Trait Aggressiveness Modulates Neurophysiological
Correlates of Laboratory-induced Reactive
Aggression in Humans
Ulrike M. Kra¨mer1, Sarah Bu¨ttner1, Gerhard Roth2,
and Thomas F. Mu¨nte1
D
o
w
n
je
o
un
d
e
d
Abstrait
& Reactive aggression following provocation is a frequent
form of human social behavior. The neural basis of reactive
aggression, especially its control, remains poorly understood,
cependant. We conducted an event-related potential (ERP) étude
using a competitive reaction time task that elicits aggression
through provocation. Participants were selected from a larger
sample because of extreme scores in trait aggressiveness,
yielding high and low trait aggressive groups. As each trial in
the task is separated into a decision phase, during which the
punishment level for the opponent is set, and an outcome
phase, during which the punishment is applied or received, nous
were able to disentangle provocation-related and evaluation-
related modulations of the ERPs during the aggressive inter-
action. Spécifiquement, we observed an enhanced frontal negativity
during the decision phase under high provocation that was
positively correlated with the participants’ ability to refrain
from retaliation. This held true for high trait aggressive par-
ticipants only, pointing to a higher need for inhibitory and
control processes in these people when provoked. During the
outcome phase, we detected a mediofrontal negativity in loss
compared to win trials, resembling previous ERP findings to
negative feedback stimuli, which have been linked to the eval-
uation of an outcome’s valence. This mediofrontal negativity
was differentially pronounced in aggressive and nonaggressive
participants: Nonaggressive participants showed only a slightly
smaller mediofrontal negativity in win than in loss trials, sug-
gesting that for them punishing the opponent had a similar
negative valence as being punished. &
INTRODUCTION
Aggression is a common behavior in both humans and
animals that can be defined as ‘‘any behavior directed
toward the goal of harming or injuring another living
being that is motivated to avoid such treatment’’ (Baron,
1977). Although neuroscientific research on aggression
has had a strong focus on the relationship between psy-
chopathology (par exemple., antisocial personality disorder) et
brain regions (Blair, 2004; Anderson, Bechara, Damasio,
Tranel, & Damasio, 1999), aggressive behavior is fre-
quent also among neurologically and psychologically
healthy people, asking for an explanation of its causes
and underlying mechanisms.
In a recent review, Anderson and Bushman (2002)
presented a general model of aggressive behavior (Gen-
eral Aggression Model [GAM]), stating that situational
variables (par exemple, provocation or pain), ensemble
with personal variables (such as traits, sex, etc.), influ-
ence aggressive behavior through the mediating effects
de la cognition, affect, and arousal. The outcome of these
cognitive and affective processes are appraisal and deci-
1Otto-von-Guericke-University, Magdeburg, Allemagne, 2Univer-
sity of Bremen, Bremen, Allemagne
sion processes, which finally lead to either thoughtful or
impulsive actions, depending on the invested amount of
cognitive control and effort. The GAM accounts for the
differential effects of, par exemple, cognitive primes (tel
as pictures of weapons) or pain on aggressive behavior
and affective reactions (Lindsay & Anderson, 2000). Comment-
jamais, simply observing an aggressive action reveals little
about the underlying motivations and decision pro-
cesse (Bushman & Anderson, 2001) or about its neural
underpinnings. The present study is thus aimed at
tapping into the decision and evaluation processes
during an aggressive social interaction by taking a closer
look at the time course of neurophysiological responses
(event-related potentials, ERPs).
A reliable method to elicit aggression in an experi-
mental setting is a competitive reaction time task, lequel
entails punishment of the loser by the winner as sug-
gested by Taylor (1967). Through provocation (selection
of different punishment levels by the opponent), le
Taylor Aggression Paradigm (TAP) seeks to elicit ag-
gression, operationalized as the intensity of punishment
administered to the opponent. The paradigm’s conver-
gent validity, as well as its discriminant and external
validity, has been found to be high (Anderson, Lindsay,
& Bushman, 1999; Giancola & Zeichner, 1995; Bernstein,
D 2008 Massachusetts Institute of Technology
Journal des neurosciences cognitives 20:8, pp. 1464–1477
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
0
je
8
e
–
1
p
4
d
6
F
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
.
t
/
.
.
F
o
n
1
8
M.
un
oui
2
0
2
1
Richardson, & Hammock, 1987). As each single trial
comprises a decision phase during which the participant
is required to set the prospective punishment for the op-
ponent, and an outcome phase during which the punish-
ment is applied or received, the paradigm in conjunction
with neurophysiological recordings enables us to delin-
eate the neural correlates of the different decision and
evaluation processes underlying reactive aggression.
In two recent functional magnetic resonance imaging
(IRMf) études, modified versions of the TAP were used
to characterize the neural correlates of social reactive ag-
gression (Kra¨mer, Jansma, Tempelmann, & Mu¨nte, 2007;
Lotze, Veit, Anders, & Birbaumer, 2007). Presenting one
opponent who turned from nice to unfair during the
experiment, Lotze et al. (2007) could demonstrate dif-
ferent roles of the ventral and dorsal medial prefrontal
cortex in reactive aggression, with the former thought to
be related to affective processes such as compassion and
the latter involved in cognitive processes elicited by
more intense social interaction processes. By introduc-
ing two opponents—one highly and one less provoking—
Kra¨mer et al. (2007) were able to dissociate general so-
cial interaction processes and cognitive and motivational
mechanisms specific for reactive aggression, reflected
in activations in the dorsal and rostral parts of the ante-
rior cingulate gyrus (ACC) and the striatum.
Although, up to now, no ERP study has taken a direct
look at the temporal dynamics of an aggressive inter-
action, studies related to social interaction, inhibition,
and the processing of feedback hint at the ERP com-
ponents that could be of interest for the present study.
In a first pilot study employing the TAP (unpublished
data), we observed modulations of specific ERP compo-
nents during the decision phase. En particulier, we de-
tected provocation-related differences in the time range
of the N2 (maximum at 250–300 msec) and in a later
right fronto-lateral negativity (400–600 msec).
The N2, a fronto-centrally distributed component, is ob-
served in experimental settings calling for an inhibition
of motor responses as in go/no-go tasks (Falkenstein,
Hoormann, & Hohnsbein, 1999; Pfefferbaum, Ford, Weller,
& Kopell, 1985) or in situations of stimulus–response in-
compatibility (van Veen & Carter, 2002). Evidence from
animal and human studies suggest that the N2 in go/no-
go tasks reflects inhibitory mechanisms emanating from
areas in the prefrontal cortex (Pliszka, Liotti, & Woldorff,
2000; Sasaki & Gemba, 1986). Children with attention-
deficit/hyperactivity disorder showed a reduced right
frontal N2 in a stop-signal task, related to their core def-
icit in inhibitory control (Pliszka et al., 2000). Results of
an fMRI study suggest a widely distributed cortical net-
work including the dorsolateral and inferior frontal lobe
underlying response inhibition (Garavan, Ross, & Stein,
1999). D'autre part, tasks calling for the moni-
toring for response competition, such as the Eriksen
Flanker task, give rise to a fronto-central negativity that
similarly has been termed N2 but for which the ante-
rior cingulate gyrus has been identified as the likely
generator (Nieuwenhuis, Yeung, van den Wildenberg, &
Ridderinkhof, 2003; van Veen & Carter, 2002). In the
paradigm used in the present study, provocation may
lead to both a higher need for monitoring processes and
inhibition of predominant aggressive reactions during
the selection of the prospective punishment. This is
suggested by the GAM, which refers to cognitive control
and monitoring processes that might prevent an aggres-
sive retaliation (Anderson & Bushman, 2002). De plus,
the results from a previous fMRI study on reactive
aggression (Kra¨mer et al., 2007) demonstrated a recruit-
ment of cognitive control processes in response to a
higher level of provocation. Donc, N2-like negativ-
ities are of special interest in the decision phase.
For the outcome phase, the feedback-related negativ-
ity is of relevance (FRN, in the literature also referred
to as mediofrontal negativity [MFN]) (Mu¨ller, Mo¨ller,
Rodriguez-Fornells, & Mu¨nte, 2005; Gehring & Willoughby,
2002; Miltner, Brun, & Coles, 1997). The FRN is an
enhanced negative def lection following negative in
comparison to positive feedback, with an onset at about
250 msec and a fronto-central maximum. Converging
evidence suggests that the FRN reflects the evaluation
of the valence of an outcome and is thus sensitive to the
motivational significance of ongoing events. Source mod-
eling studies have suggested the rostral anterior cingu-
late, the posterior cingulate gyrus (Mu¨ller et al., 2005;
Nieuwenhuis, Slagter, von Geusau, Heslenfeld, & Holroyd,
2005), and right superior frontal gyrus (Nieuwenhuis
et coll., 2005) as generators of the FRN. The FRN can pro-
vide insight into the participants’ motivational and eval-
uative processes in reaction to the provocation und
punishment.
As suggested by the GAM, both personality (as traits)
and situative factors (as provocation) influence the
cognitive and motivational processes associated with
aggressive acts. Personality traits that have been shown
to affect someone’s proneness to aggression include her
impulsivity, ou
or his trait aggressiveness, narcissism,
emotional susceptibility (Bettencourt, Talley, Benjamin,
& Valentine, 2006; Anderson & Bushman, 2002). Trait
aggressiveness has been shown to affect aggressive
behavior both under neutral and provoking conditions
(Bettencourt et al., 2006). It is thus together with
irritability the personality variable with the most reliable
influence on laboratory-induced aggressive behavior.
