The Phenomenal Contents and Neural Correlates
of Spontaneous Thoughts across Wakefulness,
NREM Sleep, and REM Sleep
Lampros Perogamvros1,2,3, Benjamin Baird1, Mitja Seibold4, Brady Riedner1,
Melanie Boly1, and Giulio Tononi1
D
o
w
n
je
o
un
d
e
d
F
r
o
m
Abstrait
■ Thoughts occur during wake as well as during dreaming
dormir. Using experience sampling combined with high-density
EEG, we investigated the phenomenal qualities and neural cor-
relates of spontaneously occurring thoughts across wakeful-
ness, non-rapid eye movement (NREM) dormir, and REM sleep.
Across all states, thoughts were associated with activation of a
region of the midcingulate cortex. Thoughts during wakeful-
ness additionally involved a medial prefrontal region, lequel
was associated with metacognitive thoughts during wake. Phe-
nomenologically, waking thoughts had more metacognitive
content than thoughts during both NREM and REM sleep,
whereas thoughts during REM sleep had a more social content.
Ensemble, these results point to a core neural substrate for
pensées, regardless of behavioral state, within the midcingu-
late cortex, and suggest that medial prefrontal regions may con-
tribute to metacognitive content in waking thoughts. ■
INTRODUCTION
Thoughts are a central feature of waking life, si
they are directed toward accomplishing a particular task
or whether they occur spontaneously. Experience sam-
pling experiments have found that spontaneous thoughts
during wakefulness occur often, with a frequency of oc-
currence of as high as 30–50% (Killingsworth & Gilbert,
2010; Kane et al., 2007). Although the underlying neural
mechanisms of thought are not well understood, neuro-
imaging studies using fMRI and PET have found neural
correlates of spontaneous thought in a network of areas
including the default mode network (DMN) et le
executive control network (reviewed in Fox, Spreng,
Ellamil, Andrews-Hanna, & Christoff, 2015).
Thoughts occur not only in wakefulness but also dur-
ing sleep, often in combination with perceptual experi-
ences, in the form of dreams. Although there may be
differences between thoughts that occur during wake-
fulness and dreaming, including reduced metacognitive
insight (Nir & Tononi, 2010), several studies have sug-
gested that the overall phenomenology of consciousness
is remarkably similar across sleep and wake (Kahan &
LaBerge, 2011; Kahan, LaBerge, Levitan, & Zimbardo,
1997). Par exemple, both states draw on proximal and
distal memory sources (Fox, Nijeboer, Solomonova,
1University of Wisconsin-Madison, 2Geneva University Hospitals,
3University of Geneva, 4University of Amsterdam
© 2017 Massachusetts Institute of Technology
Domhoff, & Christoff, 2013; Nielsen & Stenstrom, 2005)
and frequently involve thoughts regarding daily personal
concerns (Andrews-Hanna, Reidler, Huang, & Buckner,
2010; Smallwood, Fitzgerald, Miles, & Phillips, 2009;
Cartwright, Agargun, Kirkby, & Friedman, 2006; Nielsen
& Stenstrom, 2005). The continuity of cognition across
wake and sleep has led some researchers to suggest that
there may be a common neurophysiological substrate of
thought across waking and dreaming (Domhoff & Fox,
2015; Fox et al., 2013; Wamsley, 2013; Llinas & Pare,
1991).
Physiologically, an important distinguishing feature of
sleep and wakefulness is that during sleep individuals
are largely disconnected from the environment, and as
a consequence, external sensory stimuli have little influ-
ence on the content of experience. Cependant, it is inter-
esting to note that a similar reduction in cortical
responses to the external environment, as measured by
an attenuation of sensory-evoked responses or reduced
phase-locking to perceptual stimuli, has also been ob-
served during mind-wandering (Baird, Smallwood, Lutz,
& Schooler, 2014; Braboszcz & Delorme, 2011; Kam
et coll., 2011; Smallwood, Beach, Schooler, & Handy,
2008). Another distinguishing feature of sleep and wake
thought is that, outside of rare instances of lucidity (La
Berge, Nagel, Dement, & Zarcone, 1981), dreaming par-
ticipants have little or no deliberate cognitive control
over the content of thought, which is not always the case
during wake, even during mind-wandering (Christoff,
Irving, Fox, Spreng, & Andrews-Hanna, 2016).
Journal des neurosciences cognitives 29:10, pp. 1766–1777
est ce que je:10.1162/jocn_a_01155
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
un
/
_
j
0
o
1
c
1
n
5
5
_
un
p
_
d
0
1
b
1
oui
5
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
/
.
t
F
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
To the best of our knowledge, no study to date has si-
multaneously investigated the phenomenal content and
neural correlates of thought across wakefulness and sleep
states. Although thoughts across wakefulness and sleep
exhibit some continuity of content, they occur within
brain states having very different neurophysiological
and neuromodulatory profiles ( Jones, 2005). It is there-
fore an open question whether there is a core neural sub-
strate to thought across wakefulness and sleep. Dans ce
étude, we aimed at examining the neural substrate and
phenomenology of thought across quiet resting wakeful-
ness, non-rapid eye movements (NREM) dormir, and REM
dormir. Throughout all three states, brain activity was con-
tinuously recorded using high-density EEG (hd-EEG),
and participants were intermittently prompted to report
the contents of consciousness using thought sampling
(wake; Smallwood & Schooler, 2006; Klinger & Cox,
1987) or serial awakenings (dormir; Siclari, Larocque, Postle,
& Tononi, 2013).
MÉTHODES
Study Participants
Sixty-nine healthy individuals (23 men, age = 43.14 ±
12.94 années, 25–64 [mean ± SD, range]) participated in
the experiment. All participants had no history of neuro-
logical disorder. Signed informed consent was obtained
from all participants before the experiment, and ethical
approval for the study was obtained from the University
of Wisconsin-Madison institutional review board.
Procedures
Spontaneous Thought Task during Wakefulness
Participants were asked to rest their eyes on a fixation
cross in the center of a computer screen for approximate-
ly 30 min. At pseudorandom intervals (environ
once per minute), participants were prompted with a
sound and were instructed to report the last thing going
through their mind (any images, pensées, feelings, emo-
tion) just before the sound. Following this open-ended
report, they were asked to rate the content of their expe-
rience on a thought scale (thinking or reasoning) ranging
depuis 0 (no thoughts) à 5 (maximally thought-like).
Participants were also asked to rate on a scale from 0 (ab-
sence) à 5 (maximum) the degree of the cognitive ef-
fort in their experience (par exemple., trying to think through a
problem or accomplish a particular task). Examples of
what we operationally defined as “thoughts” were given
to participants before testing, which included, for exam-
ple, invariant concepts, ideas, memories, and decisions.
Serial Awakenings during Sleep
Experience sampling during sleep was accomplished
using the “serial awakening” method, which is described
in detail elsewhere (Siclari et al., 2013). In brief, partici-
pants were awakened throughout the night while sleep-
ing in the sleep laboratory and were asked to report
si, just before the awakening, they were dreaming
of anything. If participants reported having a dream expe-
rience, they were asked to describe its most recent con-
tent (“the last thing going through your mind before the
alarm sound”) and then underwent a structured inter-
view via intercom. Spécifiquement, participants were again
asked to rate their experience on a “thought” scale rang-
ing from 0 (no thoughts) à 5 (maximally thought-like),
as well as to rate their experience on several other di-
mensions, including their level of voluntary control over
the content of the experience, the richness and complex-
ity of the experience, the cognitive effort put in the expe-
rience, the duration and capacity to recall the experience.
Awakenings were performed at intervals of at least
20 min, in N2 or REM sleep, using a computerized alarm
sound. Participants were required to have been asleep for
a minimum of 10 min and must have been in a stable
sleep stage for a minimum of 5 min for an awakening
to occur.
