A Brain Mechanism for Facilitation of Insight
by Positive Affect
Karuna Subramaniam, John Kounios, Todd B. Parrish,
and Mark Jung-Beeman
Astratto
& Previous research has shown that people solve insight or
creative problems better when in a positive mood (assessed
or induced), although the precise mechanisms and neural sub-
strates of this facilitation remain unclear. We assessed mood and
personality variables in 79 participants before they attempted
to solve problems that can be solved by either an insight or an
analytic strategy. Participants higher in positive mood solved
more problems, and specifically more with insight, compared
with participants lower in positive mood. fMRI was performed on
27 of the participants while they solved problems. Positive mood
(and to a lesser extent and in the opposite direction, anxiety)
was associated with changes in brain activity during a prepara-
tory interval preceding each solved problem; modulation of pre-
paratory activity in several areas biased people to solve either
with insight or analytically. Analyses examined whether (UN) posi-
tive mood modulated activity in brain areas showing respon-
sivity during preparation; (B) positive mood modulated activity
in areas showing stronger activity for insight than noninsight
trials either during preparation or solution; E (C) insight ef-
fects occurred in areas that showed mood-related effects during
preparation. Across three analyses, the ACC showed sensitivity
to both mood and insight, demonstrating that positive mood
alters preparatory activity in ACC, biasing participants to engage
in processing conducive to insight solving. This result suggests
that positive mood enhances insight, at least in part, by mod-
ulating attention and cognitive control mechanisms via ACC,
perhaps enhancing sensitivity to detect non-prepotent solution
candidates. &
INTRODUCTION
This article investigates the neural basis of one way
that affect modulates cognition. Specifically, we report
changes in brain activity, as measured with fMRI, Quello
occur as affect modulates problem-solving strategies.
People can solve problems through methodical, analytic
processing, through insight, or through some mix of
both (for recent reviews, see Bowden, Jung-Beeman,
Fleck, & Kounios, 2005; Gilhooly & Murphy, 2005).
These two strategies or (sets of ) processes can co-occur,
sovrapposizione, and interact, yet they are phenomenologically,
behaviorally, and neurologically distinct, as described
below. It has previously been demonstrated that posi-
tive affect (PAPÀ) specifically facilitates people’s ability to
solve creative or ‘‘insight problems,’’ that is, problems
that are more often solved with insight (Rowe, Hirsch,
& Anderson, 2007; Amabile, Barsade, Mueller, & Staw,
2005; Isen, 1999UN, 1999B; Estrada, Young, & Isen, 1994;
Isen, Daubman, & Nowicki, 1987). Therefore, observing
brain activity associated with shifts of problem-solving
approaches in different affective states provides fertile
ground for examining the neural mechanisms of emotion–
cognition interactions. Here, we show that distinct affect
states change actual cognitive organization to modulate
Northwestern University, Evanston, IL
problem-solving processes beyond the well-documented
mood–memory congruency effect (Teasdale & Fogarty,
1979).
The distinction between insight and analytic solving
has been anecdotally recognized for millennia and has
been the subject of scientific inquiry for nearly a cen-
tury (per esempio., Duncker, 1945; Maier, 1930; Kohler, 1917). UN
plethora of behavioral evidence details how these two
solving processes differ. Analytic processing involves de-
liberate application of strategies and operations to grad-
ually approach solution. Insight, which is considered a
type of creative cognition, is the process through which
people suddenly and unexpectedly achieve solution
through processes that are not consciously reportable.
Insight solutions tend to involve conceptual reorganiza-
zione, often occurring after solvers overcome an impasse
in their solving effort, and are suddenly able to recognize
distant or atypical relations between problem elements
that had previously eluded them (Gilhooly & Murphy,
2005; Smith & Kounios, 1996; Schooler & Melcher, 1995;
Weisberg, 1994; Schooler, Ohlsson, & Brooks, 1993;
Metcalfe & Weibe, 1987; Metcalfe, 1986). When solution
is achieved, these factors combine to create a unique phe-
nomenological experience, termed the Aha! or Eureka!
moment.
PA has been shown to facilitate insight and creative
problem solving across a broad range of settings (Rowe
D 2008 Istituto di Tecnologia del Massachussetts
Journal of Cognitive Neuroscience 21:3, pag. 415–432
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
/
.
.
F
T
.
o
N
1
8
M
UN
sì
2
0
2
1
et al., 2007; Ashby, Isen, & Turken, 1999; Isen, 1999B;
Estrada, Isen, & Young, 1997; Isen et al., 1987). One
description of this effect is that PA enhances cognitive
flexibility in various settings, such as in classifying ma-
terial (Isen & Daubman, 1984),
in negotiation tasks
(Carnevale & Isen, 1986), in medical diagnoses (Estrada
et al., 1994), and in creative problem-solving tasks
(Isen, Mitzi, Johnson, Mertz, & Robinson, 1985; Isen &
Daubman, 1984). Various explanations have been pro-
posed to explain this facilitation (also see Discussion
section). Briefly, one hypothesis is that PA promotes a
more global scope of attention (Bolte, Goschke, & Kuhl,
2003; Gasper & Clore, 2002), enhancing access to distant
or unusual associations (Friedman, Fishbein, Fo¨rster,
& Werth, 2003; Federmeier, Kirson, Moreno, & Kutas,
2001; Isen et al., 1985), which facilitates creative solu-
tions to classic insight problems such as Duncker’s
(1945) candle task (Isen et al., 1987) and improves per-
formance (Rowe et al., 2007; Isen et al., 1987) on the
Remote Associates Test (Mednick, 1962). Another hy-
pothesis is that PA enhances switching between global
and local attentional modes (Baumann & Kuhl, 2005)
or between strategies (Dreisbach & Goschke, 2004), O,
allo stesso modo, that it enhances selection of different perspec-
tives (Ashby et al., 1999).
In contrasto, negative affect (NA) states such as anxiety
and depression have been associated with deficits in atten-
tional and cognitive control mechanisms (Bishop, Duncan,
Brett, & Lawrence, 2004; Mayberg et al., 1999), often in-
ducing a narrow scope of attention (Easterbrook, 1959).
Therefore, anxiety in particular should impede cognitive
flexibility, problem restructuring, and insight solving.
This study extends the existing literature in two ways.
Primo, we examine not just the facility in solving a
particular type of problem, but how mood modulates
which strategy, insight or analytic, is preferred (or suc-
cessful). Secondo, we measure brain activity as people
solve these problems to observe the neural mechanisms
of insight problem solving that are modulated by mood.
Insight and analytic problem solving are associated
with different patterns of brain activity, measured with
both fMRI and EEG, both at the moment people achieve
solution ( Jung-Beeman et al., 2004) and as people pre-
pare for each new problem (Kounios et al., 2006). For
one thing, the right hemisphere (RH), generally, seems
to make stronger contributions as people process in-
sight problems and recognize their solutions (Bowden
& Jung-Beeman, 2003UN; Beeman & Bowden, 2000;
Bowden & Beeman, 1998). More specifically, compared
with solving problems without insight, solving with
insight involves stronger activity in right temporal re-
gions thought to be important for integrating distant
semantic associations ( Jung-Beeman et al., 2004). Addi-
tional brain regions showed similar but weaker ‘‘insight
effects’’ in the earlier study but manifested strong effects
in the current study; these include anterior cingulate,
posterior cingulate cortex (PCC), parahippocampal cor-
tex (PHC), right superior frontal gyrus (SFG), and right
inferior parietal lobe (IPL).
Additionally, during a brief preparation period prior to
the presentation of each problem, various brain regions
are more active prior to problems solved with insight
than prior to problems solved without insight (Kounios
et al., 2006). Questo è, different patterns of brain activity
are conducive to solving the subsequent problem with
insight versus analytic processing. The distinguishing
areas include bilateral temporal areas involved with se-
mantic processing, PCC putatively involved in attention,
and ACC thought to be important for cognitive control.
Così, each of these areas represents a reasonable can-
didate for affect-induced modulation of insight problem
risolvere. The left temporal cortex is more adept at pre-
paring to retrieve many close prepotent associations,
whereas activity in the right temporal cortex enhances
the readiness to pursue weaker associations ( Jung-
Beeman, 2005). D'altra parte, the posterior cingu-
late is thought to be involved in visuospatial expectancy
(Small et al., 2003), and the anterior cingulate is more
likely to be involved in cognitive control and possibly
in switching between solution candidates (or other
thought processes), which is likely an important com-
ponent of insight.
Anterior Cingulate and Insight Processes
We have demonstrated that the rostral portion of the
dorsal anterior cingulate cortex (dACC; Brodmann’s area
[BA] 9, 24, 32) showed a sustained increase in neural
activity during the preparatory interval before partici-
pants actually see problems, and stronger ACC activity
occurs prior to trials solved with insight than those
solved more analytically (Kounios et al., 2006).
We hypothesized that insights would involve greater
cognitive control and restructuring processes, and that
the dACC would be involved in the shift and the se-
lection of a new solution path. In tasks involving re-
sponse competition, cognitive control is thought to be
important for the monitoring of competing responses
( Weissman, Giesbrecht, Song, Mangun, & Woldorff,
2003; Van Veen, Cohen, Botvinick, Stenger, & Carter,
2001; MacDonald, Cohen, Stenger, & Carter, 2000), In
overcoming prepotent responses when strategic pro-
cesses were less engaged and conflict was high (Carter
et al., 2000), and in shifting attention (Davis et al., 2005;
Dreisbach & Goschke, 2004; Kondo, Osaka, & Osaka,
2004). Such cognitive control mechanisms could be
critical for insight because they enable problem solvers
to detect competing solution candidates, rely less on dom-
inant associations or strategies, and/or enable shifting
attention from a prepotent but irrelevant association to
the less potent, but correct, association. This could be an
important component of what insight researchers vari-
ously term cognitive restructuring and flexibility or
‘‘breaking set’’ and ‘‘overcoming functional fixedness.’’
416
Journal of Cognitive Neuroscience
Volume 21, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
T
.
F
/
.
.
o
N
1
8
M
UN
sì
2
0
2
1
Anterior Cingulate, PAPÀ, and Insight
One possible mechanism by which PA could facilitate
insight is through cognitive restructuring processes. PAPÀ
is likely to facilitate insight by increasing a person’s
ability to switch and select alternative cognitive perspec-
tives (Baumann & Kuhl, 2005; Dreisbach & Goschke,
2004; Isen, 1999B), reducing perseveration on one par-
ticular solution candidate or solving approach, così
increasing the probability of engaging in various cogni-
tive restructuring processes. We propose that PA could
modulate activity in ACC (Lane, Reiman, Axelrod, Yun,
& Holmes, 1998) to make it more open to detecting
competing (weak) activations, biasing a shift toward in-
sight solutions. The modulated ACC activity might fa-
cilitate one or a combination of mechanisms such as
switching between global and local processing modes of
Attenzione (Baumann & Kuhl, 2005), switching from ir-
relevant to relevant solving strategies, and/or selecting
the correct solution (Dreisbach & Goschke, 2004).
ACC appears to be a particularly promising site for in-
teractions between cognitive processes and affect states.
