The Default Mode of Human Brain Function Primes the
Intentional Stance
Robert P. Spunt1, Meghan L. Meyer2, and Matthew D. Lieberman2
Abstrait
■ Humans readily adopt an intentional stance to other people,
comprehending their behavior as guided by unobservable mental
states such as belief, désir, and intention. We used fMRI in healthy
adults to test the hypothesis that this stance is primed by the de-
fault mode of human brain function present when the mind is at
repos. We report three findings that support this hypothesis. D'abord,
brain regions activated by actively adopting an intentional rather
than nonintentional stance to a social stimulus were anatomically
similar to those demonstrating default responses to fixation base-
line in the same task. Deuxième, moment-to-moment variation in
default activity during fixation in the dorsomedial PFC was re-
lated to the ease with which participants applied an intentional—
but not nonintentional—stance to a social stimulus presented mo-
ments later. Enfin, individuals who showed stronger dorsomedial
PFC activity at baseline in a separate task were generally more ef-
ficient when adopting the intentional stance and reported having
greater social skills. These results identify a biological basis for the
human tendency to adopt the intentional stance. More broadly,
they suggest that the brain’s default mode of function may have
evolved, in part, as a response to life in a social world. ■
INTRODUCTION
Humans have a seemingly irresistible tendency to con-
ceive the actions of others as intentional and guided by
beliefs and desires (Rosset, 2008; Uleman, Adil Saribay, &
González, 2008; Mesoudi, Whiten, & Dunbar, 2006;
Dennett, 1989; Vallacher & Wegner, 1987; Heider &
Simmel, 1944). This intentional stance toward other
humans is already apparent in the first year of life
(Kovacs, Teglas, & Endress, 2010) and eventually be-
comes so automatized that it is effortlessly adopted to
understand the behavior of not just other humans but
also pets and iPhones (Epley, Waytz, & Cacioppo, 2007).
The importance of the intentional stance is highlighted
by the enormous difficulties faced by those who are not
predisposed to it, such as individuals with an autism
spectrum disorder (Senju, 2012). Although the tendency
to engage the intentional stance is regarded as essential to
human sociality, the neurobiological basis of this pre-
paredness remains a mystery.
Numerous functional neuroimaging studies in humans
have demonstrated that a psychological process at the
core of the intentional stance—mental state inference—
is reliably associated with a set of cortical regions com-
monly referred to as the theory-of-mind or mentalizing
réseau (Amodio & Frith, 2006; Saxe, Carey, & Kanwisher,
2004; Gallagher & Frith, 2003; Happé et al., 1996; Fletcher
et coll., 1995; Goel, Grafman, Sadato, & Hallett, 1995). Pour
instance, our own work has shown that regions of this net-
1California Institute of Technology, 2Université de Californie,
Les anges
© 2015 Massachusetts Institute of Technology
travail, namely dorsomedial PFC (dmPFC), precuneus, TPJ,
and anterior superior temporal sulcus (STS), show a supra-
modal association with the use of mental state concepts
to produce and evaluate explanations of others’ actions
and emotional behavior (Spunt & Adolphs, 2014; Spunt
& Lieberman, 2012un, 2012b; Spunt, Satpute, & Lieberman,
2011). Although these studies outline the functional neuro-
anatomy of experimentally induced mental state inference,
it remains unknown why the human mind seems naturally
primed to adopt the intentional stance in the first place.
The explanation considered here is motivated by two
empirical facts about the human brain. The first is that
most of the brain’s energy budget is consumed not by
activity evoked by specific cognitive tasks (par exemple., mental
arithmetic) but by spontaneous ongoing activity that is
most notable when the brain is at rest (Raichle, 2010).
This spontaneous activity is most prominent in a dis-
tributed cortical network commonly referred to as the
default-mode network (DMN; Greicius, Krasnow, Reiss,
& Menon, 2003; Mazoyer et al., 2001; Raichle et al.,
2001; Binder et al., 1999; Shulman et al., 1997). Given
that the brain consumes a disproportionate amount of
the energy available to the body (Attwell & Laughlin,
2001), it is likely that the persistent activity of the DMN
during periods of rest serves important adaptive func-
tion (Andrews-Hanna, Smallwood, & Spreng, 2014).
Ici, we consider a function of the DMN that is sug-
gested by a second fact about the human brain: Le
anatomical boundaries of the DMN largely correspond
with the neuroanatomy associated with adopting the
intentional stance (Mars et al., 2012; Schilbach et al.,
2012; Spreng, Mar, & Kim, 2009; Schilbach, Eickhoff,
Journal des neurosciences cognitives 27:6, pp. 1116–1124
est ce que je:10.1162/jocn_a_00785
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
6
2
7
1
/
1
6
1
/
6
1
1
1
9
1
4
6
9
/
1
1
1
7
3
8
o
3
c
2
n
6
_
6
un
/
_
j
0
o
0
c
7
n
8
5
_
un
p
_
d
0
0
b
7
oui
8
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
.
F
/
t
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Rotarska-Jagiela, Fink, & Vogeley, 2008). Surtout, ce
anatomical coincidence does not permit the conclusion
that spontaneous DMN at rest is functionally relevant
for adopting the intentional stance in response to social
stimuli. This is because regional brain activity observed
under different conditions (par exemple., resting vs. inferring a
mental state) may reflect different underlying processes
(Poldrack, 2006). Ainsi, this anatomical coincidence
raises an important yet unanswered question: Fait
spontaneous activity in the DMN during periods of men-
tal rest prime the intentional stance, preparing us to con-
ceive others as minds and not merely bodies?
A handful of neuroimaging studies already illustrate
that intraindividual variability in the neural (Fox,
Snyder, Zacks, & Raichle, 2006; Arieli, Sterkin, Grinvald, &
Aertsen, 1996) and behavioral (Callard & Margulies, 2014;
Hsieh, Colas, & Kanwisher, 2012; Fox, Snyder, Vincent, &
Raichle, 2007) response to a nonsocial stimuli can be par-
tially explained by spontaneous brain activity occurring in
the resting periods before stimulus onset. Building on this
logic, we designed a novel fMRI task to test the hypothesis
that default activity in the DMN functions to prepare the
mind to adopt the intentional stance to social stimuli. Ce
hypothesis also has strong theoretical ties to large body of
research on priming and accessibility in social and cogni-
tive psychology (Tulving & Schacter, 1990; Higgins, 1989;
Neely, 1977), which reliably observes that the efficiency of
evaluating a target stimulus (par exemple., the word “DOCTOR”) est
increased by recent exposure to a conceptually related
priming stimulus (par exemple., the word “NURSE”). Ainsi, notre
hypothesis can be elaborated as follows: If spontaneous
activity in the DMN between stimulus events involves
mental operations that are similar to those involved when
adopting the intentional stance, then spontaneous DMN
activity before encountering a social stimulus may make
it easier to adopt an intentional (rather than noninten-
tional) stance to that stimulus. If this is true, then sustained
activity in the DMN during periods of rest might serve as an
endogenous prime that makes an intentional stance the
default strategy for making sense of the social world.
MÉTHODES
Participants
Twenty-one right-handed participants (10 men, 11 femmes;
âge moyen = 22.86 années, age range = 18–31 years) étaient
recruited from the University of California, Les anges
(UCLA) participant pool and provided written informed
consent according to the procedures of the UCLA institu-
tional review board. All participants were native English
speakers and were not taking psychotropic medications
at the time of the study.
