The Regulation of Cognitive Control following Rostral
Anterior Cingulate Cortex Lesion in Humans
Giuseppe di Pellegrino1,2, Elisa Ciaramelli1,2,
and Elisabetta La`davas1,2
Abstrait
& The contribution of the medial prefrontal cortex, partic-
ularly the anterior cingulate cortex (ACC), to cognitive control
remains controversial. Ici, we examined whether the rostral
ACC is necessary for reactive adjustments in cognitive con-
trol following the occurrence of response conflict [Botvinick,
M.. M., Plus courageux, T. S., Barch, D. M., Carter, C. S., & Cohen, J.. D.
Conflict monitoring and cognitive control. Psychological Re-
voir, 108, 624–652, 2001]. To this end, we assessed 8 patients
with focal lesions involving the rostral sector of the ACC (rACC
patients), 6 patients with lesions outside the frontal cortex
(non-FC patients), et 11 healthy subjects on a variant of
the Simon task in which levels of conflict were manipulated
on a trial-by-trial basis. More specifically, we compared Simon
effects (c'est à dire., the difference in performance between congruent
and incongruent trials) on trials that were preceded by high-
conflit (c'est à dire., incongruent) trials with those on trials that were
preceded by low-conflict (c'est à dire., congruent) trials. Normal con-
trols and non-FC patients showed a reduction of the Simon
effect when the preceding trial was incongruent, suggestive of
an increase in cognitive control in response to the occurrence
of response conflict. In contrast, rACC patients attained com-
parable Simon effects following congruent and incongruent
events, indicating a failure to modulate their performance de-
pending on the conflict level generated by the preceding trial.
En outre, damage to the rostral ACC impaired the poster-
ror slowing, a further behavioral phenomenon indicating re-
active adjustments in cognitive control. These results provide
insights into the functional organization of the medial prefron-
tal cortex in humans and its role in the dynamic regulation of
cognitive control. &
INTRODUCTION
Many views of cognition posit the existence of executive
or supervisory control mechanisms that guide and flex-
ibly adapt behavior to current goals or intentions (par exemple.,
Umilta`, 1988; Baddeley, 1986; Norman & Shallice, 1986;
Shiffrin & Schneider, 1977). Cognitive control is mainly
exerted when we need to ignore interfering stimuli, ou
overcome prepotent responses, particularly in novel and
complex tasks. A number of theories have been pro-
posed for how executive control might achieve this
(Miller & Cohen, 2001; Désimone & Duncan, 1995;
Cohen & Servan-Schreiber, 1992). Recently, cependant,
the debate has centered on how the cognitive system
determines the need to recruit cognitive control, et
dynamically regulates its influence on processing. C'est
argued that adjustments of top-down control occur on-
line on the basis of the amount of conflict induced by
competing stimuli or responses. One prominent theory
(Botvinick, Cohen, & Carter, 2004; Botvinick, Plus courageux,
Barch, Carter, & Cohen, 2001) proposes that the medial
frontal cortex (MFC), particularly the anterior cingulate
1Universita` di Bologna, Italy, 2Centro Studi e Ricerche in Neuro-
scienze Cognitive, Cesena, Italy
cortex (ACC), constantly monitors for response con-
flicts in information processing, triggering other systems
(housed in the lateral prefrontal cortex [PFC]) to im-
plement strategic processes when conflict occurs. Al-
though the MFC has been consistently implicated in
situations demanding such performance-monitoring ac-
tivité (Ridderinkhof, Ullsperger, Vieille femme, & Nieuwenhuis,
2004), uncertainties remain regarding which specific sub-
area within the MFC is essential for this process (voir
Rushworth, Kennerley, & Walton, 2005).
Neuroanatomical studies reveal that the medial sur-
face of the human frontal lobe is highly differentiated
in terms of cytoarchitecture and connectivity with other
brain regions (Picard & Strick, 1996). The ACC, in par-
particulier, can be parsed into two major subdivisions.
Firstly, the rostral ACC, which lies anterior and ventral
to the genu of the corpus callosum, and has dense
projection to limbic areas, including the orbito-frontal
insula, and amygdala (O¨ ngu¨r & Prix, 2000).
cortex,
Deuxièmement, the dorsal ACC, which is located above the
corpus callosum, and connects with the lateral PFC and
motor systems (Luppino, Rozzi, Calzavara, Matelli, 2003;
Paus, 2001). Based on evidence from functional imag-
ing studies, it has been suggested that the rostral and
dorsal subregions of the ACC subserve distinct affective
D 2007 Massachusetts Institute of Technology
Journal des neurosciences cognitives 19:2, pp. 275–286
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
un
/
d
m
e
je
d
t
F
r
p
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
m
un
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
t
n
je
c
/
e
un
–
r
p
t
d
je
c
1
je
9
e
2
–
2
p
7
d
5
F
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
oui
7
g
.
toi
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
.
.
.
.
F
t
.
o
n
1
8
M.
un
oui
2
0
2
1
and cognitive functions, respectivement (Allman, Hakeem,
Erwin, Nimchinsky, & Hof, 2001; Paus, 2001; Bush, Luu,
& Posner, 2000).
Consistent with this hypothesis, several functional
neuroimaging studies have demonstrated enhanced ac-
tivity in the dorsal ACC during a variety of cognitively
demanding tasks, specifically those involving response
conflit, such as the Stroop, the Eriksen, the go/no-go,
and Simon tasks (for a review, see Botvinick et al., 2004;
Rushworth, Walton, Kennerley, & Bannerman, 2004;
Paus, 2001). Cependant, despite the ubiquity of the dorsal
ACC activation in functional studies, lesion data do not
convincingly implicate this brain region in the detection
and resolution of response conflict (Baird et al., 2006;
Swick, & Jovanovich, 2002; Swick & Turken, 2002; Stuss,
Floden, Alexander, Levine, & Katz, 2001; Vendrell et al.,
1995). Notably, executive functions and performance
monitoring can be entirely normal despite severe dam-
age of the dorsal ACC (Fellows & Farah, 2005; Critchley
et coll., 2003).
The rostral ACC and the adjoining PFC, by contrast,
have been primarily implicated in the evaluation of emo-
tional and reward-related information (Phillips, Drevets,
Rauch & voie, 2003un; Whalen et al., 1998; Devinsky,
Morrell, & Vogt, 1995), and the way in which this in-
formation guides decision making (Bechara, Tranel, &
Damasio, 2000). Humans with lesion including these
prefrontal regions have been typically described as
apathetic and unconcerned when committing errors
(Eslinger & Damasio, 1985), and demonstrate abnormal
control of autonomic arousal states (Critchley, 2005).
De plus, functional abnormalities in the rostral ACC
have been reported in patients suffering from psychiat-
ric illness, such as schizophrenia and mood disorders
(Laurens, Ngan, Bates, Kiehl, & Liddle, 2003; Phillips,
Drevets, Rauch & voie, 2003b; Drevets et al., 1997).
Cependant, the affective–cognitive parcellation of the
ACC remains a contentious subject, and a substantial
body of evidence suggests that each region may not be
functionally exclusive (Critchley, 2005; Davis et al., 2005;
Eisenberger & Lieberman, 2004). Par exemple, several
electrophysiological studies have suggested that the
error-related negativity (ERN), a scalp potential com-
monly taken as an index of performance monitoring
(Falkenstein, Hohnsbein, & Hoormann, 1991), engages
both the ‘‘affective’’ rostral and ‘‘cognitive’’ dorsal por-
tions of the ACC (Luu, Tucker, Derryberry, Reed, &
Poulsen, 2003; Luu, Flaisch, & Tucker, 2000). De plus,
a number of brain imaging studies have revealed the
involvement of both subdivisions of the ACC in pro-
cessing conflicts between different information process-
ing pathways ( Wager, En Ionie, Forgeron, & Nichols, 2005;
Dreher & Grafman, 2003; Menon, Adleman, Blanc,
Glover, & Reiss, 2001; Kiehl, Forgeron, Hare, & Liddle,
2000). Fait intéressant,
in control demanding tasks, le
dorsal ACC activity increases while the rostral ACC ac-
intercon-
tivity decreases,
implying close functional
nection between ACC subregions in the regulation of
cognition (Polli et al., 2005; Bush et al., 1998; Drevets &
Raichle, 1998).
