presentations (32 presentations to-
tal), and test blocks were identical to those in the baseline
run. Anticorrelated runs were identical to correlated runs,
except that the polarity of all dots in the left eye was re-
versed during the adaptation blocks. During adaptation
blocks, we instructed participants to fixate on the central
fixation square while performing a Vernier discrimination
task (Preston, Li, Kourtzi, & Welchman, 2008). During test
blocks, we instructed observers to maintain fixation and
limit blinks. cRDSs were used as the test stimulus for all
conditions, providing an equal test of the effects of adap-
tazione. Although viewing either aRDS or cRDS can produce
an electrophysiological response, cRDSs were used as the
test stimulus as they evoke a larger response than aRDSs
(Petrig, Julesz, Kropfl, Baumgartner, & Anliker, 1981;
Braddick et al., 1980), providing a better signal-to-noise ra-
tio (SNR) and thus a more sensitive measure of neural
change resulting from adaptation.
EEG
Electroencephalography data were acquired from all 33
participants with a 64-channel cap (BrainCap, Brain
Products GmbH). Data were recorded using BrainVision
Recorder software. Caps were fitted with 61 Ag/AgCl elec-
trodes positioned according to the standard 10–20 sys-
tem (Fp1 Fp2 F3 F4 C3 C4 P3 P4 O1 O2 F7 F8 T7 T8
P7 P8 Fz Cz Pz IO FC1 FC2 CP1 CP2 FC5 FC6 CP5 CP6
FT9 FT10 F1 F2 C1 C2 P1 P2 AF3 AF4 FC3 FC4 CP3
CP4 PO3 PO4 F5 F6 C5 C6 P5 P6 AF7 AF8 FT7 FT8 TP7
TP8 PO7 PO8 Fpz CPz POz Oz). Data were acquired with
a reference electrode at FCz. Electrooculograms were
also acquired, using two pairs of bipolar electrodes
placed horizontally and vertically around the left eye.
Data were high-pass filtered online at 0.1 Hz and ac-
quired with a 1-Hkz sampling rate. Temporal markers
were sent from the stimulus presentation computer to
mark the onset of the stimulus. These timings were vali-
dated using a pair of photodiodes attached to the two
stimulus presentation screens.
Preprocessing and analyses were performed in MATLAB
using the EEGLAB toolbox (Delorme & Makeig, 2004) E
custom in-house scripts. Data were first filtered offline with
a 1-Hz high-pass and a 40 Hz low-pass filter. For the SSVEP
analysis, each epoch was extracted around the test du-
ration to include a period of 29 sec (2 sec before the first
test stimulus onset to 2 sec after the offset of the final test
stimulus); così, there were 12 epochs per adaptation con-
dition and five in the baseline, all of which were included
in the analysis. For the ERP analysis, epochs were ex-
tracted around the stimulus onset to include a 1-sec pres-
timulus and 1-sec poststimulus period (384 epochs per
adaptation condition). The poststimulus period, Perciò,
did not include any data from the next stimulus presenta-
zione. Epochs used in the ERP analysis were visually in-
spected, and artifactual epochs were rejected (excluding
eye movements). For one participant, two channels (TP9
and TP10) were interpolated. All data were re-referenced
to an average reference across all channels, and then inde-
pendent component analysis (ICA) decomposition was ap-
plied for the purpose of artifact identification. Resultant
ICA components were visually inspected before rejection.
102
Journal of Cognitive Neuroscience
Volume 32, Numero 1
Only components reflecting eye movements and other
likely muscle artifacts were removed. These components
were identified by characteristic features in the compo-
nent time course and power spectrum, in addition to
their frontal topography.
There were 12 blocks in each adaptation run and five
blocks in the baseline run; così, we matched the length
of data in each calculation by including data from the first
half of the adaptation epochs (4–13.5 sec) E 22.8 sec of
data from the baseline epochs (2–24.8 sec). Questo è, we
included 12 blocks × 9.5 sec of data from the adaptation
runs (144 sec total) and five blocks × 22.8 sec of data
from the baseline run (144 sec total). We avoided in-
cluding data immediately from stimulus onset to avoid
contamination by any onset evoked potential. Fourier
transformation of the data was performed on the full
concatenated data recording; no window was applied to
the data before transformation.
Eye Tracking
Owing to the bespoke experimental setup (cioè., recording
eye position from behind one-way mirrors in a haploscope),
the eye tracker would occasionally fail to track participants’
eyes for an entire block. To draw within-participant com-
parisons, we only included the participants for whom data
were available for both experimental conditions in the eye
tracking analysis (n = 21). Before analysis, eye movement
data were screened to remove noisy and/or spurious re-
cordings. To remove spurious significant differences in
the time course between conditions, a cluster correction
was applied. Clusters were defined by the sum of their
constituent (absolute) t values and compared with a null
hypothesis distribution of clusters produced by shuffling
the condition labels (1000 permutations). Clusters below
the 95th percentile of the null hypothesis distribution
were disregarded.
Significance Testing
The significance of differences between data from differ-
ent conditions was assessed using the repeated-measures
ANOVA and the paired t test, and the significance of dif-
ferences between data from different experiments was
assessed using the independent t test. The normality as-
sumption was tested with the Shapiro–Wilk test of nor-
mality. Excluding the time-series analyses, there were
16 t tests performed. For six of these, the raw data vio-
lated the assumption of normality. Allo stesso modo, the raw data
used in the repeated-measures ANOVA violated the as-
sumption of normality. Così, before assessing the differ-
ences between groups for these tests, we normalized the
data using a log transform. We report the results of the
normalized data, although for all cases the pattern of re-
sults was the same for the raw data.
RESULTS
SSVEP Analysis of Test Stimuli
If mismatches between the two eyes evoke inhibitory ac-
attività, we would expect that selectively reducing the re-
sponsiveness of the neural mechanisms that respond to
mismatches (through adaptation) would lead to less in-
hibitory activity and thus relatively more excitability. A
test this idea, observers were initially adapted to binocu-
lar mismatches by viewing aRDS for a prolonged period
(64 sec). Then, following adaptation, participants viewed
a cRDS composed of black and white blobs depicting an
annulus that was either near or far relative to the back-
ground. We used the ERP evoked by changing the depth
sign of the annulus (from near to far or far to near) as an
index of stereoscopic related activity and the ERP evoked
by refreshing the positions of the dots as an index of non-
stereoscopic-related activity. Similar to previous work
(Cottereau, McKee, Ales, et al., 2012; Cottereau, McKee, &
Norcia, 2012; Cottereau, McKee, Ales, & Norcia, 2011), we
rapidly changed the stimuli at two frequencies, questo è,
4 Hz for depth sign and 20 Hz for dot refresh, producing
two distinct SSVEPs. We measured the activity evoked by
the stimulus changes by performing a Fourier transform
on the data, converting it from the time domain to the
frequency domain and taking the SNR between the peak
at the target frequencies (4 E 20 Hz) and the base-
line noise in the spectrum (from bins either side of the
target frequency). For comparison, we also measured
observers’ SSVEP in response to the test stimulus follow-
ing adaptation to cRDS and without prior adaptation
(baseline).
