Uncoupling Sensation and Perception in
Human Time Processing
Nicola Binetti1, Alessandro Tomassini2, Karl Friston3, and Sven Bestmann3,4
Abstrait
■ Timing emerges from a hierarchy of computations ranging
from early encoding of physical duration (time sensation) à
abstract time representations (time perception) suitable for stor-
age and decisional processes. Cependant, the neural basis of the per-
ceptual experience of time remains elusive. To address this, nous
dissociate brain activity uniquely related to lower-level sensory
and higher-order perceptual timing operations, using event-
related fMRI. Participants compared subsecond (500 msec) sinu-
soidal gratings drifting with constant velocity (standard) against
two probe stimuli: (1) control gratings drifting at constant velocity
ou (2) accelerating gratings, which induced illusory shortening of
temps. We tested two probe intervals: a 500-msec duration (Short)
and a longer duration required for an accelerating probe to be per-
ceived as long as the standard (Long—individually determined).
On each trial, participants classified the probe as shorter or longer
than the standard. This allowed for comparison of trials with an
“Objective” (physical) or “Subjective” (perceived) difference in
duration, based on participant classifications. Objective duration
revealed responses in bilateral early extrastriate areas, extending
to higher visual areas in the fusiform gyrus (at more lenient
thresholds). Par contre, Subjective duration was reflected by
distributed responses in a cortical/subcortical areas. This com-
prised the left superior frontal gyrus and the left cerebellum,
and a wider set of common timing areas including the BG, pariétal
cortex, and posterior cingulate cortex. These results suggest two
functionally independent timing stages: early extraction of dura-
tion information in sensory cortices and Subjective experience
of duration in a higher-order cortical–subcortical timing areas. ■
INTRODUCTION
Duration estimation is a fundamental ability for successful
interactions with our environment. Evaluating and com-
paring durations of sensory events is thought to require
the interplay between parietal, premotor, cingulate, et
prefrontal cortices and subcortical regions in the BG,
cerebellum, and thalamus (Merchant & Yarrow, 2016;
Coull, Nazarian, & Vidal, 2008). Although research has
shed light on various processes that enable our sense of
duration, such as the extraction and representation of
duration information, or the mnemonic and decisional
processes required for duration comparison and classifica-
tion, the biological basis of the subjective experience of
time remains scarcely understood (Wittmann & Meissner,
2018; Trojano, Caccavale, De Bellis, & Crisci, 2017; Bueti &
Macaluso, 2011; Wittmann, 2009).
Sensory input conveys durational information, with evi-
dence from neural network modeling (Buonomano,
Bramen, & Khodadadifar, 2009), brain stimulation (Salvioni,
Murray, Kalmbach, & Bueti, 2013), and neuroimaging studies
1UCL Interaction Centre, University College London, 2MRC
Cognition and Brain Sciences Unit, University of Cambridge,
3Wellcome Centre for Human Neuroimaging, UCL Queen
Square Institute of Neurology, 4Department of Movement and
Clinical Neurosciences, UCL Queen Square Institute of
Neurologie, University College London
(Bueti, Bahrami, Walsh, & Rees, 2010; Bueti & Macaluso,
2010), suggesting that early striate and extrastriate ( V5/
MT) regions play a key role in temporal encoding and
STM, independent of low-level visual feature processing.
The ability to compare durations of stimuli belonging to
different sensory modalities or to reproduce the duration
of a sensory stimulus with an equivalently timed motor
response implies that duration information can be repre-
sented in a more abstract format. Several models of time
perception propose that time is quantified by means of a
linear accumulation of timing evidence (Ivry & Richardson,
2002; Rosenbaum, 2002; Wing, 2002; Treisman, 1963). Ce
seems compatible with the build-up of activity before an
event, which covaries with stimulus duration, as seen in
nonhuman primate ( Janssen & Shadlen, 2005; Leon &
Shadlen, 2003) and human electrophysiology recordings
(Pfeuty, Ragot, & Pouthas, 2005; Macar & Vidal, 2004;
Pouthas, Garnero, Ferrandez, & Renault, 2000).
En effet, imaging studies reveal multiple regions sensi-
tive to time accumulation when contrasting timing tasks
with equivalent nontiming controls. Par exemple, differ-
ences in visual stimulus duration lead to larger activity in
the anterior portion of the SMA (preSMA) and the ACC
(Pouthas et al., 2005). Bueti and Macaluso (2011) assessed
the relationship between the subjective experience of
time and corresponding neural responses in a more direct
approach for identifying the biological substrates of time
© 2020 Massachusetts Institute of Technology. Published under a
Creative Commons Attribution 4.0 International (CC PAR 4.0) Licence.
Journal des neurosciences cognitives 32:7, pp. 1369–1380
https://doi.org/10.1162/jocn_a_01557
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
7
e
1
–
3
p
6
d
9
F
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
un
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
/
t
F
.
o
n
0
5
M.
un
oui
2
0
2
1
accumulation (Bueti & Macaluso, 2011). The authors
observed that activity in the right putamen, right mid insula,
and right mid/superior temporal cortex correlated with an
illusory lengthening of time, suggesting a distributed corti-
cal and subcortical time accumulation network. These
findings are consistent with recent models proposing that
timekeeping is distributed and carried out in parallel. Pour
example, the striatal beat frequency model suggests that
striatal spiny neurons act as temporal integrators of corti-
cal and thalamic oscillatory clock signals (Buhusi & Meck,
2005; Matell, Meck, & Nicolelis, 2003) across both duration
estimation and duration reproduction tasks and indepen-
dently of sensory modality.
Duration discrimination further relies on working memory
processes to store and compare duration of nonsimulta-
neous stimuli. Lateral premotor and right inferior frontal
cortices have been linked to the mnemonic components
of comparison/reproduction timing tasks (Pouthas et al.,
2005; Schubotz & von Cramon, 2001; Kawashima et al.,
1999). Although typically observed in timing research,
SMA is more generically linked to working memory, typically
recruited in perceptual and motor tasks (Rao, Mayer, &
Harrington, 2001; Kawashima et al., 1999; Rao et al., 1997).
Early auditory (Franssen, Vandierendonck, & Van Hiel,
2006) and visual (Salvioni et al., 2013) processing areas
are involved in maintaining STM traces of temporal infor-
mation in working memory. Enfin, the comparison of
these memory traces must lead to a categorical decision
of which stimulus lasted longer (Lindbergh & Kieffaber,
2013). Studies have linked the cingulate cortex to these
decision-making stages in duration discrimination tasks
(Pouthas et al., 2005; Rao et al., 2001).
Cependant, there is still a limited understanding of what
subset of timing areas uniquely contribute to our subjec-
tive experience of time (Hayashi, van der Zwaag, Bueti, &
Kanai, 2018; Trojano et al., 2017; Wittmann, Van Wassenhove,
Craig, & Paulus, 2010; Wittmann & Paulus, 2008). Ici, nous
address at which stage—within a timing hierarchy—our
perceptual experience of duration (c'est à dire., a duration “percept”)
emerges. To this end, we contrasted activity related to the
low-level encoding of duration features embedded in a sen-
sory signal (sensitive to differences in time—duration sen-
sation) from activity related to the subjective, higher-order
perception of duration (sensitive to perceived differences
in time—duration perception), which inform categorical
duration judgments.
Participants evaluated the duration of sequential drifting
sinusoidal gratings in a classic standard/probe comparison
task. Participants compared subsecond (500 msec) sinu-
soidal gratings drifting with constant velocity (standard)
against two probe stimuli: (1) control gratings drifting at
constant velocity or (2) accelerating gratings, which in-
duced an illusory shortening of time (Bruno, Ayhan, &
Johnston, 2015; Sasaki, Yamamoto, & Miura, 2013; Binetti,
Lecce, & Doricchi, 2012; Matthieu, 2011). The inclusion of
accelerating stimuli dissociated objective and subjective
duration effects. On each trial, participants classified the
probe as lasting shorter or longer than the standard.
This allowed for identifying brain activity uniquely related
to an objective difference in probe duration (Objectively
Long vs. Objectively Short) or to a subjective difference
in probe duration (Subjectively Short vs. Subjectively Long),
based on participants’ duration classifications. Although
there is no dedicated sensor for time, duration is a feature
that characterizes a physical stimulus. We hypothesized
that visual areas, which are sensitive to features embedded
within a sensory signal, should primarily respond to varia-
tions in objective duration (Salvioni et al., 2013; Bueti
et coll., 2010), c'est, reflecting duration sensation. This ex-
tracted information would then inform a conscious per-
cept of time, c'est, duration perception. Objective and
perceived duration, cependant, do not always agree, as evi-
denced by time distortions that highlight the constructive
nature of timing (Eagleman, 2008). We reasoned that the
subjective experience of duration would engage higher-
order areas and in some measure should be less sensitive
to objective differences in duration.
We found two independent (c'est à dire., sans chevauchement) sets,
one sensitive to objective duration and comprising bilat-
eral early extrastriate areas and one sensitive to subjective
duration, including the left superior frontal gyrus and the
left cerebellum, and to a lesser degree the BG, pariétal
cortex, and posterior cingulate cortex. Direct comparisons
of Subjective and Objective activations confirmed a pat-
tern of activation that was consistent with that observed
in the Subjective contrast and not with the Objective con-
trast, thus further supporting the notion of independent
sets of brain activations. These data suggest two function-
ally independent, hierarchically segregated systems for
sensory and perceptual representations of duration.
MÉTHODES
Participants
Twenty-eight participants were recruited in the study
(19 femmes, age = 27.7 ± 6.3 années, range = 20–48 years).
All participants had normal or corrected-to-normal vision.
