Active Ignoring in Early Visual Cortex
Helen E. Payne and Harriet A. Allen
Astratto
■ Selective attention is critical for controlling the input to
mental processes. Attentional mechanisms act not only to select
relevant stimuli but also to exclude irrelevant stimuli. There is
evidence that we can actively ignore irrelevant information. Noi
measured neural activity relating to successfully ignoring dis-
tracters (using preview search) and found increases in both
the precuneus and primary visual cortex during preparation
to ignore distracters. We also found reductions in activity in
fronto-parietal regions while previewing distracters and a re-
duction in activity in early visual cortex during search when a
subset of items was successfully excluded from search, both
associated with precuneus activity. These results are consis-
tent with the proposal that actively excluding distractions has
two components: an initial stage where distracters are encoded,
and a subsequent stage where further processing of these items
is inhibited. Our findings suggest that it is the precuneus that
controls this process and can modulate activity in visual cortex
as early as V1. ■
INTRODUCTION
Efficient mental processing requires that we select from
the world those stimuli that are behaviorally relevant to
our current goals and to ignore those objects that are ir-
relevant. Attentional selection can enhance the neural
processing of attended stimuli, manifested as improve-
ments in reaction time, accuracy, and target detection/
discriminability of cued items (Carrasco, Penpeci-Talgar,
& Eckstein, 2000; Yeshurun & Carrasco, 1999; Posner,
Snyder, & Davidson, 1980). It can also suppress process-
ing of signals evoked by irrelevant stimuli and locations
(Sylvester, Jack, Corbetta, & Shulman, 2008; Ruff & Driver,
2006; Serences, Yantis, Culberson, & Awh, 2004). Fur-
thermore, fMRI studies reveal not only a stimulus evoked
response to an attended target but also increases in ac-
tivation in the portions of visual cortex that represent
the anticipated location of the stimulus, questo è, prepara-
tory activity (per esempio., Macaluso, Eimer, Frith, & Driver, 2003;
Hopfinger, Buonocore, & Mangun, 2000; Ress, Backus, &
Heeger, 2000; Kastner, Pinsk, De Weerd, Desimone, &
Ungerleider, 1999). This preparatory activity is thought to
bias the visual areas to favor the processing of the subse-
quent expected target. In the present study, we are inter-
ested in the complementary effect, that of preparation to
ignore.
Excluding unhelpful or irrelevant stimuli is clearly advan-
tageous. Several behavioral experiments have shown that
if the locations of distracters are known, then their detri-
mental effects on target processing are reduced (Ruff &
Driver, 2006; Serences et al., 2004), and this is linked to
increased neural activity in visual cortex. Control of atten-
University of Birmingham, UK
tion both to targets and away from distracters is likely to
involve a network of fronto-parietal brain regions. Brain re-
gions consistently activated during attentional preparation
following a cue include the intraparietal sulcus, frontal eye
fields, and the superior parietal lobule (SPL) (Sylvester et al.,
2008; Ruff & Driver, 2006; Macaluso et al., 2003; Corbetta,
Kincade, Ollinger, McAvoy, & Shulman, 2000; Hopfinger
et al., 2000; Kastner et al., 1999; Shulman et al., 1999). It
is thought that this network of brain regions is important
for generating biasing signals that modulate activity in visual
cortex (for a review, see Pessoa, Kastner, & Ungerleider,
2003). Given that parts of this network are likely to guide
attention toward targets and away from known distracters,
it is critical to separate these two processes. The preview
search paradigm (Watson & Humphreys, 1997) used here
allows the to-be ignored items to be separated in time
from the attended items. The preview distracters are pre-
sented prior to the addition of the remaining distracters
and a target to the display. Search performance improves
compared to trials where all items are shown at the same
time (a full-set search; Watson & Humphreys, 1997).
Watson and Humphreys (1997) argued that observers
actively apply top–down inhibition to the locations of
previewed distracters, which they termed visual mark-
ing (see also Braithwaite & Humphreys, 2003; Olivers &
Humphreys, 2003; Watson, 2001). Other accounts have
proposed no inhibition of old items and have placed em-
phasis on attention to the new items. Transient luminance
onsets of the new search items may automatically capture
Attenzione (Donk & Theeuwes, 2001) or temporal segmen-
tation between the old and new items may guide attention
( Jiang, Chun, & Marks, 2002). Behavioral evidence from dual-
task studies, on the other hand, (Humphreys, Watson, &
Jolicœur, 2002) and probe-dot detection studies (Humphreys,
© 2011 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 23:8, pag. 2046–2058
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
3
l
8
e
–
2
P
0
D
4
F
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
C
6
N
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
P
C
D
N
.
B
sì
2
0
G
1
tu
0
e
.
S
T
2
o
1
N
5
6
0
2
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
/
F
.
.
T
o
N
1
8
M
UN
sì
2
0
2
1
Stalmann, & Olivers, 2004; Olivers & Humphreys, 2002;
Watson & Humphreys, 2000) suggests old items in the
preview search task are actively inhibited. Watson and
Humphreys (2000) conducted a standard preview search
task but on a minority of trials participants were cued to
detect a probe dot. Detection of probes presented at the
locations of previewed distracters was impaired (vs. SU
new distracters), suggesting that information from the
locations of the previewed distracters was inhibited. Simi-
larly, comparing cueing to preview search showed that
participants were better able to exclude previewed items
than noncued items (Allen & Humphreys, 2007UN). Effec-
tively, the visual system reduces the contrast of success-
fully previewed and ignored items (Allen & Humphreys,
2007B).
Preview search, Perciò, offers a way to investigate
neural mechanisms underlying ignoring known distracters.
Recent neuroimaging studies (Dent, Allen, & Humphreys,
in press; Allen, Humphreys, & Matthews, 2008; Olivers,
Smith, Matthews, & Humphreys, 2005; Pollmann et al.,
2003) compared neural activity in response to preview
trials to that in response to nonpreview search baseline
trials. Trials consisted of two displays. The preview con-
dition first display was a true preview; items remained on
the screen when the new items appeared. In the baseline
conditions, the items in the first display disappeared and
were replaced by distracters in different locations. Così,
the first displays of preview trials and baseline trials were
visually identical but the attentional set of the participants
was different between the conditions; the first display in
baseline trials required only passive viewing, whereas the
preview trials would reveal the processes involved with
ignoring stimuli. These studies have consistently demon-
strated enhanced neural activation in posterior parietal cor-
tex (in particular, the SPL and the precuneus) for preview
trials relative to baseline. The SPL/precuneus is proposed
to set up a spatial representation of the old previewed
distracters so that these items are biased in favor of the
subsequently presented new search items. This is sup-
ported by computational modeling (Mavritsaki, Allen, &
Humphreys, 2009). Mavritsaki et al. (2009) used the spik-
ing Search over Time and Space (sSoTS) model to analyze
the preview search fMRI data from Allen et al. (2008) by
including inhibition (suppressing old distracters) and exci-
tazione (anticipation for target) as regressors and found
that the activation in the precuneus reported by Allen
et al. could be predicted by the inhibition.
Here we investigate what effect these parietal activa-
tions have on visually responsive cortex by presenting
stimuli to different retinal locations (cioè., the four visual
field quadrants). It is an open question as to what effect
previewing distracters has on sensory brain areas. Tradi-
tional models of visual attention would predict that un-
attended stimuli (cioè., uncued) would lead to a decrease
in neural activation in brain areas specialized for vision
(Gazzaley, Cooney, McEvoy, Knight, & DʼEsposito, 2005;
Smith, Singh, & Greenlee, 2000). In contrasto, when pre-
viewing face distracters, Allen et al. (2008) found an in-
crease in activation in face processing areas, even during
the initial preview display. This enhanced neural activation
in response to the preview trials may reflect an active
ignoring process that is distinct from passive viewing (O
simply attending elsewhere) of the same stimuli.
If a similar pattern of activation is found in early visual
cortex as in category-specific regions, then this might be
viewed as a signature for active ignoring. Given that at-
tending to a stimulus at a particular spatial location will en-
hance striate cortex blood oxygenation level dependent
(BOLD) activation (per esempio., Gandhi, Heeger, & Boynton, 1999;
Martínez et al., 1999; Somers, Dale, Seiffert, & Tootell,
1999), it is important to relate any activation change in
early visual areas with successful ignoring rather than sim-
ply the intent to ignore. Secondo, we investigate the func-
tional relationship between activity in precuneus regions
and changes in activity in visual cortex. Previously, it has
been assumed that if parietal and visual areas both change
in activation in preview trials, then one must drive the
other. Here, we take this one step further and look for
brain regions where activation appears to be part of a
functional network and link this to successfully ignoring
previewed items.