Although the results of these studies suggest that a role
in executive processes extends to the rostral portion of
the cingulate, it is currently unclear whether this brain
structure is essential for those functions. Ainsi, functional
imaging studies can suggest merely the involvement of a
brain region in a functional system, but cannot confirm
whether an individual component is necessary for nor-
mal functioning. À ce jour, only few neuropsychological
studies have explicitly looked at compensatory adjust-
ments in control following cingulate damage (Fellows &
Farah, 2005; Swick & Jovanovich, 2002). None of these
studies have focused on the rostral sector of the ACC.
Nor have they confirmed whether the ACC performs an
essential role in conflict monitoring and on-line mobili-
zation of cognitive control.
Dans cette étude, we evaluated whether the rostral ACC is
necessary for the on-line modulation of control process
depending on the strength of response conflict. To in-
vestigate this, 8 patients with damage to the rostral ACC
and the adjacent medial PFC (rACC patients), 6 patients
with lesions outside the frontal lobe (non-FC patients),
et 11 healthy control subjects were tested on a version
of the Simon task in which levels of conflict and cog-
nitive control were manipulated on a trial-by-trial basis.
Using the Simon task (Simon, 1969), it is possible to
study how the cognitive system handles conflicts between
competing task dimensions. In this paradigm, sujets
must respond with a left or right keypress based on the
color (or shape) of a stimulus that is randomly presented
to the left or right of fixation. Although this stimulus
location is irrelevant for the task, reaction times (RTs) sont
faster and error rates are lower when the stimulus and
response location correspond (congruent trials), que
when they do not (incongruent trials). Interference (ou
Simon) effects (par exemple., the difference in performance be-
tween congruent and incongruent trials) are usually
explained in terms of conflict between two parallel routes
of response selection (Kornblum, Hasbroucq, & Osman,
1990). Typiquement, it is argued that there is an indirect route
that determines the correct response on the basis of the
relevant stimulus feature (par exemple., color), and a direct route
that automatically primes the response corresponding to
the position of the stimulus. Ainsi, if stimulus position
and correct response do not match (par exemple.,
left-sided
stimulus requires right-sided response), a response con-
flict arises, and the resolution of this conflict delays re-
sponse execution.
Critique, recent findings on the Simon task strongly
indicate that the prior context influences the size and
sign of the interference effects in subsequent trials. Plus
specifically, it has been reported that the Simon effect
decreases ( Wuhr & Ansorge, 2005), disappears (Stu¨rmer,
Leuthold, Soetens, Schro¨ter, & Sommer, 2002), or even
reverses (Hommel, Proctor, & Vu, 2004) when the pre-
276
Journal des neurosciences cognitives
Volume 19, Nombre 2
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
un
/
d
m
e
je
d
t
F
r
p
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
m
un
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
t
n
je
c
/
e
un
–
r
p
t
d
je
c
1
je
9
e
2
–
2
p
7
d
5
F
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
oui
7
g
.
toi
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
F
t
.
.
.
.
.
o
n
1
8
M.
un
oui
2
0
2
1
vious trial (trial n(cid:1)1) is incongruent, compared to when
trial n(cid:1)1 is congruent. Similar sequential effects have
also been demonstrated for other conflict tasks, tel que
the Stroop and Eriksen task (Egner, & Hirsh, 2005;
Kerns et al., 2004; Botvinick, Nystrom, Fissell, Carter, &
Cohen, 1999; Gratton, Coles, & Donchin, 1992). Accord-
ing to the conflict monitoring hypothesis (Botvinick
et coll., 1999, 2001), these sequential dependencies of
interference effects can be readily interpreted as an
example of task-induced regulation of control. On this
voir, whenever a conflict is detected, compensatory
adjustments take place, and more control is allocated
to a particular task. The purpose of these adjustments is
to eliminate, or at least reduce, the influence of the
irrelevant information, hence, an overall reduction of
the interference effect results. Some authors, cependant,
have challenged this account, suggesting that adjust-
ments effects are completely confounded with the pres-
ence of stimulus–response repetitions in successive
trials, and therefore, can be explained by bottom-up,
binding, or priming effects (Hommel et al., 2004; Mayr,
Awh, & Laurey, 2003; Notebaert, Soetens, & Melis, 2001).
Several recent studies, on the other hand, controlled for
such stimulus or response repeats and still observed
sequential modulations, making it difficult to attribute
these effects entirely to perceptual or motor priming
(Ullsperger, Bylsma, & Botvinick, 2005; Wuhr & Ansorge,
2005; Kerns et al., 2004).
The Simon paradigm elicits a sufficient number of
errors to allow examination of another indicator of
conf lict-mediated adjustments in control, posterror
slowing. This phenomenon refers to the fact that par-
ticipants performing speeded response tasks tend to
slow down following the occurrence of errors (Laming,
1968; Rabbitt, 1966). According to the conflict monitor-
ing model, errors are frequently associated with a high
degree of response conflict. This is a consequence of
the competition between the just executed incorrect
response and the (posterror) activation of the correct
response resulting from continued processing of the
stimulus (Yeung, Botvinick, & Cohen, 2004; Botvinick
et coll., 2001). Following errors, activation of the conflict-
related control process tends to shift the system toward
a more conservative strategy, producing slower but
more accurate responding in subsequent trials. Ici,
as in the case of sequential effects, conflict monitoring
provides crucial information in regulating cognitive pro-
cessing based on an ongoing evaluation of performance.
Such trial-by-trial sequential modulation of interfer-
ence effect and posterror slowing provides an excellent
means of investigating reactive, conflict-triggered adjust-
ments in top-down control in patients with rostral ACC
lesions. Accordingly, we directly compared Simon effects
following congruent and incongruent trials. If the ros-
tral ACC provides adaptability in performance according
to changing task demands, then strategic modulation
of the Simon interference effect depending on the na-
ture of the preceding trial (congruent vs. incongruent)
should not be seen in rACC patients. Par contre, normal
participants and non-FC patients would be expected to
show a significant decrease of the Simon interference
effect after incongruent compared to congruent trials.
De plus, rostral ACC damage is expected to result in
reduced adjustments (slowing) of the response genera-
tion process after commission of errors.
MÉTHODES
Participants
Three groups of subjects participated in the study: (un)
a group of patients with focal lesions centered on the
rostral ACC and the adjoining ventromedial PFC (rACC
group, n = 8, âge moyen = 55 années, SD = 6.1); (b) un
control group of patients with focal damage sparing
the frontal cortex (non-FC group, n = 6, âge moyen =
66.8 années, SD = 8.1); et (c) a control group of healthy
sujets (n = 11, âge moyen = 53.4 années, SD = 7.7), âge,
éducation, and sex ratio matched with the ACC group.
Brain-damaged patients were recruited from the Cen-
tre for Studies and Research in Cognitive Neuroscience in
Cesena. They were selected on the basis of the location
of their lesion evident on computed tomography (CT) ou
magnetic resonance imaging (IRM) scans. In rACC pa-
patients, lesions were the result of a ruptured aneurysm
of the anterior communicating artery. Included patients
were those who had lesion restricted to the rostral por-
tion of medial surface of the frontal lobe, and with no
other diagnosis likely to affect cognition or interfere with
the participation in the study (par exemple., significant psychiatric
maladie, alcohol misuse, history of cerebrovascular dis-
ease, focal neurological examination). In no case did pa-
tients selected for the study contain lesions extending
into the lateral PFC. Lesions were traced from CT or MRI
scans on standard templates (Damasio & Damasio, 1989)
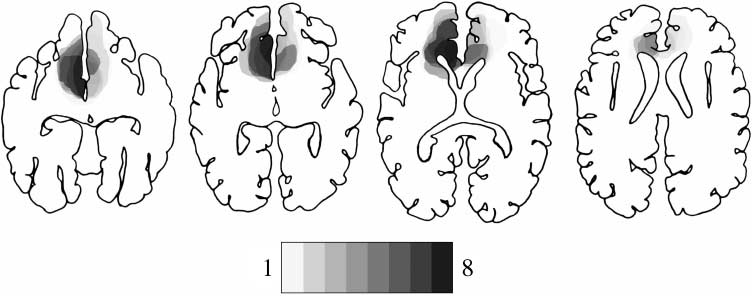
by a neurologist. Chiffre 1 shows the extent and over-
lap of the brain lesions in the rostral ACC group. Le
Brodmann’s areas affected in this group were areas 10,
12, 32, 24, with region of maximal overlap occurring in
Brodmann’s areas 32 and 24a–c (c'est à dire., the rostral portion
of the ACC), where all cases had lesions.
