Transcranial Cerebellar Direct Current Stimulation
Enhances Verb Generation but Not Verb
Naming in Poststroke Aphasia
Paola Marangolo1,2, Valentina Fiori2, Carlo Caltagirone2,3,
Francesca Pisano1, and Alberto Priori4
Abstrait
■ Although the role of the cerebellum in motor function is
well recognized, its involvement in the lexical domain remains
to be further elucidated. En effet, it has not yet been clarified
whether the cerebellum is a language structure per se or whether
it contributes to language processing when other cognitive com-
ponents (par exemple., cognitive effort, working memory) are required by
the language task. Neuromodulation studies on healthy partici-
pants have suggested that cerebellar transcranial direct current
stimulation (tDCS) is a valuable tool to modulate cognitive func-
tion. Cependant, so far, only a single case study has investigated
whether cerebellar stimulation enhances language recovery in
aphasic individuals. In a randomized, crossover, double-blind de-
sign, we explored the effect of cerebellar tDCS coupled with lan-
guage treatment for verb improvement in 12 aphasic individuals.
Each participant received cerebellar tDCS (20 min, 2 mA) in four
experimental conditions: (1) right cathodal and (2) sham stimu-
lation during a verb generation task and (3) right cathodal and
(4) sham stimulation during a verb naming task. Each ex-
perimental condition was run in five consecutive daily sessions
over 4 weeks. At the end of treatment, a significant improve-
ment was found after cathodal stimulation only in the verb gen-
eration task. No significant differences were present for verb
naming among the two conditions. We hypothesize that cere-
bellar tDCS is a viable tool for recovery from aphasia but only
when the language task, such as verb generation, also demands
the activation of nonlinguistic strategies. ■
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
un
_
0
1
2
0
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
INTRODUCTION
During the past two decades, converging neuroscientific
evidence has largely documented that the human cere-
bellum contributes to a much wider range of higher-level
cerebral functions than previously accepted. En effet,
bien que, traditionally, there has been a unanimous
agreement that the cerebellum is primarily involved in
autonomic and somatic motor processes (Leiner, 2010;
Schmahmann, 2010; Strick, Dum, & Fiez, 2009; De Smet,
Baillieux, De Deyn, Mariën, & Paquier, 2007; Holmes,
1939), particularly after aphasia reports, there has been
a rapidly increasing interest in the cerebellum’s role in
cognition (Reeber, Otis, & Sillitoe, 2013; Manto & Haines,
2012; Strick et al., 2009). En effet, several linguistic dis-
orders after acquired cerebellar lesions have been docu-
mented (De Smet et al., 2007), such as impaired verbal
fluency (Meinzer, Yetim, McMahon, & de Zubicaray, 2016;
Stoodley & Schmahmann, 2009; Richter et al., 2007;
Leggio, Silveri, Petrosini, & Molinari, 2000; Schmahmann &
Sherman, 1998; Molinari, Leggio, & Silveri, 1997; Appollonio,
Grafman, Schwartz, Massaquoi, & Hallett, 1993; Akshoomoff,
1Università Federico II, Naples, Italy, 2IRCCS Fondazione Santa
Lucia, Rome, Italy, 3Università degli Studi di Roma Tor Vergata,
Rome, Italy, 4Università degli Studi di Milano, Milan, Italy
© 2017 Massachusetts Institute of Technology
Courchesne, Presse, & Iragui, 1992), agrammatism
(Schmahmann & Sherman, 1998; Molinari et al., 1997;
Mariën et al., 1996), and naming difficulties (Fabbro,
Moretti, & Bava, 2000; Gasparini et al., 1999; Schmahmann
& Sherman, 1998). On the basis of these findings, quelques
authors have assumed that the cerebellum represents an
“inter-area functional coordinator subserving precisely
timed sequential organization of verbal sentences” (Zettin,
Cappa, D’amico, Rago, & Perino, 1997; Silveri, Leggio, &
Molinari, 1994). En effet, this process might be compro-
mised in patients with cerebellar lesions (Molinari et al.,
1997). Several other cases of aphasia, predominantly, as a
result of right cerebellar lesions, characterized by prevailing
verbal fluency disturbances, have been described (Mariën
& Beaton, 2014; Stoodley & Schmahmann, 2009; Mariën,
Engelborghs, Pickut, & De Deyn, 2000; Gasparini et al.,
1999). The frequent co-occurrence of a right cerebellar
lesion and aphasia led some authors to hypothesize the
existence of a “lateralized linguistic cerebellum” (Mariën
et coll., 1996, 2000, 2014). According to Mariën et al. (1996,
2000, 2014), the aphasic disorder reflects a “diaschisis”
phenomenon whereby the damage of the right cerebel-
lum causes a hypofunction of the left frontal cortical areas,
“home” of our language representation (Mariën et al.,
1996, 2000, 2014). The cerebellum would thus have a role
Journal des neurosciences cognitives 30:2, pp. 188–199
est ce que je:10.1162/jocn_a_01201
in linguistic representation but only through its connec-
tions with the left frontal cerebral language areas (mais
see Gasparini et al., 1999). In line with this hypothesis,
several neuroimaging reports on healthy participants have
confirmed the activation of the posterior lateral area of
the right cerebellum together with an activation of the
left frontal cortex during different linguistic tasks (Chen,
Ho, & Desmond, 2014; Stoodley & Schmahmann, 2009;
McDermott, Petersen, Watson, & Ojemann, 2003; Gurd
et coll., 2002; Ojemann et al., 1998; Schlösser et al., 1998),
but many conclusions about the role of the cerebellum
in language originate from applying word generation
tasks (Stoodley, Valera, & Schmahmann, 2010, 2012; Frings
et coll., 2006; Petersen, Fox, Posner, Mintun, & Raichle,
1989). Petersen and coworkers (1989) reported the first
nonmotor linguistic PET activation study in which par-
ticipants were requested to produce a verb semantically
associated to a presented noun. In contrast to the control
condition in which the nouns only had to be read or
merely repeated, the verb generation condition activated
the right lateral cerebellum and a number of left frontal
régions. En effet, this task, which reflects the capacity to
generate words according to a given semantic category,
requires a large amount of cognitive effort, and it is gen-
erally considered to depend on a close cooperation be-
tween verbal, executive, and working memory functions,
which rely on frontal lobes (Stoodley & Schmahmann,
2009; Bellebaum & Daum, 2007; Gottwald, Mihajlovic,
Wilde, & Mehdorn, 2003; Schmahmann & Sherman, 1998;
Grafman et al., 1992). Thompson-Schill and colleagues
(1998) have suggested that verb generation is a more “dif-
ficult” task than naming because it requires the selection
of a response from among multiple competitors due to
the association strength between an object (noun) and its
corresponding verbs (par exemple., knife → “cut,” “spread,” “sharp,»
“stab”).
Despite variations on the original task design, several
other studies have consistently reproduced activation of
the right lateral cerebellum during word generation tasks
(Grabowski et al., 1996; Martine, Haxby, Lalonde, Wiggs,
& Ungerleider, 1995; Raichle et al., 1994). Leiner, Leiner,
and Dow (1989) interpreted the simultaneous activation
of the right cerebellum and the left Broca’s language area
during word generation as “the reflection of accelerated
transmission of signals between these two centers during
word finding.” Consistent with this assumption, quelques
neuroimaging studies on aphasic individuals have pro-
vided evidence in favor of a close connection between
the right cerebellar activity and the activation of the
contralateral left frontal regions. En effet, aphasic patients
showed an abnormal response from the right cerebellum
due to the absence of inputs from the damaged left
frontal regions (Connor et al., 2006) and a reactivation
of the right cerebellar area after language recovery due
to a recruitment of the left perilesional frontal cortex
(Marangolo et al., 2016; Heath et al., 2013; Szaflarski,
Allendorfer, Banks, Vannest, & Holland, 2013).
Parallel to this increasing interest in the role of the
cerebellum in cognition, in more recent years, non-
invasive brain stimulation techniques, such as trans-
cranial direct current stimulation (tDCS), have been used
to modulate cognitive functions, such as working memory,
attention, et la langue (Lefaucheur et al., 2016; Nitsche
& Paulus, 2011).
The assumption on which tDCS is based is that a con-
stant, weak, and continuous current is able to alter the
firing rate of the neurons. It has been proposed that
cerebellar tDCS is most likely to produce its effects by
polarizing Purkinje cells (see Van Dun, Bodranghien,
Mariën, & Manto, 2016; Pope & Miall, 2014) and changing
the levels/pattern of activity in the deep cerebellar output
nuclei, thereby also affecting distant plasticity in human
cortical areas (Grimaldi et al., 2016; Van Dun et al., 2016).
En effet, it has been shown that, whereas anodal stimu-
lation, through its excitatory effects, increases the dis-
charge from the Purkinje cells, augmenting the inhibition
of the facilitatory pathways from the cerebellar nuclei to
the cerebral cortex, cathodal stimulation exerts the oppo-
site effect, through a disinhibition of Purkinje cells, et
activates the frontal cerebral cortex (Pope, 2015; Pope &
Miall, 2012; Galea & Celnik, 2009).