For the present study, we thus selected student partic-
ipants on the basis of their score in an aggression
questionnaire (Hampel & Selg, 1975), yielding two
groups of high (HT) and low trait (LT) aggressive
participants. This design affords us with the possibility
to delineate the impact of both personality (trait aggres-
sion) and situative factors (provocation) on cognitive
and motivational processes during an aggressive inter-
action. Besides the impact of traits, the search for sex
differences in aggression has generated a considerable
Kra¨mer et al.
1465
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
0
je
8
e
–
1
p
4
d
6
F
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
.
.
.
/
F
t
o
n
1
8
M.
un
oui
2
0
2
1
amount of research literature, with controversial results,
cependant (Bettencourt & Miller, 1996; Eagly & Steffen,
1986; Frodi, Macaulay, & Thome, 1977). Although re-
searchers largely agree on a preponderance of men in
physical aggression (Anderson & Bushman, 2002; Eagly
& Steffen, 1986), they acknowledge a dramatic reduction
of sex differences under conditions of provocation
(Bettencourt & Miller, 1996; Frodi et al., 1977; Taylor
& Epstein, 1967). In the present study, we therefore
included both women and men to be able to control for
putative sex differences.
MÉTHODES
Participants
Data of the trait aggressiveness questionnaire were
obtained from 231 economics students (129 femmes;
âge moyen = 22.6 années, SD = 1.9 années). As psychology
students are familiar with this kind of experiment and
can be expected to get suspicious, we decided to se-
lect subjects from a sample of economics students, OMS
were unfamiliar with psychological experiments. Based
on their general aggression score (see below for fur-
ther explanation), we selected participants with extreme
valeurs (low/high) for the brain potential experiment.
Forty-nine young, healthy students (24 femmes; mean
age = 22.9 années, SD = 1.9 années) participated in the
electroencephalogram (EEG) study after giving informed
consent. All had normal or corrected-to-normal vision
and were free of neuropsychiatric disorders. Nine par-
ticipants were excluded due to extensive eye movement
or muscle artifacts. Ainsi, 40 participants (21 femmes)
were included in the analyses. The study was performed
in agreement with the Declaration of Helsinki and
approved by the ethics committee of the University of
Magdeburg. Participants received money or course cred-
it for taking part.
Questionnaire
Participants for the ERP study were selected based on
their trait aggressiveness, assessed with a German inven-
tory for the assessment of factors of aggression (FAF,
Fragebogen zur Erfassung von Aggressivita¨tsfaktoren)
(Hampel & Selg, 1975). With this questionnaire, five
subscales (spontaneous aggression, reactive aggression,
impulsiveness, autoaggression, aggression inhibition)
and a control scale (openness) can be obtained. Spon-
taneous aggression (19 items) refers to unrestrained
verbal or physical aggression, a typical item is, ‘‘I some-
times like to taunt others.’’ Items of the reactive ag-
gression scale (13 items) ask for aggressive reactions
to some kind of provocation or unfairness, tel que. ‘‘If
someone provokes me, I want to punish him badly.’’
Items of the impulsivity scale (13 items) deal with the
affective component of aggression, as ‘‘I flare up quickly,
but get over it quickly.’’ The sum of the scales ‘‘sponta-
neous aggression,’’ ‘‘reactive aggression,’’ and ‘‘impul-
siveness’’ gives a reliable measure for outwardly directed
aggression (internal consistency Cronbach’s alpha =
.85) and was thus used for selection of HT and LT
aggressive participants. The sum score has been proven
to be significantly different between both adolescent and
adult violent criminals on the one hand and nonviolent
controls on the other hand (Hampel & Selg, 1975),
providing evidence for its external validity.
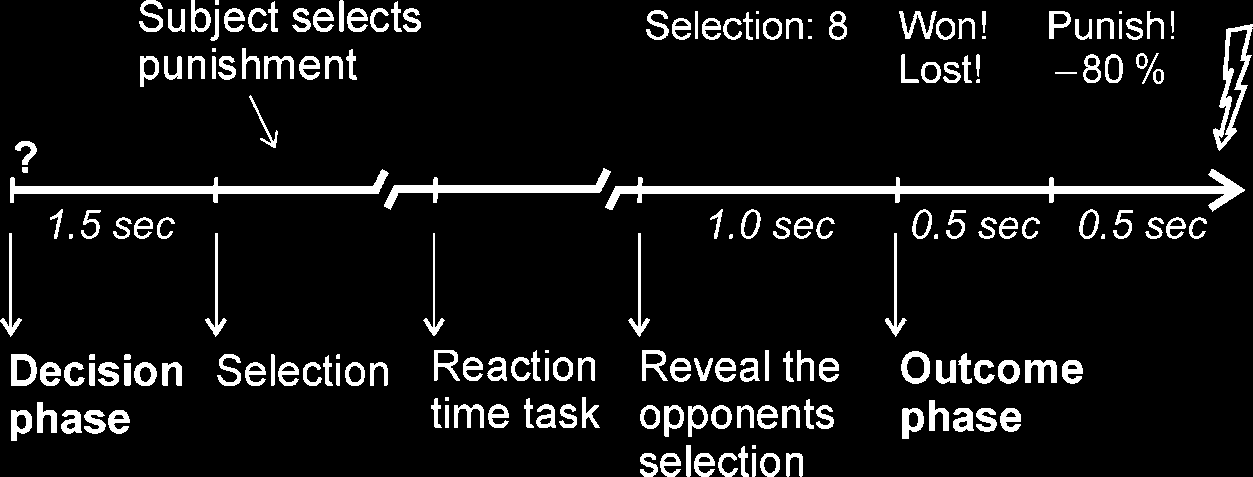
Task and Procedure
Aggression was elicited and assessed using a modified
version of the TAP (Taylor, 1967). Participants were
instructed that they were playing successive competitive
reaction time trials against one of two opponents in
alternating blocks. The opponents (one man, one wom-
un), confederates of the experimenters, met the partic-
ipant prior to the experiment to jointly listen to the
instructions: They were told that whoever lost would
be punished by the opponent with a highly aversive
bruit. The severity of the punishment, that is the vol-
ume of the noise, had to be selected for each trial on
a range from 1 à 8. En plus, participants would
lose a corresponding amount of money (lowest noise—
10 cents, highest noise—40 cents) to assure the partic-
ipants’ motivation to avoid the punishment. En fait,
selections of the putative opponents and outcome of
the trials (50% winning and losing trials in each block)
were under the control of the experimenter. The exper-
iment comprised eight blocks of 40 trials each, yielding a
total of 320 trials. Participants were told that the oppo-
nents would play in alternating blocks and rest during
the others. At the end of the experiment, participants
were completely debriefed about the deception and the
experiment’s motivation. We always introduced one
man and one woman as opponents, as the sex of the
target is known to affect the level of aggressiveness
(Taylor & Epstein, 1967). As the respective target’s sex
was not revealed during the experiment, this effect
could not influence the behavior, cependant.
At the beginning of each trial, participants were asked
to consider the punishment for the opponent in the
upcoming trial (in the following referred to as decision
phase; duration of 1.5 sec). A prompt to press the
respective keyboard button followed. After the reaction
time task proper (a visual cue, which prompted the
players to press a button as fast as possible), the se-
lection of the opponent was presented: In half of the
blocks, the opponent mainly selected a low punishment
(level 1–4 in 80% of the trials; blocks of low provoca-
tion); in the remaining blocks, the opponent selected
mainly a high punishment (level 5–8 in 80% of the trials;
blocks of high provocation). Ainsi, the participant got
the impression of playing blockwise against one fair and
one rather unfair opponent. Enfin, feedback was given
1466
Journal des neurosciences cognitives
Volume 20, Nombre 8
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
0
je
8
e
–
1
p
4
d
6
F
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
.
F
/
.
t
.
o
n
1
8
M.
un
oui
2
0
2
1
whether the participant had won or lost (in the follow-
ing referred to as outcome phase, duration of 500 msec).
On win trials, they had to elicit the noise punishment for
the opponent by button press; on loss trials, they were
exposed to the aversive noise via headphones. Dans 10% de
the reaction time task, ‘‘no-go’’ trials were introduced to
ensure the participants’ attention. These trials were
followed by feedback on the successful or failed inhibi-
tion. False alarms were punished by the experimenter
with monetary subtraction, whereas successful
inhibi-
tions did not yield any consequences. The feedback
phase of these trials was excluded from any further
analyses, as it was unrelated to any punishment or re-
taliation. Prior to the experiment, the volume of the
loudest noise was adapted to the participant’s individual
threshold for perceiving noises as aversive. Intertrial
interval was 3 sec. After each of the eight blocks, ils
were allowed to rest. Stimulus presentation and behav-
ioral data acquisition were controlled with Presentation
software (www.neurobehavioralsystems.com). The ex-
periment, including preparation and debriefing, had a
duration of 90 min (for the timing of each trial, see also
Chiffre 1).
EEG Recordings
The EEG was recorded from 27 tin electrodes mounted
in an elastic cap (Easycap; positions: Fp1/2, F3/4, C3/4,
P3/4, O1/2, F7/8, T7/8, CP1/2, P7/8, FC1/2, FC5/6, CP5/6,
Fz, Cz, Pz, with reference electrodes placed on the right
and left mastoids. During recording, all scalp electrodes
were referenced against the right mastoid and off-line re-
referenced against the algebraic mean of the activity at
the two mastoid processes. Electrode impedances were
kept below 5 k(cid:1). To monitor horizontal eye move-
ments, electrodes were placed on the outer canthus of
the right and left eyes. Vertical eye movements and
blinks were monitored by an electrode placed below
the right eye referenced to the right mastoid. EEG and
electrooculogram were recorded continuously with a
band pass of 0.095 à 70 Hz and digitized with a sam-
pling rate of 250 Hz.
