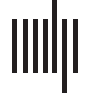REVIEW
Atypical core-periphery brain dynamics in autism
Dipanjan Roy
1
and Lucina Q. Uddin
2
1Cognitive Brain Dynamics Lab, National Brain Research Centre, Manesar, India
2Département de psychologie, University of Miami, Coral Gables, FL, Etats-Unis
Mots clés: Core-periphery dynamics, Atypical timescales, Caudate, Core and contextual symptom
severity, Sensory-motor network, Restricted and repetitive behaviors
ABSTRAIT
The intrinsic function of the human brain is dynamic, giving rise to numerous behavioral
subtypes that fluctuate distinctively at multiple timescales. One of the key dynamical
processes that takes place in the brain is the interaction between core-periphery brain
régions, which undergoes constant fluctuations associated with developmental time frames.
Core-periphery dynamical changes associated with macroscale brain network dynamics span
multiple timescales and may lead to atypical behavior and clinical symptoms. Par exemple,
recent evidence suggests that brain regions with shorter intrinsic timescales are located at the
periphery of brain networks (par exemple., sensorimotor hand, face areas) and are implicated in
perception and movement. On the contrary, brain regions with longer timescales are core
hub regions. These hubs are important for regulating interactions between the brain and the
body during self-related cognition and emotion. Dans cette revue, we summarize a large body of
converging evidence derived from time-resolved fMRI studies in autism to characterize
atypical core-periphery brain dynamics and how they relate to core and contextual sensory
and cognitive profiles.
INTRODUCTION
Sensory Processing in Autism
Perhaps the most remarkable feature of autism spectrum disorder (ASD) is profound behavioral
diversity across different individuals, which pertains to all factors involved in interactions with
the physical and social environment (Baron-Cohen, Ashwin, Ashwin, Tavassoli, & Chakrabarti,
2009; Blakemore, Burnett, & Dahl, 2010; Bolton, Morgenroth, Preti, & Van De Ville, 2020;
Lawson et al., 2015; Robertson & Baron-Cohen, 2017; Shafritz, Dichter, Baranek, & Belger,
2008; Uddin, 2021). This diversity underlies variability in personality, physiology, and mental
capacity, which in turn are sculpted by not only complex biological influences (par exemple., med-
ication, genetic and epigenetic factors) but also various sociocultural factors (par exemple., multilin-
gual environments, social learning, trauma; Baron-Cohen et al., 2009; Baum et al., 2017;
Bolton et al., 2020; Robertson & Baron-Cohen, 2017; Uddin, 2021). Developmental re-
search suggests that sensory symptoms manifest early in life, and contribute unique variance
to the diagnostic criteria of autism (Andreae, 2019; Chen, Nomi, Uddin, Duan, & Chen, 2017;
Ciarrusta et al., 2019; Courchesne, Campbell, & Solso, 2011; Uddin, 2021). Neuroimaging
evidence suggests that sensory symptoms originate from differences in low-level processing
in sensory-dedicated regions in the autistic brain, and offers insight into circuit-level alter-
ations (Abbott et al., 2018; Alaerts et al., 2015; Alaerts, Swinnen, & Wenderoth, 2016; Alaerts
et autres. 2014; Anderson et al., 2011; Baum et al., 2017; Collignon et al., 2013; Courchesne et al.,
2011; Uddin, 2021). Studying the brain at rest has demonstrated that although the environment
un accès ouvert
journal
Citation: Roy, D., & Uddin, L. Q. (2021).
Atypical core-periphery brain dynamics
in autism. Neurosciences en réseau, 5(2),
295–321. https://est ce que je.org/10.1162
/netn_a_00181
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00181
Reçu: 2 Septembre 2020
Accepté: 31 Décembre 2020
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Dipanjan Roy
dipanjan.nbrc@gov.in
Éditeur de manipulation:
Olaf Sporns
droits d'auteur: © 2021
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
un
_
0
0
1
8
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
has an influence, the brain operates intrinsically and is modulated by, rather than controlled by,
the external world (Baum et al., 2017; Bolton et al., 2020; Liégeois et al., 2019; Lin et al., 2016;
Liu, Liao, Xia, & Il, 2018). This modulation is a recursive process between the brain and the
environment mediated by perception and action (Friston, 2009; Kiebel, Daunizeau, & Friston,
2008). This process is highly dynamic, as are both the environment and the brain (Bolton et al.,
2020; Friston, 2009; Huang et al., 2015).
Individuals with severe autism usually have intellectual impairments and develop little spo-
ken language. There are subgroups of autistic individuals who may have average or above-
average IQ, but who still struggle with more subtle aspects of communication, such as body
langue (Jasmin et al., 2019; Ostrolenk, Bao, Mottron, Collignon, & Bertone, 2019; Robertson
& Baron-Cohen, 2017; Supekar et al., 2013; Uddin, 2021; Uddin et al., 2015). In addition to
social difficulties, many individuals with autism show restricted and repetitive behaviors (RRB)
and sensory abnormalities (SA), and have narrow interests (Huang et al., 2015; Jao Keehn et al.,
2019; Jao Keehn et al., 2017; Jasmin et al., 2019; Kana, Keller, Minshew, & Just, 2007;
Manning, Tibber, Charman, Dakin, & 2015; Mash, Reiter, Linke, Townsend, & Müller, 2018;
McKinnon et al., 2019; Mottron, Belleville, Rouleau, & Collignon, 2014; Moul, Cauchi, Hawes,
Brennan, & Dadds, 2015; Robertson & Baron-Cohen, 2017; Uddin, 2021).
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
un
_
0
0
1
8
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The brain as a whole shows less coordinated activity in autism, and one way of classifying
subtypes that can include hyper- and hyporeactivity to sensory environments or unusual inter-
est in sensory aspects of the environment could be to use brain network-based classification
méthodes (Harlalka, Bapi, Vinod, & Roy, 2018, 2019; Nomi, Bolt, Ezie, Uddin, & Heller, 2017;
Uddin et al., 2013). There is a growing body of evidence further suggesting that individual brain
regions work in a less cohesive manner in autism, with widely distributed timescales and hi-
erarchical organization of brain networks (Atasoy, Donnelly, & Pearson, 2016; Chaudhuri,
Knoblauch, Gariel, Kennedy, & Wang, 2015; Gollo, Roberts, & Cocchi, 2017; Gollo, Zalesky,
Hutchison, van den Heuvel, & Breakspear, 2015; Harlalka et al., 2019; Hasson, Lequel, Vallines,
Heeger, & Rubin, 2008; Hong et al., 2019; Kumar et al., 2016; Lin et al., 2016; Londres, 2018;
Nomi, Bolt, et coll., 2017; Nomi & Uddin, 2015; Nomi, Vij, et coll., 2017; Oldham & Fornito,
2019; Pillai & Jirsa, 2017; Preti & Van De Ville, 2019; Raut, Snyder, & Raichle, 2020; P.. Wang
et coll., 2019; S. Wang et al., 2015; Watanabe & Rees, 2017; Watanabe, Rees, & Masuda,
2019). Here we use the terms core and periphery (outside the core) brain regions when we
refer to atypical timescales, flexibility, cohesion, dispersion, and functional gradients based on
their hierarchical organization and differences in network dynamics (Chaudhuri et al., 2015;
Gollo, 2019; Gollo et al., 2017; Gollo et al., 2015; Hasson et al., 2008; Hong et al., 2019;
van den Heuvel, Kahn, Goñi, & Sporns, 2012; P.. Wang et al., 2019; S. Wang et al., 2015).