Pope and Miall (2012) have suggested that one crucial
factor for cerebellar tDCS impact is task difficulty. In their
étude, three groups of 22 participants each performed
the paced auditory serial addition task and a variant of
this task called the paced auditory serial subtraction task
(PASST), together with a verb generation task, before
and after anodal, cathodal, or sham tDCS over the right
cerebellum. The authors reported an effect on the dif-
ficult PASST but not on the easier paced auditory serial
addition task. Fait intéressant, an improvement in the PASST
and a reduction in verbal response latencies in verb gen-
eration were observed after cathodal right cerebellar
tDCS, whereas no effect of anodal stimulation was found
(Pope & Miall, 2012). According to Pope and Miall (2012),
right cerebellum stimulation has influenced working
memory and attention abilities differently depending on
task difficulty. Ainsi, the cerebellum is capable of releasing
cognitive resources by disinhibition of the left prefrontal
régions, enhancing performance only when the task is
cognitively demanding (Pope & Miall, 2012).
Contrary to these findings, in a group of healthy partic-
ipants, Turkeltaub, Swears, D’Mello, and Stoodley (2016)
showed that both anodal and cathodal stimulation over
the right cerebellum improves word generation but the
effects were found using a different task, namely, a pho-
nemic fluency task. Following Pope and Miall’s suggestion
(Pope & Miall, 2012, 2014), the authors hypothesized that
cerebellar tDCS did not act directly on the language func-
tion per se but on the executive control and response
selection components required by the generation task
(Turkeltaub et al., 2016).
À ce jour, only a single case study has investigated
whether cerebellar tDCS leads to recovery from aphasia.
Marangolo et al.
189
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
un
_
0
1
2
0
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
In a patient with a large bilateral frontoparietal and insu-
lar infarct, Sebastian et al. (2016) found that both anodal
and sham coupled with language treatment resulted in im-
proved spelling to dictation for trained and untrained words
immediately after and 2 months post-treatment but
the improvement was greater with anodal tDCS than with
sham especially for untrained items. Although the results
are interesting and suggest a therapeutic potential of cer-
ebellar tDCS for language recovery, we believe that any
final conclusion deserves further investigations. En effet,
as the authors also pointed out, a crucial limitation of their
study was that it includes a single case with a large bilateral
dommage, which is not a lesion typically observed in the apha-
sic population. En outre, in their experimental design,
active tDCS followed sham; thus, any extra benefits of tDCS
might be due to having a second treatment after already
having had the first treatment (Sebastian et al., 2016).
Dans cette étude, we aimed to verify the role of cerebellar
tDCS in language processing in a group of 12 aphasic par-
ticipants with left unilateral damage by contrasting two dif-
ferent language tasks with different demands in terms of
cognitive effort: a verb naming (VN) task and a verb gen-
eration task. En effet, with respect to VN in which the pro-
duction of the correct answer is facilitated by the presented
picture, verb generation, because of some combination of
both retrieval and competition demands (Snyder, Banich,
& Munakata, 2011), relies on different cognitive strategies
(Ackermann, Mathiak, & Riecker, 2007; Justus, Ravizza,
Fiez, & Ivry, 2005).
Because cathodal stimulation, reducing the inhibition
of the Purkinje cells, favors increased excitability of the
left frontal language areas (Pope & Miall, 2014; Connor
et coll., 2006), in the present work, two experimental con-
ditions were used: right cathodal and sham cerebellar
tDCS. On the basis of previous findings, we expected
to find that cathodal stimulation would lead to a greater
improvement in verb retrieval with respect to the sham
condition only in the verb generation task.
MÉTHODES
Participants
Twelve left-brain-damaged participants (six men and six
femmes) with chronic aphasia were included in the study
(voir la figure 1). Inclusion criteria were native Italian
conférencier, premorbid right-handedness (Oldfield, 1971),
a single left-hemispheric stroke at least 6 months before
the investigation, mild nonfluent aphasia with no articu-
latory difficulties, preserved basic comprehension skills
(so as to allow them to be engaged in verbal exchanges
with the therapist), and no attentive or memory deficits
that might bias their performance. The data analyzed in
the current study were collected in accordance with the
Declaration of Helsinki and the institutional review board
of the IRCCS Fondazione Santa Lucia, Rome, Italy. Before
participation, all patients signed informed consent forms.
Clinical Data
The aphasic disorders were assessed using standardized
language tests (the Battery for the Analysis of Aphasic
Disorders test; Miceli, Laudanna, Burani, & Capasso, 1994;
De Renzi & Vignolo, 1962). All patients were classified as
nonfluent aphasics as they had reduced verbal output
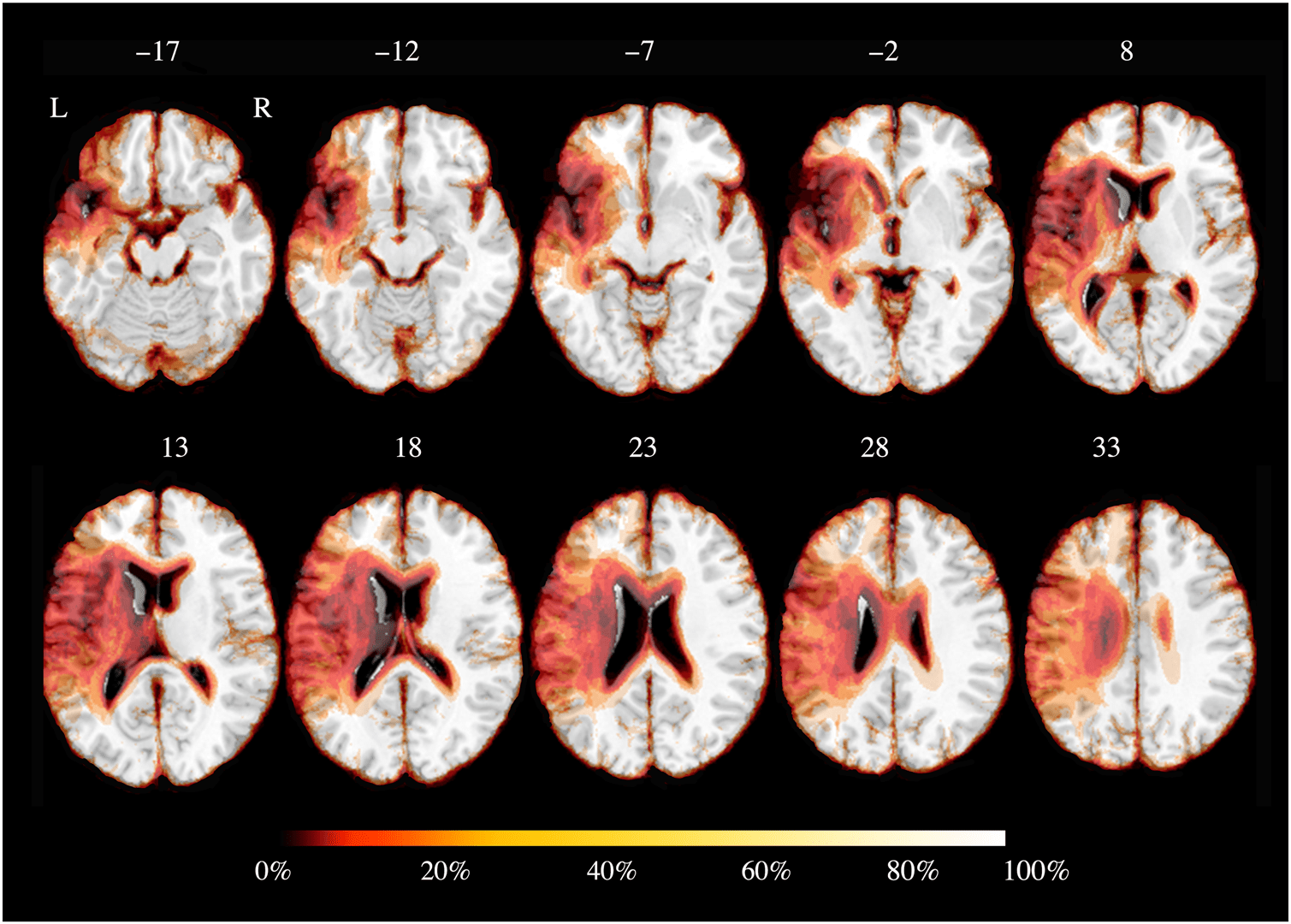
Chiffre 1. Brain parenchyma
overlap across patients. Color
bar refers to the amount of
saved voxels, implying 0%
being related to the total
absence of tissue and 100%
being related to the total
presence of tissue. Comme indiqué,
lesion extent included the
temporal lobe, the inferior
frontal gyrus, the insula, et,
partially, the postcentral
and precentral gyrus. Axial
coordinates refer to the
standard space (MNI152).