Content Analysis
To phenomenologically compare thoughts in wakeful-
ness with those in NREM and REM sleep, reports that
were scored by participants between 3 et 5 on the
thought scale (see Procedures) were considered high-
thought reports. High-thought reports were additionally
classified by two independent raters in eight categories on
a scale from 0 (absence of the category) à 5 (maximum
in this category): (1) metacognition, (2) social focus,
(3) positive emotions, (4) negative emotions, (5) past
focus, (6) present focus, (7) future focus, et (8) bizarre-
ness. The definitions of these categories are given in
Table A1. In total, 264 dream reports (157 in N2 and
107 in REM sleep) et 869 waking reports were scored.
Of those, 141 dream reports (71 in N2 and 70 in REM
dormir) et 652 waking reports were high-thought trials.
The average interrater reliability was 79% for waking data
et 82% for dream data, representing an acceptable inter-
rater reliability for both states (Hallgren, 2012; Schredl,
2010b). Because of the non-normal distribution of the data
(Shapiro–Wilk normality test p < .05; Table A2), Wilcoxon
signed-rank tests were performed between averaged
scores for each participant on each of the categories for
high-thought trials in wakefulness versus N2 sleep, wake-
fulness versus REM sleep, and N2 versus REM sleep.
Wilcoxon signed-rank tests were also performed for the
“cognitive effort” category, which was another dimension
in which participants self-evaluated in both wakefulness
and sleep. Finally, scores in certain categories in which par-
ticipants self-evaluated during sleep (voluntary control over
the content of the experience, richness and complexity of
the experience, recall of the experience, duration of the
experience) in the N2 and REM reports were averaged
Perogamvros et al.
1767
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
/
f
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
for each of the 69 participants, and Wilcoxon signed-rank
tests were performed between N2 and REM sleep scores
for each participant. Multiple comparisons were performed
for the 31 independent tests using the Holm–Bonferroni
method (Holm, 1979), and p values were adjusted at a
p < .05 significance level. To study the relation between
the cognitive effort and thought variables across states, a
linear mixed model was used to account for repeated mea-
sures with varied numbers of repeated observations within
participants. Mixed model construction and mixed model
boostrapping were performed with the lme4 package
(Bates, Maechler, Bolker, & Walker, 2015) in the R environ-
ment (R Development Core Team, 2006).
Participants Selected for the hd-EEG Analysis
From the initial group of 69 participants, 13 individuals
were selected for the EEG analysis of wake and N2 stages
(six men, age = 40.6 ± 11.9 years, 25–59 [mean ± SD,
range]). We selected participants from this data set who
had trials with both low level of thoughts (0–2) and trials
with high level of thoughts (3–5) in both wakefulness and
N2 sleep. Ten different participants were included in the
EEG analysis of REM sleep (6 men, age = 37.7 ±
11.5 years, 27–64 [mean ± SD, range]) based on the
same criterion.
EEG Recordings
Recordings were made at the University of Wisconsin-
Madison Center for Sleep Medicine and Sleep Research
( WisconsinSleep) using a 256-channel hd-EEG system
(Electrical Geodesics, Inc., Eugene, OR) combined with
Alice Sleepware (Philips Respironics, Murrysville, PA).
Additional polysomnography channels were used to re-
cord and monitor eye movements and submental electro-
myography during sleep. Sleep scoring was performed
over 30-sec epochs according to standard criteria (Iber,
Ancoli-Israel, Chesson, & Quan, 2007).
EEG Preprocessing
The EEG signal was sampled at 500 Hz and band-pass fil-
tered offline between 1 and 50 Hz. The 1-Hz high-pass
threshold was used due to sweating artifacts in some par-
ticipants, which caused intermittent high-amplitude
(>300 μV ) slow frequency oscillatory activity around
0.3 Hz. Noisy channels and epochs containing artifactual
activity were visually identified and removed. Ocular,
muscular, and electrocardiograph artifacts were removed
with independent component analysis using EEGLAB
routines (Delorme & Makeig, 2004). Only independent
component analysis components with specific activity
patterns and component maps characteristic of artifactual
activity were removed (Jung et al., 2000). Previously re-
moved noisy channels were interpolated using spherical
splines (EEGLAB). Enfin, EEG data were referenced to
the average of all electrodes.
EEG Signal Analysis
Source Localization
The cleaned, filtered, and average-referenced EEG signal
corresponding to the 20 sec before the alarm sound in
both wakefulness and sleep was extracted and analysed
at the source level. Source modeling was performed
using the GeoSource software (Electrical Geodesics,
Inc.). A four-shell head model based on the Montreal
Neurological Institute atlas and a standard coregistered
set of electrode positions were used to construct the for-
ward model. The source space was restricted to 2447 di-
poles in three dimensions that were distributed over 7 ×
7 × 7 mm cortical voxels. The inverse matrix was com-
puted using the standardized low-resolution brain elec-
tromagnetic tomography constraint. A Tikhonov
−1) was applied to
regularization procedure (λ = 10
account for the variability in the signal-to-noise ratio
(Pascual-Marqui, 2002). We computed spectral power
density using the Welch’s modified periodogram method
(implemented with the pwelch function in MATLAB; Le
MathWorks, Inc., Natick, MA) in 2-sec Hamming windows
(8 segments, 50% overlap) to decompose the source sig-
nals into frequency bands of interest before taking the
norm across dimension to produce a single power value
for each dipole.
Statistical Analysis
Statistical analysis was carried out in MATLAB. To com-
pare brain activity between low-thought and high-
thought trials, source-space power was averaged within
standard frequency bands (Delta: 1–4 Hz, Theta: 4–
8 Hz, Alpha: 8–12 Hz, Sigma: 12–18 Hz, Beta: 18–
25 Hz, Gamma: 25–50 Hz). We then averaged the power
values within low-thought and high-thought trials for
each participant and for each frequency band and stage
(wakefulness, N2 sleep, and REM sleep) separately.
Group level analyses used paired two-sample t tests
(two-tailed) between the low-thought and high-thought
conditions in wake, NREM sleep, and REM sleep, tests
that were performed separately for each frequency band
and thresholded at corrected p < .05 using nonparamet-
ric threshold-free cluster enhancement (TFCE; weighing
parameters E = 0.5 and H = 2; Mensen & Khatami, 2013;
Smith & Nichols, 2009).
RESULTS
Phenomenological Results
We examined potential differences and similarities in the
phenomenology of thoughts across wake and sleep
states. High-thought trials were found to be more
1768
Journal of Cognitive Neuroscience
Volume 29, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
.
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
frequent in wake (75%), compared with N2 sleep (45%)
and REM sleep (65%). Mean values across the phenome-
nological categories are shown in Table A3. Higher scores
on the thought scale were significantly associated with re-
ported higher cognitive effort across all three states (β =
0.36, p < .0001) and in each state separately (wake: β =
0.37, p < .0001; N2: β = 0.42, p < .0001; REM: β = 0.41,
p < .0001).
Wakefulness versus N2 Sleep
Thoughts in wakefulness were rated significantly higher
than thoughts occurring in N2 sleep in metacognition
( p = .01, Z = 3.51; Table 1). No other significant differ-
ences were found in the comparisons at a Holm–Bonferroni
corrected significance level.
Wakefulness versus REM Sleep
Thoughts in wakefulness were significantly higher than
thoughts occurring in REM sleep in metacognition
( p = .008, Z = 3.58). REM thoughts were significantly
higher in social focus ( p = .001, Z = −4.06) and in pos-
itive emotions ( p = .002, Z = −3.92; Table 1). Thoughts
in REM sleep were also rated more “effortful” than
thoughts in wakefulness ( p = .02, Z = −3.29). No other
significant differences were found in the comparisons at a
Holm–Bonferroni corrected significance level.