Informed consent was obtained from all participants
before starting the experiment. Five participants were
excluded from the analysis of functional data because of
poor psychometric fits (with left tail >.25 and right tail
<.75 probability) for Accelerating probe data collected
during the structural scan. The functional data of the re-
maining 23 participants is reported below (15 women,
age = 28.5 ± 6.5 years, range = 20–48 years). The study
was approved by the University College London ethics com-
mittee and was in agreement with research guidelines and
regulations. Experimental protocols conformed to the
guidelines of the Declaration of Helsinki.
Apparatus
Visual stimuli were back-projected with an LCD projector
displaying at 1024 × 760 (60 Hz) onto a translucent screen at
1370
Journal of Cognitive Neuroscience
Volume 32, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
3
p
6
d
9
f
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
a
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
/
f
.
t
o
n
0
5
M
a
y
2
0
2
1
end of the MRI bore. Participants viewed the screen through
a mirror mounted on a 32-channel head coil. Participants
wore silicone earplugs and overear headphones through
which they received instructions. Stimulus presentation
and response collection were implemented on MATLAB
2011a (The MathWorks), running the Cogent toolbox library
(www.vislab.ucl.ac.uk/cogent.php).
Task and Stimuli
Participants performed duration comparisons in a visual
Standard–Probe binary choice task. Participants were tested
on two variants of the task, the first during a 12-min struc-
tural scan (Phase 1) and the second during two 22-min
functional scan runs (Phase 2).
Identifying the Accelerating Drift Timing Bias for
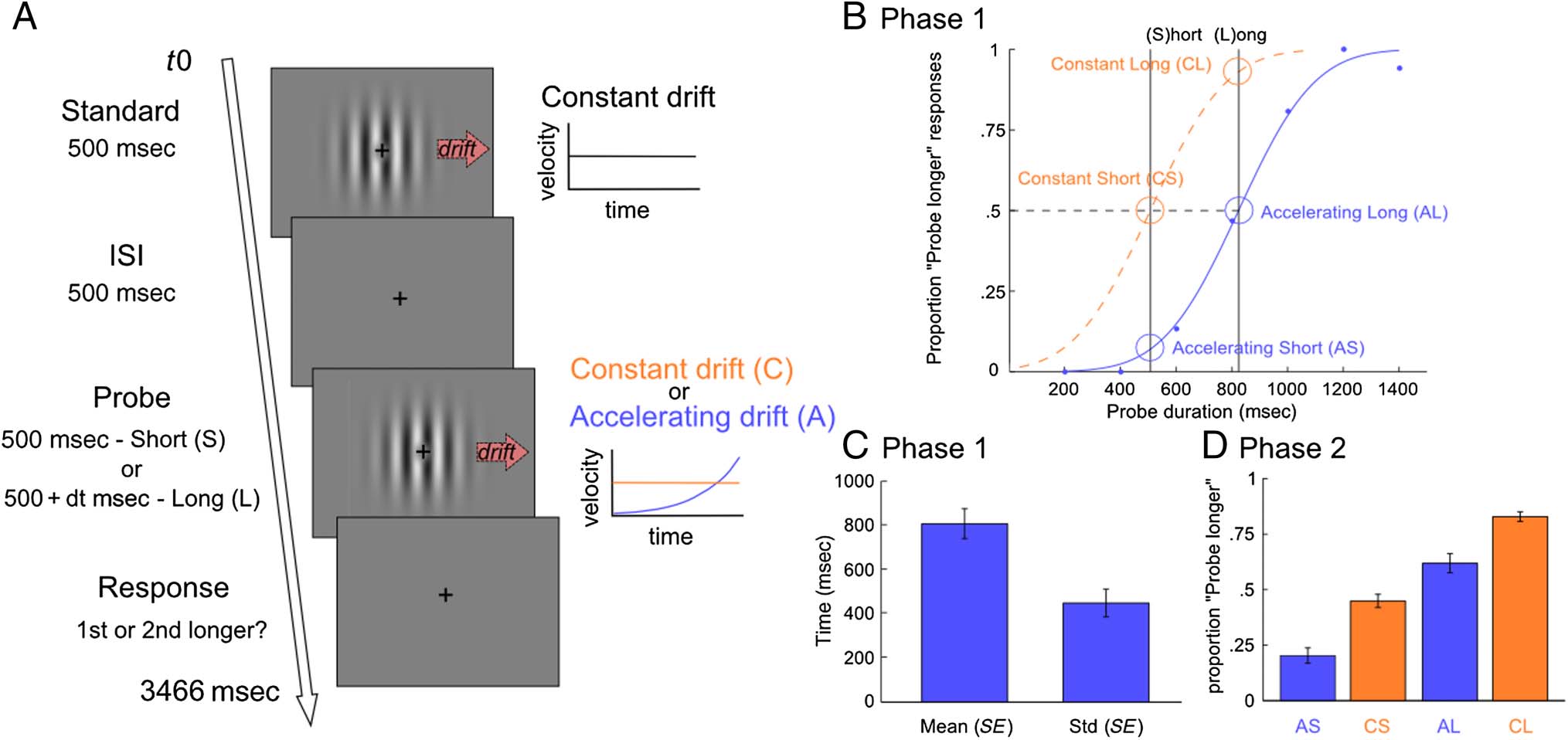
Perceptually Consistent Timing Categories (Phase 1)
Participants sequentially viewed Standard and Probe dura-
tions (Figure 1A), delivered through horizontally drifting
sinusoidal carriers set within a stationary Gaussian enve-
lope (drifting Gabor patch encompassing approximately
5° of visual angle, with a 0.6 cpd spatial frequency). The
central portion of the patch was occupied by a Gaussian
blob, which encompassed 0.5° of visual angle, on top of
which a fixation cross was overlaid. The Standard stimulus
drifted rightward with Constant velocity (0.42 rad dis-
placement per draw cycle) for 500 msec. The Probe stim-
uli were characterized by Constant or Accelerating drift
and variable duration (for animation lasting nFrames equiv-
alent to 500 msec or required to match point of subjective
equality [PSE], drift was approximated by a fourth-degree
polynomial function, y = 976.87 × x4 – 49620 × x3 +
9.1337e+05 × x2 − 6.2571e+06 × x + 1.0829e+07, for x
was between 1 and 30 in nFrames steps and y was succes-
sively scaled to guarantee that the Probe’s average drift
velocity was equal to the Standard drift velocity). Horizontal
shifts progressively increased (accelerated) across draw
cycles, with an average velocity that matched that of the
Constant drift Standard. Previous studies reveal that accel-
erating stimuli appear to last shorter than stimuli of equal
duration that move at constant velocity (Bruno et al., 2015;
Sasaki et al., 2013; Binetti et al., 2012; Matthews, 2011).
Probe duration was selected based on randomly perturbed
estimates of the PSE, yielded by a QUEST adaptive staircase
routine across 200 trials ( Watson & Pelli, 1983). Stimuli
were separated by a 500-msec ISI, where only the central
fixation cross was displayed. At the end of the trial, partici-
pants were required to indicate with a right hand button
press which stimulus, the first or second, lasted longer.
Standard and Probe stimulus order was counterbalanced
across trials. The purpose of this task was that of identifying
the PSE of the Accelerating drift stimuli for each participant,
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
3
p
6
d
9
f
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
a
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
t
.
f
/
o
n
0
5
M
a
y
2
0
2
1
Figure 1. (A) Time course of events in trial during functional scans. The Standard grating drifted rightward with Constant velocity for 500 msec, whereas
the Probe grating had Constant or Accelerating drift, lasting 500 msec or a duration that matched the participant’s PSE for accelerating stimuli. Stimuli were
separated by a 500-msec ISI. At the end of the trial, participants indicated which stimulus lasted longer (first or second). Standard and Probe order
was counterbalanced across trials. (B) During the structural scan (Phase 1), we identified the participant’s PSE for Accelerating stimuli, that is, duration
required for an Accelerating probe to appear equally long to the Constant standard. During the functional scans (Phase 2), participants were presented
Constant (C) and Accelerating (A) stimuli lasting 500 msec (S) or longer durations equivalent to the participant’s PSE (L), yielding four stimulus
categories: CS, CL, AS and AL. (C) Mean and variability (standard deviation) of the PSE required for Accelerating stimuli to be perceived equally long to
the 500-msec Constant drift Standard, as calculated during the structural scan (Phase 1). Error bars depict the SEM. (D) Proportion of “Probe lasted
longer” responses for the four stimulus categories presented during the functional scans (Phase 2).
Binetti et al.
1371
that is, the duration required for the Accelerating stimulus
to be perceived equally long to the Constant drift Standard.
This was aimed at generating duration categories that were
perceptually consistent across participants. We predicted,
based on previous observations (Binetti et al., 2012;
Matthews, 2011), that the Accelerating drift would pro-
duce a PSE greater than the 500-msec Standard duration,
indicating a compression of perceived duration, that is, the
Probe would have to last longer than the Standard in order
for the two to appear equally long.
yielded an estimate of the participant’s PSE for Accelerating
drift, that is, the duration required for an Accelerating Probe
to be perceived equally long to the Constant drift Standard.
A PSE larger than the 500-msec Standard would indicate that
accelerating drift induces a compression of time as the
Probe has to last longer than the Standard in order for the
two to appear equally long. The PSE values for Accelerating
drift, calculated for each participant, determined the dura-
tion of Long stimuli presented in Phase 2. Behavioral data
were analyzed with JASP 0.8.1.1 (https://jasp-stats.org/).