By using the preview search paradigm, we are able to
separate changes in activation related to successful ignor-
ing from changes related to the target. We follow the logic
of Dent et al. (in press), Allen et al. (2008), and Pollmann
et al. (2003), and include catch trials to measure the pure
neural activity associated with previewing informative (cioè.,
in the preview condition) and uninformative (cioè., in the
baseline condition) first displays without any contamina-
tion from the neural activity arising from the search dis-
plays. This is comparable to experiments (Macaluso et al.,
2003; Hopfinger et al., 2000; Kastner et al., 1999) that sep-
arate the neural activity associated with the cue (cioè., IL
preparatory activity) with that associated with the target.
METHODS
Participants
Eighteen paid participants (14 women, 18–35 years old,
M = 24.3 years) gave written informed consent in accor-
dance with the ethical procedures of the Birmingham Uni-
versity Imaging Centre, Birmingham, UK. All had normal
or corrected-to-normal vision.
Stimuli and Apparatus
Experiments were created in Matlab (The Mathworks, Natick,
MA) using the Psychophysics Toolbox (Brainard, 1997;
Pelli, 1997). The distracter stimuli were white uppercase
Lʼs, presented at four different orientations on a black
background (0°, 90°, 180°, and 270°; Guarda la figura 1). The tar-
get stimulus was a white uppercase T, presented either
Payne and Allen
2047
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
3
l
8
e
–
2
P
0
D
4
F
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
C
6
N
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
P
C
D
N
.
B
sì
2
0
G
1
tu
0
e
.
S
T
2
o
1
N
5
6
0
2
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
F
.
.
T
.
/
o
N
1
8
M
UN
sì
2
0
2
1
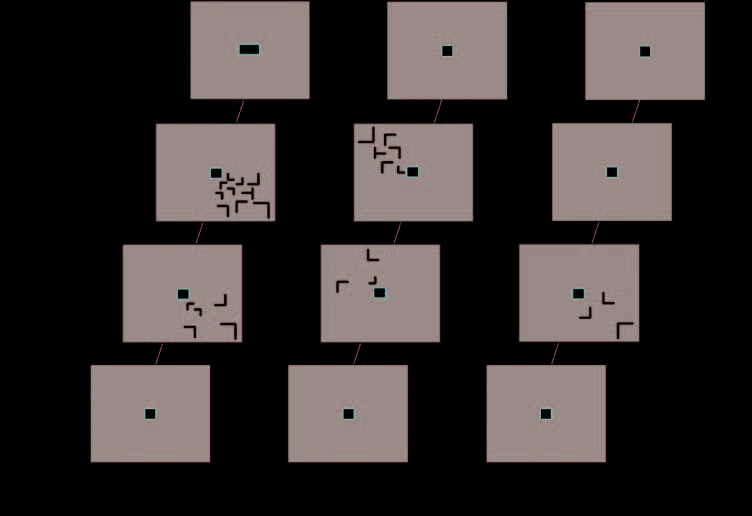
Figura 1. Experimental
procedure and stimuli.
Participants were instructed
to fixate on the central square
throughout the entire scan.
(UN) A preview search trial
(10 items). The distracters in
the first display remained on
the screen when the remainder
of the distracters and the
target (a T tilted ±90° from
vertical) appeared in the search
display. (B) A full search trial
(6 items). The first display was
uninformative; the distracters
offset at the start of the search
display and were replaced with
new distracters and a target
in the same quadrant. IL
target always appeared in the
search display for the preview
search and full search trials.
(C) A preview-only/dummy
trial. No search display was
presented for these trials.
During the end fixation for the
preview search and full search
trials, the fixation square
provided feedback; it changed
to a rectangle if the response
was incorrect (UN).
90° right or 90° left of vertical (randomly on each trial). IL
two line components making up each letter were identical
in length. The fixation consisted of a centrally located red
square (0.27° × 0.27°, at a distance of 65 cm).
Possible stimulus locations were arranged on a circular
140-cell virtual matrix that consisted of eight concentric cir-
cular grids with radii of 1°, 2.1°, 3.2°, 4.9°, 7°, 9.1°, 11.2°,
and 14°. Stimuli were scaled according to the human cor-
tical magnification factor (Dougherty et al., 2003; Horton
& Hoyt, 1991), resulting in 4, 8, 16, 12, 20, 24, 32, E 24
cells per circular grid. The stimuli presented on the three
innermost circular grids subtended 0.63° × 0.63°, il prossimo
four grids presented stimuli subtending 1.37° × 1.37°, E
the outermost ring presented stimuli subtending 2.41° ×
2.41°. The display was divided along the vertical and hor-
izontal meridians to create four quadrants resulting in 35
possible stimulus locations per quadrant. Stimuli were ran-
domly assigned to cells in one quadrant for each trial, E
were positioned in the center of each cell. There were two
set sizes of 6 E 10 stimulus items.
Behavioral Methods
Participants completed four experimental scans (each 14 min
26 sec). Each scan consisted of two blocks of trials: a pre-
view block and a full (baseline) block. There were two
types of trial in the preview block: preview search (n = 20)
and preview only (n = 12). A preview search trial was
composed of two consecutive 2-sec displays. The first was
the preview display which presented half of the distracter
items (either 3 O 5) followed by the second, search, dis-
play, where the remainder of the distracter items (either
2 O 4) plus the target (always present) joined the pre-
viewed items on the screen. Participants indicated, us-
ing a response box held in the right hand, whether the T
was tilted leftward or rightward. If a response was not
made within 2 sec, it was counted as being incorrect. UN
preview-only trial consisted only of the initial preview dis-
play followed by 2 sec of fixation. Participants did not know
when these trials would occur and were instructed not
to respond on these trials. All trials began and finished with
a 1-sec fixation, and feedback was given during the end fixa-
tion for a preview search trial via a change in shape of the
fixation marker (see Figure 1A and C). Trial order and ITI
length (between 4 E 12 sec) were randomly selected
separately for each participant using Optseq2 (http://surfer.
nmr.mgh.harvard.edu/optseq). For each combination of
set size and quadrant there were 10 preview search trials
E 6 preview-only trials across all four scans.
There were also two trial types in the full block: full
search and dummy trials (see Figure 1B and C). These
trials were identical to those in the preview block in terms
of the trial numbers, trial sequence, set sizes, and presen-
tation quadrants. Tuttavia, for the full search trials, IL 3
O 5 distracters presented in the first display disappeared
with the onset of the search display and were replaced by
5 O 9 new distracters (plus the target) in the same quad-
rant. The final number of items in the search display of the
2048
Journal of Cognitive Neuroscience
Volume 23, Numero 8
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
3
l
8
e
–
2
P
0
D
4
F
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
C
6
N
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
P
C
D
N
.
B
sì
2
0
G
1
tu
0
e
.
S
T
2
o
1
N
5
6
0
2
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
T
/
.
F
.
.
o
N
1
8
M
UN
sì
2
0
2
1
full search condition matched the final number in the pre-
view search condition. The dummy trials were visually the
same as preview-only trials.
The order of the two blocks within a scan was counter-
balanced across participants and scans. Each scan began
and ended with 30 sec of fixation. Prior to each block,
there was a 3-sec instruction indicating the following
block type. The two blocks were separated by a 31-sec fix-
ation screen. Participants were instructed to fixate the cen-
tral square for the entirety of the scan, and to use their
peripheral vision to perform the task. Participants were en-
couraged to actively ignore the previewed distracters in
the preview block and were informed that it would not
be useful to ignore the distracters in the first displays of
the trials in the full block. All participants undertook a
practice session outside the scanner prior to the experi-
ment (average correct performance = 84%).
fMRI Methods
Data were acquired using a 3-T Philips Achieva MRI scanner.
Participants lay in the scanner and viewed the projector
screen through a tilted mirror on the eight-channel SENSE
head coil. If necessary, participants wore MRI-compatible
glasses to correct vision. The BOLD signal was measured
using a T2*-weighted echo-planar imaging sequence (con
32 ascending slices, a repetition time of 2000 msec, a time
to echo of 35 msec, a flip angle of 85°, and a resolution of
2.5 mm3). A T1-weighted high-resolution anatomical scan
(1 mm3) was acquired during the same session.
The fMRI Expert Analysis Tool (FEAT) Version 4.0.4 (part
of FMRIBʼs software library, available at www.fmrib.ox.ac.
uk/fsl) was used to process and analyze the data. Prepro-
cessing of each functional scan involved head motion cor-
rection (absolute mean displacements per scan averaged
across participants were 0.48, 0.56, 0.46, E 0.58 mm),
slice-timing correction, nonbrain removal, spatial smooth-
ing (5 mm full width at half maximum Gaussian kernel),
intensity normalization and high-pass Gaussian-weighted
temporal filtering (sigma = 50 sec). Each participantʼs
functional dataset was registered with their anatomical im-
age and then transformed into MNI space. Unexpected
noise and artifacts were removed using Probabilistic Inde-
pendent Component Analysis (Beckmann & Smith, 2004)
implemented in MELODIC (Multivariate Exploratory Linear
Decomposition into Independent Components), part of
FMRIBʼs software library. Only 25 (of a total of 72) del
search scans required component removal, and of these
scans, an average 3.8% of components were removed.
fMRI Analysis
General linear modeling analysis was conducted for each
scan using FILM with local autocorrelation correction
(Woolrich, Ripley, Brady, & Smith, 2001). A design matrix
was created with 16 regressors representing each experi-
mental condition according to the following factorial de-
sign: 4 (trial type: preview search, preview only, full search,
and dummy) by 4 (quadrant: lower-left, lower-right, superiore-
left, and upper-right). Data were collapsed across set size.