N2 Sleep versus REM Sleep
REM thoughts contained significantly higher cognitive ef-
fort ( p = .04, Z = −3.05) than N2 thoughts. REM
thoughts also lasted longer ( p = .001, Z = −4.25), were
more easily recalled ( p = .0001, Z = −4.48), and were
more rich and complex ( p = .0001, Z = −4.84), than N2
thoughts. No other significant differences were found in
the comparisons at a Holm–Bonferroni corrected signifi-
cance level.
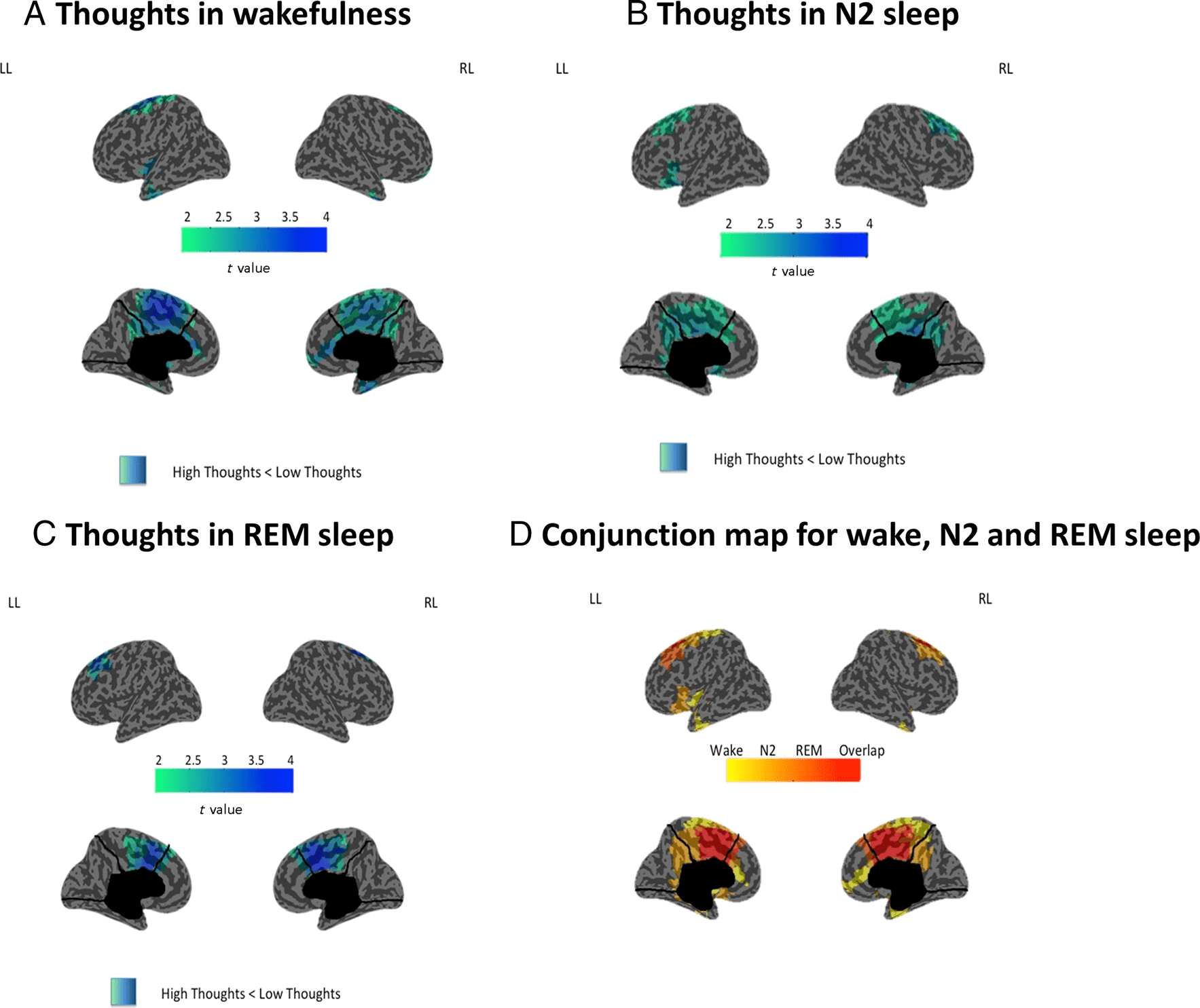
Topographical Results
Across all three states (wakefulness, REM sleep, and
NREM sleep), we found that high-thought reports com-
pared with low-thought reports had decreased delta
power (1–4 Hz), which was maximal over the midcingu-
late cortex (Figure 1). No significant changes were found
for other frequency bands. During wake, topographical
differences extended to the posterior cingulate cortex,
premotor cortex, and the medial prefrontal cortex (mPFC;
Figure 1A). In N2 sleep, decreased delta power in high-
thought reports compared with low-thought reports was
again maximal over the midcingulate cortex and again in-
cluded posterior cingulate cortex and premotor cortex but
did not extend to frontal cortex (Figure 1B). In REM sleep,
decreased delta power during high-thought reports was
again maximal over the midcingulate cortex but had a
more restricted topography, which did not include either
Table 1. Phenomenological Analysis of Thoughts across Wakefulness and Sleep in All Participants
Wake vs. N2
Wake vs. REM
N2 vs. REM
Metacognition
Social focus
Positive emotions
Negative emotions
Past focus
Present focus
Future focus
Bizarreness
Cognitive effort
Duration
Richness/complexity
Voluntary control
Recall of experience
puncor
.0004
.03
.74
.90
.88
.76
.60
.94
.80
n/a
n/a
n/a
n/a
pcor
.01*
.63
1
1
1
1
1
1
1
n/a
n/a
n/a
n/a
puncor
.0003
.00004
.00008
1
.29
.08
.26
.27
.001
n/a
n/a
n/a
n/a
pcor
.008*
.001*
.002*
1
1
1
1
1
.02*
n/a
n/a
n/a
n/a
puncor
pcor
.66
.19
.003
.04
.26
.38
.39
.32
.002
.00002
.00001
.91
1
1
.06
.8
1
1
1
1
.04*
.001*
.0001*
1
.000007
.0001*
Wilcoxon signed-rank tests between high-thought trials in wakefulness, N2 sleep, and REM sleep.
*Significant difference after correction for multiple comparisons for 31 independent tests using the Holm–Bonferroni method and adjustment of
p values at a .05 significance level.
Perogamvros et al.
1769
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Figure 1. The neural correlates of thoughts across wakefulness and sleep. Inflated cortical maps illustrating the cortical distribution of t values (two-
tailed, paired t tests, p < .05, TFCE-corrected) for the contrast between trials with high thought and low thought at the source level for delta
power (1–4 Hz) for (A) wakefulness (n = 13), (B) N2 sleep (n = 13), (C) REM sleep (n = 10). (D) Conjunction map showing the differences (yellow
to orange) and overlap (red) of the topographical maps contrasting high-thought and low-thought trials across wakefulness, N2 sleep, REM sleep.
posterior cingulate cortex or frontal cortex (Figure 1C). A
conjunction map showing the differences and overlap of
the topographical maps contrasting high-thought and low-
thought trials across the three states is shown in
Figure 1D. The midcingulate cortex is a region of overlap
among all three states.
