Act Now, Play Later: Temporal Expectations Regarding
the Onset of Self-initiated Sensations Can Be
Modified with Behavioral Training
Ruth B. Elijah, Mike E. Le Pelley, and Thomas J. Whitford
Astratto
■ Mechanisms of motor-sensory prediction are dependent
on expectations regarding when self-generated feedback will
occur. Existing behavioral and electrophysiological research sug-
gests that we have a default expectation for immediate sensory
feedback after executing an action. Tuttavia, studies investigat-
ing the adaptability of this temporal expectation have been lim-
ited in their ability to differentiate modified expectations per se
from effects of stimulus repetition. Here, we use a novel, within-
participant procedure that allowed us to disentangle the effect
of repetition from expectation and allowed us to determine
whether the default assumption for immediate feedback is fixed
and resistant to modification or is amenable to change with ex-
perience. While EEG was recorded, 45 participants completed a
task in which they repeatedly pressed a button to produce a
tone that occurred immediately after the button press (imme-
diate training) or after a 100-msec delay (delayed training).
The results revealed significant differences in the patterns of
cortical change across the two training conditions. Specifically,
there was a significant reduction in the cortical response to
tones across delayed training blocks but no significant change
across immediate training blocks. Inoltre, experience with
delayed training did not result in increased cortical activity in
response to immediate feedback. These findings suggest that
experience with action–sensation delays broadens the window
of temporal expectations, allowing for the simultaneous antici-
pation of both delayed and immediate motor-sensory feedback.
This research provides insights into the mechanisms underlying
motor-sensory prediction and may represent a novel therapeutic
avenue for psychotic symptoms, which are ostensibly associated
with sensory prediction abnormalities. ■
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
J
o
C
N
_
UN
_
0
1
2
6
9
P
D
.
F
B
e
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
INTRODUCTION
The ability to determine the origin of sensations is impor-
tant for our adaptive behavior. Per esempio, the sound of
footsteps in a dark parking lot is not alarming if we deter-
mine that we have created the sound ourselves but could
be a cause for alarm if we determine that the footsteps
are generated externally by another agent (Stetson, Cui,
Montague, & Eagleman, 2006). A critical distinction that
our perceptual system makes when attempting to distin-
guish between self-generated and externally generated
sensations relates to their predictability. In contrast to
externally generated sensations, self-generated sensations
are typically more predictable in nature. This inherent
predictability means that the sensory consequences of
self-initiated movements can be anticipated before their
occurrence (Roussel, Hughes, & Waszak, 2013; Bays,
Flanagan, & Wolpert, 2006; Wolpert, Ghahramani, &
Jordan, 1995). This predictability is associated with a de-
crease in the perceived salience of sensations compared
with equivalent sensations that are externally produced, UN
phenomenon known as sensory attenuation (Hughes,
Desantis, & Waszak, 2013UN; Ford & Mathalon, 2004;
University of New South Wales
© 2018 Istituto di Tecnologia del Massachussetts
Blakemore, Wolpert, & Frith, 1998). Sensory attenuation
has been formalized in feed-forward models of motor con-
trol, which postulate that, when an action is self-initiated,
an efference copy of the motor command is produced
and used to predict the sensory consequences of that action
(Crapse & Sommer, 2008; Wolpert & Miall, 1996; Wolpert
et al., 1995; von Holst, 1954). This efference copy acts to
attenuate the (predictable) sensory experience resulting
from a self-generated action, relative to when the sensory
experience is produced by an external agent and hence is
less predictable.
An assumption of the forward model is that self-produced
sensations that are not accurately predicted in terms of
their physical and temporal characteristics will be subject
to less attenuation than self-produced sensations, Quale
are better predicted (Roussel et al., 2013; Blakemore, Frith,
& Wolpert, 1999). There is substantial evidence in support
of this hypothesis (Hughes, Desantis, & Waszak, 2013B;
Behroozmand, Liu, & Larson, 2011; Bäss, Jacobsen, &
Schröger, 2008; Heinks-Maldonado, Nagarajan, & Houde,
2006; Bays, Wolpert, & Flanagan, 2005; Heinks-Maldonado,
Mathalon, Grigio, & Ford, 2005; McClure, Berns, & Montague,
2003; Houde, Nagarajan, Sekihara, & Merzenich, 2002;
Blakemore et al., 1999). One aspect of sensory prediction
involves anticipating when sensations will occur (Arnal &
Journal of Cognitive Neuroscience 30:8, pag. 1145–1156
doi:10.1162/jocn_a_01269
Giraud, 2012; McClure et al., 2003). Several studies have
investigated how modifying the onset of self-initiated sen-
sations alters the degree of sensory attenuation (Oestreich
et al., 2016; Whitford et al., 2011; Aliu, Houde, & Nagarajan,
2009; Bäss et al., 2008; Bays et al., 2005; Blakemore et al.,
1999). In these studies, participants pressed a button to
either produce a sound (Oestreich et al., 2016; Whitford
et al., 2011; Aliu et al., 2009; Bäss et al., 2008) or receive
tactile stimulation (Bays et al., 2005; Blakemore et al.,
1999). In these studies, the onset of sensory feedback was
manipulated; it ranged from occurring immediately after
the button press (cioè., 0-msec delay) or after some delay
ranging from 50 A 500 msec. Consistently across these
studies, it was found that imposing a delay between the
action and resultant sensation caused a reduction in the
amount of sensory attenuation compared with immediate
feedback, with larger delays associated with lower levels of
sensory attenuation. These results suggest that we have a
default expectation that sensory feedback will follow imme-
diately from self-generated actions and that deviations
from this expectation result in prediction errors that can
produce an increase in the salience of the experienced
sensazioni (van Laarhoven, Stekelenburg, & Vroomen,
2017; Bendixen, SanMiguel, & Schröger, 2012).
The aim of this study was to investigate whether the
expectation that sensations follow immediately from ac-
tions is fixed or whether it can be modified with experi-
ence. Predictive coding accounts of sensory attenuation
argue that prediction errors, such as those caused by de-
layed sensory feedback, are critical teaching signals that
facilitate new learning and recalibrate or update our sen-
sory expectations (Bogacz, 2017; Marrone, Adams, Parees,
Edwards, & Friston, 2013; Friston, 2005; Rao & Ballard,
1999). According to such accounts, repeated exposure to
temporally delayed sensory feedback could lead to a
recalibration of the expectation that “sensations follow
immediately from actions.” However, this raises the ques-
tion of whether all sensory expectations are malleable as a
function of experience or whether more weight is placed
on certain “fundamental” predictions (such as the pre-
diction that sensations follow immediately from actions),
such that these predictions are resistant to modification
on the basis of experience.
On the one hand, there seems to be clear value in the
perceptual system updating its expectations in response
to changes in perceptual experience. Per esempio, visuo-
motor recalibrations take place via trial-and-error learning
when reaching for objects or when walking while wearing
laterally displacing prism glasses; this also results in a
negative aftereffect when the glasses are removed (move-
ment biases opposite to the displacement of the glasses),
which again results in visuomotor adjustment with expe-
rience to maintain goal-directed actions (Hatada, Miall, &
Rossetti, 2006; Morton & Bastian, 2004). Inoltre,
with regard to the temporal association between actions
and sensations, Stetson et al. (2006) have shown that
repeated exposure to a delay between a button press and
a subsequent visual stimulus results in a change in tempo-
ral order judgments, such that participants were more likely
to judge a visual stimulus that occurred immediately after
an action as occurring before the button press. In other
parole, participants appeared to adopt a new baseline for
their temporal expectations consistent with their experi-
ence of delayed feedback from their actions. Stetson
et al. (2006) suggested that these temporal recalibrations
are adaptive and occur across the life span. Per esempio,
in the haptic system, the delay between actions and resul-
tant sensations changes as we grow and our limbs elongate
(per esempio., when touching your thumb to a surface, the resultant
reafference takes longer to reach the brain if your arm is
50 cm long than if it is 10 cm long). The implication is that
accurate and efficient sensory modulation would benefit
from updating predictions regarding the timing of sensory
signals across the course of development.
An alternate hypothesis is that we hold relatively rigid
expectations regarding certain key predictions, ad esempio
the prediction that “sensations follow immediately from
actions.” Such predictions develop from a lifetime of
experience and, as such, might be expected to be less
susceptible to change. There is evidence to suggest that
we can hold fixed heuristics to optimize information pro-
cessazione. Per esempio, in the perceptual domain, the axiom
that “light shines from above” seems to be inherent or
fixed as it governs perceptual experience early in devel-
opment and is resistant to modification with experience
(Champion & Adams, 2007; Kleffner & Ramachandran,
1992; Hershberger, 1970; Hess, 1950). Research has
shown that newborn chicks reared in an environment
where light shone from above or light shone from below
preferentially pecked to pictures of seeds where the
shadows were consistent with a light source shining from
above, irrespective of the environment they were reared
in and when controlling for learning to peck based on
depth cues (Hershberger, 1970; Hess, 1950). Inoltre,
Champion and Adams (2007) used a visual search task in
humans to show that visual–haptic training of stimuli with
different lighting orientations did not result in a recalib-
ration of the prior belief that “light shines from above,"
as evidenced by no change in visual search performance
after training. This suggests that a prior of “light shines
from above,” by default, governs perception at the pre-
attentive stage of processing, despite training experience
to the contrary. Extending from this parameter, it is con-
ceivable that the expectation that “sensations follow
immediately from actions” may be an unmalleable percep-
tual rule that governs sensory experience and that persists
even despite evidence to the contrary.