Besides its involvement in modulating cognitive pro-
cesses via attention shifting, conflict detection, risposta
competition, and/or selection mechanisms (Badre &
Wagner, 2004; Botvinick, Cohen, & Carter, 2004; Kerns
et al., 2004; Dreher & Grafman, 2003; Ruff, Woodward,
Laurens, & Liddle, 2001; Bush, Luu, & Posner, 2000),
ACC also appears to be involved in emotional processes
(Bush et al., 2000; Mayberg et al., 1999; Drevets & Raichle,
1998; Whalen et al., 1998). Functional neuroimaging
studies show overlapping activation patterns within
ACC between cognitive and affective tasks (Fichtenholtz
et al., 2004; Teasdale et al., 1999; Lane et al., 1998; Papez,
1937). Electrophysiological studies have identified a pop-
ulation of dACC neurons that show increased activity to
high- versus low-conflict Stroop tasks, including those
with emotional valence (Davis et al., 2005). Inoltre,
cytoarchitectonic studies suggest the involvement of
specialized spindle cells of BA 24 that integrate cognitive
input with emotional overtones (Nimchinsky et al., 1999).
Given ACC’s involvement in cognitive control and
emotional processes and our prior evidence that activity
in ACC prior to solving problems is associated with
solution strategy, we predict that affect states will mod-
ulate ACC activation and thereby influence insight (versus
analytic)-solving processes. Specifically, we hypothesize
that PA states will increase activity in ACC before the
actual problem onset, biasing the solver toward cognitive
processing that is relatively conducive to insight.
Hemispheric Asymmetries, Affect, and Insight
Another possibility can be derived from the following
considerations: (1) RH processing seems to make strong
contributions to insight solving overall ( Jung-Beeman
et al., 2004; Bowden & Beeman, 1998); (2) RH semantic
processing activates or maintains activation of a broader
set of semantic associations than does LH semantic pro-
cessazione (Faust & Mashal, 2007; Beeman et al., 1994;
Chiarello, 1991), and these broad associations seem very
relevant for solving with insight; (3) positive mood
seems to broaden the overall pattern of semantic assoc-
iations (Federmeier et al., 2001; Isen et al., 1985); (4) global
or broad attention is associated with RH visual process-
ing, creative problem solving (Ansburg & Hill, 2003), E
positive mood (Rowe et al., 2007; Gasper & Clore, 2002);
E (5) inducing an approach regulatory focus (with low
arousal) increases measures of relative RH activation as
well as facilitating creative problem solving (Friedman &
Fo¨rster, 2005). Così, it remains hypothetically possible
that PA will directly increase overall activity in the RH,
specifically in the right superior temporal gyrus (STG),
che è, cytoarchitectonically more suited than the left
STG at integrating distant semantic associates via coarse
semantic coding (for a review, see Jung-Beeman, 2005).
Tuttavia, such an effect might seem to contradict some
established associations between positive mood (or ap-
proach focus) and leftward asymmetries in electroenceph-
alographic activity (Herrington, Mohanty, Koven, Fisher,
& Stewart, 2005; Davidson, 1992; Tomarken, Davidson,
Wheeler, & Doss, 1992). Inoltre, to us, it seems intui-
tively more likely that a global characteristic like positive
mood would either modulate all semantic processing (In
both hemispheres) to broaden the scope of semantic asso-
ciations or, more likely, to modulate attention or cognitive
control mechanisms that make solvers better able to de-
tect (and utilize) remote associations that are only weakly
active (perhaps, mostly due to RH semantic processing).
Experiment
Insight typically occurs when people initially focus on
an incorrect but dominant association (per esempio., in Figure 1,
ache can form compounds with tooth and heart but not
potato) and need to overcome this impasse and switch
to the correct solving strategy to be able to reach a sud-
den (Aha!) understanding of the solution ( Jung-Beeman
et al., 2004; Bowden & Jung-Beeman, 2003UN). In many
studies of insight solving, problems have typically been
classified a priori, as either ‘‘insight problems’’ or ‘‘non-
insight problems’’ (Weisberg, 1994); but because any
problem can be solved through insight, through straight-
forward (incremental, strategic) problem solving, O
through a combination of both (Bowden et al., 2005),
the a priori ‘‘insight’’ classification is not definitive.
We exploit this feature by asking participants to report
directly which strategy they used predominantly to
achieve solutions to directly contrast trials that lead to
insight solutions versus those that lead to noninsight
solutions. This enables us to examine insight versus
noninsight processing while holding task and stimulus
type constant. Participants were presented with a large
set of compound remote associate (CRA) problems
Subramaniam et al.
417
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
.
T
/
.
F
.
o
N
1
8
M
UN
sì
2
0
2
1
formed consent, all participants completed mood state
inventories for the Positive and Negative Affect Schedule
(PANAS), the State-Trait Anxiety Inventory (STAI), and a
variety of other personality inventories measuring more
stable individual traits (the Behavioral Inhibition Scale–
Behavioral Activation Scale, the Neuroticism subscale
for the Big 5 Personality Mini-Markers, and the Magical
Ideation Scale as an indicator of schizotypy). The mood
state inventories (PANAS and STAI), given to all par-
ticipants just before they performed the CRA task,
measured the extent that participants were currently ex-
periencing a positive (PANAS) or anxious mood (STAI).
We examined correlations between all mood and per-
sonality scores and various problem-solving measures
(solving rate and proportion of problems solved with
insight) as well as fMRI signal change.
After these questionnaires, 52 participants performed
the problem-solving task outside the scanner, providing
only behavioral data, E 30 participants performed the
problem-solving task in the scanner. Data from three
participants were excluded—due to poor fMRI signal in
two of the participants and due to one participant pro-
viding only two analytic responses.
Problem-solving Paradigm
We measured insight and analytical solving of 135 CRA
problems (Bowden & Jung-Beeman, 2003B), adapted
from a test of creative cognition (Mednick, 1962). For
each problem, participants see three problem words
(tooth, potato, and heart) and must generate a solution
(sweet) that can form a compound word or phrase with
each problem word (sweet tooth, sweet potato, sweet-
heart). The solution word can precede or follow each
problem word. Like most problems (even classic ‘‘in-
sight problems’’), these problems can be solved either
with insight or through more methodical or analytical
processes. We relied on participants’ trial-by-trial judg-
ments to determine the type of processing that led to
each solution. This method has reliably shown consis-
tent differences in behavior (Bowden & Jung-Beeman,
2003UN; Beeman & Bowden, 2000; Bowden & Beeman,
1998) and in brain activity (Kounios et al., 2006, 2008;
Jung-Beeman et al., 2004). For instance, in our prior EEG
study, the neural processes biasing the sudden (Aha!)
that led up to an insight solution were associated with
increased neural activity (less alpha power) peaking over
midfrontal cortex and bilateral temporal cortices for in-
sight versus analytical preparatory processes (Kounios
et al., 2006). Using a different population sample and
methodology, fMRI signal corroborated the EEG find-
ing, specifically isolating ACC as the medial frontal
region that revealed increased neural activity for insight
versus noninsight preparatory processing, and also
showed increased activity within the bilateral temporal
cortical areas revealed during EEG (Kounios et al., 2006).
In another study, about a third of a second prior to the
Figura 1. Sequence of events within a trial of the CRA task. Each
trial began with a central fixation cross, signaling the onset of the
preparation interval, which lasted for a variable 0–8 sec, after which
the problem was presented. Participants pressed the response buttons
bimanually if/when they achieved solution, verbalized the solution
at the Solution prompt, and then reported whether they solved the
trial with or without insight at the Insight prompt. The intervals
between these events were jittered for a variable 0–8 sec.
(Bowden & Jung-Beeman, 2003B). Similar problems
(Mednick, 1962) are often used as ‘‘insight problems’’
or for creative problem solving (Isen, 1999UN; Isen et al.,
1987), and the ability to solve them correlates with the
ability to solve other classic insight problems (Duncker,
1945; Maier, 1930). Tuttavia, they can be solved either
analytically or with insight (Bowden & Jung-Beeman,
2003UN; for a review, see Bowden et al., 2005). The type of
processing involved in successfully solving these prob-
lems varies across trials (Kounios et al., 2006; Jung-
Beeman et al., 2004) and across individuals (Kounios
et al., 2008), making this paradigm a strong candidate
for investigating how affect can, in a general (piuttosto che
in only a mood-consistent) modo, modulate higher level
cognition. Specifically, we examine the brain basis of how
PA and anxiety modulate solving strategy, tipping the
balance of processes toward insight or analytic strategies.
Mood state (including PA and NA and anxiety) and per-
sonality measures, gathered prior to the experimental
session, were related to performance and neuroimaging
measures. For the participants who underwent fMRI scan-
ning, we identified brain regions involved in various
aspects of problem solving and correlated the signal
change in these regions with the mood and personality
indices, as well as identifying areas that showed contrast
in the brain activity during problem preparation between
high and low positive mood individuals.
METHODS
Participants and Procedure
Tutto 79 participants were neurologically healthy, right-
handed, and native speakers of English. After giving in-
418
Journal of Cognitive Neuroscience
Volume 21, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
F
.
.
T
/
.
o
N
1
8
M
UN
sì
2
0
2
1
insight solution button press, a burst of EEG gamma
activity in the right anterior superior temporal gyrus
(aSTG) corresponded to the increase in fMRI solution-
related signal within the same region ( Jung-Beeman
et al., 2004). This RH activation likely reflects the pro-
cessing and integration of a broad range of semantic as-
sociations leading to solution ( Jung-Beeman et al., 2004;
Bowden & Jung-Beeman, 2003UN; Bowden & Beeman,
1998; Beeman et al., 1994).
Prior to the current experiment, participants received
instructions to make insight/noninsight judgments, em-
phasizing that they should respond ‘‘insight’’
if they
achieved solution suddenly and surprisingly, possibly by
switching their train of thought just prior to solution,
and that as soon as they thought of the solution can-
didate, they were instantly confident it was the solution.
In contrasto, they should respond ‘‘noninsight’’ if they
achieved solution incrementally or by some analytical
strategy, Per esempio, by strategically retrieving candi-
dates and testing them out.
Each trial began with a fixation cross that remained
on the screen for a variable rest period (from 0, 2, 4, 6,
O 8 sec, randomized across all trials), during which
participants prepared for the next trial (Figura 1). Such
variable delays were used to jitter the events and to
optimize deconvolution of the fMRI signal from suc-
cessive events. After this preparation period, the three
problem words (tooth, potato, and heart) were pre-
sented on the screen (horizontally centered, just above,
at, and just below central fixation) and persisted until
participants solved the problem or a 15-sec time limit
was reached. Participants attempted to produce a sin-
gle solution word (sweet) that could form a compound
word with each of the problem words. If participants
solved the problem, they made a bimanual button press
(to avoid biasing laterality of cortical activity) by pressing
the two outer buttons with a finger on each hand when
they arrived at the solution; after a variable (0–8 sec)
delay, a solution prompt appeared, and participants
verbalized the solution. After another variable delay
(0–8 sec), an insight prompt (‘‘Insight?’’) appeared,
and participants pressed the two outer buttons with a
finger on each hand if they had reached the solution
with an insight, or they pressed the two inner buttons if
they had reached the solution through analytic nonin-
sight means. After the insight/analytical solution rating,
or after 15 sec elapsed on unsolved trials, il prossimo
preparation period began.
Image Acquisition
Thirty fMRI participants performed the CRA task dur-
ing scanning, which for all participants occurred in the
same Siemens Trio (3 T) scanner and eight-channel
head coil, with the same scanning protocol, at North-
western’s Center for Advanced MRI. Head motion was
restricted with plastic calipers built into the coil and a
vacuum pillow. The functional imaging sequence was op-
timized for detection of the BOLD effect (Ogawa et al.,
1992) including local shimming and 8 sec of scanning
prior to data collection to allow the MR signal to reach
equilibrium. Functional
imaging used a gradient-echo
echo-planar sequence (TR = 2 sec for thirty-eight 3-mm
slices, TE = 20 msec, matrix size = 64 (cid:1) 64 in 220-mm
field of view). Participants solved problems during four
scans of 10 min 20 sec and a final fifth scan that was
truncated when participants finished solving problems.