Off-line eye and muscle artifacts were rejected auto-
matically. For nine participants with extensive blinks, eye
movements were corrected with the technique of Sec-
ond Order Blind Identification ( Joyce, Gorodnitsky, &
Kutas, 2004). Stimulus-locked ERPs were obtained sep-
arately for the different phases of the trial (décision
phase: time-locked to the prompt to consider punish-
ment for the opponent in the upcoming trial; outcome
phase: time-locked to the feedback). The epochs were
1024 msec long, including a 100-msec baseline.
Data Analysis
Unless otherwise specified, only those 80% of trials
were analyzed that followed a block-congruent selec-
tion of the opponent, c'est, a high selection in a block
avec 80% highly provoking trials and vice versa. Selec-
tions of the participants under high and low provoca-
tion and reaction times were scored and compared
with repeated measures analysis of variance (ANOVA;
within-subject factor provocation: high vs. faible; entre-
subject factor FAF: high vs. low trait aggressive, c'est à dire., HT
vs. LT).
As we observed that some participants’ behavior was
contrary to their questionnaire score (c'est à dire., LT partici-
pants showed highly aggressive behavior and vice versa),
we tested whether the neural responses depended rath-
er on the experimentally induced aggression. To this
end, we performed a median-split in each of the two
groupes (LT and HT) based on participants’ overall ag-
gressive behavior in the paradigm (mean selection in all
trials). We thus yielded groups with low and high
experimentally induced aggression, referred to as LE
and HE, respectivement (see Figure 2C). Note that as the
median of selections in the two groups were different
(LT = 4.23 and HT = 5.20), the groups LT–HE and HT–
LE are in fact overlapping with respect to their behavior.
We will thus consider the between-subject factor behav-
ior (LE vs. HE) only in interaction with the between-
subject factor FAF (LT vs. HT).
Event-related potentials were analyzed separately for
the decision and outcome phase. For the decision phase,
ERPs were quantified by a mean amplitude measure at
different electrode positions. Based on the results of
our previous pilot study, we used 12 électrodes (F3/4,
C3/4, P3/4, F7/8, T7/8, P7/8) to yield three factors:
Chiffre 1. Time line for a
single trial, a trial of high
provocation (opponent selects
8) is depicted an example.
The duration of the selection
and reaction time task phases
depended on the participants’
reaction time. The intertrial
interval had a duration of 3 sec.
Kra¨mer et al.
1467
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
0
je
8
e
–
1
p
4
d
6
F
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
/
F
.
.
.
t
o
n
1
8
M.
un
oui
2
0
2
1
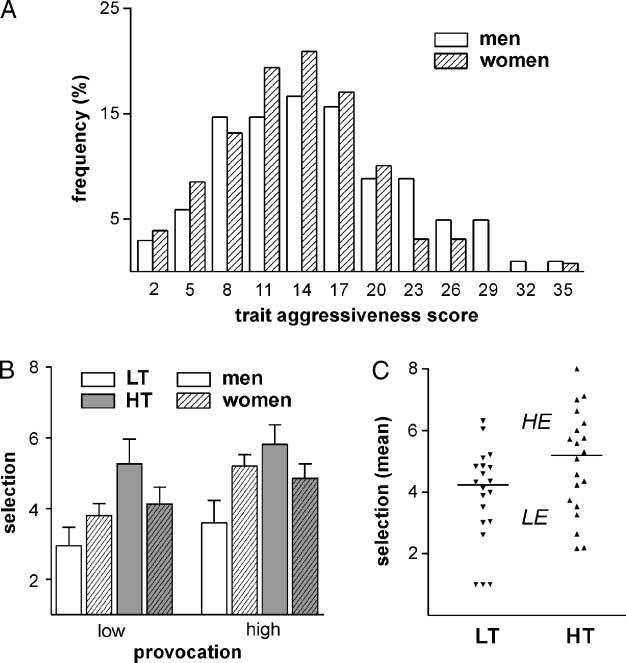
Chiffre 2. Behavioral results.
Depicted first is the
distribution of the
aggressiveness scores of
men and women in the
questionnaire sample
(n = 231). Each bar represents
the percentage for a score
range of three points, le
numbers refer to the mean of
the respective range (c'est à dire., ‘‘2’’
moyens: aggression score of 1
à 3; UN). Shown are the mean
selections under low (gauche) et
high provocation (droite) dans le
different groups (B). Blanc
boxes show low trait aggressive
(LT), gray boxes high trait
aggressive (HT) participants.
Blank boxes show men’s data,
whereas lined boxes show
women’s data. C shows the
distribution of the mean
punishment selection in both
groupes (HT and LT). The small
lines mark the groups’ median,
Et ainsi, the cut value to
yield groups of low (LE) et
haut (HE) experimentally
induced aggression.
anteriority (frontal, central, posterior; in the following
referred to as ANT), hemisphere (droite, gauche; referred to
as HEM), and laterality (parasagittal and temporal; concernant-
ferred to as LAT). Two components were analyzed: un
negative deflection around 300 msec (‘‘N2,’’ time win-
dow 270–300 msec) and a late frontal negativity quan-
tified by a mean amplitude measure in the time window
350–600 msec. The ANOVA thus comprised the between-
subject factors FAF (HT vs. LT) and behavior (HE vs.
LE) and the within-subject factors provocation (high vs.
low provocation trial), ANT, HEM, and LAT. For all
statistical effects involving more than one degree of free-
dom in the numerator, the Huynh and Feldt (1976)
correction was applied to correct for possible viola-
tions of the sphericity assumption. The corrected prob-
abilities, together with the corresponding >-values, sont
reported.
For the outcome phase, we examined the FRN. As the
FRN is superimposed on the large positive deflection,
we filtered the data with a band-pass filter of 4–10 Hz to
remove the slow waves (see Luu & Tucker, 2001 for a
similar procedure). As the FRN is known to be maximal
at fronto-central sites, we subjected the participants’
mean amplitude between 270 et 300 msec at Fz to a
repeated measures ANOVA with the between-subject
factors FAF and behavior and the within-subject factors
feedback (won vs. lost) and provocation (high vs. faible
provocation trial). In this analysis, we included all trials
and refer with high and low provocation to the actual
selection of the opponent, presented directly before the
feedback. Losing under high provocation thus means
that the participant had to expect a high punishment
and vice versa.
RÉSULTATS
Questionnaire Data
The FAF mean score (see above for selected factors) était
13.19 (SD = 6.57), and higher for men (mean = 14.3,
SD = 7.23) than for women [mean = 12.3, SD = 5.88;
t(229) = 2.32, p = .021; see Figure 2A for the distribu-
tion]. From this sample, 24 HT and 25 LT aggressive
participants were selected for the EEG study, of whom
nine had to be discarded due to extensive artifacts. Le
mean score for the remaining 20 HT aggressive partic-
ipants (11 femmes) était 23.8 (range 19 à 34, corre-
sponding to a percentile > 82.0, with respect to the
complete sample), and for the 20 LT aggressive partic-
ipants (10 femmes) était 3.9 (range 0 à 7, percentile <
18.2). In this sample, only a small sex effect could be ob-
served [main effect of sex: F(1, 36) = 4.052, p = .052],
with no differences between high and low aggressive
participants (interaction FAF (cid:1) Sex: F < 1).
1468
Journal of Cognitive Neuroscience
Volume 20, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
8
e
-
1
p
4
d
6
f
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
f
.
.
.
/
t
o
n
1
8
M
a
y
2
0
2
1
Behavioral Data
Participants selected higher punishments in blocks of
high provocation compared to blocks of low provoca-
tion [main effect provocation: F(1, 38) = 42.91, p < .001].
HT aggressive participants showed more aggressive
behavior [main effect FAF: F(1, 38) = 4.42, p = .042].
However, a weak-to-middle correlation of FAF score and
average punishment selection confirmed the aforemen-
tioned observation of trait-incongruent behavior in
some participants (r = .317, p = .046). The level of
selected punishments increased over time, when com-
paring the four blocks in each condition, resulting in a
significant main effect of block [repeated measures
ANOVA with FAF, provocation, and block: main effect
block, F(3, 114) = 3.52, p = .023]. No interaction with
the level of provocation or the FAF was detected (both
p > .2). The average level of selections under high
provocation was in the first block 4.62 (61.74) et en
the last block 4.95 (62.0), whereas under low provoca-
tion it increased from 3.76 (61.65) à 4.19 (62.09).
Fait intéressant, the effect of trait aggressiveness on the
aggressive behavior was observed in the male partic-
ipants only, whereas HT and LT aggressive women
showed similar behavior. This was confirmed with a
repeated measures ANOVA with the between-subject
factors FAF and sex and the within-subject factor prov-
ocation [interaction FAF (cid:1) Sex: F(1, 36) = 5.37, p = .026;
main effect FAF in men: p < .05, in women: p > .10].