As such, the current study aimed to determine whether
expectations regarding the timing of self-generated
sensations are modifiable with training. Participants com-
pleted a task in which they repeatedly pressed a button
to produce a tone, while EEG was continuously recorded.
All participants underwent two training conditions, IL
order of which was randomized between participants. In
1146
Journal of Cognitive Neuroscience
Volume 30, Numero 8
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
J
o
C
N
_
UN
_
0
1
2
6
9
P
D
.
F
B
e
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
the first condition, the tone was presented immediately
subsequent to the button press; in the second condition,
the tone was presented 100 msec subsequent to the
button press. The dependent variable in this study was
the amplitude of the N1 component of the ERP elicited
by the tone. This component was of particular interest be-
cause prior research has shown that (other factors being
equal) louder sounds evoke larger N1 amplitudes than
softer sounds (Simmons, Nathan, Berger, & Allen, 2011;
Mulert et al., 2005; Näätänen & Picton, 1987). The implica-
tion is that N1 amplitude provides a proxy measure of the
perceived loudness of an auditory sensation and has been
used as a standard measure of sensory suppression in past
research (Timm, Schönwiesner, Schröger, & SanMiguel,
2016; SanMiguel, Todd, & Schröger, 2013; Bäss, Horváth,
Jacobsen, & Schröger, 2011; Ford et al., 2001). Given that
the focus of the current study is not on sensory suppression
per se (cioè., a difference in N1 amplitude from a task where
participants passively listen to tones) but rather differences
in the predictability of self-generated sensations (cioè., UN
difference in N1 amplitude between delayed and imme-
diate tones), it is also worth noting that N1 has been used
to index sensory expectations, whereby unexpected stim-
uli typically evoke a larger N1 than expected stimuli (van
Laarhoven et al., 2017; Oestreich et al., 2016; Hughes et al.,
2013UN; Lange, 2011; Bäss et al., 2008; Heinks-Maldonado
et al., 2006; Heinks-Maldonado et al., 2005).
It was hypothesized that, before training, temporally
delayed tones would be unpredicted by the motor-
perceptual system, in keeping with the default assump-
tion that “sensations follow immediately from actions.”
This would be reflected in a larger N1 amplitude elicited
by delayed tones compared with immediate tones, be-
fore training. Tuttavia, if participants updated their sen-
sory expectations with experience, then over the course
of training, participants should learn to expect a delayed
outcome after their actions, resulting in more accurate
predictions on the timing of sensory feedback. This would
be reflected in reduced N1 amplitude over the course of
formazione. In contrasto, if the expectation that “sensations
immediately follow actions” was fixed and unmalleable,
then delayed tones should remain poorly predicted even
after extensive training, and correspondingly, N1 amplitude
would be expected to remain relatively constant over
formazione. The immediate training condition provides a
necessary control for the possibility that N1 amplitude
changes purely as a function of repeated exposure to a
tone (Todorovic & de Lange, 2012).
METHODS
Participants
Fifty University of New South Wales Sydney students
participated for course credit. Five participants were
excluded because of equipment failures (n = 2) and low
signal-to-noise ratios (n = 3). The remaining 45 partici-
pants had a mean age of 19.4 years (SD = 3.2 years, range =
17–28 years), con 24 women. There were 37 right-handed,
3 ambidextrous, E 5 left-handed individuals, as mea-
sured by the Edinburgh Handedness Inventory (Ransil &
Schachter, 1994). This study was approved by the Univer-
sity of New South Wales Sydney Human Research Ethics
Advisory Panel (Psychology).
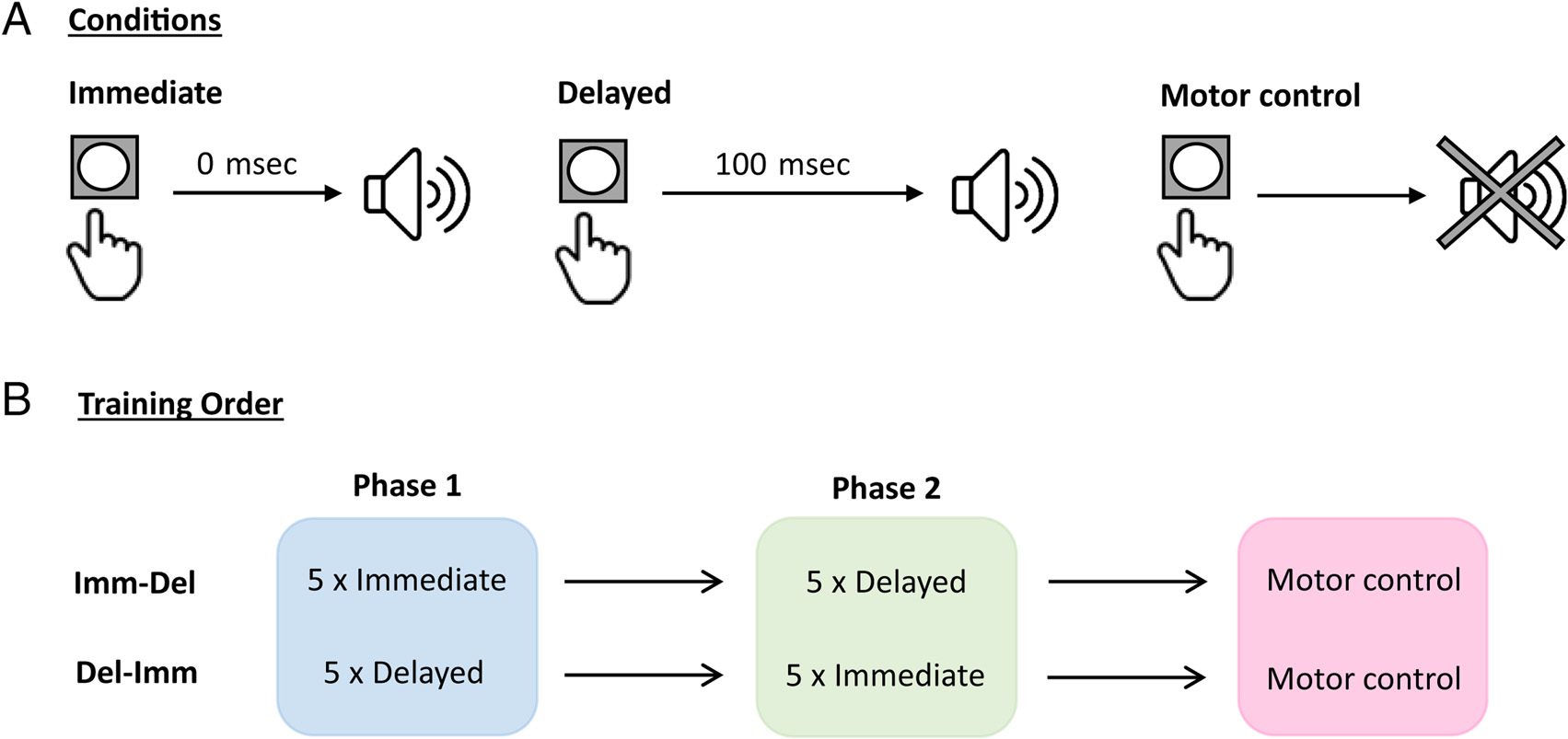
Stimuli and Procedure
The experiment was composed of a button-press-for-tone
compito (Figure 1A). Participants were required to press a
button on a response pad (Model RB-530; Cedrus Corpo-
ration) with their dominant hand at a time of their choos-
ing so long as it was subsequent to the presentation of a
fixation cross on the monitor. The purpose of the fixation
cross was to standardize the block length and to limit the
occurrence of eyeblinks that were coincident with the tone
(see below); it was emphasized to participants that the task
was not speeded and that they should not press the button
as soon as the fixation cross appeared. Pressing the button
produced a pure tone (500 Hz, 100-msec duration, 5-msec
rise-and-fall time), which occurred 17 msec after the button
press (immediate tone) O 117 msec after the button press
(delayed tone). The tone played binaurally over headphones
(Sennheiser HD 201) at approximately 70 dB SPL. The fixation
cross disappeared from the screen 1500 msec after tone
offset, signaling the end of the trial. The next trial began
after a random interval of between 1000 E 3000 msec.
Each of the first 10 blocks of the experiment consisted of
60 button-press-for-tone trials. Each block was homoge-
nous in that it contained only immediate or delayed tones.
Each block took approximately 6 min to complete.