190
Journal des neurosciences cognitives
Volume 30, Nombre 2
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
un
_
0
1
2
0
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
in spontaneous speech. Their utterances were short,
and they were mainly characterized by omissions of verbs
as well as errors in verb inflection. Their basic compre-
hension skills were preserved, but they still have difficulty
in comprehending complex materials (mean = 17/36,
cutoff score = 29/36, <29 impaired performance on the
Token test; De Renzi & Vignolo, 1962). In the noun and
VN task, moderate-to-severe word finding difficulties
were still present. A neuropsychological battery of tests
was also administered, which excluded presence of
attention (i.e., alertness, sustained and selective atten-
tion), working memory digit span), executive
function deficits that might confound data (Zimmermann
& Fimm, 1994; Spinnler Tognoni, 1987; see Table 1).
Materials
Sixty pictures verbs to sing, write) for VN
task 60 nouns associated a correspondent verb
(i.e., pen → verb generation task were
selected. For each stimuli were subdivided
into two lists 30 items, matched frequency
(VN list 1: mean =28,
SD SD =31; VN 2: > .05 for each
comparison) and length ( VN list 1: mean = 8, SD = 1;
VN list 2: mean = 8, SD = 2; nouns list 1: mean = 7,
SD = 2; nouns list 2: mean = 7, SD = 2; unpaired t test,
p > .05 for each comparison; Bertinetto et al., 2005). Le
lists were also matched for imageability (estimated on the
basis of a sample of 30 healthy participants along a 7-point
scale, depuis 1 = no imageability to 7 = clear imageability
[VN list 1: mean = 6, SD = 1; VN list 2: mean = 6, SD = 1;
nouns list 1: mean = 6, SD = 1; nouns list 2: mean = 6,
SD = 1; unpaired t test, p > .05 for each comparison]).
The correlations between the above variables were not
significant among the lists suggesting that each measure
represented an independent attribute (frequency vs.
length, VN list 1: r = −.22, p = .25; frequency vs. image-
ability, VN list 1: r = −.13, p = .50; imageability vs. length,
VN list 1: r = −.08, p = .66; frequency vs. length, VN list 2:
r = −.29, p = .12; frequency vs. imageability, VN list 2: r =
−.23, p = .23; imageability vs. length, VN list 2: r = .03, p =
.86; frequency vs. length, nouns list 1: r = −.28, p = .14;
frequency vs. imageability, nouns list 1: r = −.23, p = .23;
imageability vs. length, nouns list 1: r = .29, p = .12; fre-
quency vs. length, nouns list 2: r = −.32, p = .09; fre-
quency vs. imageability, nouns list 2: r = .13, p = .50;
imageability vs. length, nouns list 2: r = .18; p = .34).
Procedure
Cerebellar tDCS
tDCS was applied using a battery-driven EMS (Bologna,
Italy) programmable direct current stimulator with a pair
of surface-soaked sponge electrodes (5 × 7 cm). A con-
stant current of 2-mA intensity was applied through the
cathode on the right cerebellar cortex, 1 cm under and
4 cm lateral to the inion (approximately comparable with
the projection of the cerebellar lobule VII into the scalp)
pour 20 min, whereas the reference electrode was posi-
tioned over the right shoulder on the deltoid muscle
(Pope & Miall, 2012). If applied according to safety guide-
lines, tDCS is considered to be a safe brain stimulation
technique with minor adverse effects (Lefaucheur et al.,
2016; Fregni et al., 2015). For each task ( VN vs. verb gen-
eration), two different stimulation conditions were car-
ried out: (1) cathodal and (2) sham. Sham stimulation
was performed exactly like the cathodal condition, mais
the stimulator was turned off after 30 sec (Gandiga,
Hummel, & Cohen, 2006). Ainsi, we had four different
experimental conditions: (1) right cathodal cerebellar
tDCS for VN, (2) sham for VN, (3) right cathodal cerebel-
lar tDCS for verb generation, et (4) sham for verb gen-
eration. For each task, le 60 stimuli were subdivided
into two lists of 30 items, each matched for frequency,
length, and imageability. The assignment of each list of stim-
uli was randomized across the two conditions (cathodal vs.
sham). All patients underwent the four experimental con-
ditions whose order was randomized across participants.
To ensure the double-blind procedure, both the experi-
menter and the patient were blinded regarding the stimu-
lation condition, and the stimulator was turned on/off
by another person. At the end of each experimental condi-
tion, participants were asked if they were aware of which
condition (real or sham) they were in. We inferred that
all participants well tolerated the stimulation by inter-
preting their spontaneous report as well as the results
from a questionnaire completed by the participant at
the end of each experimental condition (see Fertonani,
Rosini, Cotelli, Rossini, & Miniussi, 2010, for the question-
naire). Itch was the most commonly reported sensation
with light (16% of the participants) to moderate (83% de
the participants) intensity. Participants reported that the
sensation started at the beginning of the stimulation and
stopped after a few minutes both during the real and/or
sham stimulation. Ainsi, none of the participants was able
to distinguish between the two conditions. A paired t test
did not show any significant difference in the participants’
perception of sensation between the real and sham con-
ditions ( p > .05).
Treatment
Once the electrodes were placed, participants performed
the two tasks while they received 20 min of cerebellar
tDCS. Each stimulation condition was performed in five
consecutive daily sessions over 1 week with 6 days of
intersession interval. The order of item presentation
was randomized across sessions. During the VN task,
participants were asked to name aloud each picture that
appeared on the PC screen (screen size = 15 in., viewing
distance = 1 m) pour 20 sec preceded by a fixation point,
which lasted 800 msec (see also Fiori et al., 2013, for a
similar procedure). If the participant failed or did not
Marangolo et al.
191
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
un
_
0
1
2
0
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
4
5
:
)
t
o
t
(
t
t
un
d
e
n
je
un
t
s
toi
S
6
6
:
)
t
o
t
(
t
t
un
e
v
je
t
c
e
e
S
je
0
1
0
1
5
:
M.
W
9
9
:
)
t
o
t
(
s
s
e
n
t
r
e
UN
je
6
3
/
4
1
0
0
1
0
0
1
4
6
0
9
s
h
t
n
o
m
3
r
un
e
oui
1
3
1
6
7
:
)
t
o
t
(
t
t
un
d
e
n
je
un
t
s
toi
S
4
9
:
)
t
o
t
(
t
t
un
e
v
je
t
c
e
e
S
je
4
3
:
)
t
o
t
(
t
t
un
d
e
n
je
un
t
s
toi
S
8
2
:
)
t
o
t
(
t
t
un
e
v
je
t
c
e
e
S
je
4
:
M.
W
0
3
:
)
t
o
t
(
s
s
e
n
t
r
e
UN
je
6
3
/
4
1
0
0
1
0
0
1
0
2
0
3
s
h
t
n
o
m
0
1
r
un
e
oui
1
8
5
2
.
9
4
:
M.
W
5
6
:
)
t
o
t
(
s
s
e
n
t
r
e
UN
je
6
3
/
5
1
0
0
1
0
0
1
0
4
5
4
s
h
t
n
o
m
7
r
un
e
oui
1
3
1
0
7
:
)
t
o
t
(
t
t
un
d
e
n
je
un
t
s
toi
S
0
6
:
)
t
o
t
(
t
t
un
e
v
je
t
c
e
e
S
je
5
,
2
1
5
:
M.
W
9
8
:
)
t
o
t
(
s
s
e
n
t
r
e
UN
je
6
3
/
1
2
0
0
1
0
0
1
7
6
7
7
s
h
t
n
o
m
9
r
un
e
oui
1
8
1
5
7
,
0
1
5
:
M.
W
5
3
:
)
t
o
t
(
s
s
e
n
t
r
e
UN
je
6
3
/
4
1
0
0
1
0
0
1
4
6
7
6
s
h
t
n
o
m
9
s
r
un
e
oui
2
3
1
0
3
:
)
t
o
t
(
t
t
un
d
e
n
je
un
t
s
toi
S
5
3
:
)
t
o
t
(
t
t
un
e
v
je
t
c
e
e
S
je
5
,
1
1
5
:
M.
W
5
9
:
)
t
o
t
(
s
s
e
n
t
r
e
UN
je
6
3
/
1
2
0
0
1
0
0
1
5
6
0
7
s
h
t
n
o
m
9
r
un
e
oui
1
3
1
0
8
:
)
t
o
t
(
t
t
un
d
e
n
je
un
t
s
toi
S
0
7
:
)
t
o
t
(
t
t
un
e
v
je
t
c
e
e
S
je
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
/
j
/
o
c
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
un
_
0
1
2
0
1
p
d
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
5
7
:
)
t
o
t
(
t
t
un
d
e
n
je
un
t
s
toi
S
5
5
:
)
t
o
t
(
t
t
un
e
v
je
t
c
e
e
S
je
5
7
,
9
4
:
M.