Women tended to react stronger on the provocation as
indicated by a marginally significant interaction of Sex (cid:1)
Provocation [F(1, 36) = 3.44, p = .072; for the behav-
ioral results, see also Figure 2B]. Participants’ mean
reaction time to select the noise level was 770.4 msec
(SD = 361.7), with no differences between conditions
and the groups (all main effects and interactions:
p > .05). Participants’ mean reaction time on the target
était 257.3 msec (SD = 82.8), which again did not differ
between the provocation level and the groups (all main
effects and interactions, p > .1). During debriefing,
participants reported to have been more aggressive
under high than under low provocation [F(1, 35) =
41.43, p < .01]. Again, an interaction of FAF and sex
indicated that trait aggressiveness was a significant pre-
dictor of reported aggression in men, but not so in
women [F(1, 35) = 4.79, p = .035; main effect FAF in
men: p < .05, in women: p > .5].
ERPs in the Decision Phase
ERPs in the decision phase were characterized by the
typical N1 and P2 components, followed by an extended
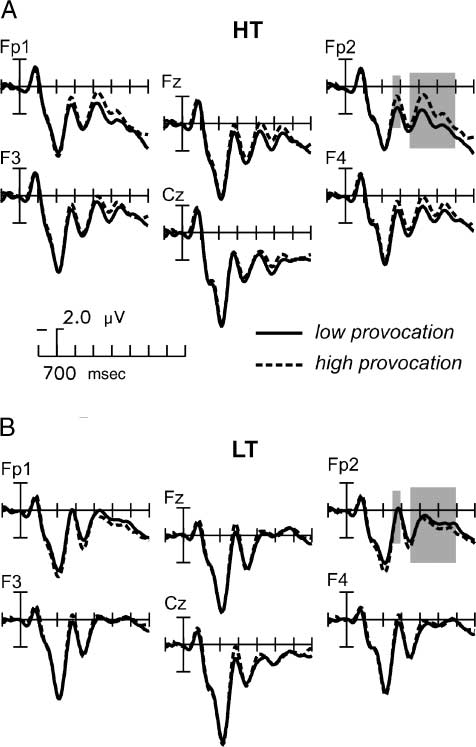
negativity. In high provocation trials, a right-lateralized,
frontal negative deflection of the ERP was observable
first in the time range of the N2 (250–300 msec) et
then from 350 msec on. Visual inspection suggested that
this held true for the high trait aggressive (HT) partic-
ipants only (Figure 3A and B; voir la figure 6 for the
topography of these effects). En effet, in the HT group,
high provocation elicited an enlarged negativity com-
pared to low provocation, indicated by the interaction
Provocation (cid:1) ANT (cid:1) FAF [F(2, 72) = 3.86, p = .045, > =
0.6572; see Figure 3A]. This effect was maximal at frontal
sites and peaked around 270–300 msec. No such effect
was detected in the LT group (HT: Provocation (cid:1) ANT,
p < .01; LT: Provocation (cid:1) ANT, p > .10; see Figure 3B).
Fait intéressant, this effect was seen only for those HT ag-
gressive participants, OMS, nevertheless, did not behave
aggressively in the experiment (c'est à dire., the HT–LE group),
as indicated by a (albeit marginal) significant interaction
FAF (cid:1) Behavior (cid:1) Provocation (cid:1) ANT [F(2, 72) = 3.42,
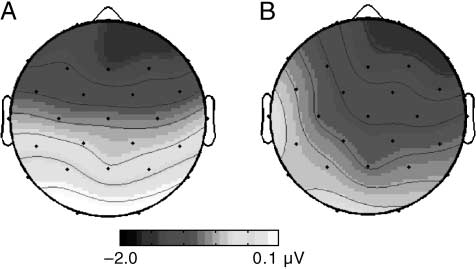
p = .0549, > = 0.6572; see Figures 4 et 6].
As mentioned previously, ERP effects resembling the
N2 in the current experiment have repeatedly been linked
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
0
je
8
e
–
1
p
4
d
6
F
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
t
/
.
.
F
.
o
n
1
8
M.
un
oui
2
0
2
1
Chiffre 3. Grand-average ERPs in the decision phase. For illustration,
the ERPs were low-pass filtered (high cutoff 12 Hz). Depicted are
the ERPs in (UN) the high trait aggressive group (HT) et (B) le
low trait aggressive group (LT). Low provocation trials are shown
with solid lines, high provocation trials are shown with dashed lines.
Marked are the analyzed time windows for the two effects in the
decision phase (early and late frontal negativity).
Kra¨mer et al.
1469
ginally significant ( p = .060). As the FAF questionnaire
contains an inhibition subscale, we checked whether the
participants’ score on this scale did predict their behavior
and/or the fronto-lateral negativity. Both the correlation
with the aggressive behavior and with the magnitude of
the neurophysiological effect were not significant, comment-
jamais (both r < .1, p > .2).
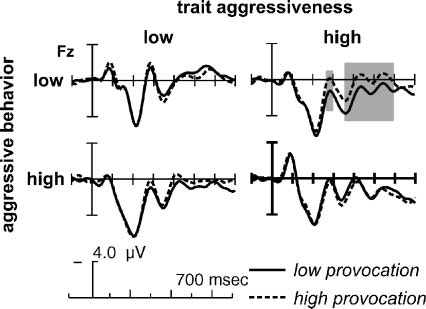
Following the early frontal negativity, we observed an
enhanced late frontal negativity for high compared to
low provocation trials. This effect was lateralized with
a maximum over right frontal electrodes. Encore, ce
effect was differentially affected by the participant’s trait
aggressiveness and experimentally induced aggressive
behavior. We found in the HT aggressive group an en-
hanced frontal, right-lateralized negativity only for the
HT–LE participants [high FAF: Behavior (cid:1) Provocation (cid:1)
ANT (cid:1) HEM: F(2, 36) = 3.60, p = .038, > = 0.9818; voir
also Figure 4 and Figure 6]. The significant correlation of
the participants’ average aggressive behavior and the
amplitude difference at Fz further supported this finding
(r = .48, p = .032). No such relation was found for the
LT aggressive participants (all interactions of involving
Provocation (cid:1) Behavior: p > .1; correlation r = (cid:2).172,
p > .1), yielding a significant interaction effect in the
moderator analyses ( p = .042). Néanmoins, the inter-
action of the late frontal negativity effect with the
between-subject factor FAF in the overall ANOVA was
only marginally significant [Provocation (cid:1) ANT (cid:1) HEM (cid:1)
FAF (cid:1) Behavior: F(2, 72) = 2.50, p = .0992].
The fact that the early and the later negativity during
the decision phase had a similar fronto-lateral distribu-
tion and were similarly related to the participants’
behavior suggests that it might be rather one underlying
extended frontal negativity than two separate effects. Si
this holds true, no topographic differences should be
detectable comparing the early and later effect. Nous
therefore subjected the vector-normalized data to a
repeated measures ANOVA to probe for interactions of
temps (270–300 msec vs. 350–600 msec) with provocation
and electrode position (factors ANT, HEM, and LAT)
(McCarthy & Wood, 1985). As both observed effects were
strongest for the HT–LE participants, we performed those
analyses with this group only. No significant interactions
Chiffre 4. Grand-average ERPs at Fz in the decision phase. Pour
illustration, the ERPs were low-pass filtered (high cutoff 12 Hz).
Depicted are the ERPs in the four different groups: faible (LE, upper
part) and high (HE, lower part) aggressively behaving participants
in the low trait aggressive group (LT, gauche) and the high trait
aggressive group (HT, droite). Low provocation trials are shown
with solid lines, high provocation trials with dashed lines. Marked
are the analyzed time windows for the two effects in the decision
phase (early and late frontal negativity).
to inhibitory processes (Pliszka et al., 2000; Pfefferbaum
et coll., 1985). Assuming that the higher negativity reflects
inhibitory processes, it should be largest for those HT
aggressive participants who inhibit their predominant
reaction to get back to the opponent, namely, ceux
who show low aggressive behavior during the actual
experiment. To further probe this hypothesis, we com-
puted the mean amplitude of the difference waves (haut
minus low provocation) entre 270 et 300 msec at Fz,
where the maximum of the negativity was detected, et
tested its relationship with the participants’ mean selec-
tion, reflecting their average level of aggressive behavior
(irrespective of the level of provocation). Although a clear
correlation between aggressive behavior in the experi-
ment and the enhanced negativity emerged for the HT
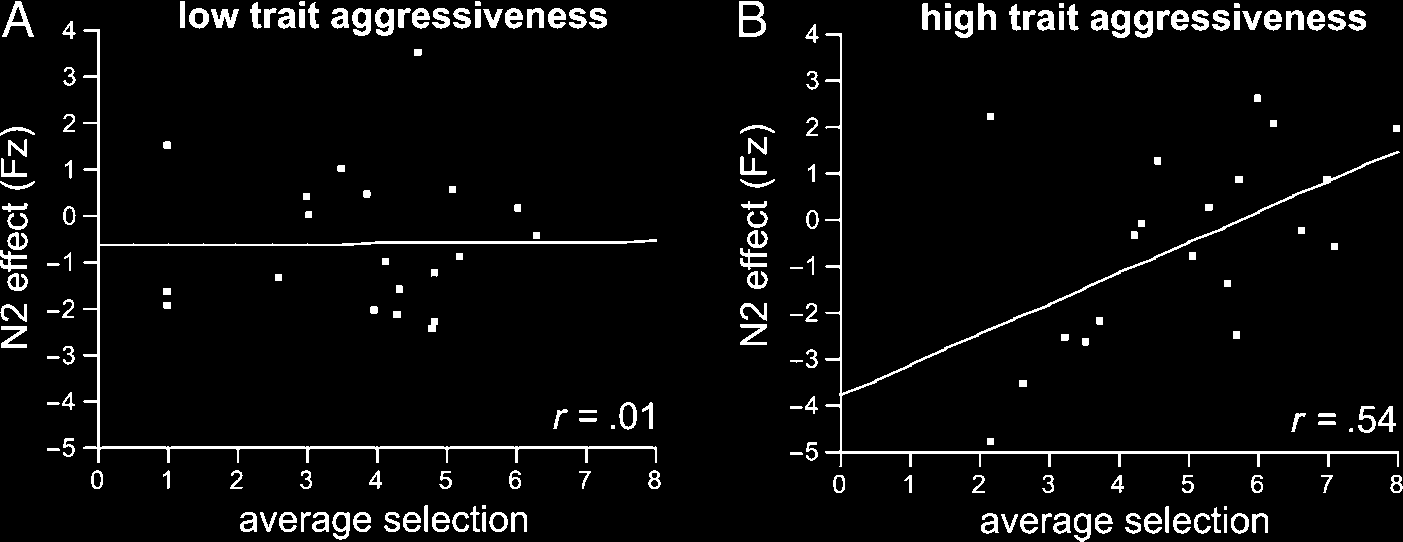
aggressive participants (r = .54, p = .014), a similar
relation was absent in the LT aggressive group (r = .01;
see also Figure 5). The interaction effect in the moderator
analyse (Cohen, Cohen, West, & Aiken, 2003) was mar-
Chiffre 5. Correlation of
aggressive behavior (mean
selection over all blocks)
and early negativity effect
(amplitude of difference high
minus low provocation trials).