Judgment Task
The primary experimental task (Figure 1A) involved mak-
ing speeded yes/no judgments under three conditions.
Mind-focused judgments evoked the intentional stance
by asking participants to evaluate the appropriateness
of a sentence describing the mental state of a person in
a photograph. Body-focused judgments featured the
same photographs but evoked a nonintentional stance
by asking participants to evaluate a sentence providing
a physical description of the person who is performing
an action. In numerous published studies, we have
shown that conceptually similar manipulations robustly
and selectively modulate activity in the regions of the
brain associated with mental state reasoning (Spunt &
Adolphs, 2014; Spunt & Lieberman, 2012un, 2012b; Spunt
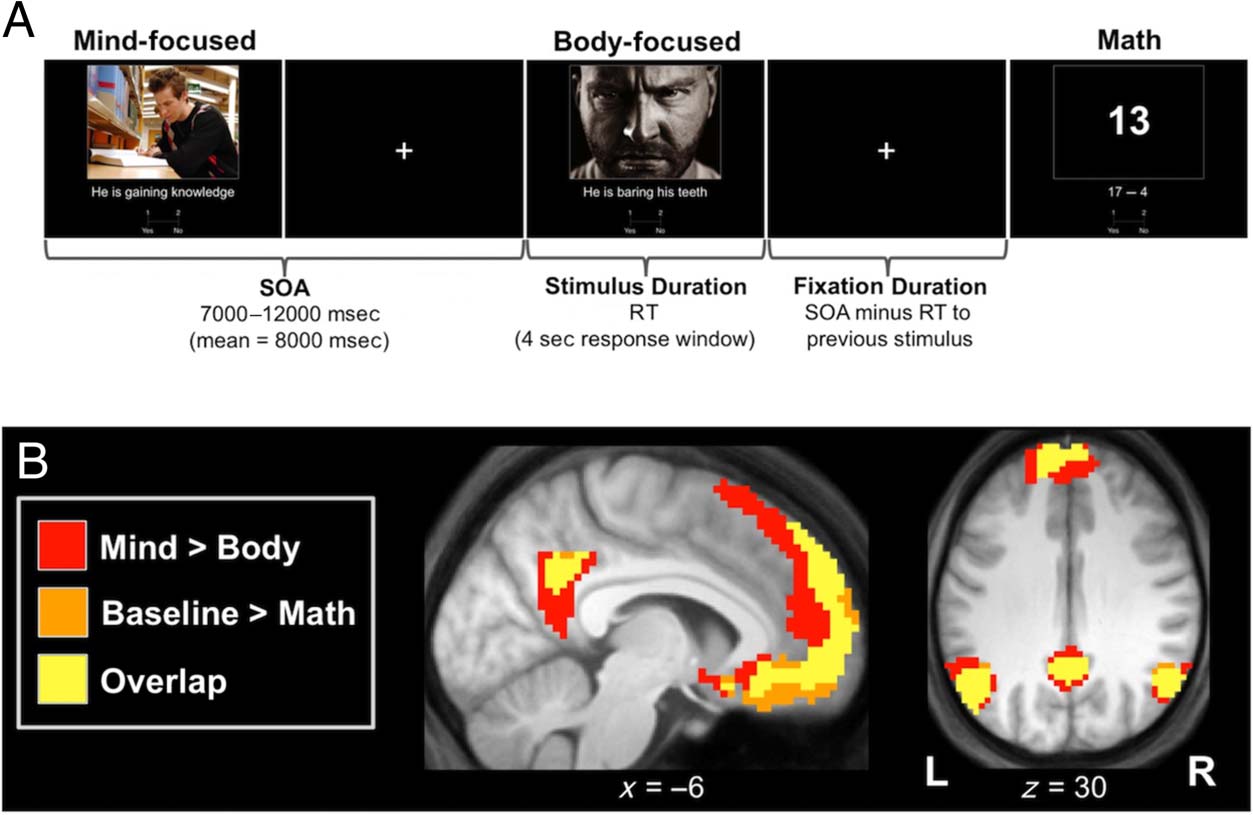
Chiffre 1. (UN) Schematic of the
event-related design used to
manipulate social judgments
focused on either a person’s
mind or body. To isolate
spontaneous DMN activity
related to the fixation baseline
periods dividing each trial,
structurally similar mathematical
judgments were interleaved
with these social judgments.
Although the examples used to
illustrate the mind-focused and
body-focused judgments feature
different photographs, tous
photographs were the object of
one mind-focused judgment and
one body-focused judgment.
(B) Anatomical overlap of brain
networks associated with mind-
focused judgments and the
default mode. Individual
contrasts of interest were first thresholded so that all remaining clusters were significant at an FWE rate of 0.05. These maps were then binarized,
assigned a color as indicated, and overlaid on the group mean anatomical image. See Table 2 for regions surviving a test of these contrasts against the
conjunction null. L = left; R = right.
Spunt, Meyer, and Lieberman
1117
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
6
2
7
1
/
1
6
1
/
6
1
1
1
9
1
4
6
9
/
1
1
1
7
3
8
o
3
c
2
n
6
_
6
un
/
_
j
0
o
0
c
7
n
8
5
_
un
p
_
d
0
0
b
7
oui
8
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
F
t
.
/
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
et coll., 2011). Enfin, math judgments were entirely non-
social and asked participants to evaluate arithmetical
expressions. Mental arithmetic is a cognitive task known
to reliably suppress activity in the DMN (Mazoyer et al.,
2001) and would thus provide a method for independently
defining regions demonstrating high activity during the
fixation baseline period that preceded each judgment.
The mind-focused and body-focused conditions fea-
tured 40 naturalistic photographs of people performing
goal-directed actions and/or displaying expressions of
emotion. Each photograph was paired with two sen-
tences, one that described an inference about the per-
son’s state of mind and one that described a physical
feature of their behavior. For both conditions, 70% de
the sentences were intended to provide an accurate or
plausible description, whereas the remaining 30% étaient
intended to provide an inaccurate or implausible descrip-
tion. The sentences featured in the two conditions were
matched on length (mean number of characters: descrip-
tive statements = 23.58, inferential statements = 23.23).
To create the stimuli used in the experiment, each sen-
tence was paired with its corresponding photograph in a
single image (image size = 800 × 600 pixels, photograph
size = 509 × 382 pixels, font height = 33 pixels, black
background with white foreground). En outre, a 2-point
yes/no scale was added to the bottom of each image.
Enfin, the arithmetic condition featured the 20 integers
depuis 10 à 29, each of which was paired with two arith-
metical expressions (70/30 correct/incorrect), one per-
forming addition (par exemple., 14 + 2) and one performing
subtraction (par exemple., 20 − 4). The formatting for the final arith-
metic stimuli was the same as that used for the social stim-
uli, with the target integers (font height = 96 pixels) printed
in the center of a rectangular white line with the same
dimensions as the photographs used in the social conditions.
In a separate laboratory-based study, 72 undergradu-
ates (30 men, 42 femmes; âge moyen = 20.64 années, SD =
3.64 années) from the UCLA performed the judgment task
while seated at a computer station. When examining
normative data on the individual stimuli, all stimuli elicited
an accuracy rate of at least 79.5%. En effet, the accuracy
ranges for the individual stimuli across the three conditions
were very similar (why = 80.7–100.0%, how = 79.5–
100.0%, math = 79.5–100.0%). Surtout, a repeated-
measures ANOVA revealed no significant effect of Judgment
type on accuracy rates, F(2, 142) = 0.754, p = .47.