Dissecting Activity Sensitive to Objective and Subjective
Differences in Duration (Phase 2)
fMRI Data Acquisition
After completing the structural scan, participants performed
a similar task during two functional scan runs. The task
varied with respect to the previous one in two respects:
(1) Participants now estimated both Accelerating and
Constant drift probes, presented in alternating blocks of
six trials, and (2) only two Probe durations were tested
(Figure 1A, B): a 500-msec duration (Short—equivalent to
the standard duration) and the duration required for an
Accelerating probe to appear equally long to the standard
(>500 msec, Long—the PSE determined on an individual
basis in Phase 1). This yielded four stimulus combinations:
Constant-Short (CS), Accelerating-Short (AS), Constant-
Long (CL), and Accelerating-Long (AL). To maintain atten-
tional set, incidental Constant and Accelerating stimuli at
two flanker durations were included (Short flankers and
Long flankers—the duration required for an Accelerating
probe to generate 10% et 90% of “Probe longer”
réponses, respectivement; not shown in Figure 1). Ce
ensured the participants were engaged by occasionally ex-
posing them to easy trials (in which Standard and Probe
were noticeably different). Probe duration (Short, Long, et
flankers) was randomly selected across trials. Participants
were presented 140 Short probes (70 Constant drift +
70 Accelerating drift), 140 Long probes (70 Constant drift +
70 Accelerating drift), et 56 flankers (28 Short Constant
drift flankers + 28 Long Accelerating drift flankers), for a
total of 336 trials per run. Every 48 trials, participants had
a 12-sec resting period where only the central fixation cross
was displayed (six resting periods per run). Phase 2 était
aimed at identifying brain activity uniquely related to an
Objective difference in probe duration (Long vs. Short
probes) or to a Subjective difference in probe duration,
based on duration classifications (Subjectively Short vs.
Subjectively Long), revealing responses linked to duration
sensation and perception, respectivement.
Behavioral Data Analysis
Randomly perturbed QUEST estimates were binned (seven
linearly spaced time bins ranging between 100 et 1500 msec),
and the proportion of “Probe longer” responses per time bin
was calculated. We fit each participant’s responses with a cumu-
lative Gaussian (Figure 1B). Le 50% point of this function
Whole-brain 3-D gradient-echo EPI (Lutti, Thomas, Hutton,
& Weiskopf, 2013) data were acquired on a Siemens Trio 3T
scanner equipped with a 32-channel head coil for signal
reception. Functional data were acquired over two sessions
(2249 volumes total per participant). Acquisition parameters
were as follows: 3 mm isotropic resolution, echo time =
15.85 et 34.39 msec (multiecho acquisition), repetition
time = 1.1 sec per volume. Each volume was acquired with
an acceleration of 3 in the through-slab direction and a
speed-up factor of 2 in the in-plane direction, et le
images were reconstructed using the GRAPPA algorithm
(Griswold et al., 2002) as implemented on the scanner
console. The first five volumes were discarded to allow
for steady-state magnetization. A flip angle of 15° was used.
Field maps were acquired before the functional runs for
subsequent correction of geometric distortions in EPI data
at high field strength using a multiecho gradient-echo
with 3-mm isotropic resolution and echo times of 10 msec/
12.46 msec (short/long, respectivement). Whole-brain T1-weighted
anatomical images of each participant’s brain were acquired
using an optimized 3-D modified driven equilibrium Fourier
transform imaging sequence (Deichmann, Schwarzbauer, &
Tourneur, 2004) with 1-mm isotropic resolution. The sequence
used an inversion time of 910 msec, echo time = 2.48 msec,
repetition time = 7.92 msec, and flip angle = 16°.
fMRI Data Analysis
Data were analyzed using SPM12 (Wellcome Trust Centre for
Neuroimaging, University College London). Preprocessing
included realignment and unwarping using individual field
maps, coregistration of EPI to individual anatomical images,
spatial normalization to the Montreal Neurological Institute
(MNI) espace, and spatial smoothing using an 8-mm FWHM
Gaussian kernel.
We disambiguate Objective and Subjective duration by
contrasting stimuli based on their physical duration (Long—
L vs. Short—S) or based on the classification of duration
as indicated through participant responses (Probe judged
shorter—0 vs. Probe judged longer—1). The general linear
model included 13 event-related regressors modeling
responses to each event in our 2 × 2 × 2 factorial design.
The first eight involved combinations of stimulus type
1372
Journal des neurosciences cognitives
Volume 32, Nombre 7
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
7
e
1
–
3
p
6
d
9
F
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
un
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
t
.
/
F
o
n
0
5
M.
un
oui
2
0
2
1
(Accelerating—A/Constant drift—C), stimulus duration
(Short—S/Long—L), and perceived duration (Perceived
Short—0/Long—1), modeled relative to the onset of the
Probe stimulus. En plus, the onset of short (Regressor 9)
and long (Regressor 10) flanker stimuli, the onset of
(Regressor 11) eye blinks, (Regressor 12) pauses, et le
onset participants’ button presses (Regressor 13) were in-
cluded. All event-related responses were modeled as stick
les fonctions; apart from pauses that were modeled as contin-
uous 12-sec events. In addition to event regressors, a total
de 23 nuisance regressors were included to account for mo-
tion and physiological effects of no interest. A physiological
noise model was constructed using an in-house MATLAB
toolbox (Hutton et al., 2011) to remove variance accounted
for by cardiac and respiratory responses. The model com-
prised 17 physiological regressors in total: 6 for respiratory
phase, 1 for respiratory volume, et 10 for cardiac phase.
Enfin, to discount motion-related artifacts that eluded rigid
body motion correction, the six motion regressors obtained
during realignment were included.
In this paper, we focus on the simple main effects of
Objective and Subjective duration (Short vs. Long) par
testing two contrasts. Objective duration was explored by
contrasting (CL1 + AL1 + CL0 + AL0)–(CS1 + AS1 + CS0 +
AS0). Subjective duration was explored by contrasting stimuli
based on the participant duration classifications, indepen-
dently of their objective duration: Subjective contrast (CS1
+ AS1 + CL1 + AL1)–(CS0 + AS0 + CL0 + AL0). Pour
whole-brain analyses, statistical parametric maps of each
contrast were characterized in terms of significant clusters,
using a cluster-defining threshold of p = .001; le 0.05
family-wise error (FWE)-corrected critical cluster size was
100 voxels. We also report additional findings descriptively
at p = .001, uncorrected for subsequent confirmation.
We also directly contrasted Subjective and Objective dura-
tion to identify activations that significantly differed between
these conditions.
Experimental data and codes used in the study are avail-
able upon direct request, consistent with University
College London’s guidelines and regulations.
RÉSULTATS
Behavioral Data
We computed the PSE for Accelerating drift stimuli. Comme
anticipated, we found that Accelerating drift induced a
substantial compression of perceived time: on average,
Accelerating drift stimuli had to last 806 ± 324 msec to
be perceived equally long to the 500-msec Constant drift
Standard (Figure 1C).
We submitted proportion of “Probe longer” responses to
a repeated-measures ANOVA with Stimulus Type as within-
subject factor (AS vs. CS vs. AL vs. CL; Figure 1D) and par-
ticipant Age (binned, above and below mean) and Sex as
covariables. The analysis revealed a main effect of Stimulus
Type, F(3, 57) = 51.99, p < .001, ηp
2 = .71; we observed
no modulatory effect of Age, F(3, 57) = .2, p = .89, ηp
2 =
.003, or Sex, F(3, 57) = 1, p = .4, ηp
2 = .01, on Stimulus
Type. t-Test comparisons showed that, relative to
Constant drift stimuli, Accelerating drift stimuli yielded
significantly smaller proportions of “Probe longer”
responses, both for Short (AS vs. CS: t(22) = −4.6, p <
.0001, d = −.96) and Long (AL vs. CL: t(22) = −4.78,
p < .00001, d = −.99) durations.
We also tested whether proportions of “Probe longer”
responses for AS, CS, AL, and CL stimuli differed from
chance-level performance (.5 proportion of “Probe longer
responses”). Bonferroni corrected t tests revealed that,
despite lasting the same as the Standard, the proportion
of “Probe longer” responses for AS stimuli significantly
differed from chance level, t(22) = −8.61, p < .00001, d =
−1.79, due to an illusory shortening of time induced by
acceleration. Unsurprisingly, proportion of “Probe longer”
responses for CL stimuli also significantly differed from
chance level, t(22) = 14.83, p < .00001, d = 3.09, as these
did not accelerate and physically lasted longer than the
Standard. Proportion of “Probe longer” responses for AL
stimuli showed a borderline significant difference from
chance level, t(22) = 2.73, p = .049, d = .57. Because
Long durations were selected based on each participant’s
PSE for accelerating stimuli observed in Phase 1, AL stimuli
were perceived on average similar in duration to the
Standard despite their physical difference in duration.
Finally, proportion of “Probe longer” responses for CS
stimuli, which shared duration and drift profile with the
Standard, did not significantly differ from chance level,
t(22) = −1.74, p = .38, d = −.36.