The left side shows the
(nonsignificant) correlation
in the LT aggressive group,
the right side shows the
correlation in the HT
aggressive group.
1470
Journal des neurosciences cognitives
Volume 20, Nombre 8
D
o
w
n
je
o
un
d
e
d
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
F
r
o
m
D
h
o
t
w
t
n
p
o
:
un
/
d
/
e
m
d
je
F
t
r
o
p
m
r
c
h
.
s
p
je
je
d
v
je
e
r
e
r
c
c
t
.
h
m
un
je
r
e
.
d
toi
c
o
o
m
c
/
n
j
un
o
r
c
t
je
n
c
/
e
un
–
p
r
d
t
je
2
c
0
je
8
e
–
1
p
4
d
6
F
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
oui
2
0
g
0
toi
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
F
e
m
b
oui
b
e
g
r
toi
2
0
e
2
s
3
t
/
j
.
t
F
.
/
.
o
n
1
8
M.
un
oui
2
0
2
1
albeit still significant, difference for losing and winning
trials [Feedback: F(1, 18) = 10.09, p = .005]. This obser-
vation suggests that punishing the opponent had a sim-
ilar negative valence for nonaggressive participants as
being punished.
Cependant, as argued previously, a group analysis based
on a median-split is problematic, as it assigns partici-
pants to different groups who show, in fact, quite similar
behavior, Et ainsi, may yield misleading results. This is
also true for the present set of data as the median of
punishment selection was different in the HT and LT
aggression groups. Ainsi, when comparing the HE and
LE participants, the groups are, in fact, overlapping with
respect to their behavior.
To therefore further probe the relation between the
participants’ aggressive behavior and their feedback
evaluation, we compared the upper and lower third
regarding the aggressive behavior (each group with
n = 13, average selection >5.2 and <3.8, respectively).
This was done irrespective of the participants’ trait ag-
gressiveness. This analysis confirmed our finding of a
modulated FRN [Feedback (cid:1) Third: F(1, 24) = 7.53, p =
.011]. Although the negativity had a similar amplitude
for win and loss trials in the lowest third (amplitude at
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
8
e
-
1
p
4
d
6
f
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
.
/
t
f
o
n
1
8
M
a
y
2
0
2
1
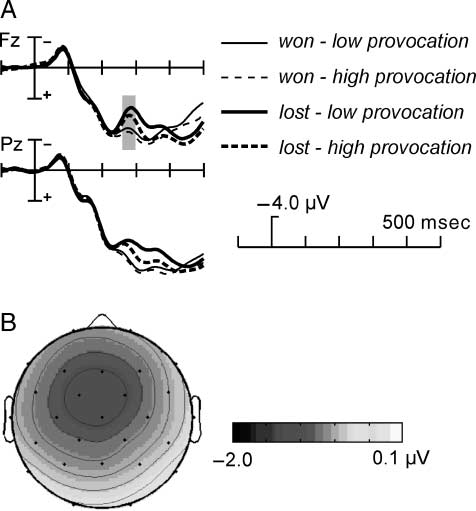
Figure 7. (A) Grand-average ERPs at Fz and Pz in the outcome phase.
For illustration, the ERPs were low-pass filtered (high cutoff 12 Hz).
Depicted are the ERPs in the whole sample (n = 40). At Fz, the
time window of the maximum of the FRN is marked in gray. Depicted
are the ERPs for the four different conditions: win (thin lines) and
lost trials (thick lines), high provocation (dashed lines) and low
provocation trials (solid lines). (B) Map showing the FRN effect in
the complete sample (band-pass filtered 4–10 Hz). The map depicts
the mean amplitude of the difference between negative and positive
feedback in the time window 270–300 msec.
Kra¨mer et al.
1471
Figure 6. Maps showing the early (A) and the late frontal negativity
(B) in the HT–LE group. The maps depict the mean amplitude of
the difference between high and low provocation in the time window
270–300 msec and 350–600 msec.
were detected (all p > .05), further supporting the
interpretation as one prolonged negativity.
As we observed significant sex differences in the
behavioral data, we conducted post hoc analyses to test
whether these behavioral effects were reflected in the
neurophysiological data. To this end, we subjected the
mean amplitude for both the early (270–300 msec) et
late time window (350–600 msec) to a repeated mea-
sures ANOVA with the factors sex, FAF, provocation,
ANT, HEM, and LAT. In contrast to the behavioral data,
we did not observe any sex differences in the neuro-
physiological data, neither regarding the early nor the
late frontal negativity (main effects of factor sex and all
interactions, p > .05).
ERPs in the Outcome Phase
ERPs following negative feedback in both high and low
provocation trials showed a clear MFN peaking around
300 msec in comparison to positive feedback [main
effect of feedback at Fz: F(1, 36) = 39.45, p < .001;
see Figure 7]. The FRN was independent of the level of
provocation, that is, the magnitude of punishment to be
expected (interaction Feedback (cid:1) Provocation, p > .1).
The difference between positive and negative feed-
back was differentially pronounced in the different
groupes, reflected by the interaction of Feedback (cid:1)
FAF (cid:1) Behavior [F(1, 36) = 4.40, p = .043; voir
Chiffre 8]. En fait, for the participants in the HT–LE
group, the difference between negative and positive
feedback was even absent (main effect of feedback,
p > .1). Fait intéressant, it seemed that for both the HT–
LE and LT–LE participants, positive feedback also elicited
an MFN that was only slightly smaller than after negative
feedback. This was confirmed by the interaction of
Feedback (cid:1) Behavior [F(1, 36) = 7.15, p = .01]. Al-
though HE participants showed a clear difference be-
tween negative and positive feedback [Feedback: F(1,
18) = 29.58, p < .001], LE participants showed a smaller,
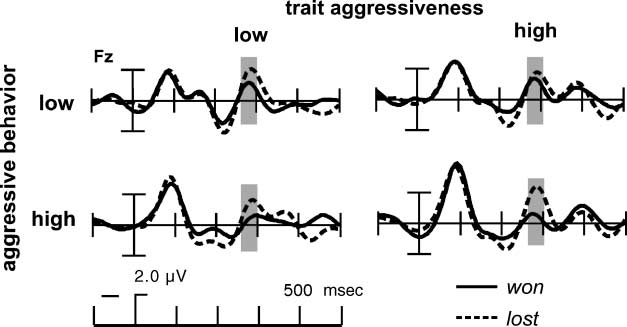
Figure 8. Grand-average ERPs
at Fz in the outcome phase,
separately for the four groups.
The ERPs were band-pass
filtered (4–10 Hz) to remove
the underlying slow positive
waves. Depicted are the ERPs
in the different groups: low
(LT, left) and high (HT, right)
trait aggressive, separately for
low (LE, upper part) and high
(HE, lower part) aggressive
behavior. ERPs following
positive feedback are shown
with solid lines, following
negative feedback with
dashed lines. The time
window used for the FRN
analyses is marked in gray.
Fz, wins: (cid:2)0.95 AV; losses (cid:2)1.36 AV; p > .1), it showed a
significant modulation by the feedback in the most
aggressive group (highest third; wins (cid:2)0.25 AV; pertes
(cid:2)1.83 AV; p < .005).
As the FRN has been related to the perceived valence
of a stimulus (Yeung, Holroyd, & Cohen, 2005), our
findings suggest that winning (and thus punishing the
opponent) was associated with negative feelings for the
less aggressive participants. This could reflect their
empathic feelings for the punished opponents (Lotze
et al., 2007). One might thus speculate that this emo-
tional modulation of the punishment’s evaluation is
driving the regulation of aggressive behavior in less
aggressive participants. This should cause lower punish-
ment selections especially after winning trials in these
participants. Indeed, nonaggressive participants selected
lower punishments after win than after loss trials, in high
provocation blocks at least (mean selection after win-
ning: 2.74, after losing: 3.11; p < .05). This difference
was not observable in low provocation blocks, most
probably due to a floor effect (main selection after win-
ning: 2.11, after losing: 2.19). Highly aggressive partici-
pants did not exhibit this modulation of aggressiveness
after winning, resulting in the threefold interaction of
Feedback (cid:1) Provocation (cid:1) Third on the selection in
the consecutive trial [F(1, 24) = 4.14, p = .046].