Each functional scan was synchronized with the onset
of the first problem in that block of trials; timing of
subsequent trials was response dependent and not
synchronized with image acquisition. Anatomical high-
resolution images were acquired in the same plane, con
T1-weighted images parallel to the AC–PC plane.
Image Analysis
Functional and anatomical
images were coregistered
through time, spatially smoothed with a 7.5-mm Gaussian
kernel, and fit to a common template. Within each run,
voxels were eliminated if the signal magnitude changed
more than 20% across successive TRs, or if the mean
signal level was below a noise threshold. Functional data
were transformed (Collins, Neelin, Peters, & Evans, 1994)
to a standard stereotaxic atlas (Talairach & Tournoux,
1988) with a voxel size of 2.5 mm3. The data were
analyzed using general linear model analysis, as imple-
mented in AFNI ( Ward, http://afni.nimh.nih.gov/afni),
that extracted average estimated responses to each trial
type, correcting for linear drift and removing signal
changes correlated with head motion as well as signal
attributed to other temporally adjacent events to ensure
that signal could be isolated to the event of interest. For
esempio, when extracting signal related to preparation
events, we included in the analysis the preceding insight
ratings, the subsequent problem onsets, and the sub-
sequent solutions to factor out signal more closely tied
to those events than to the preparation event. Signal was
estimated for all time points (TRs 0–10) within the same
modello, without regard to any presumed hemodynamic
response function.
The primary focus of this report was fMRI signal,
hence brain activity, corresponding to the preparation
intervals. We examined changes in BOLD signal after the
onset of this preparation period in three ways:
(UN) Areas that turned on, questo è, changed their activity,
during preparation. We examined overall responsivity
corresponding to the preparation interval, manifested
as a rise and fall of BOLD signal from onset of the
preparation period to peak response and back down
to baseline. Specifically, for every voxel, signal cor-
responding to the peak of the preparation period
(TRs 4, 5, E 6 after onset of preparation period; for
comparison, there was a peak signal in motor cortex
Subramaniam et al.
419
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
F
T
.
.
.
/
o
N
1
8
M
UN
sì
2
0
2
1
at TR 3, corresponding to the button press from the
insight-rating preceding the preparation period) era
contrasted with signal corresponding to the points
preceding and after the preparation period (TRs 1, 9,
E 10). We identified regions of signal change that
were consistent across all 27 participants, with a sig-
nificance threshold combining t values ( P < .005) and
cluster size (at least 1500 mm3 in volume). The dACC,
the PCC, and the right angular gyrus (AG) clusters
exceeded the above criteria, increasing preparatory ac-
tivity. Of all these statistically reliable clusters (func-
tionally defined ROIs), the dACC and the right AG
were the only two ROIs where preparatory respon-
sivity strongly correlated with positive mood across all
27 participants.
Because any changes (up or down) in activity could
be meaningful, to be thorough we also examined areas
that exhibited deactivation, that is, a fall and a rise of
signal corresponding to the preparation interval. The left
and the right inferior frontal gyrus (IFG) showed sys-
tematic preparatory deactivation in which the mean
signal for the expected preparatory peak hemodynamic
signal (i.e., TRs 4, 5, 6) was significantly lower than the
mean baseline signal (i.e., the first TR and last two TRs).
Neither of these areas exhibited correlations between
signal change and mood.
(B) Areas that showed insight-specific activity during
preparation or solution. Peak preparatory signal spe-
cific to insight trials was calculated by comparing the
difference between insight and analytic preparation
events for each participant by extracting the mean
signal within the three TRs (TRs 4, 5, and 6) corre-
sponding to the expected preparatory hemodynamic
peak. For comparison, the preceding insight-rating
button press elicited peak signal in motor cortex at
4 sec, just prior to the preparation onset peak signal
(6 sec) for each participant. Similarly, peak insight
solution-related signal was calculated by examining
differences between insight and analytic solution
events for each participant by examining the mean
signal within the three TRs (TRs 3, 4, and 5—we
chose an early time window to minimize contami-
nation from postsolution activity) corresponding to
the expected peak signal leading up to the solution
point (see Figure 9 for comparison). The subsequent
button press elicited peak signal in motor cortex
(10 sec) at the solution point. The significance thresh-
old combined cluster size and t values for each voxel
within a cluster (set at least 500 mm3 in volume) in
which each voxel was reliably different across par-
ticipants, t(26) = 3.09, p < .005 uncorrected, for
insight versus noninsight preparation and for insight
versus noninsight solutions. ACC, PCC,
left STG,
and right MTG ROI clusters exceeded these criteria,
manifesting stronger preparatory peak signal for in-
sight versus analytical trials. Several regions showed
stronger peak signal for insight versus analytical so-
lutions including ACC, PCC, right PHC, left MTG, right
MTG, right IPL, and right SFG.
We then extracted the mean preparatory hemody-
namic responsivity signal for each participant, as de-
scribed by item A, within the regions that showed an
insight effect at preparation, and within the regions that
showed an insight effect at solution, as described above.
We correlated this preparatory responsivity within these
‘‘insight’’ regions with positive mood (PA–NA) and
anxiety (STAI) scores. Of all the ROIs defined by the
insight effect that corresponded to the time window at
preparation and the time window leading up to the
solution point, only ACC ROI manifested strong corre-
lations between overall preparatory signal change and
positive mood.
(C) Areas that showed mood differences in activity during
preparation. To examine how individual differences
in affect state influenced successful preparation pre-
ceding solved trials, a whole-brain analysis identified
regions in which the eight participants highest in PA
showed different signal during preparation (as de-
scribed in A) than did the eight participants lowest
in PA. The dACC, ventral ACC (vACC), and PCC all
exceeded significance criteria, t(14) = 3.32, p = .005,
v > 500 mm3, all showing stronger preparatory ac-
tivation for subjects high in PA than for participants
low in PA.
The functional overlap, illustrated in a convergence
map, between all the three analyses occurred only with-
in the dACC at ((cid:2)2, 42, 22). The analysis with the least
stringent significance threshold corresponded to a p <
.005, combined with a cluster size of at least 500 mm3.
Thus, the functional overlap between all three analyses,
manifesting activation only within the dACC, suggests a
much lower probability of a type I error.
In a final set of analyses, we examined whether insight
effects (stronger peak signal for insight than for nonin-
sight trials, across all 27 participants) occurred in any of
the ROIs defined by the positive mood preparatory ef-
fect (item C). We contrasted peak fMRI signal for insight
versus noninsight preparation periods (defined above)
as well as insight versus noninsight solutions (at the TRs
corresponding to the last 2 sec of processing prior to
solutions). Within these ROIs, consistently stronger sig-
nal for insight than for analytic preparatory events oc-
curred only within the dACC. Similarly, within these
mood-sensitive ROIs, stronger signal for insight than
for analytic solutions occurred only in the dACC. None
of the mood-sensitive ROIs showed stronger signal for
analytic than insight trials at preparation or solution. In-
sight versus analytic signal was not enhanced by positive
mood at any other time points (all p > .2).
420
Journal of Cognitive Neuroscience
Volume 21, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
/
.
.
F
T
.
o
N
1
8
M
UN
sì
2
0
2
1
RESULTS
Behavioral Measures
Participants correctly solved 41.0% (SD = 11.4) del
problems and identified 50.8% (SD = 16.3) of their
solutions as insight (mean response time = 6.57 sec,
SD = 1.31) E 46.8% (SD = 16.2) of their solutions as
analytic/without insight (mean response time = 7.35 sec,
SD = 1.23), reliably slower than the insight responses
[T(78) = 3.60, P << .001]. Of trials with responses,
3.96% (SD = 2.52) were errors.
We examined how affect, assessed by a variety of state,
trait, and personality questionnaires, related to problem-
solving behavior. The range of scores on the affective
scales was somewhat limited. In particular, only 5 of
79 participants had a score higher than 20 on the NA
scale, which ranges from 10 to 50. However, some par-
ticipants had a high score on both the PA and the NA
scales, consistent with the assertion that the PA and the
NA scales are orthogonal (Watson, Clark, & Tellegen,
1988). How should we compare the mood of a person
scoring high on PA and NA with the mood of a person
scoring high on PA but low on NA? Although results
were as strong (sometimes stronger) if we used strict PA
scores, we took into account NA scores by using PA–NA
as an index of positive mood.
Consistent with prior studies, positive mood modu-
lated solving rates: the top third most positive (PA–NA)
participants (mean PA–NA score = 24.0, SD = 3.77; see
Table 1) solved more problems (mean solved = 60.0;
mean solution response time = 6.66 sec) than did the
bottom third or least positive mood participants (mean
PA–NA score = 5.35; mean solved = 51.3; mean solution
response time = 7.19 sec), t(50) = 2.24, p < .05.
Positive mood was also related to which type of strat-
egy, by self-report, led to solutions. As predicted, the
number of insights differed significantly across the three
levels of positive mood [F(2,76) = 7.364, p = .001]. By
contrast, the number of problems solved analytically,
that is, without insight, did not differ [F(2,76) = 1.485,
p = .233]. Therefore, positive mood specifically facili-
tated insights but did not change the rate of analytical
solutions (Figure 3A). Specifically, the highest positive
mood participants solved more problems with insight
(mean insights = 34.5; mean insight response time =
6.12 sec) than did the lowest positive mood participants
(mean insights = 21.9; mean insight response time =
7.31 sec), t(50) = [3.96], p < .0005. Overall, a regression
analysis (partialing out all other mood and personality
variables) showed that positive mood (PA–NA) was di-
rectly correlated with insight solving [r(77) = .40, p <
.005; Figure 2].
Anxiety had the opposite effect (see Figure 3B) where
the third of participants highest in anxiety (mean STAI
score = 42.1, SD = 3.77) solved fewer problems with
insight (mean insights = 24.1; mean insight response
time = 6.12 sec) than did the third of participants,
t(50) = [2.75], p < .01, lowest in anxiety (mean STAI
score = 24.7; mean insights = 33.1; mean insight re-
sponse time = 7.31 sec), and anxiety was inversely cor-
related with solving with insight [r(77) = (cid:2).34, p < .005;
Figure 2]. However, anxiety did not have a reliable ef-
fect on overall solving rates (top versus bottom third),
t(50) = [1.277], p = .207. Anxiety enhanced the propor-
tion of solutions achieved analytically without insight,
t(50) = [2.189], p = .033, but did not reliably change
the raw number of analytical solutions, t(50) = [1.235],
p = .222.
Imaging Measures
We conducted three analyses to examine the neural
basis of the interaction between positive mood and in-
sight solving. In these analyses, we showed that PA mod-
ulates participants’ preproblem preparatory brain states
to specifically facilitate insight solutions by enhancing sig-
nal within the rostral region of the dACC (see conver-
gence map in Figure 7). These preparatory brain states
were assessed by examining fMRI signal corresponding
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
3
e
-
4
p
1
d
5
f
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
c
5
n
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
5
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
f
.
t
/
.
Table 1. Behavior: Positive Mood Enhances Solving Performance and Solving with Insight while Anxiety Inhibits Solving with
Insight
Out of 135 Problems
All 79 participants
High positive mood participants
Low positive mood participants
High anxious participants
Low anxious participants
Average Solved
Number
Average Insight
Number
Average Noninsight
Number
Solve % Insight % Noninsight %
55.3
60.0*
51.3
52.3
57.1
28.1
34.5***
21.9
24.1**
33.1
25.9
24.9
29.0
27.1
23.5
41.0
50.8
46.8
44.4*
57.5***
41.5***
38.0
38.7
42.3
42.7
46.1**
57.9
56.5
51.8*
41.2
For all 79 participants tested, mean number of overall solutions, solutions with insight, and analytical noninsight solutions are given for each
participant group (n = 26); high versus low positive mood was calculated using PA–NA scores from the PANAS inventory; high- versus low-anxiety
scores from the STAI inventory (*p < .05, **p < .01, ***p < .0005). Solved percentages were calculated out of 135 trials; insight and analytical
percentages were calculated out of the total solved number.
o
n
1
8
M
a
y
2
0
2
1
Subramaniam et al.