During functional MRI scanning, le 120 trials (40 esprit-
focused, 40 body-focused, 40 math) were presented to
participants in an event-related design (Figure 1A). Chaque
trial was presented for a maximum duration of 4 sec, et
RT to trial onset was recorded at participant response. If
the participant responded before 4 sec elapsing, the exper-
imental stimulus was replaced with a fixation crosshair
stimulus, which remained onscreen until the onset of the
next trial. The order and onset of trials were optimized for
estimation efficiency using custom MATLAB software (Le
MathWorks, Natick, MA). Trial order was constrained so
that the maximum number of consecutive trials from the
same condition was 2. Onsets were constrained so that
the SOA had a mean of 7.5 sec (min = 6.5 sec, max =
9.5 sec).
The following procedures were used to prepare all
participants for task performance. Before entering the
scanner, participants were told they would perform a task
requiring them to make judgments about people and
numbers. They were then shown two trials from each
of the three conditions. For people trials, they were told
to indicate whether the bottom statement is a good
description of what they see happening in the photo-
graph. For number trials, they were told to indicate
whether the bottom statement equals the number in
the box. For both trial types, participants were told to
respond quickly and accurately. Immediately before start-
ing the task in the scanner, participants were shown a
screen with the same instructions and were given the
opportunity to ask questions before beginning.
Match-to-Sample Task
After performance of the judgment task described above,
participants performed a blocked visual match-to-sample
task that would allow us to independently assess DMN
activation levels in each participant. For each trial, partic-
ipants judged which of two shapes matched a target
shape in both shape and orientation (see Figure 2A for
an example trial; image size = 800 × 600 pixels, shape
height = 94 pixels, black background with white fore-
ground). Participants had 2 sec to respond to each trial,
and trials were presented in blocks of nine. The onset
and offset of each block featured brief cues (1 sec) dans-
structing participants to “Get Ready!” or “Relax!», respecter-
tivement. Each block was preceded and followed by a 20-sec
rest period featuring a fixation cross centered onscreen.
Stimulus Presentation and Response Recording
For both tasks, stimuli were presented using the MATLAB
Psychophysics Toolbox (version 3.0.9; Brainard, 1997).
Participants viewed the stimuli through LCD goggles
(800 × 600 pixels) and made their responses with a
button box using their right-hand index and middle
fingers.
Personality Measures
Before their scanning session, all participants were asked
to complete an online survey that included two person-
ality questionnaires that were examined for this study.
This study was not specifically designed to examine indi-
vidual differences and indeed is underpowered in this re-
spect (Button et al., 2013; Yarkoni, 2009). Ainsi, nous
clarify that these analyses were conducted only to pro-
vide additional constraint on interpreting the effects ob-
served in our primary within-subject analysis.
1118
Journal des neurosciences cognitives
Volume 27, Nombre 6
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
6
2
7
1
/
1
6
1
/
6
1
1
1
9
1
4
6
9
/
1
1
1
7
3
8
o
3
c
2
n
6
_
6
un
/
_
j
0
o
0
c
7
n
8
5
_
un
p
_
d
0
0
b
7
oui
8
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
/
t
.
F
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
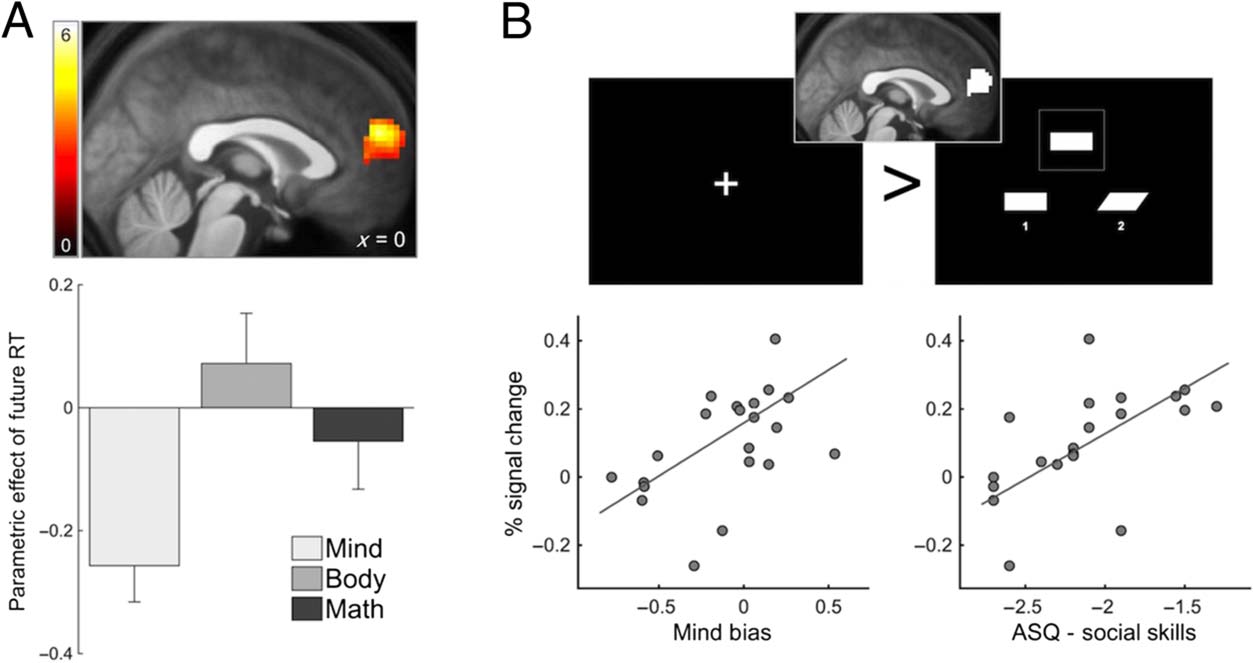
Chiffre 2. (UN) The region of
dmPFC whose response to the
fixation period preceding
accurate mind-focused
judgments was negatively
associated with RT to those
judgments (initially identified
with a cluster-level FWE rate of
0.05 and shown at p < .01
uncorrected to show extent).
Plotted is the region’s mean
parametric effect for the three
conditions. (B) Sample screens
from blocks of the match-to-
sample task that participants
performed after the primary
judgment task. Data from
this task were used to
independently estimate the magnitude of spontaneous resting activity in the dmPFC region (inset) that was found to prime mind-focused judgments
previously. Individual variation in the rest-related response of this region in the match-to-sample task was predicted by two individual measures
tied to the intentional stance: mind bias, a general bias to respond more efficiently to mind-focused compared with body-focused judgments,
and social skills, measured with the ASQ.
First, the autism spectrum quotient (ASQ) is a 50-item
scale designed to measure behaviors and preferences
associated with autism spectrum disorders (Baron-Cohen,
Wheelwright, Skinner, Martin, & Clubley, 2001). Although
this study’s participants completed the full scale, our in-
terest was only in the two 10-item subscales directly rele-
vant to social cognition: social skills (α = .70; e.g., “I find it
difficult to work out people’s intentions”) and communi-
cation (α = .24; e.g., “I am often the last to understand the
point of a joke”). Given that responses to the communi-
cation subscale demonstrated poor reliability, they were
not retained for further analysis. Second, participants
completed the 12-Item Daydream Frequency Scale
(DFS) from the Imaginal Process Inventory (Singer &
Antrobus, 1972; α = .83; e.g., “I am the kind of person
whose thoughts often wander”), which has been used in
previous neuroimaging studies to establish the relation-
ship between DMN function and mind wandering (Mason
et al., 2007). Because of participant noncompliance, ASQ
data were available for only 20 participants, whereas DFS
data were available for only 19 participants.
bias (Figure 2). This was achieved by subtracting the
mean RT for accurate body-focused trials from the mean
RT to accurate mind-focused trials and normalizing the
result by their pooled standard deviation. Hence, a posi-
tive mind bias indicates more efficient performance on
mind-focused relative to body-focused trials.