Similarly to the decision phase, we performed post
hoc analyses to probe for sex differences in the neuro-
physiological data. To this end, we subjected the mean
amplitude at Fz for the FRN to a repeated measures
ANOVA with the factors FAF, sex, feedback, and provo-
cation. Again, no differential neurophysiological re-
sponses comparing men and women were detected.
DISCUSSION
The present study is the first to record neurophysiolog-
ical responses during an aggressive interaction with the
aim to tap into the decision processes and evaluation
processes involved and to demonstrate their modulation
by trait aggressiveness. Provocation-related modulations
of specific ERP components that were differentially
pronounced for participants showing HT and LT ag-
gressiveness may provide insight into the mechanisms
subserving aggression and expand findings from mere
behavioral studies.
Behavioral Data
Both behavioral effects and results of the postexperi-
mental questionnaire showed a clear provocation effect
in the study, despite the fact that a modified version of
the TAP with an increased number of trials was em-
ployed. We also made behavioral observations that are
not directly reflected by neurophysiological data. In con-
trast to the male participants, the trait aggressiveness
was not predictive of the actual aggressive behavior in
women. In fact, the women’s behavior appeared more
driven by the provocation level in the experiment rather
than by their trait aggressiveness. This is in line with
previous studies which, in general, have found a weaker
relation between trait aggressiveness and actual behavior
for women (Giancola, 2003). The tendency of a stronger
provocation effect in women is corroborated by earlier
studies on laboratory-induced aggression (Bettencourt &
Miller, 1996; Taylor & Epstein, 1967). These behavioral
differences were not directly reflected by neurophysio-
logical differences. Thus, further studies are needed to
specify the neural correlates of sex differences in aggres-
sive behavior.
Decision Phase
During the decision phase, we observed an enhanced
frontal negativity (‘‘N2’’) in high compared to low
provocation trials. This effect was detectable only in
HT aggressive participants and was correlated with the
1472
Journal of Cognitive Neuroscience
Volume 20, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
8
e
-
1
p
4
d
6
f
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
t
f
.
.
/
.
o
n
1
8
M
a
y
2
0
2
1
actual aggressive behavior, such that HT aggressive
participants who, nevertheless, behaved nonaggressively
showed the highest effect. No such effect was seen in
the LT aggressive participants. Negativities of a similar
temporal onset and distribution have been observed in a
number of situations and have been interpreted as re-
flecting response conflict (van Veen & Carter, 2002) or
inhibitory processes (Falkenstein et al., 1999; Pfefferbaum
et al., 1985). As participants did not have to respond
during the decision phase, the observed negativity can
neither be due to the detection of a conflict between
two motor responses nor be related to the inhibition
of a predominant motor response. Rather, we propose
that this effect is reflecting general monitoring or in-
hibition processes unrelated to the motor output itself
(Bruin & Wijers, 2002). The lack of significant correla-
tions between the inhibition subscale of the FAF and
the participants’ behavior or neurophysiological re-
sponse might argue against this association with inhib-
itory processes. However, the FAF inhibition score has
failed to differentiate between violent criminals and
controls, and rather assesses the knowledge of moral
rules and degree of conscientiousness (Hampel & Selg,
1975). It might thus be unrelated to the inhibitory
mechanisms necessitated in a social provocation.
Following this early provocation-related effect, we
observed an enhanced late fronto-lateral negativity un-
der high compared to low provocation. This effect
showed a similar distribution and relation to the partic-
ipants’ aggressiveness as the earlier effect. It was also
observable for HT aggressive participants only and cor-
related with their level of aggressive behavior. Further
analyses of the topography confirmed the similarity of
both effects. This indicates that both effects are probably
reflecting an extended right frontal negativity, although
further studies will be needed to probe possible func-
tional and anatomical differences. It might well be that
this extended negativity reflects the prolonged activity
of those same neural generators that are involved in
response competition and inhibitory processes. How-
ever, for the present study, we will, for descriptive pur-
poses only, refer to this extended frontal negativity as
decision-related negativity (DRN), in parallel to the later
discussed FRN.
The fact, that the DRN was largest in HT aggressive
participants, who behaved nonaggressively in the actual
experiment (see Figures 4 and 5), suggests that a
conflict between different motivations might drive this
effect: having the predisposition to get back to a pro-
voking opponent, but willing to prevent escalation at the
same time. As no self-report data were assessed that
could confirm this association with a conflict, future
studies will be needed to further substantiate this argu-
ment. However, given the similarity of the neurophysi-
ological response with data from ‘‘pure’’ cognitive tasks,
this goes in line with recent observations that similar
(although not necessarily the same) mechanisms are
engaged by cognition–emotion competition as by con-
flicts within the cognitive domain (Etkin, Egner, Peraza,
Kandel, & Hirsch, 2006; Sanfey, Rilling, Aronson, Nystrom,
& Cohen, 2003). In an fMRI study on the neural basis of
decision making in the ultimatum game, for instance,
unfair offers yielded a higher blood oxygenation level-
dependent response in ACC (Sanfey et al., 2003). The
authors explain this with a detection of a conflict be-
tween the emotional response to unfairness and the
cognitive motivation to maximize money. Results in an
‘‘emotional’’ Stroop task also point to a similar role of
ACC in emotional conflict resolution (Etkin et al., 2006),
although the activation was in more rostral parts of
ACC, compared to the dorsal activations found typically
in situations entailing stimulus–response incompatibili-
ties (Botvinick, Nystrom, Fissell, Carter, & Cohen, 1999).
Similar to these findings, the DRN in the present study
might reflect a conflict between the emotional response
to provocation and the cognitive motivation to refrain
from retaliation. Moreover, these results might be useful
to extend abovementioned fMRI data, as they can speak
also to the similarities or differences in the temporal dy-
namics of the neural correlates in purely cognitive tasks
or in tasks entailing emotion–cognition interferences.
However, the DRN might also reflect inhibitory pro-
cesses and self-regulation that takes place in these
participants (Knoch & Fehr, 2007), as suggested by the
negative correlation between the DRN and aggressive
retaliation. This would be in line with the aforemen-
tioned studies of the N2 in the context of response
inhibition (Pliszka et al., 2000; Falkenstein et al., 1999).
As pointed out previously, the present experimental
setting was quite different to the usual response inhibi-
tion tasks. Moreover, although the DRN had a similar
onset as the classical N2, a prolonged duration was
observable, which distinguishes it from results in go/
no-go or stop-signal studies. Recent fMRI studies have
extended these results by demonstrating a role for the
right PFC also in self-control and inhibition within the
scope of complex decision making and social interac-
tions (Knoch & Fehr, 2007; Knoch et al., 2006). More
research is clearly needed to establish the interpretation
of the DRN in terms of inhibition and its relation to
social decision making in the right PFC.
Although the exact functional meaning of the ob-
served DRN needs to be specified in further studies,
the involvement of executive processes in the control of
aggressive behavior is clearly implicated by this effect. As
we observed the DRN in the HT–LE group only, the
association of this effect with executive functioning
might be counterintuitive, given the usually reported
lower executive functions in HT aggressive people
(Giancola, 2004; Morgan & Lilienfeld, 2000). However,
an inverse relationship between trait aggressiveness and
executive functions has been reported particularly re-
garding clinical groups as persons with antisocial per-
sonality disorder or conduct disorder (Morgan &
Kra¨mer et al.
1473
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
8
e
-
1
p
4
d
6
f
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
/
.
t
f
o
n
1
8
M
a
y
2
0
2
1
Lilienfeld, 2000). With respect to healthy people, an effect
of executive functions on laboratory-induced aggression
has been demonstrated, which was differentially pro-
nounced for men and women and under intoxicated and
sober conditions (Giancola, 2004; Hoaken, Shaughnessy,
& Pihl, 2003). The participants of the present study were
students (i.e., mentally healthy, high-functioning individ-
uals), thus a weaker correlation of executive functions
and trait aggressiveness can be expected. Moreover, the
current results do not exclude that the LT group had, in
general, better executive functions. However, the DRN
suggests that only the participants prone to aggressive
behavior had to recruit executive functions when pro-
voked to refrain from retaliation.
This allows us to make differential predictions of the
effects of manipulations that interfere with executive
functions (e.g., alcohol intoxication, dual-task settings).
As the LT aggressive group stayed at ease during the
provocation without higher cognitive effort, interference
with their executive functions should have little effect on
aggressive behavior. In contrast, one would predict
higher aggression also in the HT–LE participants under
conditions interfering with their cognitive control pro-
cesses. In fact, a similar behavioral effect has been
observed by Giancola (2002), who showed that alcohol
consumption led to a more pronounced increase of ag-
gressive behavior in men with higher as opposed to lower
levels of trait anger. There is evidence suggesting that the
alcohol–aggression relationship is mainly driven by a
disruption of executive functions (Hoaken, Giancola, &
Pihl, 1998). We propose that the DRN effect in the current
study is the neural correlate of aggression-controlling
executive processes. This is in line with an fMRI study
employing the TAP (Kra¨mer et al., 2007), which reported
prefrontal activations during the decision phase, reflect-
ing the enhanced cognitive processing associated with
aggressive retaliation.
Outcome Phase
During the outcome phase, when participants got feed-
back whether they could administer the punishment to
the opponents or were exposed to the aversive noise
themselves, again, specific modulations of the neuro-
physiological responses by situational and personality
factors were observed.
Negative feedback, that is, the warning that the par-
ticipant lost and would be punished, elicited a fronto-
centrally located enhanced negativity, peaking at about
300 msec. This FRN was independent of the actual level
of provocation (i.e., the to-be-expected punishment).