W
0
6
:
)
t
o
t
(
s
s
e
n
t
r
e
UN
je
6
3
/
4
1
t
s
e
T
g
n
je
t
r
o
S
s
'
je
g
je
e
W
,
0
5
,
4
=
F
F
o
t
toi
C
(
)
d
e
r
je
un
p
m
je
0
5
,
4
<
=
f
f
o
t
u
C
(
M
W
,
2
±
5
s
e
i
t
i
l
i
b
A
l
a
n
o
i
t
n
e
t
t
A
e
l
i
t
n
e
c
r
e
P
n
i
s
e
r
o
c
S
(
=
f
f
o
t
u
C
(
T
T
,
6
3
/
9
2
)
d
e
r
i
a
p
m
I
5
<
)
d
e
r
i
a
p
m
I
5
<
)
d
e
r
i
a
p
m
I
9
2
<
C
V
)
%
(
0
0
1
C
N
)
%
(
0
0
1
N
V
)
%
(
0
2
N
N
)
%
(
8
4
s
r
a
e
y
2
6
1
t
s
o
P
e
m
i
T
t
e
s
n
O
l
e
v
e
L
c
u
d
E
)
s
r
a
e
Y
(
e
g
A
)
s
r
a
e
Y
(
s
t
n
e
i
t
a
P
c
i
s
a
h
p
A
t
n
e
u
l
f
n
o
N
2
1
e
h
t
f
o
a
t
a
D
l
a
c
i
n
i
l
C
d
n
a
c
i
h
p
a
r
g
o
m
e
d
o
i
c
o
S
.
1
e
l
b
a
T
0
5
1
6
6
4
5
6
8
6
7
5
0
7
P
1
2
3
4
5
6
7
192
Journal of Cognitive Neuroscience
Volume 30, Number 2
1
1
0
1
5
7
,
9
5
:
M
W
4
9
:
)
t
o
t
(
s
s
e
n
t
r
e
A
l
6
3
/
4
1
0
0
1
0
0
1
0
4
3
3
s
h
t
n
o
m
2
r
a
e
y
1
8
1
5
5
:
)
t
o
t
(
t
t
a
d
e
n
i
a
t
s
u
S
0
5
:
)
t
o
t
(
t
t
a
e
v
i
t
c
e
e
S
l
0
6
:
)
t
o
t
(
t
t
a
d
e
n
i
a
t
s
u
S
4
8
:
)
t
o
t
(
t
t
a
e
v
i
t
c
e
e
S
l
5
:
M
W
7
9
:
)
t
o
t
(
s
s
e
n
t
r
e
A
l
6
3
/
3
2
0
0
1
0
0
1
0
3
0
3
s
h
t
n
o
m
6
r
a
e
y
1
8
8
4
1
5
8
9
1
8
:
)
t
o
t
(
t
t
a
d
e
n
i
a
t
s
u
S
0
5
:
)
t
o
t
(
t
t
a
e
v
i
t
c
e
e
S
l
5
,
0
1
5
:
M
W
8
7
:
)
t
o
t
(
s
s
e
n
t
r
e
A
l
6
3
/
4
1
0
0
1
0
0
1
5
2
3
4
h
t
n
o
m
5
r
a
e
y
1
0
1
1
6
1
1
5
7
,
0
1
6
:
M
W
0
9
:
)
t
o
t
(
s
s
e
n
t
r
e
A
l
6
3
/
4
1
0
0
1
0
0
1
1
7
0
7
h
t
n
o
m
1
s
r
a
e
y
3
3
1
6
5
2
1
=
T
T
;
)
4
9
9
1
,
.
l
a
t
e
i
l
e
c
i
M
:
t
s
e
t
s
r
e
d
r
o
s
i
D
c
i
s
a
h
p
A
’
e
r
a
s
e
r
o
c
s
w
a
r
,
t
n
e
i
t
a
p
h
c
a
e
r
o
F
:
)
7
8
9
1
,
i
n
o
n
g
o
T
&
l
r
e
n
n
p
S
i
;
n
o
i
s
r
e
v
n
a
i
l
a
t
I
(
t
s
e
t
g
n
i
t
r
o
s
s
l
’
g
i
e
W
l
w
o
e
b
s
e
r
o
c
s
;
e
l
i
t
n
e
c
r
e
p
n
i
d
e
t
r
o
p
e
r
e
r
a
s
e
r
o
c
s
s
t
n
e
i
t
a
p
:
)
4
9
9
1
,
i
m
m
F
&
n
n
a
m
r
e
m
m
Z
(
i
s
e
i
t
i
l
i
b
a
l
a
n
o
i
t
n
e
t
t
A
.
y
r
o
m
e
m
g
n
k
r
o
w
=
M
W
i
;
n
o
i
t
n
e
t
t
a
=
t
t
a
;
)
2
6
9
1
,
l
o
o
n
g
i
V
&
i
z
n
e
R
e
D
(
6
3
/
9
2
=
f
f
o
t
u
c
,
t
s
e
t
n
e
k
o
T
.
)
7
8
9
1
,
i
n
o
n
g
o
T
&
l
r
e
n
n
p
S
i
;
7
8
9
1
,
.
l
a
t
e
i
n
i
s
r
O
,
n
a
p
s
t
i
g
i
d
,
.
e
.
i
(
d
e
r
i
a
p
m
i
d
e
r
e
d
i
s
n
o
c
e
r
a
e
l
i
t
n
e
c
r
e
p
h
t
f
i
f
e
h
t
.
d
e
t
r
o
p
e
r
2
6
:
)
t
o
t
(
t
t
a
d
e
n
i
a
t
s
u
S
5
8
:
)
t
o
t
(
t
t
a
e
v
i
t
c
e
e
S
l
f
o
s
i
s
y
l
a
n
A
e
h
t
r
o
f
y
r
e
t
t
a
B
(
n
o
i
s
n
e
h
e
r
p
m
o
c
b
r
e
v
=
C
V
;
n
o
i
s
n
e
h
e
r
p
m
o
c
n
u
o
n
=
C
N
i
;
g
n
m
a
n
n
u
o
n
=
N
N
;
l
e
v
e
l
l
a
n
o
i
t
a
c
u
d
e
=
l
e
v
e
L
c
u
d
E
;
s
t
n
a
p
i
c
i
t
r
a
p
=
P
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
a
_
0
1
2
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Marangolo et al.
193
0
8
:
)
t
o
t
(
t
t
a
d
e
n
i
a
t
s
u
S
0
6
:
)
t
o
t
(
t
t
a
e
v
i
t
c
e
e
S
l
6
:
M
W
0
9
:
)
t
o
t
(
s
s
e
n
t
r
e
A
l
6
3
/
3
2
0
0
1
0
0
1
0
6
5
5
s
h
t
n
o
m
6
r
a
e
y
1
3
1
0
6
0
1
answer within 20 sec, the corresponding written name was
presented below the picture for 5 sec and the participant
was asked to read the word aloud. The pair of stimuli re-
mained on the screen until the participant read the word or
5 sec elapsed. In all cases, participants were able to
correctly read the word. During the verb generation task,
the examiner orally presented a noun (i.e., trampoline) and
participants were required to produce within 20 sec the
most appropriate corresponding verb (i.e., to jump). Be-
cause previous studies have reported activation of the right
lateral cerebellum during a verb generation task indepen-
dently of the input modality (Richter et al., 2004; Petersen
et al., 1989), we chose to orally present the noun to pre-
vent participants from “task errors” such as reading the
noun by presenting a written noun (Thompson-Schill et al.,
1998) or naming the object by presenting the object
picture (Kurland, Reber, & Stokes, 2014).
For both tasks, the examiner manually recorded the
response on a separate sheet. If the participant did not
respond within the 20-sec interval, the program auto-
matically presented the subsequent picture or noun.
Vocal RTs were calculated from the presentation of the
picture or the noun to the pronunciation of the first
phoneme through Audacity 2.1.2 Software. RTs were re-
corded only if the participants responded within 20 sec.
Data Analysis
Before the experiment, the two lists of stimuli were pre-
sented to a group of 30 healthy individuals (15 men and
15 women) matched for age (40–75 years) and educa-
tional level (13–17 years) to the aphasic group. Each par-
ticipant was asked to produce for each presented verb for
the VN task and noun for the verb generation task the
most appropriate corresponding verb, with no inter-
ference from the examiner. Only those verbs that elicited
at least 80% of agreement among participants in the two
lists were considered correct responses and therefore
used for a response accuracy analysis in the aphasic
group.
Data were analyzed with STATISTICA 10 (StatSoft, Inc.,
Tulsa, OK). Statistical analyses were performed with two
separate ANOVAs, respectively, for response accuracies
and vocal RTs with three within-participant factors: Task
(verb generation vs. VN), Condition (cathodal stimulation
vs. sham), and Time (baseline [T0] vs. end of the treat-
ment [T5] vs. follow-up [FU]). If the ANOVA showed sig-
nificant effects, respective post hoc Bonferroni tests were
conducted.