Specifically, studies have suggested a distinction between a network periphery containing sen-
sory and motor regions with more locally clustered connectivity, and a rich-club “core” that ag-
gregates long-range connections and serves as a backbone for transmodal integration, giving rise
to behavior and cognition (Deco, Kringelbach, Jirsa, & Ritter, 2017; Gollo, 2019; Gollo et al.,
2017; Gollo et al., 2015; Griffa & van den Heuvel, 2018; Harlalka et al., 2019; Hasson et al.,
2008; Hilgetag & Goulas, 2020; Hong et al., 2019; Lin et al., 2016; Rashid et al., 2018;
Shafritz et al., 2008; P.. Wang et al., 2019; Watanabe & Rees, 2017).
Perturbation of resting-state brain dynamics and distortion of
timescales of sensory-
processing regions in individuals with autism compared with that of people without autism
may shed light on core and contextual neural processing and links with symptom severity in
the disorder (Andreae, 2019; Baron-Cohen et al., 2009; Cerliani et al., 2015; Foxe et al., 2015;
Harlalka et al., 2019; Hasson et al., 2008; Henry, Dichter, & Gates, 2018; Jao Keehn et al.,
296
Network core:
A set of densely connected brain
regions that aggregate long-range
connectivity and serve as a backbone
for polymodal integration.
Network periphery:
Brain networks primarily consisting
of sensory and motor regions, avec
more locally clustered connectivity.
Flexibility:
Flexibility is a network metric that
characterizes the modular changes in
each brain area throughout the scan
period. Cependant, this dynamic
measure does not capture
community affiliation.
Cohesion strength:
In a network of brain areas, node
strength is estimated as a cohesion
matrix where the edge weights of the
network denote the number of times
a pair of nodes changes to the
identical community affiliation.
Rich-club nodes:
Strongly connected nodes in a
réseau. If there is a core of such
nodes with a high node degree that
receives a very large proportion of
connections and that are more
densely interconnected among
themselves than lower degree nodes
in the network, they form a rich club.
Autrement dit, the high-degree
nodes are densely connected hubs
in the brain networks and form an
exclusive club.
Transmodal cortex:
This overarching system is thought
to facilitate abstract, higher order
cognitive functions by segregating
information processing of the
sensory environment from more
self-generated and internally oriented
cognition emerging in transmodal,
integrative cortices.
Neurosciences en réseau
Core-periphery brain network dynamics in autism
2019; Jao Keehn et al., 2017; Jasmin et al., 2019; Mash et al., 2018; McKinnon et al., 2019;
Mottron et al., 2014; Moul et al., 2015). Recent functional connectivity studies in autism
further highlight two classes of relationships between functional connectivity among various
brain regions during core (state invariant) versus contextual (state-dependent) neural process-
ing (Jasmin et al., 2019; Robertson & Baron-Cohen, 2017).
Recent evidence further suggests that genetic mutations trigger brain reorganization in in-
dividuals with a low plasticity threshold in autism, mostly within sensory and extrasensory
regions sensitive to cortical reallocations (Cheng, Rolls, Gu, Zhang, & Feng, 2015; Hahamy,
Behrmann, & Malach, 2015; Mottron et al., 2014). These changes may account for the cog-
nitive enhancements and reduced social expertise associated with autism (Mottron et al.,
2014). Enhanced but normal plasticity may underlie non-syndromic autism, whereas syn-
dromic autism may occur when a triggering mutation or event produces an altered plastic
reaction, also resulting in intellectual disability and dysmorphism in addition to autism. Dif-
language regions) account for
ferences in the target of brain reorganization (perceptual vs.
the main differences linking neocortical, cognitive, and genetic variability in autism that was
proposed earlier as the trigger-threshold-target (TTT) model (Mottron et al., 2014). According
to this model, the regions that are the most susceptible to reorganization in autism (the multi-
modal association regions) are also those that have the largest variability in terms of connec-
tivité (structural and functional) among typical developing individuals (Figure 1A). The highest
interindividual differences in resting-state connectivity are in the multimodal association cor-
texand the lowest are in the unimodal sensory and motor cortices (as displayed in Figures 1A
and 1E). En outre, the same unimodal sensory regions, particularly lateral occipital cortex
(LOC), exhibit greater gyrification index and volumetry in autistic individuals compared with
typically developing individuals (as shown in Figures 1B and 1F). The same set of brain re-
gions exhibits enhanced resting-state functional connectivity differences between autistic and
typically developing individuals (Figure 1C) and high MEG connectivity based on coherence
in the left parietal regions during visual processing (Figures 1G). Fait intéressant, the same set
of brain regions also exhibits greater BOLD activity differences with neurotypicals during vi-
sual processing (as shown in Figures 1D), and the same set of brain regions are also involved
in cross-modal plasticity in non-autistic, sensory-impaired individuals (Figures 1H; Mottron
et coll., 2014). This evidence of greater variability in the left visuo-motor superior parietal cortex
and in the left associative visual areas in the autistic group compared with the control group,
and idiosyncrasy over associative visual-motor areas and greater sensory functional connec-
tivité, is prevalent throughout the autism literature (Alaerts et al., 2015; Alaerts et al., 2016;
Alaerts et al., 2014; Foxe et al., 2015; Cerliani et al., 2015; Cheng et al., 2015; Ciarrusta et al.,
2019; Hahamy et al., 2015; Harlalka et al., 2018; Henry et al., 2018; Keown et al., 2017;
King et al., 2019; Lynch et al., 2013; Mash et al., 2018). En résumé, the brain regions with
largest cross-modal plasticity such as posterior superior temporal sulcus (pSTS), fusiform face
area (FFA), precuneus, posterior cingulate cortex (PCC), and posterior parietal cortex (PPC),
which are involved in perceptual processing, overlap with regions that are the most variable
and most plastic in neurotypical individuals (Mottron et al., 2014). This overlap suggests a
general mechanism for neuroplasticity, which mostly involves brain regions that are highly
susceptible to reorganization. In contrast, primary sensory regions are less flexible and likely
more hardwired, requiring a high degree of neural constraints because of their topographic
(par exemple., retinotopic/tonotopic) organization (Mottron et al., 2014).