Figure 1B shows the experimental design. Participants
were initially given verbal instructions on how to perform
the task and were encouraged to time their eyeblinks to
when the fixation cross was not on screen. They then
completed five practice trials of pressing for an immedi-
ate tone. After this, participants completed 10 formazione
blocks, consisting of five blocks with immediate tones
(immediate training condition) and five blocks with de-
layed tones (delayed training condition). Participants who
were randomly assigned to the Imm-Del group (n = 24)
completed five immediate tone blocks followed by five de-
layed tone blocks; participants in the Del-Imm group (n =
21) completed five delayed tone blocks followed by five
immediate tone blocks. Participants took brief, self-paced
breaks between blocks, and reminder instructions were
displayed on-screen before beginning each block (questi
instructions were identical for immediate and delayed
tone blocks and made no reference to any delay between
button press and tone). After training, all participants
underwent one block (60 trials) of a motor control con-
dizione. The motor control condition was identical to the
immediate and delayed blocks, except that pressing the
button did not result in a tone being played. The instruc-
tions and fixation cross were presented on a 24-in. BenQ
Elijah, Le Pelley, and Whitford
1147
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
J
o
C
N
_
UN
_
0
1
2
6
9
P
D
.
F
B
e
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
l
D
o
w
N
o
UN
D
e
D
F
R
o
M
H
T
T
P
:
/
/
D
io
R
e
C
T
.
M
io
T
.
e
D
tu
/
J
/
o
C
N
UN
R
T
io
C
e
–
P
D
l
F
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
J
o
C
N
_
UN
_
0
1
2
6
9
P
D
.
F
B
e
G
tu
e
S
T
T
o
N
0
8
S
e
P
e
M
B
e
R
2
0
2
3
Figura 1. (UN) A schematic of the three different trial types in the experiment. In all trial types, participants pressed a button. Pressing the button
resulted in a tone either 17 msec after the button press (immediate condition), 117 msec after the button press (delayed condition), or no tone
(motor control condition). (B) Training order across the experiment. Participants were trained to immediate tones first and then delayed tones
(Imm-Del group) or delayed tones first and then immediate tones (Del-Imm group).
XL2420T monitor (1920 × 1080 resolution, 120-Hz refresh
rate). The background color of the screen was middle
gray throughout the experiment; the text instructions
and the fixation cross were white. All stimuli were pre-
sented using MATLAB R2012b with Psychophysics Toolbox
extensions (Kleiner, Brainard, & Pelli, 2007; Brainard, 1997;
Pelli, 1997).
EEG Acquisition and Processing
EEG data were collected using a BioSemi ActiView system
with a 2048-Hz sample rate, a 417-Hz bandwidth (3 dB),
and a 24-dB/octave roll-off (Biosemi, 2012). The Ag/Ag-Cl
electrodes were connected to all 64 cap channels, con
additional electrodes attached to the mastoids and nose
as well as electrodes placed 1 cm from the outer canthi of
both eyes and 1 cm under the left eye to monitor hori-
zontal and vertical eye movements. Online referencing
was to sensors located in the parietal region of the cap
(CMS and DRL). The DC offsets were kept below 25 mV.
EEG data were processed using BrainVision Analyser
(v.2.1). The data were rereferenced offline to the average
of linked mastoids and were then passed through a 0.5-
to 30-Hz Butterworth bandpass filter. For all tone condi-
zioni, in each trial, an 800-msec epoch was created,
which covered 200 msec before tone onset to 600 msec
after tone onset. The same was done for the motor
condition, except that the epoch was created around
the button press. Eyeblink artifacts were corrected using
the method of Gratton, Coles, and Donchin (1983), E
epochs were rejected if the peaks were ±100 μV or had a
maximum gradient of >50 μV. The remaining epochs
were then averaged to create a waveform for each block.
A fast Fourier independent component analysis correc-
tion was applied to headphone-induced artifacts, Quale
manifest as a sharp positive deflection at approximately
15 msec after tone onset. The average waveform of the
motor control block was subtracted from the immediate
tone blocks to produce a motor-corrected waveform to
the tone alone, consistent with previous sensory suppres-
sion literature (Mifsud, Beesley, Watson, & Whitford,
2016; Ford, Palzes, Roach, & Mathalon, 2014; SanMiguel
et al., 2013; Bäss et al., 2011). The same procedure was
applied to the data from delayed blocks, except that the
motor control waveform was time-shifted 100 msec be-
fore applying the motor correction, to match the timing
of the onset of the button press, as has been done previ-
ously (Elijah, Le Pelley, & Whitford, 2016). The prestimu-
lus phase was baseline corrected in the interval of −200
A 0 msec.
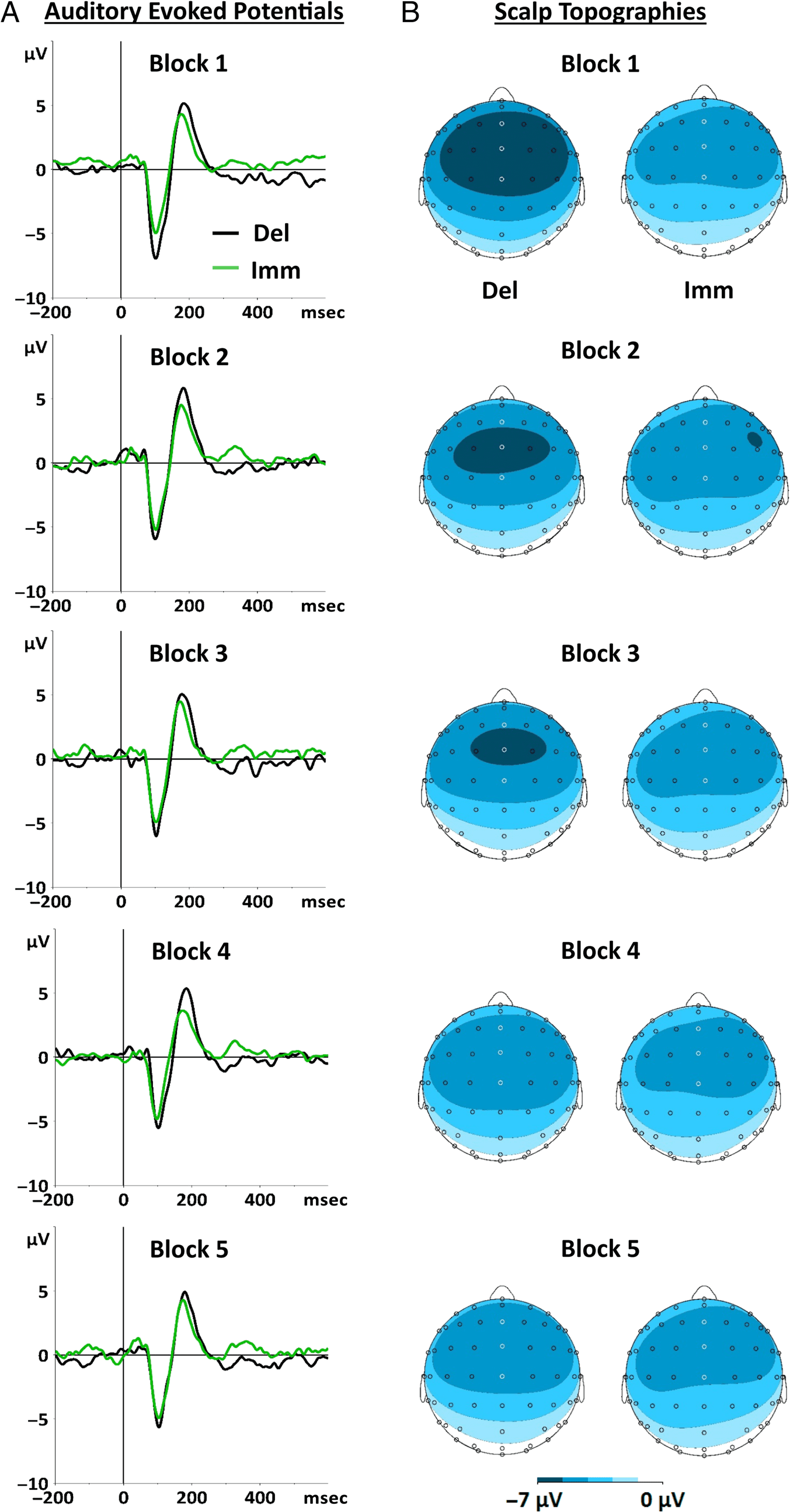
Scalp topographies (Figure 4B) indicated that the N1
component was maximal at frontocentral sites, che è
consistent with previous auditory suppression research
( Whitford et al., 2017; Ford et al., 2014; Saupe, Widmann,
Trujillo-Barreto, & Schröger, 2013). We analyzed data
from an array of electrodes at which activation was
maximal (Fz, FCz, and Cz), as is common practice in audi-
tory ERP studies (Bednark, Poonian, Palghat, McFadyen,
& Cunnington, 2015; Ford et al., 2014; Hughes et al.,
2013B; Saupe et al., 2013; Luck, 2005). The amplitude of
the auditory N1 was defined for each participant (for each
condition) as the mean activity in a 20-msec time window
centered around the most negative point on each par-
ticipant’s average waveform, in the time window of 50–
150 msec. Allo stesso modo, scalp topographies were generated
by extracting activity over a 20-msec time window around
1148
Journal of Cognitive Neuroscience
Volume 30, Numero 8
the most negative point between 50 E 150 msec on
grand-averaged waveforms for each condition. The time
windows for delayed tones for Blocks 1–5 averaged across
all participants were 92–112, 90–110, 93–113, 92–112, E
94–114 msec, rispettivamente. The time windows for imme-
diate tones for Blocks 1–5 were 92–112, 92–112, 92–112,
88–108, and 94–114 msec, rispettivamente. In all figures
plotting N1 amplitude, the within-participant SEM was
used to represent variability (Cousineau, 2005).