The mean mind bias in the group was −0.11 (SD =
0.34), indicating that, on average, accurate body-focused
RTs were faster than mind-focused RTs. However, there
was considerable interindividual variability (scores
ranged from −0.78 to 0.54), which we capitalized on in
the individual difference analyses. Importantly, this vari-
ability is unlikely because of a general speed–accuracy
tradeoff, as mind bias showed a nonsignificant positive
association with accuracy to mind-focused judgments (r =
.33) and a nonsignificant negative association with accuracy
to body-focused judgments (r = −.20; to address negative
skewness, accuracy scores were Box-Cox transformed for
this analysis). This supports the validity of using this as a
measure of individual differences in the relative ease of
adopting an intentional rather than nonintentional stance
to other human beings.
Behavior Analysis
MATLAB was used to compute performance on both
tasks. For the judgment task, response accuracy was near
ceiling for both the mind-focused and body-focused con-
ditions (results presented below). Therefore, our behav-
ioral analysis focused on RT to accurate trials as a
measure of processing efficiency. To eliminate the influ-
ence of outliers, we removed trials to which RT deviated
from the mean by 3 SDs (the cutoff was computed for
each trial separately using a leave-one-out procedure).
Then, for each participant, we computed a measure of
the difference in processing efficiency for mind-focused
compared with body-focused trials, which we term mind
Image Acquisition
Imaging data were acquired using a Siemens Trio 3.0-T MRI
scanner (Erlangen, Germany) at the UCLA Ahmanson-
Lovelace Brainmapping Center. For each participant, we
acquired 590 functional T2*-weighted EPI volumes (slice
thickness = 3 mm, gap = 1 mm, 36 slices, repetition time
[TR] = 2000 msec, echo time [TE] = 25 msec, flip angle =
90°, matrix = 64 × 64, field of view = 200 mm). The judg-
ment task was performed in two runs (each acquiring
230 volumes). The match-to-sample task was performed
in a single run (130 volumes). We also acquired a T2-
weighted matched-bandwidth anatomical scan (same
Spunt, Meyer, and Lieberman
1119
D
o
w
n
l
o
a
d
e
d
f
r
o
m
l
l
/
/
/
/
j
t
t
f
/
i
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
i
n
t
o
p
a
r
d
c
e
.
d
s
f
i
r
o
l
m
v
e
h
r
c
p
h
a
d
i
i
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
u
c
n
o
/
c
a
n
r
a
t
r
i
t
i
c
c
l
e
e
-
p
-
d
p
d
2
f
7
/
6
2
7
1
/
1
6
1
/
6
1
1
1
9
1
4
6
9
/
1
1
1
7
3
8
o
3
c
2
n
6
_
6
a
/
_
j
0
o
0
c
7
n
8
5
_
a
p
_
d
0
0
b
7
y
8
g
5
u
.
e
p
s
t
d
o
f
n
b
0
y
8
S
M
e
I
p
T
e
m
L
i
b
b
e
r
r
a
2
r
0
2
i
3
e
s
/
j
f
.
/
t
u
s
e
r
o
n
1
7
M
a
y
2
0
2
1
parameters as EPIs, except TR = 5000 msec, TE = 34 msec,
flip angle = 90°, matrix = 128 × 128) and a T1-weighted
magnetization-prepared rapid-acquisition gradient echo
anatomical scan (slice thickness = 1 mm, 176 slices, TR =
2530 msec, TE = 3.31 msec, flip angle = 7°, matrix = 256 ×
256, field of view = 256 mm).
Image Analysis
Functional data were analyzed using SPM (SPM8; Wellcome
Department of Cognitive Neurology, London, U.K.) ope-
rating in MATLAB. Before statistical analysis, each partici-
pant’s EPI volumes were subjected to the following
preprocessing steps: (1) EPI volumes were corrected for
slice-timing differences; (2) within each run, each EPI
volume was realigned to the first EPI volume of the run;
(3) the T1 structural volume was coregistered to the EPI
time series by initially registering the T2 structural volume
to the mean EPI and then registering the T1 to the T2; (4)
the group-wise DARTEL registration method included in
SPM8 (Ashburner, 2007) was used to normalize the T1
structural volume to a common group-specific space (with
subsequent affine registration to Montreal Neurological
Institute [MNI] space); and (5) normalization of all EPI
volumes to MNI space using the deformation flow fields
generated in the previous step, which simultaneously
resampled volumes (3 mm isotropic) and applied spatial
smoothing (Gaussian kernel of 8 mm, FWHM).
Single-participant Contrast Estimation
A general linear model was used to estimate the effects of
interest for each task. We defined three such models, one
for the match-to-sample task and two for the judgment
task. All models used the canonical (double-gamma) hemo-
dynamic response function for convolution and modeled
serial correlations as an AR(1) process. Moreover, as co-
variates of no interest, all models included the six motion
parameters from image realignment as well as regressors
modeling time points where in-brain global signal change
exceeded 2.5 SDs of the mean global signal change or
where estimated motion exceeded 0.5-mm translation or
0.5° rotation (cutoffs were computed for each time point
separately after excluding the time point from the distribu-
tion). Finally, high-pass filtering was applied using a cutoff
period of 100 sec.
The match-to-sample task was modeled using a single
fixed-epoch regressor modeling shape matching blocks.
The first judgment task model was set up to allow the
simple comparison of the task-evoked activity when par-
ticipants responded accurately to each of the three judg-
ment conditions. For each condition, a variable epoch
model was used (Grinband, Wager, Lindquist, Ferrera,
& Hirsch, 2008) with the epoch for each trial spanning
stimulus onset to participant response. Additional covar-
iates of no interest included regressors modeling inaccu-
rate and no-response trials.
The second judgment task model was set up to test the
hypothesis that, during the course of task performance,
DMN activation to the resting period preceding each trial
is predictive of the ease with which participants make ac-
curate mind-focused (but not body-focused) judgments
about people. In the description to follow, the term pre-
trial response (PTR) will be used to refer to the evoked
response to the offset of the trial that precedes a given
trial. In other words, the PTR models the brain’s response
to the onset of the fixation baseline period that divided
the offset and onset of sequential trials. We modeled the
PTR for each condition separately using an impulse func-
tion placed at the onset of the fixation period. Next, we
modulated the amplitude of the evoked PTR by RT to the
next trial. We omitted PTRs for trials featuring outlier RTs
(criteria described above) and removed variance in the
RT parameter explained by a binary variable coding
whether the accurate response to each trial was to accept
or reject the statement paired with the photograph. To
constrain interpretation of the PTR × RT parametric re-
gressors, multiple regressors of no interest were included
in the model: (1) the unmodulated (i.e., time-invariant)
response to the PTR for each condition, (2) the PTR for
each condition modulated by the duration of the pretrial
interval, (3) the PTR for each condition modulated by a
binary variable indexing whether the preceding trial was
from the same condition, and (4) the variable epoch re-
sponse to the trials themselves (modeled separately for
each condition). To additionally minimize the influence
of task-evoked effects, we estimated this model on the
residuals from the first judgment task model (described
above).1
Group-level Analysis
Except for the ROI analysis described below, all group-
level effects were investigated by subjecting participants’
contrast images for the effects of interest into one-sample
t tests. To test the conjunction null, a minimum statistic
image (Nichols, Brett, Andersson, Wager, & Poline, 2005)
was computed from the mind-focused > body-focused
and rest > math statistical images produced by these
one-sample t tests.