Imaging Analyses
Our general linear model design enabled contrasting
stimuli based on differences in Objective duration
(Objectively Long = L vs. Objectively Short = S) or based
on the classification of duration provided by participant
responses on a trial-by-trial basis, Subjective duration
(Subjectively Long = 1 vs. Subjectively Short = 0). This
design accounts for low-level feature processing, atten-
tional, decisional, and motoric task demands, which are
equally shared across conditions. We performed two con-
trasts to identify brain activity uniquely related to Objective
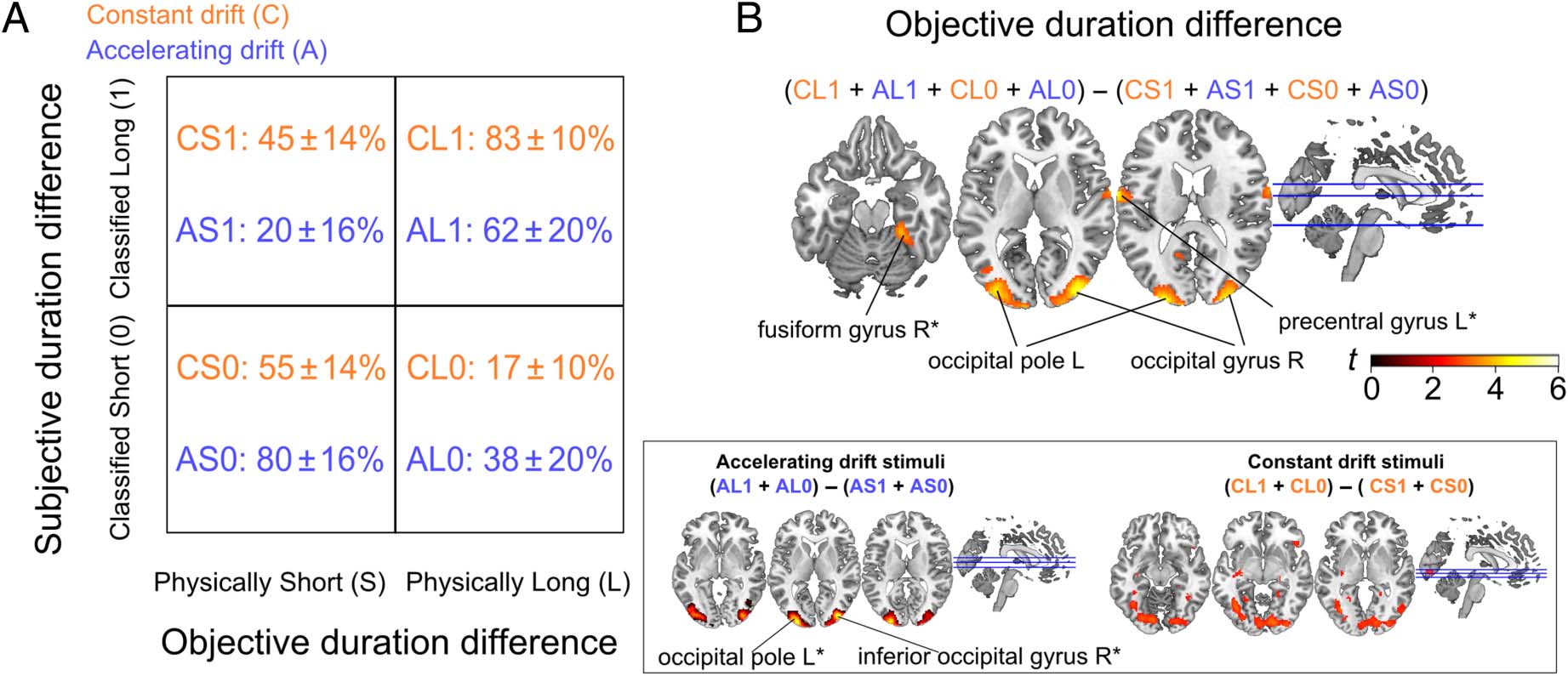
and Subjective differences in duration (Figure 2A).
Objective Duration Contrast Reveals Activity Bilateral
Early Visual Extrastriate Areas
Objective duration was explored by contrasting longer
versus shorter stimuli, independently of participant re-
sponses (Figure 2B, top). Activated areas were confined
to visual regions including right inferior occipital gyrus ( p =
.002 FWE cluster-corrected; peak coordinate [36 −88 6],
t(1, 22) = 4.96) and the left occipital pole ( p = .003 FWE
cluster-corrected; peak coordinate [−22 −96 14], t(1, 22) =
4.82). For exploratory purposes, we also evaluated the
Binetti et al.
1373
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
3
p
6
d
9
f
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
a
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
.
t
/
o
n
0
5
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
3
p
6
d
9
f
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
a
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
Figure 2. (A) Objective and Subjective duration contrasts, based on the subjective (columns) or objective stimulus duration (rows). Percentages
depict the distribution of Short (S) and Long (L) Control (C) and Accelerating (A) stimuli across Objective/Subjective duration categories. (B) Axial
slices depicting activations for the contrast identifying objective duration difference, collapsed across stimuli categories (top). Lower panels show the
activity changes within each stimulus category. Activation images thresholded for t scores > 2.8 (all stimuli and Accelerating stimuli) and t scores >
2.3 (Constant stimuli). *p < .001, uncorrected.
Objective duration contrast at a more lenient p < .001 thresh-
old (uncorrected), which showed additional activity in the left
precentral gyrus (peak coordinate [−66 −4 16], t(1, 22) =
4.53) and the right fusiform gyrus (peak coordinate [24
−36 −20], t(1, 22) = 4).
We further explored Objective duration for Accelerating and
Constant stimuli separately (Figure 2B, bottom). Accelerating
drift stimuli ((AL1 + AL0) − (AS1 + AS0)) revealed a cluster
of activation in the right inferior occipital gyrus ( p = .005
FWE cluster-corrected; p = .046 voxelwise FWE correction;
peak coordinate [−36 −86 2], t(1, 22) = 6.18) and in the
left occipital pole ( p < .001 FWE cluster-corrected; peak
coordinate [−24 −96 8], t(1, 22) = 5.67). For Constant drift
stimuli ((CL1 + CL0) − (CS1+CS0)), no regions reached
either FWE cluster-corrected significance ( p < .05) or
significance at more lenient statistical thresholds ( p < .001,
uncorrected). We explored Objective duration for Constant
drift stimuli at a more lenient threshold ( p < .01 uncor-
rected), which showed an equivalent pattern of activity
to the Accelerating drift stimuli, with a cluster of activation
in the left and right occipital gyri. Although these patterns
of activity were equivalent for Accelerating and Constant
drift stimuli, the former were more effective at revealing
differences in time sensation.
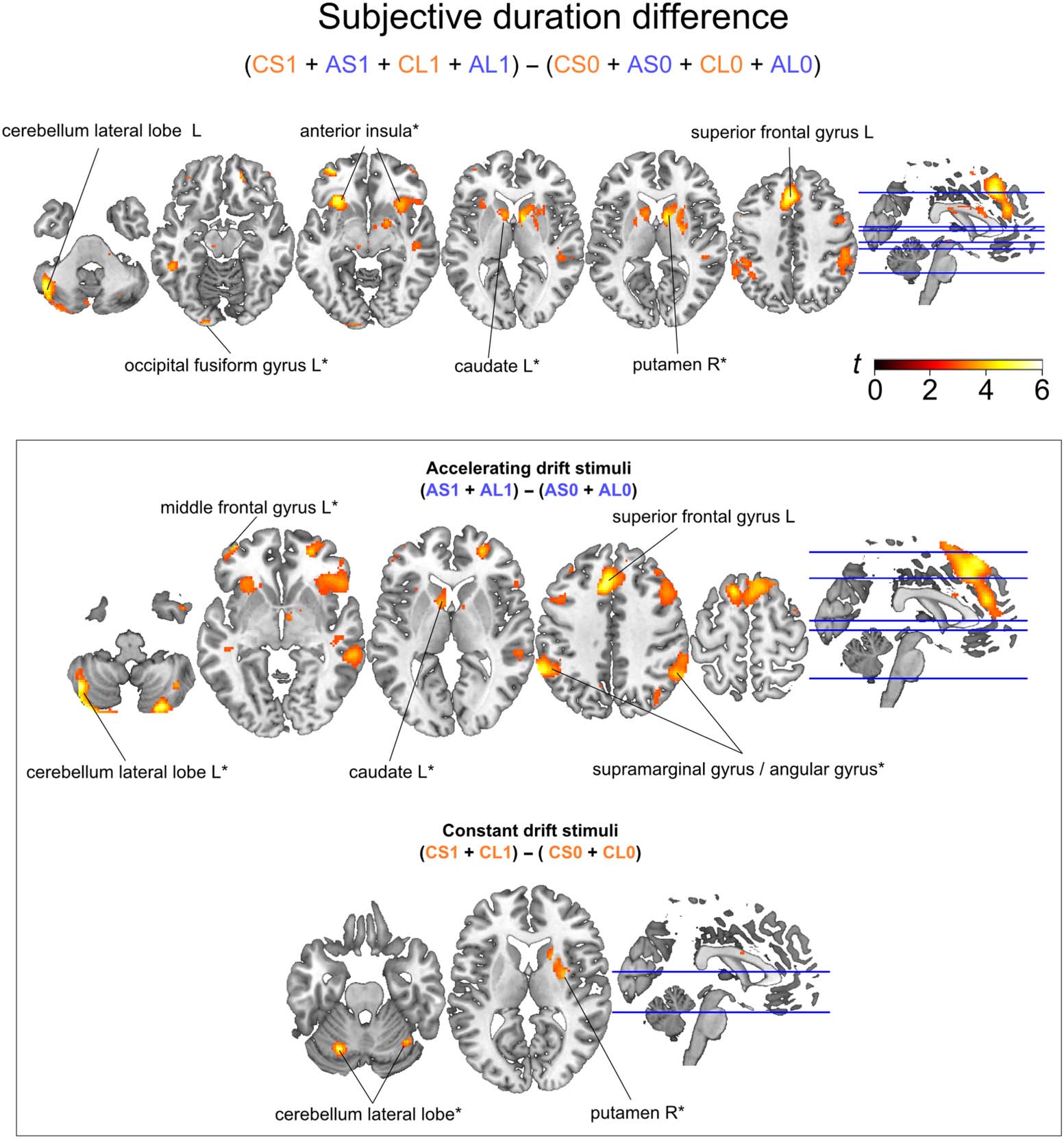
Subjective Duration Contrast Reveals Activity in the Left
Superior Frontal Gyrus and the Left Cerebellum
Subjective duration was explored by contrasting stimuli
based on a participant’s duration classification, indepen-
dently of their physical duration (Figure 3, top). We iden-
tified a cluster of activation in the medial segment of the
left superior frontal gyrus ( p = .002 FWE cluster-
corrected; peak coordinate [−2 28 42], t(1, 22) = 5.61)
and on the left lateral lobe of the cerebellum ( p = .041
FWE cluster-corrected; peak coordinate [−46 −60 −38],
t(1, 22) = 5.17). In addition, at a more lenient statistical
threshold (i.e., p < .001 uncorrected), activations includ-
ed the right putamen (peak coordinate [26 −6 8], t(1, 22) =
3.89), the left caudate ([−8 8 4], t(1, 22) = 3.81), the right
anterior insula ([32 18 −8], t(1, 22) = 3.93), the left parietal
cortex (supramarginal gyrus [−46 −50 56], t(1, 22) = 4.3)
and the left occipital fusiform gyrus ([−16 −98 −14],
t(1, 22) = 3.81). We explored Subjective duration for each
stimulus drift type separately (Constant and Accelerating;
Figure 3, bottom). Accelerating drift stimuli ((AS1 + AL1)
− (AS0 + AL0)) showed a cluster of activation in the
medial segment of the left superior frontal gyrus ( p <
.001 FWE cluster-corrected; peak coordinate [−4 26 42],
t(1, 22) = 5.69) and marginally significant activations ( p <
.001 uncorrected) in the left and right lateral lobes of the
cerebellum (left: peak coordinate [−44 −62 −44], t(1, 22)
= 5.81; right: peak coordinate [26 −80 −46], t(1, 22) = 5.41),
left superior frontal gyrus (peak coordinate [−14 18 62],
t(1, 22) = 4.65), left middle frontal gyrus (peak
coordinate [−40 56 −4], t(1, 22) = 4.27), right supra-
marginal gyrus (peak coordinate [60 −40 38], t(1, 22) =
3.53), right angular gyrus (peak coordinate [54 −52 40], t
(1, 22) = 4.19), and left caudate (peak coordinate [−8 8 4], t
(1, 22) = 3.8), all areas generally associated with tasks that
require integration of somatosensory signals and timing
tasks (Jones & Jahanshahi, 2011; Buhusi & Meck, 2005;
Nenadic et al., 2003; Rao et al., 2001; Ivry & Keele,
1989). More specifically, the BG and pFC are believed to
be involved in magnitude representations used in percep-
tual timing tasks (Buhusi & Meck, 2005). Constant drift
1374
Journal of Cognitive Neuroscience
Volume 32, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
3
p
6
d
9
f
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
a
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
.