RESULTS
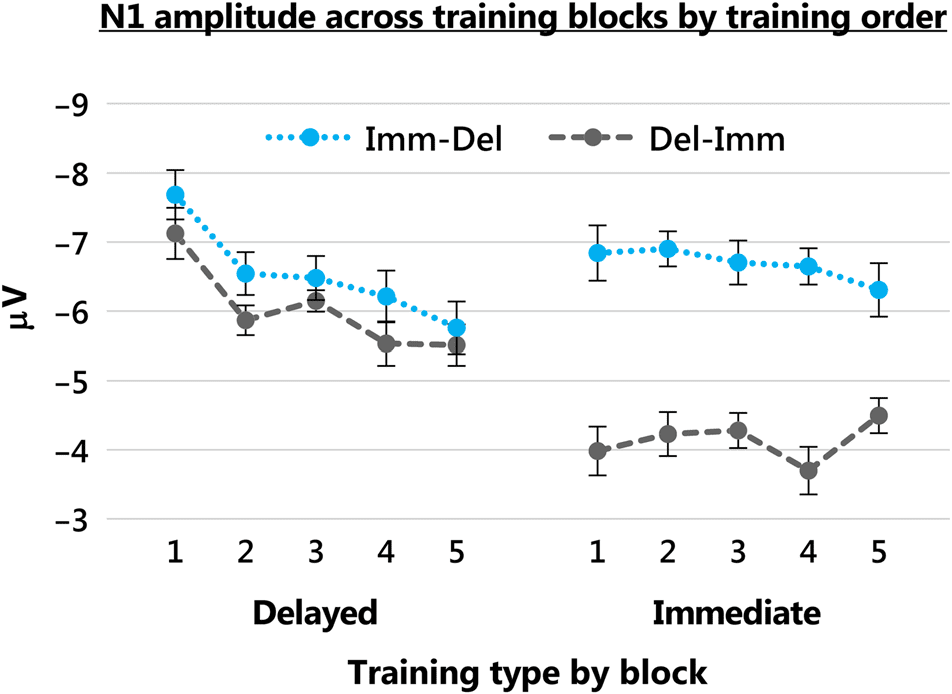
Figura 2 displays the mean N1 amplitude elicited by im-
mediate and delayed tones across the five blocks of train-
ing, separated by training order. Mean N1 amplitude was
analyzed using a 2 × 2 × 5 mixed ANOVA with factors
of Training order (Imm-Del or Del-Imm), Training type
(delayed or immediate training), and Block (Training
Blocks 1–5). Given that we were interested in the change
in N1 across multiple blocks, this analysis used orthogonal
linear trend contrasts.
There was a significant main effect of Training type, F(1,
43) = 4.160, p = .048, ηp
2 = .088, and Block, F(1, 43) =
9.084, p = .004, ηp
2 = .174. Critically, the Training type ×
Block interaction was significant, F(1, 43) = 7.683, p =
.008, ηp
2 = .152, indicating that the influence of training
on N1 amplitude depended on the delay experienced
between button press and tone. Figura 2 shows that N1
amplitude tended to decrease across the course of
training with delayed tones but remained relatively stable
across training with immediate tones. In particolare, this pattern
of change across blocks did not depend on the order
Figura 2. N1 amplitude (microvolts) across blocks of training (1–5) per
participants in the Imm-Del group (who were first trained with the
immediate tone, followed by training with the delayed tone) and the
Del-Imm group (who were first trained with the delayed tone, followed
by training with the immediate tone). Performance in blocks with the
delayed tone is shown on the left-hand side of the figure; performance
in blocks with the immediate tone is shown on the right. Error bars
show within-participant SEM.
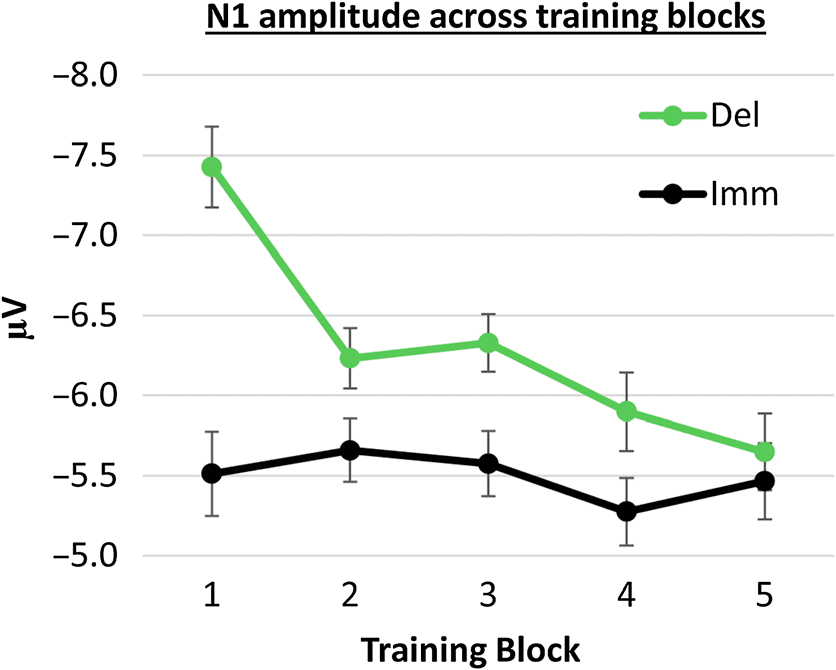
Figura 3. N1 amplitude (microvolts) across blocks of training with
delayed tones (black line) and immediate tones (green line) collapsed
across training order. Error bars show within-participant SEM.
of training, as indicated by a nonsignificant three-way
interaction (Training order × Training type × Block: F(1,
43) = 0.24, p = .63, ηp
2 = .005). That said, the order of
training did have an impact on N1 amplitude: There was
a significant main effect of Training order, F(1, 43) =
4.317, p = .044, ηp
2 = .091, that was qualified by a Training
order × Training type interaction, F(1, 43) = 5.644, p =
.022, ηp
2 = .116. Further exploration of this interaction with
simple effects revealed that, collapsing across training
blocks, whereas N1 amplitude to delayed tones did not differ
significantly as a function of Training order, F(1, 43) = 0.261,
p = .612, ηp
2 = .006, N1 amplitude to immediate tones
was significantly greater in the Imm-Del group than in the
Del-Imm group, F(1, 43) = 13.180, p = .001, ηp
2 = .235
(Guarda la figura 2). The Training order × Block interaction was
not significant, F(1, 43) = 0.731, p = .397, ηp
2 = .017.
Because the three-way interaction was not significant, we
collapsed across the two training order groups (Del-Imm
and Imm-Del) in a follow-up analysis, which was aimed at
unpacking the Training type × Block interaction. Figura 3
shows the resulting mean N1 amplitudes for immediate and
delayed tones across training blocks, and Figure 4 shows
the corresponding ERPs (UN) and scalp topographies (B).
Consistent with the pattern observed in Figure 3, trend
analysis revealed a significant linear decrease in N1 ampli-
tude across delayed training blocks, F(1, 44) = 14.293,
P < .001, ηp
2 = .245, but no significant linear trend in N1
amplitude across immediate training blocks, F(1, 44) =
0.324, p = .572, ηp
2 = .007. Furthermore, a targeted
comparison of data at the first block of training between
each tone type revealed a significantly higher N1 ampli-
tude for delayed tones compared with immediate tones,
F(1, 43) = 11.182, p = .002, ηp
2 = .203. There was no sig-
nificant difference between the two tone types at the final
block of training, F(1, 43) = 0.108, p = .744, ηp
2 = .002.
To further investigate the change in sensory modula-
tion when switching from one timing expectation to
Elijah, Le Pelley, and Whitford
1149
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
j
o
c
n
_
a
_
0
1
2
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. (A) The ERPs for
delayed (black) and immediate
(green) tones for each training
block. The 0-msec point on the
x axis represents tone onset,
and the negative deflection
at approximately 100 msec
post-tone-onset is the N1
component. (B) Scalp
topographies for the N1
component for delayed and
immediate tones for each
training block. The time
windows for Blocks 1–5 for
delayed tones were 92–112,
90–110, 93–113, 92–112, and
94–114 msec, respectively,
and for immediate tones were
92–112, 92–112, 92–112,
88–108, and 94–114 msec,
respectively.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
j
o
c
n
_
a
_
0
1
2
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
1150
Journal of Cognitive Neuroscience
Volume 30, Number 8
et al., 2009; Bäss et al., 2008). Consistent with the litera-
ture, our results showed that, at the beginning of the rel-
evant training phase, participants’ N1 amplitudes were
larger to delayed tones compared with immediate tones
(see Figure 3), suggesting that delayed tones were un-
predicted relative to immediate tones. This finding adds
to the growing body of literature that suggests that, by
default, we have a prior expectation for immediate sensory
feedback after our own willed actions (Friston, 2005; Niemi
& Näätänen, 1981). The primary purpose of the current
study was to determine whether this expectation that
“sensations follow immediately from actions” is amenable
to modification with training or whether it is fixed and un-
changeable with experience. We demonstrated that, with
repeated exposure to a delayed action–sensation contin-
gency (i.e., a willed button press for a tone presented
100 msec later), there was a significant decrease in the
N1 amplitude elicited by the tone over the course of train-
ing. This suggests that participants were able to learn to
anticipate delayed feedback from their actions, which was
reflected in the extent to which the cortical response to
these tones was attenuated with training. In other words,
we suggest that there was an update of the sensory expec-
tation as to “when” auditory sensations were predicted to
result from a willed action.