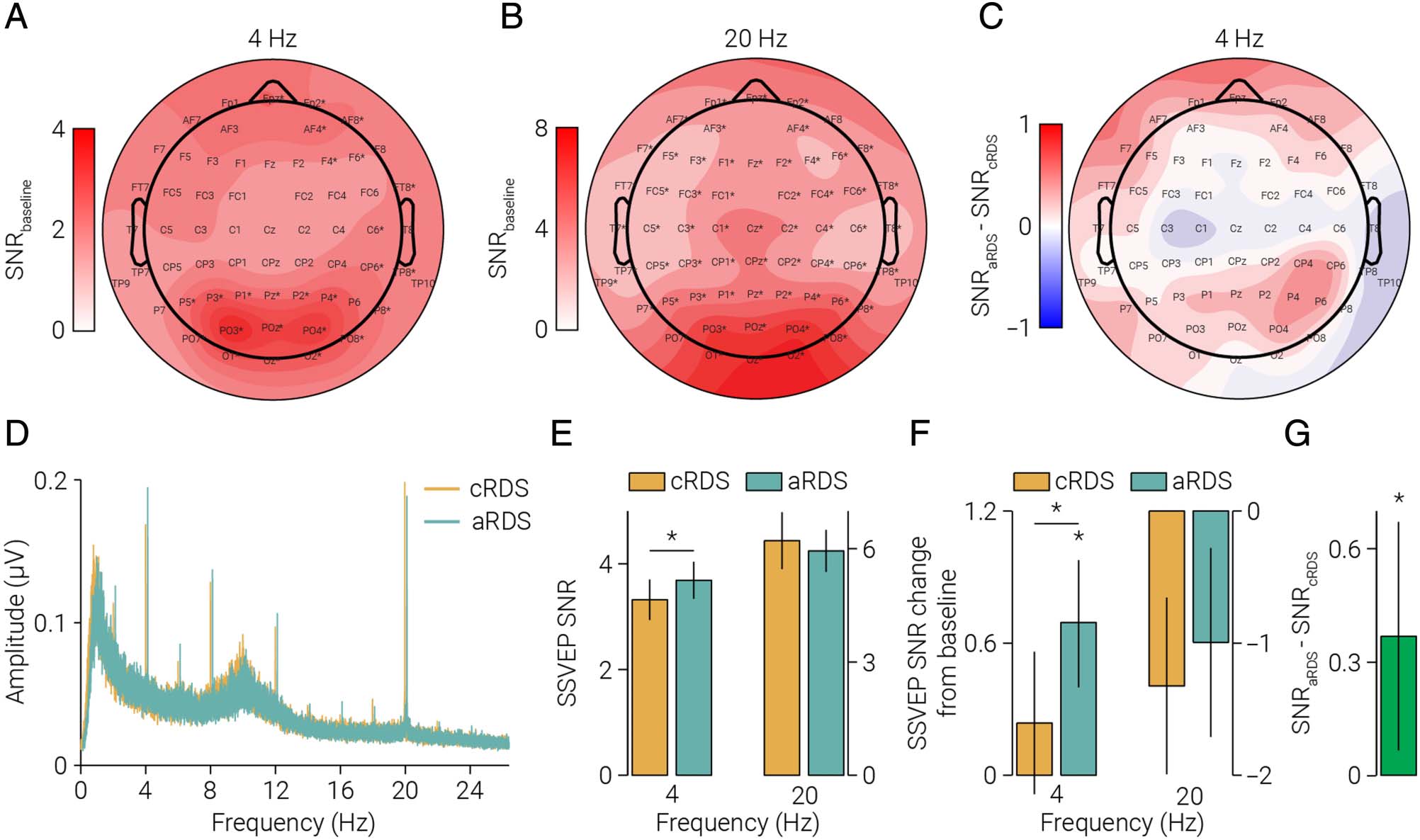
To guide electrode selection, we computed the 4-Hz
SNR of each sensor during the baseline test run. As antic-
ipated, we found the highest SNR for occipital and pari-
etal sensors (Figure 2A); we found a similar pattern of
activity at 20 Hz (Figure 2B). Visual inspection of the
topographies suggests the peak activity of the 20-Hz
response was more occipital than the 4-Hz response.
Based on these results and previous electrophysiological
work on binocular disparity (Cottereau, McKee, & Norcia,
2012; Cottereau et al., 2011), we included all parietal and
occipital (cioè., Oz, O1, O2, POz, PO3, PO4, PO7, PO8, Pz,
P1, P2, P3, P4, P5, P6, P7, and P8) sensors in the main
SSVEP analysis.
Overall, we observed differences in activity in response
to the test following adaptation to cRDS and aRDS
(Figure 2C, D). To test the prediction that prolonged
viewing of binocular mismatches increases neural excit-
ability, we computed the average SSVEP SNR across pari-
etal and occipital sensors and compared responses at the
depth change frequency (4 Hz). In line with the predic-
zione, we found that SSVEP SNR was significantly higher
following adaptation to aRDS compared with cRDS
(paired t test, all participants: T(30) = 2.49, p=.019; par-
ticipants with baseline: T(21) = 2.77, p = .011; Figure 2E;
19 Di 31 participants showed the effect). A possible
Rideaux, Michael, and Welchman
103
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
.
T
/
o
N
0
5
M
UN
sì
2
0
2
1
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
.