RESULTS
Accuracy
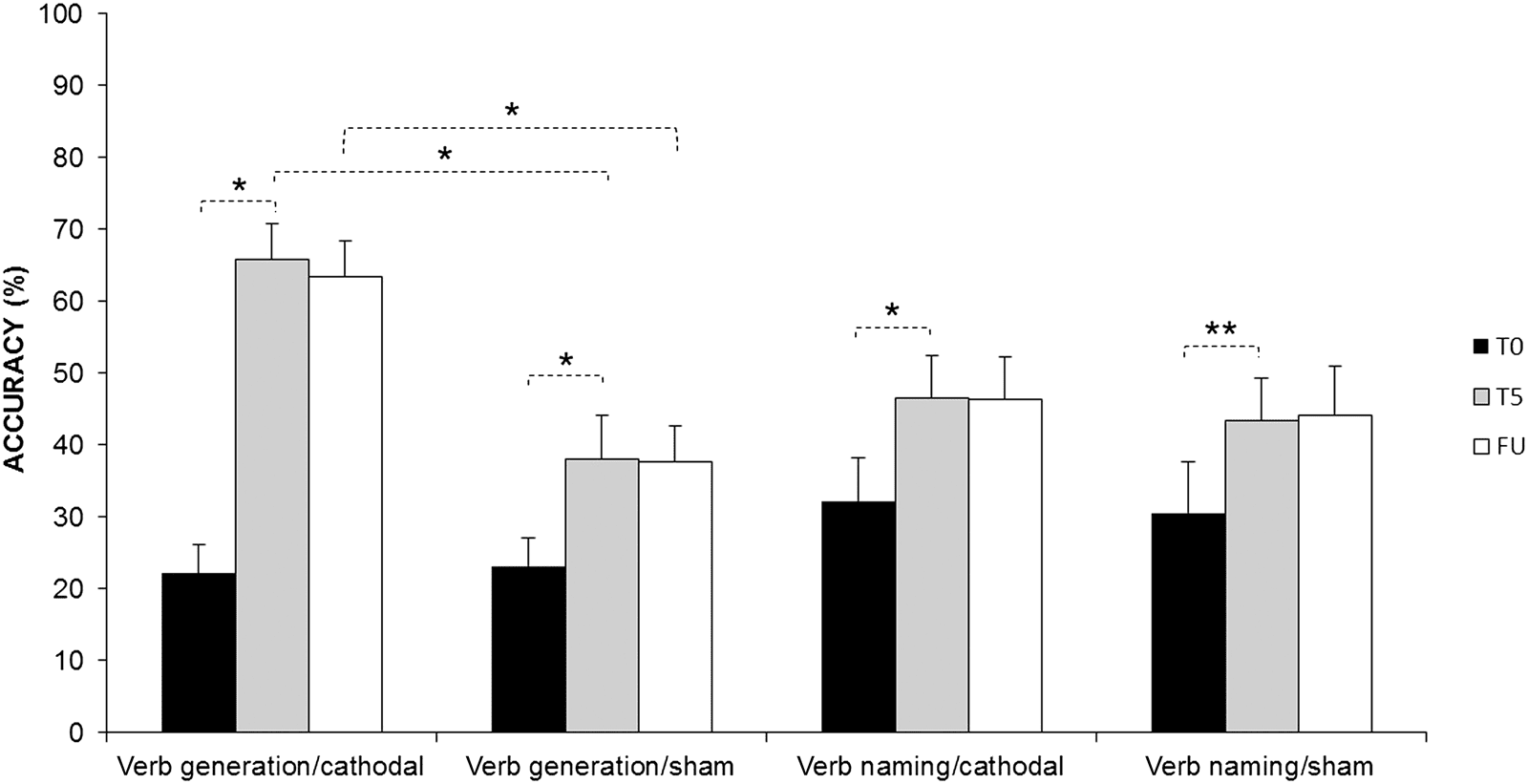
The analysis showed a significant effect of Condition (F(1,
11) = 13.88, p < .01) and Time (F(2, 22) = 77.94, p <
.001). The interaction Task × Condition × Time was also
significant (F(2, 22) = 16.21, p < .001). Indeed, although
all experimental conditions led to a significant greater
percentage of correct responses at the end of treatment
(T5) compared with the baseline (T0; verb generation:
difference between T5 and T0, cathodal = 44%, p <
.001; sham = 15%, p = .001; VN: difference between
T5 and T0, cathodal = 15%, p = .001; sham = 12%, p =
.007), at the end of treatment, only in the verb generation
task, the percentage of correct responses was greater after
cathodal stimulation compared with sham (cathodal vs.
sham = 28%, p < .001), and this difference persisted at
FU (cathodal vs. sham = 25%, p < .001). No differences
between the two stimulation conditions were found for
the naming task (cathodal vs. sham: T5 = 4%, p = 1;
cathodal vs. sham: FU = 2%, p = 1; see Figure 2).
Figure 2. Mean percentage of response accuracy for verb generation and VN task at baseline (T0), at the end of the treatment (T5), and at FU for the
cathodal and sham conditions, respectively (*p ≤ .001, **p < .01). Error bars represent SEM.
194
Journal of Cognitive Neuroscience
Volume 30, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
a
_
0
1
2
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Table 2. Mean Number of Errors in the Verb Generation and VN Task for Cathodal and Sham tDCS, Respectively (±SD)
Verb Generation, Cathodal
Verb Generation, Sham
VN, Cathodal
VN, Sham
Type of Errors
No responses
Semantic paraphasias
Unrelated verb responses
T0
19 (±5)
3 (±2)
2 (±2)
T5
7 (±3)
2 (±1)
2 (±2)
T0
T5
T0
T5
T0
T5
18 (±6)
13 (±7)
17 (±6)
13 (±6)
18 (±7)
12 (±6)
3 (±2)
2 (±2)
3 (±2)
3 (±3)
2 (±1)
2 (±2)
2 (±2)
3 (±2)
1 (±1)
1 (±2)
1 (±1)
1 (±2)
To further investigate if tDCS had a different impact
on the participant’s response, we classified the errors
made by each participant in all experimental conditions.
As shown in Table 2, errors were (1) no responses, (2)
semantic paraphasias, and (3) unrelated verb responses,
but at baseline (T0), for both the VN task and the verb gen-
eration task, errors were predominantly “no responses.”
Thus, we conducted an ANOVA on the number of “no
responses” with three within-participant factors: Task
(verb generation vs. VN), Condition (cathodal vs. sham),
and Time (baseline [T0] vs. end of the treatment [T5]).
The analysis revealed a significant interaction of Task ×
Condition × Time (F(1, 11) = 8.29, p = .01). Indeed,
although all experimental conditions led to a lower num-
ber of “no responses” at the end of treatment (T5) com-
pared with the baseline (T0; verb generation: difference
between T5 and T0, cathodal = 12, p < .001; sham = 5,
p = .04; VN: difference between T5 and T0, cathodal =
5, p = .04; sham = 6, p = .03), at the end of treatment
(T5), only in the verb generation task, the number of “no
responses” was lower after cathodal stimulation compared
with sham (cathodal vs. sham = −6, p = .02). No dif-
ferences between the two stimulation conditions were
found for the VN task (cathodal vs. sham: T5 = 1, p = 1).
Thus, these results resembled those previously found for
the accuracy data.