Taken together, these findings suggest that the highest interindividual differences in resting-
state connectivity are in the multimodal association cortex and the lowest are in the uni-
modal sensory and motor cortices, following a cortical hierarchy. This gradient of connectivity
297
Heteromodal association cortex:
A region that receives input from
multiple sensory or multimodal
domaines. These areas, y compris
the prefrontal cortices,
occipito-parieto-temporal junction,
multisensory areas, and superior
cortex temporal, are considered to
play role in higher level cognition
and context-dependent social
traitement.
Neurosciences en réseau
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
un
_
0
0
1
8
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
un
_
0
0
1
8
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 1.
Interindividual variability and functional connectivity between autistic and neurotypi-
cals in polymodal brain regions. (UN) Interindividual variability in resting-state functional connec-
tivity in neurotypical individuals. Positive and negative resting-state correlation values below the
global mean value are displayed in warm and cool colors, respectivement. (B) Regions showing greater
cortical gyrification in autistic individuals compared with neurotypical individuals. (C) Regions of
enhanced resting-state local connectivity are displayed, with greater connectivity in the autistic in-
dividuals than in neurotypicals (in warm colors), and regions of lower connectivity (in cool colors).
(D) Regions showing greater activity in autistic individuals than in neurotypical individuals when
processing visual stimuli (whole-brain FDR corrected). (E) The localization of the peak activation
patterns is shown in autistic individuals (blue) and exhibits higher variability than in neurotypical
individuals (red). (F) Clusters of brain structure alterations (differences in gray or white matter) être-
tween autistic and neurotypical individuals (whole-brain FDR corrected). (G) Regions where high
MEG connectivity with the right parietal region (yellow circle, coherence analysis) is associated
with high reading ability (darker color represents stronger correlation). (H) Regions of differences
in multisensory activity between visually impaired and sighted individuals when processing audi-
tory information (whole-brain FWE corrected). Figure adapted and modified with permission from
Mottron et al. (2014).
strength from early sensory areas to polymodal cortices may directly impact whole-brain dy-
namism and higher order multisensory integration in autism (Fu et al., 2019; Guo et al., 2017;
Guo et al., 2020; Harlalka et al., 2019; Hong et al., 2019; King et al., 2019; Naïr, Treiber,
Shukla, Shih, & Müller, 2013; Noel, De Niear, Stevenson, Alais, & Wallace, 2017; Nomi &
Uddin, 2015; Ostrolenk et al., 2019; Watanabe & Rees, 2017; Watanabe et al., 2019).
Maturation of Core-Periphery Brain Networks in ASD: Structural Changes
Brain network maturation occurs as early as the neonatal stage, showing sharp increases
entre 6 et 9 years of age, then follows a protracted development throughout young adult-
hood, becoming largely mature by age 20; cependant, there are also many changes that con-
tinue to occur throughout adulthood and into old age (Ciarrusta et al., 2019; Courchesne
et coll., 2011; Foxe et al., 2015; Henry et al., 2018; Oldham & Fornito, 2019; Olson et al., 2020).
Neurosciences en réseau
298
Core-periphery brain network dynamics in autism
Functional degeneracy:
Well-known characteristics of a
biological system whose elements
that are structurally different to
perform the same function or
yield the same output.
Default mode network:
Network of brain regions known to
be active during the resting condition
but that typically deactivate during
the task condition. It is commonly
related to self-referential thinking
and daydreaming and represents
intrinsic activity of the brain.
Early brain volume overgrowth during infancy and the toddler years has been observed in autis-
tic children, followed by an accelerated rate of decline in size and perhaps degeneration from
adolescence to late middle age (Ciarrusta et al., 2019; Courchesne et al., 2011). Previous longi-
tudinal and cross-sectional MRI studies reported maturation-related anatomical abnormalities
in ASD, including overgrowth in early life but accelerated decline during adolescence and
young adulthood (Abbott et al., 2018; Alaerts et al., 2014; Anderson et al., 2011). The frontal
lobe, implicated in functions with high social and executive demands, showed the most severe
enlargement in ASD beginning between 2 et 3 years of age, and frontal gray matter developed
at an atypical growth rate in children with ASD (Anderson et al., 2011). Cortical thickness stud-
ies also clarified abnormal longitudinal neurodevelopmental trends with regional specificity in
individuals with ASD, suggesting that cortical development in ASD first undergoes an expan-
sion at a high rate in early childhood, then undergoes accelerated thinning until adolescence,
and finally protracted thinning in early adulthood (Abbott et al., 2018; Anderson et al., 2011;
Courchesne et al., 2011). Plus loin, diffusion-weighted imaging (DWI) studies examined frac-
tional anisotropy (FA), mean diffusivity, and structural connectivity (SC) using tractography to
characterize various stages of the adult life span (Baum et al., 2017; Dajani et al., 2020; Huang
et coll., 2015). Volumetric studies suggest an early period of brain overgrowth in ASD followed
by slowed growth during later childhood when the typically developing braincatches up with
that of the autistic brain in terms of volume (Courchesne et al., 2011). De plus, a recent study
found that modular organization and small-world attributes are evident at birth, with several
important topological metrics increasing monotonically during development (Huang et al.,
2015). Most significant increases of regional nodes occur in the posterior cingulate cortex, un
hub region of the default mode network (DMN). Positive correlations exist between nodal effi-
ciencies and FA values of the white matter tracts, while correlations between efficiencies and
FA values vary among many brain regions. These results reveal substantial topological reorga-
nization of human brain structural networks through infancy and childhood, which is likely to
be the outcome of both heterogeneous strengthening of the major white matter tracts and prun-
ing of other axonal fibers. The anatomical connectivity alterations during early development
and increased modular segregations between anatomical brain areas and large-scale brain net-
works mediate development of executive functions at youth (Baum et al., 2017). Fait intéressant,
both cross-sectional and longitudinal data support the increased segregation-with-maturation
and decrease thereafter story (Baum et al., 2017; Dajani et al., 2020; Huang et al., 2015;
Keown et al., 2017; Lawson et al., 2015). These findings further set the stage for conducting
more targeted investigation into multidimensional brain and behavioral links spanning neu-
rodevelopmental processes that support executive functions, mémoire, and salience process-
ing (Keown et al., 2017; Khambhati, Medaglia, Karuza, Thompson-Schill, & Bassett, 2018;
Lawson et al., 2015; Liégeois et al., 2019; Preti & Van De Ville, 2019; Rosenthal et al., 2013;
Xia et al., 2018).