F
/
T
o
N
0
5
M
UN
sì
2
0
2
1
Figura 2. SSVEP in response to the test stimulus. (A–B) Topographic map showing the (UN) 4 Hz and (B) 20 Hz SSVEP SNR in response to the test
stimulus without prior adaptation. (C) Same as A, but for the difference in response to the test stimulus following adaptation to either correlated or
aRDSs across. (D) The SSVEP SNR response spectra (averaged across parietal and occipital sensors) to the test stimulus following adaptation to
cRDS/aRDS, for all participants (n = 31). The aRDS spectrum is horizontally offset to facilitate comparison with the cRDS spectra. (E) Same as C, Ma
isolating the depth alternation (4 Hz) and dot refresh (20 Hz) frequencies. A repeated-measures ANOVA of SSVEP SNR revealed a main effect of
frequency (4/20 Hz; F(1, 30) = 14.59, p = 6.2e−4) and an interaction between frequency and adaptation (cRDS/aRDS; F(1, 30) = 7.19, p = .012),
but no main effect of adaptation (F(1, 30) = 2.76, p = .107). For participants who completed the baseline measurement (n = 15), F shows
the same as D, but referenced to baseline SSVEP amplitude. We found numerical differences in the same direction between conditions at
8 E 12 Hz harmonics, but these did not reach thresholds for statistical significance (paired t test, 8 Hz: T(30) = 1.50, p = .144; 12 Hz: T(30) =
1.40, p = .171). Consistent with previous work showing that looming stimuli are more salient than receding stimuli (Franconeri & Simons,
2003), we found peaks (significantly above the level of noise) at the asymmetric harmonics (2 E 6 Hz) indicating a difference in response to
the near and far stimuli (t test, 2 Hz: cRDS, T(30) = 7.09, p = 3.5e−8, aRDS, T(30) = 7.49, p = 1.2e−8; 6 Hz: cRDS, T(30) = 9.91, p = 4.1e−11,
aRDS, T(30) = 6.34, p = 2.7e−7). Tuttavia, we found no difference in the amplitude of these peaks between adaptation conditions (paired t test,
2 Hz: T(30) = 0.87, p = .389; 6 Hz: T(30) = 0.58, p = .564). (G) The average paired difference in 4 Hz SSVEP SNR between aRDS/cRDS
conditions. Asterisks in A and B indicate sensors with SNR significantly than >1 following Bonferroni correction (N = 64, α = 7.8e−8); asterisks
in E–G indicate significant differences. Error bars in E–F indicate SEM. Error bars in G indicate 95% confidence intervals.
concern is that the difference in SSVEP SNR between ad-
aptation conditions was due to a decrease in excitability
following adaptation to cRDS, rather than an increase fol-
lowing adaptation to aRDS. Tuttavia, we found no evi-
dence for this: Whereas adaptation to aRDS significantly
increased SSVEP SNR relative to baseline (paired t test, T
(21) = 2.39, p = .026), adaptation to cRDS produced no
significant change (paired t test, T(21) = 0.74, p = .469;
Figure 2F); these results are consistent with the interpre-
tation that adaptation to binocular mismatches increased
neural excitability.
Another possible concern is that the increased excit-
ability we observed in the primary visual cortex following
adaptation to aRDS is not specific to stereopsis, but a
generalized effect. Tuttavia, we found no evidence for
a difference in SSVEP amplitude at the frequency of the
dot refresh (20 Hz) between aRDS and cRDS adaptation
conditions (paired t test, T(30) = 0.60, p = .552; Figure 2E),
suggesting that the effect relates to changes in depth and
not luminance. Another possible concern is that the effect
might be driven by a subset of participants with low SSVEP
amplitude across all conditions. Specifically, SSVEP ampli-
tude measurements are less reliable at low values; così, UN
subset of unreliable values could potentially yield a false
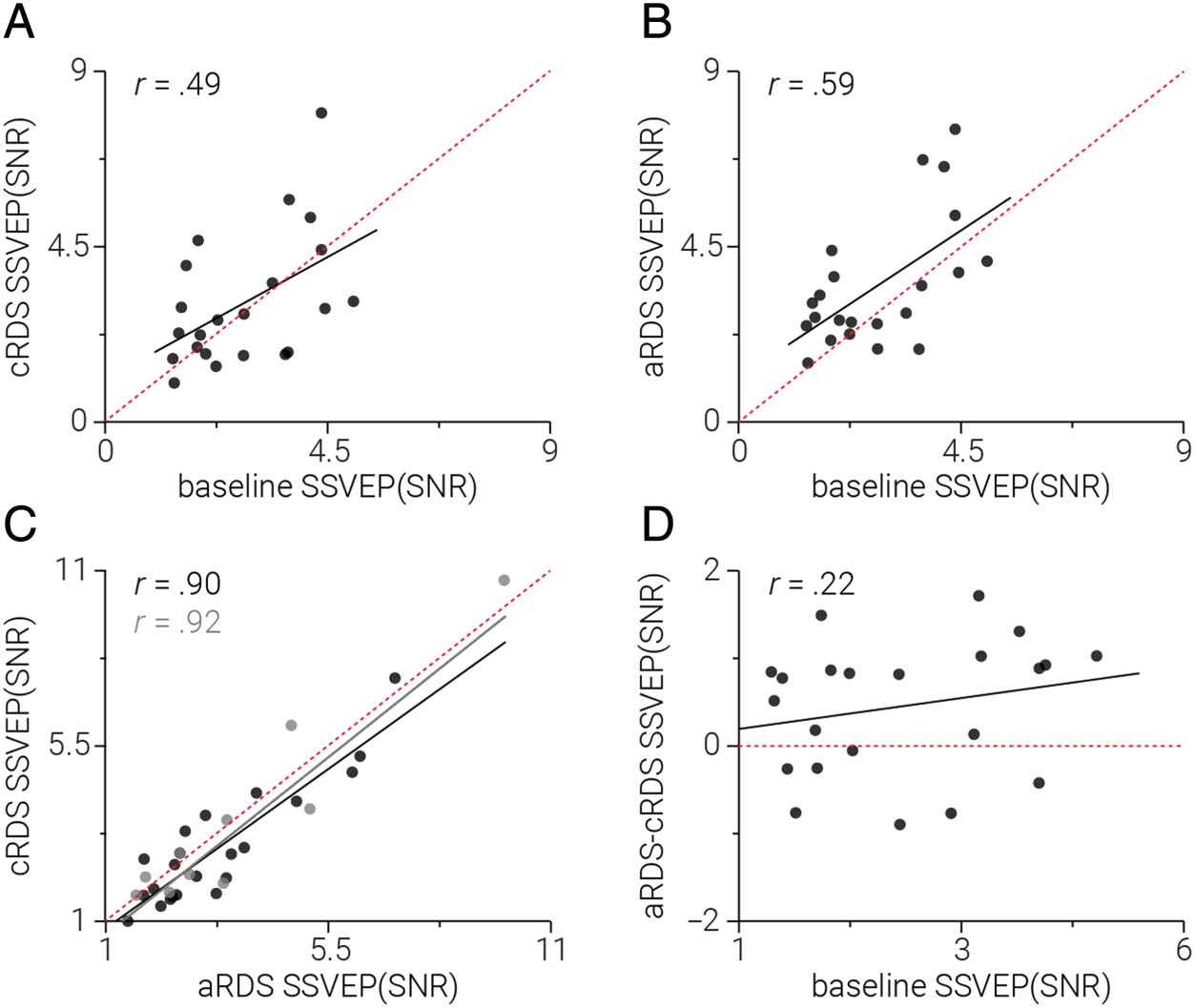
positive. Tuttavia, although we found significant correla-
tions between baseline, cRDS, and aRDS SSVEP amplitude
(Pearson correlation: participants with baseline: n = 22,
baseline–cRDS: r = .489, p = .021; baseline–aRDS: r =
.596, p = .003; cRDS–aRDS: r = .897, p = 1.5e−13; Tutto
participants, n = 31: cRDS–aRDS: r = .92, p = 1.5e−13;
Figure 3A–C), we found no evidence of a relationship be-
tween baseline amplitude and the difference between aRDS
104
Journal of Cognitive Neuroscience
Volume 32, Numero 1
Figura 3. Relationship between
SSVEP SNR across conditions.