All analyses were interrogated using a cluster-level
family-wise error (FWE) taux de 0.05 with a cluster-forming
voxel-level p value of .001 (uncorrected). Regions of
activation were labeled based on a combination of visual
comparison to functional regions identified in existing
meta-analyses (Denny, Kober, Wager, & Ochsner, 2012;
Mar, 2011; Caspers, Zilles, Laird, & Eickhoff, 2010;
Lieberman, 2010; Carrington & Bailey, 2009; Van Overwalle
& Baetens, 2009) and by reference to probabilistic cyto-
architectonic maps of the human brain using the SPM
anatomy toolbox (Eickhoff et al., 2005). For visual pre-
phrase, thresholded t statistic maps were overlaid on
the average of the participants’ T1-weighted anatomical
images.
1120
Journal des neurosciences cognitives
Volume 27, Nombre 6
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
6
2
7
1
/
1
6
1
/
6
1
1
1
9
1
4
6
9
/
1
1
1
7
3
8
o
3
c
2
n
6
_
6
un
/
_
j
0
o
0
c
7
n
8
5
_
un
p
_
d
0
0
b
7
oui
8
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
F
/
t
.
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Definition of dmPFC ROI
The dmPFC ROI used in the individual difference analy-
ses was defined using the cluster observed in the group-
level parametric effect of pretrial activity on RTs to correct
mind-focused judgments (Figure 2B; peak t = 5.707; x =
3, y = 51, z = 21). Given that this analysis was conducted
within a mask of regions showing the conjunction effect
in the first neuroimaging analysis, this ROI necessarily
overlaps with both the task-negative effect observed in
the rest > math contrast and the task-positive effect
observed in the mind-focused > body-focused contrast.
To account for interindividual variability in the anatomical
locus of the estimated dmPFC response, the ROI was
defined using an uncorrected threshold of p < .01. The
resulting 136-voxel ROI was used to extract data from
the match-to-sample task in all participants.
RESULTS
Performance Results
Mean accuracy and RT for each condition are shown in
Table 1. For the two conditions demanding social judg-
ments, response accuracy was high (mind-focused: M =
96.79%, SD = 5.25%; body-focused: M = 96.19%, SD =
3.32%) and did not significantly differ by condition,
t(20) = 0.446, p = .66. Similarly, RT to correct trials
(mind-focused: M = 2.02 sec, SD = 0.30 sec; body-focused:
M = 1.96 sec, SD = 0.27 sec) did not significantly differ by
condition, t(20) = 1.479, p = .16.
Within-subject Neuroimaging Results
To confirm that the brain regions associated with the
intentional stance were also associated with the DMN, we
tested the conjunction (minimum statistic) of two whole-
brain contrasts: mind-focused compared with body-focused
judgments and fixation baseline (i.e., rest) compared
with math trials. Consistent with published meta-analyses
(Schilbach et al., 2012; Spreng et al., 2009), this revealed
common functional responses in the dmPFC and ventro-
medial PFC, the TPJ bilaterally, the anterior STS, and the
precuneus/posterior cingulate cortex (Table 2). The medial
and transverse slices in Figure 1B show widespread, dis-
tributed correspondence between these two ostensibly
unrelated contrasts. The overlap spans the major nodes of
Table 1. Performance Results for the Three Conditions in the
Primary Judgment Task (N = 21)
Judgment Condition
Mind-focused
Body-focused
Math
Table 2. Peak Coordinates from Significant Clusters Observed
When Testing against the Conjunction Null for the Contrasts
Mind-focused > Body-focused and Rest > Math (N = 21,
Whole-brain Search with a Cluster-level FWE Rate of 0.05)
Region Name
L/R Extent
t
Coordonnées INM
X
−9
18
−3
oui
57
42
z
30
48
54 −12
750
7.041
–
–
5.460
5.243
168
7.408 −51 −66
96
6.175
54 −63
30
33
102
120
6.095 −60
−6 −18
5.328
−6 −51
36
Dorsomedial PFC
Ventromedial PFC
TPJ
Anterior STS
Precuneus/PCC
L
R.
L
L
R.
L
L
Coordinates are all local maxima observed, which were separated by at
least 20 mm. X, oui, and z are MNI coordinates in the left–right, anterior–
posterior, and inferior–superior dimensions, respectivement. PCC = poste-
rior cingulate cortex.
both networks in the medial frontoparietal, temporoparietal,
and anterior temporal cortices.
The anatomical correspondence of the two cognitive
states suggests that DMN activity during rest may prime
the intentional stance to social stimuli. If the DMN activ-
ity during rest primes the intentional stance, we should
observe that, as the magnitude of its pretrial response in-
creases, the time it takes to produce a correct response on
subsequent mind-focused trials should decrease. When re-
stricting the search to the regions of overlap identified in
the previous analysis, we observed such an effect in one
area of the DMN, the dmPFC (Figure 2A; peak: t = 5.71,
x = 3, y = 51, z = 21; extent = 50 voxels).2 The region
of dmPFC identified is anatomically similar to those ob-
served in numerous neuroimaging studies highlighting
the importance of the dmPFC to mental state inference
(Amodio & Frith, 2006; Gallagher & Frith, 2003; Happé
et coll., 1996; Fletcher et al., 1995; Goel et al., 1995). As is
evident in the plot shown in Figure 2A, this priming effect
is specific to mind-focused trials in our study. En fait, le
priming effect for mind-focused judgments was significantly
stronger than the same effect estimated for body-focused
judgments, which featured the same set of social stimuli
(peak: t = 5.91, x = 3, y = 54, z = 21; extent = 34 voxels).
Enfin, no regions within the overlap were found to
exhibit a significant priming effect on RTs to either body-
focused or math-focused trials, and in all three conditions,
there were no regions that showed an antipriming effect,
c'est, pretrial activity that positively correlated with RTs.
Measure
Mean
SD
Mean
SD
Mean
SD
Between-subject Neuroimaging Results
Accuracy (%)
96.79
RT (sec)
2.02
5.25
0.31
96.19
3.32
94.05
4.90
1.96
0.27
1.85
0.32
The evidence so far demonstrates that, within the same
individual, transient changes in spontaneous dmPFC
activity over time prime more efficient responses to
Spunt, Meyer, and Lieberman
1121
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
6
2
7
1
/
1
6
1
/
6
1
1
1
9
1
4
6
9
/
1
1
1
7
3
8
o
3
c
2
n
6
_
6
un
/
_
j
0
o
0
c
7
n
8
5
_
un
p
_
d
0
0
b
7
oui
8
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
F
t
/
.
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
judgments requiring the intentional stance. To corrobo-
rate this transient priming effect, we examined the extent
to which individual differences in rest-related responses
during the match-to-sample scan can be predicted by
individual differences in our measure of mind bias, que
est, the relative speed with which participants executed
accurate mind-focused and body-focused judgments,
averaged across trials. We examined the extent to which
this measure could predict individual variation in the
amplitude of the rest-related activity in dmPFC ROI
defined based on the priming effect observed above.