Vocal RTs
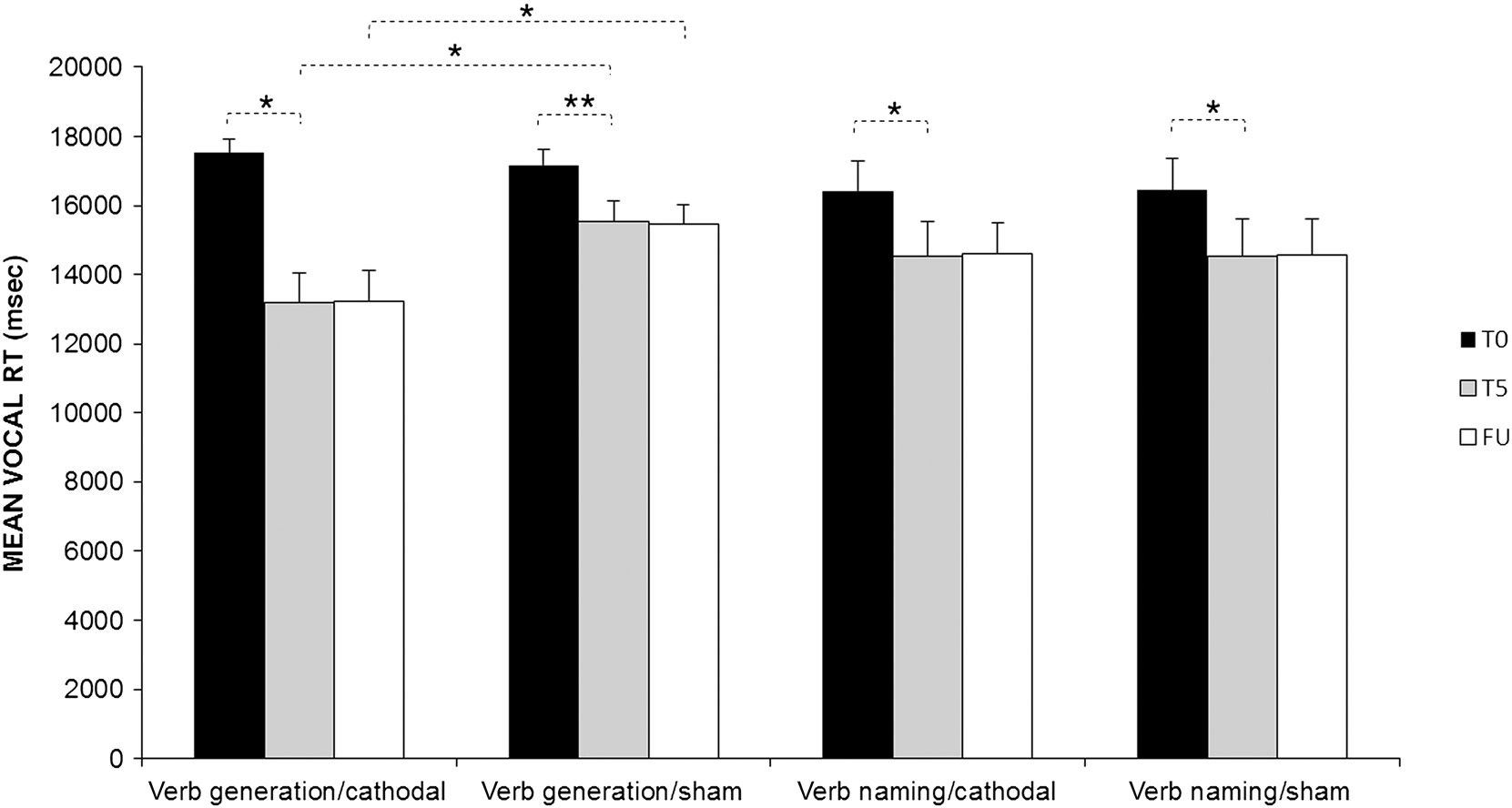
The analysis showed a significant effect of Condition (F(1,
11) = 6.20, p = .03) and Time (F(2, 22) = 40.24, p <
.001). The interaction Task × Condition × Time was also
significant (F(2, 22) = 11.12, p < .001). Indeed, although
all experimental conditions led to faster vocal RTs at the
end of treatment (T5) compared with baseline (T0; verb
generation: difference between T5 and T0, cathodal =
4334 msec, p < .001; sham = 1644 msec, p = .003; VN:
difference between T5 and T0, cathodal = 1897 msec, p =
.001; sham = 1932 msec, p < .001), at the end of treat-
ment, only in the verb generation task, vocal RTs were
faster after cathodal stimulation compared with sham
(cathodal vs. sham = −2350 msec, p < .001), and this dif-
ference persisted at FU (cathodal vs. sham = −2250 msec,
p < .001). No differences between the two stimulation
conditions were found for the VN task (cathodal vs. sham:
T5 = 10 msec, p = 1; cathodal vs. sham: FU = 23 msec,
p = 1; see Figure 3).
Figure 3. Mean vocal RTs for verb generation and VN at baseline (T0), at the end of the treatment (T5), and at FU for the cathodal and sham
conditions, respectively (*p ≤ .001, **p < .01). Error bars represent SEM.
Marangolo et al.
195
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
a
_
0
1
2
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
DISCUSSION
The aim of this study was to investigate whether cere-
bellar tDCS coupled with language training improves
verb retrieval in nonfluent chronic aphasic individuals.
Our findings showed that cathodal stimulation differ-
ently affected verb recovery depending on the language
task. Indeed, at the end of treatment, only the verb
generation task led to a significant improvement in
verb retrieval. Moreover, FU testing showed that these
effects lasted over 1 week after the intervention. This
specificity argues against an effect simply due to en-
hanced cognitive arousal, which should have influenced
both language tasks.
As stated in the Introduction, several studies have
already supported the hypothesis that the cerebellum
plays a role in language processing but depends on task
demands (Pope & Miall, 2012, 2014; Stoodley et al., 2010,
2012; Ackermann et al., 2007). Ackermann et al. (2007)
have argued that nonlinguistic aspects of task perfor-
mance, such as the amount of effort or the degree of
automaticity, might account for cerebellar involvement
during verb generation tasks. Similarly, Stoodley and
Schmahmann (2009) have claimed that the cerebellum
takes part not in the language function per se but only
when the task is cognitively demanding and therefore en-
gages other cognitive components, such as working mem-
ory and/or executive functions (Stoodley & Schmahmann,
2009). Indeed, apart from motor control and higher-
order aspects of speech production, a variety of studies
point to a contribution of the cerebellum to executive
and memory tasks (Ackermann et al., 2007). Because
the paradigm of verb generation involves the production
and selection of different verbal responses (Thompson-
Schill et al., 1998), prearticulatory rehearsal processes are
engaged as well, which rely to working memory processes
(Ackermann et al., 2007; Helmuth, Ivry, & Shimizu, 1997).
Indeed, our choice to directly compare cerebellar tDCS
effects in two verb production tasks was made taking into
account the substantial differences between the two tasks.
Like confrontation naming, verb generation is a seman-
tic association task in which the participant has to pro-
duce a verb strictly associated to a given noun. Much of
the cognitive demand between the two tasks is shared,
including semantic and lexical retrieval processes and
the planning, execution, and monitoring of speech pro-
duction (e.g., Levelt, 1989). However, whereas verb gen-
eration requires the patient to creatively link a noun to a
verb choosing among competing response alternatives
(Thompson-Schill et al., 1998), in VN, the correct answer
is univocally determined by the presented picture and
the task is one of the earliest linguistic skills developmen-
tally mastered and thus is an overlearned task (Herholz
et al., 1997). Interestingly, although verb generation is a
task more cognitive demanding than VN and persons
with aphasia generally experience greatest difficulty with
verb generation (Martin & Cheng, 2006; Thompson-Schill
et al., 1998), our aphasic patients benefited only for this
task after right cerebellar cathodal stimulation.
Although our data are only behavioural, we might spec-
ulate that right cathodal cerebellar stimulation, through a
disinhibition of the Purkinje cells, has favored the engage-
ment of the left frontal areas, which, in turn, enhanced the
activation of executive and memory components required
by the verb generation task (Pope & Miall, 2014; Connor
et al., 2006; Mariën, Engelborghs, Fabbro, & De Deyn,
2001; Mariën et al., 1996). Indeed, most of our patients
had a partial damage to the left frontal areas (see Figure 1);
thus, the hypothesis can be advanced that subregions of
the left frontal cortex took part in verb recovery. Accord-
ingly, several studies have already shown that the same
facilitatory patterns may be observed in a verbal fluency
task after cathodal cerebellar tDCS or anodal stimulation
over the frontal cortex (Pope & Miall, 2012; Iyer et al.,
2005). Confirming evidence for a functional relationship
between the left frontal cortex and the cerebellum comes
also from a recent study combining bilateral tDCS and
resting state fMRI in a group of left-brain-damaged popu-
lation (Marangolo et al., 2016). Indeed, in nine chronic
aphasic patients, Marangolo et al. (2016) found that bilat-
eral anodic stimulation over the left inferior frontal area
and cathodal contralesional stimulation over its right homo-
logue coupled with an intensive language treatment led
to functional connectivity changes within the left damaged
hemisphere, together with the cerebellum (Marangolo
et al., 2016). In agreement with our hypothesis that the
cerebellum is functionally connected to the language net-
work, very recently, D’Mello, Turkeltaub, and Stoodley
(2017) acquired behavioral and resting state fMRI data, dur-
ing a sentence completion task, before and after cerebellar
tDCS in a group of healthy adults. Relative to sham, anodal
tDCS increased activation in the right cerebellum only
when the preceding context in the sentence modulated
the predictability of the target word (predictive sentences).
In the same study (D’Mello et al., 2017), functional connec-
tivity changes were also found in the left language areas,
including the left inferior frontal gyrus. Thus, these data
showed that cerebellar neuromodulation specifically alters
activation patterns during semantic prediction tasks
(D’Mello et al., 2017). Similarly, in our work, cerebellar
tDCS improved the generation of highly predictable verbs
semantically associated to the presented nouns.
It might be finally argued that the effects found were an
artifact of linguistic variables, so that, at the end of treatment,
the verbs produced in the generation task had higher
frequency and/or were shorter (in terms of number of
phonemes) than the verbs produced in the naming task.
However, statistical analyses performed to control for those
factors did not show any significant difference between
the correct responses given in the two tasks (verb genera-
tion: mean frequency = 32, SD = 9; mean length = 8,
SD = 1; VN: mean frequency = 31, SD = 8; mean length = 8,
SD = 0; t tests, p = .72 and p = .69, respectively, for
frequency and length).
196
Journal of Cognitive Neuroscience
Volume 30, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
a
_
0
1
2
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
In conclusion, although our data deserve further inves-
tigations, they suggest that cerebellar tDCS might be a
viable tool for enhancing language recovery in chronic
aphasia. Because our results point to potential thera-
peutic benefits of cerebellar stimulation only for complex
language tasks, we believe that these findings have im-
portant implications for aphasia. Indeed, they address
the possibility that the cerebellum supports cognitive
functions that are important for language recovery.
Acknowledgments
We are extremely grateful to Dr. Tommaso Gili for his help in
drawing Figure 1.