Notably, the reduction in N1 amplitude over the
course of delayed training cannot simply be a conse-
quence of habituation due to repeated exposure to the
tone (Todorovic & de Lange, 2012). If habituation were
the sole source of any change in N1 amplitude, then we
would expect a similar decrease in N1 amplitude over the
course of training to both immediate and delayed tones.
However, we found a significant interaction between the
training tone (immediate vs. delayed) and exposure
(Training Blocks 1–5), whereby the reduction in N1 am-
plitude with repeated exposure to the tone was more
marked in the delayed training condition compared with
the immediate training condition, which showed no sig-
nificant reduction in N1 amplitude over training blocks
(shown most clearly in Figure 3). In other words, the re-
duction in N1 amplitude in the delayed training condition
was apparent over and above any change that occurred in
the immediate training condition. This lack of significant
change with immediate training is critical, as it suggests
that repetition per se was not the determinant of change;
rather, we can conclude that participants’ temporal
expectations were modified with the experience of de-
layed feedback. Further evidence for the importance of
the action–sensation duration came from a secondary
analysis whereby the change in N1 amplitude with a switch
in training depended critically on the type of training be-
fore and after the switch (see Figure 5). After repeated
experience with a particular type of tone (delayed or im-
mediate), there was no difference in N1 amplitude be-
tween delayed and immediate tones; however, after a
switch in the experienced tone type, a difference in N1 am-
plitude between delayed and immediate tones emerged.
Elijah, Le Pelley, and Whitford
1151
Figure 5. N1 amplitude (microvolts) across blocks after a switch in
training type. This figure compares N1 amplitude in the last block of
training in Phase 1 with N1 amplitude in the first block of training in
Phase 2. The Del-Imm line (dashed gray line) represents participants
who were trained to the delayed tone first followed by the immediate
tone. The Imm-Del line (dotted blue line) represents participants
who were trained to the immediate tone first followed by the delayed
tone. Error bars show within-participant SEM.
another, a secondary analysis was run that focused on the
period of transition between training types. Figure 5
shows N1 amplitude to the tone in the fifth training block
(the final block of Phase 1 of the task, before the delay
type switched) and in the sixth training block (the first
block of Phase 2, after the delay type switched). These
data were analyzed using a 2 × 2 mixed ANOVA with fac-
tors of Block (the final block of Phase 1 vs. the first block
of Phase 2) and Training order (Del-Imm vs. Imm-Del).
This revealed a significant main effect of Training order,
F(1, 43) = 7.911, p = .007, ηp
2 = .155, with a larger N1
amplitude in the Del-Imm condition. There was no sig-
nificant main effect of Block, F(1, 43) = 0.018, p =
.894, ηp
2 < .001. Importantly, there was a significant Train-
ing order × Block interaction, F(1, 43) = 6.472, p = .015,
ηp
2 = .131. Analysis of simple effects found no significant
difference in N1 amplitude between delayed and imme-
diate tones in the final block of Phase 1, F(1, 43) = 0.537,
p = .468, ηp
2 = .012, but a significantly smaller N1 ampli-
tude for immediate tones compared with delayed tones
in the first block of Phase 2, F(1, 43) = 18.298, p < .001,
ηp
2 = .809. Orthogonal comparisons revealed that the
change in N1 amplitude from Phase 1 to Phase 2 fell short
of significance for both the Del-Imm group, F(1, 20) =
3.87, p = .063, ηp
2 = .162, and the Imm-Del group, F(1,
23) = 2.793, p = .108, ηp
2 = .108.
DISCUSSION
There is substantial evidence that auditory stimuli that
follow immediately from self-initiated actions normatively
evoke smaller N1 amplitudes compared with physically
identical stimuli that occur after a delay (Elijah et al.,
2016; Oestreich et al., 2016; Whitford et al., 2011; Aliu
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
j
o
c
n
_
a
_
0
1
2
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
This result suggests that our recent sensory experience
(in this case, for delayed or immediate feedback after
the button press) modulates subsequent sensory atten-
uation. Taken together, these results provide strong evi-
dence that sensory expectations regarding the timing of
self-generated sensations are, at least to some extent,
malleable given new temporal information.
The current findings add to previous research that has
investigated temporal adaptation to delayed, self-initiated
sensations (Cao, Veniero, Thut, & Gross, 2017; Elijah et al.,
2016; Aliu et al., 2009). Aliu et al. (2009) investigated
auditory suppression changes with repeated exposure to
delayed sensory feedback using MEG. Participants under-
went four blocks of pressing a button for a 100-msec
delayed tone, and suppression to these tones was calcu-
lated as the difference between the cortical response to
passively presented tones and delayed, self-initiated tones.
Aliu et al. found an increase in cortical suppression of the
m100 component (the MEG equivalent of the N1 compo-
nent in EEG) across repeated exposure to the delayed
tone. Cao et al. (2017) showed the same training effect
on the m100 component with delayed action effects and
showed that this temporal adaptation was modulated by
the cerebellum. These existing studies are consistent with
the idea that expectations for delayed sensory feedback
from an action can be updated.
An important methodological issue in the sensory
attenuation literature relates to the validity of comparing
“active” and “passive” conditions. Most previous studies
in the field have focused on comparing an “active” con-
dition, in which tones are generated by a self-initiated
motor action, with a “passive” condition, in which tones
are externally generated without any action. This active-
versus-passive comparison is problematic, as it intro-
duces potential confounds that may render findings
equivocal (Horváth, 2015; Hughes et al., 2013a). For ex-
ample, attention may differ between conditions: In a pas-
sive condition, participants’ attention is solely focused on
the tone, whereas in an active condition, attention may
be divided between the action and the tone. In light of
previous research indicating that attention can influence
N1 amplitude, whereby the more attention given to a
stimulus, the larger the N1 it elicits, (Saupe et al., 2013;
Timm, SanMiguel, Saupe, & Schröger, 2013), it is pos-
sible that studies employing active-versus-passive com-
parisons may be tapping changes in attention rather
than changes in temporal expectations. Furthermore,
the fact that an active condition involves motor-related
activity (from the button press) whereas the passive con-
dition does not is also potentially problematic. Although
movement-related activity is typically corrected during
EEG processing (by measuring the motor-evoked poten-
tial generated in the motor-only condition and subtract-
ing this from the active condition), this correction is
suboptimal as it makes the (potentially erroneous) as-
sumption that the auditory- and motor-evoked activity
is additive in nature (Horváth, 2015). As such, any ob-
served differences in N1 amplitude between active and
passive conditions could potentially be due to differences
in motor-related activity (or the correction of such activ-
ity), rather than differences in motor–auditory prediction
per se. In contrast, the current study relied on a compar-
ison between two active conditions (in which participants
pressed a button for either immediate or delayed tones).
In this design, factors such as attention and motor activity
are more closely matched for the critical comparison, and
thus we can be more confident that the observed differ-
ence between the two conditions reflects a difference in
temporal expectations.
The current findings are also consistent with previous
work from our own laboratory in which we investigated
people’s ability to modify temporal expectations with de-
layed training (Elijah et al., 2016). Like the current study,
this previous research was based on a comparison be-
tween two active conditions. However, Elijah et al. (2016)
used a fully between-participant design: One group of
participants received training in which button presses pro-
duced a delayed tone, whereas another group was trained
with immediate tones. Pretraining and posttraining mea-
sures of N1 amplitude elicited by delayed and immediate
tones were used to assess the effectiveness of training in
each group. We showed that training with delayed tones
led to an elimination of the baseline difference in N1 am-
plitude between immediate and delayed tones. Training
with immediate tones, in contrast, resulted in these base-
line differences being retained, posttraining. However, as a
product of its between-participant design, differences in
the type of training received (i.e., immediate or delayed
tones) were confounded with the order in which the criti-
cal test blocks were administered. This precluded a direct
statistical comparison of the two groups, because it meant
that different types of test trials were confounded with
participants’ level of experience with the task. This limited
our ability to directly compare the effects of immediate
versus delayed training. In contrast, the within-participant
design of the current study allowed us to directly compare
the effects of training with immediate or delayed tones,
because all participants completed both training conditions
(in counterbalanced order). As such, this study is, to our
knowledge, the first to directly compare the differential
effects of training temporally expected (immediate) and
unexpected (delayed) tones on sensory attenuation. This
direct comparison demonstrates that attenuation of N1
amplitude is indeed dependent on temporal experience,
rather than merely reflecting habituation, or attentional
and/or motor-related differences between conditions.