421
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
3
e
-
4
p
1
d
5
f
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
c
5
n
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
5
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
.
f
t
/
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Scatterplots for all 79 participants indicating the relation between percent of solutions achieved by insight and (A) positive mood
(PA–NA) and (B) anxiety (STAI), both presented in standardized z scores for illustration purposes, with regression lines and values obtained
from multiple regression including all mood and personality measures.
to the variable 0–8 sec rest between the end of one trial
and the beginning of the next three-word problem while
participants fixate on a centrally located cross and pre-
pare for the next problem (Kounios et al., 2006).
Figure 3. Subgroups of participants by high, medium, and low
(A) positive mood and (B) anxiety scores, illustrating the number
of correct solutions achieved with and without insight.
(A) Do Brain Regions Showing Signal Change
at Preparation Show Mood Effects?
As described in the Methods section, we first identified
ROIs that showed changes in neural activity across all
preparatory periods preceding trials that participants
subsequently solved (Figure 4, Table 2). Across all par-
ticipants, we then examined whether this preparatory
activity correlated with PA, anxiety, solving rates, or
solving strategy (solving with insight or noninsight). This
analysis enabled us to investigate if certain regions that
‘‘turned on’’ at preparation were modulated by positive
mood and anxiety states.
As illustrated by Table 2A, three areas showed in-
creased activation during preparation: dACC, PCC, and
the right AG. In two of these regions, as positive mood
increased across all 27 participants, so did the amount
of preparatory activity: in the ACC [r(25) = .41, p < .05;
see Figure 4C] and in the right AG [r(25) = .40, p < .05].
Preparatory activity in the rostral dACC also inversely
correlated with anxiety, but this correlation was not
statistically reliable [r(25) = (cid:2).34, p = .08; Table 2].
Preparatory activity in the PCC showed a mild but
nonsignificant positive correlation with overall propor-
tion of problems solved [r(25) = .36, p = .06; Table 2],
but no correlation with positive mood.
Hypothetically, deactivations could be equally impor-
tant to increases in activation. So, for completeness,
we performed the same analyses looking at areas that
deactivated during preparation. Two areas showed sys-
tematic deactivation compared with baseline: the left
and the right IFG. This deactivation during preparation
was negatively correlated with the overall proportion of
problem solved [left IFG: r(25) = (cid:2).40, p < .05; right
IFG: r(25) = (cid:2).50, p < .05; Table 2] but did not
422
Journal of Cognitive Neuroscience
Volume 21, Number 3
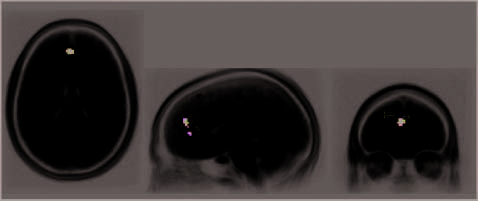
preparatory activity then correlated with mood in re-
gions showing specific insight effects. As mentioned
above, the areas showing overall increased responsivity
at preparation included the dACC, the PCC, and the
right AG. Each of these areas, therefore, represents a
good candidate for preparatory activity predicting over-
all solution-related activity. To examine where prepara-
tory activity predicted overall solution-related activity,
we identified regions that showed solution-related re-
sponsivity, similar to the way we defined preparatory
ROIs as described by item A (Methods section). For in-
stance, we defined solution-related ROIs by subtracting
the mean signal across the three TRs corresponding
to baseline solution-related signal (TRs 1, 6, and 7) from
the mean signal across the three TRs (TRs 3, 4, and 5)
corresponding to peak signal leading up to the solu-
tion (see Figure 9 for comparison). These solution-
related functional ROIs would, therefore,
indicate re-
gions of the brain that ‘‘turned on’’ upon arriving at
solution. We then looked back at preparatory respon-
sivity within these solution-active ROIs. We found that
as preparatory activity increased, so did solution-related
responsivity, within one region only: the region of the
dACC. This analysis demonstrates that preparatory ac-
tivity within the dACC partially predicted overall solving
activity.
(B) Do Brain Regions Showing Insight Specific Activity
at Either Preparation or Solution Correlate with Mood?
We next examined whether preparatory activity corre-
lated with mood in regions identified as showing insight-
specific processing (see item B, Methods section). We
identified ROIs that showed an ‘‘insight effect,’’ that is,
stronger peak signal for insight versus analytical processes,
either during preparation (Kounios et al., 2006) or leading
up to solution (as in Jung-Beeman et al., 2004). Within
these ‘‘insight effect’’ regions, we examined whether over-
all preparatory responsivity (from preparation onset to
peak response and back down to baseline) was modulated
by positive mood states.
Regions that showed this ‘‘insight effect’’ at preparation—
stronger signal during preparation preceding problems
that were eventually solved with insight than during
preparation preceding analytic solutions—included the
ACC, the PCC, and the right and left MTG (Table 2C),
as previously described (Kounios et al., 2006). Within
these ROIs, positive mood correlated (across all 27 par-
ticipants) with preparatory responsivity only in ACC
[r(25) = .40, p < .05; Table 2C]. This preparatory activ-
ity in ACC also inversely correlated with anxiety [r(25) =
(cid:2).40, p < .05]. Moreover, the peak of this preparatory
activity in ACC correlated with the overall proportion of
problems solved [r(25) = .37, p = .05]. Positive mood
did not correlate with preparatory activity observed in
other areas showing insight effects during preparation
Subramaniam et al.
423
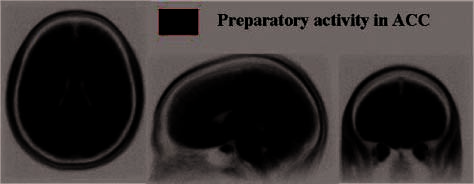
Figure 4. (A) The ROIs within the dACC (see Table 2 for coordinates)
showing strongly increased signal ( p < .0001), across all 27 participants,
corresponding to the preparation interval, superimposed on the
averaged normalized structural image of all participants. Brain images
show (left to right) axial, sagittal, and coronal images (with left
hemisphere on left of axial and coronal images). (B) The average
signal change across this dACC region for the 20 sec after onset of the
preparation interval (which lasted 0–8 sec). (C) Scatterplot illustrating
the correlation between positive mood and increased preparatory
activity in this dACC region (peak–baseline) across all 27 participants.
correlate with any mood variables ( ps > .20). Questo
analysis (item A, Methods section), Perciò, demon-
strates that among the ROIs showing changes in neural
activity at preparation, only the dACC and right AG
increased activation with positive mood.
Does Brain Activity at Preparation Predict Brain
Activity at Solution?
We examined whether preparatory brain activity pre-
dicted overall solution brain activity, and whether this
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
.
T
.
.
F
/
o
N
1
8
M
UN
sì
2
0
2
1
Tavolo 2. Neuroimaging: Positive Mood States Predict Increased Preparatory Activity in ACC to Enhance Solving with Insight
Correlations
Structure
Positive Mood Anxiety Solve %
BA
UN. Preparatory Activity
Center
Coordinates
X
Y
Z
Volume
(mm)
Mean %
Significance
Max %
Significance Mean t Max t
l. IFG
R. IFG
ACC
PCC
R. AG
0.22
0.2
0.41*
0.29
0.40*
(cid:2)0.24 (cid:2)0.40*
9, 6
4375 (cid:2)42
3
(cid:2)0.26 (cid:2)0.50*
6
(cid:2)0.34
(cid:2)0.32
(cid:2)0.22
0.27
0.36
9, 32
31, 23
0.41* 40
3219
1562
6641
6031
47 (cid:2)13
1
47
(cid:2)1 (cid:2)48
47 (cid:2)60
B. High Positive Mood Preparatory State > Low Positive Mood Preparatory State
dACC
vACC
PCC
0.50*
0.44*
0.40*
(cid:2)0.44*
(cid:2)0.47*
0.20
0.21
32, 9
24
3188
0
938 (cid:2)10
36
26
(cid:2)0.34 (cid:2)0.23
31, 30
875
6 (cid:2)44
26
41
13
33
39
21
0
25
C. Insight Preparation > Analytical Preparation
(cid:2)0.28
0.10
31
6641 (cid:2)24 (cid:2)10 (cid:2)10
PCC
ACC
L p.M/STG
0.23
0.40*
0.22
(cid:2)0.40*
0.37* 32
(cid:2)0.41*
0.21
22, 19, 39
R. p.M/STG
(cid:2)0.16
(cid:2)0.15 (cid:2)0.10
39, 37
D. Insight Solution > Analytical Solution
1047
797
562
2156
2047
1984
1984
(cid:2)3
50
46
43
59
69
57 (cid:2)33
2 (cid:2)42
5
1
24
2
34
20 (cid:2)11 (cid:2)13
(cid:2)4
36
3
0.27
0.27
(cid:2)0.38*
(cid:2)0.24
0.02
0.26
(cid:2)0.05
(cid:2)0.01 (cid:2)0.01
21, 22
31
34
0.45*
(cid:2)0.44*
0.37* 24, 32
0.22
0.18
0.24
0.18
(cid:2)0.27
(cid:2)0.16
0.25
0.30
0.08
21
9
40
1516 (cid:2)59 (cid:2)19 (cid:2)5
1234
703
8
51
55 (cid:2)39
28
38
R M/STG
PCC
R. PHC
ACC
l. M/STG
R. SFG
R. IPL
(cid:2)0.07
(cid:2)0.06
0.1
0.05
0.07
(cid:2)0.11
(cid:2)0.1
0.15
0.1
0.1
0.08
0.07
0.05
0.07
0.08
0.08
0.07
0.08
0.08
0.08
0.09
0.05
0.09
0.06
0.12
0.09
0.1
0.09
0.1
0.07
0.07
0.1
0.12
0.12
0.13
0.1
0.11
0.07
6.6
5.5
5.5
3.5
3.4
3.8
4
3.8
3.5
3.5
3.4
3.4
4
4
3.8
3.4
3.4
3.4
3.4
9.1
7
6.5
4.4
4.2
5.3
6.2
4.6
4.4
4.4
4.2
4.2
4.8
4.9
5.6
4.1
4
4.2
4.1
Each value in the correlations section is a correlation value of either positive mood (PA–NA), anxiety (STAI), or overall solving proportion with
activity in the corresponding cluster that represents the signal difference between the contrasted conditions as a percent of average signal within
the cluster (*P < .05). (A) ROIs identifying significant signal change within the three TRs corresponding to the expected peak preparatory signal
(i.e., TRs starting at 6 through 12 sec) compared with the first and last two TRs corresponding to the baseline preparation signal. (B) the positive
mood preparatory ROIs with increased fMRI preparatory activity for the top eight participants highest in positive mood than the bottom eight
participants lowest in positive mood. (C) ROIs with stronger fMRI peak signal for insight preparation than for analytical noninsight preparation. (D)
ROIs with stronger fMRI signal within the three TRs corresponding to the expected peak signal just prior to insight solutions than for analytical
solutions. No clusters showed the opposite effect at this strict threshold.
[PCC: r(25) = .23, p = .27; left MTG: r(25) = .22, p = .27;
right MTG: r(25) = (cid:2).16, p = .42].