Maturation of Core-Periphery Brain Networks and Functional Connectivity
Emerging evidence suggests that ASD is associated with atypical trajectories of brain matu-
ration (Guo et al., 2017). This has been indexed by decreased spontaneous low-frequency
fluctuation (ALFF) of BOLD amplitude in the right precuneus and left middle occipital gyrus
during all developmental stages (Guo et al., 2017). Significant diagnosis by age interactions is
mediated by medial prefrontal cortex (mPFC), a key node of the DMN, with lowered ALFF
in autistic children but higher ALFF in autistic adolescents and adults (Guo et al., 2017).
More specifically, the quadratic changes of ALFF associated with increasing age in mPFC in
Neurosciences en réseau
299
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
un
_
0
0
1
8
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
Executive control network:
Large-scale control network
consisting of flexible hubs that
regulate distributed systems (par exemple.,
visuel, limbic, moteur) according to
current task goals.
neurotypicals were largely absent in ASD. En plus, abnormal ALFF values in ASD-related
brain regions predicted social deficits in ASD (Guo et al., 2017).
Recent functional connectivity evidence further suggests that in neonates with and without
a family history of ASD, those with a family history had significantly higher neural activity
in the right fusiform and left parietal cortex (Ciarrusta et al., 2019). En outre, the pattern
of age-related changes in spontaneous activity in the cingulate and insula was disrupted in
infants with a family history of ASD (Ciarrusta et al., 2019). En outre, stronger functional
connectivity and the degree of overconnectivity between visual and sensorimotor networks
were associated with greater autism symptoms in toddlers (B. Chen et al., 2020; Ciarrusta
et coll., 2019).
Significant hypoconnectivity has been observed in adolescents, especially in the DMN,
while younger children exhibit both hyper- and hypoconnectivity (Harlalka et al., 2018). Fur-
thermore, few recent studies highlight the importance of age stratification to test the devel-
opmental hypothesis that hyperconnectivity of brain networks may be more characteristic of
young children with ASD, while hypoconnectivity may be more prevalent in adolescents and
adultes (Harlalka et al., 2018, 2019; Henry et al., 2018; Nomi & Uddin, 2015). In one study,
the authors found that in the youngest cohort (âge 11 et sous), children with ASD exhib-
ited hyperconnectivity within large-scale brain networks including the DMN, salience network
(SN), and executive control network (ECN) as well as decreased between-network connectiv-
ity compared with age-matched neurotypicals. In contrast, adolescents with ASD (in the age
range 11–18) did not differ from neurotypicals in within-network connectivity, yet showed de-
creased between-network connectivity compared with neurotypicals (Nomi & Uddin, 2015).
Adults with ASD showed no within- or between-network significant differences in functional
network connectivity compared with age-matched controls (Nomi & Uddin, 2015). Taken to-
gether, atypical development of functional connectivity patterns in key sensory and higher
order multimodal brain regions may index vulnerability for autism.
Many previous cross-sectional functional connectivity studies reported that individuals with
ASD exhibited atypical developmental trajectories of DMN connectivity and frontostriatal con-
nectivity across childhood and adolescence, and a significant interaction between diagnosis
and age was observed in several core DMN regions, such as the mPFC, PCC, and precuneus
(Cheng et al., 2015; Delmonte, Gallagher, O’Hanlon, McGrath, & Balsters, 2013; Guo et al.,
2017; Harlalka et al., 2018; Henry et al., 2018). Functional connectivity circuits of the pSTS,
a core hub region implicated in sociocognitive processing, has also been shown to exhibit
atypical developmental trajectories in ASD (Alaerts et al., 2015; Alaerts et al., 2016).
Research examining developmental changes in large-scale network functional connectiv-
ity demonstrated that individuals with ASD exhibited different abnormal patterns of within-
and between-network connectivity during different developmental stages (Guo et al., 2017;
Harlalka et al., 2018; Henry et al., 2018; Nomi & Uddin, 2015). In spite of site-specific het-
erogeneity and inclusion of nonoverlapping samples in the studies listed in Table 2 from the
Autism Brain Imaging Data Exchange (ABIDE), there is increasing convergence in support of
a developmental model accounting for the age-specific over- and underconnectivity findings
in ASD. This model posits that childhood autism is characterized by brain hyperconnectivity,
whereas adolescent and adulthood autism is characterized by brain hypoconnectivity (B. Chen
et coll., 2020; Delmonte et al., 2013; Guo et al., 2017; Guo et al., 2019; Harlalka et al., 2018;
Henry et al., 2018; Huang et al., 2015; Nomi & Uddin, 2015). These findings suggest atypical
cortical developmental trajectories across the life span, and highlight the importance of taking
Neurosciences en réseau
300
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
un
_
0
0
1
8
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
different developmental stages into account when exploring the potential neural mechanisms
of ASD (Gollo et al., 2017; Hahamy et al., 2015; Henry et al., 2018; Naik, Banerjee, Bapi,
Deco, & Roy, 2017; Naik, Subbareddy, Banerjee, Roy, & Bapi, 2017).
Another interesting recent study evaluating age and gender effects jointly on intrinsic func-
tional connectivity found that the ASD group was characterized by an increase in regional
segregation into distinct functional networks, followed by a marked decrease in segregation
across time. The opposite pattern was observed for neurotypical individuals, suggesting that
segregation of functional networks persists into adulthood in typical development but not in
ASD (Henry et al., 2018). A similar pattern was observed in integration results: Individuals
with ASD evidenced increased functional integration over development, whereas the oppo-
site was observed for typically developing (TD) individuals. These findings help to clarify prior
work that showed hypoconnectivity in ASD for adults within various large-scale brain networks
(DMN, SN, ECN), as well as between networks (Kana et al., 2007; Lynch et al., 2013; Supekar
et coll., 2013; Uddin et al., 2013), whereas the opposite has been found for children with ASD
(Anderson et al., 2011; Guo et al., 2020; Harlalka et al., 2018, 2019). Cependant, complemen-
tary to the above findings, it was also observed that there was an overall idiosyncratic pattern
distributed over the whole brain in adults with ASD, with a lack of segregation of functional
networks and a higher integration of the component regions of interest in ASD (Hahamy et al.,
2015). Ainsi, hypoconnectivity within networks may partially be explained by a lack of differ-
entiation in adulthood and increased connectivity within sensory areas (B. Chen et al., 2020).