Preview search and full search trial regressors were defined
as the duration from the onset of the first display to the
response time. Preview-only and dummy trial regressors
were the duration of the first display (cioè., 2 sec). Trials with
incorrect responses were modeled as regressors of no in-
terest as were the six movement parameters obtained dur-
ing motion correction. All regressors were convolved with
a gamma function.
Contrasts of interest included comparing preview-only
trials with dummy trials for each quadrant separately. Simi-
larly, contrasts were created to compare preview search
with full search trials for each quadrant. Contrasts were com-
bined across runs for each participant using fixed effects
analysis. Group analysis was conducted using FMRIBʼs Lo-
cal Analysis of Mixed Effects ( Woolrich, Behrens, Bedell,
Jenkinson, & Smith, 2004; Beckmann, Jenkinson, & Smith,
2003). Z (Gaussianized T/F) statistic images were thresh-
olded using clusters with Z > 2.1 and a (corrected) cluster
significance threshold of p < .05 (Worsley, Evans, Marrett,
& Neelin, 1992) or an extent threshold of k > 50 and a
significance threshold of p = .05. Group analyses included
a prethreshold mask to limit results to the gray matter.
Time-course data were extracted from each run for each
participant using the Perl Event-related Average Time-
course Extraction tool (www.jonaskaplan.com/peate).
Psychophysiological Interaction Analysis
We conducted psychophysiological interaction (PPI) anal-
yses to examine brain activity functionally connected to
activations identified with the main analyses. To create
the source regions for the PPI analysis, the most significant
voxel from each brain region was identified and a 6-mm
spherical ROI was centered on this voxel. The mean activ-
ity time courses were extracted from the source regions of
each of the participantʼs scans. The first level of an indi-
vidual PPI analysis was conducted on each scan separately
and included 16 regressors that represented the interac-
tion between the time course of the source region and
IL 16 experimental conditions. Contrasts of interest were
preview search–full search and preview only–dummy for
each quadrant (and vice versa). Contrast images were en-
tered into a fixed effects analysis to average data within
each participant and a group-level mixed effects analysis
Z > 1.7 and p < .05 was conducted (as above; Worsley
et al., 1992).
RESULTS
Behavioral Data
Data were collapsed across the separate search scans and
analyses were performed on the preview search trials and
full search trials separately for each set size.
Payne and Allen
2049
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
8
e
-
2
p
0
d
4
f
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
c
6
n
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
6
0
2
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
/
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Table 1. Proportion of Correct Responses for Each Search
Condition and Set Size
Set Size
Full Search
Preview Search
0.84
0.77
0.89
0.86
6
10
Accuracy
We recorded RTs and accuracy. All participants achieved
75% (or more) correct responses (Table 1). A repeated
measures ANOVA with factors of condition (full search,
preview search) and set size (6, 10 items) revealed sig-
nificant main effects of condition [F(1, 17) = 27.7, p =
.00006, partial η2 = 0.62] and set size [F(1, 17) = 15.8,
p = .001, partial η2 = 0.48], and no significant interac-
tion [F(1, 17) = 1.6, p = .22, partial η2 = 0.87]. Because
there was a response deadline in the experiment (2 sec,
after which responses were recorded as errors), the sig-
nificant effects of condition and set size were not un-
expected. RTs were longer in both the full condition and
at the larger set size, thus there was a speed–accuracy
tradeoff.
Reaction Times
We used an adjusted response time measure (RTadj) in-
stead of RT as the dependent measure due to the re-
stricted response period. We divided the average correct
RT for each participant, condition, and set size combina-
tion by the proportion correct for that combination. We
used this adjustment in a recent study (Allen et al., 2008)
where the error rates were also inflated by a response
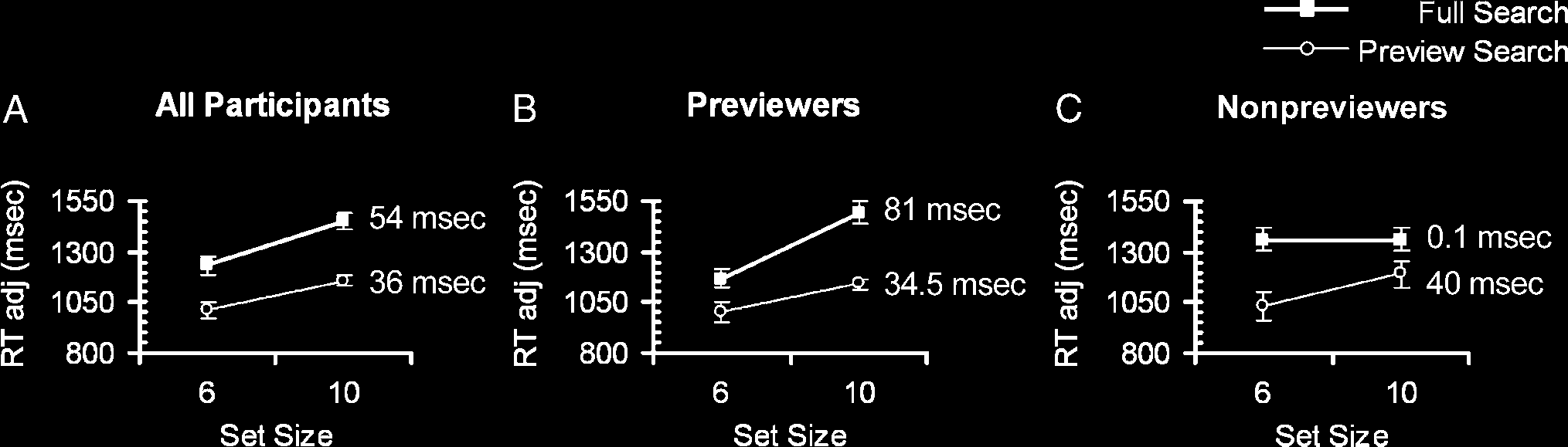
deadline. Figure 2A plots the RTadj against set size for
the full search and preview search conditions.
RTadj data for the correct trials were entered into a re-
peated measures ANOVA with main factors of condition
(preview search, full search) and set size (6, 10 items). De-
spite there being no significant interaction between condi-
tion and set size [F(1, 17) = 1.9, p = .19, partial η2 = 0.1],
performance on full search was significantly slower than
that in preview search [F(1, 17) = 54, p = .000001, partial
η2 = 0.76], suggesting an advantage of the preview dis-
play. Participants were slower with more display items
[F(1, 17) = 33.6, p = .00002, partial η2 = 0.66].
Inspection of individual participant data revealed var-
iations in preview benefit across participants, enabling
us to categorize participants as “previewers” and “non-
previewers.” A total of 12 of the 18 participants were classi-
fied as previewers, identified strictly as those displaying the
standard preview benefit ( Watson & Humphreys, 1997)
measured in terms of an improvement of search efficiency
(in terms of time per item) in the preview condition com-
pared to the full condition. An ANOVA revealed a signifi-
cant interaction between condition and set size for these
participants [F(1, 11) = 15.4, p = .002, partial η2 = 0.58;
Figure 2B]. There was also a significant interaction be-
tween condition and set size for the non-previewers [F(1,
5) = 14.2, p = .013, partial η2 = 0.74], although the full
and preview search slope functions did not conform to
the standard preview benefit (Figure 2C).