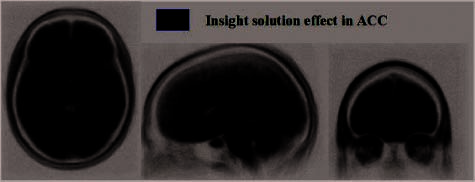
We next examined whether positive mood modulated
preparatory activity in areas that showed an ‘‘insight ef-
fect’’ at solution (see Figure 9). We identified several re-
gions showing insight effects at solution, that is, stronger
signal for insight solutions than for noninsight solutions.
These ROIs included the right aSTG, the ACC, the PCC,
the right PHC, the bilateral MTG (stronger in right than
left), the right SFG, and the right IPL (Table 2D). These
data, with more participants and better imaging protocols,
match well with earlier results showing smaller effects, but
in the same general regions, with right aSTG again show-
ing the largest effect ( Jung-Beeman et al., 2004). Within all
these ROIs showing insight effects at solution, preparatory
activity correlated with positive mood only within ACC
[r(25) = .45, p < .05; see Figure 5, Table 2D]. Again,
ACC preparatory activity negatively correlated with anxiety
424
Journal of Cognitive Neuroscience
Volume 21, Number 3
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
3
e
-
4
p
1
d
5
f
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
c
5
n
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
5
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
f
.
t
.
.
/
o
n
1
8
M
a
y
2
0
2
1
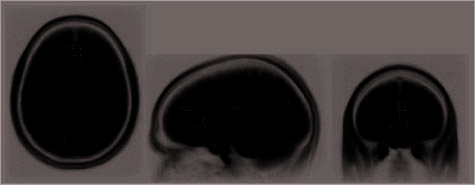
Figure 6. All ROIs showing stronger signal change (peak–baseline)
corresponding to the preparation interval for high positive mood than
for low positive mood participants ( p < .005). Reliable clusters include
dACC and vACC as well as PCC. (No reliable clusters showed the
reverse, that is, stronger signal for low positive mood participants.)
mood than in high positive mood participants. ACC and
PCC showed more preparatory responsivity for the eight
participants highest versus the eight participants lowest
in positive mood. In ACC region showing a mood group
effect across all trials (specifically, the rostral portion
of the dACC; see Figure 6), the preparation signal was
stronger, across all participants, preceding problems
subsequently solved with insight than preceding prob-
lems subsequently solved analytically, t(26) = [2.3],
p = .03 (see Figure 8). In contrast, the PCC region that
showed stronger preparation signal for the high positive
than for the low positive mood participants did not
show any insight effect during preparation (t < 1.0).
We then tested whether these same regions (showing
mood effects during preparation) showed insight effects
leading up to solution. Indeed, across all participants,
there was stronger fMRI signal for insight solutions than
for noninsight solutions in the dACC, t(26) = [3.97],
p < .0005 (see Figure 9), the vACC, t(26) = [3.8], p <
.001, and the PCC, t(26) = [3.8], p < .001. These effects
were not due to making the insight rating at the end of
each trial, as there were no effects within any of these
ROIs on the BOLD signal corresponding to the insight
rating button press (all t values <1.2).
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
3
e
-
4
p
1
d
5
f
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
c
5
n
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
5
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
t
f
.
.
/
o
n
1
8
M
a
y
2
0
2
1
Figure 7. Convergence map showing all voxels within each of
the three types of analyses: Voxels showing reliable signal change
(peak–baseline) corresponding to preparation (blue); voxels showing
both preparation activity and insight solution effects (green); voxels
showing both preparation activity and stronger preparation signal in
high than in low positive mood participants (purple); and voxels
showing all three effects (black).
Subramaniam et al.
425
Figure 5. (A) The ROI within the rostral ACC showing stronger
signal for insight ( p < .001) than for noninsight solutions (as in
Jung-Beeman et al., 2004) across all 27 participants. Brain images
show (left to right) axial, sagittal, and coronal images (with left
hemisphere on left of axial and coronal images). (B) Scatterplot
illustrating the correlation between positive mood and increased
preparatory activity (peak–baseline) in this rostral ACC region
showing an insight solution effect across all 27 participants.
[r(25) = (cid:2).44, p < .05], whereas preparatory peak signal
positively correlated with the overall proportion of prob-
lems solved [r(25) = .37, p = .05].
(C) Are Brain Regions Showing Positive Mood Effects
during Preparation Involved in Solving with Insight?
In the above analyses, we identified ROIs by overall
preparatory responsivity (item A, Methods section) and
by insight effects (item B, Methods section) and then
found that preparatory activity within ACC ROIs specif-
ically consistently correlated with positive mood across
all participants. Analysis C (Methods section) does the
converse, first identifying ROIs that show mood effects
in preparation for all trials, then determining whether
an insight effect (stronger signal prior to insight solu-
tions than prior to noninsight solutions) occurred within
these ROIs. The positive mood preparatory effect in-
dicated which brain regions manifest increased pre-
paratory responsivity, across all trials, for the eight
participants highest in positive mood compared with
the eight participants lowest in positive mood, regard-
less of whether the hemodynamic response demonstrat-
ed a rise and fall of signal (Figure 6).1 For instance, some
areas showed decreasing activity during preparation (left
and right IFG) but more rapid decreases in low positive
Thus, some brain areas—particularly ACC—in which
positive mood modulated activity during the prepara-
tion for upcoming trials do seem especially involved in
processing that leads to insight solutions. The functional
overlap of areas showing both mood and insight effects
is illustrated in a convergence map (Figure 7), which
shows that only the rostral portion of the dACC mani-
fests the mood–insight correspondence in all three anal-
yses described above.
DISCUSSION
Participants higher in positive mood showed different
patterns of brain activity during preparation periods
preceding each solved problem and solved more prob-
lems overall compared with participants lower in posi-
tive mood. The mood-related facilitation in solving was
limited to solving with insight, as high positive mood
participants solved many more problems with insight
and somewhat fewer without insight compared with the
low positive mood participants. The results reported
above used PA–NA scores as an index of positive mood
and are maintained or stronger when using PA alone
as the index of positive mood. In regression analyses
with all mood and personality measures, PA yielded a
nominally stronger correlation with insight percentage
[r(77) = .41, p < .0005] than did PA–NA [r(77) = .40,
p < .0005]. Furthermore, the same pattern of HRF peaks
and group differences were attained if PA was used
rather than PA–NA. However, some subjects scored
high on both PA and NA (consistent with prior litera-
ture claiming PA and NA scores on the PANAS inventory
are orthogonal; e.g., Watson et al., 1988), so it is unclear
whether they should be considered high in positive
mood. Therefore, we decided to consistently use PA
minus NA scores throughout all the analyses.
Interestingly, as positive mood seemed to be increasing
overall solving productivity, as well as shifting the type of
processing employed to specifically facilitate insight solv-
ing, anxiety had somewhat the opposite effect, decreasing
insight solutions, but not affecting solving performance
as reliably or as consistently as positive mood.
The experimental paradigm relies on retrospective
self-report measures to categorize solutions as insight
versus noninsight. It is, thus,
important to note that
positive mood affected not just whether participants re-
ported insight, but also their overall ability to solve prob-
lems (higher positive mood participants actually solved
more of these problems, and all the ‘‘extra’’ solutions
were reported to be with insight). Thus, mood affects
solving behavior.
This trial-by-trial reporting method does not as-
sume participants solve problems with insight based on
a priori categorization of the problems. (In pilot re-
search, participants report that they solved classic in-
sight problems with insight about 65% of the time—with
some analytic and ‘‘other’’ solutions—and report that
they solve classic analytic problems with insight about
25% of the time). Even if we did rely on relatively more
‘‘objective’’ measures such as GSR measures or warmth
ratings, we would still have more confidence in self-
report measures. For instance, if a subject reports to
have had an insight but shows gradual continuous
changes in warmth ratings as he or she progresses
toward the solution rather than the sudden discontinu-
ous jump associated with insights upon reaching the
solution, we would still have more trust in the subject’s
self-report assessment rather than warmth ratings.
Moreover, in prior studies, participants manifest dif-
ferent patterns of behavior and neural activity when they
report solving (or recognizing solutions) with insight
compared with when they report solving without in-
sight. For example, recognizing solutions with insight
occurs faster and with more priming of solutions (sug-
gesting semantic activation of the solution prior to solv-
ing) than recognizing solutions without insight (e.g.,
Bowden & Jung-Beeman, 2003a).
Within the current study, the different solution cate-
gories were associated with qualitatively distinct patterns
of brain activation preceding solution (see Figure 9),
including differently shaped hemodynamic response
functions; yet there were no consistent differences at
the point of insight judgments. This suggests that the
decisions were based on some differences in prior pro-
cessing leading up to solutions rather than post hoc
decisions.
Also, both the high and the low positive mood groups
showed identical solution latency patterns (in this exper-
iment, slightly faster insight than noninsight solutions)
and parallel hemodynamic responses in fMRI signal with-
in each category (insight vs. noninsight), suggesting that
high and low positive mood participants used roughly
the same processes and decision-making criteria for iden-
tifying insight and noninsight solutions.
Besides affecting behavior, positive mood also corre-
lated with brain activity as people prepared for each new
problem (in the task-free preparation interval). Specifi-
cally, we examined brain regions that changed activity
during this preparation period, regions that showed
insight effects (more activity during insight than non-
insight trials) during this preparation period, and re-
gions that showed insight effects at solution. Across all
these analyses, only dACC consistently showed brain
activity during this (resting) preparation interval that
increased as positive mood increased (see Table 2, Fig-
ure 7). The corollary was also true: ACC region that was
more responsive (showed greater increase of fMRI signal
corresponding to the preparatory period) in highly pos-
itive than in less positive participants also showed in-
sight effects across all participants (Figures 8 and 9). All
these affect-related effects occurred despite a somewhat
limited range of variability in affect (particularly in terms
of NA).
426
Journal of Cognitive Neuroscience
Volume 21, Number 3
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
3
e
-
4
p
1
d
5
f
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
c
5
n
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
5
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
f
.
.
/
.
t
o
n
1
8
M
a
y
2
0
2
1
Figure 8. Insight preparation
effect in mood-sensitive ROI:
average percent signal change
over time, corresponding to
the preparation interval across
all voxels in the ROI showing a
reliable mood effect (stronger
preparation activity in the
high than in the low positive
mood participants). The blue
line shows signal change for
preparation prior to problems
solved with insight; the pink
line shows preparation signal
prior to problems solved
without insight; and the green
line shows the difference,
which was near-constant
throughout the epoch. The
green-shaded region (i.e., TRs
starting at 6 sec through TR
ending at 12 sec), showing stronger signal for insight versus noninsight preparation (*p < .05), corresponds to the peak signal for the
preparation period. For comparison, the preceding button press elicited peak signal in the motor cortex (M1) at 4 sec, which corresponded
to the insight rating button press from the prior trial.
Thus, we have strongly demonstrated that positive
mood is reliably associated with preparatory states that
increase responsivity in the rostral dACC, and that this
modulation is associated with processing that leads to
insight solutions. We are not arguing that the activation
in ACC represents a neural correlate of positive mood
or that positive mood states induce insight. We are con-
cluding that positive mood is one factor that enhances
activity in the rostral dACC, and that this mediates the
shift toward insight solutions.
The precise mechanism by which positive mood
facilitates insights through correspondingly modulating
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
3
e
-
4
p
1
d
5
f
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
c
5
n
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
5
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
f
/
t
.
.
o
n
1
8
M
a
y
2
0
2
1
Subramaniam et al.