As for non-FC patients, their lesions were the result of
infarction or the removal of a meningioma (1 patient)
involving the cerebral cortex outside the frontal lobe,
mostly in the medial and lateral temporal lobe (n = 5).
The Brodmann’s areas affected in this group were areas
21, 22, 28, 36, 37, 38, 39.
Patients were not receiving psychoactive drugs at the
time of testing, and were all more than a year postonset.
They gave informed consent to participate in the study
according to the Declaration of Helsinki (International
Committee of Medical Journal Editors, 1991) et le
Ethical Committee of the Department of Psychology,
University of Bologna. Tableau 1 shows demographic data,
di Pellegrino, Ciaramelli, and La`davas
277
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
un
/
d
m
e
je
d
t
F
r
p
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
m
un
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
t
n
je
c
/
e
un
–
r
p
t
d
je
c
1
je
9
e
2
–
2
p
7
d
5
F
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
oui
7
g
.
toi
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
F
.
.
.
.
.
t
o
n
1
8
M.
un
oui
2
0
2
1
Chiffre 1. Location and
degree of overlap of brain
lesions. The figure shows the
lesions of the eight subjects
with rostral ACC lesion.
Lesions are projected on the
same four axial templates
following the method
developed by Damasio and
Damasio (1989). Progressively
darker shades denote the
degree to which lesions
involve the same brain regions,
as indicated in the legend.
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
un
/
d
m
e
je
d
t
F
r
p
o
r
m
c
.
h
s
je
p
je
v
d
e
je
r
r
e
c
c
h
t
.
m
un
je
r
e
.
d
c
toi
o
m
o
/
c
j
n
o
un
c
r
t
n
je
c
/
e
un
–
r
p
t
d
je
c
1
je
9
e
2
–
2
p
7
d
5
F
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
oui
7
g
.
toi
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
F
e
m
b
b
oui
e
r
g
2
toi
0
e
2
s
3
t
/
j
.
.
.
t
.
.
F
o
n
1
8
M.
un
oui
2
0
2
1
Tableau 1. Demographic, Clinique, and Lesion Data of the Two Patient Groups
Sex
Age at Test (Years)
Éducation
Side of Lesion
Etiology
Description of Lesion
MMSE Score
Rostral ACC Patient
1
2
3
4
5
6
7
8
M.
M.
M.
F
F
M.
F
M.
Non-FC Patient
1
2
3
4
5
6
M.
F
F
F
F
M.
49
56
56
62
52
66
53
46
58
75
61
82
77
58
8
13
13
8
19
5
8
8
13
10
5
5
8
8
L
B
R.
L
B
L
R.
R.
R.
R.
L
R.
R.
L
AcoA
Rostral ACC
Aneurysm
VmPFC
AcoA
Rostral ACC
Aneurysm
VmPFC
AcoA
Rostral ACC
Aneurysm
VmPFC
AcoA
Rostral ACC
Aneurysm
VmPFC
AcoA
Rostral ACC
Aneurysm
VmPFC
AcoA
Rostral ACC
Aneurysm
VmPFC
AcoA
Rostral ACC
Aneurysm
VmPFC
AcoA
Rostral ACC
Aneurysm
VmPFC
Infarction
Mesial Temporal
Infarction
Temporal Parietal
Infarction
Lateral Temporal
Tumor
Mesial Temporal
Infarction
Temporal pole
Infarction
Insula
Basal ganglia
26
27
24
24
27
23
23
24
25
23
–
23
24
26
M = male; F = female; L = left; R = right; B = bilateral; ACoA = Anterior Communicating Artery; ACC = Anterior Cingulate Cortex; VmPFC =
Ventromedial prefrontal cortex; MMSE = Mini-Mental State Examination.
278
Journal des neurosciences cognitives
Volume 19, Nombre 2
lesion side, etiology, lesion description, as well as the
Mini-Mental Status Examination score (MMSE; Folstein,
Robins, & Helzer, 1983) for each patient.
Normal participants were healthy volunteers who were
not taking psychoactive medication and were free of
current or past psychiatric or neurological illness as de-
termined by history. Normal controls scored at least 28
out of 30 on the MMSE. There were no significant differ-
ences between ACC patients and normal controls on the
measured demographic variables ( p > .7 in all cases).
Stimuli and Apparatus
In each experiment, stimuli were displayed on a 21-in. col-
or VGA monitor (1024 (cid:2) 768 spatial resolution, 16 color
bit) situated on a table top. An IBM-compatible Pentium IV
computer, running E-Prime software (Schneider, Eschman,
& Zuccolotto, 2002), controlled the presentation of stimu-
li, timing operation, and data collection. The subjects
were seated approximately 60 cm away from the screen.
All stimuli were presented on a black background. UN
fixation display, consisting of a central fixation cross (sub-
tending 0.48 (cid:2) 0.48) was present for the entire duration
du procès, except during the intertrial interval (ITI). Le
central cross was positioned at eye level, along the sub-
jects’ midline. Target stimuli were green or red circle out-
lines (4.58 in diameter), indicating left or right responses,
respectivement. Each target stimulus appeared approxi-
mately 68 either on the left or right of central fixation.
Responses were made by pressing the ‘‘Alt’’ key (on the
left side of the keyboard) with the index finger of the left
hand if a green circle was presented, and the ‘‘Ctrl’’ key
(on the right side of the keyboard) with the index finger
of the right hand, if a red circle was shown.
Procedure
All experiments took place in a dimly lit room. Sujets
were seated in front of the computer, which presented
the task instructions. These were also summarized by
the experimenter, after subjects had read them, to en-
sure that they had been understood.
Sequences of two consecutive trials were created,
somewhat arbitrarily, by increasing the ITI after every
second trial. In each sequence, the first trial was treated
as the prime or ‘‘previous trial,’’ and the second trial
as the probe or ‘‘current trial.’’ This procedure allowed
us to connect previous and current trials as closely as
possible, and to prevent effects of a trial sequence from
influencing the processing of the following ones.
When target stimuli appeared, participants were re-
quired to press the key corresponding to the color of
the circle, while ignoring the physical location of the
target stimuli. At the beginning of each trial sequence,
the fixation display appeared for 1000 msec, acting as
a warning signal. Alors, the first target stimulus (S1)
was presented until one of the keys was pressed (R1),
ou 3000 msec had elapsed, whichever occurred first.
Following a fixed interval of 1000 msec, the second
target stimulus (S2) was shown for up to 3000 msec
until a response (R2) was given. This started an ITI of
2000 msec during which the screen was blank. If par-
ticipants responded with the wrong key, or were slower
que 3000 msec after S1 or S2, it was considered re-
spectively an error or omission. No feedback was given,
and omission and error trials were not repeated.
Target stimuli were presented randomly in the left
and right visual space with equal probability, such that
the side of the first stimulus did not predict the location
of the second stimulus. The participants were informed
that there was no relationship between the S1 and S2
stimuli, and were instructed to respond as quickly as
possible while maintaining high levels of accuracy.
Design
Each participant received one practice block and two ex-
perimental blocks in two separate sessions. Every block
was composed of four repetitions of 16 randomly inter-
mixed unique trial sequences, resulting from the facto-
rial combination of two S1 positions (left vs. droite), (cid:2) deux
R1 locations (left vs. droite, correlated with S1 color), (cid:2)
two S2 positions (left vs. droite), (cid:2) two R2 locations (gauche
vs. droite, correlated with S2 color). Congruent and in-
congruent trials were in equal proportion. Trial se-
quences were classified according to the congruency
of the previous and current trial, generating four equi-
probable trial sequences: congruent–congruent (C–C),
congruent–incongruent (C–I),
incongruent–congruent
(I–C), and incongruent–incongruent (I–I). The depen-
dent variables were RTs (msec) and error rates. Quand
necessary, multiple comparisons were conducted using
the Newman –Keuls test. The level of significance was
set to p < .05 for all analyses.