This is consistent with previous studies showing that the
FRN is unaffected by the punishment’s magnitude
(Yeung & Sanfey, 2004). However, the difference be-
tween ERPs following positive and negative feedback
was related to the participants’ trait aggressiveness and
their aggressive behavior in the experiment. Although
highly aggressive participants showed a clear difference
between losing and winning trials, this difference was
smaller in the nonaggressive participants. In fact, an
FRN, although slightly smaller than in the loss trials,
seemed to be present in win trials also in the nonag-
gressive participants. To the extent to which the FRN
can be seen as a direct reflection of the perceived
valence of a stimulus, this suggests that winning (and
thus being able to punish the opponent) is associated
with negative feelings for these participants.
This interpretation is supported by recent reports of
an FRN elicited by observing another person losing
money in a gambling task, depending on the affective
engagement of the observer (Fukushima & Hiraki, 2006;
Yu & Zhou, 2006). This suggests that the FRN can be
driven by empathic involvement in the outcome of
actions for another person, thus extending earlier stud-
ies showing a modulation of the FRN by the person’s
involvement (Yeung et al.,
motivation and emotional
2005). As the FRN is believed to be generated in ACC,
this fits well with the hypotheses that this region is
critical for the integration of emotional and cognitive
information in the control of actions (Paus, 2001). The
finding of an enhanced FRN in win trials in nonaggres-
sive participants might thus be taken as an indicator of
their empathy with the to-be-punished opponent. This
dovetails with the results of the aforementioned fMRI
study of reactive aggression (Lotze et al., 2007), demon-
strating higher activation in the ventromedial PFC in less
callous (thus supposedly more empathic) participants
during the observation of the opponent suffering. This
observation is especially interesting, as one might argue
that such an emotional modulation of the punishment’s
evaluation is causing the regulation of aggressive behav-
ior in participants. Indeed,
in the present study, less
aggressive subjects selected lower punishments, in par-
ticular, in trials following wins. This further supports the
impact of the outcome’s emotional evaluation on the
aggressive behavior. Further studies relating this effect
with, for instance, self-report data on individuals’ empa-
thy will be needed to clarify the kind of emotional
evaluation that exerts an impact here.
Implications for a Model of Aggression
The neurophysiological findings can be used to further
specify the General Aggression Model (Anderson &
Bushman, 2002). A trivial prediction is, of course, that
people with HT aggressiveness should show more ag-
gressive behavior, especially when provoked. Although
this was true for our participants’ average behavior, we
also observed participants showing nonaggressive behav-
ior irrespective of their high predisposition for aggres-
sion. Although the model acknowledges that individual
differences in decision and evaluation processes may
modulate aggressive behavior, the present data provide
evidence for differences in executive control processes
1474
Journal of Cognitive Neuroscience
Volume 20, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
8
e
-
1
p
4
d
6
f
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
.
t
/
f
.
o
n
1
8
M
a
y
2
0
2
1
instigated by the frontal cortex that modulate aggressive
behavior under provocation in HT aggressive partici-
pants. No such provocation effect on cognitive control
processes was found in the LT aggressive group, likely
because their predisposition to react in a nonaggressive
way does not necessitate the recruitment of executive
control functions to ameliorate aggressive reactions.
Our data thereby specify the role of executive func-
tions in modulating aggressive behavior that has been
postulated in previous studies (Hoaken et al., 2003;
Blair, 2001). Based mainly on studies with sociopathic
patients, different models have been proposed that tap
into the regulatory impact of executive functions on
reactive aggression (Blair, 2001). Thus, some models
stress the executive function of ‘‘inhibitory control,’’
explaining aggressive outbursts as failed inhibitions of
violent impulses (Krakowski, 2003). This is corroborated
by ERP studies demonstrating reduced inhibitory control
in psychopathics, reflected in a diminished no-go-related
N2 component or impaired performance (Munro et al.,
2007; Kiehl, Smith, Hare, & Liddle, 2000). Other models
rather point at a disturbed interplay of executive func-
tions and emotion (Blair, 2001). The social response re-
versal model, for instance, states, that the orbito-frontal
cortex is modulating social behavior on the basis of social
cues such as angry facial expressions (Blair, 2004). Accord-
ingly, aggressive behavior in patients with orbito-frontal
lesions is ascribed to their deficits in processing angry fa-
cial expressions.
Although inhibitory control models cannot account
for a range of psychopathological observations, especial-
ly concerning impulsivity, the present data stress the
relevance of inhibition in the control of aggressive
tendencies in healthy participants. This is implicated
by the relation of the DRN and the participants’ ability
to refrain from retaliation. Inconsistent findings on
inhibition deficits in sociopathic patients might well be
due to differences in pathology or experimental tasks
employed. With the present data, we suggest one way to
clarify the relation of inhibitory processes and aggres-
sion and extend it to healthy, high-functioning people.
The modulation of the FRN by empathy for the
opponent, on the other hand, points to a mechanism
how emotional evaluation affects the regulation of ag-
gressive behavior. As pointed out previously, it might
well be that the outcome’s evaluation is driving the
enhanced executive control processes during the deci-
sion phase, triggered by a change in reward expectancies
(Blair, 2004; Rolls, 2000). Further studies will have to
clarify the interaction of other personality traits (namely
empathy) and executive functions in the regulation of
reactive aggression. This extends results from an fMRI study
on reactive aggression (Kra¨mer et al., 2007). The authors
report enhanced blood oxygenation level-dependent
responses in the dorsal striatum and in the dorsal part
of ACC during the decision for retaliation. The dorsal
ACC activation is related to the processing of reward
expectations on the one hand and possible behavioral
costs (by risking higher punishments) on the other
hand. This emphasizes the role of action monitoring
and processing of reward contingencies in the control of
aggression. Moreover, our data indicate that not only
angry facial expressions, as suggested by Blair (2001),
but also empathic feelings might have an impact on
social response reversal mechanisms, in healthy humans
at least.
Acknowledgments
This work was supported by grants from the BMBF and from
the DFG to T. F. M. We thank Joachim Weimann, Department
for his
of Economics of
cooperation in recruiting the participants.
the University of Magdeburg,
Reprint requests should be sent to Thomas F. Mu¨nte, De-
partment of Neuropsychology, Otto-von-Guericke University,
Universita¨tsplatz 2, 39106 Magdeburg, Germany, or via e-mail:
thomas.muente@medizin.ovgu.de.
REFERENCES
Anderson, C. A., & Bushman, B. J. (2002). Human aggression.
Annual Review of Psychology, 53, 27–51.
Anderson, C. A., Lindsay, J. J., & Bushman, B. J. (1999).
Research in the psychological laboratory: Truth or triviality?
Current Directions in Psychological Science, 8, 3–9.
Anderson, S. W., Bechara, A., Damasio, H., Tranel, D., &
Damasio, A. R. (1999). Impairment of social and moral
behavior related to early damage in human prefrontal
cortex. Nature Neuroscience, 2, 1032–1037.
Baron, R. A. (1977). Human aggression. New York: Plenum.
Bernstein, S., Richardson, D., & Hammock, G. (1987).
Convergent and discrimant validity of the Taylor and Buss
measures of physical aggression. Aggressive Behavior, 13,
15–24.
Bettencourt, B. A., & Miller, N. (1996). Gender differences in
aggression as a function of provocation: A meta-analysis.
Psychological Bulletin, 119, 422–447.
Bettencourt, B. A., Talley, A., Benjamin, A. J., & Valentine, J.
(2006). Personality and aggressive behavior under provoking
and neutral conditions: A meta-analytic review.
Psychological Bulletin, 132, 751–777.
Blair, R. J. R. (2001). Neurocognitive models of aggression,
the antisocial personality disorders, and psychopathy.
Journal of Neurology, Neurosurgery and Psychiatry, 71,
727–731.
Blair, R. J. R. (2004). The roles of orbital frontal cortex in the
modulation of antisocial behavior. Brain and Cognition, 55,
198–208.
Botvinick, M., Nystrom, L. E., Fissell, K., Carter, C. S.,
& Cohen, J. D. (1999). Conflict monitoring versus
selection-for-action in anterior cingulate cortex. Nature,
402, 179–181.
Bruin, K. J., & Wijers, A. A. (2002). Inhibition, response mode,
and stimulus probability: A comparative event-related
potential study. Clinical Neurophysiology, 113, 1172–1182.
Bushman, B. J., & Anderson, C. A. (2001). Is it time to pull the
plug on the hostile versus instrumental aggression
dichotomy. Psychological Review, 108, 273–279.
Cohen, J., Cohen, P., West, S. G., & Aiken, L. S. (2003). Applied
multiple regression/correlation analysis for the behavioral
sciences (3rd ed.). Mahwah, NJ: Erlbaum.
Kra¨mer et al.
1475
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
8
e
-
1
p
4
d
6
f
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
/
.
t
.
o
n
1
8
M
a
y
2
0
2
1
Eagly, A. H., & Steffen, V. J. (1986). Gender and aggressive
behavior: A meta-analytic review of the social psychological
literature. Psychological Bulletin, 100, 309–330.
Etkin, A., Egner, T., Peraza, D. M., Kandel, E. R., & Hirsch, J.
(2006). Resolving emotional conflict: A role for the rostral
anterior cingulate cortex in modulating activity in the
amygdala. Neuron, 51, 871–882.