(UN) SSVEP SNR measured in the
response to the test stimulus
following cRDS adaptation as a
function of SSVEP SNR without
adaptation. (B) Same as A, Ma
following aRDS adaptation. (C)
Same as B, but as a function of
SSVEP SNR following cRDS
adaptation. Black and gray dots
indicate participants who did
and did not complete the
baseline condition, rispettivamente.
Black and gray lines indicate the
least squares regression not
including (n = 22) E
including (n = 33) participants
who did not complete the
baseline, rispettivamente. The red
dotted lines in A–C indicate the
first diagonal ( y = x). (D) IL
difference in the effect of cRDS
and aRDS adaptation on SSVEP
SNR as a function of SSVEP SNR
without adaptation.
and cRDS (Pearson correlation, n = 22, r = .219, p = .327;
Figure 3D).
Finalmente, another possible explanation for the effect
might be that observers’ attentional allocation during the
adaptation period varied between conditions. Tuttavia,
we found no evidence for a difference in performance
on the attentionally demanding Vernier task between
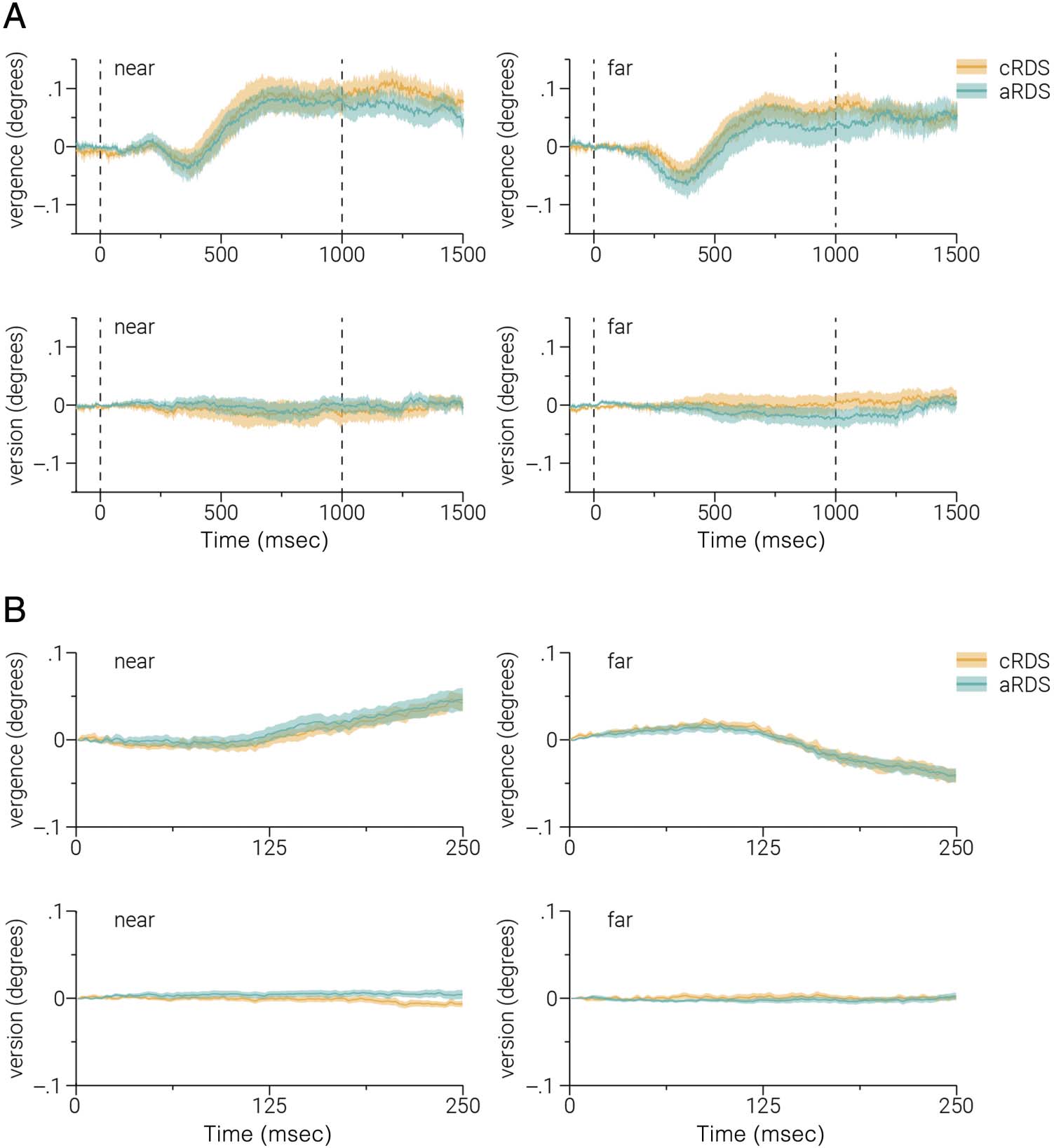
conditions (paired t test, T(30) = 1.22, p = .23). Sim-
ilarly, we found no evidence for a difference in eye move-
ments in response to either the adaptation or test stimuli
(Figure 4A, B).
Although the strongest response to the 4-Hz depth
alternation of the test stimulus in the baseline condition
was in posterior sensors (Figure 2A), there was a region
of elevated SNR in anterior frontal electrodes. This activ-
ity was also reflected in the topography of condition dif-
ferences (Figure 2C). Eye-tracking results show this is
unlikely to be due to differences in eye movements.
Piuttosto, consistent with previous work (Cottereau et al.,
2011), this activity likely reflects the distributed involve-
ment of multiple neural structures in the processing of
binocular disparity.