RESULTS
Response times exceeding the range of 150–2000 msec
were discarded from analysis. This cutoff procedure
resulted in the exclusion of 2% of responses for normal
controls, and 3.8% and 4.1% for the rACC patients and
the non-FC patients, respectively. This outlier analysis
was done prior to all statistical analyses in all experi-
ments for previous and current trial RTs, and therefore
will not be mentioned again below. Of the remaining
data, correct median RTs and percentages of errors were
computed for each group of participants in each con-
dition of the previous and current trial.
We first tested whether the three participant groups
differed on overall task performance through a two-
factorial analysis of variance (ANOVA) on correct RTs,
with congruency (congruent and incongruent) as a
within-subject factor, and group (normal controls, rACC
di Pellegrino, Ciaramelli, and La`davas
279
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
2
-
2
p
7
d
5
f
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
.
.
t
.
.
.
o
n
1
8
M
a
y
2
0
2
1
patients, and non-FC patients) as a between-subject fac-
tor, pooling over previous and current trials. The main
effect of congruency and group were both significant
[F(1,22) = 130.4, p < .0001, and F(2,22) = 17.8, p <
.0001, respectively]. Of particular relevance, the effect
of congruency interacted with group [F(2,22) = 6.1,
p < .007], reflecting a smaller Simon effect (e.g., in-
congruent minus congruent RTs) in normal controls
(31 msec) relative to both non-FC (47 msec) and rACC
patients (65 msec). However, the Simon effect did not
differ between patient groups [F(1,12) = 1.6, p = .2].
Likewise, the Simon effect in error rate was significantly
higher in non-FC (1.1%) and rACC patients (1.3%) than
in healthy participants [0.4%; F(2,22) = 6.7, p < .005],
but the two patient groups did not differ ( p = .5).
significant [F(2,22) = 10.04, p < .001]. Thus, normal
control participants displayed significantly faster RTs
(561 msec) than did rACC (678 msec) and non-FC pa-
tients (726 msec; p < .007 in all cases). However, no
significant differences were observed between patient
groups ( p > .05). There was also a significant main ef-
fect of congruency [F(1,22) = 49.36, p < .001], with
faster RTs on congruent (623 msec) than incongruent
trials (686 msec), confirming the presence of the basic
Simon effect. The interaction between group and con-
gruency was not significant [F(2,22) = 1.82, p = .20].
Error rates followed a similar pattern. There was a sig-
nificant main effect of group [F(2,22) = 7.23, p< 0.003], a
significant effect of congruency [F(1,22) = 30.5, p < .001],
and no interaction between group and congruency
[F(2,22) = 2.2, p = .12].
Previous Trial
For previous trials, a two-factorial ANOVA, with congru-
ency (congruent and incongruent) as a within-subject
factor, and group (normal controls, rACC patients, and
non-FC patients) as a between-subject factor, was con-
ducted on the RT data. The main effect of group was
Current Trial
Incorrect RTs of the current trial, as well as RTs following
an error in the previous trial, were excluded from further
analyses. The remaining RTs (see Figure 2, left panel)
were subjected to a three-factorial ANOVA, with previous
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
2
-
2
p
7
d
5
f
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
.
t
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
Figure 2. Mean reaction times for congruent (C) and incongruent (I) current trials, reported separately for trials coming after congruent
(Previous C) and incongruent trials (Previous I) in the three participant groups (rACC = rostral anterior cingulate patients; non-FC = nonfrontal
patients; NC = normal controls). Error bars show standard error of the mean. Left panel depicts data from all current trials. Right panel
depicts data from 50% of current trials in which neither color of stimuli nor responses of the preceding trials were repeated.
280
Journal of Cognitive Neuroscience
Volume 19, Number 2
congruency (congruent and incongruent) and current
congruency (also, congruent and incongruent) as within-
subject factors, and group (normal controls, rACC pa-
tients, and non-FC patients) as a between-subject factor.
The main effect of group was significant [F(2,22) =
17.15, p < .0001], with normal controls responding fast-
er (516 msec) than either group of patients (671 and
659 msec, for non-FC and rACC patients, respectively).
There was also a main effect of current congruency
[F(1,22) = 82.67, p < .0001], reflecting shorter RTs
on congruent (584 msec) than on incongruent trials
(619 msec; e.g., standard Simon effect). The effect of
previous congruency was not significant [F(1,22) = 1.01,
p = .3].
Of particular interest in the present context were the
sequential influences of the Simon effect from a previ-
ous trial. The analysis revealed that the interaction
between previous congruency and current congruency
was significant [F(1,22) = 59.41, p < .0001], and its de-
tails were consistent with the conflict adaptation effect.
Thus, the Simon effect (e.g., incongruent minus con-
gruent RTs) was large after congruent trials (68 msec)
but tended to become negative after incongruent trials
((cid:1)4.5 msec). There was also a significant interaction be-
tween group and current congruency [F(2,22) = 9.33,
p = .001].
Of most importance for the present purposes, however,
there was a significant three-way interaction between
group, previous congruency, and current congruency
[F(2,22) = 5.00, p < .01], suggesting that the sequen-
tial modulation of the Simon effect differed across
groups. This interaction was further explored with two-
way ANOVAs performed separately on RTs from each
group of participants. Significant interactions between
previous congruency and current congruency were noted
for normal controls [F(1,10) = 33.93, p < .0001] and non-
FC patients [F(1,5) = 74.33, p < .001], but not for the
rACC group [F(1,7) = 3.11, p = .12]. Planned com-
parisons revealed that the Simon effect was significantly
larger following congruent trials than following incon-
gruent trials in normal control participants (57 msec vs.
(cid:1)23 msec) and non-FC patients (76 msec vs. (cid:1)28 msec),
but not in rACC patients (70 msec vs. 38 msec).
Error percentages (shown in Table 2) were also sub-
jected to a three-way ANOVA. The main effect of group
was significant [F(2,22) = 20.02, p < .0001]. Normal
controls tended to produce fewer errors (1%) than both
non-FC (2.4%) and rACC patients (2.2%). The significant
main effect of current congruency [F(1,22) = 43.47,
p < .0001] indicated a Simon effect in error rates, with
fewer errors on congruent trials (1.5%) than on incon-
gruent ones (2.4%). Moreover, the significant interaction
between previous congruency and current congruency
[F(1,22) = 22.15, p < .0001] signaled a sequential
modulation of the Simon effect. In particular, a signifi-
cant Simon effect in error percentages occurred after
congruent trials (1.7% of accuracy difference between
Table 2. Mean Percentages of Errors for Congruent (C) and
Incongruent (I) Current Trials, Reported Separately for Trials
Coming after Congruent (Previous C) and Incongruent Trials
(Previous I) in the Three Participant Groups
Previous C
Previous I
Group
Current C
Current I
Current C
Current I
rACC
non-FC
NC
1.5
1.3
0.6
3
3.7
1.8
1.7
2.5
1.1
2.9
2.1
0.6
rACC = rostral anterior cingulate patients; non-FC = nonfrontal pa-
tients; NC = normal controls.
incongruent and congruent trials) but not after incon-
gruent trials (0.1%). The three-way interaction was not
significant [F(2,22) = 3.0, p = .07]. Nevertheless, for
completeness, we also conducted planned comparisons
of the sequence effect for each individual group of
participants. We found a reliable sequential modulation
of the Simon effect in normal controls [1.2% vs. (cid:1)0.5%
of error rate difference after congruent vs. incongruent
trials, respectively; F(1,10) = 8.05, p < .01] and non-FC
patients [2.3% vs. (cid:1)0.3%; F(1,5) = 11.03, p < .02], but
not in rACC patients [1.6% vs.1.1%; F(1,7) = 3.50, p = .1].
Therefore, accuracy data corroborated the RT data.