There are very few studies that have examined the relationship between age-related change
in intrinsic functional connectivity and gender in ASD and TD (Guo et al., 2017; Harlalka
et coll., 2018; Henry et al., 2018). The majority of resting-state fMRI studies of autism have
focused on characterizing intrinsic large-scale brain network organization in adolescent and
adult males, barring a few studies that have given some consideration to both age and gender
(Guo et al., 2017; Harlalka et al., 2018; Henry et al., 2018; Lai et al., 2017). A large majority
of these studies found that ASD exhibits increased functional integration at the expense of de-
creased functional segregation (Abbott et al., 2018; Alaerts et al., 2015; Alaerts et al., 2016;
Alaerts et al., 2014; Cerliani et al., 2015; Cheng et al., 2015; Delmonte et al., 2013; Harlalka
et coll., 2018; Henry et al., 2018; Keown et al., 2017).
In adolescents with ASD, there is a
significant decrease in modularity, suggesting a less robust modular organization, and an in-
crease in participation coefficient, suggesting more random integration and widely distributed
connection edges (Harlalka et al., 2018; Henry et al., 2018; Keown et al., 2017). Modularity
decreased nonlinearly in the ASD group with age, as evidenced by an increase and then a
decrease over development. Age effects on modularity were localized to the somatosensory
réseau (Henry et al., 2018). En outre, there is significant hypoconnectivity observed in
the adolescent group, especially in the DMN (Harlalka et al., 2018), while children showed
both hyper- and hypoconnectivity (Harlalka et al., 2018; Henry et al., 2018). While the find-
ings were applied at the global level, they were not equally robust across all networks and
in one case (c'est à dire., greater cohesion within the ventral attention network in ASD) even reversed
(Keown et al., 2017; shown in Figures 2 et 3). Among those studies that investigated gender
effects in an age-constrained manner was one reporting that TD female individuals had higher
quadratic effects of age on modularity that trended toward significantly different from male
individuals with ASD, and males and females showed a differential neural expression of ASD,
characterized by predominantly hypoconnectivity patterns in males with ASD (compared with
TD males), and hyperconnectivity in females with ASD (compared with TD females; Alaerts
et coll., 2016; Harlalka et al., 2018; Henry et al., 2018). As higher order interaction and nonlin-
ear regression effects are notoriously underpowered even for large sample sizes, these results
Modularity:
A graph theoretic measure for the
strength of division of a network into
modules (or communities). Networks
with high modularity have dense
connections between the nodes
within modules, but sparse
connections between nodes in
different modules.
Neurosciences en réseau
301
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
un
_
0
0
1
8
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
un
_
0
0
1
8
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
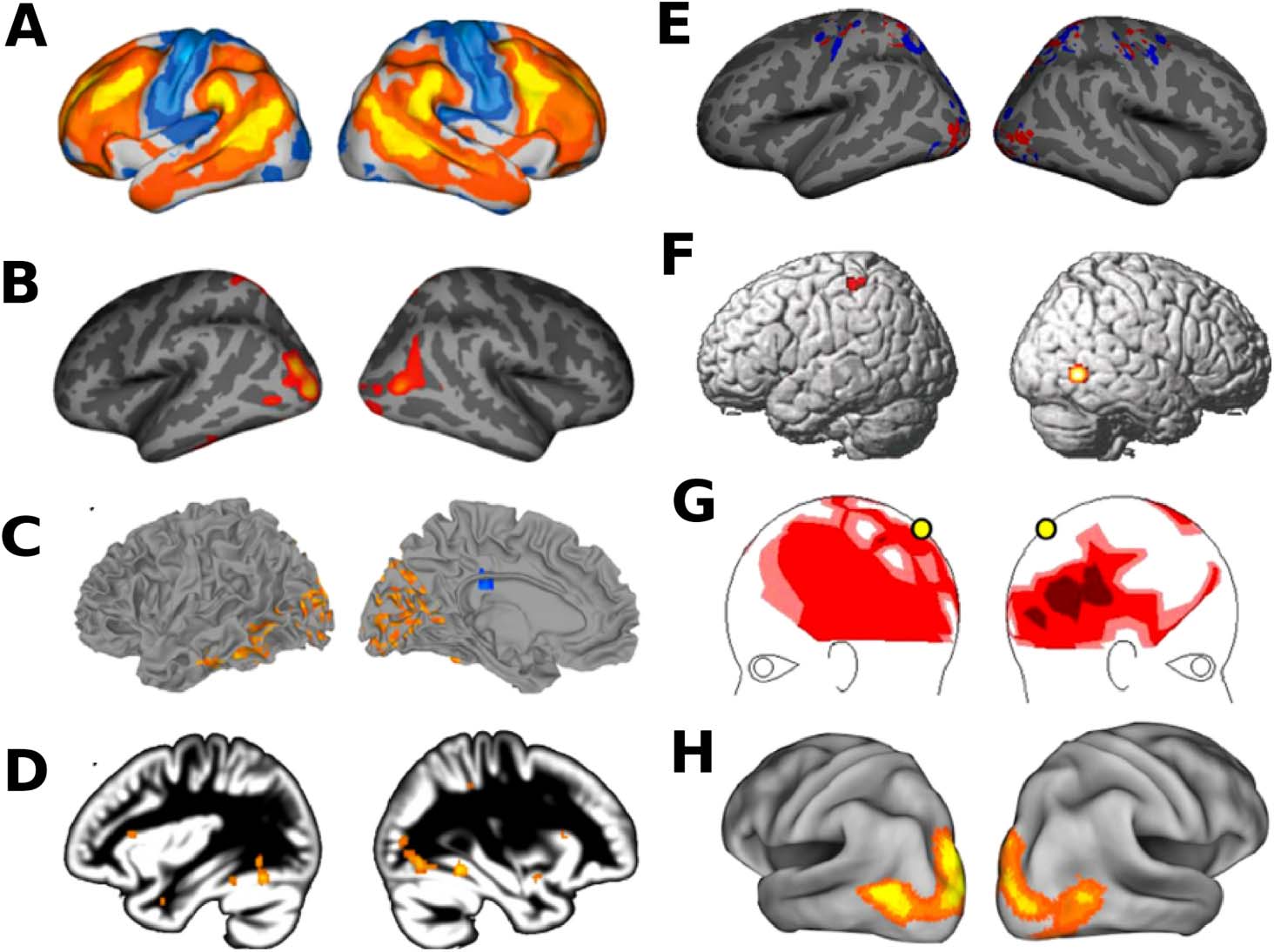
Chiffre 2. Hypervariant ASD connections estimated using dFCVar matrix. The majority of con-
nections in children are long-range, while the adults exhibit hypervariability in dFC in both middle-
range and long-range connections. Adolescents are seen to have majority short-range connections
exhibiting hypervariability. Figure adapted with permission from Harlalka et al. (2019).
in terms of reduced age and gender-specific trends in differentiation and functional special-
ization of brain networks should be interpreted with caution (Courchesne et al., 2011; King
et coll., 2019).