ERP Analysis of Adaptation Stimuli
The primary SSVEP analysis revealed a difference in stereo-
scopic event-related neural responsiveness following
adaptation to aRDS but not cRDS, relative to baseline,
consistent with the prediction that adaptation to binocular
mismatches increases neural excitability. If the cRDS/aRDS
adaptation stimuli have different functional consequences,
this indicates that these stimuli evoked different patterns
of activity during adaptation. Although the central aim of
the experiment was to test the consequences of adapta-
tion to cRDS/aRDS, the ERPs evoked by these stimuli dur-
ing the adaptation period may inform the mechanism by
which different effects of adaptation were produced. Così,
in an exploratory analysis, we computed the difference be-
tween cRDS/aRDS ERPs (averaged across all presentations,
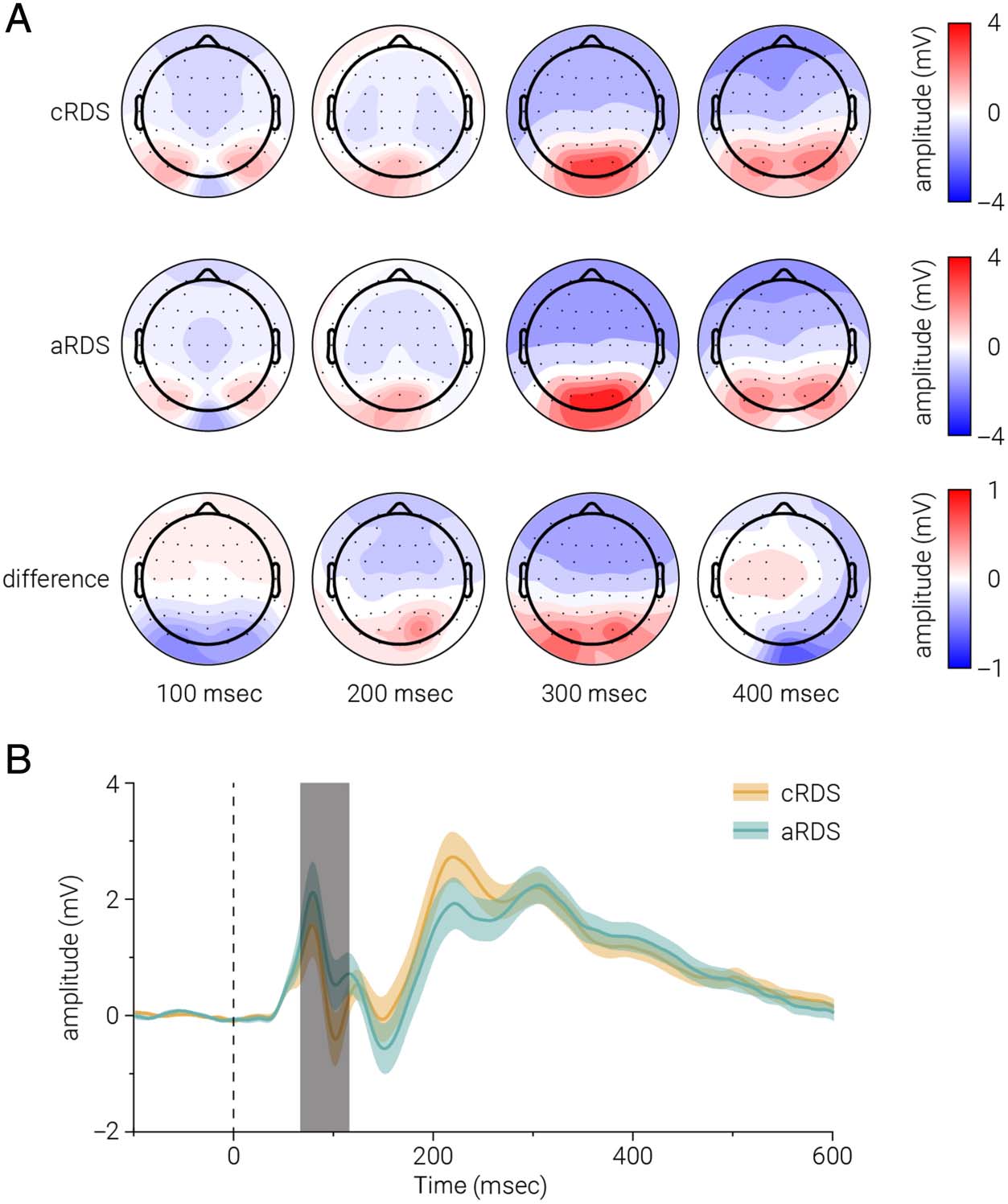
across all sensors). We found that the sensors that showed
the greatest difference were regionally similar to those
used in the SSVEP analysis (Figure 5A). Given the topo-
graphic similarity of differences in activity evoked by test
and adaptor stimuli between conditions, we then com-
pared the pooled activity of adaptor ERPs from the same
(occipital and parietal) sensors used in the previous anal-
ysis. We found ERP amplitude at time points consistent
with the P1 component were significantly smaller in re-
sponse to aRDS compared with cRDS, but the amplitude
of those consistent with the N1 component were signifi-
cantly larger (paired t test, peak t(30) = 3.54, p = .001;
Figure 5B).
Anticorrelated Test Stimulus
We used a cRDS test stimulus, instead of an aRDS, A
measure changes in neural excitability following adapta-
tion to anticorrelated images as cRDS evoke a larger re-
sponse than aRDS (Petrig et al., 1981; Braddick et al.,
1980), providing a better SNR and thus a more sensitive
measure of neural change. Tuttavia, a possible concern
is that the reduction in SSVEP amplitude we observed in
response to the change in the depth of a cRDS following
adaptation to anticorrelation was influenced by the
Rideaux, Michael, and Welchman
105
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
T
.
F
o
N
0
5
M
UN
sì
2
0
2
1
Figura 4. Comparison of eye
movements between (cRDS/
aRDS) adaptation conditions.
We assessed whether the
adaptation effects observed
could be explained by
differences in eye movements
during stimulus presentation by
comparing observers’ eye
position during the adaptation
and test periods between cRDS
and aRDS adaptation
conditions. (UN) Average
vergence and version eye
movements for near and far
stimuli presented during the
cRDS and aRDS adaptation
periods. (B) Same as A, but for
test periods. The dashed lines in
A indicate stimulus onset and
offset; color-shaded regions
indicate ±1 SEM.
difference between adaptation and test stimuli. Questo è,
the cRDS test stimulus evoked a reduced neural response
following adaptation to aRDS compared with cRDS
because the difference between adaptation and test stim-
ulus was more pronounced in the aRDS condition. A
test this possibility, we repeated the experiment, on a
new cohort of participants, using an aRDS test stimulus
(Figure 6A).