Repetition Priming vs. Conflict Adaptation
As already noted in the Introduction, it is controversial
whether sequential modulation of interference effects
truly reflect task-induced variations in top-down control,
or depend on bottom-up repetition priming effects. Re-
cently, Mayr et al. (2003) demonstrated (in an Eriksen
flanker task) that when trial repetitions were excluded
from the data analysis, there was no reduction of the
interference effect after incongruent trials. They noted
that subjects may be faster on incongruent trials pre-
ceded by incongruent trials (I–I) than on incongruent
trials preceded by congruent trials (C–I) because half
of the I–I transitions but none of the C–I transitions
involved exact stimulus (and thus response) repetitions.
Therefore, they suggested that the substantial RT bene-
fits observed after incongruent trials may simply reflect
repetition priming, rather than adaptation of top-down
control. Given these considerations, it is important to
explore whether our results can be accounted for in
terms of repetition priming. To this end, 50% of the C–C
and I–I sequences that involved complete repetitions
(e.g., sequences in which stimulus color, stimulus posi-
tion, and the response repeat), and 50% of the C–I and
I–C that involved partial repetitions (e.g., sequences in
which stimulus color and response repeat, while stimu-
lus position changes) were eliminated from the dataset
(see Figure 2, right panel). Additional analyses focusing
on the sequential modulations of the Simon effect were
di Pellegrino, Ciaramelli, and La`davas
281
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
2
-
2
p
7
d
5
f
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
t
f
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
then performed on the remaining RTs. As before, the
analyses showed significant interactions between previ-
ous congruency and current congruency for normal
controls [F(1,10) = 21.71, p < .001] and non-FC patients
[F(1,5) = 21.43, p < .005], but not for the rACC group
[F(1,7) = 0.14, p = .7]. These interactions signaled that
a pronounced, positive Simon effect was present follow-
ing congruent trials, but it reversed following incongru-
ent trials in normal controls (77 msec vs. (cid:1)27 msec) and
non-FC patients (68 msec vs. (cid:1)17 msec). By contrast, a
robust Simon effect occurred in rACC patients, regard-
less of congruency of previous trial (68 msec vs. 61 msec,
following congruent vs. incongruent trials, respectively).
Posterror Slowing
To test the prediction that rACC patients are impaired in
posterror slowing, we performed an ANOVA with group
as a between-subject factor, and trial (trial after error and
trial after correct response) as a within-subject factor.
The normal control group was limited to those par-
ticipants who made more than three errors (n = 9). For
all groups, correct responses on trials following errors
were slower than following correct trials [F(2,20) =
20.3, p < .001]. Most importantly, there was a signifi-
cant interaction between group and trial [F(2,20) =
3.94, p < .003]. Planned comparisons revealed that
normal controls and non-FC patients exhibited a size-
able, significant posterror slowing (76 msec, p <
.001, and 63 msec, p < .003, respectively). In contrast,
rACC patients showed a nonsignificant posterror slowing
(8 msec, p = .6).
DISCUSSION
The role of the more rostral aspect of the medial PFC
in executive processes is not well understood. Classical
neurological evidence suggests that patients with lesion
involving this region appear to be intellectually unim-
paired, and demonstrate apparently normal performance
on a host of neuropsychological measures (Teuber, 1964;
Rylander, 1947). More recent studies have essentially
confirmed these early observations (Bechara et al.,
2000; Eslinger & Damasio, 1985), but they also demon-
strate that some of these patients show behavioral deficits
on difficult, attention-demanding tasks (Burgess, Veitch,
de Lacy Costello, & Shallice, 2000). In the present study,
we examined whether the rostral ACC and the adjoining
medial PFC is necessary for rapid, on-line adjustments in
cognitive control, following the occurrence of response
conflict. Specifically, subjects were tested in a variant of
the Simon task in which interference effects were mea-
sured as a function of conflict level of the preceding trial.
Less interference was expected following events associat-
ed with response conflict, arguably due to increased
focusing of top-down control. The performance of indi-
viduals with focal damage to the rostral ACC was com-
pared with that of age-matched normal controls and
patients who had brain damage outside the frontal lobe.
Our neuropsychological data provide compelling new
evidence that the rostral ACC is critical for the dynamic
regulation of cognitive control. Rostral ACC patients
failed to appropriately modulate their performance in
response to the amount of conflict generated by the
task, showing a sizeable Simon interference effect not
only after congruent trials (which do not induce re-
sponse conflict) but also after incongruent trials (which
induce response conflict). By contrast, both healthy
controls and non-FC patients showed abolition of inter-
ference effects following incongruent events, thus re-
vealing that they were able to actively monitor their
performance and use this information to intensify the
cognitive control allocated to the task.
Because the two patient groups had comparable
interference effects after the occurrence of congruent
trials, as well as on n(cid:1)1 (or previous) trials, it appears
unlikely that the inefficiency of rACC patients after
incongruent trials can be explained entirely by group
differences in mental effort or motivation. Moreover,
control analyses suggested that group differences in
the sequential modulation of the Simon effect cannot
be attributed to low-level priming effects from one trial
to the next (Mayr et al., 2003), but truly reflect differ-
ences in compensatory adjustments of cognitive control
( Wuhr & Ansorge, 2005; Kerns et al., 2004; Stu¨rmer
et al., 2002). Indeed, our pattern of results persists even
when repetition of identical stimuli (and responses) in
trial sequences was excluded from analysis. Finally,
both healthy participants and non-FC patients tended
to adopt a more cautious response mode after errors.
This was not the case for rACC patients, who exhibited
a near absence of posterror slowing. Together, these
findings reinforce the proposal, put forth by Botvinick
et al. (2001), that both postconflict and posterror com-
pensatory adjustments in performance may depend on
a single functional mechanism.
These results have several implications for theories of
executive control and ACC function. We first discuss how
rostral ACC damage can disrupt context-sensitive control
adjustments. Cognitive theorists distinguish between
two executive functions involved in the flexible modu-
lation of control: an evaluative function, which detects
conflict between competing behavioral responses, and
a strategic function, which implements control adjust-
ments for conflict reduction. According to one perspec-
tive (the conflict monitoring hypothesis, Botvinick et al.,
2001), the ACC plays a critical role in conflict and error
monitoring, but it is not responsible for the allocation of
control. Conversely, the selection for action hypothesis
(Posner & DiGirolamo, 1998; Posner & Petersen, 1990)
maintains that the ACC implements cognitive control
directly by biasing or selecting task-relevant responses
against strong habitual behaviors. It is difficult, however,
282
Journal of Cognitive Neuroscience
Volume 19, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
2
-
2
p
7
d
5
f
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
f
.
.
.
t
.
o
n
1
8
M
a
y
2
0
2
1
to adjudicate between these two alternative views based
on current lesion data. This is because impairments of
either evaluative or strategic processes would result in
the same pattern of behavioral effects, namely, lack of
context-dependent adjustments in behavior. It is im-
portant to note, however, that the selection for action
hypothesis would predict that rACC patients should
exhibit performance decrements in all incongruent con-
ditions, irrespective of previous context, due to difficul-
ties in overcoming task-irrelevant, prepotent responses.
Our behavioral findings do not support such a predic-
tion. When compared to non-FC patients, the rACC
group did not display greater interference effects and/
or higher error rates in incongruent trials that were pre-
ceded by congruent ones. This seems to suggest that
the rostral ACC itself does not contribute to attentional
allocation. Accordingly, both behavioral and imaging
studies have shown that the lateral PFC is most critical
in processing response selection via top-down control of
other brain regions (Egner & Hirsh, 2005; Kerns et al.,
2004; Gehring & Knight, 2002; Miller & Cohen, 2001;
Vendrell et al., 1995). Instead, the current observations
fit more closely with a conflict monitoring view, accord-
ing to which ACC lesions would specifically impair the
patients’ ability to shift toward more focused or con-
servative behavior on the basis of an evaluation of their
own performance.
As previously discussed, and in contrast to the dorsal
ACC and its association with cognitive activity, the rostral
ACC and adjacent areas have been identified primarily
with emotional, motivational, and reward-related pro-
cessing (Bush et al., 2000; Devinsky et al., 1995). The
current data, however, challenge this view by demon-
strating that the rostral, ‘‘affective’’ ACC is also critically
involved in the ongoing adjustment of cognitive control.