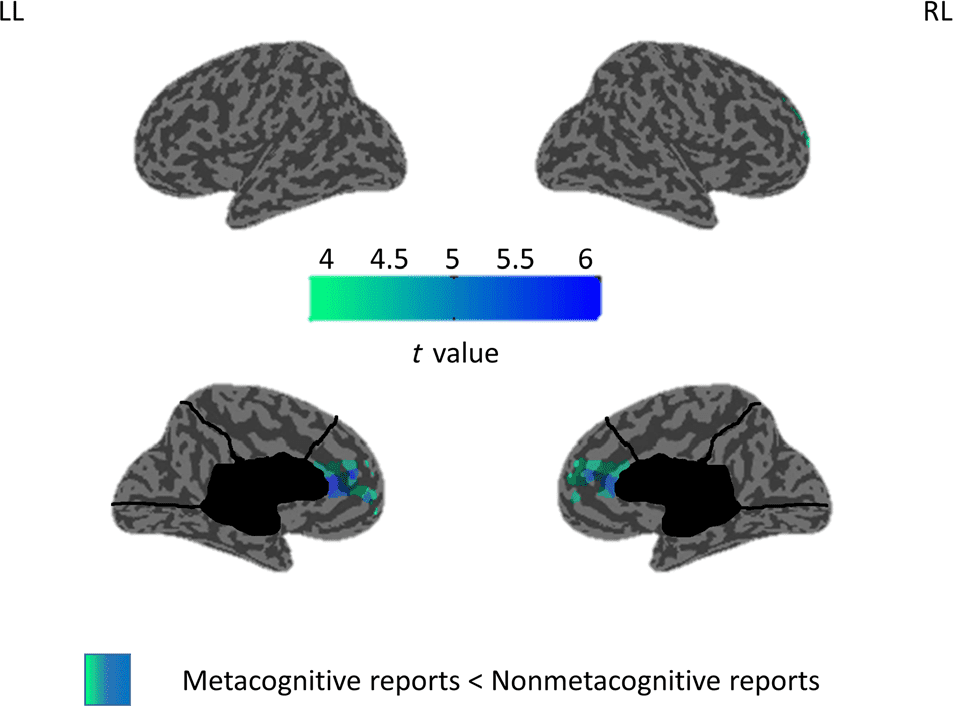
Metacognitive versus Nonmetacognitive Thoughts
in Wake
Given that metacognition was a major phenomenological
difference in thoughts between wakefulness and sleep,
we performed a supplementary EEG analysis, in which
we compared metacognitive thoughts (averaged score
1–5 between the two independent scorers in the “meta-
cognition” category), with thoughts involving no meta-
cognition (score 0) in wakefulness. We found higher
delta power (1–4 Hz) over a frontal region corresponding
to the mPFC in the reports with no metacognition com-
pared with the metacognitive trials (Figure 2). No signif-
icant changes were found for other frequency bands.
DISCUSSION
In this study, we investigated the neural correlates of
spontaneous thought across wakefulness, NREM sleep,
and REM sleep. The results indicate that experiences
characterized by high levels of thought were associated
with the activation of the midcingulate region across
all three states. Waking thoughts also involved a medial
prefrontal region, whose activation was associated with
higher metacognitive content than dreaming thoughts.
Aside from metacognitive content, spontaneous thoughts
in wakefulness did not differ from thoughts in NREM and
REM sleep in most phenomenological categories.
The Phenomenology of Spontaneous Thoughts in
Wake, NREM Sleep, and REM Sleep
Content analysis revealed that thoughts in wakefulness
did not differ from thoughts in NREM and REM sleep in
most categories. First, the proportion of episodic memo-
ries and thoughts about the present or future were similar
1770
Journal of Cognitive Neuroscience
Volume 29, Number 10
imagining events from the future or past of the individual
(Karapanagiotidis, Bernhardt, Jefferies, & Smallwood,
2017). Further investigation will be needed to determine
whether these frontal regions are directly involved in expe-
riential aspects of metacognitive thinking or are instead re-
cruited when metacognition poses additional cognitive
demands with no direct experiential correspondence.
We also observed that REM dreaming thoughts have
greater social focus than waking thoughts, as reported
previously (McNamara, McLaren, Smith, Brown, &
Stickgold, 2005). A possible explanation for this finding
is that typical dreams expose the person to richer envi-
ronments with more opportunities for diverse social in-
teractions than typical daydreams. Dreaming exposure
to rich social stimuli may permit an offline enhancement
of social cognition (Revonsuo, Tuominen, & Valli, 2015)
and of the so-called theory of mind (Fox et al., 2013;
Perogamvros, Dang-Vu, Desseilles, & Schwartz, 2013).
Another interesting finding is that cognitive effort was
higher in REM dreaming than in wakefulness and NREM
dreaming, even though thinking was more frequent in
wake than N2/REM sleep.
The Neural Correlates of Spontaneous Thought in
Wake, NREM Sleep, and REM Sleep
The present experiment highlights a midcingulate region
as a neural correlate of spontaneous thought across
wakefulness, NREM sleep, and REM sleep. Decreased
delta power was consistently observed in this region
when participants reported that their experience in-
volved thinking both during an experience-sampling
task in wakefulness and during serial awakenings from
sleep. The negative peak of slow waves in the EEG delta
frequency range (<4 Hz) is associated with neuronal
down states, during which neurons become hyperpolar-
ized and cease firing (Steriade, Timofeev, & Grenier,
2001), as confirmed by intracranial recordings in humans
(Nir et al., 2011). In contrast, EEG activation, associated
with decreased delta power, is associated with the recovery
of neural activity (Nir et al., 2011). The occurrence of
bistability and neuronal down states have been linked to
the loss of consciousness during both sleep and anesthe-
sia (Sachdev et al., 2015; Purdon et al., 2013; Tononi &
Massimini, 2008). Because down states lead to the break-
down of stable causal interactions among neurons (Pigorini
et al., 2015), a cortical area undergoing down states is
hypothesized to not contribute specific contents to con-
scious experiences (Tononi, Boly, Massimini, & Koch,
2016). Thus, our findings suggest that conscious thoughts
are less likely to be reported when midcingulate regions
are at least partially inactivated as indicated by the occur-
rence of slow waves. Although the occurrence of bistabil-
ity between “ON” and “OFF” periods is typical of NREM
sleep, recent studies have demonstrated the occurrence
of local EEG fluctuations in both the delta (Sachdev
et al., 2015) and theta range (Bernardi et al., 2015; Hung
Perogamvros et al.
1771
Figure 2. Metacognitive versus nonmetacognitive reports in wake.
Inflated cortical map illustrating the topographical distribution of
t values for the contrast between trials with no metacognition and
metacognition at the source level for delta power (1–4 Hz) in wake (last
20 sec before the alarm sound). Only significant differences at the
p < .05 level, obtained after correction for multiple comparisons, are
shown (two-tailed, paired t tests, seven participants).
across states. This result is consistent with the idea that
dreaming, similar to daydreaming, frequently involves past
memories, current concerns, and future plans (Wamsley,
2013; Perogamvros & Schwartz, 2012; Baird, Smallwood,
& Schooler, 2011). Moreover, we did not observe a dif-
ference in bizarreness between waking and dreaming
thoughts. Although we are hesitant to overinterpret a
null effect, one possible implication of this result is that,
although dreaming, particularly REM dreaming, frequently
involves bizarre elements (Schredl, 2010a), thoughts in
particular may be less bizarre and more consistent across
sleep and wake states (Fox et al., 2013; Wamsley, 2013;
Domhoff, 2007) than previously acknowledged (Williams,
Merritt, Rittenhouse, & Hobson, 1992).
However, waking thoughts differed from dreaming
thoughts, whether occurring in NREM sleep or in REM
sleep, by having higher metacognitive content. This finding
is partly consistent with previous work (Kahan & LaBerge,
2011), which found that metacognition about one’s own
thoughts, feelings, and behavior was one of the few dimen-
sions in which reported dreaming and waking experiences
differ. Our results suggest that the higher metacognitive
content of waking thoughts may be related to the involve-
ment of the mPFC (Figure 1D). Metacognitive reports com-
pared with nonmetacognitive reports in wake (Figure 2)
were associated with activation (reduced delta) of mPFC,
consistent with previous studies, which have found mPFC
involvement in metacognition (e.g., Baird, Smallwood,
Gorgolewski, & Margulies, 2013; Fleming, Huijgen, &
Dolan, 2012). The implication of this structure in metacog-
nition is also supported by a recent study that found
increased functional connectivity between the hippo-
campus and mPFC during a metacognitive process of
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
.
f
/
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
et al., 2013) in awake humans, often associated with tran-
sient behavioral impairments (Bernardi et al., 2015). Similar
local low-frequency fluctuations have been observed in
rodents, where they are associated with neuronal OFF
periods and behavioral misses ( Vyazovskiy et al., 2011).