More precisely, we suggest that the specific reduction in
N1 amplitude that resulted from training with delayed
tones, but not immediate tones, reflected the updating of
temporal expectations, which enabled the anticipation of
delayed feedback from self-initiated actions.
A further advantage of the current within-participant
design over previous designs is that it allowed us to exam-
ine more closely the influence of switching participants’
1152
Journal of Cognitive Neuroscience
Volume 30, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
j
o
c
n
_
a
_
0
1
2
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
temporal experience—from immediate action effects to
delayed action effects and vice versa—on their temporal
expectations. At a theoretical level, it could be the case
that experience of action–sensation delays results in the
change of the expectation that “sensations follow immedi-
ately from actions” to an expectation of “sensations follow
from actions after a specific delay” (i.e., there is no longer
a default expectation for immediate feedback). Another
possibility is that the experience of action–sensation
delays leads to the formation of a new expectation (that
“delayed sensations result from actions in the current con-
text”), and this new expectation comes to dominate over
the existing, default “immediate” expectation (that re-
mains intact and unchanged) based on contextual infor-
mation (i.e., recent experience of delayed feedback). In
either of these cases, we would expect to see an increase
in N1 amplitude to immediate feedback (indicating a vio-
lation of expectations) after repeated experience of de-
layed feedback. However, this was not the pattern we
observed, as illustrated in Figure 5. On the contrary, in an
analysis targeted at the period of transition between the
two training types, we found evidence that a switch in
training type from delayed to immediate tones did not
result in a larger N1 amplitude for immediate tones. Indeed,
when analyzing over the whole training period, we found
that N1 amplitude elicited by immediate tones was sig-
nificantly smaller when participants had received previous
training with delayed tones, compared with when they
had not, as shown in Figure 2.
These findings are inconsistent with an account under
which the effect of training with delays is simply to shift
the mean expectation of action–sensation delays, such
that immediate tones are unexpected after prior training
with delayed tones. At a general level, our data suggest
that prior experience of delays results in greater sensory
attenuation of both delayed and immediate tones in the
future. One possibility is that the effect of training with
delays might widen the range of delays that is expected,
rather than shifting the mean expectation (see also van
Laarhoven et al., 2017; Shadmehr, Smith, & Krakauer,
2010). More specifically, our results are consistent with
an account in which experience with delayed feedback
triggers a recalibration of action–sensation expectations,
which nevertheless retains a privileged status for imme-
diate sensory feedback. This account can be described as
follows. All participants begin the experiment with the
default expectation that “sensations follow immediately
from actions,” and immediate sensations will be subject
to greater sensory attenuation. For the Imm-Del group,
the first training phase involves immediate tones, and
there is no violation of the default expectation and thus
no trigger to recalibrate it. When this group then encoun-
ters delayed tones in the second training phase, the default
expectation is violated, and recalibration is triggered—
resulting in increased sensory attenuation of delayed
tones as training proceeds, such as we observed empiri-
cally. In contrast, participants in the Del-Imm group expe-
rience expectancy-violating delayed tones during the first
training phase. This immediately triggers recalibration,
thus increasing sensory attenuation of delayed tones as
training proceeds (resulting again in a steadily decreasing
N1 amplitude over training with delayed tones). Critically,
our data suggest that the perceptual system may accom-
plish this via a “global” increase in suppression over the
range of the delay: Additional suppression is applied over
the whole range from 0 to 100 msec (as illustrated in the
Figure 6 schematic). This has the effect of achieving atten-
uation of the now-predictable delayed tones, resulting
again in a steadily decreasing N1 amplitude over training
with delayed tones. However, as a side effect, when im-
mediate tones are subsequently presented in the second
training phase for the Del-Imm group, they are now sub-
ject to both the attenuation resulting from the default
expectation plus the additional suppression that occurred
as a result of the recalibration process. Hence, for these
participants, N1 response to immediate tones will be
especially small, as we observed empirically.
The idea that we still expect immediate sensations
from our actions despite experience with delayed
feedback raises the question of whether expectations
for immediate feedback are “hardwired” or occur be-
cause of a lifetime of experience. For example, as noted
earlier, previous research suggests that raising chicks in
an environment where light shines from below does
not seem to influence chicks’ prior belief that “light
shines from above,” suggesting that this prior is innate
and resistant to modification (Hershberger, 1970; Hess,
1950). It could be the case that the same rigidity is appar-
ent in our belief that sensations follow immediately from
actions. That said, it should be noted that the delay
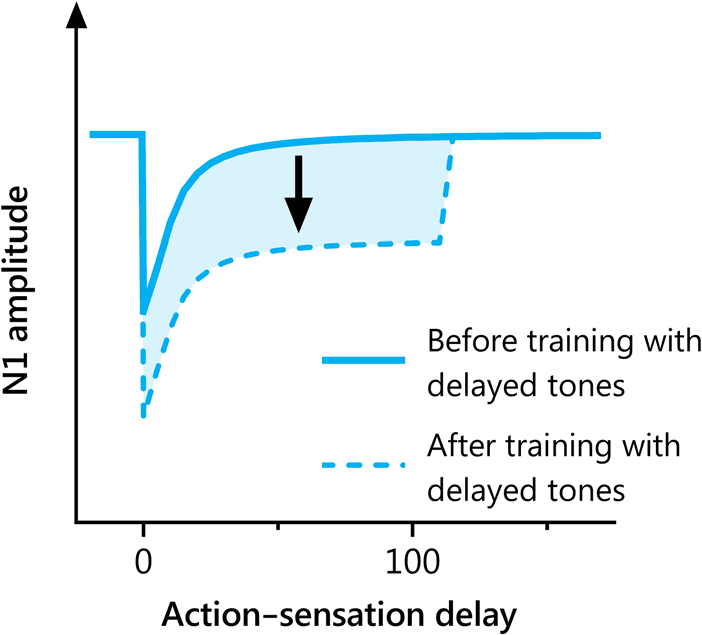
Figure 6. Schematic illustration of the effect of delayed training on N1
amplitude elicited by tones across a range of delays. Before training
with 100-msec delayed tones, immediate tones (0 msec) elicit a smaller
N1 amplitude than delayed tones (100 msec) because of the default
expectation for immediate sensory feedback. The shaded region shows
the additional sensory attenuation that is triggered by training with a
delayed action–sensation relationship. This training has the effect of
reducing N1 amplitude for both delayed and immediate tones.
Elijah, Le Pelley, and Whitford
1153
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
j
o
c
n
_
a
_
0
1
2
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
manipulation in our task altered only a subset of the
participants’ experiences. Specifically, although we intro-
duced a delay between button press and tone, actions pe-
ripheral to this task would have still produced immediate
sensory feedback. For example, if the participants shifted
in their seat, this would create immediate auditory and
somatosensory feedback; likewise, head movements
would create immediate visual feedback. In this sense,
the expectation that “sensations follow immediately from
actions” would continue to receive partial confirmation
even during the delayed training condition. Hence, it
seems plausible that, under these conditions, participants
could retain a privileged status for immediate auditory
feedback, while simultaneously adjusting their expecta-
tions for delayed auditory feedback (as illustrated in Fig-
ure 6). This implies that our procedure, if anything,
probably underestimates the ability of the sensorimotor
system to adapt to “pure” changes in experienced action–
sensation delays (i.e., training in which there are no “dis-
confirmatory” trials). Finally, it raises the question of
whether the adaptation of sensory attenuation that is
produced by training is specific to the action–sensation
relationship learned in the training context (e.g., a button
press results in a 500-Hz tone in the psychophysiology
laboratory) or whether the effects of training will gener-
alize to other stimuli or contexts and possibly even other
sensory modalities. This remains a question for future
research.
In the discussion above, we have offered a simplistic
account of the mechanisms that might underlie adapt-
ability of temporal expectations. In reality, the situation
is undoubtedly more nuanced; for example, the neural
processing engaged by tasks such as this would be much
more complex than simply categorizing temporal infor-
mation as “delayed” or “immediate” based solely on past
experience and using this information as a basis for sen-
sory prediction. Instead, noise in the system, and in the
world more broadly, would mean that people would ex-
perience a range of delays over a narrow window, based
on what they have experienced in the past (Shadmehr
et al., 2010; Wolpert & Miall, 1996; Wolpert et al.,
1995). For example, during development, the conduction
delays associated with nerve impulses initiated by periph-
eral tactile stimulation would change with limb growth
(Campbell, Ward, & Swift, 1981). To account for such
variation in the system, it is likely that the normative level
of expected delay falls within a certain range. As such, it
may only be when a set of experiences consistently fall
outside this range (e.g., 100-msec delayed auditory feed-
back in the case of the current experiment) that a re-
calibration process is triggered. Nevertheless, predictive
coding accounts suggest that priors are based on proba-
bility estimates of the likelihood of outcomes based on
the current state and the context, not only on past expe-
rience (Bogacz, 2017; Brown et al., 2013; Friston, 2005;
Rao & Ballard, 1999). Consequently, recalibration of the
expectation for delayed feedback may be very specific to
the action–effect association that is learnt and may not
generalize to experiences outside this experimental con-
text. As such, it may be the case that, for recalibration to
trigger an expansion of the normative temporal window
more permanently, the physical system must change
(i.e., limb growth, diseases processes) so that most feed-
back from actions consistently occur within a new time
frame. Otherwise, recalibration may be specific to the
context in which the temporal association between the
action and the event is learnt.