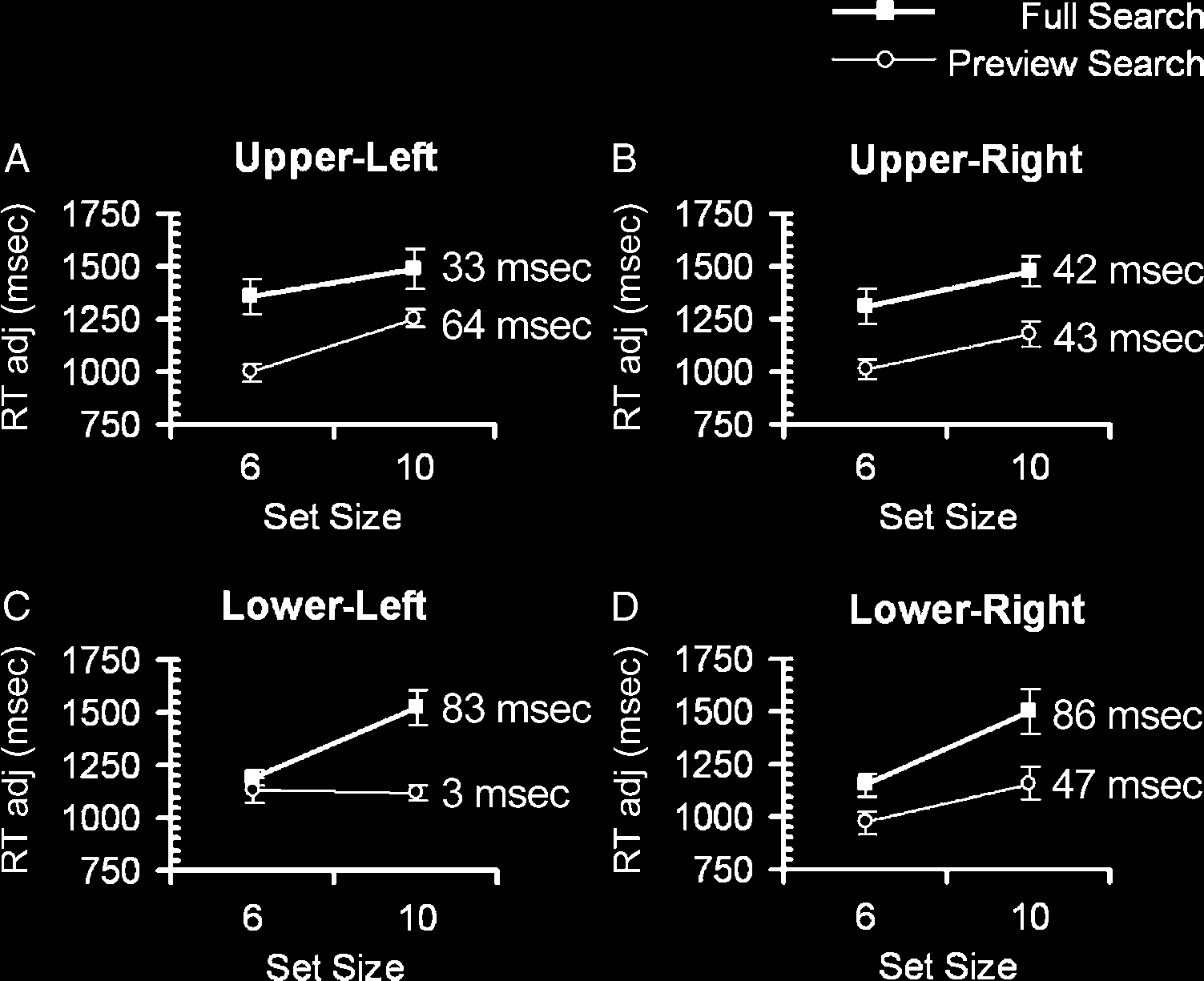
In a similar vein, we observed that there was considerable
variation in preview benefit across quadrants (Figure 3A–
D). For each participant, we calculated RTadj for each
quadrant, set size, and search condition combination, and
separate ANOVAs for each quadrant (including all partici-
pants) revealed a significant interaction between condition
and display size for only the lower-left quadrant [F(1, 17) =
12, p = .003, partial η2 = 0.41]. Thus, the best preview
benefit was found in the lower-left quadrant where search
efficiency for the preview search condition was significantly
better than the efficiency in the full search condition. Fur-
thermore, 13 of the 18 participants displayed a clear pre-
view benefit in terms of slope differences between the
conditions in the lower-left quadrant. Nine participants
previewed in the lower-right and upper-right quadrants,
whereas only six participants previewed in the upper-left
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
8
e
-
2
p
0
d
4
f
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
c
6
n
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
6
0
2
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
t
.
.
f
/
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Adjusted reaction times (RTadj) plotted against set size for the full (filled squares) and preview (unfilled circles) search conditions
averaged across (A) all participants (n = 18), (B) previewers (n = 12), and (C) non-previewers (n = 6). RTadj is measured by RT/proportion correct.
The slope function in terms of time (msec) per search item for each condition is reported. Vertical bars represent ±1 standard error.
2050
Journal of Cognitive Neuroscience
Volume 23, Number 8
Figure 3. Adjusted reaction
times (RTadj) plotted against
set size for the full (filled
squares) and preview
(unfilled circles) search
conditions averaged across
all participants (n = 18).
The search slopes
are plotted for each
quadrant separately:
(A) upper-left quadrant,
(B) upper-right quadrant,
(C) lower-left quadrant, and
(D) lower-right quadrant.
The slope function in terms
of time (msec) per search
item for each condition
is reported. Vertical bars
represent ±1 standard error.
quadrant. Note that the search slopes in the lower-right
quadrant (Figure 3D) indicate a standard preview benefit
for this quadrant, although this is not significant ( p = .26).
It is unclear why there are stronger preview benefits in the
lower visual field (although it is consistent with attention
studies that report a lower visual field advantage for tasks in-
volving attention, e.g., He, Cavanagh, & Intriligator, 1996). It
is possible that the unusual viewing conditions made it easier
(or more difficult) to direct attention to (or from) certain lo-
cations. We used the differences in preview benefit between
quadrants, as well as between previewing and nonpreview-
ing participants, to search for BOLD activity specifically
linked to successful preview by incorporating these behav-
ioral variations into the fMRI analyses. Because the behav-
ioral preview benefit was stronger in some quadrants and
participants, this allows us to find the neural signature relat-
ing to a successful preview benefit while removing activity
relating to merely searching the stimuli or intent to preview.
full search/dummy trials. Preview activity in behavioral
non-previewers was deducted from preview activity in be-
havioral previewers (Table 2). Supporting previous findings
(Dent et al., in press; Allen et al., 2008; Olivers et al., 2005;
Pollmann et al., 2003), we find activation in the precuneus
and SPL that corresponds with ignoring the preview display.
Regardless of what attentional task a participant intends
to perform, they will only be successful if the correct neural
messages are passed to earlier cortical stages. If the intent is
to attend to a particular target, but the appropriate stimulus-
specific neurons are not modulated, then the target will
not be enhanced, for example. Here, only some of our par-
ticipants and quadrants generated a preview benefit (see
above). Thus, we can assume that the activation patterns
that underlie the preview benefit will be stronger in the
more successful preview trials. In our data, regions such as
the SPL were more activated when previewers (as opposed
to non-previewers) had greater preview benefit, which sug-
gested that this was the case (Supplementary Figure).
Imaging Data
fMRI data from all scans from one participant and from
two scans belonging to another participant were discarded
due to excessive head movement.
Preview-related Neural Activity
First, we identified brain areas showing preview-related
activity. Preview search/only trials were contrasted against
Linking Preview-related Neural Activity with Behavioral
Preview Benefit
Here, the critical comparison is between preview-only and
dummy trials as this reveals processes involved with active
ignoring without contamination from the search displays.
For each participant and quadrant separately, dummy tri-
als were deducted from preview-only trials. Because we
were interested in trials where the intent to preview was
Payne and Allen
2051
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
8
e
-
2
p
0
d
4
f
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
c
6
n
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
6
0
2
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
.
.
/
f
.
o
n
1
8
M
a
y
2
0
2
1
Table 2. Areas of Significant Clusters of Preview-related Activity, Averaged across Quadrants, for Behavioral Previewers Compared to
Non-previewers (Extent Threshold = 50, p = .05)
Contrast
Structure
Location x, y, z (mm)
Z Score
Volume ( Voxels)
Preview (search and only)–full
L Lateral occipital cortex/precuneus
(search and dummy)
L Lateral occipital cortex
L Precuneus
L Precentral gyrus
L Supramarginal gyrus
L Superior frontal gyrus
Preview only–dummy
L Lateral occipital cortex/precuneus
R Precentral gyrus
R Lateral occipital cortex/SPL
L Lateral occipital cortex
L Cingulate gyrus
R Cingulate gyrus
Preview search–full search
L Lateral occipital cortex/SPL
L Postcentral gyrus
L Lateral occipital cortex
L Precentral gyrus
R Middle temporal gyrus
−24, −64, 32
−42, −74, 16
−12, −54, 56
−24, −6, 46
−54, −24, 26
0, 32, 52
−22, −64, 32
48, 4, 26
36, −62, 52
−42, −74, 16
−6, −28, 34
10, −42, 38
−28, −62, 60
−52, −26, 50
−40, −74, 16
−24, −6, 48
46, −46, 12
2.56
2.84
2.14
3
2.33
2.22
3.07
2.25
2.73
2.41
2.44
2.14
2.64
2.38
2.54
3.44
2.7
239
115
75
67
67
59
414
194
80
71
50
50
189
100
92
91
69
successfully carried out, quadrants that participants pre-
viewed in were compared to those quadrants in which
participants did not preview. There were three clusters
where activity was greater for previewers compared to non-
previewers (Table 3, Figure 4).