Reprint requests should be sent to Paola Marangolo, Diparti-
mento di Studi Umanistici, Università degli Studi di Napoli
Federico II, Via Porta di Massa, 1, 80133 Napoli, Italia, or via
e-mail: paola.marangolo@gmail.com.
REFERENCES
Ackermann, H., Mathiak, K., & Riecker, A. (2007). The
contribution of the cerebellum to speech production and
speech perception: Clinical and functional imaging data.
Cerebellum, 6, 202–213.
Akshoomoff, N. A., Courchesne, E., Press, G. A., & Iragui, V.
(1992). Contribution of the cerebellum to
neuropsychological functioning: Evidence from a case of
cerebellar degenerative disorder. Neuropsychologia, 30,
315–328.
Appollonio, I. M., Grafman, J., Schwartz, V., Massaquoi, S., &
Hallett, M. (1993). Memory in patients with cerebellar
degeneration. Neurology, 43, 1536–1544.
Bellebaum, C., & Daum, I. (2007). Cerebellar involvement in
executive control. Cerebellum, 6, 184–192.
Bertinetto, P. M., Burani, C., Laudanna, A., Marconi, L., Ratti, D.,
Rolando, C., et al. (2005). Corpus e Lessico di Frequenza
dell’Italiano Scritto (CoLFIS). Istituto di Linguistica
Computazionale, Unità staccata di Genova. http://linguistica.
sns.it/CoLFIS/CoLFIS home.htm.
Chen, S.-H. A., Ho, M.-H. R., & Desmond, J. E. (2014). A meta-
analysis of cerebellar contributions to higher cognition from
PET and fMRI studies. Human Brain Mapping, 35, 593–615.
Connor, L. T., Braby, T. D., Snyder, A. Z., Lewis, C., Blasi, V., &
Corbetta, M. (2006). Cerebellar activity switches hemispheres
with cerebral recovery in aphasia. Neuropsychologia, 44,
171–177.
De Renzi, E., & Vignolo, L. A. (1962). The Token test: A sensitive
test to detect receptive disturbances in aphasics. Brain, 85,
665–678.
De Smet, H. J., Baillieux, H., De Deyn, P. P., Mariën, P., &
Paquier, P. (2007). The cerebellum and language: The story
so far. Folia Phoniatrica et Logopaedica, 59, 165–170.
D’Mello, A. M., Turkeltaub, P. E., & Stoodley, C. J. (2017).
Cerebellar tDCS modulates neural circuits during semantic
prediction: A combined tDCS-fMRI study. Journal of
Neuroscience, 37, 1604–1613.
Fabbro, F., Moretti, R., & Bava, A. (2000). Language
impairments in patients with cerebellar lesions. Journal of
Neurolinguistics, 13, 173–188.
Fertonani, A., Rosini, S., Cotelli, M., Rossini, P. M., & Miniussi, C.
(2010). Naming facilitation induced by transcranial direct
current stimulation. Behavioural Brain Research, 208,
311–318.
Fiori, V., Cipollari, S., Di Paola, M., Razzano, C., Caltagirone, C.,
& Marangolo, P. (2013). tDCS stimulation segregates words
in the brain: Evidence from aphasia. Frontiers in Human
Neuroscience, 7, 269.
Fregni, F., Nitsche, M. A., Loo, C. K., Brunoni, A. R., Marangolo,
P., Leite, J., et al. (2015). Regulatory considerations for the
clinical and research use of transcranial direct current
stimulation (tDCS): Review and recommendations from an
expert panel. Clinical Research and Regulatory Affairs, 32,
22–35.
Frings, M., Dimitrova, A., Schorn, C. F., Elles, H. G., Hein-Kropp,
C., Gizewski, E. R., et al. (2006). Cerebellar involvement in
verb generation: An fMRI study. Neuroscience Letters, 409,
19–23.
Galea, J. M., & Celnik, P. (2009). Brain polarization enhances
the formation and retention of motor memories. Journal of
Neurophysiology, 102, 294–301.
Gandiga, P. C., Hummel, F. C., & Cohen, L. G. (2006).
Transcranial DC stimulation (tDCS): A tool for double-blind
sham-controlled clinical studies in brain stimulation. Clinical
Neurophysiology, 117, 845–850.
Gasparini, M., Di Piero, V., Ciccarelli, O., Cacioppo, M. M.,
Pantano, P., & Lenzi, G. L. (1999). Linguistic impairment after
right cerebellar stroke: A case report. European Journal of
Neurology, 6, 353–356.
Gottwald, B., Mihajlovic, Z., Wilde, B., & Mehdorn, H. M. (2003).
Does the cerebellum contribute to specific aspects of
attention? Neuropsychologia, 41, 1452–1460.
Grabowski, T. J., Frank, R. J., Brown, C. K., Damasio, H., Ponto,
L. L., Watkins, G. L., et al. (1996). Reliability of PET activation
across statistical methods, subject groups, and sample sizes.
Human Brain Mapping, 4, 23–46.
Grafman, J., Litvan, I., Massaquoi, S., Stewart, M., Sirigu, A., &
Hallett, M. (1992). Cognitive planning deficit in patients with
cerebellar atrophy. Neurology, 42, 1493–1496.
Grimaldi, G., Argyropoulos, G. P., Bastian, A., Cortes, M., Davis,
N. J., Edwards, D. J., et al. (2016). Cerebellar transcranial
direct current stimulation (ctDCS): A novel approach to
understanding cerebellar function in health and disease.
Neuroscientist, 22, 83–97.
Gurd, J. M., Amunts, K., Weiss, P. H., Zafiris, O., Zilles, K.,
Marshall, J. C., et al. (2002). Posterior parietal cortex is
implicated in continuous switching between verbal fluency
tasks: An fMRI study with clinical implications. Brain, 125,
1024–1038.
Heath, S., McMahon, K. L., Nickels, L., Angwin, A., MacDonald,
A. D., van Hees, S., et al. (2013). Facilitation of naming in
aphasia with auditory repetition: An investigation of
neurocognitive mechanisms. Neuropsychologia, 51, 1534–1548.
Helmuth, L. L., Ivry, R. B., & Shimizu, N. (1997). Preserved
performance by cerebellar patients on tests of word
generation, discrimination learning, and attention. Learning
and Memory, 3, 456–474.
Herholz, K., Reulen, H. J., von Stockhausen, H. M., Thiel, A.,
Ilmberger, J., Kessler, J., et al. (1997). Preoperative activation
and intraoperative stimulation of language-related areas in
patients with glioma. Neurosurgery, 41, 1253–1260.
Holmes, G. (1939). The cerebellum of man. Brain, 62, 1–30.
Iyer, M. B., Mattu, U., Grafman, J., Lomarev, M., Sato, S., &
Wassermann, E. M. (2005). Safety and cognitive effect of
frontal DC brain polarization in healthy individuals.
Neurology, 64, 872–875.
Justus, T., Ravizza, S. M., Fiez, J. A., & Ivry, R. B. (2005).
Reduced phonological similarity effects in patients with
damage to the cerebellum. Brain and Language, 95,
304–318.
Marangolo et al.
197
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
a
_
0
1
2
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Kurland, J., Reber, A., & Stokes, P. (2014). Beyond picture
naming: Norms and patient data for a verb-generation task.
American Journal of Speech-Language Pathology, 23,
S259–S270.
Lefaucheur, J.-P., Antal, A., Ayache, S. S., Benninger, D. H.,
Brunelin, J., Cogiamanian, F., et al. (2016). Evidence-based
guidelines on the therapeutic use of transcranial direct
current stimulation (tDCS). Clinical Neurophysiology, 128,
56–92.
Leggio, M. G., Silveri, M. C., Petrosini, L., & Molinari, M. (2000).
Phonological grouping is specifically affected in cerebellar
patients: A verbal fluency study. Journal of Neurology,
Neurosurgery and Psychiatry, 69, 102–106.
Leiner, H. C. (2010). Solving the mystery of the human
cerebellum. Neuropsychology Review, 20, 229–235.
Leiner, H. C., Leiner, A. L., & Dow, R. S. (1989). Reappraising
the cerebellum: What does the hindbrain contribute to
the forebrain? Behavioral Neuroscience, 103, 998–1008.
Levelt, W. J. M. (1989). Speaking: From intention to
articulation. Cambridge, MA: MIT Press.
Manto, M., & Haines, D. (2012). Cerebellar research: Two
centuries of discoveries. Cerebellum, 11, 446–448.
Marangolo, P., Fiori, V., Sabatini, U., De Pasquale, G., Razzano,
C., Caltagirone, C., et al. (2016). Bilateral transcranial
direct current stimulation language treatment enhances
functional connectivity in the left hemisphere: Preliminary
data from aphasia. Journal of Cognitive Neuroscience, 28,
724–738.
Mariën, P., Ackermann, H., Adamaszek, M., Barwood, C. H. S.,
Beaton, A., Desmond, J., et al. (2014). Consensus paper:
Language and the cerebellum: An ongoing enigma. Cerebellum,
13, 386–410.