In line with the results presented so far, variation in
baseline activity in dmPFC was positively predicted by var-
iation in mind bias (r19 = .50, p = .021, 95% CIbootstrapped
[0.21, 0.70]; Figure 2D). De plus, this relationship is
robust when controlling for individual differences in
both performance on the match-to-sample task and self-
reported mind wandering as measured using the DFS
(rpartial = .51, p = .039). En outre, the same individuals
who exhibited greater dmPFC activity during rest also
scored higher on a self-report measure of the social skills
that are commonly impaired in individuals with an autism
spectrum disorder (r18 = .57, p = .009, 95% CIbootstrapped
[0.15, 0.78]; Figure 2D), and this relationship also remains
after controlling for match-to-sample task performance
and DFS scores (rpartial = .52, p = .032). Ainsi, individuals
who exhibited greater activity in dmPFC while at rest (com-
pared with while performing a speeded match-to-sample
task) showed a general processing advantage for adopting
an intentional (rather than nonintentional) stance to peo-
ple and reported having higher levels of everyday social
expertise.
DISCUSSION
Taken together, the findings reported here suggest that
the default mode of human brain function, perhaps cen-
tralized to the dmPFC, primes the intentional stance to
social stimuli. Just as the word “face” primes people to
initially see the Ruben’s illusion as faces rather than a
vase, spontaneous DMN activity before a social interac-
tion may prime the mind to treat others as minds rather
than simply bodies extended in space. Drawing on a psy-
chological theory and method on priming, we reasoned
que, if spontaneous DMN activity features mental opera-
tions that are utilized when adopting the intentional
position, DMN activity should make it easier to adopt the
intentional stance in the event that another person is en-
countered. We found evidence that variability in sponta-
neous dmPFC activity both within and across participants
has a priming-like effect that is selective for mind-focused
judgments of other people. We offer this as strong evi-
dence that DMN activity in between moments of cogni-
tive activity is the biological basis for the powerful human
tendency to adopt the intentional stance.
This study was motivated by an observation that has
now been made many times before in the literature:
The functional neuroanatomy of mental-state reasoning
and the resting state are remarkably similar (Mars et al.,
2012; Spreng et al., 2009; Schilbach et al., 2008; Buckner
& Carroll, 2007). Encore, to our knowledge, this is the first
study to identify the widespread neuroanatomical overlap
of the two networks in the same set of participants and
using the time series of brain activity measured in a single
behavioral task. This allowed us to demonstrate that the
very same voxels that show a task-negative effect (deactiva-
tion to math judgments) can also show a task-positive effect
(activation to mind-focused judgments). This compellingly
highlights the fallacy implied by labeling the DMN a
“task-negative” network (Meyer, Spunt, Berkman, Taylor,
& Lieberman, 2012; Spreng, 2012). Whether regions of the
DMN show “task-negative” or “task-positive” effects depends
on cognitive requirements of the task at hand.
Le plus important, this is the first study to provide direct
evidence that stimulus-independent activity in DMN re-
gions is functionally consequential for the execution of
stimulus-dependent mental state inferences. This estab-
lishes the mechanism by which individual differences in
resting baseline activation (Kennedy, Redcay, & Courchesne,
2006) and connectivity (Li, Peut, & Liu, 2014) would be
associated with variability in both typical and atypical social
functioning. De plus, it suggests that the early maturation
of the DMN may be functionally critical in early develop-
ment, providing children with a “jump start” on acquiring
the psychological skills necessary for understanding a
complex and heterogeneous social world.
Bien sûr, these findings should not be taken to imply
that social cognition is the only domain in which the
DMN makes a functional contribution. En fait, the DMN
can be functionally divided into at least two subsystems
(Andrews-Hanna et al., 2014; Andrews-Hanna, Reidler,
Sepulcre, Poulin, & Buckner, 2010). The first subsystem
is primarily localized to medial temporal lobe (MTL)
structures, whereas the second, termed the dmPFC sub-
système, includes the TPJ, lateral and polar temporal cortex,
and an area of the dmPFC that is anatomically consistent
with the region of dmPFC highlighted by this study. Le
MTL subsystem is not reliably observed in studies of men-
tal state reasoning; hence, we had no strong reason to hy-
pothesize either an anatomical or functional relationship
of this system with mind-focused judgments. Bien sûr,
default activity in the MTL subsystem likely does serve
adaptive functions, par exemple, in the consolidation of
long-term memories (Wig et al., 2008).
Given that the DMN activity is metabollically costly,
widely distributed in the cortex, and highly sensitive to
both the presence and type of task demand, it should
be no surprise that this network would have functional
consequences in multiple domains. A related but wholly
separate question regards the reasons why the DMN
evolved in the first place. Evidence suggests that the
DMN is a basic and phylogenetically old feature of human
cortical function: The basic elements of the DMN can be
observed in human neonates (Fransson et al., 2007), et
1122
Journal des neurosciences cognitives
Volume 27, Nombre 6
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
6
2
7
1
/
1
6
1
/
6
1
1
1
9
1
4
6
9
/
1
1
1
7
3
8
o
3
c
2
n
6
_
6
un
/
_
j
0
o
0
c
7
n
8
5
_
un
p
_
d
0
0
b
7
oui
8
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
/
F
t
.
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
similar default networks have been observed in chimpan-
zees (Rilling et al., 2007) and monkeys ( Vincent et al.,
2007). In light of these observations, we suggest our re-
sults converge with theories proposing that primate intel-
ligence evolved as a response to the enormous demands
imposed on the brain by living in increasingly large and
complex social groups (Sallet et al., 2011; Cheney &
Seyfarth, 2008; Dunbar, 1998). The data we present here
suggest that the DMN and its activity in between mo-
ments of directed thought may be evolution’s solution
to the problem of other minds. Evolution seems to have
made a “bet” that the best thing to do with any spare
moment is to get ready to see the world in terms of other
minds. This bet has allowed human beings to get
together in groups and achieve far more than ever would
have been possible separately.
Remerciements
We acknowledge Catherine Mulvenna for help with data collec-
tion, and Uta Frith, Nathan Spreng, Lucina Uddin, and Jamil
Zaki for helpful comments on an earlier version of the paper.
Reprint requests should be sent to Matthew D. Lieberman, De-
partment of Psychology, 1285 Franz Hall, UCLA, Les anges,
Californie 90095-1563, ou par e-mail: lieber@ucla.edu.
Remarques
1. We note that the PTR effects reported in the main text are
also observed when estimating them on the nonresidualized
time series with a model including the task-related effects.
2. This region is also observed in the whole-brain analysis
(peak: t = 6.34, x = 6, y = 48, z = 21; extent = 83 voxels).
Cependant, given that our analysis was specifically designed to
investigate the functional implications of the neuroanatomical
overlap of the resting state and the intentional stance, all re-
maining analyses restrict the search to the mask of regions
showing evidence of such overlap in this study (see Methods
for further details).
RÉFÉRENCES
Amodio, D., & Frith, C. (2006). Meeting of minds: The medial
frontal cortex and social cognition. Nature Reviews
Neurosciences, 7, 268–277.
Andrews-Hanna, J., Reidler, J., Sepulcre, J., Poulin, R., &
Buckner, R.. (2010). Functional–anatomic fractionation of the
brain’s default network. Neurone, 65, 550–562.
Andrews-Hanna, J., Smallwood, J., & Spreng, R.. (2014). Le
default network and self-generated thought: Component
processes, dynamic control, and clinical relevance. Annales de
the New York Academy of Sciences, 1316, 29–52.