Moreover, local slow waves and neuronal OFF periods have
been recently discovered also in REM sleep (Funk, Honjoh,
Rodriguez, Cirelli, & Tononi, 2016).
So far, studies examining the neural correlates of spon-
taneous thoughts had focused on the waking state (re-
viewed in Fox et al., 2015) and employed fMRI or PET,
methods that have lower temporal resolution than hd-
EEG. Brain areas related to spontaneous thought pro-
cesses were mainly part of the DMN, which encompasses
mPFC, precuneus/posterior cingulate cortex, and bilateral
inferior parietal lobule, as well as the medial-temporal
lobe/parahippocampal cortex (O’Callaghan, Shine, Lewis,
Andrews-Hanna, & Irish, 2015; Bernhardt et al., 2014;
Stawarczyk, Majerus, Maquet, & D’Argembeau, 2011;
Dumontheil, Gilbert, Frith, & Burgess, 2010; Christoff,
Gordon, Smallwood, Smith, & Schooler, 2009; Pagnoni,
Cekic, & Guo, 2008; Mason et al., 2007; McKiernan,
D’Angelo, Kaufman, & Binder, 2006; Christoff, Ream, &
Gabrieli, 2004). However, non-DMN structures have also
been implicated in spontaneous thought processes,
such as the dorsal ACC, midcingulate cortex, secondary
somatosensory cortex, insula, rostrolateral pFC, and
temporopolar cortex (Fox et al., 2015; Christoff et al.,
2009), as well as dorsolateral pFC (Smallwood, Brown,
Baird, & Schooler, 2012).
By considering the neural correlates of spontaneous
thought across wake, NREM sleep, and REM sleep, our
study highlights a more restricted brain region centered
on midcingulate cortex bilaterally. This region, which cor-
responds to large part of Brodmann’s area 24 (Palomero-
Gallagher, Vogt, Schleicher, Mayberg, & Zilles, 2009), lies
anatomically between the anterior and the posterior cin-
gulate cortex (Palomero-Gallagher et al., 2009; Vogt, Hof,
& Vogt, 2004). Functional studies have previously as-
sociated neural activity in this region with various uncon-
scious or conscious processes such as conflict monitoring
(Carter et al., 1998), readiness for action (Hoffstaedter,
Grefkes, Zilles, & Eickhoff, 2013), reward processing
(Parvizi, Rangarajan, Shirer, Desai, & Greicius, 2013; Bush
et al., 2002; Shima & Tanji, 1998), and processing of pain
and negative affect (Shackman et al., 2011). Consistent
with our findings, fMRI studies had also found that mid-
cingulate cortex is recruited, along with other regions,
during spontaneous thought in wakefulness (Bernhardt
et al., 2014; Hasenkamp, Wilson-Mendenhall, Duncan,
& Barsalou, 2012; Christoff et al., 2009). Specifically,
Christoff et al. (2009) demonstrated that episodes of
mind-wandering showed activation of the midcingulate
cortex, precuneus, and TPJ when compared with task-
related episodes. Hasenkamp et al. (2012) also observed
activity in the midcingulate cortex during self-reported
mind-wandering. Complementing these results, Bernhardt
et al. (2014) found that task-unrelated thoughts under low-
demanding conditions were associated with increased
cortical thickness of mPFC and midcingulate cortex. In
our experiments, midcingulate cortex was highlighted by
hd-EEG contrasts between high and low spontaneous
thought across all three behavioral states of wake, NREM
sleep, and REM sleep. Moreover, midcingulate cortex was
the only region to emerge as a neural correlate of sponta-
neous thought during REM sleep. Contrasts within NREM
sleep highlighted again midcingulate cortex as well as an
adjacent region of posterior cingulate cortex. Contrasts
within wake added premotor cortex, posterior cingulate
cortex, and mPFC. The more restricted and specific neural
correlates of thought were observed with contrasts within
sleep. This finding is consistent with the prediction that
sleep reduces confounding factors extraneous to conscious
thought, because sleeping participants, unlike awake ones,
are disconnected from the external environment, perform
no task, and have little or no cognitive control over the
content and form of the thought experience.
The fact that midcingulate cortex alone and not adja-
cent posterior cingulate cortex emerged as a neural cor-
relate of spontaneous thought in REM sleep deserves
some comment. It has been suggested that posterior cin-
gulate cortex, a core region of the DMN, may support in-
ternally generated thought (Fox et al., 2015; Leech,
Kamourieh, Beckmann, & Sharp, 2011), due to its capac-
ity to integrate information from different long-term
memory systems of the temporal lobe (Smallwood
et al., 2016; Andrews-Hanna et al., 2010). Our results in
wake and NREM also point to posterior cingulate cortex
as a possible neural correlate of spontaneous thought.
During REM sleep, however, posterior cingulate cortex
is deactivated compared with wakefulness or NREM
states (Fox et al., 2013; Braun et al., 1997; Maquet
et al., 1996). Because individuals are still perfectly capa-
ble of spontaneous thought during REM sleep, one inter-
pretation of these results is that the recruitment of
posterior cingulate cortex may not be a necessary condi-
tion for having conscious thoughts, and its involvement
in thought may be indirect. Alternatively, thought-related
changes in activity in the posterior cingulate region may
still occur during REM sleep but may be dampened and
thus more difficult to detect. Yet another possibility is
that REM sleep thoughts may lack some specific features
mediated by posterior cingulate cortex; however, in the
current analysis, we did not observe consistent differences
between REM thoughts and both N2 and wake thoughts
along a single content dimension.
The midcingulate cortex, especially its anterior portion,
has been associated with the so-called salience network,
which mediates appropriate responses to environmental
or internal stimuli of significant valence (Parvizi et al.,
2013; Seeley et al., 2007; Bush et al., 2002; Shima & Tanji,
1998; Williams & Goldman-Rakic, 1998). High activity in this
region could mean that the content of high-thought trials is
more salient for the participant than the one of low-thought
1772
Journal of Cognitive Neuroscience
Volume 29, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
t
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
trials. This region also receives dopaminergic input from
the ventral tegmental area (Hollerman & Schultz, 1998),
in line with suggestions that dreaming may be related to
the activation of cortical and subcortical reward structures,
as in the Reward Activation Model (Perogamvros &
Schwartz, 2012). Combined EEG/fMRI studies of dreaming
are needed to further investigate the hypothesis that cor-
tical and subcortical reward structures may be implicated
in dreaming, because hd-EEG is inadequate for accurately
localizing current sources in deep structures. Similarly, a
possible involvement of subcortical structures in the
medial- temporal lobe (hippocampus, parahippocampus)
in triggering spontaneous thoughts (Christoff et al., 2016;
Ellamil et al., 2016) cannot be ruled out. On the other hand,
cortical regions such as the inferior parietal lobule and
posterior insula, which had been proposed as possible early
generators of thought (Ellamil et al., 2016), were not
highlighted by our contrasts even though they are typically
accessible to hd-EEG.
In summary, hd-EEG topographical contrasts as well as
content analysis support the conclusion that conscious
thoughts share similar phenomenological features and
neural correlates across wakefulness, NREM, and REM
sleep, despite the otherwise different neurophysiological
profiles of these states. This finding is broadly consistent
with the claim that common brain regions are involved in
both daydreaming and dreaming (or rather, in REM sleep,
as most neuroimaging studies so far did not investigate
dreaming per se; Domhoff & Fox, 2015; Fox et al., 2013;
Nir & Tononi, 2010; Llinas & Pare, 1991). However, our
results also point to several important differences in
thoughts across waking and sleep states. The most nota-
ble one is that significant differences in prefrontal acti-
vation were not observed during NREM and REM sleep
thoughts, in contrast to thoughts during wake. Corre-
spondingly, the frequency of metacognitive thoughts is
also reduced in dreams, whereas the metacognitive con-
tent of thoughts during wake is associated with the acti-
vation of mPFC. Together, these observations point to
the hypothesis that prefrontal regions may either support
specific types of conscious thoughts (i.e., those involving
metacognitive content) or support unconscious functions
that support metacognitive thoughts (Fleming, Dolan, &
Frith, 2012).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
f
.