427
Figure 9. Insight solution
effect in mood-sensitive ROI:
average percent signal change
over time, corresponding to
the solution interval (i.e., 0 sec
corresponds to a solution
event 2 sec prior to the
solution button press) across
all voxels in the ROI showing
a reliable mood effect at
preparation. The blue line
shows signal change prior to
problems solved with insight;
the pink line shows signal
change prior to problems
solved without insight; and
the green line shows the
difference, which was
near-constant throughout
the epoch. The green-shaded
region (TR starting at
4 sec through TR ending at
10 sec) showing greater
solution-related signal (*p <
.0005) for insight versus
noninsight trials corresponds
to the peak signal leading
up to the solution. For
comparison, the subsequent
solution button press elicited
peak signal in the motor
cortex (M1) at 10 sec (i.e.,
8 sec after the button press).
cognitive control processes within ACC is not entirely
obvious. Cognitive control is itself a multifaceted con-
cept,
involving the recruitment of frontal regions—
including the dACC, but also DLPFC, particularly in the
LH—implicated in the detection of competing re-
sponses, overcoming prepotent response tendencies,
and switching attention to select the correct response
(Hedden & Gabrieli, 2006; Kondo et al., 2004; Weissman,
Warner, & Woldorff, 2004; Weissman et al., 2003; Carter
et al., 2000). ACC, specifically, has been implicated in
several processes, such as error detection (Carter et al.,
1998) or conflict monitoring (Botvinick et al., 2004;
Kerns et al., 2004; Weissman et al., 2003).
We did not examine conflict monitoring in our study
per se, and this study was not designed to tease apart
the exact role of ACC in cognitive control. However, we
favor a view by which ACC is involved in monitoring not
just conflict but a variety of competing responses, such
as multiple associations or strategies involved in solv-
ing problems. One way of putting it is that ACC sets a
parameter of detecting such competing activations that
allows either task shielding (ignoring other stimuli or
thoughts to remain focused) or task switching (detect-
ing competing stimuli, so that other components of cog-
nitive control networks can switch attention to them;
see Dreisbach & Goschke, 2004). One mechanism by
which PA facilitates insight is by increasing this parameter
for detecting multiple competing associations, which
provides the solver a better chance of suddenly switch-
ing attention to the correct solution (or to solution-
related information), thus facilitating insights. In line with
our ‘‘competing activation’’ hypothesis, we think that
PA enhances insights by possibly enhancing the detection
of semantic associations (Rowe et al., 2007) facilitating
shorter solution RTs, which would also partly explain why
insight trials tended to be slightly faster than noninsight
trials. In contrast, if insights only involved greater conflict
monitoring, we would predict longer RTs for insight ver-
sus noninsight trials in our task.
PA previously has been linked to modulation of
cognitive control processes to enhance cognitive flexi-
bility, at the expense of perseveration or maintained
focus (Dreisbach & Goschke, 2004). Further, prior the-
oretical explanations have attributed increases in cogni-
tive flexibility to the effect of PA at enhancing phasic
dopaminergic activity in the ACC and the pFC (Ashby,
Valentin, & Turken, 2002; Ashby et al., 1999), consistent
with other models of dopamine’s effect on cognitive con-
trol (e.g., Daw, O’Doherty, Dayan, Seymour, & Dolan,
2006; Braver, Barch, & Cohen, 1999).
When people encounter a problem to solve (or any
input to understand), they frequently engage multiple
possible solving mechanisms. However, under various
circumstances, different mechanisms are favored—due
to individual states or traits or due to the problem itself
(which is why some problems are more likely to be solved
with insight and others more analytically; Bowden et al.,
2005; Ansburg & Hill, 2003; Oelling & Knoblich, 2003). PA
likely shifts the balance of which mechanisms will be most
effective. As noted in the introduction, solving problems
with insight requires cognitive flexibility (hence cognitive
control) because it benefits from ‘‘cognitive restructur-
ing’’ of the problem, enabling the solver to pursue a new
strategy or a new set of associations. Several putative
mechanisms could explain (in whole or in part) how PA
enhances such flexibility. It may alter the selection pro-
cess through which information enters working memory
(Ashby et al., 1999, 2002); it may tip the balance toward
a more global focus of attention (Gasper & Clore, 2002)
or a broader attention to both external visual space
and internal conceptual space (Rowe et al., 2007) allow-
ing more problem elements to simultaneously influence
solution efforts; and it may facilitate switching between
different modes of attention (Baumann & Kuhl, 2005;
Kondo et al., 2004) or switching from irrelevant to rel-
evant solving strategies (Dreisbach & Goschke, 2004).
These putative mechanisms may overlap or may work in
combination. The bottom line is that solvers appear to
be better able to switch from pursuing a dominant but
errant set of associations to a solution-relevant set.
Note that such a proposal does not mean that PA
facilitates solutions by directly enhancing access to a
broader range of semantic associations, for example, by
increasing RH semantic processing. Recall that another
hypothetical mechanism by which positive mood could
facilitate insight would be through enhanced RH pro-
cessing, given the demonstrated importance of RH se-
mantic processing for processing a broad set of semantic
associations (Chiarello, 1998; Beeman et al., 1994) gen-
erally and for insight solutions specifically ( Jung-Beeman
et al., 2004; Bowden & Beeman, 1998). Several pieces
of evidence suggest that PA could enhance relative RH
activation. First, PA increases sensitivity to a larger range
of semantic associations (Fredrickson & Branigan, 2005;
Federmeier et al., 2001), which, as noted, is character-
istic of RH semantic processing. Second, induced posi-
tive mood increases a global focus of attention (Gasper
& Clore, 2002), which is usually associated with RH vi-
sual attention, whereas a local focus of attention is asso-
ciated with LH processing. Third, inducing an approach
regulatory focus (which is often associated with PA)
enhances both overall RH activation, as measured by
a line-bisection task, and creativity (e.g., Friedman &
Fo¨rster, 2005). Finally, compared with people who
solve anagrams analytically, people who solve with in-
sight show increased brain activity at rest in mostly right-
lateralized regions according to resting-state EEG (Kounios
et al., 2008). However, a great deal of research using
frontal asymmetries during resting-state EEG associates
LH activity with PA or approach regulatory focus (e.g.,
Sutton & Davidson, 1997). Further, effects that shift pro-
cessing toward biases that are associated with one or the
other hemisphere could occur due to modulation of
medial attention or cognitive control-related processes.
428
Journal of Cognitive Neuroscience
Volume 21, Number 3
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
1
l
3
e
-
4
p
1
d
5
f
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
c
5
n
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
p
c
d
n
.
b
2
y
0
g
0
u
9
e
.
s
t
2
o
1
n
0
5
0
7
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
e
0
2
s
3
t
/
j
.
/
.
f
t
.
o
n
1
8
M
a
y
2
0
2
1
Regardless, in the current experiment, PA did corre-
late with signal change during the preparation period
in one lateral (rather than midline) cortical region, the
AG of the RH; however, this area did not show other
mood-related effects nor did it show an ‘‘insight effect’’
(stronger activity for insight than noninsight trials) at
either solution or preparation period. Rather than sim-
ply increasing RH semantic processing, it appears that
PA heightens solvers’ sensitivity to solution-relevant pro-
cessing, which may often occur within the RH semantic
processing network (Jung-Beeman, 2005), working in co-
operation with cognitive control processes in the frontal
cortex to make the switch to converge to the correct
solution. Still, it remains possible that a wider range of
assessed (or induced) PA would reveal enhanced RH
relative activation associated with a high positive mood.
There are several potential alternative explanations
that can be considered and rejected. First, one might
wonder whether positive mood did not alter the pro-
cessing that led to solution but instead simply affected
participants’ willingness to label a solution as ‘‘insight.’’
This is unlikely, as we mentioned earlier, because par-
ticipants higher in positive mood actually solved more
problems than participants lower in positive mood—
they solved more with insight and almost equally as
many without insight as the lower positive mood group.
Moreover, the high and low positive mood subgroups
showed similar solution reaction times for insight versus
noninsight solutions (for both groups, slightly faster
insight than noninsight solutions). Furthermore, both
subgroups showed nearly identical hemodynamic re-
sponses for insight solutions and likewise for noninsight
solutions; that is, the solution types differed but the
groups did not, suggesting that both groups used the
same processes for solutions they labeled as insight.
Given that insight solutions were (in this study) faster
than noninsight solutions, the possibility arises that
participants higher in positive mood were more likely
to adopt simpler decision heuristics before responding
that they achieved solution. For instance, positive mood
has been suggested to the use of ‘‘satisficing’’ rather
than optimizing solving strategy (Kaufmann & Vosburg,
1997) or even suggested to be related to reduced overall
cognitive capacity (Mackie & Worth, 1989). However,
such a strategy should lead to more premature and in-
correct responses, that is, trials on which participants
press the button indicating solution, but then give an
incorrect response. Yet high and low positive mood
participants gave equally few incorrect responses ( p >
.20); Infatti, in other studies, participants who demon-
strate a preference to solve without insight are more
likely to make incorrect responses (Kounios et al., 2008).
Another possibility to consider is that PA enhances all
neural activity (or perhaps enhances hemodynamic re-
sponse, such as caffeine does), and that the PA-associated
enhancements during preparation only occur in ACC be-
cause that is the primary area showing increased signal
during that epoch. Tuttavia, we observed no PA-related
enhancement of signal change in brain areas showing
large responses corresponding to either problem onset
or solution (per esempio., the insight effect in right aSTG was no
bigger in high positive mood than in low positive mood
participants).
Given that the ‘‘Aha!’’ experience has an affective
component, we also considered the possibility that
differences during the preparation period were rem-
nants of activity from the preceding trial. Immediately
before the preparation period, participants made their
insight versus noninsight rating of the prior trial (if it was
solved). Tuttavia, hemodynamic responses directly re-
lated to these ratings did not differ depending on the
type of rating made (no reliable clusters of activation
were observed). The enhanced activation of dACC also
did not relate to whether the prior trial was solved at all,
so it was not a form of increased attention in response to
failure or error evaluation on the prior trial (Bush et al.,
2000).
The difference between insight preparation and non-
insight preparation cannot be attributed to simple lack
of attention because we analyzed only preparation peri-
ods preceding problems that were solved, not solved
versus unsolved problems. Inoltre, the mood-related
difference in preparation activity within the dACC was
not attributable to increased arousal (Critchley, Tang,
Glaser, Butterworth, & Dolan, 2005) because if anything,
it was inversely related to anxiety. If increased arousal
drove the effect, then it should be stronger in high-
rather than low-anxiety participants. Infatti, given the
inverse relation between positive mood and anxiety, it is
possible that some effects discussed here could be
attributable to lack of anxiety (Beversdorf, White,
Chever, Hughes, & Bornstein, 2002; Beversdorf, Hughes,
Steinberg, Lewis, & Heilman, 1999) rather than presence
of positive mood. Tuttavia, all behavioral and neuro-
imaging measures correlated more consistently with
increasing positive mood than with decreasing anxiety,
whereas few of the effects correlated with the anxiety
measure. Further, the effects of PA have been shown to
be distinct from ‘‘affectless arousal’’ (Isen et al., 1987). If
anything, arousal is thought to impede creativity, facili-
tating a narrow range of attention and perseveration on
the prepotent response, thereby inhibiting overall cog-
nitive flexibility (Kischka et al., 1996; Martindale, 1995;
Easterbrook, 1959).
Finalmente, others have noted increased activation during
what they term the default state of attention in MPFC
(including dACC) and PCC (Raichle et al., 2001). It is at
least possible that mood-associated changes in ACC in
the current study reflect modulation of a default state
rete. Tuttavia, we have no assessment of such de-
fault activation in the current study, so it would be a leap
to make solid claims one way or the other.