t
o
n
0
5
M
a
y
2
0
2
1
Figure 3. Subjective duration contrast activations, collapsed across stimuli categories (top) or within each stimulus category separately (bottom).
Activation images thresholded for t scores > 2.8. *p < .001, uncorrected, for display purposes.
stimuli ((CS1 + CL1) − (CS0 + CL0)) revealed only mar-
ginally significant activations on the left and right lateral
lobe of the cerebellum ( p < .001 uncorrected, left: peak
coordinate [−22 −66 −24], t(1, 22) = 5.51; right: peak
coordinate [36 −60 −24,] t(1, 22) = 3.86) and the right
putamen ( p < .001 uncorrected, peak coordinate [30 −2
10], t(1, 22) = 3.93).
Subjective–Objective Duration Direct Comparison
Confirms Differences in Frontal–Medial Areas
Objective and Subjective activations were directly contrasted
to formally assess differences between networks uniquely
processing timing sensation and perception, identified by
the simple main Objective and Subjective contrasts. Paired-
sample t tests revealed a cluster of activity in frontal–medial
areas, peaking in dorsal ACC BA 32 ( p < .001 FWE cluster-
corrected; peak coordinate [−12 32 24], t(1, 22) = 5.22).
This cluster included left superior frontal gyrus, which was
the strongest activation observed in the Subjective contrast
( p < .001 FWE cluster-corrected; peak coordinate [−4 26
40], t(1, 22) = 5.22). At a more lenient statistical threshold
(i.e., p < .001 uncorrected), we observed a pattern of acti-
vations that were consistent with what observed in the
Subjective duration contrast, notably including the insula
(left: peak coordinate [−32 −20 −8], t(1, 22) = 5.08; right:
peak coordinate [32 18 −10], t(1, 22) = 3.89) and the left
lobe of the cerebellum (peak coordinate [−26 −86 −42],
t(1, 22) = 5.08; Figure 4).
DISCUSSION
We identified brain activity uniquely sensitive to Objective
differences in duration (time sensation) and Subjective
differences in duration (time perception) while accounting
for low-level feature processing, attentional, decisional, and
Binetti et al.
1375
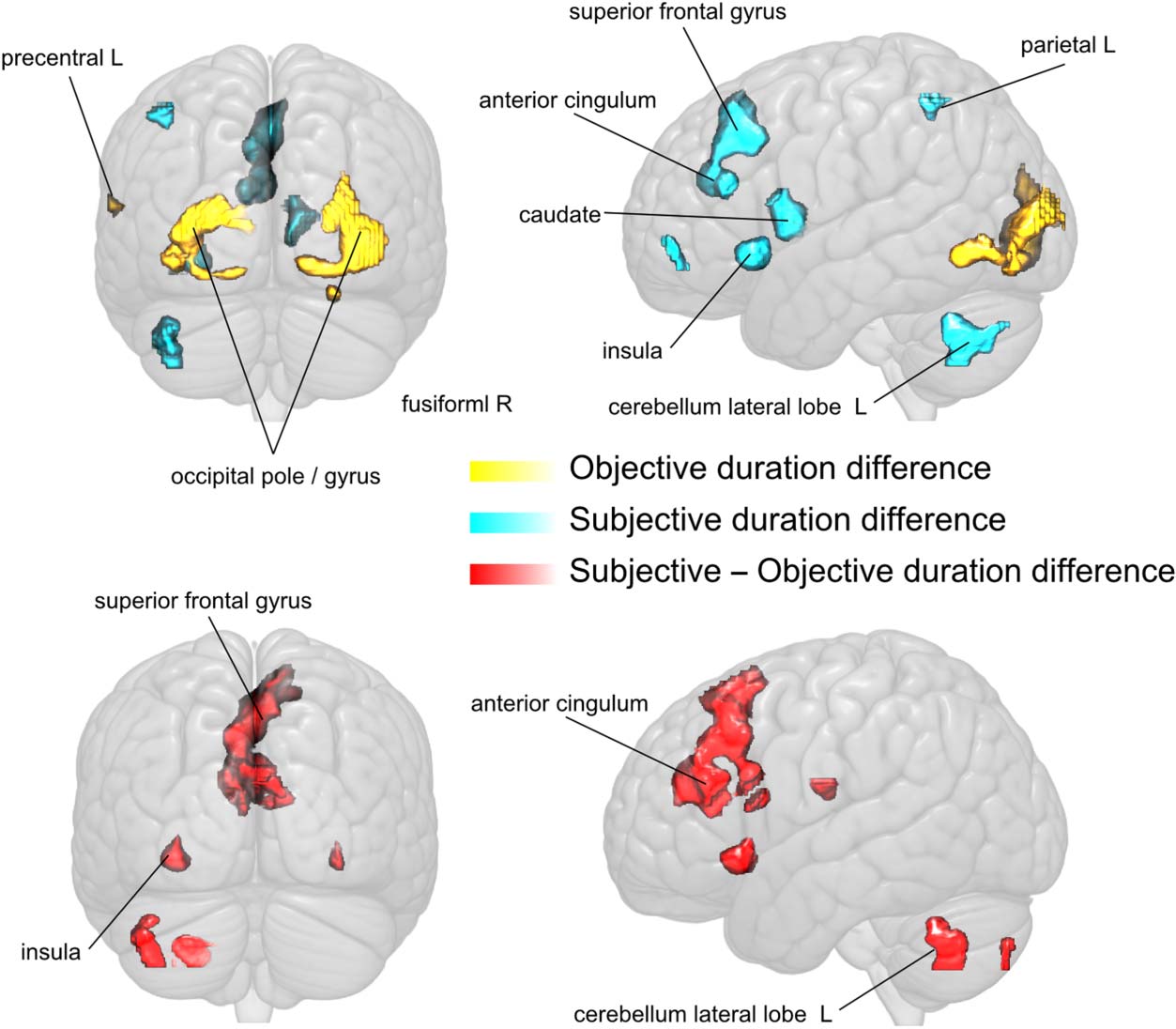
Figure 4. Rendered,
semi-transparent view depicting
brain regions sensitive to
Objective (yellow) and
Subjective (blue) duration
(t scores > 2.8 and Objective
cluster size > 12 voxels or
Subjective cluster size > 35
voxels).
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
o
h
w
t
t
n
p
o
:
un
/
d
/
e
m
d
je
F
r
t
o
p
m
r
c
h
.
s
p
je
je
d
v
je
r
e
e
r
c
t
c
.
m
h
un
je
e
r
d
.
toi
c
o
o
m
c
n
/
j
un
o
r
t
c
je
c
n
e
/
–
un
p
r
d
t
je
3
c
2
je
7
e
1
–
3
p
6
d
9
F
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
un
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
oui
n
g
_
toi
un
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
F
p
e
b
m
oui
b
e
g
r
toi
2
e
0
s
2
t
3
/
j
F
t
.
/
o
n
0
5
M.
un
oui
2
0
2
1
motoric task demands, which were balanced across condi-
tion. We found that Objective duration was reflected in
activity increases in early visual areas (left occipital pole and
right inferior occipital gyrus), whereas Subjective duration
was reflected in activity increases across a cortical/subcortical
network comprising the left superior frontal gyrus and the
left lateral cerebellum (Tableau 1). A more lenient threshold
( p < .001, uncorrected) revealed additional activity in-
creases in the BG (right putamen, left caudate), the right
supramarginal gyrus, and right angular gyrus, all areas typi-
cally associated with perceptual timing tasks. Interestingly,
there was no overlap between these networks, suggesting a
clear dichotomy between activity linked to Objective and
Subjective duration (Figure 4).