This suggests that a functional separation of the rostral
and dorsal ACC along emotional versus cognitive lines
may be an oversimplification. Neuroimaging studies
showing activation within the dorsal ACC (the alleged
cognitive division) in response to emotional and painful
stimuli also argue against a clear-cut functional segrega-
tion within the ACC and the medial PFC (Rainville, 2002;
Ploghaus et al., 1999).
In the majority of previous imaging studies, the
response associated with cognitive conflict has been
localized to the dorsal ACC. However, the present
findings suggest that other regions in the frontal cortex
may be critical for determining when cognitive control is
needed (Swick & Turken, 2002; Gehring & Knight,
2000). Consistent with our results, recent research has
suggested that the conflict-dependent activation may
lie rostrally within the ACC (Milham & Banich, 2005).
Moreover, several authors emphasize the role of the
rostral ACC in processing errors and monitoring the
outcomes of actions (Nieuwenhuis, Slagter, Alting von
Geusau, Heslenfeld, & Holroyd, 2005; Braver, Barch,
Gray, Molfese, & Snyder, 2001; Kiehl et al., 2000). Also
consistent with this evidence, patients with damage to
the ventromedial PFC, including the rostral ACC, show
reduced error-related brain potentials while responding
in a flanker task, which suggests a close relation be-
tween performance monitoring, as reflected by the ERN,
and the rostral ACC (Stemmer, Segalowitz, Witzke, &
Scho¨nle, 2003). Thus, current data and previous physi-
ological and neuropsychological findings converge in in-
dicating that a role in conflict processing and cognitive
control extends to the rostral region of the ACC.
One may then ask what the specific role of the rostral
ACC in conflict processing and control might be. Multi-
ple evidence suggests that rostral ACC is responsive to
a variety of emotional signals (such as errors, negative
feedback, pain, and monetary loss) that predict negative
outcome, unless the level of cognitive control
is ap-
propriately intensified (Aston-Jones & Cohen, 2005). Al-
though a characterization of the rostral ACC as a purely
it seems
‘‘emotional’’ sector is clearly inappropriate,
plausible to suggest that this area is involved in medi-
ating the interaction between cognitive functioning and
affective, motivational, and autonomic processes neces-
sary for the guidance of adaptive behavior (Critchley,
2005; Simpson, Drevets, Snyder, Gusnard, & Raichle,
2001; Bechara et al, 2000; Drevets & Raichle, 1998).
Thus, when lapses in performance are detected, signals
from rostral ACC can activate both affective and cogni-
tive systems, perhaps via prominent cingulate connec-
tions with the amygdala and brainstem nuclei, such as
the locus coeruleus. Recent findings suggest that the
locus coeruleus, in addition to its role in arousal, can
have precise effects in regulating higher-level cognitive
functions, for instance, by enhancing task-specific con-
trol mechanisms in the PFC. This pathway, possibly com-
plemented by direct projections from the dorsal ACC to
the PFC, may ensure rapid change in behavior according
to emergent demands (Aston-Jones & Cohen, 2005).
The observed impairment of dynamic adjustment in
cognitive control
is highly consistent with the well-
known effects of medial prefrontal lesion, which typical-
ly causes patients to suffer from behavioral rigidity and
perseveration (Robbins, 2005; Fellows & Farah, 2003).
On this view, perseveration may result from the patients’
inability to modify their behavior in response to an eval-
uation of their own performance, due to a failure in
detecting internal stimuli (e.g., conflict) that predict un-
favorable outcomes.
The current findings also have direct implications for
our understanding of the nature several psychiatric dis-
orders. For instance, the disordered monitoring and
regulation of self-generated behavior in schizophrenia
bears some striking similarities with the cognitive defi-
cits evidenced in ventromedial PFC patients (Cohen,
& Servan-Schreiber, 1992). Consistent with this, neuro-
imaging studies have suggested that schizophrenic pa-
tients demonstrate relative underactivity in the rostral
ACC associated with impaired performance in conflict
di Pellegrino, Ciaramelli, and La`davas
283
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
2
-
2
p
7
d
5
f
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
t
f
.
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
tasks (Laurens et al., 2003; Carter, Mintun, Nichols, &
Cohen, 1997). Another relevant example comes from the
strong correlation between obsessive–compulsive disor-
ders (OCD) symptoms and the presence of hyperactivity
in the rostral ACC and medial PFC. Although these
findings were initially interpreted as a nonspecific result
of increased anxiety, more recent studies have shown
that a dysfunctional conflict monitoring system is a
critical aspect of OCD (Ursu, Stenger, Shear, Jones, &
Carter, 2003), which might explain why OCD patients
experience the need to monitor and correct their ac-
tions repeatedly (Pitman, 1987).
‘‘emotion’’ and dorsal
In summary, we report that the rostral ACC and the
adjacent ventromedial PFC play a crucial role in conflict
monitoring and on-line, plastic adjustment of perform-
ance. These findings have important implications for
the anatomy of cognitive control, as well as for our
understanding of medial frontal functions. Thus, our re-
sults impact on the simple scheme of the ACC divided
into rostral
‘‘cognition’’ parts.
The data also highlight the importance of lesion studies
for establishing cognitive neuroanatomy, given that
the rostral ACC lies outside the regions commonly
activated in functional imaging studies of control. Final-
ly, the present findings appear compatible with the
view wherein the rostral ACC is involved in the mecha-
nisms by which mental processes are integrated by
emotional signals. This proposal is, of course, specula-
tive, and further research will be needed to directly eval-
uate its merit.
Acknowledgments
We thank Eric van Diessen for his help in collecting part of the
data, and Dr. Oliver Turnbull for his comments on a draft of
the manuscript.
Reprint requests should be sent to Giuseppe di Pellegrino,
Dipartimento di Psicologia, Universita` di Bologna, Viale Berti
Pichat 5 - 40127 Bologna, Italy, or via e-mail: g.dipellegrino@
unibo.it.
REFERENCES
Allman, J. M., Hakeem, A., Erwin, J. M., Nimchinsky, E.,
& Hof, P. (2001). The anterior cingulate cortex. The
evolution of an interface between emotion and cognition.
Annals of the New York Academy of Science, 935,
107–117.
Aston-Jones, G., & Cohen, J. D. (2005). An integrative theory
of locus coeruleus–norepinephrine function: Adaptive
gain and optimal performance. Annual Review of
Neuroscience, 28, 403–450.
Baddeley, A. (1986). Working memory. Oxford: Psychology
Press.
Baird, A., Dewar, B. K., Critchley, H., Gilbert, S. J., Dolan, R. J.,
& Cipolotti, L. (2006). Cognitive functioning after medial
frontal lobe damage including the anterior cingulate cortex:
A preliminary investigation. Brain and Cognition, 60,
166–175.
Bechara, A., Tranel, D., & Damasio, H. (2000). Characterization
of the decision-making deficit of patients with
ventromedial prefrontal cortex lesions. Brain, 123,
2189–2202.
Botvinick, M. M., Braver, T. S., Barch, D. M., Carter, C. S.,
& Cohen, J. D. (2001). Conflict monitoring and cognitive
control. Psychological Review, 108, 624–652.
Botvinick, M. M., Cohen, J. D., & Carter, C. S. (2004).
Conflict monitoring and anterior cingulate cortex:
An update. Trends in Cognitive Sciences, 8, 539–546.
Botvinick, M. M., Nystrom, L. E., Fissell, K., Carter, C. S.,
& Cohen, J. D. (1999). Conflict monitoring versus
selection-for-action in anterior cingulate. Nature, 402,
179–181.
Braver, T. S., Barch, D. M., Gray, J. R., Molfese, D. L., &
Snyder, A. (2001). Anterior cingulate cortex and response
conflict: Effects of frequency, inhibition and errors. Cerebral
Cortex, 11, 825–836.
Burgess, P. W., Veitch, E., de Lacy Costello, A., & Shallice, T.
(2000). The cognitive and neuroanatomical correlates of
multitasking. Neuropsychologia, 38, 848–863.
Bush, G., Luu, P., & Posner, M. I. (2000). Cognitive and
emotional influences in anterior cingulate cortex. Trends
in Cognitive Sciences, 4, 215–222.
Bush, G., Whalen, P. J., Rosen, B. R., Jenike, M. A.,
McInerney, S. C., & Rauch, S. L. (1998). The counting
Stroop: An interference task specialized for functional
neuroimaging-validation study with functional MRI.