We included data from the same sensors used in the
SSVEP analysis of the main experiment (cioè., Oz, O1,
O2, POz, PO3, PO4, PO7, PO8, Pz, P1, P2, P3, P4, P5,
P6, P7, and P8). The SSVEP SNR response spectra for
adaptation conditions are shown in Figure 5B. Although
there is a clear peak at the dot refresh frequency (20 Hz),
there is little discernible peak at the depth alternation fre-
quency (4 Hz); contrast this with the spectra from the
main experiment where a peaks can be clearly resolved
at 4 Hz and its harmonics (Figure 2D). Infatti, a compar-
ison of the SSVEP SNR at 4 Hz between the baseline con-
dition here and in the main experiment show that the
amplitude was significantly lower here, in response to
an aRDS test stimulus (independent t test, T(43) = 6.51,
p = 7.3e−8). We found no significant difference in the
4-Hz SSVEP SNR between aRDS and cRDS adaptation con-
ditions (paired t test, T(21) = 1.46, p = .159). Tuttavia, if
there were differential effects of adaptation to cRDS and
aRDS on the potential evoked by the aRDS test stimulus,
given the low amplitude of the signal, it is unlikely that
this difference could be detected.
Although we did not find a reliable signal in response
to the depth alternation of the anticorrelated test stimu-
lus, we were able to reliably measure the ERP in response
to the anticorrelated stimulus during the adaptation
phase. This is likely because in the adaptation phase,
the depth alternation was concurrent with the stimulus
presentation; così, the ERP reflected the response to
both the change in depth and the stimulus onset. By con-
trast, in the test phase, the stimulus was presented con-
stantly whereas depth alternated at 4 Hz; così, the signal
at 4 Hz reflects the response to depth alternation in
106
Journal of Cognitive Neuroscience
Volume 32, Numero 1
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
/
.
T
o
N
0
5
M
UN
sì
2
0
2
1
Figura 5. ERPs in response to
the adaptation stimuli. (UN)
Topographic maps showing
average neural activity in
response to the (cRDS/aRDS)
adaptation stimuli and the
difference between them at 100,
200, 300, E 400 msec after
stimulus onset. (B) Neural
activity in response to the
(cRDS/aRDS) adaptation
stimuli, averaged over the
occipital and parietal sensors, COME
a function of time. The dashed
line indicates stimulus onset;
color-shaded regions indicate
±1 SEM, and gray-shaded bar
indicates significant differences
between conditions.
isolation. These results suggest that the ERP measured in
the adaptation phase (at least for the anticorrelation con-
dizione) was primarily a response to stimulus onset.
DISCUSSION
Electrophysiological recordings from macaque visual cor-
tex ( Janssen et al., 2003; Tsao et al., 2003; Cumming &
Parker, 1997) and psychophysical work with humans
(Katyal et al., 2018; Kingdom et al., 2018) has revealed
the existence of cortical mechanisms tuned to mismatched
features between the left and right eyes. Although the ev-
idence supporting the existence of neurons tuned to mis-
matches is extensive, our understanding of their role in
binocular vision remains limited. Here, we provide evi-
dence that neural activity in the visual cortex may facilitate
Figura 6. Stimuli and results from the control experiment. (UN) Example of the aRDS test stimuli used in the control experiment. (B)The SSVEP
SNR response spectra (averaged across parietal and occipital sensors) to the aRDS test stimulus following adaptation to cRDS/aRDS, for all
participants (n = 22). The aRDS spectrum is horizontally offset to facilitate comparison with the cRDS spectra.
Rideaux, Michael, and Welchman
107
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
.
T
F
/
o
N
0
5
M
UN
sì
2
0
2
1
binocular vision through inhibition. In particular, we show
that adaptation to mismatched binocular stimuli, questo è,
aRDS, produces increased excitability in the visual cortex
in response to changes in depth.
Prolonged/repeated exposure typically produces a re-
duction in the responsiveness of stimulated neurons, Quello
È, adaptation. Così, one might expect adaptation to
aRDS to reduce the net responsiveness of the visual cor-
tex in response to a change in depth. Alternatively, one
might anticipate that adaptation to aRDS to have no ef-
fect on the response to a cRDS, due to the perceptual
dissimilarity of these stimuli, questo è, aRDS do not pro-
duce a percept of depth. Tuttavia, in contrast to these
intuitive hypotheses, we found that adaptation to aRDS
yields an increase in excitability. Although these results
may seem surprising, they are consistent with the notion
that neurons tuned to binocular mismatches can facilitate
stereopsis by suppressing unlikely perceptual interpreta-
zioni (Goncalves & Welchman, 2017).
The results of the SSVEP analysis of activity evoked by
cRDS stimuli in the test period suggested that prolonged
viewing of cRDS and aRDS resulted in adaptation of
different neural ensembles or similar neural ensembles
adapted to different extents. Analysis of the ERPs evoked
by the adaptor stimuli confirmed this, revealing different
patterns of activity corresponding to cRDS and aRDS
stimuli. Specifically, we found differences between the
P1 and N1 components. P1 and N1 components are
thought to represent activity relating to early sensory pro-
cesses; così, the differences in amplitude of these com-
ponents in response to aRDS and cRDS may reflect the
differences in the engagement of excitatory vs. inhibitory
mechanisms. The amplitude of N1 has been shown to in-
crease with attentional allocation (Hillyard & Anllo-Vento,
2002; Polich, 1986; Van Voorhis & Hillyard, 1977; Haider,
Spong, & Lindsley, 1964). As the amplitude of the N1
component was higher for aRDS, this may signal that ob-
servers allocated more attention to the aRDS than the
cRDS, Per esempio, because of its perceptual peculiarity.
Tuttavia, the results from the concurrent (attentionally
demanding) task suggest otherwise: No differences in ac-
curacy or response time were observed between condi-
zioni. Inoltre, no difference in amplitude between
adaptation conditions was found for the P1 component,
which is also modulated by attention (Hillyard & Anllo-
Vento, 2002; Heinze et al., 1994).