Arieli, UN., Sterkin, UN., Grinvald, UN., & Aertsen, UN. (1996). Dynamics
of ongoing activity: Explanation of the large variability in
evoked cortical responses. Science, 273, 1868–1871.
Ashburner, J.. (2007). A fast diffeomorphic image registration
algorithme. Neuroimage, 38, 95–113.
Attwell, D., & Laughlin, S. B. (2001). An energy budget for
signaling in the grey matter of the brain. Journal of Cerebral
Blood Flow & Metabolism, 21, 1133–1145.
Baron-Cohen, S., Wheelwright, S., Skinner, R., Martine, J., &
Clubley, E. (2001). The autism-spectrum quotient (AQ):
Evidence from Asperger syndrome/high-functioning autism,
males and females, scientists and mathematicians. Journal de
Autism Developmental Disorders, 31, 5–17.
Binder, J., Frost, J., Hammeke, T., Bellgowan, P., Rao, S., & Cox,
R.. (1999). Conceptual processing during the conscious
resting state. A functional MRI study. Journal of Cognitive
Neurosciences, 11, 80–95.
Brainard, D. H. (1997). The psychophysics toolbox. Spatial
Vision, 10, 433–436.
Buckner, R., & Carroll, D. (2007). Self-projection and the brain.
Tendances des sciences cognitives, 11, 49–57.
Button, K., Ioannidis, J., Mokrysz, C., Nosek, B., Flint, J.,
Robinson, E., et autres. (2013). Power failure: Why small sample
size undermines the reliability of neuroscience. Nature
Reviews Neuroscience, 14, 365–376.
Callard, F., & Margulies, D. S. (2014). What we talk about when
we talk about the default mode network. Frontiers in
Human Neuroscience, 8, 619.
Carrington, S., & Bailey, UN. (2009). Are there theory of mind
regions in the brain? A review of the neuroimaging literature.
Human Brain Mapping, 30, 2313–2335.
Caspers, S., Zilles, K., Laird, UN., & Eickhoff, S. (2010). ALE meta-
analysis of action observation and imitation in the human
brain. Neuroimage, 50, 1148–1167.
Cheney, D. L., & Seyfarth, R.. M.. (2008). Baboon metaphysics:
The evolution of a social mind (1st ed.). Chicago: University
of Chicago Press.
Dennett, D. C. (1989). The intentional stance. Cambridge, MA:
AVEC Presse.
Denny, B., Kober, H., Wager, T., & Ochsner, K. (2012). A meta-
analysis of functional neuroimaging studies of self- et autre
judgments reveals a spatial gradient for mentalizing in medial
prefrontal cortex. Journal des neurosciences cognitives, 24,
1742–1752.
Dunbar, R.. je. M.. (1998). The social brain hypothesis.
Evolutionary Anthropology, 9, 178–190.
Eickhoff, S., Stephan, K., Mohlberg, H., Grefkes, C., Fink, G.,
Amunts, K., et autres. (2005). A new SPM toolbox for combining
probabilistic cytoarchitectonic maps and functional imaging
data. Neuroimage, 25, 1325–1335.
Epley, N., Waytz, UN., & Cacioppo, J.. (2007). On seeing human: UN
three-factor theory of anthropomorphism. Psychological
Review, 114, 864–886.
Fletcher, P.. C., Happe, F., Frith, U., Boulanger, S. C., Dolan, R.. J.,
Frackowiak, R.. S. J., et autres. (1995). Other minds in the brain: UN
functional imaging study of “theory of mind” in story
comprehension. Cognition, 57, 109–128.
Fox, M., Snyder, UN., Vincent, J., & Raichle, M.. (2007). Intrinsic
fluctuations within cortical systems account for intertrial
variability in human behavior. Neurone, 56, 171–184.
Fox, M., Snyder, UN., Zacks, J., & Raichle, M.. (2006). Coherent
spontaneous activity accounts for trial-to-trial variability in
human evoked brain responses. Neurosciences naturelles, 9,
23–25.
Fransson, P., Skiold, B., Horsch, S., Nordell, UN., Blennow, M.,
Lagercrantz, H., et autres. (2007). Resting-state networks in the
infant brain. Proceedings of the National Academy of
les sciences, USA., 104, 15531–15536.
Gallagher, H. L., & Frith, C. D. (2003). Functional imaging
of “theory of mind.” Trends in Cognitive Sciences, 7, 77–83.
Goel, V., Grafman, J., Sadato, N., & Hallett, M.. (1995). Modeling
other minds. NeuroReport, 6, 1741–1746.
Greicius, M., Krasnow, B., Reiss, UN., & Menon, V. (2003).
Functional connectivity in the resting brain: A network
analysis of the default mode hypothesis. Actes du
Académie nationale des sciences, USA., 100, 253–258.
Grinband, J., Wager, T., Lindquist, M., Ferrera, V., & Hirsch, J..
(2008). Detection of time-varying signals in event-related
fMRI designs. Neuroimage, 43, 509–520.
Spunt, Meyer, and Lieberman
1123
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
t
t
F
/
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
6
2
7
1
/
1
6
1
/
6
1
1
1
9
1
4
6
9
/
1
1
1
7
3
8
o
3
c
2
n
6
_
6
un
/
_
j
0
o
0
c
7
n
8
5
_
un
p
_
d
0
0
b
7
oui
8
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
t
/
F
.
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1
Happé, F., Ehlers, S., Fletcher, P., Frith, U., Johansson, M.,
Gillberg, C., et autres. (1996). “Theory of mind” in the brain.
Evidence from a PET scan study of Asperger syndrome.
NeuroReport, 8, 197–201.
Heider, F., & Simmel, M.. (1944). An experimental study of apparent
behavior. The American Journal of Psychology, 57, 243.
Higgins, E. T. (1989). Knowledge accessibility and activation:
Subjectivity and suffering from unconscious sources. In J. S.
Uleman & J.. UN. Bargh (Éd.), Unintended thought ( Vol. 3,
pp. 75–123). New York: Guilford Press.
Hsieh, P., Colas, J., & Kanwisher, N. (2012). Pre-stimulus pattern
of activity in the fusiform face area predicts face percepts
during binocular rivalry. Neuropsychologie, 50, 522–529.
Kennedy, D., Redcay, E., & Courchesne, E. (2006). Failing to
deactivate: Resting functional abnormalities in autism.
Actes de l'Académie nationale des sciences, USA.,
103, 8275–8280.
Kovacs, UN., Teglas, E., & Endress, UN. (2010). The social sense:
Susceptibility to others’ beliefs in human infants and adults.
Science, 330, 1830–1834.
Li, W., Peut, X., & Liu, C. (2014). The default mode network and
social understanding of others: What do brain connectivity
studies tell us. Frontiers in Human Neuroscience, 8, 74.
Lieberman, M.. (2010). Social cognitive neuroscience. In S. T.
Fiske, D. T. Gilbert, & G. Lindzey (Éd.), Handbook of social
psychologie (5th ed., pp. 143–193). New York: McGraw-Hill.
Mar, R.. (2011). The neural bases of social cognition and story
comprehension. Revue annuelle de psychologie, 62, 103–134.
Mars, R., Neubert, F., Noonan, M., Sallet, J., Toni, JE., &
Rushworth, M.. (2012). On the relationship between the
“default mode network” and the “social brain”. Frontiers in
Human Neuroscience, 6, 189.