Efficient functioning of specialized sensorimotor and cognitive networks relies on two com-
plementary organizing principles: functional segregation (or differentiation), emphasizing the
degree to which different regions or networks are specialized, and functional integration, concernant-
ferring to the communication between regions within a specialized network (Abbott et al.,
2018; Anderson et al., 2011; Harlalka et al., 2018; Henry et al., 2018; Keown et al., 2017;
Nomi & Uddin, 2015; Padmanabhan, Lynch, Schaer, & Menon, 2017; Ray, Hajare, Roy, &
Banerjee, 2020; Reiter et al., 2019). The differential relationship between modularity and age
seen in ASD was in a large part due to the peripheral networks (somato-sensorimotor and vi-
sual networks). This result from localization analysis suggests that the somatosensory network
drives, at least in part, the increase in modularity across time seen in neurotypicals relative
Dans une étude récente, it was fur-
to those with ASD (Harlalka et al., 2018; Henry et al., 2018).
ther demonstrated in toddlers with ASD that the degree of overconnectivity between visual
and sensorimotor networks was associated with greater autism symptoms, and an age-related
weakening of the visual-auditory between-network connectivity was observed in the ASD but
not the TD group (B. Chen et al., 2020).
Ainsi, differentiation and specification of regions related to the visual and somato-
sensorimotor network appears to contribute greatly to functional connectivity changes across
development. Cependant, a network knockout approach was used to isolate the influence of spe-
cific functional networks, a simple leave-one-out process demonstrating that the somatosen-
sory cortex had no effect on global efficiency models (Harlalka et al., 2018; Henry et al., 2018;
Keown et al., 2017), which suggests that the differences in global efficiency between ASD and
Neurosciences en réseau
302
Core-periphery brain network dynamics in autism
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
un
_
0
0
1
8
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
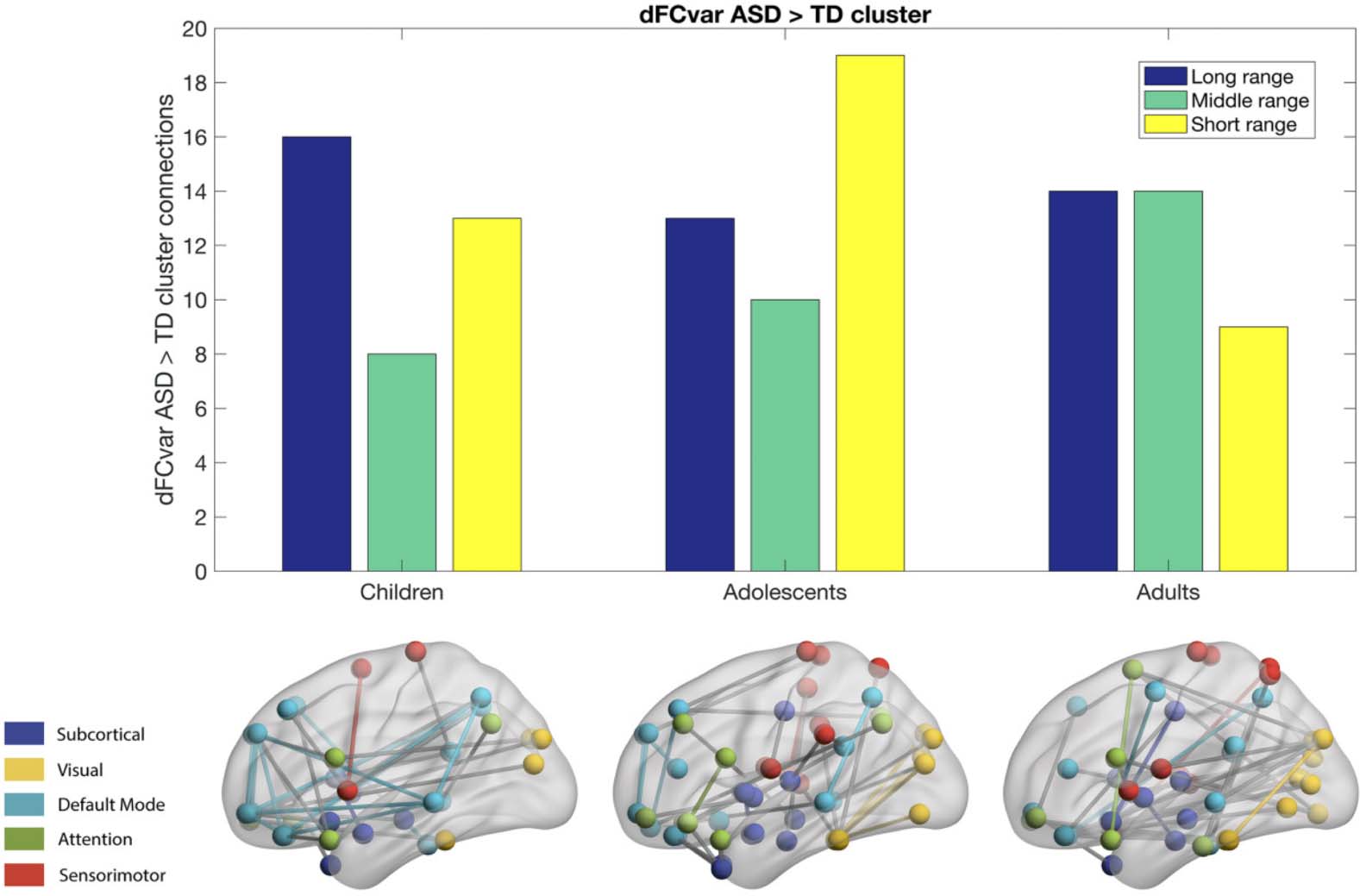
Chiffre 3. Globally atypical network flexibility of brain modules in autism. (UN, B) Connection density (UN) and strength (B) between each pair
of networks. Group averages are shown for the TD group in the lower left triangle and for the ASD group in the upper right triangle. Réseau
connections with lower density or strength are denoted by dashes (bold indicates p < 0.05 after FDR correction). (C) Approximate location of
nodes with highest betweenness centrality in the TD group (yellow), and nodes with greater (red) or reduced (blue) betweenness centrality in
ASD (all p < 0.05, uncorrected). (D) Brain plot of areas showing significant effect of age, diagnosis by age, and interaction effect on flexibility,
cohesion strength, and disjointness, respectively. Typically developing, TD; autism spectrum disorder, ASD. Figure adapted with permission
from Harlalka et al. (2019) and Keown et al. (2017).