Timing Is Achieved throughout a Distributed
Cortical and Subcortical Hierarchy
Timing relies on mechanisms that guarantee a quasi-linear
correspondence between objective and perceived dura-
tion, at least within the millisecond-to-second range. As
with all perceptual systems, timing is constructive, where
sensory and nonsensory (e.g., contextual, prior experience)
information are integrated across processing stages. Indeed,
neuroimaging, brain lesion, and neurophysiological evi-
dence reveal a highly modular and distributed architecture
(Merchant, Harrington, & Meck, 2013). Furthermore, recent
studies reveal a chronotopic organization of time (Harvey,
Dumoulin, Fracasso, & Paul, 2020; Protopapa et al., 2019).
Adaptation studies show duration selectivity in neural popu-
lations of the inferior parietal lobule, the posterior temporal
cortex, the middle frontal gyrus, the middle cingulate cortex,
the caudate, putamen, the inferior temporal gyri (Hayashi
et al., 2015), and the SMA (Protopapa et al., 2019). Given
the repeated presentation of two durations in our study
(500 and ∼800 msec—average PSE), a subset of activations
(parietal, cingulate cortex, putamen, and caudate) may
additionally reflect adaptation-like effects uncovered in
these previous studies.
Primate electrophysiology (Shuler & Bear, 2006; Ghose
& Maunsell, 2002) and human neuroimaging (Bueti et al.,
2010; Bueti & Macaluso, 2010) studies have shown that
time information modulates activity in primary visual areas,
suggesting an involvement of sensory cortices in memory
and encoding stages of time perception (Salvioni et al.,
2013). Studies contrasting timing against carefully matched
nontiming tasks have identified the cerebellum and
BG/striatum as candidate time accumulation structures,
whereas frontal and parietal areas are more likely involved
with attentional, working memory and decisional aspects
of the task (Ferrandez et al., 2003; Nenadic et al., 2003;
Belin et al., 2002; Rao et al., 2001). Physiological and imaging
studies have narrowed down brain activity that is modu-
lated by time and therefore more specifically linked to
timekeeping. Direct recordings in macaque parietal cortex
show a build-up of response frequency as a function of
duration ( Janssen & Shadlen, 2005). Similarly, amplitude
of evoked brain responses originating in frontal brain areas
(contingent negative variation) covary with elapsed dura-
tion (Herbst, Chaumon, Penney, & Busch, 2015; Pfeuty
et al., 2005). These findings suggest that areas involved with
the accumulation of timing evidence should be sensitive
1376
Journal of Cognitive Neuroscience
Volume 32, Number 7
Table 1. FWE Cluster-corrected Peak Coordinates of Activation for Objective and Subjective Duration Contrasts and
Objective/Subjective Duration for Accelerating Stimuli
Contrast
Objective
Objective
Subjective
Subjective
Objective
(Accelerating stimuli)
Objective
(accelerating stimuli)
Subjective
(accelerating stimuli)
Subjective
(accelerating stimuli)
Location in Stereotaxic
Space: MNI
Cluster Size
pFWE
t Value (Peak)
Label (Neuromorphometrics)
38 −88 6
−22 −96 14
−2 28 42
−46 −60 −38
36 −86 2
712
622
608
294
512
.002
4.96
4.82
5.61
5.17
.003
.002
.041
.005
Right IOG (inferior
occipital gyrus)
Left OP (occipital pole)
Left MSFG (superior frontal
gyrus medial segment)
Left cerebellum exterior
6.18 Also
pfwe = 0.046
voxelwise
Right IOG (inferior
occipital gyrus)
−24 −96 8
886
p < .001
5.67
Left OP (occipital pole)
−44 −62 −44
367
.055
5.81
Left cerebellum exterior
−4 26 42
1285
p < .001
5.696
Left MSFG (superior frontal
gyrus medial segment)
to differences in duration, which is a feature expected from
hypothetical time accumulation mechanisms.
Objective or Subjective Differences in Duration as
Handles of Time Accumulation Mechanisms
Based on this evidence, Pouthas and coworkers (2005) inves-
tigated what areas within a timing network were specifically
modulated by duration, aimed at identifying physiological
correlates of time accumulator/s (Pouthas et al., 2005). A
timing versus nontiming task contrast identified a timing
network comprising the frontal, mesiofrontal (preSMA),
parietal cortices, and BG. This was followed by contrasting
Long versus Short stimuli, revealing activity sensitive to
difference in duration in the preSMA, the ACC, the right
inferior frontal gyrus (corresponding to Broca’s area),
the lateral premotor cortex bilaterally, and the right caudate
nucleus. PreSMA and the caudate nucleus were identified as
candidate accumulator mechanisms, consistently with neu-
ropsychological (Jahanshahi et al., 2010; Malapani, Deweer,
& Gibbon, 2002; Harrington, Haaland, & Hermanowitz,
1998) and pharmacological (Coull, Hwang, Leyton, &
Dagher, 2012) evidence linking frontostriatal dopaminergic
activity to timing. The authors argued that the ACC activa-
tion reflected attentional control related to the response
selection component of the task, where longer durations
require longer sustaining of attention ( Wu et al., 2017;
Peru, Pavesi, & Campello, 2004).
As with all constructive processes, our perception of dura-
tion is not necessarily veridical. This is clearly evidenced by
time distortions induced by endogenous factors such as fluc-
tuations of arousal (Binetti, Harrison, Mareschal, & Johnston,
2017) or dopamine levels (Marinho et al., 2018) or through
manipulations of stimulus features, such as stimulus number,
size, or luminance (Xuan, Zhang, He, & Chen, 2007) or
motion dynamics (Binetti et al., 2012; Matthews, 2011;
Kanai, Paffen, Hogendoorn, & Verstraten, 2006). Based
on this, perceived duration, opposed to objective duration
of a stimulus, should provide a more robust handle on
timekeeping. Bueti and Macaluso (2011) focused on the
relationship between the perceptual experience of time
and corresponding neural activity by inducing illusory
time dilation via visual motion (faster moving stimuli =
time overestimation) of stimuli in a duration reproduction
task (Bueti & Macaluso, 2011). Contrasting the timing task
versus a nontiming control revealed activity in the puta-
men, the mid-insula, the SMA, the mid/superior temporal
gyri, the right VI lobule of the cerebellum, and the TPJ.
Importantly, the authors identified activity that covaried
with perceptual duration estimates (reproduction errors)
in the right putamen, the right mid-insula, and the superior
temporal cortex, revealing accumulation behavior across
various brain structures. However, a limitation of studies
relying on timing versus nontiming comparisons is that,
despite being carefully matched in difficulty and sensory
features, they can introduce discrepancies in attentional,
strategic, or decisional demands (Pouthas et al., 2005).
Disentangling Activity Uniquely Sensitive to
Objective and Subjective Differences in Duration
In this study, we combined approaches comparing Ob-
jective and Subjective differences in duration, within a
single protocol strictly focused on comparisons within
Binetti et al.
1377
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
3
p
6
d
9
f
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
a
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
.
/
o
n
0
5
M
a
y
2
0
2
1
timing conditions. Opposed to prior studies, this ap-
proach did not rely on a nontemporal control task.
Both Objective and Subjective contrasts accounted for
processing of low-level visual features of stimuli,
attentional, decisional, and motoric task demands,
which were equally shared across conditions.
Objective duration was associated with activity increases
in early visual areas (left occipital pole and right inferior
occipital gyrus), corresponding to bilateral early extrastriate
area, as previously reported in animal (Onoe et al., 2001) and
human imaging studies (Bueti et al., 2010). Importantly,
early sensory areas have been associated with the extraction
of time features from the sensory signal (Salvioni et al.,
2013), and Objective differences in duration did not modu-
late activity in areas beyond the visual system that are
involved with cognitive (i.e., nonsensory) components of
timing. For exploratory purposes, we investigated brain
activity at a more lenient threshold ( p < .001, uncorrected).
We found activations in the left precentral gyrus, which has
been frequently documented in time estimation (Ortuño,
Guillén-Grima, López-García, Gómez, & Pla, 2011) and re-
production tasks ( Jech, Dušek, Wackermann, & Vymazal,
2005), as well as the right fusiform gyrus, which has been
linked to attention and working memory in visual duration
discrimination tasks (Pouthas et al., 2000). The fusiform
gyrus therefore was the latest visual processing stage whose
activity was modulated by objective duration information.
We observed no activity in V5, which was expected, given
that stimuli of different durations were equated in average
velocity.
Subjective duration was reflected in activity increases in
the medial segment of the left superior frontal gyrus, on
the border between the dorsal anterior cingulate and the
pFC, and the left cerebellum. The dorsal anterior cingu-
late is involved in reward-based decision-making (Bush
et al., 2002) and in focusing attention toward task-relevant
features ( Weissman, Gopalakrishnan, Hazlett, & Woldorff,
2004), whereas the pFC has been frequently linked to
timing performance in neuropsychological (Koch, Oliveri,
Carlesimo, & Caltagirone, 2002) and brain stimulation
studies (Koch, Oliveri, Torriero, & Caltagirone, 2003) and
has been associated with attention to time information
(Coull, Vidal, Nazarian, & Macar, 2004). Notably, both the
superior frontal gyrus (Murai, Whitaker, & Yotsumoto, 2016;
Jones & Jahanshahi, 2011) and the cerebellum (Murai et al.,
2016; Lewis & Miall, 2003; Ivry & Keele, 1989) are com-
monly identified in standard/probe comparisons (Wiener,
Turkeltaub, & Coslett, 2010). At more lenient thresholds
( p < .001, uncorrected), we observed activations in the pa-
rietal cortex, a hub for abstract representations of quanti-
ties (Bueti & Walsh, 2009), posterior cingulate cortex, the
angular gyrus, and the BG (right putamen and left cau-
date), all areas associated with perceptual timing tasks
(Nenadic et al., 2003; Onoe et al., 2001; Rao et al., 2001;
Ivry & Keele, 1989). Notably, we also observed activations
in the anterior insula, which has been previously observed
in imaging (Bueti & Macaluso, 2011; Wittmann et al., 2010)
and brain lesion (Trojano et al., 2017) timing studies. The
insula is believed to mediate the subjective awareness of
duration through the integration of interoceptive signals
(Wittmann & Meissner, 2018; Craig, 2009a, 2009b). Also,
a recent meta-analysis on conscious visual awareness revealed
a distributed network involving the superior frontal gyrus,
the caudate, insula, and the fusiform gyrus (Bisenius, Trapp,
Neumann, & Schroeter, 2015), all areas observed in the
subjective duration contrast. Importantly, Objective and
Subjective contrasts yielded independent, nonoverlap-
ping sets of activations, confirmed by direct comparisons
of Subjective and Objective activations. This suggests
functional independence of sensory and perceptual timing
stages, which has also been reported in the frequency
domain in an MEG study (Noguchi & Kakigi, 2006).