Whether default state or task-related preparation,
positive mood enhances activity within dACC in a manner
Subramaniam et al.
429
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
.
/
.
F
T
.
o
N
1
8
M
UN
sì
2
0
2
1
conducive to solving with insight. This modulation may
promote a more global (Gasper & Clore, 2002) or diffuse
focus of attention, which has previously been linked to
improved insight or creative problem solving (Rowe et al.,
2007; Ansburg & Hill, 2003). Così, we believe that one
mechanism by which positive mood facilitates the shift
toward an insight is by modulating ACC activity, at both
the preparation and the solution time periods,
in un
manner that enhances the detection of multiple compet-
ing associations. Therefore, a solver focused on an incor-
rect association (or solution path) is better ‘‘prepared’’ to
detect and to switch attention to the correct association;
if this attention suddenly brings the correct solution into
awareness, the solver experiences an ‘‘Aha!’’
Conclusions
We examined the relation between various mood states
(including positive mood and anxiety) and personality
measures, assessed prior to the experiment, and brain
activity immediately preceding and during problem solv-
ing. We found that positive mood enhanced overall
solving for these insight-like verbal problems and par-
ticularly increased the likelihood of solving them with
insight. We demonstrated that these effects were related
to brain activity in ACC during the preparation interval
prior to each trial. Specifically, activity increased in ACC
more for high positive than for low positive mood
participants. ACC was the only region showing sensitivity
to multiple measures of this mood–insight association,
providing strong evidence that positive mood states
alter preparatory activity in ACC biasing participants to
engage in problem processing that is conducive to
solving with insight. These results have important im-
plications for neural accounts of both general analytic
problem solving and creative insight solving. Previous
research has demonstrated that positive mood broadens
the scope of attention to both external visual space and
internal conceptual space (Rowe et al., 2007). IL
current work illustrates a neural basis for this modula-
tion of problem solving by positive mood. Further, Esso
suggests that positive mood enhances insight and crea-
tive problem solving, at least in part, by modulating at-
tentional and cognitive control mechanisms within ACC
to allow more sensitivity to detect competing solution
candidates.
Ringraziamenti
This research was supported in part by NIH/NIDCD through
grants R01 DC-04052 to Mark Jung-Beeman and R01 DC-04818
to John Kounios. We thank Edward Bowden, Paul Reber,
Nondas Loudas, Jason Haberman, Zoe Clancy, Joel Voss, E
John Rudoy for their assistance and input on this project.
Brain Mapping Group, Northwestern University, 2029 Sheridan
Road, Evanston, IL 60208-2710, or via e-mail: k-subramaniam@
northwestern.edu; mjungbee@northwestern.edu.
Note
1. Although the top third of participants would technically
be nine participants, matching PA scores made it impossible
to use more than eight participants on either end of the
distribution.
REFERENCES
Amabile, T. M., Barsade, S. G., Mueller, J. S., & Staw, B. M.
(2005). Affect and creativity at work. Administrative Science
Trimestrale, 50, 367–403.
Ansburg, P. I., & Hill, K. (2003). Creative and analytic thinkers
differ in their use of attentional resources. Personality and
Individual Differences, 34, 1141–1152.
Ashby, F. G., Isen, UN. M., & Turken, U. (1999). UN
neuropsychological theory of positive affect and its influence
on cognition. Psychological Review, 106, 529–550.
Ashby, F. G., Valentin, V. V., & Turken, M. (2002). The effects of
positive affect and arousal on working memory and
executive attention: Neurobiology and computational
models. In S. Moore & M. Oaksford (Eds.), Emotional
cognition: From brain to behaviour (pag. 245–287).
Amsterdam: John Benjamins.
Badre, D., & Wagner, UN. D. (2004). Selection, integration,
and conflict monitoring: Assessing the nature and
generality of prefrontal control mechanisms. Neuron, 41,
473–487.
Baumann, N., & Kuhl, J. (2005). Positive affect and flexibility:
Overcoming the precedence of global over local processing
of visual information. Motivation and Emotion, 29,
123–134.
Beeman, M. J., & Bowden, E. M. (2000). The right hemisphere
maintains solution-related activation for yet-to-be solved
insight problems. Memory & Cognition, 28, 1231–1241.
Beeman, M. J., Friedman, R. B., Grafman, J., Perez, E.,
Diamond, S., & Lindsay, M. B. (1994). Summation priming
and coarse semantic coding in the right hemisphere.
Journal of Cognitive Neuroscience, 6, 26–45.
Beversdorf, D. Q., Hughes, J. D., Steinberg, B. A., Lewis, l. D.,
& Heilman, K. M. (1999). Noradrenergic modulation of
cognitive flexibility in problem solving. NeuroReport, 10,
2763–2767.
Beversdorf, D. Q., White, D. M., Chever, D. C., Hughes, J. D., &
Bornstein, R. UN. (2002). Central B-adrenergic modulation of
cognitive flexibility. NeuroReport, 13, 2505–2507.
Bishop, S., Duncan, H., Brett, M., & Lawrence, UN. D. (2004).
Prefrontal cortical function and anxiety: Controlling
attention to threat-related stimuli. Nature Neuroscience,
7, 184–188.
Bolte, A., Goschke, T., & Kuhl, J. (2003). Emotion and
intuition: Effects of positive and negative mood on implicit
judgments of semantic coherence. Psychological Science,
14, 416–421.
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004). Conflict
monitoring and anterior cingulate cortex: An update. Trends
in Cognitive Sciences, 8, 539–546.
Bowden, E. M., & Beeman, M. J. (1998). Getting the right idea:
RH contributions to solving insight problems. Psicologico
Scienza, 9, 435–440.
Bowden, E. M., & Jung-Beeman, M. (2003UN). Aha! Insight
Reprint requests should be sent to Karuna Subramaniam or
Mark Jung Beeman, Department of Psychology and Cognitive
experience correlates with solution activation in the RH.
Psychonomic Bulletin and Review, 10, 730–737.
430
Journal of Cognitive Neuroscience
Volume 21, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
F
.
.
.
T
/
o
N
1
8
M
UN
sì
2
0
2
1
Bowden, E. M., & Jung-Beeman, M. (2003B). Normative data
for 144 compound remote associate problems. Behavior
Research Methods, Instruments, & Computers, 35, 634–639.
Bowden, E. M., Jung-Beeman, M., Fleck, J., & Kounios, J.
(2005). New approaches to demystifying insight. Trend in
Cognitive Sciences, 9, 322–328.
Braver, T. S., Barch, D. M., & Cohen, J. D. (1999). Cognition
and control in schizophrenia: A computational model of
dopamine and prefrontal function. Biological Psychiatry,
46, 312–328.
Bush, G., Luu, P., & Posner, M. (2000). Cognitive and
emotional influence in anterior cingulate cortex. Trends in
Cognitive Sciences, 4, 215–222.
Carnevale, P. J. D., & Isen, UN. M. (1986). The influence of
positive affect and visual access on the discovery of
integrative solutions in bilateral negotiation. Organizational
Behavior and Human Decision Processes, 37, 1–13.
Carter, C. S., Braver, T. S., Barch, D. M., Botvinick, M. M., Noll,
D., & Cohen, J. D. (1998). Anterior cingulate cortex, error
detection and the online monitoring of performance.
Scienza, 280, 747–749.
Carter, C. S., McDonald, UN. M., Botvinick, M., Ross, l. L.,
Stenger, V. A., Noll, D., et al. (2000). Parsing executive
processes: Strategic vs. evaluative functions of the anterior
cingulate cortex. Proceedings of the National Academy of
Scienze, U.S.A., 97, 1944–1948.
Chiarello, C. (1991). Interpretation of word meanings in the
cerebral hemispheres: One is not enough. In P. J.
Schwanenflugel (Ed.), The psychology of word meanings
(pag. 251–275). Hillsdale, NJ: Erlbaum.
Chiarello, C. (1998). On codes of meaning and meaning of
codes: Semantic access and retrieval with and between
hemispheres. In M. J. Beeman & C. Chiarello (Eds.), Right
hemisphere language comprehension: Perspectives from
cognitive neuroscience (pag. 141–160). Mahwah, NJ:
Erlbaum.
Collins, D. L., Neelin, P., Peters, T. M., & Evans, UN. C. (1994).
Automatic 3D intersubject registration of MR volumetric data
in standardized Talairach space. Journal of Computer
Assisted Tomography, 18, 192–205.
Critchley, H. D., Tang, J., Glaser, D., Butterworth, B., & Dolan,
R. J. (2005). Anterior cingulate activity during error and
autonomic response. Neuroimage, 27, 885–895.
Davidson, R. J. (1992). Emotion and affective style:
Hemispheric substrates. Psychological Science, 3, 39–43.
Davis, K. D., Taylor, K. S., Hutchison, W. D., Dostrovsky, J. O.,
McAndrews, M. P., Richter, E. O., et al. (2005). Umano
anterior cingulate cortex neurons encode cognitive and
emotional demands. Journal of Neuroscience, 25,
8402–8406.
Daw, N. D., O’Doherty, J. P., Dayan, P., Seymour, B., & Dolan,
R. J. (2006). Cortical substrates for exploratory decisions in
humans. Letters to Nature, 44, 876–879.
Dreher, J. C., & Grafman, J. (2003). Dissociating the roles of the
rostral anterior cingulate and the lateral prefrontal cortices
in performing two tasks simultaneously or successively.
Cerebral Cortex, 13, 329–339.
Dreisbach, G., & Goschke, T. (2004). How PA modulates
cognitive control: Reduced perseveration at the cost of
increased distractibility. Journal of Experimental
Psychology: Apprendimento, Memory, and Cognition, 30, 343–353.
Drevets, W. C., & Raichle, M. E. (1998). Reciprocal suppression
of regional cerebral blood flow during emotional versus
higher cognitive processes; implications for interaction
between emotion and cognition. Cognitive Emotion, 12,
353–385.
Duncker, K. (1945). On problem-solving. Psicologico
Monographs, 58 (5, Whole No. 270).
Easterbrook, J. UN. (1959). The effect of emotion on cue
utilization and the organization of behavior. Psicologico
Review, 66, 183–201.
Estrada, C. A., Isen, UN. M., & Young, M. (1997). PA facilitates
integration of information and decreases anchoring in
reasoning among physicians. Organizational and Human
Decision Processes, 72, 117–135.
Estrada, C. A., Young, M., & Isen, UN. M. (1994). Positive affect
influences creative problems solving and reported source of
practice satisfaction in physicians. Motivation & Emotion,
18, 285–299.
Faust, M., & Mashal, N. (2007). The role of the right cerebral
hemisphere in processing novel metaphoric expressions
taken from poetry: A divided visual field study.
Neuropsychologia, 45, 860–870.
Federmeier, K. D., Kirson, D. A., Moreno, E. M., & Kutas, M.
(2001). Effects of transient, mild mood states on semantic
memory organization and use: An event-related potential
investigation in humans. Neuroscience Letters, 305, 149–152.
Fichtenholtz, H. M., Dean, H. L., Dillon, D. G., Yamasaki, H.,
McCarthy, G., & LaBar, K. S. (2004). Emotion–attention
network interactions during a visual oddball task. Cognitive
Brain Research, 20, 67–80.
Fredrickson, B., & Branigan, C. (2005). Positive emotions
broaden the scope of attention and thought-action
repertoires. Cognition & Emotion, 19, 313–332.
Friedman, R., Fishbein, A., Fo¨rster, J., & Werth, l. (2003).
Attentional priming effects on creativity. Creativity Research
Journal, 15, 277–286.
Friedman, R., & Fo¨rster, J. (2005). Effects of motivational cues
on perceptual asymmetry: Implications for creativity and
analytical problem solving. Journal of Personality and
Social Psychology, 88, 263–275.