Human Brain Mapping, 6, 270–282.
Carter, C. S., Mintun, M., Nichols, T., & Cohen, J. D.
(1997). Anterior cingulate gyrus dysfunction and selective
attention deficits in schizophrenia: [15O]H2O PET study
during single-trial Stroop task performance. American
Journal of Psychiatry, 154, 1670–1675.
Cohen, J. D., & Servan-Schreiber, D. (1992). Context, cortex,
and dopamine: A connectionist approach to behavior
and biology in schizophrenia. Psychological Review, 99,
45–77.
Critchley, H. (2005). Neural mechanisms of autonomic,
affective, and cognitive integration. Journal of Comparative
Neurology, 493, 154–166.
Critchley, H., Mathias, C. J., Josephs, O., O’Doherty, J.,
Zanini, S., Dewar, B. K., et al. (2003). Human cingulate
cortex and autonomic control: Converging neuroimaging
and clinical evidence. Brain, 126, 2139–2152.
Damasio, H., & Damasio, A. (1989). Lesions analysis in
neuropsychology. New York: Oxford University Press.
Davis, K. D., Taylor, K. S., Hutchison, W. D., Dostrovsky,
J. O., McAndrews, M. P., Richter, E. O., et al. (2005).
Human anterior cingulate cortex neurons encode cognitive
and emotional demands. Journal of Neuroscience, 25,
8402–8406.
Desimone, R., & Duncan, J. (1995). Neural mechanisms of
selective visual attention. Annual Review of Neuroscience,
18, 193–222.
Devinsky, O., Morrell M. J., & Vogt, B. A. (1995). Contributions
of anterior cingulate cortex to behaviour. Brain, 118,
279–306.
Dreher, J. C., & Grafman, J. (2003). Dissociating the roles
of the rostral anterior cingulate and the lateral prefrontal
cortices in performing two tasks simultaneously or
successively. Cerebral Cortex, 13, 329–339.
Drevets, W. C., Price, J. L., Simpson, J. R., Jr., Todd, R. D.,
Reich, T., Vannier, M., et al. (1997). Subgenual prefrontal
cortex abnormalities in mood disorders. Nature, 386,
824–827.
Drevets, W. C., & Raichle, M. E. (1998). Reciprocal suppression
of regional cerebral blood flow during emotional versus
284
Journal of Cognitive Neuroscience
Volume 19, Number 2
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
2
-
2
p
7
d
5
f
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
t
.
.
.
.
.
o
n
1
8
M
a
y
2
0
2
1
higher cognitive processes: Implications for interactions
between emotion and cognition. Cognition and Emotion,
12, 353–385.
Egner, T., & Hirsh, J. (2005). Cognitive control mechanisms
resolve conflict through cortical amplification of
task-relevant information. Nature Neuroscience, 12,
1784–1790.
Eisenberger, N. I., & Lieberman, M. D. (2004). Why
rejection hurts: A common neural alarm system for
physical and social pain. Trends in Cognitive Sciences, 8,
294–300.
Eslinger, P. J., & Damasio, A. R. (1985). Severe disturbance
of higher cognition after bilateral frontal lobe ablation:
Patient EVR. Neurology, 35, 1731–1741.
Falkenstein, M., Hohnsbein, J., & Hoormann, J. (1991).
Effects of crossmodal divided attention on late ERP
components. II. Error processing in choice reaction tasks.
Electroencephalography and Clinical Neurophysiology,
78, 447–455.
Fellows, L. K., & Farah, M. J. (2003). Ventromedial frontal
cortex mediates affective shifting in humans: Evidence
from a reversal learning paradigm. Brain, 126, 1830–1836.
Fellows, L. K., & Farah, M. J. (2005). Is anterior cingulate cortex
necessary for cognitive control? Brain, 128, 788–796.
Folstein, M. F., Robins, L. N., & Helzer, J. E. (1983). The
Mini-Mental State Examination. Archives of General
Psychiatry, 40, 812.
Gehring, W. J., & Knight, R. T. (2000). Prefrontal–cingulate
interactions in action monitoring. Nature Neuroscience,
3, 516–520.
Gehring, W. J., & Knight, R. T. (2002). Lateral prefrontal
damage affects processing selection but not attention
switching. Brain Research, Cognitive Brain Research,
13, 267–279.
Gratton, G., Coles, M. G., & Donchin, E. (1992). Optimizing
the use of information: Strategic control of activation of
responses. Journal of Experimental Psychology: General,
121, 480–506.
Hommel, B., Proctor, R. W., & Vu, K. P. (2004). A
feature-integration account of sequential effects in the
Simon task. Psychological Research, 68, 1–17.
Kerns, J. G., Cohen, J. D., MacDonald, A. W., III, Cho, R. Y.,
Stenger, V. A., & Carter, C. S. (2004). Anterior cingulate
conflict monitoring and adjustments in control. Science,
303, 1023–1026.
Kiehl, K. A., Smith, A. M., Hare, R. D., & Liddle, P. F. (2000).
An event-related potential investigation of response
inhibition in schizophrenia and psychopathy. Biological
Psychiatry, 48, 210–221.
Kornblum, S., Hasbroucq, T., & Osman, A. (1990).
Dimensional overlap: Cognitive basis for stimulus–response
compatibility—A model and taxonomy. Psychological
Review, 97, 253–270.
errors and feedback in the process of action regulation.
Psychological Science, 14, 47–53.
Mayr, U., Awh, E., & Laurey, P. (2003). Conflict adaptation
effects in the absence of executive control. Nature
Neuroscience, 6, 450–452.
Menon, V., Adleman, N. E., White, C. D., Glover, G. H., &
Reiss, A. L. (2001). Error-related brain activation during
a Go/NoGo response inhibition task. Human Brain
Mapping, 12, 131–143.
Milham, M. P., & Banich, M. T. (2005). Anterior cingulate
cortex: An fMRI analysis of conflict specificity and
functional differentiation. Human Brain Mapping, 25,
328–335.
Miller, E. K., & Cohen, J. D. (2001). An integrative theory
of prefrontal cortex function. Annual Review of
Neuroscience, 24, 167–202.
Nieuwenhuis, S., Slagter, H. A., Alting von Geusau, N. J.,
Heslenfeld, D. J., & Holroyd, C. B. (2005). Knowing
good from bad: Differential activation of human cortical
areas by positive and negative outcomes. European
Journal of Neuroscience, 21, 3161–3168.
Norman, D., & Shallice, T. (1986). Attention to action:
Willed and automatic control of behavior. In R. Davidson,
G. Schwartz, & D. Shapiro (Eds.), Consciousness and
self regulation: Advances in research and theory
(Vol. 4, pp. 1–18). New York: Plenum Press.
Notebaert, W., Soetens, E., & Melis, A. (2001). Sequential
analysis of a Simon task: Evidence for an attention-shift
account. Psychological Research, 65, 174–180.
O¨ ngu¨r, D., & Price, J. L. (2000). The organization of networks
within the orbital and medial prefrontal cortex of rats,
monkeys and humans. Cerebral Cortex, 10, 206–219.
Paus, T. (2001). Primate anterior cingulate cortex: Where
motor control, drive and cognition interface. Nature
Reviews Neuroscience, 2, 417–424.
Phillips, M. L., Drevets, W. C., Rauch, S. L., & Lane, R.
(2003a). Neurobiology of emotion perception I: The
neural basis of normal emotion perception. Biological
Psychiatry, 54, 504–514.
Phillips, M. L., Drevets, W. C., Rauch, S. L., & Lane, R. (2003b).
Neurobiology of emotion perception: II. Implications
for major psychiatric disorders. Biological Psychiatry,
54, 515–528.
Picard, N., & Strick, P. L. (1996). Motor areas of the medial
wall: A review of their location and functional activation.
Cerebral Cortex, 6, 342–353.
Pitman, R. K. (1987). A cybernetic model of obsessive–
compulsive psychopathology. Comprehensive Psychiatry,
28, 334–343.
Ploghaus, A., Tracey, I., Gati, J. S., Clare, S., Menon, R. S.,
Matthews, P. M., et al. (1999). Dissociating pain from
its anticipation in the human brain. Science, 284,
1979–1981.