Falkenstein, M., Hoormann, J., & Hohnsbein, J. (1999). ERP
components in Go/NoGo tasks and their relation to
inhibition. Acta Psychologica, 101, 267–291.
Frodi, A., Macaulay, J., & Thome, P. R. (1977). Are women
always less aggressive than men? A review of the
experimental literature. Psychological Bulletin, 84,
634–660.
Fukushima, H., & Hiraki, K. (2006). Perceiving an opponent’s
loss: Gender-related differences in the medial-frontal
negativity. Social, Cognitive and Affective Neuroscience, 1,
149–157.
Garavan, H., Ross, T. J., & Stein, E. A. (1999). Right hemispheric
dominance of inhibitory control: An event-related functional
MRI study. Proceedings of the National Academy of
Sciences, U.S.A., 96, 8301–8306.
Gehring, W. J., & Willoughby, A. R. (2002). The medial frontal
cortex and the rapid processing of monetary gains and
losses. Science, 295, 2279–2282.
Giancola, P. R. (2002). The influence of trait anger on
the alcohol–aggression relation in men and women.
Alcoholism: Clinical and Experimental Research, 26,
1350–1358.
Giancola, P. R. (2003). The moderating effects of
dispositional empathy on alcohol-related aggression
in men and women. Journal of Abnormal Psychology, 112,
275–281.
Giancola, P. R. (2004). Executive functioning and
alcohol-related aggression. Journal of Abnormal
Psychology, 113, 541–555.
Giancola, P. R., & Zeichner, A. (1995). Construct validity of a
competitive reaction-time aggression paradigm. Aggressive
Behavior, 21, 199–204.
Krakowski, M. (2003). Violence and serotonin: Influence of
impulse control, affect regulation, and social functioning.
Journal of Neuropsychiatry and Clinical Neurosciences,
15, 294–305.
Kra¨mer, U. M., Jansma, H., Tempelmann, C., & Mu¨nte, T. F.
(2007). Tit-for-tat: The neural basis of reactive aggression.
Neuroimage, 38, 203–211.
Lindsay, J. J., & Anderson, C. A. (2000). From antecedent
conditions to violent actions: A general affective aggression
model. Personality and Social Psychology Bulletin, 26,
533–547.
Lotze, M., Veit, R., Anders, S., & Birbaumer, N. (2007). Evidence
for a different role of the ventral and dorsal medial
prefrontal cortex for social reactive aggression: An
interactive fMRI study. Neuroimage, 34, 470–478.
Luu, P., & Tucker, D. M. (2001). Regulating action: Alternating
activation of midline frontal and motor cortical networks.
Clinical Neurophysiology, 112, 1295–1306.
McCarthy, G., & Wood, C. C. (1985). Scalp distributions of
event-related potentials: An ambiguity associated with
analysis of variance models. Electroencephalography and
Clinical Neurophysiology, 62, 203–208.
Miltner, W. H. R., Braun, C. H., & Coles, M. G. H. (1997).
Event-related brain potentials following incorrect feedback
in a time-estimation task: Evidence for a ‘‘generic’’ neural
system for error-detection. Journal of Cognitive
Neuroscience, 9, 788–798.
Morgan, A. B., & Lilienfeld, S. O. (2000). A meta-analytic
review of the relation between antisocial behavior and
neuropsychological measures of executive function.
Clinical Psychology Review, 20, 113–136.
Mu¨ller, S. V., Mo¨ller, J., Rodriguez-Fornells, A., & Mu¨nte, T. F.
(2005). Brain potentials related to self-generated and
external information used for performance monitoring.
Clinical Neurophysiology, 116, 63–74.
Munro, G. E. S., Dywan, J., Harris, G. T., McKee, S., Unsal, A.,
& Segalowitz, S. J. (2007). Response inhibition in
psychopathy: The frontal N2 and P3. Neuroscience Letters,
418, 149–153.
Hampel, R., & Selg, H. (1975). Fragebogen zur Erfassung von
Nieuwenhuis, S., Slagter, H. A., von Geusau, N. J. A.,
Aggressivita¨tsfaktoren. Goettingen: Hogrefe.
Hoaken, P. N., Giancola, P. R., & Pihl, R. O. (1998). Executive
cognitive functions as mediators of alcohol-related
aggression. Alcohol and Alcoholism, 33, 47–54.
Heslenfeld, D. J., & Holroyd, C. B. (2005). Knowing good
from bad: Differential activation of human cortical areas by
positive and negative outcomes. European Journal of
Neuroscience, 21, 3161–3168.
Hoaken, P. N., Shaughnessy, V. K., & Pihl, R. O. (2003).
Nieuwenhuis, S., Yeung, N., van den Wildenberg, W., &
Executive cognitive functioning and aggression: Is it an issue
of impulsivity? Aggressive Behavior, 29, 15–30.
Huynh, H., & Feldt, L. S. (1976). Estimation of the box
correction for degrees of freedom from sample data in
randomized block and splitsplot designs. Journal of
Educational Statistics, 1, 69–82.
Joyce, C. A., Gorodnitsky, I. F., & Kutas, M. (2004). Automatic
removal of eye movement and blink artifacts from EEG data
using blind component separation. Psychophysiology, 41,
313–325.
Kiehl, K. A., Smith, A. M., Hare, R. D., & Liddle, P. F. (2000). An
event-related potential investigation of response inhibition
in schizophrenia and psychopathy. Biological Psychiatry,
48, 210–221.
Knoch, D., & Fehr, E. (2007). Resisting the power of
temptations. The right prefrontal cortex and self-control.
Annals of the New York Academy of Sciences, 1104,
123–134.
Ridderinkhof, K. R. (2003). Electrophysiological correlates of
anterior cingulate function in a go/no-go task: Effects of
response conflict and trial type frequency. Cognitive,
Affective, & Behavioral Neuroscience, 3, 17–26.
Paus, T. (2001). Primate anterior cingulate cortex: Where
motor control, drive and cognition interface. Nature
Reviews Neuroscience, 2, 417–424.
Pfefferbaum, A., Ford, J. M., Weller, B. J., & Kopell, B. S. (1985).
ERPs to response production and inhibition.
Electroencephalography and Clinical Neurophysiology, 60,
423–434.
Pliszka, S. R., Liotti, M., & Woldorff, M. G. (2000). Inhibitory
control in children with attention-deficit/hyperactivity
disorder: Event-related potentials identify the processing
component and timing of an impaired right-frontal
response-inhibition mechanism. Biological Psychiatry, 48,
238–246.
Rolls, E. T. (2000). The orbitofrontal cortex and reward.
Knoch, D., Gianotti, L. R. R., Pascual-Leone, A., Treyer, V.,
Cerebral Cortex, 10, 284–294.
Regard, M., Hohmann, M., et al. (2006). Disruption of right
prefrontal cortex by low-frequency repetitive transcranial
magnetic stimulation induces risk-taking behavior. Journal
of Neuroscience, 26, 6469–6472.
Sanfey, A. G., Rilling, J. K., Aronson, J. A., Nystrom, L. E., &
Cohen, J. D. (2003). The neural basis of economic
decision-making in the ultimatum game. Science, 300,
1755–1758.
1476
Journal of Cognitive Neuroscience
Volume 20, Number 8
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
8
e
-
1
p
4
d
6
f
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
/
f
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
Sasaki, K., & Gemba, H. (1986). Electrical activity in the
prefrontal cortex specific to no-go reaction of conditioned
hand movement with colour discrimination in the monkey.
Experimental Brain Research, 64, 603–606.
Taylor, S. P. (1967). Aggressive behavior and physiological
arousal as a function of provocation and the tendency to
inhibit aggression. Journal of Personality, 35, 297–310.
Taylor, S. P., & Epstein, S. (1967). Aggression as a function
of the interaction of the sex of the aggressor and the
sex of the victim. Journal of Personality, 35, 474–485.
van Veen, V., & Carter, C. S. (2002). The timing of
action-monitoring processes in the anterior cingulate
cortex. Journal of Cognitive Neuroscience, 14, 593–602.
Yeung, N., Holroyd, C. B., & Cohen, J. D. (2005). ERP correlates
of feedback and reward processing in the presence and
absence of response choice. Cerebral Cortex, 15, 535–544.
Yeung, N., & Sanfey, A. G. (2004). Independent coding of
reward magnitude and valence in the human brain. Journal
of Neuroscience, 24, 6258–6264.
Yu, R., & Zhou, X. (2006). Brain responses to outcomes of
one’s own and other’s performance in a gambling task.
NeuroReport, 17, 1747–1751.
D
o
w
n
l
o
a
d
e
d
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
f
r
o
m
D
h
o
t
w
t
n
p
o
:
a
/
d
/
e
m
d
i
f
t
r
o
p
m
r
c
h
.
s
p
i
l
d
v
i
e
r
e
r
c
c
t
.
h
m
a
i
r
e
.
d
u
c
o
o
m
c
/
n
j
a
o
r
c
t
i
n
c
/
e
a
-
p
r
d
t
i
2
c
0
l
8
e
-
1
p
4
d
6
f
4
/
1
2
9
0
3
/
6
8
9
/
7
1
4
4
o
6
c
4
n
/
1
2
0
7
0
5
8
9
6
2
5
0
1
1
0
/
3
j
o
p
c
d
n
.
b
y
2
0
g
0
u
8
e
.
s
t
2
o
0
n
1
0
0
3
7
.
S
p
e
d
p
f
e
m
b
y
b
e
g
r
u
2
0
e
2
s
3
t
/
j
.
f
t
.
.
/
o
n
1
8
M
a
y
2
0
2
1
Kra¨mer et al.
1477