Supporting previous findings (Dent et al., in press; Allen
et al., 2008; Olivers et al., 2005; Pollmann et al., 2003), we
found activation in the left and right precuneus corre-
sponding to ignoring the preview display. Secondly, the
results indicate that actively ignoring visual information
leads to signal changes in early visual cortex.
To investigate whether actively preparing to ignore in-
creased activation in the precuneus and visual cortex, we
extracted the mean BOLD activity from these areas for the
preview only–dummy contrast for each quadrant and par-
ticipant and correlated these signals with the magnitude of
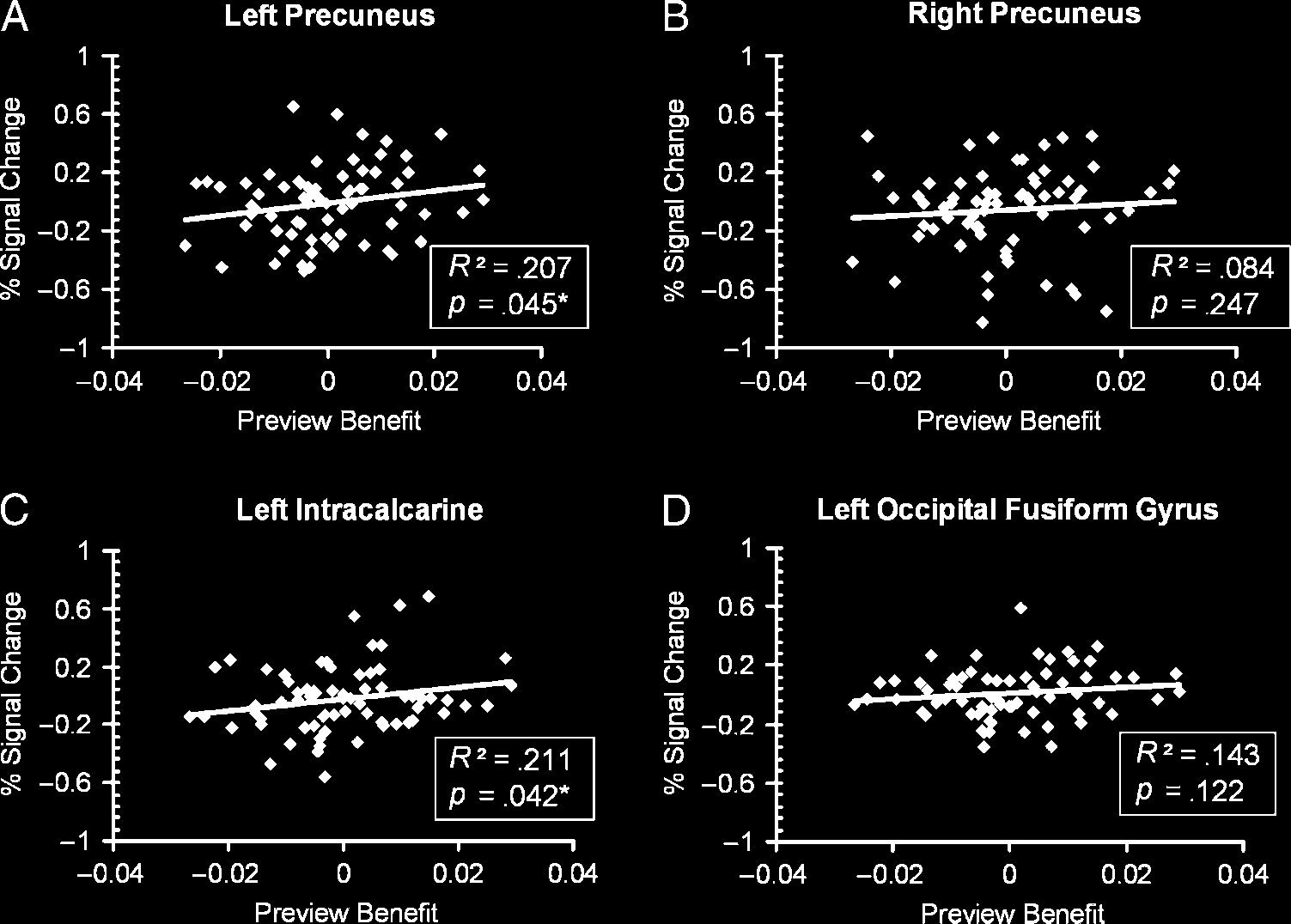
behavioral preview benefit (Figure 5A–D). We extracted
the mean BOLD signal (using Featquery) from spherical
ROIs of 6 mm radius centered on the maximally activated
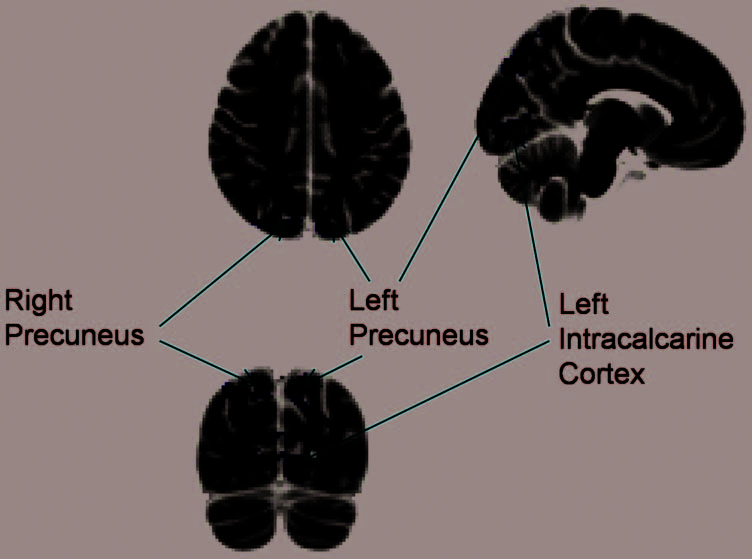
peak voxels in the left precuneus (−8, −78, 40), right pre-
cuneus (14, −82, 42), left intracalcarine cortex (−10, −90,
−2), and left occipital fusiform gyrus (−16, −88, −10). To
calculate the magnitude of behavioral preview benefit for
each participant and quadrant, we used Equation 1:
Preview Benefit ¼ Efficiency : Full Search þ c
Efficiency : Preview Search þ c
ð1Þ
where c is a constant added to reflect that slopes occasion-
ally decreased with increasing set size in the preview con-
dition. The behavioral preview benefit was significantly
correlated with activation in left intracalcarine cortex and
the left precuneus but not with the right precuneus and
left occipital fusiform gyrus.
We further examined the fMRI data in response to preview-
only and dummy trials presented to the quadrant (lower-
left) that showed the strongest benefit from preview.
This contrast revealed significantly greater preview activ-
ity for the behavioral previewers compared to the non-
previewers in occipital cortex (Table 4). Importantly, the
focus of activation in visual cortex is located above the
calcarine sulcus in the right hemisphere, that is, contra-
lateral to the visual field of stimulation, and thus, corre-
sponds to the known retinotopic anatomy of early visual
cortex.
As predicted by the behavioral data, there was no signif-
icant preview-related neural activity from the other presen-
tation quadrants. However, with a threshold of p = .01
(uncorrected, k > 50), an area of early visual cortical activ-
ità (−18, −104, 0; 110 voxels) related to successful pre-
viewing was identified in the left hemisphere for the
lower-right quadrant (cioè., contralateral to the visual field
of stimulation). There were no (k > 50) areas of activation
in visual regions for the upper-left and upper-right quad-
rant analyses using the uncorrected threshold.
2052
Journal of Cognitive Neuroscience
Volume 23, Numero 8
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
3
l
8
e
–
2
P
0
D
4
F
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
C
6
N
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
P
C
D
N
.
B
sì
2
0
G
1
tu
0
e
.
S
T
2
o
1
N
5
6
0
2
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
0
e
2
S
3
T
/
j
.
.
.