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Perogamvros et al.
1773
APPENDIX
Table A1. Definitions of Categories Used for Scoring Dream
and Waking Thoughts
Metacognition: thinking or reflecting about one’s experience
Social focus: experience related to other people or social
entities
Positive emotions: presence of positive emotions, such as
happiness, excitement, curiosity, etc.
Negative emotions: presence of negative emotions, such as
sadness, anger, boredom, etc.
Past focus: experience related to one’s autobiographical
(episodic) past events
Present focus: thoughts about something that is happening
right now
Future focus: thoughts about the future
Bizarreness: presence of impossible, unlikely or inconsistent
features in the experience
Table A2. Shapiro–Wilk Normality Test for the Categories Used
in Content Analysis
Shapiro–Wilk
Metacognition N2
Metacognition wake
Metacognition REM
Social N2
Social wake
Social REM
Positive emotion N2
Positive emotion W
Positive emotion REM
Negative emotion N2
Negative emotion W
Negative emotion REM
Past N2
Past W
Past REM
Present N2
Present W
Present REM
Future N2
Future W
Future REM
Bizarre N2
Bizarre W
Bizarre REM
Statistic
.221
.872
.188
.687
.941
.911
.791
.875
.932
.446
.689
.690
.576
.820
.592
.953
.948
.890
.635
.858
.557
.458
.615
.580
Sig.
<.001*
.055
<.001*
<.001*
.467
.021*
.005*
.060
.067
<.001*
<.001*
<.001*
<.001*
.12*
<.001*
.652
.573
.007
<.001*
.037
<.001*
<.001*
<.001*
<.001*
Several variables have non-normal distribution (marked with *p < .05).
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
/
.
f
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
1774
Journal of Cognitive Neuroscience
Volume 29, Number 10
Table A3. Mean Numbers (Scores) and Standard Deviations of Number of Words in the Report as well as Content Categories across
Wake, N2 Sleep, and REM Sleep
Words
Metacognition
Social focus
Positive emotions
Negative emotions
Past focus
Present focus
Future focus
Bizarreness
Cognitive effort
Duration
Richness/complexity
Recall of experience
Voluntary control
Wake
15.8 (±10.8)
0.27 (±0.25)
0.88 (±0.76)
0.61 (±0.55)
0.62 (±0.57)
0.56 (±0.43)
1.88 (±0.69)
0.63 (±0.59)
0.11 (±0.25)
0.76 (±0.67)
n/a
n/a
n/a
n/a
N2
14.1 (±7.3)
0.03 (±0.14)
1.66 (±1.66)
0.73 (±1.01)
0.50 (±0.86)
0.61 (±0.51)
1.95 (±0.68)
0.55 (±1.09)
0.17 (±0.47)
0.94 (±0.89)
3.03 (±0.79)
2.35 (±0.81)
2.67 (±0.91)
0.56 (±0.80)
REM
16.1 (±8.4)
0.007 (±0.04)
2.69 (±1.74)
1.90 (±1.35)
1.09 (±0.73)
0.47 (±0.92)
2.30 (±0.75)
0.45 (±0.91)
0.21 (±0.69)
1.98 (±1.29)
3.53 (±0.87)
3.06 (±0.88)
3.24 (±0.90)
0.58 (±0.86)
Acknowledgments
This work was supported by Swiss National Science Foundation
grant 155120 (L. P.), NIH/NCCAM P01AT004952 (G. T.), NIH/
NIMH 5P20MH077967 (G. T.), Tiny Blue Dot Inc. grant
MSN196438/AAC1335 (G. T.), and NIH/NINDS F32NS089348
(B. B.). The authors thank Stephanie Jones, David Bachhuber,
Anna Castelnovo, Francesca Siclari, Amelia Cayo, Chiara Cirelli,
William Marshall, Gary Garcia Molina, Armand Mensen, Poorang
Nori, Ana Maria Vascan, and the undergraduate research assis-
tants for help with data collection, sleep scoring, technical assis-
tance, and helpful discussions.
Reprint requests should be sent to Giulio Tononi, Department of
Psychiatry, University of Wisconsin-Madison, 6001 Research Park
Blvd., Madison, WI 53719, or via e-mail: gtononi@wisc.edu.
REFERENCES
Andrews-Hanna, J. R., Reidler, J. S., Huang, C., & Buckner,
R. L. (2010). Evidence for the default network’s role in
spontaneous cognition. Journal of Neurophysiology, 104,
322–335.
Baird, B., Smallwood, J., Gorgolewski, K. J., & Margulies, D. S.
(2013). Medial and lateral networks in anterior prefrontal
cortex support metacognitive ability for memory and
perception. Journal of Neuroscience, 33, 16657–16665.
Baird, B., Smallwood, J., Lutz, A., & Schooler, J. W. (2014).
The decoupled mind: Mind-wandering disrupts cortical
phase-locking to perceptual events. Journal of Cognitive
Neuroscience, 26, 2596–2607.
Baird, B., Smallwood, J., & Schooler, J. W. (2011). Back to
the future: Autobiographical planning and the functionality
of mind-wandering. Consciousness and Cognition, 20,
1604–1611.
Bates, D., Maechler, M., Bolker, B., & Walker, S. (2015). Fitting
linear mixed-effects models using lme4. Journal of Statistical
Software, 67, 1–48.
Bernardi, G., Siclari, F., Yu, X., Zennig, C., Bellesi, M., Ricciardi,
E., et al. (2015). Neural and behavioral correlates of extended
training during sleep deprivation in humans: Evidence for
local, task-specific effects. Journal of Neuroscience, 35,
4487–4500.
Bernhardt, B. C., Smallwood, J., Tusche, A., Ruby, F. J., Engen,
H. G., Steinbeis, N., et al. (2014). Medial prefrontal and
anterior cingulate cortical thickness predicts shared
individual differences in self-generated thought and temporal
discounting. Neuroimage, 90, 290–297.
Braboszcz, C., & Delorme, A. (2011). Lost in thoughts: Neural
markers of low alertness during mind wandering.
Neuroimage, 54, 3040–3047.
Braun, A. R., Balkin, T. J., Wesenten, N. J., Carson, R. E., Varga,
M., Baldwin, P., et al. (1997). Regional cerebral blood flow
throughout the sleep-wake cycle. An H2(15)O PET study.
Brain, 120, 1173–1197.
Bush, G., Vogt, B. A., Holmes, J., Dale, A. M., Greve, D., Jenike,
M. A., et al. (2002). Dorsal anterior cingulate cortex: A role in
reward-based decision making. Proceedings of the National
Academy of Sciences, U.S.A., 99, 523–528.
Carter, C. S., Braver, T. S., Barch, D. M., Botvinick, M. M., Noll,
D., & Cohen, J. D. (1998). Anterior cingulate cortex, error
detection, and the online monitoring of performance.
Science, 280, 747–749.
Cartwright, R., Agargun, M. Y., Kirkby, J., & Friedman, J. K.
(2006). Relation of dreams to waking concerns. Psychiatry
Research, 141, 261–270.
Christoff, K., Gordon, A. M., Smallwood, J., Smith, R., &
Schooler, J. W. (2009). Experience sampling during fMRI
reveals default network and executive system contributions
to mind wandering. Proceedings of the National Academy of
Sciences, U.S.A., 106, 8719–8724.
Perogamvros et al.