The current finding that sensory attenuation is, to
some extent, modifiable with experience may provide a
potential avenue for treating clinical populations who
show dysfunctions in sensory attenuation, such as pa-
tients with schizophrenia. Research has consistently
shown that patients with schizophrenia show subnormal
differences in N1 amplitude between self-generated and
externally generated sounds (Ford et al., 2014; Whitford
et al., 2011; Ford & Mathalon, 2004). This deficit in sen-
sory suppression provides a potential explanation for
some of the most characteristic symptoms of schizophre-
nia, such as delusions of control whereby misplaced
salience attributed to willed actions leads to misinter-
pretation of these self-initiated movements as being con-
trolled by an external force (Fletcher & Frith, 2009; Frith,
2005; Feinberg & Guazzelli, 1999; Feinberg, 1978). There
is further evidence to suggest that this disruption in sen-
sory attenuation may be due to deficits in predicting the
timing of self-generated sensations (Oestreich et al.,
2016; Whitford et al., 2011), possibly due to structural
damage to white matter (Whitford et al., 2011). If tempo-
ral expectations are modifiable with training—as sug-
gested by the results of this study—retraining patients’
expectations regarding the timing of self-generated sensa-
tions may be a viable avenue of treatment for normalizing
their sensory attenuation deficits. Furthermore, research
suggests that neurocognitive dysfunctions in schizophrenia
can be remediated with cognitive and behavioral training
(Dale et al., 2016; Penadés et al., 2013; Subramaniam
et al., 2012; Vinogradov, Fisher, & de Villers-Sidani, 2012).
If the characteristic symptoms of schizophrenia indeed re-
flect sensory attenuation abnormalities, then normalizing
these abnormalities by means of behavioral training might
be expected to be clinically therapeutic and thus worthy of
investigation in future studies (Whitford, Ford, Mathalon,
Kubicki, & Shenton, 2012).
In conclusion, this study provides evidence that the
sensory expectation that “sensations follow immediately
from actions” can be modified with experience. However,
this flexibility is conditional; the current data suggest that,
although it is possible to learn to expect delayed feedback
from our actions with training, we concurrently continue to
expect immediate feedback, despite this training. These
findings are consistent with the idea that the experience
of temporally delayed sensations after actions results in a
broadening of the temporal window over which sensations
are expected to occur after a self-generated action.
1154
Journal of Cognitive Neuroscience
Volume 30, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
j
o
c
n
_
a
_
0
1
2
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Acknowledgments
We thank the two anonymous reviewers for their helpful and
insightful comments. Ruth B. Elijah is supported by an Australian
Postgraduate Award. The study was funded in part by Discovery
Projects from the Australian Research Council (DP140104394
and DP170103094) awarded to Thomas J. Whitford and Mike.
E. Le Pelley. Thomas J. Whitford is supported by an NHMRC
Career Development Fellowship (APP1090507).
Reprint requests should be sent to Ruth B. Elijah, School of
Psychology, The University of New South Wales, Sydney, NSW
2052, Australia, or via e-mail: ruth.elijah@unsw.edu.au.
REFERENCES
Aliu, S. O., Houde, J. F., & Nagarajan, S. S. (2009). Motor-induced
suppression of the auditory cortex. Journal of Cognitive
Neuroscience, 21, 791–802.
Arnal, L. H., & Giraud, A.-L. (2012). Cortical oscillations and
sensory predictions. Trends in Cognitive Sciences, 16, 390–398.
Bäss, P., Horváth, J., Jacobsen, T., & Schröger, E. (2011).
Selective suppression of self-initiated sounds in an auditory
stream: An ERP study. Psychophysiology, 48, 1276–1283.
Bäss, P., Jacobsen, T., & Schröger, E. (2008). Suppression of
the auditory N1 event-related potential component with
unpredictable self-initiated tones: Evidence for internal
forward models with dynamic stimulation. International
Journal of Psychophysiology, 70, 137–143.
Bays, P. M., Flanagan, J. R., & Wolpert, D. M. (2006). Attenuation
of self-generated tactile sensations is predictive, not
postdictive. PLoS Biology, 4, e28.
Bays, P. M., Wolpert, D. M., & Flanagan, J. R. (2005). Perception
of the consequences of self-action is temporally tuned and
event driven. Current Biology, 15, 1125–1128.
Bednark, J. G., Poonian, S. K., Palghat, K., McFadyen, J., &
Cunnington, R. (2015). Identity-specific predictions and
implicit measures of agency. Psychology of Consciousness:
Theory, Research, and Practice, 2, 253–268.
Behroozmand, R., Liu, H., & Larson, C. R. (2011). Time-dependent
neural processing of auditory feedback during voice pitch error
detection. Journal of Cognitive Neuroscience, 23, 1205–1217.
Bendixen, A., SanMiguel, I., & Schröger, E. (2012). Early
electrophysiological indicators for predictive processing in
audition: A review. International Journal of
Psychophysiology, 83, 120–131.
Biosemi. (2012). ActiView701 [computer software]. Retrieved
from http://www.biosemi.com/download_actiview.htm/.
Blakemore, S.-J., Frith, C. D., & Wolpert, D. M. (1999). Spatio-
temporal prediction modulates the perception of self-produced
stimuli. Journal of Cognitive Neuroscience, 11, 551–559.
Blakemore, S.-J., Wolpert, D. M., & Frith, C. D. (1998). Central
cancellation of self-produced tickle sensation. Nature
Neuroscience, 1, 635–640.
Bogacz, R. (2017). A tutorial on the free-energy framework for
modelling perception and learning. Journal of Mathematical
Psychology, 76, 198–211.
Brainard, D. H. (1997). The Psychophysics Toolbox. Spatial
Vision, 10, 433–436.
Brown, H., Adams, R. A., Parees, I., Edwards, M., & Friston, K.
(2013). Active inference, sensory attenuation and illusions.
Cognitive Processing, 14, 411–427.
Campbell, W. W., Ward, L. C., & Swift, T. R. (1981). Nerve
conduction velocity varies inversely with height. Muscle &
Nerve, 4, 520–523.
Cao, L., Veniero, D., Thut, G., & Gross, J. (2017). Role of the
cerebellum in adaptation to delayed action effects. Current
Biology, 27, 2442–2451.
Champion, R. A., & Adams, W. J. (2007). Modification of the
convexity prior but not the light-from-above prior in visual
search with shaded objects. Journal of Vision, 7, 10.
Cousineau, D. (2005). Confidence intervals in within-subject
designs: A simpler solution to Loftus and Masson’s method.
Tutorials in Quantitative Methods for Psychology, 1, 42–45.
Crapse, T. B., & Sommer, M. A. (2008). Corollary discharge across
the animal kingdom. Nature Reviews Neuroscience, 9, 587–600.
Dale, C. L., Brown, E. G., Fisher, M., Herman, A. B., Dowling,
A. F., Hinkley, L. B., et al. (2016). Auditory cortical plasticity
drives training-induced cognitive changes in schizophrenia.
Schizophrenia Bulletin, 42, 220–228.
Elijah, R. B., Le Pelley, M. E., & Whitford, T. J. (2016). Modifying
temporal expectations: Changing cortical responsivity to
delayed self-initiated sensations with training. Biological
Psychology, 120, 88–95.
Feinberg, I. (1978). Efference copy and corollary discharge:
Implications for thinking and its disorders. Schizophrenia
Bulletin, 4, 636–640.
Feinberg, I., & Guazzelli, M. (1999). Schizophrenia—A disorder
of the corollary discharge systems that integrate the motor
systems of thought with the sensory systems of
consciousness. British Journal of Psychiatry, 174, 196–204.
Fletcher, P. C., & Frith, C. D. (2009). Perceiving is believing: A
Bayesian approach to explaining the positive symptoms of
schizophrenia. Nature Reviews Neuroscience, 10, 48–58.
Ford, J. M., & Mathalon, D. H. (2004). Electrophysiological
evidence of corollary discharge dysfunction in schizophrenia
during talking and thinking. Journal of Psychiatric Research,
38, 37–46.
Ford, J. M., Mathalon, D. H., Heinks, T., Kalba, S., Faustman,
W. O., & Roth, W. T. (2001). Neurophysiological evidence of
corollary discharge dysfunction in schizophrenia. American
Journal of Psychiatry, 158, 2069–2071.
Ford, J. M., Palzes, V. A., Roach, B. J., & Mathalon, D. H. (2014).
Did I do that? Abnormal predictive processes in schizophrenia
when button pressing to deliver a tone. Schizophrenia
Bulletin, 40, 804–812.
Friston, K. (2005). A theory of cortical responses. Philosophical
Transactions of the Royal Society of London, Series B:
Biological Sciences, 360, 815–836.