/
F
T
o
N
1
8
M
UN
sì
2
0
2
1
Tavolo 3. Three Clusters of Significant Activations for the Preview-only–Dummy Contrast for Quadrants in Which Participants
Behaviorally Previewed Compared to Quadrants in Which Participants Did Not Preview (Cluster Threshold Z = 2.1, P < .05)
Structure
Left Occipital
Location x, y, z (mm)
Z Score
Volume ( Voxels)
L Occipital fusiform gyrus extending to:
−16, −88, −10
3.45
1556
–L Intracalcarine cortex
–L Lateral occipital cortex
–L Lingual gyrus
Left Medial
−10, −90, −2; 0, −94, 4
−44, −72, −10; −36, −86, 2
−6, −76, −8
L Occipital cortex extending to:
−30, −92, 24
3.76
645
–L Precuneus
−8, −78, 40; −10, −76, 54;
−6, −70, 56; −6, −80, 52
–L Lateral occipital cortex
−16, −86, 42
Right Medial
R Lateral occipital cortex extending to:
44, −58, 54
3.64
1199
–R Angular gyrus
–R Precuneus
–R Lateral occipital cortex
42, −54, 56; 50, −52, 56
14, −82, 42
50, −58, 52; 26, −70, 46
Psychophysiological Interaction
We performed functional connectivity analyses to identify
the contribution of the precuneus to activity in other brain
regions. We examined whether the coupling between the
precuneus and other brain areas differed depending on
whether the observers were actively ignoring items in the
first display (i.e., preview condition) compared to when
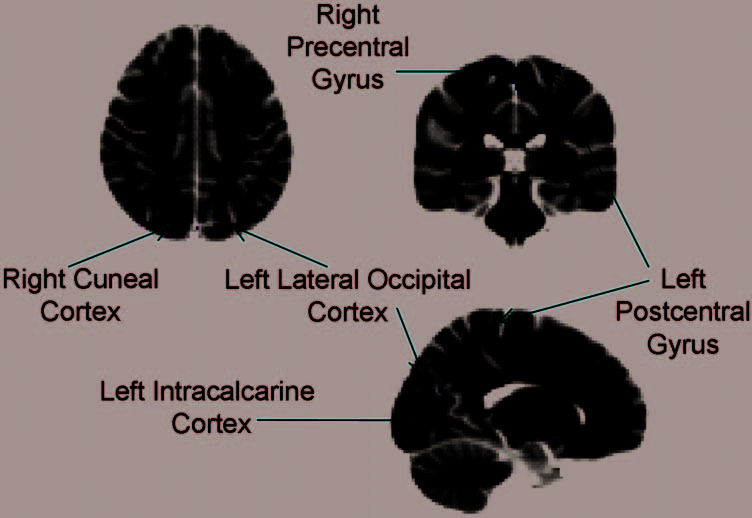
Figure 4. Preview-related
group activation revealed from
the preview only–dummy
contrast for quadrants in which
participants behaviorally
previewed compared to those
quadrants in which participants
did not preview (presented on
the MNI template brain).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
8
e
-
2
p
0
d
4
f
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
c
6
n
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
6
0
2
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
f
t
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Payne and Allen
2053
Figure 5. The magnitude of
behavioral preview benefit
plotted against mean BOLD
signal extracted from the
(A) left precuneus, (B) right
precuneus, (C) left
intracalcarine cortex, and
(D) left occipital fusiform gyrus
for the preview only–dummy
contrast for each participant
and quadrant. One-tailed
Pearsonʼs R2 and p values are
indicated for each area. Each
point represents data from each
quadrant for each participant
(n = 68). *p < .05.
they were not (i.e., full condition). We seeded the separate
PPI analyses in the left (centered at −8, −78, 40) and right
precuneus (centered at 14, −82, 42). The PPI analysis using
the right precuneus as the source region revealed that
there was differential activation within regions of the
fronto-parietal attention network depending on whether
participants were undertaking the dummy condition or
the preview-only condition and whether participants were
previewers or non-previewers. We found that for those par-
ticipants who benefited from the preview, the right pre-
cuneus interacted significantly with the postcentral and
the precentral gyri when participants were completing
the dummy condition in comparison to the preview-only
condition (see Table 5 and Figure 6). Similar clusters of ac-
tivation (R precentral gyrus = 14, −30, 70; L postcentral
gyrus = −56, −18, 34) were found for the left precuneus
source region with a threshold of p = .01 (uncorrected,
k > 50). Precuneus activity during preview-only trials,
relative to dummy trials, was thus related to a decrease
in preview-related activation in attentional control areas
(Corbetta & Shulman, 2002).
For the PPI analyses contrasting full search with pre-
view search, for those participants who benefited from
the preview, both the left and right precuneus interacted
significantly with visual cortical regions when participants
were completing the full search condition in comparison
to the preview search condition (Tavolo 5, Figura 6). Pre-
cuneus activity during preview search trials was thus re-
lated to a decrease in search-related activation in visual
cortex (relative to full search trials). Of particular interest
here is that there is significant activation in early visual
cortex, specifically in intracalcarine cortex (−16, −70,
4). Other areas of activation that overlap across the two
analyses include left lateral occipital cortex and right cu-
neal cortex.
DISCUSSION
Extending previous studies, we found that (Dent et al., In
press; Allen et al., 2008; Olivers et al., 2005; Pollmann
et al., 2003) precuneus activation was associated with suc-
cessfully ignoring distractors. Secondo, preparatory-related
Tavolo 4. Areas of Significant Activations for the Preview Only–Dummy Contrast in the Lower-left Quadrant for Behavioral
Previewers Compared to Non-previewers (Cluster Threshold Z = 2.1, P < .05)
Structure
Location x, y, z (mm)
R Intracalcarine cortex extending to:
–R Occipital pole
–R Lingual gyrus
–L Lingual gyrus
6, −82, 10
8, −92, 20
8, −74, −4; 4, −82, −8
−4, −74, −6
Z Score
3.33
Volume ( Voxels)
901
2054
Journal of Cognitive Neuroscience
Volume 23, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
8
e
-
2
p
0
d
4
f
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
c
6
n
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
6
0
2
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
t
f
.
.
o
n
1
8
M
a
y
2
0
2
1
Table 5. Areas That Significantly Interact with the L Precuneus and the R Precuneus for the Dummy/Full Search–Preview-only/
Preview Search Contrasts for Quadrants in Which Participants Behaviorally Previewed Compared to Quadrants in Which
Participants Did Not Preview (Cluster Threshold Z = 1.7, p < .05)
Contrast
Structure
Location x, y, z (mm)
Z Score
Volume ( Voxels)
Dummy–preview
Source region: R Precuneus
only
L Postcentral gyrus extending to:
−32, −36, 70
3.45
1466
–R Precentral gyrus
24, −22, 62; 10, −30, 74;
–R Postcentral gyrus
Full search–
preview search
Source region: L Precuneus
L Cuneal cortex extending to:
–L Intracalcarine cortex
–L Lateral occipital cortex
–R Cuneal cortex
Source region: R Precuneus
22, −26, 74
16, −30, 70
−8, −88, 24
−16, −70, 4
−22, −82, 36; −28, −72, 20
8, −84, 40; 18, −72, 24
3.58
4712
L Lateral occipital cortex extending to:
−24, −82, 38
4.13
5628
–L Intracalcarine cortex
–R Cuneal cortex
–R Lateral occipital cortex
−16, −78, 2; −16, −70, 4
18, −70, 22
16, −84, 38
activity also increased neural activity in early visual cortex
during this active ignoring process. Third, precentral and
postcentral gyri activation (part of the fronto-parietal atten-
tion network), driven by the precuneus, was reduced when
successful previewers were preparing to ignore distracters.
Fourth, search-related activation in early visual cortex,
driven by the precuneus, was reduced when participants
were able to successfully ignore early presented distracters.
Figure 6. Results of PPI
analyses. Yellow: BOLD activity
driven by the right precuneus
when previewing participants
(relative to non-previewers)
completed the dummy trials
relative to the preview-only
trials. Green: BOLD activity
driven by both the left
precuneus and the right
precuneus when previewing
participants (relative to
non-previewers) completed
the full search trials relative
to the preview search trials.
Activations are presented on
the MNI template brain.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
8
e
-
2
p
0
d
4
f
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
c
6
n
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
6
0
2
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
/
f
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Payne and Allen
2055
Preview-related Preparatory Activity in Early
Visual Cortex
Previous studies have demonstrated preparatory activity in
visual cortex for targets (Macaluso et al., 2003; Hopfinger
et al., 2000; Kastner et al., 1999) and distracters (Ruff &
Driver, 2006; Serences et al., 2004). Here, preparatory
activity associated with previewing irrelevant distracters
occurred in early visual cortex and could be easily sepa-
rated from target-related activity. Our results demonstrate
increased neural activity in primary visual cortex (in likely
V1 identified using WFU PickAtlas: Maldjian, Laurienti,
Kraft, & Burdette, 2003) that was quadrant specific. Con-
sistent with the finding that increased preview-related
BOLD activation in early visual cortex is correlated with
the magnitude of behavioral preview benefit, the trials
presented to the upper visual quadrants did not display
a behavioral preview benefit or enhanced neural activity.