Laming, D. R. J. (1968). Information theory of choice-reaction
Polli, F. E., Barton, J. J., Cain, M. S., Thakkar, K. N., Rauch,
times. London: Academic Press.
Laurens, K. R., Ngan, E. T., Bates, A. T., Kiehl, K. A., & Liddle,
P. F. (2003). Rostral anterior cingulate cortex dysfunction
during error processing in schizophrenia. Brain, 126,
610–622.
Luppino, G., Rozzi, S., Calzavara, R., & Matelli, M. (2003).
Prefrontal and agranular cingulate projections to the
dorsal premotor areas F2 and F7 in the macaque monkey.
European Journal of Neuroscience, 17, 559–578.
Luu, P., Flaisch, T., & Tucker, D. M. (2000). Medial frontal
cortex in action monitoring. Journal of Neuroscience,
20, 464–469.
Luu, P., Tucker, D. M., Derryberry, D., Reed, M., &
Poulsen, C. (2003). Electrophysiological responses to
S. L., & Manoach, D. S. (2005). Rostral and dorsal anterior
cingulate cortex make dissociable contributions during
antisaccade error commission. Proceedings of the National
Academy of Sciences, U.S.A., 102, 15700–15705.
Posner, M. I., & Di Girolamo, G. J. (1998). Executive attention:
Conflict, target detection, and cognitive control. In R.
Parasuraman (Ed.), The attentive brain (pp. 401–423).
Cambridge: MIT Press.
Posner, M. I., & Petersen, S. E. (1990). The attention system
of the human brain. Annual Review of Neuroscience, 13,
25–42.
Rabbitt, P. M. A. (1966). Errors and error correction in
choice-response tasks. Journal of Experimental Psychology,
71, 264–272.
di Pellegrino, Ciaramelli, and La`davas
285
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
2
-
2
p
7
d
5
f
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
.
.
t
.
.
f
.
o
n
1
8
M
a
y
2
0
2
1
Rainville, P. (2002). Brain mechanisms of pain affect and
pain modulation. Current Opinion in Neurobiology, 12,
195–204.
Ridderinkhof, K. R., Ullsperger, M., Crone, E. A., &
Nieuwenhuis, S. (2004). The role of the medial frontal
cortex in cognitive control. Science, 306, 443–447.
Robbins, T. W. (2005). Chemistry of the mind: Neurochemical
modulation of prefrontal cortical function. Journal of
Comparative Neurology, 493, 140–146.
Rushworth, M. F., Kennerley, S. W., & Walton, M. E. (2005).
Cognitive neuroscience: Resolving conflict in and over
the medial frontal cortex. Current Biology, 15, R54–R56.
Rushworth, M. F., Walton, M. E., Kennerley, S. W., &
Bannerman, D. M. (2004). Action sets and decisions in
the medial frontal cortex. Trends in Cognitive Sciences,
8, 410–417.
Rylander, G. (1947). Personality analysis before and after
frontal lobotomy. In J. F. Fulton, C. D. Aring, & B. S.
Wortis (Eds.), Research publications association for
research in nervous and mental disease: The frontal
lobe (pp. 691–705). Baltimore: Williams and Wilkins.
Schneider, W., Eschman, A., & Zuccolotto, A. (2002). E-prime
reference guide. Pittsburgh: Psychology Software Tool Inc.
Shiffrin, R. M., & Schneider, W. (1977). Controlled and
automatic human information processing: 1. Detection,
search, and attention. Psychological Review, 84, 127–190.
Simon, J. R. (1969). Reactions towards the source of
stimulation. Journal of Experimental Psychology, 81,
174–176.
Simpson, J. R., Jr., Snyder, A. Z., Gusnard, D. A., & Raichle,
M. E. (2001). Emotion-induced changes in human medial
prefrontal cortex: I. During cognitive task performance.
Proceedings of the National Academy of Sciences, U.S.A.,
98, 683–687.
Stemmer, B., Segalowitz, S. J., Witzke, W., & Scho¨nle, W.
(2003). Error detection in patients with lesions to the medial
prefrontal cortex: An ERP study. Neuropsychologia, 42,
118–130.
Stu¨rmer, B., Leuthold, H., Soetens, E., Schro¨ter, H., & Sommer,
W. (2002). Control over location-based response activation
in the Simon task: Behavioral and electrophysiological
evidence. Journal of Experimental Psychology: Human
Perception and Performance, 28, 1345–1363.
Stuss, D. T., Floden, T., Alexander, M. P., Levine, B., &
Katz, D. (2001). Stroop performance in focal lesion
patients: Dissociation of processes and frontal lobe
location. Neuropsychologia, 39, 771–786.
Swick, D., & Jovanovic, J. (2002). Anterior cingulate cortex
and the Stroop task: Neuropsychological evidence for
topographic specificity. Neuropsychologia, 40, 1240–1253.
Swick, D., & Turken, A. U. (2002). Dissociation between
conflict detection and error monitoring in the human
anterior cingulate cortex. Proceedings of the National
Academy of Sciences, U.S.A., 99, 16345–16349.
Teuber, H. L. (1964). The riddle of the frontal lobe function
in man. In J. M. Warren & K. Akert (Eds.), The frontal
granular cortex and behavior (pp. 410–414). New York:
McGraw Hill.
Ullsperger, M., Bylsma, L. M., & Botvinick, M. M. (2005). The
conflict adaptation effect: It’s not just priming. Cognitive,
Affective & Behavioral Neuroscience, 5, 467–472.
Umilta`, C. (1988). The control operations of consciousness.
In A. J. Marcel & E. Bisiach (Eds.), Consciousness in
contemporary science (pp. 334–356). Oxford: Oxford
University Press.
Ursu, S., Stenger, V. A., Shear, M. K., Jones, M. R., &
Carter, C. S. (2003). Overactive action monitoring in
obsessive–compulsive disorder: Evidence from functional
magnetic resonance imaging. Psychological Science, 14,
347–353.
Vendrell, P., Junque, C., Pujol, J., Jurado, M. A., Molet, J.,
& Grafman, J. (1995). The role of prefrontal regions in
the Stroop task. Neuropsychologia, 33, 341–352.
Wager, T. D., Jonides, J., Smith, E. E., & Nichols, T. E.
(2005). Toward a taxonomy of attention shifting:
Individual differences in fMRI during multiple shift types.
Cognitive, Affective & Behavioural Neuroscience, 5,
127–143.
Whalen, P. J., Bush, G., McNally, R. J., Wilhelm, S., McInerney,
S. C., Jenike, M. A., et al. (1998). The emotional counting
Stroop paradigm: A functional magnetic resonance
imaging probe of the anterior cingulate affective division.
Biological Psychiatry, 44, 1219–1228.
Wuhr, P., & Ansorge, U. (2005). Exploring trial-by-trial
modulations of the Simon effect. Quarterly Journal
of Experimental Psychology, 58, 705–731.
Yeung, N., Botvinick, M. M., & Cohen, J. D. (2004). The
neural basis of error detection: Conflict monitoring and
error-related negativity. Psychological Review, 111,
931–959.
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
D
h
o
t
w
t
p
n
:
o
/
a
/
d
m
e
i
d
t
f
r
p
o
r
m
c
.
h
s
i
p
l
v
d
e
i
r
r
e
c
c
h
t
.
m
a
i
r
e
.
d
c
u
o
m
o
/
c
j
n
o
a
c
r
t
n
i
c
/
e
a
-
r
p
t
d
i
c
1
l
9
e
2
-
2
p
7
d
5
f
/
1
1
9
9
3
6
/
2
2
0
/
7
2
7
o
5
c
/
n
1
2
7
0
5
0
6
7
5
1
8
9
2
/
2
j
2
o
7
c
5
n
p
.
d
2
0
b
0
y
7
g
.
u
1
e
9
s
.
t
2
o
.
n
2
0
7
8
5
S
.
p
e
p
d
f
e
m
b
b
y
e
r
g
2
u
0
e
2
s
3
t
/
j
f
.
.
.
.
.
t
o
n
1
8
M
a
y
2
0
2
1
286
Journal of Cognitive Neuroscience
Volume 19, Number 2