Frith, C. (2005). The neural basis of hallucinations and
delusions. Comptes Rendus Biologies, 328, 169–175.
Gratton, G., Coles, M. G., & Donchin, E. (1983). A new method
for off-line removal of ocular artifact. Electroencephalography
and Clinical Neurophysiology, 55, 468–484.
Hatada, Y., Miall, R. C., & Rossetti, Y. (2006). Long lasting
aftereffect of a single prism adaptation: Directionally biased
shift in proprioception and late onset shift of internal
egocentric reference frame. Experimental Brain Research,
174, 189–198.
Heinks-Maldonado, T. H., Mathalon, D. H., Gray, M., & Ford,
J. M. (2005). Fine-tuning of auditory cortex during speech
production. Psychophysiology, 42, 180–190.
Heinks-Maldonado, T. H., Nagarajan, S. S., & Houde, J. F. (2006).
Magnetoencephalographic evidence for a precise forward
model in speech production. NeuroReport, 17, 1375–1379.
Hershberger, W. (1970). Attached-shadow orientation
perceived as depth by chickens reared in an environment
illuminated from below. Journal of Comparative and
Physiological Psychology, 73, 407–411.
Hess, E. H. (1950). Development of the chick’s responses to
light and shade cues of depth. Journal of Comparative and
Physiological Psychology, 43, 112–122.
Horváth, J. (2015). Action-related auditory ERP attenuation:
Paradigms and hypotheses. Brain Research, 1626, 54–65.
Houde, J. F., Nagarajan, S. S., Sekihara, K., & Merzenich, M. M.
(2002). Modulation of the auditory cortex during speech:
Elijah, Le Pelley, and Whitford
1155
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
j
o
c
n
_
a
_
0
1
2
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
An MEG study. Journal of Cognitive Neuroscience, 14,
1125–1138.
Hughes, G., Desantis, A., & Waszak, F. (2013a). Mechanisms of
intentional binding and sensory attenuation: The role of
temporal prediction, temporal control, identity prediction,
and motor prediction. Psychological Bulletin, 139, 133–151.
Hughes, G., Desantis, A., & Waszak, F. (2013b). Attenuation
of auditory N1 results from identity-specific action-effect
prediction. European Journal of Neuroscience, 37, 1152–1158.
Kleffner, D. A., & Ramachandran, V. S. (1992). On the perception
of shape from shading. Perception & Psychophysics, 52, 18–36.
Kleiner, M., Brainard, D. H., & Pelli, D. G. (2007). What’s new in
Psychtoolbox-3? Perception, 36, ECVP Abstract Supplement.
Lange, K. (2011). The reduced N1 to self-generated tones:
An effect of temporal predictability? Psychophysiology, 48,
1088–1095.
Luck, S. J. (2005). An introduction to the event-related potential
technique. Cambridge, MA: MIT Press.
McClure, S. M., Berns, G. S., & Montague, P. R. (2003). Temporal
prediction errors in a passive learning task activate human
striatum. Neuron, 38, 339–346.
Mifsud, N. G., Beesley, T., Watson, T. L., & Whitford, T. J.
(2016). Attenuation of auditory evoked potentials for hand
and eye-initiated sounds. Biological Psychology, 120, 61–68.
Morton, S. M., & Bastian, A. J. (2004). Prism adaptation during
walking generalizes to reaching and requires the cerebellum.
Journal of Neurophysiology, 92, 2497–2509.
Mulert, C., Jäger, L., Propp, S., Karch, S., Störmann, S., Pogarell,
O., et al. (2005). Sound level dependence of the primary
auditory cortex: Simultaneous measurement with 61-channel
EEG and fMRI. Neuroimage, 28, 49–58.
Näätänen, R., & Picton, T. (1987). The N1 wave of the human
electric and magnetic response to sound: A review and an
analysis of the component structure. Psychophysiology, 24,
375–425.
Niemi, P., & Näätänen, R. (1981). Foreperiod and simple reaction
time. Psychological Bulletin, 89, 133–162.
unspecific N1 component of the auditory ERP. Psychophysiology,
50, 334–343.
Saupe, K., Widmann, A., Trujillo-Barreto, N. J., & Schröger, E.
(2013). Sensorial suppression of self-generated sounds and
its dependence on attention. International Journal of
Psychophysiology, 90, 300–310.
Simmons, J. G., Nathan, P. J., Berger, G., & Allen, N. B. (2011).
Chronic modulation of serotonergic neurotransmission with
sertraline attenuates the loudness dependence of the auditory
evoked potential in healthy participants. Psychopharmacology,
217, 101–110.
Shadmehr, R., Smith, M. A., & Krakauer, J. W. (2010). Error
correction, sensory prediction, and adaptation in motor
control. Annual Review of Neuroscience, 33, 89–108.
Stetson, C., Cui, X., Montague, P. R., & Eagleman, D. M. (2006).
Motor-sensory recalibration leads to an illusory reversal of
action and sensation. Neuron, 51, 651–659.
Subramaniam, K., Luks, T. L., Fisher, M., Simpson, G. V.,
Nagarajan, S., & Vinogradov, S. (2012). Computerized
cognitive training restores neural activity within the
reality monitoring network in schizophrenia. Neuron, 73,
842–853.
Timm, J., SanMiguel, I., Saupe, K., & Schröger, E. (2013). The
N1-suppression effect for self-initiated sounds is independent
of attention. BMC Neuroscience, 14, 2.
Timm, J., Schönwiesner, M., Schröger, E., & SanMiguel, I. (2016).
Sensory suppression of brain responses to self-generated
sounds is observed with and without the perception of agency.
Cortex, 80, 5–20.
Todorovic, A., & de Lange, F. P. (2012). Repetition suppression
and expectation suppression are dissociable in time in
early auditory evoked fields. Journal of Neuroscience, 32,
13389–13395.
van Laarhoven, T., Stekelenburg, J. J., & Vroomen, J. (2017).
Temporal and identity prediction in visual-auditory events:
Electrophysiological evidence from stimulus omissions.
Brain Research, 1661, 79–87.
Oestreich, L. K. L., Mifsud, N. G., Ford, J. M., Roach, B. J.,
Vinogradov, S., Fisher, M., & de Villers-Sidani, E. (2012).
Mathalon, D. H., & Whitford, T. J. (2016). Cortical suppression
to delayed self-initiated auditory stimuli in schizotypy:
Neurophysiological evidence for a continuum of psychosis.
Clinical EEG and Neuroscience, 47, 3–10.
Pelli, D. G. (1997). The VideoToolbox software for visual
psychophysics: Transforming numbers into movies. Spatial
Vision, 10, 437–442.
Penadés, R., Pujol, N., Catalán, R., Massana, G., Rametti, G.,
García-Rizo, C., et al. (2013). Brain effects of cognitive
remediation therapy in schizophrenia: A structural and
functional neuroimaging study. Biological Psychiatry, 73,
1015–1023.
Ransil, B. J., & Schachter, S. C. (1994). Test–retest reliability of
the Edinburgh Handedness Inventory and global handedness
preference measurements, and their correlation. Perceptual
and Motor Skills, 79, 1355–1372.
Rao, R. P. N., & Ballard, D. H. (1999). Predictive coding in the
visual cortex: A functional interpretation of some extra-classical
receptive-field effects. Nature Neuroscience, 2, 79–87.
Roussel, C., Hughes, G., & Waszak, F. (2013). A preactivation
account of sensory attenuation. Neuropsychologia, 51, 922–929.
SanMiguel, I., Todd, J., & Schröger, E. (2013). Sensory suppression
effects to self-initiated sounds reflect the attenuation of the
Cognitive training for impaired neural systems in
neuropsychiatric illness. Neuropsychopharmacology
Reviews, 37, 43–76.
von Holst, E. (1954). Relations between the central nervous
system and the peripheral organs. British Journal of Animal
Behaviour, 2, 89–94.
Whitford, T. J., Ford, J. M., Mathalon, D. H., Kubicki, M., &
Shenton, M. E. (2012). Schizophrenia, myelination, and
delayed corollary discharges: A hypothesis. Schizophrenia
Bulletin, 38, 486–494.
Whitford, T. J., Jack, B. N., Pearson, D., Griffiths, O., Luque, D.,
Harris, A. W. F., et al. (2017). Neurophysiological evidence
of efference copies to inner speech. eLife, 6, e28197.
Whitford, T. J., Mathalon, D. H., Shenton, M. E., Roach, B. J.,
Bammer, R., Adcock, R. A., et al. (2011). Electrophysiological
and diffusion tensor imaging evidence of delayed corollary
discharges in patients with schizophrenia. Psychological
Medicine, 41, 959–969.
Wolpert, D. M., Ghahramani, Z., & Jordan, M. I. (1995). An
internal model for sensorimotor integration. Science, 269,
1880–1882.
Wolpert, D. M., & Miall, R. C. (1996). Forward models for
physiological motor control. Neural Networks, 9, 1265–1279.
1156
Journal of Cognitive Neuroscience
Volume 30, Number 8
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
3
0
8
1
1
4
5
1
7
8
7
5
5
8
/
/
j
o
c
n
_
a
_
0
1
2
6
9
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3