The asymmetry of preview performance between upper
and lower visual quadrants may be explained by a com-
monly reported behavioral finding that visual processing
is enhanced in the lower visual field compared to the
upper visual field (e.g., Danckert & Goodale, 2001; Rubin,
Nakayama, & Shapley, 1996). In particular, it has been re-
ported that there is an advantage for attentional process-
ing in the lower visual field (He et al., 1996). The portion
of visual cortex in the macaque monkey brain that repre-
sents the lower visual field projects considerably more to
posterior parietal cortex, an area involved with spatial at-
tention, than does the portion that represents the upper
visual field (Maunsell & Newsome, 1987). This may explain
the lower visual field advantage with attentional tasks in
humans. However, the asymmetry seen here is weak with
only the lower-left quadrant showing a significant advan-
tage for the preview condition. Further research is re-
quired to assess the specificity of the preview benefit to
visual field location.
Precuneus Activity
There was a bilateral increase in precuneus neural activity
for the preview-only trials relative to the dummy trials. The
precuneus is consistently activated (Dent et al., in press;
Allen et al., 2008; Olivers et al., 2005; Pollmann et al., 2003)
in response to preview trials, suggesting this region plays
a critical role in visual marking. This brain area is thought
to be involved with a variety of higher-level cognitive func-
tions including episodic memory, consciousness, and visual–
spatial imagery (see Cavanna & Trimble, 2006 for a review).
Allen et al. (2008) found that the precuneus was activated
both by a visual working memory task and by a preview
search task, and that the former could interfere with the
latter. This provides evidence that the precuneus is in-
volved with encoding the spatial representations of the
old, to-be-inhibited, items in memory. Indeed, a second-
ary auditory task did not interfere with the preview task
if it was presented 1 sec after the presentation of the pre-
view, whereas a secondary visual task did interfere, imply-
ing that the visual memory representation of the old
distracter items is critical (Humphreys et al., 2002).
Functional Connectivity Analysis
We report that activity in the precuneus is more con-
nected with fronto-parietal activation for the dummy con-
dition relative to the preview-only condition in successful
previewers. The pre/postcentral cortical activation we dis-
cerned has been implicated in several functional imaging
studies that have used spatial attention tasks (Donner
et al., 2000; Culham et al., 1998; see Corbetta & Shulman,
2002 for a review). Although often considered to be pas-
sively viewed, the dummy trials informed observers as to
the location of the subsequent stimulus and to prepare
for a relatively difficult search task. It seems reasonable,
therefore, that there is a strong connection with areas in-
volved in attentional control and orienting in these trials. In
support, a visual search study by Weidner, Krummenacher,
Reimann, Müller, and Fink (2008) found that decreased
target saliency increased BOLD activation in several areas
of the fronto-parietal attention network, including pre/
postcentral cortex. For preview search, it seems that these
orienting signals are comparatively weak in comparison to
the preview-related activity.
We also report that activity in the precuneus is related
to decreases in early visual cortex activation for the pre-
view search condition relative to the full search condition
in successful previewers. This is in contrast to the increase
in early visual activation for the preview-only condition rel-
ative to the dummy. Thus, it seems that in early visual cor-
tex, successful ignoring of the previewed distracters is
related to, first, an increase in activity but also an overall
down modulation of activity by the precuneus. This is con-
sistent with recent studies comparing distracter encoding
during the preview display that also show an increase in
activity associated with the preview benefit (Allen et al.,
2008; Pollmann et al., 2003). Similarly, Humphreys et al.
(2004) presented probe dots early (200 msec) and late
(800 msec) in a preview display. As with previous probe-
dot detection studies (Olivers & Humphreys, 2002; Watson
& Humphreys, 2000), late probes gave rise to longer RTs
when they appeared at the locations of the old distracters
than when they occurred at the location of the new dis-
tracters, suggesting inhibition of the old distracters. How-
ever, detection of early probes was actually facilitated,
suggesting that participants initially attend to the old dis-
tracters before inhibiting them. Similarly, results from an
event-related potential study (Belopolsky, Peterson, &
Kramer, 2005) suggest that the inhibition of old distracters
is applied toward the end of the preview interval. Of course,
such an explanation requires that a decrease in BOLD signal
means a decrease in neural activity (see Shmuel, Augath,
Oeltermann, & Logothetis, 2006).
The processes underlying the preview benefit can be
separated into two components (Humphreys et al., 2002).
2056
Journal of Cognitive Neuroscience
Volume 23, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
8
e
-
2
p
0
d
4
f
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
c
6
n
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
6
0
2
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
f
.
t
.
.
o
n
1
8
M
a
y
2
0
2
1
First, observers attend and encode the previewed dis-
tracters to form a representation of the items in memory.
Second, this representation is inhibited and the new items
are prioritized. Thus, the increased activation for pre-
view during the first display is likely to represent active
attentional encoding of the distracters. The decreased
activation for preview (relative to full search) is likely to
reflect that in the full search condition there were effec-
tively more items to search through. It is important to note,
however, that the data cannot tell us whether activation in
the preview search condition was suppressed relative to
the full search condition or whether activation in the full
search condition was increased relative to the preview
search condition.
Conclusion
We used the preview search paradigm to investigate
whether there are neural signatures relating to ignoring
known distracters. We found that in order to benefit from
the preview, observers must prepare to inhibit old dis-
tracters, leading to an increase in activation in visual and
parietal regions. Subsequently, further processing of these
items is suppressed. The precuneus is likely to be involved
with representation of the previewed distracters and our
results suggest that the precuneus modulates activity in
early visual cortex. In particular, the precuneus is involved
with inhibiting further processing in visual cortex of the
encoded, but excluded, distracters.
Braithwaite, J. J., & Humphreys, G. W. (2003). Inhibition
and anticipation in visual search: Evidence from effects of
color foreknowledge on preview search. Perception &
Psychophysics, 65, 213–237.
Carrasco, M., Penpeci-Talgar, C., & Eckstein, M. (2000).
Spatial covert attention increases contrast sensitivity across
the CSF: Support for signal enhancement. Vision Research,
40, 1203–1215.
Cavanna, A. E., & Trimble, M. R. (2006). The precuneus: A
review of its functional anatomy and behavioural correlates.
Brain, 129, 564–583.
Corbetta, M., Kincade, J. M., Ollinger, J. M., McAvoy, M. P., &
Shulman, G. L. (2000). Voluntary orienting is dissociated
from target detection in human posterior parietal
cortex. Nature Neuroscience, 3, 292–297.
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neuroscience, 3, 201–215.
Culham, J. C., Brandt, S. A., Cavanagh, P., Kanwisher, N. G.,
Dale, A. M., & Tootell, R. B. H. (1998). Cortical fMRI
activation produced by attentive tracking of moving targets.
Journal of Neurophysiology, 80, 2657–2670.
Danckert, J., & Goodale, M. A. (2001). Superior performance
for visually guided pointing in the lower visual field.
Experimental Brain Research, 137, 303–308.
Dent, K., Allen, H. A., & Humphreys, G. W. (in press).
Comparing segmentation by time and by motion in visual
search: An fMRI investigation. Journal of Cognitive
Neuroscience.
Donk, M., & Theeuwes, J. (2001). Visual marking beside the
lark: Prioritizing selection by abrupt onsets. Perception &
Psychophysics, 63, 891–900.
Donner, T., Kettermann, A., Diesch, E., Ostendorf, F., Villringer,
A., & Brandt, S. A. (2000). Involvement of the human
frontal eye field and multiple parietal areas in covert visual
selection during conjunction search. European Journal
of Neuroscience, 12, 3407–3414.
Reprint requests should be sent to Harriet A. Allen, Brain and
Behavioural Sciences, School of Psychology, University of
Birmingham, Edgbaston, Birmingham, B15 2TT, United Kingdom,
or via e-mail: h.a.allen@bham.ac.uk.
Dougherty, R. F., Koch, V. M., Brewer, A. A., Fischer, B.,
Modersitzki, J., & Wandell, B. A. (2003). Visual field
representations and locations of visual areas V1/2/3 in
human visual cortex. Journal of Vision, 3, 586–598.
REFERENCES
Allen, H. A., & Humphreys, G. W. (2007a). A psychophysical
investigation into the preview benefit in visual search.
Vision Research, 37, 735–745.
Allen, H. A., & Humphreys, G. W. (2007b). Previewing
distracters reduces their effective contrast. Vision Research,
47, 2992–3000.
Allen, H. A., Humphreys, G. W., & Matthews, P. M. (2008). A
neural marker of content-specific active ignoring. Journal of
Experimental Psychology: Human Perception and
Performance, 34, 286–297.
Gandhi, S. P., Heeger, D. J., & Boynton, G. M. (1999). Spatial
attention affects brain activity in human primary visual
cortex. Proceedings of the National Academy of Sciences,
U.S.A., 96, 3314–3319.
Gazzaley, A., Cooney, J. W., McEvoy, K., Knight, R. T., &
DʼEsposito, M. (2005). Top–down enhancement and
suppression of the magnitude and speed of neural activity.
Journal of Cognitive Neuroscience, 17, 507–517.