Gasper, K., & Clore, G. l. (2002). Attending to the big picture:
Mood and global versus local processing of visual
informazione. Psychological Science, 13, 34–40.
Gilhooly, K. J., & Murphy, P. (2005). Differentiating insight from
noninsight problems. Thinking and Reasoning, 11, 279–302.
Hedden, T., & Gabrieli, J. D. E. (2006). The ebb and flow of
attention in the human brain. Nature Neuroscience, 9, 863–865.
Herrington, J. D., Mohanty, A., Koven, N. S., Fisher, J. E., &
Stewart, J. l. (2005). Emotion modulated performance and
activity in left dorsolateral prefrontal cortex. Emotion, 5,
200–207.
Isen, UN. M. (1999UN). Positive affect. In T. Dalgleish & M. J. Energia
(Eds.), Handbook of cognition and emotion, Chapter 25
(pag. 521–539). New York: John Wiley & Sons.
Isen, UN. M. (1999B). On the relationship between affect and
creative problem solving. In S. W. Russ (Ed.), Affect, creative
experience and psychological adjustment (pag. 3–17).
Philadelphia: Brunner/Mazel.
Isen, UN. M., & Daubman, K. UN. (1984). The influence of affect
on categorization. Journal of Personality and Social
Psychology, 47, 1206–1217.
Isen, UN. M., Daubman, K. A., & Nowicki, G. P. (1987). PAPÀ
facilitates creative problem solving. Journal of Personality
and Social Psychology, 32, 112–1131.
Isen, UN. M., Johnson, M. M., Mertz, E., & Robinson, G. F.
(1985). The influence of PA on the unusualness of word
associations. Journal of Personality and Social Psychology,
48, 1413–1426.
Jung-Beeman, M. (2005). Bilateral brain processes for
comprehending natural language. Trends in Cognitive
Scienza, 9, 512–518.
Jung-Beeman, M., Bowden, E. M., Haberman, J., Frymiare, J. L.,
Arambel-Liu, S., Greenblatt, R., et al. (2004). Neural activity
observed in people solving verbal problems with insight.
Public Library of Science. Biology, 2, 500–510.
Subramaniam et al.
431
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
F
/
T
T
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
F
/
.
.
.
T
o
N
1
8
M
UN
sì
2
0
2
1
Kaufmann, G., & Vosburg, S. K. (1997). Paradoxical mood
effects on creative problem-solving. Cognition and
Emotion, 11, 151–170.
Kerns, J. G., Cohen, J. D., MacDonald, UN. W., III, Cho, R. Y.,
Stenger, V. A., & Carter, C. S. (2004). Anterior cingulate,
conflict monitoring and adjustments in control. Scienza,
303, 1023–1026.
Kischka, U., Kammer, T. H., Maier, S., Weisbrod, M., Thimm,
M., & Spitzer, M. (1996). Dopaminergic modulation of
semantic network activation. Neuropsychologia, 34,
1107–1113.
Kohler, W. (1925). Intelligenzenpru¨fungen an Anthropoiden
[The mentality of apes]. London: Routledge & Kegan Paul
(Original work published 1917).
Kondo, H., Osaka, N., & Osaka, M. (2004). Cooperation of the
anterior cingulate and dorsolateral prefrontal cortex for
attention shifting. Neuroimage, 23, 670–679.
Kounios, J., Fleck, J. L., Verde, D. L., Payne, L., Frymiare, J. L.,
Abrigo, E., et al. (2008). Neural correlates of spontaneous
thought predict problem solving by sudden insight.
Neuropsychologia, 46, 281–291.
Kounios, J., Frymiare, J. L., Bowden, E. M., Fleck, J. I.,
Subramaniam, K., Parrish, T. B., et al. (2006). The prepared
mente: Neural activity prior to problem presentation predicts
subsequent solution by sudden. Psychological Science, 17,
882–890.
Lane, R. D., Reiman, E. M., Axelrod, B., Yun, L., & Holmes, UN.
(1998). Neural correlate of emotional awareness: Evidence
of an interaction between emotion and attention in the
anterior cingulate cortex. Journal of Cognitive
Neuroscience, 10, 525–535.
MacDonald, UN. W., Cohen, J. D., Stenger, V. A., & Carter, C. S.
(2000). Dissociating the role of the dorsolateral prefrontal
cortex and anterior cingulate cortex in cognitive control.
Scienza, 288, 1835–1838.
Mackie, D. M., & Worth, l. T. (1989). Processing deficits and
the mediation of positive affect in persuasion. Journal of
Personality and Social Psychology, 57, 27–40.
Maier, N. R. F. (1930). Reasoning in humans: IO. On direction.
Journal of Comparative Psychology, 10, 115–143.
Martindale, C. (1995). Biological bases of creativity. In
R. Sternberg (Ed.), Handbook of creativity. Cambridge:
Cambridge University Press.
Mayberg, H. S., Liotti, M., Brannan, S. K., McGinnis, S.,
Mahurin, R. K., Jerabek, P. A., et al. (1999). Reciprocal
limbic-cortical function and negative mood: Converging PET
findings in depression and normal sadness. American
Journal of Psychiatry, 156, 675–682.
Mednick, S. UN. (1962). The associative basis of the creative
processi. Psychic Review, 69, 220–232.
Metcalfe, J. (1986). Premonitions of insight predict impending
error. Journal of Experimental Psychology: Apprendimento,
Memory, and Cognition, 12, 623–634.
Metcalfe, J., & Weibe, D. (1987). Intuition in insight and
noninsight problem solving. Memory and Cognition, 15,
238–246.
Nimchinsky, E. A., Gilissen, E., Allman, J. M., Perl, D. P.,
Erwin, J. M., & Hof, P. R. (1999). A neuronal morphological
type unique to humans and great apes. Proceedings
of the National Academy of Sciences, U.S.A., 96, 5268–5273.
Oelling, M., & Knoblich, G. (2003). Prioritizing the new:
Insights creates set for familiar processes. Amalienstr:
Max-Planck Institute for Psychological Research.
Ogawa, S., Tank, D. W., Menon, R., Ellermann, J., Kim, S.-G.,
Merkle, H., et al. (1992). Intrinsic signal changes
accompanying sensory stimulation: Functional brain
mapping with magnetic resonance imaging. Proceedings of
the National Academy of Sciences, U.S.A., 89, 5951–5955.
Papez, J. W. (1937). A proposed mechanism of emotion.
Archives of Neurology and Psychiatry, 38, 725–743.
Raichle, M. E., MacLeod, UN. M., Snyder, UN. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, J. l. (2001). A default mode of
brain function. Proceedings of the National Academy of
Scienze, U.S.A., 98, 676–682.
Rowe, G., Hirsch, J. B., & Anderson, UN. K. (2007). Positive affect
increases the breadth of attentional selection. Proceedings
of the National Academy of Sciences, U.S.A., 101, 383–388.
Ruff, C. C., Woodward, T. S., Laurens, K. R., & Liddle, P. F.
(2001). The role of the anterior cingulate in conflict
processing: Evidence from reverse Stroop interference.
Neuroimage, 14, 1150–1158.
Schooler, J. W., & Melcher, J. (1995). The ineffability of insight.
In S. M. Smith, T. B. Ward, & R. UN. Finke (Eds.), The creative
cognition approach (pag. 249–268). Cambridge: CON Premere.
Schooler, J. W., Ohlsson, S., & Brooks, K. (1993). Thoughts
beyond words: When language overshadows insight.
Journal of Experimental Psychology: General, 122, 166–183.
Small, D. M., Gitelman, D. R., Gregory, M. D., Nobre, UN. C.,
Parrish, T. B., & Mesulam, M. M. (2003). The posterior
cingulate and medial prefrontal cortex mediate the anticipatory
allocation of spatial attention. Neuroimage, 18, 633–641.
Smith, R. W., & Kounios, J. (1996). Sudden insight: All or none
processing revealed by speed-accuracy decomposition.
Journal of Experimental Psychology: Apprendimento, Memory,
and Cognition, 22, 1443–1462.
Sutton, S. K., & Davidson, R. J. (1997). Prefrontal brain
asymmetry: A biological substrate of the behavioral approach
and inhibition systems. Psychological Science, 8, 204–210.
Talairach, J., & Tournoux, P. (1988). Co-planar stereotaxic
atlas of the human brain. New York: Thieme.
Teasdale, J., Howard, R. J., Cox, S. G., Ha, Y., Brammer, M. J.,
Williams, S. C. R., et al. (1999). Functional MRI study of the
cognitive generation of affect. American Journal of
Psychiatry, 156, 209–215.
Teasdale, J. D., & Fogarty, S. J. (1979). Differential effects of
induced mood on the recall of pleasant and unpleasant
events from episodic memory. Journal of Abnormal
Psychology, 88, 248–257.
Tomarken, UN. J., Davidson, R. J., Wheeler, R. E., & Doss, R. C.
(1992). Individual differences in anterior brain asymmetry
and fundamental dimensions of emotion. Journal of
Personality and Social Psychology, 62, 676–687.
Van Veen, V., Cohen, J. D., Botvinick, M. M., Stenger, V. A., &
Carter, C. S. (2001). Anterior cingulated cortex, conflict
monitoring and levels of processing. Neuroimage, 14,
1302–1308.
Watson, D., Clark, l. A., & Tellegen, UN. (1988). Development
and validation of brief measures of positive and negative
affect: The PANAS scales. Journal of Personality and Social
Psychology, 54, 1063–1070.
Weisberg, R. W. (1994). Genius and madness? UN
quasi-experimental test of the hypothesis that
manic-depression increases creativity. Psicologico
Scienza, 5, 361–367.
Weissman, D. H., Giesbrecht, B., Song, UN. W., Mangun, G. R., &
Woldorff, M. G. (2003). Conflict monitoring in the human
anterior cingulate during selective attention to global and
local object features. Neuroimage, 19, 1361–168.
Weissman, D. H., Warner, l. M., & Woldorff, M. G. (2004). IL
neural mechanisms for minimising cross-modal distraction.
Journal of Neuroscience, 24, 10941–10949.
Whalen, P. J., Bush, G., McNally, R. J., Wilhelm, S., McInerney,
S. C., Jenike, M. A., et al. (1998). The emotional counting
Stroop paradigm: A functional magnetic resonance imaging
probe of the anterior cingulate affective division. Biological
Psychiatry, 44, 1219–1228.
432
Journal of Cognitive Neuroscience
Volume 21, Numero 3
D
o
w
N
l
o
UN
D
e
D
l
l
/
/
/
/
j
T
T
F
/
io
T
.
:
/
/
F
R
o
M
D
H
o
T
w
T
N
P
o
:
UN
/
D
/
e
M
D
io
F
T
R
o
P
M
R
C
H
.
S
P
io
l
D
v
io
e
R
e
R
C
C
T
.
H
M
UN
io
R
e
.
D
tu
C
o
o
M
C
/
N
j
UN
o
R
C
T
io
N
C
/
e
UN
–
P
R
D
T
io
2
C
1
l
3
e
–
4
P
1
D
5
F
1
/
9
2
3
1
7
/
4
3
4
/
3
4
o
1
C
5
N
/
1
2
0
7
0
5
9
9
9
2
7
1
1
0
5
/
7
j
o
P
C
D
N
.
B
2
sì
0
G
0
tu
9
e
.
S
T
2
o
1
N
0
5
0
7
7
.
S
P
e
D
P
F
e
M
B
sì
B
e
G
R
tu
2
e
0
2
S
3
T
/
j
/
F
.
.
T
.
o
N
1
8
M
UN
sì
2
0
2
1