Conclusion
We identify within a timing network a subset of brain areas
that are uniquely related to Objective differences in dura-
tion and Subjective differences in duration, thus disentan-
gling time sensation and perception. We argue that early
visual extrastriate regions extract duration features from
the sensory signal (time sensation), without directly medi-
ating a subjective duration experience, whereas superior
frontal gyrus and the cerebellum relate to higher-order
timekeeping, attentional and decisional processes sub-
tending the subjective experience of duration and ensuing
behavior (time perception). Importantly, time sensation
and perception arise from functionally independent pro-
cessing stages in the cortical hierarchy.
Acknowledgments
N. B. was supported through the Newton International Fellowship
(Fellowship Number FN111112) awarded by the British Academy
and the Royal Society. S. B. was supported by the European
Research Council Starter Grant (ActSelectContext; 260424).
The Wellcome Centre for Human Neuroimaging is supported
by core funding from the Wellcome trust (203147/Z/16/Z). K. F.
was funded by a Wellcome Trust Principal Research Fellowship
(Ref: 088130/Z/09/Z). We thank Peter Zeidman for his excellent
suggestions with the data analysis and interpretation of findings.
Reprint requests should be sent to Nicola Binetti, Gower St.,
Bloomsbury, WC1E 6BT, University College London, United
Kingdom, or via e-mail: nicolabinetti@gmail.com.
REFERENCES
Belin, P., McAdams, S., Thivard, L., Smith, B., Savel, S.,
Zilbovicius, M., et al. (2002). The neuroanatomical substrate
of sound duration discrimination. Neuropsychologia, 40,
1956–1964.
Binetti, N., Harrison, C., Mareschal, I., & Johnston, A. (2017).
Pupil response hazard rates predict perceived gaze durations.
Scientific Reports, 7, 3969.
Binetti, N., Lecce, F., & Doricchi, F. (2012). Time-dilation and
time-contraction in an anisochronous and anisometric visual
scenery. Journal of Vision, 12.
1378
Journal of Cognitive Neuroscience
Volume 32, Number 7
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
3
p
6
d
9
f
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
a
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
/
t
.
o
n
0
5
M
a
y
2
0
2
1
Bisenius, S., Trapp, S., Neumann, J., & Schroeter, M. L. (2015).
Identifying neural correlates of visual consciousness with
ALE meta-analyses. Neuroimage, 122, 177–187.
Bruno, A., Ayhan, I., & Johnston, A. (2015). Changes in apparent
duration follow shifts in perceptual timing. Journal of
Vision, 15, 2.
Bueti, D., Bahrami, B., Walsh, V., & Rees, G. (2010). Encoding
of temporal probabilities in the human brain. Journal of
Neuroscience, 30, 4343–4352.
Bueti, D., & Macaluso, E. (2010). Auditory temporal expectations
modulate activity in visual cortex. Neuroimage, 51, 1168–1183.
Bueti, D., & Macaluso, E. (2011). Physiological correlates of
subjective time: Evidence for the temporal accumulator
hypothesis. Neuroimage, 57, 1251–1263.
Bueti, D., & Walsh, V. (2009). The parietal cortex and the
representation of time, space, number and other magnitudes.
Philosophical Transactions of the Royal Society of London,
Series B: Biological Sciences, 364, 1831–1840.
Buhusi, C. V., & Meck, W. H. (2005). What makes us tick?
Functional and neural mechanisms of interval timing.
Nature Reviews Neuroscience, 6, 755–765.
Buonomano, D. V., Bramen, J., & Khodadadifar, M. (2009).
Influence of the interstimulus interval on temporal
processing and learning: Testing the state-dependent
network model. Philosophical Transactions of the Royal
Society of London, Series B: Biological Sciences, 364,
1865–1873.
Bush, G., Vogt, B. A., Holmes, J., Dale, A. M., Greve, D., Jenike, M. A.,
et al. (2002). Dorsal anterior cingulate cortex: A role in
reward-based decision making. Proceedings of the National
Academy of Sciences, U.S.A., 99, 523–528.
Coull, J. T., Hwang, H. J., Leyton, M., & Dagher, A. (2012).
Dopamine precursor depletion impairs timing in healthy
volunteers by attenuating activity in putamen and
supplementary motor area. Journal of Neuroscience,
32, 16704–16715.
Coull, J. T., Nazarian, B., & Vidal, F. (2008). Timing, storage, and
comparison of stimulus duration engage discrete anatomical
components of a perceptual timing network. Journal of
Cognitive Neuroscience, 20, 2185–2197.
Coull, J. T., Vidal, F., Nazarian, B., & Macar, F. (2004). Functional
anatomy of the attentional modulation of time estimation.
Science, 303, 1506–1508.
Craig, A. D. (2009a). Emotional moments across time: A
possible neural basis for time perception in the anterior
insula. Philosophical Transactions of the Royal Society of
London, Series B: Biological Sciences, 364, 1933–1942.
Craig, A. D. (2009b). How do you feel—Now? The anterior
insula and human awareness. Nature Reviews Neuroscience,
10, 59–70.
Deichmann, R., Schwarzbauer, C., & Turner, R. (2004).
Optimisation of the 3D MDEFT sequence for anatomical
brain imaging: Technical implications at 1.5 and 3 T.
Neuroimage, 21, 757–767.
Eagleman, D. M. (2008). Human time perception and its
illusions. Current Opinion in Neurobiology, 18, 131–136.
Ferrandez, A. M., Hugueville, L., Lehéricy, S., Poline, J. B.,
Marsault, C., & Pouthas, V. (2003). Basal ganglia and
supplementary motor area subtend duration perception:
An fMRI study. Neuroimage, 19, 1532–1544.
Franssen, V., Vandierendonck, A., & Van Hiel, A. (2006).
Duration estimation and the phonological loop: Articulatory
suppression and irrelevant sounds. Psychological Research,
70, 304–316.
Ghose, G. M., & Maunsell, J. H. (2002). Attentional modulation
in visual cortex depends on task timing. Nature, 419, 616–620.
Griswold, M. A., Jakob, P. M., Heidemann, R. M., Nittka, M.,
Jellus, V., Wang, J., et al. (2002). Generalized autocalibrating
partially parallel acquisitions (GRAPPA). Magnetic Resonance
in Medicine, 47, 1202–1210.
Harrington, D. L., Haaland, K. Y., & Hermanowitz, N. (1998).
Temporal processing in the basal ganglia. Neuropsychology,
12, 3–12.
Harvey, B. M., Dumoulin, S. O., Fracasso, A., & Paul, J. M.
(2020). A network of topographic maps in human association
cortex hierarchically transforms visual timing-selective
responses. Current Biology, 30, 1424–1434.
Hayashi, M. J., Ditye, T., Harada, T., Hashiguchi, M., Sadato, N.,
Carlson, S., et al. (2015). Time adaptation shows duration
selectivity in the human parietal cortex. PLoS Biology, 13,
e1002262.
Hayashi, M. J., van der Zwaag, W., Bueti, D., & Kanai, R. (2018).
Representations of time in human frontoparietal cortex.
Communications Biology, 1, 233.
Herbst, S. K., Chaumon, M., Penney, T. B., & Busch, N. A.
(2015). Flicker-induced time dilation does not modulate
EEG correlates of temporal encoding. Brain Topography,
28, 559–569.
Hutton, C., Josephs, O., Stadler, J., Featherstone, E., Reid, A.,
Speck, O., et al. (2011). The impact of physiological noise
correction on fMRI at 7 T. Neuroimage, 57, 101–112.
Ivry, R. B., & Keele, S. W. (1989). Timing functions of the
cerebellum. Journal of Cognitive Neuroscience, 1, 136–152.
Ivry, R. B., & Richardson, T. C. (2002). Temporal control and
coordination: The multiple timer model. Brain and
Cognition, 48, 117–132.
Jahanshahi, M., Jones, C. R., Zijlmans, J., Katzenschlager, R.,
Lee, L., Quinn, N., et al. (2010). Dopaminergic modulation
of striato-frontal connectivity during motor timing in
Parkinson’s disease. Brain, 133, 727–745.
Janssen, P., & Shadlen, M. N. (2005). A representation of the
hazard rate of elapsed time in macaque area LIP. Nature
Neuroscience, 8, 234–241.
Jech, R., Dušek, P., Wackermann, J., & Vymazal, J. (2005).
Cumulative blood oxygenation-level-dependent signal
changes support the ‘time accumulator’ hypothesis.
NeuroReport, 16, 1467–1471.
Jones, C. R., & Jahanshahi, M. (2011). Dopamine modulates
striato-frontal functioning during temporal processing.
Frontiers in Integrative Neuroscience, 5, 70.