He, S., Cavanagh, P., & Intriligator, J. (1996). Attentional
resolution and the locus of visual awareness. Nature, 383,
334–337.
Hopfinger, J. B., Buonocore, M. H., & Mangun, G. R. (2000).
The neural mechanisms of top–down attentional control.
Nature Neuroscience, 3, 284–291.
Beckmann, C. F., Jenkinson, M., & Smith, S. M. (2003). General
Horton, J. C., & Hoyt, W. F. (1991). The representation of
multi-level linear modelling for group analysis in fMRI.
Neuroimage, 20, 1052–1063.
Beckmann, C. F., & Smith, S. M. (2004). Probabilistic
independent component analysis for functional magnetic
resonance imaging. IEEE Transactions on Medical Imaging,
23, 137–152.
Belopolsky, A. V., Peterson, M. S., & Kramer, A. F. (2005). Visual
search in temporally segregated displays: Converging
operations in the study of the preview benefit. Cognitive
Brain Research, 24, 453–466.
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436.
the visual field in human striate cortex. Archives of
Ophthalmology, 109, 816–824.
Humphreys, G. W., Stalmann, B. J., & Olivers, C. (2004). An
analysis of the time course of attention in preview search.
Perception & Psychophysics, 66, 713–730.
Humphreys, G. W., Watson, D. G., & Jolicœur, P. (2002).
Fractionating the preview benefit in search: Dual-task
decomposition of visual marking by timing and modality.
Journal of Experimental Psychology: Human Perception
and Performance, 28, 640–660.
Jiang, Y. H., Chun, M. M., & Marks, L. E. (2002). Visual marking:
Selective attention to asynchronous temporal groups.
Payne and Allen
2057
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
8
e
-
2
p
0
d
4
f
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
c
6
n
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
6
0
2
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
t
.
.
.
/
o
n
1
8
M
a
y
2
0
2
1
Journal of Experimental Psychology: Human Perception
and Performance, 28, 717–730.
Kastner, S., Pinsk, M. A., De Weerd, P., Desimone, R., &
Ungerleider, L. G. (1999). Increased activity in human visual
cortex during directed attention in the absence of visual
stimulation. Neuron, 22, 751–761.
Macaluso, E., Eimer, M., Frith, C. D., & Driver, J. (2003).
Preparatory states in crossmodal spatial attention: Spatial
specificity and possible control mechanisms. Experimental
Brain Research, 149, 62–74.
Maldjian, J. A., Laurienti, P. J., Kraft, R. A., & Burdette, J. H.
(2003). An automated method for neuroanatomic and
cytoarchitectonic atlas-based interrogation of fMRI data
sets. Neuroimage, 19, 1233–1239.
Martínez, A., Anllo-Vento, L., Sereno, M. I., Frank, L. R., Buxton,
R. B., Dubowitz, D. J., et al. (1999). Involvement of striate
and extrastriate visual cortical areas in spatial attention.
Nature Neuroscience, 2, 364–369.
Maunsell, J. H. R., & Newsome, W. T. (1987). Visual processing
in monkey extrastriate cortex. Annual Review of
Neuroscience, 10, 363–401.
Mavritsaki, E., Allen, H., & Humphreys, G. (2009). Model based
analysis of fMRI-data: Applying the sSoTS framework to
the neural basis of preview search. In L. Paletta & J. K. Tsotsos
(Eds.), Attention in cognitive systems (pp. 124–138).
Berlin: Springer.
Olivers, C. N. L., & Humphreys, G. W. (2002). When visual
marking meets the attentional blink: More evidence for
top–down, limited-capacity inhibition. Journal of
Experimental Psychology: Human Perception and
Performance, 28, 22–42.
Olivers, C. N. L., & Humphreys, G. W. (2003). Visual marking
inhibits singleton capture. Cognitive Psychology, 47, 1–42.
Olivers, C. N. L., Smith, S., Matthews, P., & Humphreys, G. W.
(2005). Prioritizing new over old: An fMRI study of the
preview search task. Human Brain Mapping, 24, 69–78.
Pelli, D. G. (1997). The VideoToolbox software for visual
psychophysics: Transforming numbers into movies.
Spatial Vision, 10, 437–442.
Pessoa, L., Kastner, S., & Ungerleider, L. G. (2003).
Neuroimaging studies of attention: From modulation of
sensory processing to top–down control. Journal of
Neuroscience, 23, 3990–3998.
Pollmann, S., Weidner, R., Humphreys, G. W., Olivers, C. N. L.,
Müller, K., Lohmann, G., et al. (2003). Separating distractor
rejection and target detection in posterior parietal cortex:
An event-related fMRI study of visual marking. Neuroimage,
18, 310–323.
Posner, M. I., Snyder, C. R. R., & Davidson, B. J. (1980).
Attention and the detection of signals. Journal of
Experimental Psychology: General, 109, 160–174.
Ress, D., Backus, B. T., & Heeger, D. J. (2000). Activity in
primary visual cortex predicts performance in a visual
detection task. Nature Neuroscience, 3, 940–945.
Rubin, N., Nakayama, K., & Shapley, R. (1996). Enhanced
perception of illusory contours in the lower versus upper
hemifields. Science, 271, 651–653.
Ruff, C. C., & Driver, J. (2006). Attentional preparation for
a lateralized visual distractor: Behavioural and fMRI
evidence. Journal of Cognitive Neuroscience, 18,
522–538.
Serences, J. T., Yantis, S., Culberson, A., & Awh, E. (2004).
Preparatory activity in visual cortex indexes distractor
suppression during covert spatial orienting. Journal of
Neurophysiology, 92, 3538–3545.
Shmuel, A., Augath, M., Oeltermann, A., & Logothetis, N. K.
(2006). Negative functional MRI response correlates with
decreases in neuronal activity in monkey visual area V1.
Nature Neuroscience, 9, 569–577.
Shulman, G. L., Ollinger, J. M., Akbudak, E., Conturo, T. E.,
Snyder, A. Z., Petersen, S. E., et al. (1999). Areas
involved in encoding and applying directional expectations
to moving objects. Journal of Neuroscience, 19,
9480–9496.
Smith, A. T., Singh, K. D., & Greenlee, M. W. (2000).
Attentional suppression of activity in the human visual
cortex. NeuroReport, 11, 271–277.
Somers, D. C., Dale, A. M., Seiffert, A. E., & Tootell, R. B. H.
(1999). Functional MRI reveals spatially specific attentional
modulation in human primary visual cortex. Proceedings
of the National Academy of Sciences, U.S.A., 96,
1663–1668.
Sylvester, C. M., Jack, A. I., Corbetta, M., & Shulman, G. L.
(2008). Anticipatory suppression of nonattended locations
in visual cortex marks target location and predicts
perception. Journal of Neuroscience, 28, 6549–6556.
Watson, D. G. (2001). Visual marking in moving displays:
Feature-based inhibition is not necessary. Perception &
Psychophysics, 63, 74–84.
Watson, D. G., & Humphreys, G. W. (1997). Visual marking:
Prioritizing selection for new objects by top–down
attentional inhibition of old objects. Psychological Review,
104, 90–122.
Watson, D. G., & Humphreys, G. W. (2000). Visual marking:
Evidence for inhibition using a probe-dot detection
paradigm. Perception & Psychophysics, 62, 471–481.
Weidner, R., Krummenacher, J., Reimann, B., Müller, H. J.,
& Fink, G. R. (2008). Sources of top–down control in
visual search. Journal of Cognitive Neuroscience, 21,
2100–2113.
Woolrich, M. W., Behrens, T. E. J., Bedell, H. E., Jenkinson, M.,
& Smith, S. M. (2004). Multi-level linear modelling for
fMRI group analysis using Bayesian inference. Neuroimage,
21, 1732–1747.
Woolrich, M. W., Ripley, B. D., Brady, J. M., & Smith, S. M.
(2001). Temporal autocorrelation in univariate linear
modelling of fMRI data. Neuroimage, 14, 1370–1386.
Worsley, K. J., Evans, A. C., Marrett, S., & Neelin, P. (1992).
A three-dimensional statistical analysis for CBF activation
studies in human brain. Journal of Cerebral Blood Flow
and Metabolism, 12, 900–918.
Yeshurun, Y., & Carrasco, M. (1999). Spatial attention improves
performance in spatial resolution tasks. Vision Research,
39, 293–306.
2058
Journal of Cognitive Neuroscience
Volume 23, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
3
l
8
e
-
2
p
0
d
4
f
6
/
1
2
9
3
4
/
1
8
5
/
8
2
7
0
o
4
c
6
n
/
1
2
0
7
1
7
0
5
9
2
4
1
8
5
6
/
2
j
o
p
c
d
n
.
b
y
2
0
g
1
u
0
e
.
s
t
2
o
1
n
5
6
0
2
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
.
/
t
.
o
n
1
8
M
a
y
2
0
2
1