TD were not localized to the somatosensory cortex, but rather reflect a more global whole-
brain phenomenon (Henry et al., 2018; Keown et al., 2017). Longitudinal studies demonstrate
that childhood executive functions largely predict variance in autistic individuals’ adaptive be-
havior later in life (Kenny et al., 2019). Taken together, this work highlights the need for more
targeted future research and investigation of the brain mechanisms at various stages of matu-
ration in both male and female ASD to pinpoint subtypes of functional connectivity patterns
across development linking adaptive behavior, cognitive flexibility, executive task processing,
and manifestations of core and contextual deficits in ASD across the life span.
Sensory and Sociocognitive Deficits in ASD: Behavior and Neuroimaging Studies
Studies of neurotypical individuals and those with ASD have typically utilized a variety of
behavioral paradigms ranging from sensory-motor perceptual integration, attention, cognitive
flexibility and executive functions, face-to-face communication, and conversation, all of which
can be broadly categorized as (a) tasks with high social and executive demands (Jasmin et al.,
2019), and (b) tasks with high sensory but low social demand (Robertson, Martin, Baker, &
Baron-Cohen 2012).
Here, we review a sampling of behavioral studies in which individuals with autism dis-
play sensory and cognitive deficits (see Table 1 for a non-exhaustive list of examples). More
specifically, we have reviewed tasks with greater sensory demands, overt sensory-motor, repet-
itive behavior (low social demands), examining core neural features originating from abnormal
thalamic and striatal interactions and sensory input gating. We also highlight a sampling of
studies with social components such as face-to-face communication, day-to-day conversation,
Network Neuroscience
303
Core-periphery brain network dynamics in autism
Table 1.
sociocognitive processing, and mentalizing in autism
Sampling of studies examining restrictive and repetitive behavior, sensory abnormality, executive functions, communication,
Behavioral paradigms
Sensory-motor perception and integration
Visual acuity (VA) and perception
Integration of sensory input and visual reaction time
(RT)
Atypical visual saliency
Integration of motion information
Integration of motion signals and perceptual
decision
Attention, cognitive flexibility, and executive functions
Motor inhibition, decision-making, and set switching
Executive function (EF), response inhibition
Executive function (planning, inhibition,
and cognitive flexibility) and theory of
mind (false-belief understanding)
Behavioral inflexibility, attention and
executive functions
Cognitive shift, repetitive and restrictive
behavior
Multisensory processing (audio-visual
Integration)
Multisensory facilitation using sensory
integration
Eye gaze to integrate joint role of
attention and comprehension
of mental states of others
Atypical cross-modal (auditory-visual)
modulation linked to sociocommunicative
deficits
Atypical audio-visual temporal recalibration
and speech stimuli
Intelligible multisensory speech perception
Type of stimuli
References
Freiburg Visual Acuity and Contrast Test
Visual search task (Feature and conjunction type)
Ashwin, Ashwin, Rhydderch, Howells, and Baron-Cohen
(2009)
Plaisted et al. (1998)
Gaze patterns during natural scene-viewing
Directional variability in standard motion dot
coherence task
Motion discrimination task, manually indicating the
global direction of motion in a random dot across a
range of coherence level
Pelphrey, Morris, and McCarthy (2005)
Manning et al. (2015)
Robertson, Martin, Baker, and Baron-Cohen (2012)
Three different EF tasks: (a) motor-inhibition
(GO/NO-GO); (b) cognitive interference-inhibition
(spatial STROOP); and (c) set shifting (SWITCH)
Response inhibition task during alphabetic letter
matching criterion under three experimental
conditions
Longitudinal study 12-year follow-up Time 1 tested on
components of executive function (planning,
inhibition, and cognitive flexibility) and theory of mind
(false-belief understanding).
At Time 2, tested participants’ autistic features and
adaptive behavior.
Stimulus-evoked brain states involving performance of
social attention and numerical problem-solving tasks
A target detection task during which geometric shapes
(squares, triangles, or circles) were presented one at a
time. Participants were required to classify each
stimulus as a “target” or “non-target” on the basis of its
shape and respond with an appropriate button press.
Using BRIEF: Behavior Rating Inventory of Executive
Function scale to access behavior
Face-to-face structured and unstructured
communication using a modified version of the
Interest Scale questionnaire
Synchronous auditory pip during a complex visual
search task (pip-pop effect)
Nonsocial stimuli (i.e., flashes and beeps)
Stimulus presentation is based on congruent and
incongruent trials over which participant needs to
integrate information to comprehend what a virtual
actor ought to do in a given context. Social and
contextual stimuli.
Auditory (high or low pitch) and visual conditions (dot
located high or low in the display) were presented, and
participants indicated whether the stimuli were “high”
or “low”
Asynchronous audio-visual stimuli of varying levels of
complexity and performance of a simultaneity
judgment (SJ)
Integrated seen and heard speech were accessed while
the environmental noise was systematically
manipulated
Schmitz et al. (2006)
Kana et al. (2007)
Kenny et al. (2019)
Uddin et al. (2015)
Shafritz et al. (2008)
Moul et al. (2015)
Jasmin et al. (2019)
Collignon et al. (2013)
Ostrolenk et al. (2019)
Plaisted et al. (1998)
Jao Keehn et al. (2017)
Noel et al. (2017)
Foxe et al. (2015)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Metacognitive executive abilities and atypical
flexibility
Face-to-face communication, multisensory speech perception, sociocognitive processing, and mentalization
Sociocognitive response and communication
emotions, pragmatics, and sarcastic prosody that are highly context dependent (high social
demands) and used to examine contextual (state-dependent) neural features. Subcortical areas
also play a crucial role in core neural processing as highlighted above; specifically, determining
adaptive behavior, state- and trait-specific variability, and flexibility. However, only a handful
of studies have looked at their potential role in symptom severity in autism. Several stud-
ies showed decreased functional connectivity among cortical regions associated with social
functions, such as the superior temporal sulcus, medial prefrontal, temporoparietal junction,
left inferior frontal gyrus, as well as somatosensory cortex (Alaerts et al., 2015; Alaerts et al.,
Network Neuroscience
304
Core-periphery brain network dynamics in autism
2014; Anderson et al., 2011; Baum et al., 2017), with some showing simultaneously increased
functional connectivity between thalamus, striatum, and some of the same cortical regions
(Abbott et al., 2018; Alaerts et al., 2015; Anderson et al., 2011; Cerliani et al., 2015; but
see Nair et al., 2013). Findings from the task-based functional magnetic resonance imaging
literature demonstrate that the above subcortical and cortical brain regions are involved in re-
stricted and repetitive behaviors (RRB) and may represent some of the earliest biomarkers of
ASD (B. Chen et al., 2020; Ciarrusta et al., 2019; Robertson & Baron-Cohen, 2017). Previous
studies also reported that individuals with autism showed increased functional connectivity be-
tween regions in the frontal cortex (anterior cingulate cortex, ACC; middle frontal gyrus, MFG;
paracingulate gyrus, Pcg; and orbitofrontal cortex, OFC), and striatum (nucleus accumbens,
NAcc; and caudate) compared with neurotypical individuals (Abbott et al., 2018; Cerliani
et al., 2015; Delmonte et al., 2013; Jasmin et al., 2019).