1775
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
.
f
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
Christoff, K., Irving, Z. C., Fox, K. C., Spreng, R. N., & Andrews-
Hanna, J. R. (2016). Mind-wandering as spontaneous thought:
A dynamic framework. Nature Reviews. Neuroscience, 17,
718–731.
Christoff, K., Ream, J. M., & Gabrieli, J. D. (2004). Neural basis
of spontaneous thought processes. Cortex, 40, 623–630.
Delorme, A., & Makeig, S. (2004). EEGLAB: An open source
toolbox for analysis of single-trial EEG dynamics including
independent component analysis. Journal of Neuroscience
Methods, 134, 9–21.
Domhoff, G. (2007). Realistic simulation and bizarreness in
dream content: Past findings and suggestions for future
research (Vol. 2). Westport, CT: Praeger Press.
Domhoff, G. W., & Fox, K. C. (2015). Dreaming and the default
network: A review, synthesis, and counterintuitive research
proposal. Consciousness and Cognition, 33, 342–353.
Dumontheil, I., Gilbert, S. J., Frith, C. D., & Burgess, P. W.
(2010). Recruitment of lateral rostral prefrontal cortex in
spontaneous and task-related thoughts. Quarterly Journal of
Experimental Psychology (Hove), 63, 1740–1756.
Ellamil, M., Fox, K. C., Dixon, M. L., Pritchard, S., Todd, R. M.,
Thompson, E., et al. (2016). Dynamics of neural recruitment
surrounding the spontaneous arising of thoughts in
experienced mindfulness practitioners. Neuroimage, 136,
186–196.
Fleming, S. M., Dolan, R. J., & Frith, C. D. (2012).
Metacognition: Computation, biology and function.
Philosophical Transactions of the Royal Society of London,
Series B, Biological Sciences, 367, 1280–1286.
Fleming, S. M., Huijgen, J., & Dolan, R. J. (2012). Prefrontal
contributions to metacognition in perceptual decision
making. Journal of Neuroscience, 32, 6117–6125.
Fox, K. C., Nijeboer, S., Solomonova, E., Domhoff, G. W., &
Christoff, K. (2013). Dreaming as mind wandering: Evidence
from functional neuroimaging and first-person content
reports. Frontiers in Human Neuroscience, 7, 412.
Fox, K. C., Spreng, R. N., Ellamil, M., Andrews-Hanna, J. R., &
Christoff, K. (2015). The wandering brain: Meta-analysis of
functional neuroimaging studies of mind-wandering and
related spontaneous thought processes. Neuroimage, 111,
611–621.
Funk, C. M., Honjoh, S., Rodriguez, A. V., Cirelli, C., &
Tononi, G. (2016). Local slow waves in superficial layers
of primary cortical areas during REM sleep. Current Biology,
26, 396–403.
Hallgren, K. A. (2012). Computing inter-rater reliability for
observational data: An overview and tutorial. Tutorial in
Quantitative Methods for Psychology, 8, 23–34.
Hasenkamp, W., Wilson-Mendenhall, C. D., Duncan, E., &
Barsalou, L. W. (2012). Mind wandering and attention during
focused meditation: A fine-grained temporal analysis of
fluctuating cognitive states. Neuroimage, 59, 750–760.
Hoffstaedter, F., Grefkes, C., Zilles, K., & Eickhoff, S. B. (2013).
The “what” and “when” of self-initiated movements. Cerebral
Cortex, 23, 520–530.
Hollerman, J. R., & Schultz, W. (1998). Dopamine neurons
report an error in the temporal prediction of reward during
learning. Nature Neuroscience, 1, 304–309.
Holm, S. (1979). A simple sequentially rejective multiple test
procedure. Scandinavian Journal of Statistics, 6, 65–70.
Hung, C. S., Sarasso, S., Ferrarelli, F., Riedner, B., Ghilardi,
M. F., Cirelli, C., et al. (2013). Local experience-dependent
changes in the wake EEG after prolonged wakefulness. Sleep,
36, 59–72.
Iber, C., Ancoli-Israel, S., Chesson, A., & Quan, S. F. (2007). The
AASM manual for the scoring of sleep and associated events:
Rules, terminology and technical specifications (1st ed.).
Westchester, IL: American Academy of Sleep Medicine.
Jones, B. E. (2005). From waking to sleeping: Neuronal and
chemical substrates. Trends in Pharmacological Sciences,
26, 578–586.
Jung, T. P., Makeig, S., Humphries, C., Lee, T. W., McKeown, M. J.,
Iragui, V., et al. (2000). Removing electroencephalographic
artifacts by blind source separation. Psychophysiology, 37,
163–178.
Kahan, T. L., LaBerge, S., Levitan, L., & Zimbardo, P. (1997).
Similarities and differences between dreaming and waking
cognition: An exploratory study. Consciousness and
Cognition, 6, 132–147.
Kahan, T. L., & LaBerge, S. P. (2011). Dreaming and waking:
Similarities and differences revisited. Consciousness and
Cognition, 20, 494–514.
Kam, J. W., Dao, E., Farley, J., Fitzpatrick, K., Smallwood, J.,
Schooler, J. W., et al. (2011). Slow fluctuations in attentional
control of sensory cortex. Journal of Cognitive Neuroscience,
23, 460–470.
Kane, M. J., Brown, L. H., McVay, J. C., Silvia, P. J., Myin-
Germeys, I., & Kwapil, T. R. (2007). For whom the mind
wanders, and when: An experience-sampling study of
working memory and executive control in daily life.
Psychological Science, 18, 614–621.
Karapanagiotidis, T., Bernhardt, B. C., Jefferies, E., &
Smallwood, J. (2017). Tracking thoughts: Exploring the
neural architecture of mental time travel during mind-
wandering. Neuroimage, 147, 272–281.
Killingsworth, M. A., & Gilbert, D. T. (2010). A wandering mind
is an unhappy mind. Science, 330, 932.
Klinger, E., & Cox, W. M. (1987). Dimensions of thought flow
in everyday life. Imagination, Cognition and Personality,
7, 105–128.
La Berge, S. P., Nagel, L. E., Dement, W. C., & Zarcone,
V. P., Jr. (1981). Lucid dreaming verified by volitional
communication during REM sleep. Perceptual and Motor
Skills, 52, 727–732.
Leech, R., Kamourieh, S., Beckmann, C. F., & Sharp, D. J.
(2011). Fractionating the default mode network: Distinct
contributions of the ventral and dorsal posterior cingulate
cortex to cognitive control. Journal of Neuroscience, 31,
3217–3224.
Llinas, R. R., & Pare, D. (1991). Of dreaming and wakefulness.
Neuroscience, 44, 521–535.
Maquet, P., Peters, J., Aerts, J., Delfiore, G., Degueldre, C., Luxen,
A., et al. (1996). Functional neuroanatomy of human
rapid-eye-movement sleep and dreaming. Nature, 383, 163–166.
Mason, M. F., Norton, M. I., Van Horn, J. D., Wegner, D. M.,
Grafton, S. T., & Macrae, C. N. (2007). Wandering minds: The
default network and stimulus-independent thought. Science,
315, 393–395.
McKiernan, K. A., D’Angelo, B. R., Kaufman, J. N., & Binder, J. R.
(2006). Interrupting the “stream of consciousness”: An fMRI
investigation. Neuroimage, 29, 1185–1191.
McNamara, P., McLaren, D., Smith, D., Brown, A., & Stickgold,
R. (2005). A “Jekyll and Hyde” within: Aggressive versus
friendly interactions in REM and non-REM dreams.
Psychological Science, 16, 130–136.
Mensen, A., & Khatami, R. (2013). Advanced EEG analysis using
threshold-free cluster-enhancement and non-parametric
statistics. Neuroimage, 67, 111–118.
Nielsen, T. A., & Stenstrom, P. (2005). What are the memory
sources of dreaming? Nature, 437, 1286–1289.