A previous study found a reduction in SSVEP ampli-
tude in response to a drifting sinusoidal grating, following
adaptation to a grating drifting in the same direction
(Ales & Norcia, 2009). It seems reasonable to expect that
the response to the cRDS test stimulus would be reduced
following adaptation to cRDS. Given that we did not find
a reduction in excitability following adaptation to cRDS,
this may indicate that adaptation to motion has a more
pronounced neural signature than adaptation to binocu-
lar disparity. Tuttavia, there are other methodological
differences that may also account for this apparent dis-
crepancy. The stimuli used in our experiment covered
a visual region approximately 20% the size of that used
by Ales and Norcia (2009), thereby eliciting adaptation
of fewer neurons. Inoltre, we adapted participants
to and tested two disparities (±10 arcmin) all'interno del
same block, whereas Ales and Norcia (2009) separately
adapted to different directions within each block. In de-
signing the experiment, we predicted that we would find
the greatest overall effect on excitability by adapting both
crossed and uncrossed disparities. Tuttavia, our results
may indicate that the effects of adaptation are more read-
ily detected using the SSVEP method when one popula-
tion of neurons is adapted at a time. Another possible
explanation is that the correlation between left- and right-
eye images in the cRDS is more similar to that observed in
the natural environment, compared with the aRDS, Quale
has an artificially low correlation. Così, the tolerance for
cRDS may be higher than that for aRDS, and effects of ad-
aptation may be subtler. Tuttavia, we found a reduction in
the amplitude of the ERP over the course of the adaptation
period for both aRDS and cRDS, indicating that adaptation
had a measurable effect on neural activity for both types of
stimuli.
A possible concern is that the increase in SSVEP ampli-
tude following adaptation to aRDS, compared with cRDS,
was due to the aRDS stimulus being more perceptually
dissimilar to the test stimulus than the cRDS. In particu-
lar, the increased response may be due to less expec-
tation of the test stimulus in the aRDS condition. IL
results from the control experiment, in which we used
an aRDS test stimulus, provided inconclusive evidence
for this possibility, as the amplitude of the SSVEP SNR
was too low to produce reliable estimates. This is consis-
tent with previous work showing cRDS evoke a larger re-
sponse than aRDS (Petrig et al., 1981; Braddick et al.,
1980) and may suggest that the response to the cRDS
probe in the main experiment primarily originated from
areas where the correspondence problem had been
solved. Tuttavia, in the baseline condition of the main
experiment, the test stimulus was preceded by a period
in which a gray background was presented; this is argu-
ably more dissimilar from the test stimulus than either
cRDS/aRDS adaptor. Così, if SSVEP amplitude reflected
the similarity between the test stimuli and preceding
images, we would expect SSVEP amplitude to be highest
in the baseline condition where the dissimilarity was
highest. Tuttavia, we found no evidence for this: base-
line amplitude was significantly lower than that following
adaptation to aRDS.
Although EEG has relatively high temporal resolution,
the spatial resolution of the technique is limited. Così, UN
limitation of the current study is that we cannot make
precise statements about the likely neural locus of adap-
tation to binocular anticorrelation in the visual cortex.
fMRI is known to have much better spatial resolution
compared with EEG; Tuttavia, excitatory and inhibitory
activity cannot be differentiated from BOLD signal,
108
Journal of Cognitive Neuroscience
Volume 32, Numero 1
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
F
/
.
T
o
N
0
5
M
UN
sì
2
0
2
1
restricting the diagnostic utility of this technique in estab-
lishing the role of “what not” mechanisms.
It is interesting to consider the behavioral implications
of our results. Previous work has shown that adaptation
to binocular anticorrelation (also known as interocular
contrast differences) reduce observers’ sensitivity to bin-
ocular anticorrelation (Katyal et al., 2018; Kingdom et al.,
2018). We did not find a difference in performance on
the irrelevant task used to maintain observers’ eye move-
ments and attention; Tuttavia, it is possible that the in-
creased neural excitability resulting from adaptation to
binocular anticorrelation may have altered performance
on a task involving depth judgments. Future work could
test whether reducing the sensitivity of neural systems
tuned to binocular anticorrelation through adaptation in-
creases sensitivity to binocular correlation, Per esempio,
by reducing neural inhibition.
The current results have implications beyond stereop-
sis. There is theoretical and empirical evidence support-
ing the existence of neurons tuned to mismatches from
studies of stereopsis (Tsao et al., 2003; Prince, Cumming,
& Parker, 2002; DeAngelis, Ohzawa, & Freeman, 1991),
binocular rivalry (Katyal et al., 2018; Kingdom et al.,
2018; Said & Heeger, 2013), and integration of cues
within (Rideaux & Welchman, 2018; Kim, Angelaki, &
Deangelis, 2015; Nadler et al., 2013) and between senso-
ry modalities (Kim, Pitkow, Angelaki, & DeAngelis, 2016;
Gu, Angelaki, & DeAngelis, 2008; Morgan, DeAngelis, &
Angelaki, 2008). Here, we provide evidence suggesting a
role for mechanisms tuned to mismatches that may facil-
itate inference by driving suppression.
Ringraziamenti
We thank N. Goncalves for detailed discussions and comments
on the article. This work was supported by the Leverhulme
Trust (ECF-2017-573 to R. R.), the Isaac Newton Trust (17.08(o)
to R. R.), and the Wellcome Trust (095183/Z/10/Z to A. E. W.
and 206495/Z/17/Z to E. M.).
Reprint requests should be sent to Reuben Rideaux,
Department of Psychology, University of Cambridge, CB2
3EB, United Kingdom, or via e-mail: reuben.rideaux@gmail.
com.
REFERENCES
Ales, J. M., & Norcia, UN. M. (2009). Assessing direction-
specific adaptation using the steady-state visual evoked
potential: Results from EEG source imaging. Journal of
Vision, 9, 8.
Braddick, O., Atkinson, J., Julesz, B., Kropfl, W., Bodis-
Wollner, I., & Raab, E. (1980). Cortical binocularity in
infants. Nature, 288, 363–365.
Brainard, D. H. (1997). The psychophysics toolbox. Spatial
Vision, 10, 433–436.
Cornelissen, F. W., Peters, E. M., & Palmer, J. (2002). IL
Eyelink toolbox: Eye tracking with MATLAB and the
Psychophysics toolbox. Behavior Research Methods,
Instruments & Computers, 34, 613–617.
Cottereau, B. R., McKee, S. P., Ales, J. M., & Norcia, UN. M. (2011).
Disparity-tuned population responses from human visual
cortex. Journal of Neuroscience, 31, 954–965.
Cottereau, B. R., McKee, S. P., Ales, J. M., & Norcia, UN. M.
(2012). Disparity-Specific Spatial Interactions: Evidence
from EEG Source Imaging. Journal of Neuroscience, 32,
826–840.
Cottereau, B. R., McKee, S. P., & Norcia, UN. M. (2012). Bridging
the gap: Global disparity processing in the human visual
cortex. Journal of Neurophysiology, 107, 2421–2429.
Cumming, B. G., & Parker, UN. J. (1997). Responses of primary
visual cortical neurons to binocular disparity without depth
perception. Nature, 389, 280–283.