Mason, M., Norton, M., Van Horn, J., Wegner, D., Grafton, S., &
Macrae, C. (2007). Wandering minds: The default network
and stimulus-independent thought. Science, 315, 393–395.
Mazoyer, B., Zago, L., Mellet, E., Bricogne, S., Etard, O., Houde,
O., et autres. (2001). Cortical networks for working memory and
executive functions sustain the conscious resting state in
man. Brain Research Bulletin, 54, 287–298.
Mesoudi, UN., Whiten, UN., & Dunbar, R.. (2006). A bias for social
information in human cultural transmission. British Journal
of Psychology, 97, 405–423.
Meyer, M.. L., Spunt, R.. P., Berkman, E. T., Taylor, S. E., &
Lieberman, M.. D. (2012). Social working memory: An fMRI
study of parametric increases in social cognitive effort.
Actes de l'Académie nationale des sciences, 109,
1883–1888.
Neely, J.. H. (1977). Semantic priming and retrieval from lexical
mémoire: Roles of inhibitionless spreading activation and
limited-capacity attention. Journal d'expérimentation
Psychologie: General, 106, 226–254.
Nichols, T., Brett, M., Andersson, J., Wager, T., & Poline, J..
(2005). Valid conjunction inference with the minimum
statistique. Neuroimage, 25, 653–660.
Poldrack, R.. (2006). Can cognitive processes be inferred from
neuroimaging data? Tendances des sciences cognitives, 10, 59–63.
Raichle, M.. (2010). Two views of brain function. Trends in
Cognitive Sciences, 14, 180–190.
Raichle, M., Macleod, UN., Snyder, UN., Powers, W., Gusnard, D., &
Shulman, G. (2001). A default mode of brain function.
Actes de l'Académie nationale des sciences, USA.,
98, 676–682.
Rilling, J., Barks, S., Parr, L., Preuss, T., Faber, T., Pagnoni, G.,
et autres. (2007). A comparison of resting-state brain activity in
humans and chimpanzees. Actes de la Nationale
Académie des Sciences, USA., 104, 17146–17151.
Rosset, E. (2008). It’s no accident: Our bias for intentional
explanations. Cognition, 108, 771–780.
Sallet, J., Mars, R., Noonan, M., Andersson, J., O'Reilly, J., Jbabdi,
S., et autres. (2011). Social network size affects neural circuits in
macaques. Science, 334, 697–700.
Saxe, R., Carey, S., & Kanwisher, N. (2004). Understanding other
minds: Linking developmental psychology and functional
neuroimaging. Revue annuelle de psychologie, 55, 87–124.
Schilbach, L., Bzdok, D., Timmermans, B., Fox, P., Laird, UN.,
Vogeley, K., et autres. (2012). Introspective minds: Using ALE
meta-analyses to study commonalities in the neural correlates
of emotional processing, sociale & unconstrained cognition.
PLoS One, 7, e30920.
Schilbach, L., Eickhoff, S., Rotarska-Jagiela, UN., Fink, G., &
Vogeley, K. (2008). Minds at rest? Social cognition as the
default mode of cognizing and its putative relationship to the
“default system” of the brain. Consciousness and Cognition,
17, 457–467.
Senju, UN. (2012). Spontaneous theory of mind and its absence in
autism spectrum disorders. Neuroscientist, 18, 108–113.
Shulman, G., Fiez, J., Corbetta, M., Buckner, R., Miezin, F.,
Raichle, M., et autres. (1997). Common blood flow changes across
visual tasks: II. Decreases in cerebral cortex. Journal de
Neurosciences cognitives, 9, 648–663.
Chanteur, J., & Antrobus, J.. (1972). Daydreaming, imaginal
processes, and personality: A normative study. In P. Sheehan
(Ed.), The function and nature of imagery (1st ed.,
pp. 175–202). New York: Academic Press.
Spreng, R.. (2012). The fallacy of a “task-negative” network.
Frontiers in Psychology, 3, 145.
Spreng, R., Mar, R., & Kim, UN. (2009). The common neural basis
of autobiographical memory, prospection, navigation, théorie
of mind, and the default mode: A quantitative meta-analysis.
Journal des neurosciences cognitives, 21, 489–510.
Spunt, R., & Adolphs, R.. (2014). Validating the why/how
contrast for functional MRI studies of theory of mind.
Neuroimage, 99, 301–311.
Spunt, R., & Lieberman, M.. (2012un). An integrative model of the
neural systems supporting the comprehension of observed
emotional behavior. Neuroimage, 59, 3050–3059.
Spunt, R., & Lieberman, M.. (2012b). Dissociating modality-
specific and supramodal neural systems for action
understanding. Journal des neurosciences, 32, 3575–3583.
Spunt, R., Satpute, UN., & Lieberman, M.. (2011). Identifying the
what, why, and how of an observed action: An fMRI study of
mentalizing and mechanizing during action observation.
Journal des neurosciences cognitives, 23, 63–74.
Tulving, E., & Schacter, D. (1990). Priming and human memory
systèmes. Science, 247, 301–306.
Uleman, J., Adil Saribay, S., & González, C. (2008). Spontaneous
inferences, implicit impressions, and implicit theories.
Revue annuelle de psychologie, 59, 329–360.
Vallacher, R.. R., & Wegner, D. M.. (1987). What do people think
they’re doing? Action identification and human behavior.
Psychological Review, 94, 3–15.
Van Overwalle, F., & Baetens, K. (2009). Understanding others’
actions and goals by mirror and mentalizing systems: A meta-
analyse. Neuroimage, 48, 564–584.
Vincent, J., Patel, G., Fox, M., Snyder, UN., Boulanger, J., Van Essen,
D., et autres. (2007). Intrinsic functional architecture in the
anaesthetized monkey brain. Nature, 447, 83–86.
Wig, G., Grafton, S., Demos, K., Wolford, G., Petersen, S., &
Kelley, W. (2008). Medial temporal lobe bold activity at rest
predicts individual differences in memory ability in healthy
young adults. Proceedings of the National Academy of
les sciences, USA., 105, 18555–18560.
Yarkoni, T. (2009). Big correlations in little studies: Inflated
fMRI correlations reflect low statistical power-commentary on
Vul et al. (2009). Perspectives on Psychological Science, 4,
294–298.
1124
Journal des neurosciences cognitives
Volume 27, Nombre 6
D
o
w
n
je
o
un
d
e
d
F
r
o
m
je
je
/
/
/
/
j
F
/
t
t
je
t
.
:
/
/
h
t
t
p
:
/
D
/
o
m
w
je
n
t
o
p
un
r
d
c
e
.
d
s
F
je
r
o
je
m
v
e
h
r
c
p
h
un
d
je
je
r
r
e
.
c
c
t
.
o
m
m
/
j
e
d
o
toi
c
n
o
/
c
un
n
r
un
t
r
je
t
je
c
c
je
e
e
–
p
–
d
p
d
2
F
7
/
6
2
7
1
/
1
6
1
/
6
1
1
1
9
1
4
6
9
/
1
1
1
7
3
8
o
3
c
2
n
6
_
6
un
/
_
j
0
o
0
c
7
n
8
5
_
un
p
_
d
0
0
b
7
oui
8
g
5
toi
.
e
p
s
t
d
o
F
n
b
0
oui
8
S
M.
e
je
p
T
e
m
L
je
b
b
e
r
r
un
2
r
0
2
je
3
e
s
/
j
t
/
.
F
toi
s
e
r
o
n
1
7
M.
un
oui
2
0
2
1