Kanai, R., Paffen, C. L., Hogendoorn, H., & Verstraten, F. A.
(2006). Time dilation in dynamic visual display. Journal of
Vision, 6, 1421–1430.
Kawashima, R., Inoue, K., Sugiura, M., Okada, K., Ogawa, A., &
Fukuda, H. (1999). A positron emission tomography study of
self-paced finger movements at different frequencies.
Neuroscience, 92, 107–112.
Koch, G., Oliveri, M., Carlesimo, G. A., & Caltagirone, C. (2002).
Selective deficit of time perception in a patient with right
prefrontal cortex lesion. Neurology, 59, 1658–1658.
Koch, G., Oliveri, M., Torriero, S., & Caltagirone, C. (2003).
Underestimation of time perception after repetitive
transcranial magnetic stimulation. Neurology, 60,
1844–1846.
Leon, M. I., & Shadlen, M. N. (2003). Representation of time by
neurons in the posterior parietal cortex of the macaque.
Neuron, 38, 317–327.
Lewis, P. A., & Miall, R. C. (2003). Distinct systems for automatic
and cognitively controlled time measurement: Evidence from
neuroimaging. Current Opinion in Neurobiology, 13,
250–255.
Lindbergh, C. A., & Kieffaber, P. D. (2013). The neural
correlates of temporal judgments in the duration bisection
task. Neuropsychologia, 51, 191–196.
Lutti, A., Thomas, D. L., Hutton, C., & Weiskopf, N. (2013).
High-resolution functional MRI at 3 T: 3D/2D echo-planar
Binetti et al.
1379
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
3
p
6
d
9
f
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
a
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
.
/
f
t
o
n
0
5
M
a
y
2
0
2
1
imaging with optimized physiological noise correction.
Magnetic Resonance in Medicine, 69, 1657–1664.
Macar, F., & Vidal, F. (2004). Event-related potentials as indices
of time processing: A review. Journal of Psychophysiology,
18, 89–104.
Malapani, C., Deweer, B., & Gibbon, J. (2002). Separating storage
from retrieval dysfunction of temporal memory in Parkinson’s
disease. Journal of Cognitive Neuroscience, 14, 311–322.
Marinho, V., Oliveira, T., Rocha, K., Ribeiro, J., Magalhães, F.,
Bento, T., et al. (2018). The dopaminergic system dynamic in
the time perception: A review of the evidence. International
Journal of Neuroscience, 128, 262–282.
Matell, M. S., Meck, W. H., & Nicolelis, M. A. (2003). Interval
timing and the encoding of signal duration by ensembles
of cortical and striatal neurons. Behavioral Neuroscience,
117, 760–773.
Matthews, W. J. (2011). How do changes in speed affect the
perception of duration? Journal of Experimental Psychology:
Human Perception and Performance, 37, 1617–1627.
Merchant, H., Harrington, D. L., & Meck, W. H. (2013). Neural
basis of the perception and estimation of time. Annual
Review of Neuroscience, 36, 313–336.
Merchant, H., & Yarrow, K. (2016). How the motor system both
encodes and influences our sense of time. Current Opinion
in Behavioral Sciences, 8, 22–27.
Murai, Y., Whitaker, D., & Yotsumoto, Y. (2016). The centralized
and distributed nature of adaptation-induced misjudgments
of time. Current Opinion in Behavioral Sciences, 8, 117–123.
Nenadic, I., Gaser, C., Volz, H. P., Rammsayer, T., Häger, F., &
Sauer, H. (2003). Processing of temporal information and the
basal ganglia: New evidence from fMRI. Experimental Brain
Research, 148, 238–246.
Noguchi, Y., & Kakigi, R. (2006). Time representations can be
made from nontemporal information in the brain: An MEG
study. Cerebral Cortex, 16, 1797–1808.
Onoe, H., Komori, M., Onoe, K., Takechi, H., Tsukada, H., &
Watanabe, Y. (2001). Cortical networks recruited for time
perception: A monkey positron emission tomography (PET)
study. Neuroimage, 13, 37–45.
Ortuño, F., Guillén-Grima, F., López-García, P., Gómez, J., &
Pla, J. (2011). Functional neural networks of time perception:
Challenge and opportunity for schizophrenia research.
Schizophrenia Research, 125, 129–135.
Peru, A., Pavesi, G., & Campello, M. (2004). Impairment of
executive functions in a patient with a focal lesion in the
anterior cingulate cortex. Evidence from neuropsychological
assessment. Functional Neurology, 19, 107–111.
Pfeuty, M., Ragot, R., & Pouthas, V. (2005). Relationship between
CNV and timing of an upcoming event. Neuroscience Letters,
382, 106–111.
Pouthas, V., Garnero, L., Ferrandez, A. M., & Renault, B. (2000).
ERPs and PET analysis of time perception: Spatial and
temporal brain mapping during visual discrimination tasks.
Human Brain Mapping, 10, 49–60.
Pouthas, V., George, N., Poline, J. B., Pfeuty, M., VandeMoorteele,
P. F., Hugueville, L., et al. (2005). Neural network involved in
time perception: An fMRI study comparing long and short
interval estimation. Human Brain Mapping, 25, 433–441.
Protopapa, F., Hayashi, M. J., Kulashekhar, S., van der Zwaag, W.,
Battistella, G., Murray, M. M., et al. (2019). Chronotopic maps
in human supplementary motor area. PLoS Biology, 17,
e3000026.
Rao, S. M., Harrington, D. L., Haaland, K. Y., Bobholz, J. A.,
Cox, R. W., & Binder, J. R. (1997). Distributed neural systems
underlying the timing of movements. Journal of Neuroscience,
17, 5528–5535.
Rao, S. M., Mayer, A. R., & Harrington, D. L. (2001). The
evolution of brain activation during temporal processing.
Nature Neuroscience, 4, 317–323.
Rosenbaum, D. A. (2002). Time, space, and short-term memory.
Brain and Cognition, 48, 52–65.
Salvioni, P., Murray, M. M., Kalmbach, L., & Bueti, D. (2013).
How the visual brain encodes and keeps track of time.
Journal of Neuroscience, 33, 12423–12429.
Sasaki, K., Yamamoto, K., & Miura, K. (2013). The difference in
speed sequence influences perceived duration. Perception,
42, 198–207.
Schubotz, R. I., & von Cramon, D. Y. (2001). Interval and
ordinal properties of sequences are associated with distinct
premotor areas. Cerebral Cortex, 11, 210–222.
Shuler, M. G., & Bear, M. F. (2006). Reward timing in the
primary visual cortex. Science, 311, 1606–1609.
Treisman, M. (1963). Temporal discrimination and the
indifference interval. Implications for a model of the “internal
clock”. Psychological Monographs, 77, 1–31.
Trojano, L., Caccavale, M., De Bellis, F., & Crisci, C. (2017).
The brain and the subjective experience of time. A voxel
based symptom-lesion mapping study. Behavioural Brain
Research, 329, 26–34.
Watson, A. B., & Pelli, D. G. (1983). QUEST: A Bayesian adaptive
psychometric method. Perception & Psychophysics, 33, 113–120.
Weissman, D. H., Gopalakrishnan, A., Hazlett, C., & Woldorff, M.
(2004). Dorsal anterior cingulate cortex resolves conflict from
distracting stimuli by boosting attention toward relevant
events. Cerebral Cortex, 15, 229–237.
Wiener, M., Turkeltaub, P., & Coslett, H. B. (2010). The image of
time: A voxel-wise meta-analysis. Neuroimage, 49, 1728–1740.
Wing, A. M. (2002). Voluntary timing and brain function:
An information processing approach. Brain and Cognition,
48, 7–30.
Wittmann, M. (2009). The inner experience of time.
Philosophical Transactions of the Royal Society of London,
Series B: Biological Sciences, 364, 1955–1967.
Wittmann, M., & Meissner, K. (2018). The embodiment of
time: How interoception shapes the perception of time.
In M. Tsakiris & H. De Preester (Eds.), The interoceptive
mind: From homeostasis to awareness (p. 63). Oxford:
Oxford University Press.
Wittmann, M., & Paulus, M. P. (2008). Decision making,
impulsivity and time perception. Trends in Cognitive
Sciences, 12, 7–12.
Wittmann, M., Van Wassenhove, V., Craig, B., & Paulus, M. P.
(2010). The neural substrates of subjective time dilation.
Frontiers in Human Neuroscience, 4, 2.
Wu, D., Deng, H., Xiao, X., Zuo, Y., Sun, J., & Wang, Z. (2017).
Persistent neuronal activity in anterior cingulate cortex
correlates with sustained attention in rats regardless of
sensory modality. Scientific Reports, 7, 43101.
Xuan, B., Zhang, D., He, S., & Chen, X. (2007). Larger stimuli
are judged to last longer. Journal of Vision, 7, 1–5.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
o
h
w
t
t
n
p
o
:
a
/
d
/
e
m
d
i
f
r
t
o
p
m
r
c
h
.
s
p
i
l
d
v
i
r
e
e
r
c
t
c
.
m
h
a
i
e
r
d
.
u
c
o
o
m
c
n
/
j
a
o
r
t
c
i
c
n
e
/
-
a
p
r
d
t
i
3
c
2
l
7
e
1
-
3
p
6
d
9
f
2
/
0
3
1
2
3
/
5
7
9
/
2
1
3
o
6
c
9
n
_
/
a
1
_
8
0
6
1
1
5
6
5
7
7
9
p
/
d
j
o
b
c
y
n
g
_
u
a
e
_
s
0
t
1
o
5
n
5
0
7
7
.
p
S
d
e
f
p
e
b
m
y
b
e
g
r
u
2
e
0
s
2
t
3
/
j
f
t
/
.
o
n
0
5
M
a
y
2
0
2
1
1380
Journal of Cognitive Neuroscience
Volume 32, Number 7