DeAngelis, G. C., Ohzawa, I., & Freeman, R. D. (1991). Depth
is encoded in the visual cortex by a specialized receptive
field structure. Nature, 352, 156–159.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Fleet, D. J., Wagner, H., & Heeger, D. J. (1996). Neural
encoding of binocular disparity: Energy models,
positionshifts and phase shifts. Vision Research, 36,
1839–1857.
Franconeri, S. L., & Simons, D. J. (2003). Moving and looming
stimuli capture attention. Perception & Psychophysics, 65,
999–1010.
Goncalves, N. R., & Welchman, UN. E. (2017). “What not”
detectors help the brain see in depth. Current Biology, 27,
1403–1412.
Gu, Y., Angelaki, D. E., & DeAngelis, G. C. (2008). Neural
correlates of multisensory cue integration in macaque MSTd.
Nature Neuroscience, 11, 1201–1210.
Haider, M., Spong, P., & Lindsley, D. B. (1964). Attention,
vigilance, and cortical evoked-potentials in humans. Scienza,
145, 180–182.
Heinze, H. J., Mangun, G. R., Burchert, W., Hinrichs, H.,
Scholz, M., Münte, T. F., et al. (1994). Combined spatial and
temporal imaging of brain activity during visual selective
attention in humans. Nature, 372, 543–546.
Hillyard, S. A., & Anllo-Vento, l. (2002). Event-related brain
potentials in the study of visual selective attention.
Proceedings of the National Academy of Sciences, U.S.A.,
95, 781–787.
Janssen, P., Vogels, R., Liu, Y., & Orban, G. UN. (2003). At least at
the level of inferior temporal cortex, the stereo
correspondence problem is solved. Neuron, 37, 693–701.
Julesz, B., & Chang, J. J. (1976). Interaction between pools of
binocular disparity detectors tuned to different disparities.
Biological Cybernetics, 22, 107–119.
Katyal, S., Vergeer, M., Lui, S., Lui, B., & Engel, S. UN. (2018).
Conflict-sensitive neurons gate interocular suppression in
human visual cortex. Scientific Reports, 8, 1239.
Kim, H. R., Angelaki, D. E., & Deangelis, G. C. (2015). UN
novel role for visual perspective cues in the neural
computation of depth. Nature Neuroscience, 18, 129–137.
Kim, H. R., Pitkow, X., Angelaki, D. E., & DeAngelis, G. C.
(2016). A simple approach to ignoring irrelevant variables by
population decoding based on multisensory neurons.
Journal of Neurophysiology, 116, 1449–1467.
Kingdom, F. UN. A., Jennings, B. J., & Georgeson, M. UN.
(2018). Adaptation to interocular difference. Journal of
Vision, 18, 9.
Marr, D., & Poggio, T. (1976). Cooperative computation of
stereo disparity. Scienza, 194, 283–287.
Morgan, M. L., DeAngelis, G. C., & Angelaki, D. E. (2008).
Multisensory integration in macaque visual cortex depends
on cue reliability. Neuron, 59, 662–673.
Rideaux, Michael, and Welchman
109
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
.
T
F
o
N
0
5
M
UN
sì
2
0
2
1
Nadler, J. W., Barbash, D., Kim, H. R. R., Shimpi, S., Angelaki,
D. E., & DeAngelis, G. C. (2013). Joint representation of depth
from motion parallax and binocular disparity cues in macaque
area MT. Journal of Neuroscience, 33, 14061–14074.
Ohzawa, I., DeAngelis, G., & Freeman, R. (1990). Stereoscopic
depth discrimination in the visual cortex: Neurons ideally
suited as disparity detectors. Scienza, 249, 1037–1041.
Pelli, D. G. (1997). The Videotoolbox software for visual
psychophysics: Transforming numbers into movies. Spatial
Vision, 10, 437–442.
Petrig, B., Julesz, B., Kropfl, W., Baumgartner, G., & Anliker, M.
(1981). Development of stereopsis and cortical binocularity
in human infants: Electrophysiological evidence. Scienza,
213, 1402–1405.
binocular disparities in the human brain. Journal of
Neuroscience, 28, 11315–11327.
Prince, S. J., Cumming, B. G., & Parker, UN. J. (2002). Range and
mechanism of encoding of horizontal disparity in macaque
V1. Journal of Neurophysiology, 87, 209–221.
Read, J. C., & Cumming, B. G. (2007). Sensors for impossible
stimuli may solve the stereo correspondence problem.
Nature Neuroscience, 10, 1322–1328.
Rideaux, R., & Welchman, UN. E. (2018). Proscription supports
robust perceptual integration by suppression in human visual
cortex. Nature Communications, 9, 1502.
Said, C. P., & Heeger, D. J. (2013). A model of binocular rivalry
and cross-orientation suppression. PLOS Computational
Biology, 9, e1002991.
Polich, J. (1986). Attention, probability, and task demands as
Tsao, D. Y., Conway, B. R., & Livingstone, M. S. (2003).
determinants of P300 latency from auditory stimuli.
Electroencephalography and Clinical Neurophysiology, 63,
251–259.
Preston, T. J., Li, S., Kourtzi, Z., & Welchman, UN. E. (2008).
Multivoxel pattern selectivity for perceptually relevant
Receptive fields of disparity-tuned simple cells in macaque
V1. Neuron, 38, 103–114.
Van Voorhis, S., & Hillyard, S. UN. (1977). Visual evoked
potentials and selective attention to points in space.
Perception & Psychophysics, 22, 54–62.
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
o
H
w
T
N
T
P
o
:
UN
/
D
/
e
D
M
io
F
R
T
o
P
M
R
C
H
.
P
S
io
l
D
v
io
R
e
e
R
C
T
C
.
M
H
UN
io
e
R
D
.
tu
C
o
o
M
C
N
/
j
UN
o
R
T
C
io
C
N
e
/
–
UN
P
R
D
T
io
3
2
C
l
1
e
1
–
0
P
0
D
F
1
9
/
3
3
1
2
1
/
5
1
4
/
1
o
0
C
0
N
/
_
UN
1
_
8
0
6
1
1
4
4
7
1
1
9
P
/
D
j
o
B
C
sì
N
G
_
tu
UN
e
_
S
0
T
1
o
4
N
7
0
1
8
.
P
S
D
e
F
P
e
B
M
sì
B
e
G
R
tu
2
e
0
S
2
T
3
/
j
/
F
.
T
o
N
0
5
M
UN
sì
2
0
2
1
110
Journal of Cognitive Neuroscience
Volume 32, Numero 1