Mariën, P., & Beaton, A. (2014). The enigmatic linguistic
cerebellum: Clinical relevance and unanswered questions
on nonmotor speech and language deficits in cerebellar
disorders. Cerebellum & Ataxias, 1, 12.
Mariën, P., Engelborghs, S., Fabbro, F., & De Deyn, P. P. (2001).
The lateralized linguistic cerebellum: A review and a new
hypothesis. Brain and Language, 79, 580–600.
Mariën, P., Engelborghs, S., Pickut, B. A., & De Deyn, P. P.
(2000). Aphasia following cerebellar damage: Fact or fallacy?
Journal of Neurolinguistics, 13, 145–171.
Mariën, P., Saerens, J., Nanhoe, R., Moens, E., Nagels, G., Pickut,
B. A., et al. (1996). Cerebellar induced aphasia: Case report of
cerebellar induced prefrontal aphasic language phenomena
supported by SPECT findings. Journal of the Neurological
Sciences, 144, 34–43.
Martin, A., Haxby, J. V., Lalonde, F. M., Wiggs, C. L., &
Ungerleider, L. G. (1995). Discrete cortical regions associated
with knowledge of color and knowledge of action. Science,
270, 102–105.
Martin, R. C., & Cheng, Y. (2006). Selection demands versus
association strength in the verb generation task.
Psychonomic Bulletin & Review, 13, 396–401.
McDermott, K. B., Petersen, S. E., Watson, J. M., & Ojemann,
J. G. (2003). A procedure for identifying regions preferentially
activated by attention to semantic and phonological
relations using functional magnetic resonance imaging.
Neuropsychologia, 41, 293–303.
Meinzer, M., Yetim, Ö., McMahon, K., & de Zubicaray, G.
(2016). Brain mechanisms of semantic interference in spoken
word production: An anodal transcranial direct current
stimulation (atDCS) study. Brain and Language, 157–158,
72–80.
Miceli, G., Laudanna, A., Burani, C., & Capasso, R. (1994).
Batteria per l’Analisi del Deficit Afasico. BADA [BADA A
battery for the assessment of aphasic disorders]. Roma,
Italia: CEPSAG.
Molinari, M., Leggio, M. G., & Silveri, M. C. (1997). Verbal fluency
and agrammatism. International Review of Neurobiology,
41, 325–339.
Nitsche, M. A., & Paulus, W. (2011). Transcranial direct current
stimulation—Update 2011. Restorative Neurology and
Neuroscience, 29, 463–492.
Ojemann, J. G., Buckner, R. L., Akbudak, E., Snyder, A. Z.,
Ollinger, J. M., McKinstry, R. C., et al. (1998). Functional MRI
studies of word-stem completion: Reliability across
laboratories and comparison to blood flow imaging with PET.
Human Brain Mapping, 6, 203–215.
Oldfield, R. C. (1971). The assessment and analysis of handedness:
The Edinburgh inventory. Neuropsychologia, 9, 97–113.
Orsini, A., Grossi, D., Capitani, E., Laiacona, M., Papagno, C., &
Vallar, G. (1987). Verbal and spatial immediate memory span:
Normative data from 1355 adults and 112 children. Italian
Journal of Neuroscience, 8, 539–548.
Petersen, S. E., Fox, P. T., Posner, M. I., Mintun, M., & Raichle,
M. E. (1989). Positron emission tomographic studies of the
processing of singe words. Journal of Cognitive Neuroscience,
1, 153–170.
Pope, P. A. (2015). Modulating cognition using transcranial
direct current stimulation of the cerebellum. Journal of
Visualized Experiments, 96, e52302.
Pope, P. A., & Miall, R. C. (2012). Task-specific facilitation of
cognition by cathodal transcranial direct current stimulation
of the cerebellum. Brain Stimulation, 5, 84–94.
Pope, P. A., & Miall, R. C. (2014). Restoring cognitive functions
using non-invasive brain stimulation techniques in patients
with cerebellar disorders. Frontiers in Psychiatry, 5, 1–7.
Raichle, M. E., Fiez, J. A., Videen, T. O., MacLeod, A. M., Pardo,
J. V., Fox, P. T., et al. (1994). Practice-related changes in
human brain functional anatomy during nonmotor learning.
Cerebral Cortex, 4, 8–26.
Reeber, S. L., Otis, T. S., & Sillitoe, R. V. (2013). New roles for
the cerebellum in health and disease. Frontiers in Systems
Neuroscience, 7, 83.
Richter, S., Gerwig, M., Aslan, B., Wilhelm, H., Schoch, B.,
Dimitrova, A., et al. (2007). Cognitive functions in patients
with MR-defined chronic focal cerebellar lesions. Journal of
Neurology, 254, 1193–1203.
Richter, S., Kaiser, O., Hein-Kropp, C., Dimitrova, A., Gizewski,
E., Beck, A., et al. (2004). Preserved verb generation in patients
with cerebellar atrophy. Neuropsychologia, 42, 1235–1246.
Schlösser, R., Hutchinson, M., Joseffer, S., Rusinek, H., Saarimaki,
A., Stevenson, J., et al. (1998). Functional magnetic resonance
imaging of human brain activity in a verbal fluency task.
Journal of Neurology, Neurosurgery and Psychiatry, 64,
492–498.
Schmahmann, J. D. (2010). The role of the cerebellum in
cognition and emotion: Personal reflections since 1982 on
the dysmetria of thought hypothesis, and its historical
evolution from theory to therapy. Neuropsychology Review,
20, 236–260.
Schmahmann, J. D., & Sherman, J. C. (1998). The cerebellar
cognitive affective syndrome. Brain, 121, 561–579.
Sebastian, R., Saxena, S., Tsapkini, K., Faria, A. V., Long, C.,
Wright, A., et al. (2016). Cerebellar tDCS: A novel approach to
augment language treatment post-stroke. Frontiers in Human
Neuroscience, 10, 695.
Silveri, M. C., Leggio, M. G., & Molinari, M. (1994). The
cerebellum contributes to linguistic production: A case of
agrammatic speech following a right cerebellar lesion.
Neurology, 44, 2047–2050.
Snyder, H. R., Banich, M. T., & Munakata, Y. (2011). Choosing
our words: Retrieval and selection processes recruit shared
neural substrates in left ventrolateral prefrontal cortex.
Journal of Cognitive Neuroscience, 23, 3470–3482.
198
Journal of Cognitive Neuroscience
Volume 30, Number 2
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
a
_
0
1
2
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Spinnler, H., & Tognoni, G. (1987). Standardizzazione e taratura
italiana di test neuropsicologici. Milan: Masson Italia Periodici.
Stoodley, C. J., & Schmahmann, J. D. (2009). The cerebellum
and language: Evidence from patients with cerebellar
degeneration. Brain and Language, 110, 149–153.
Stoodley, C. J., Valera, E. M., & Schmahmann, J. D. (2010). An
fMRI study of intra-individual functional topography in the
human cerebellum. Behavioural Neurology, 23, 65–79.
Stoodley, C. J., Valera, E. M., & Schmahmann, J. D. (2012).
Functional topography of the cerebellum for motor and
cognitive tasks: An fMRI study. Neuroimage, 16, 1560–1570.
Strick, P. L., Dum, R. P., & Fiez, J. A. (2009). Cerebellum and
nonmotor function. Annual Review of Neuroscience, 32,
413–434.
Szaflarski, J. P., Allendorfer, J. B., Banks, C., Vannest, J., &
Holland, S. K. (2013). Recovered vs. not-recovered from post-
stroke aphasia: The contributions from the dominant and
non-dominant hemispheres. Restorative Neurology and
Neuroscience, 31, 347–360.
Thompson-Schill, S. L., Swick, D., Farah, M. J., D’Esposito,
M., Kan, I. P., & Knight, R. T. (1998). Verb generation in
patients with focal frontal lesions: A neuropsychological test
of neuroimaging findings. Proceedings of the National
Academy of Sciences, U.S.A., 95, 15855–15860.
Turkeltaub, P. E., Swears, M. K., D’Mello, A. M., & Stoodley, C. J.
(2016). Cerebellar tDCS as a novel treatment for aphasia?
Evidence from behavioral and resting-state functional
connectivity data in healthy adults. Restorative Neurology
and Neuroscience, 34, 491–505.
Van Dun, K., Bodranghien, F. C. A. A., Mariën, P., & Manto,
M. U. (2016). tDCS of the cerebellum: Where do we stand in
2016? Technical issues and critical review of the literature.
Frontiers in Human Neuroscience, 10, 199.
Zettin, M., Cappa, S., D’amico, A., Rago, R., & Perino, C.
(1997). Agrammatic speech production after a right
cerebellar haemorrhage. Neurocase, 3, 375–380.
Zimmermann, P., & Fimm, B. (1994). Tests d’évaluation de
l’attention (TEA). Würselen, Germany: Psytest.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
/
j
/
o
c
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
3
0
2
1
8
8
1
7
8
7
1
1
8
/
j
o
c
n
_
a
_
0
1
2
0
1
p
d
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Marangolo et al.
199