Nir, Y., Staba, R. J., Andrillon, T., Vyazovskiy, V. V., Cirelli, C.,
Fried, I., et al. (2011). Regional slow waves and spindles in
human sleep. Neuron, 70, 153–169.
Nir, Y., & Tononi, G. (2010). Dreaming and the brain: From
phenomenology to neurophysiology. Trends in Cognitive
Sciences, 14, 88–100.
1776
Journal of Cognitive Neuroscience
Volume 29, Number 10
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
f
/
t
t
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
O’Callaghan, C., Shine, J. M., Lewis, S. J., Andrews-Hanna, J. R., &
Irish, M. (2015). Shaped by our thoughts—A new task to assess
spontaneous cognition and its associated neural correlates in
the default network. Brain and Cognition, 93, 1–10.
Pagnoni, G., Cekic, M., & Guo, Y. (2008). “Thinking about
not-thinking”: Neural correlates of conceptual processing
during Zen meditation. PLoS One, 3, e3083.
Palomero-Gallagher, N., Vogt, B. A., Schleicher, A., Mayberg, H. S.,
& Zilles, K. (2009). Receptor architecture of human cingulate
cortex: Evaluation of the four-region neurobiological model.
Human Brain Mapping, 30, 2336–2355.
Parvizi, J., Rangarajan, V., Shirer, W. R., Desai, N., & Greicius,
M. D. (2013). The will to persevere induced by electrical
stimulation of the human cingulate gyrus. Neuron, 80,
1359–1367.
Pascual-Marqui, R. D. (2002). Standardized low-resolution brain
electromagnetic tomography (sLORETA): Technical details.
Methods and Findings in Experimental and Clinical
Pharmacology, 24(Suppl. D), 5–12.
Perogamvros, L., Dang-Vu, T. T., Desseilles, M., & Schwartz, S.
(2013). Sleep and dreaming are for important matters.
Frontiers in Psychology, 4, 474.
Perogamvros, L., & Schwartz, S. (2012). The roles of the reward
system in sleep and dreaming. Neuroscience and
Biobehavioral Reviews, 36, 1934–1951.
Pigorini, A., Sarasso, S., Proserpio, P., Szymanski, C., Arnulfo, G.,
Casarotto, S., et al. (2015). Bistability breaks-off deterministic
responses to intracortical stimulation during non-REM sleep.
Neuroimage, 112, 105–113.
Purdon, P. L., Pierce, E. T., Mukamel, E. A., Prerau, M. J., Walsh,
J. L., Wong, K. F., et al. (2013). Electroencephalogram
signatures of loss and recovery of consciousness from
propofol. Proceedings of the National Academy of Sciences,
U.S.A., 110, E1142–E1151.
R Development Core Team. 2006. R: A language and
environment for statistical computing, R Foundation for
Statistical Computing. Vienna, Austria. http://www.R-project.org.
Revonsuo, A., Tuominen, J., & Valli, K. (2015). The avatars in
the machine—Dreaming as a simulation of social reality. In T.
Metzinger & J. M. Windt (Eds.), Open MIND: 32(1). Frankfurt
am Main: MIND Group.
Sachdev, R. N., Gaspard, N., Gerrard, J. L., Hirsch, L. J., Spencer,
D. D., & Zaveri, H. P. (2015). Delta rhythm in wakefulness:
Evidence from intracranial recordings in human beings.
Journal of Neurophysiology, 114, 1248–1254.
Schredl, M. (2010a). Characteristics and contents of dreams.
International Review of Neurobiology, 92, 135–154.
Schredl, M. (2010b). Dream content analysis: Basic principles.
International Journal of Dream Research, 3, 65–73.
Seeley, W. W., Menon, V., Schatzberg, A. F., Keller, J., Glover,
G. H., Kenna, H., et al. (2007). Dissociable intrinsic connectivity
networks for salience processing and executive control.
Journal of Neuroscience, 27, 2349–2356.
Shackman, A. J., Salomons, T. V., Slagter, H. A., Fox, A. S.,
Winter, J. J., & Davidson, R. J. (2011). The integration of
negative affect, pain and cognitive control in the cingulate
cortex. Nature Reviews. Neuroscience, 12, 154–167.
Shima, K., & Tanji, J. (1998). Role for cingulate motor area cells
in voluntary movement selection based on reward. Science,
282, 1335–1338.
Siclari, F., Larocque, J. J., Postle, B. R., & Tononi, G. (2013).
Assessing sleep consciousness within subjects using a serial
awakening paradigm. Frontiers in Psychology, 4, 542.
Smallwood, J., Beach, E., Schooler, J. W., & Handy, T. C. (2008).
Going AWOL in the brain: Mind wandering reduces cortical
analysis of external events. Journal of Cognitive
Neuroscience, 20, 458–469.
Smallwood, J., Brown, K., Baird, B., & Schooler, J. W. (2012).
Cooperation between the default mode network and the
frontal-parietal network in the production of an internal train
of thought. Brain Research, 1428, 60–70.
Smallwood, J., Fitzgerald, A., Miles, L. K., & Phillips, L. H. (2009).
Shifting moods, wandering minds: Negative moods lead the
mind to wander. Emotion, 9, 271–276.
Smallwood, J., Karapanagiotidis, T., Ruby, F., Medea, B., de
Caso, I., Konishi, M., et al. (2016). Representing
representation: Integration between the temporal lobe and
the posterior cingulate influences the content and form of
spontaneous thought. PLoS One, 11, e0152272.
Smallwood, J., & Schooler, J. W. (2006). The restless mind.
Psychological Bulletin, 132, 946–958.
Smith, S. M., & Nichols, T. E. (2009). Threshold-free cluster
enhancement: Addressing problems of smoothing, threshold
dependence and localisation in cluster inference.
Neuroimage, 44, 83–98.
Stawarczyk, D., Majerus, S., Maquet, P., & D’Argembeau, A.
(2011). Neural correlates of ongoing conscious experience:
Both task-unrelatedness and stimulus-independence are
related to default network activity. PLoS One, 6, e16997.
Steriade, M., Timofeev, I., & Grenier, F. (2001). Natural waking
and sleep states: A view from inside neocortical neurons.
Journal of Neurophysiology, 85, 1969–1985.
Tononi, G., Boly, M., Massimini, M., & Koch, C. (2016). Integrated
information theory: From consciousness to its physical
substrate. Nature Reviews. Neuroscience, 17, 450–461.
Tononi, G., & Massimini, M. (2008). Why does consciousness
fade in early sleep? The Annals of the New York Academy of
Sciences, 1129, 330–334.
Vogt, B. A., Hof, P. R., & Vogt, L. J. (2004). Cingulate gyrus.
Amsterdam: Elsevier.
Vyazovskiy, V. V., Olcese, U., Hanlon, E. C., Nir, Y., Cirelli, C., &
Tononi, G. (2011). Local sleep in awake rats. Nature, 472,
443–447.
Wamsley, E. J. (2013). Dreaming, waking conscious experience,
and the resting brain: Report of subjective experience as a
tool in the cognitive neurosciences. Frontiers in Psychology,
4, 637.
Williams, J., Merritt, J., Rittenhouse, C., & Hobson, J. (1992).
Bizarreness in dreams and fantasies: Implications for the
activation-synthesis hypothesis. Consciousness and
Cognition, 1, 172–185.
Williams, S. M., & Goldman-Rakic, P. S. (1998). Widespread
origin of the primate mesofrontal dopamine system. Cerebral
Cortex, 8, 321–345.
Perogamvros et al.
1777
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
o
d
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
9
/
1
2
0
9
/
1
1
7
0
6
/
6
1
1
7
9
6
5
6
2
/
9
1
0
4
7
8
o
6
c
6
n
7
_
3
a
/
_
j
0
o
1
c
1
n
5
5
_
a
p
_
d
0
1
b
1
y
5
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
t
f
/
.
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1