Furthermore, increased functional connectivity between ACC and caudate was associated
with reduced activation to social rewards in the caudate (Delmonte et al., 2013). Greater
connectivity between the right MFG and caudate was associated with higher RRBs, and con-
nectivity between the bilateral Pcg and NAcc, and the right OFC and NAcc, was negatively as-
sociated with social and communicative deficits (Cerliani et al., 2015; Delmonte et al., 2013).
These findings indicate that abnormalities in frontostriatal circuitry potentially underlie ma-
jor deficits in ASD, social interaction and communication difficulties and RRBs (Alaerts et al.,
2015; Delmonte et al., 2013; Guo et al., 2020; Guo et al., 2019; Jasmin et al., 2019). Neu-
roimaging evidence further demonstrates that autistic adolescents show atypical activation of
the mentalizing system. Longitudinal follow-up of a group of autistic children who passed or
failed theory of mind tests revealed reduced activation of the medial prefrontal cortex and pre-
cuneus, posterior cingulate and lateral temporal cortices in children, and predicted significant
variance in young children’s adaptive behavior at the 12-year follow-up (Kenny et al., 2019).
Although the development of co-occurrence of deficits in sensory and higher order so-
ciocognitive processing in ASD is a topic of considerable interest, links between core-periphery
brain network maturation and how they contribute to behavioral variability and unique vari-
ance to the diagnostic criteria of autism are not yet firmly established. However, the studies
reviewed here provide critical insights into atypical integration of sensory input at the local
level, eventually leading to impairment manifested at the global level, addressing simultaneity
of sensory and as well as sociocognitive deficits encountered in autistic children, adolescents,
and adults.
Neural Substrates of Behavioral Variability in Autism: Predictions From Atypical
Core-Periphery Dynamics
One paradox of autism is the co-occurrence of deficits in sensory and higher order sociocogni-
tive processing (Hong et al., 2019). Here, we review whether these phenotypic patterns based
on different brain dynamics approaches (flexibility of system-level transitions, functional gra-
dients, and timescale hierarchy) all converge in relating overarching system-level imbalance—
specifically a disruption in macroscale hierarchy affecting integration and segregation of core
polymodal and peripheral unimodal networks. Task-based neuroimaging is necessary to un-
derstand the neural basis of atypical sensory and cognitive processing in several domains such
as face-to-face conversation, working memory, attention, executive processing, and percep-
tion; however, given the decreased cognitive demands and potential for reuse, resting-state
functional magnetic resonance paradigms or task-free approaches in autism have become a
promising avenue for discovery in recent years. Resting-state paradigms are also adoptable
with some degree of ease by various multimodal approaches (EEG, MEG, ECOG) for generat-
ing specific hypotheses based on characterizing normative brain dynamical patterns (Chang
& Glover, 2010; Sahoo, Pathak, Deco, Banerjee, & Roy, 2020). Moving forward, one could
Network Neuroscience
305
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
use resting-state fMRI for precise quantification of typical and atypical flexibility based on
core-periphery interactions (highly connected hub regions and small brain regions located
outside the core regions; e.g., striatum, subcortical areas) to discover whether atypical process-
ing in peripheral brain regions potentially gives rise to perturbation of large-scale macroscale
brain network dynamics (Gollo et al., 2017; Guo et al., 2020; Harlalka et al., 2019; Hong et al.,
2019; Preti & Van De Ville, 2019; Rashid et al., 2018; Vidaurre, Smith, & Woolrich, 2017;
Watanabe & Rees, 2017; Watanabe et al., 2019). Beyond revealing brain regions activated in
response to specific task conditions,
resting-state functional connectivity approaches
permit analysis of how cognitive functions emerge from precise timing and concerted activ-
ity in the specialized large-scale brain network interactions (Harlalka et al., 2019; Nomi, Vij,
et al., 2017; Preti & Van De Ville, 2019; Vidaurre et al., 2017; Watanabe & Rees, 2017;
Watanabe et al., 2019). Dynamic functional connectivity (dFC) approaches further enable
the study of moment-to-moment variability in neurotypical and autistic individuals, as docu-
mented by several recent studies (see Table 2). Furthermore, dFC variability is quantified by the
standard deviation of time-varying dynamic functional connectivity. Hence, dFCVar tracks the
changes in variability in dynamic functional connectivity between brain regions anchored in
large-scale neurocognitive networks. These measures are now frequently used to characterize
atypical hyper- and hypofunctional connectivity variability in neurodevelopmental disorders
(H. Chen et al., 2017; Liégeois et al., 2019; Gollo et al., 2017; Guo et al., 2020; Nomi, Bolt,
et al., 2017; Nomi, Vij, et al., 2017). A comprehensive review of various dynamic functional
connectivity methods and their application in psychopathology and flexible behavior is avail-
able elsewhere (Bolton et al., 2020; Uddin, 2021).
Accumulating evidence based on tracking dynamic changes in the autistic brain compared
with neurotypical individuals suggests broader qualitative agreement across findings using
open-access multisite neuroimaging data made available from ABIDE I and II (Di Martino
et al., 2017; Di Martino et al., 2014). The availability of such large open-access multicentric
datasets with unprecedented sample size and depth of phenotyping with balanced sex ratios
allows for discovering fundamental neural mechanisms in spite of various sources of hetero-
geneity (Harlalka et al., 2019; Henry et al., 2018; Hong et al., 2019; Nomi, Bolt, et al., 2017;
Rashid et al., 2018). The heterogeneity and site-related variability pose a severe caveat in the
interpretation and replicability of functional connectivity findings, which is further described
elsewhere in detail (King et al., 2019). Moreover, beyond sampling variability, the largest
part of observed “dynamics” and time-varying FC configurations during rest is attributable to
head motion (Laumann et al., 2017). Hence, while comparing the above metrics to establish
between-group differences (specifically in children with autism) and relating these static and
dynamic measures to symptom severity, one has to apply extreme caution as children with
autism generally exhibit higher levels of head motion and thus pose an important caveat in
interpretation.
However, several key challenges remain in the interpretation of functional connectivity–
based findings; in general, resting-state BOLD correlations and brain dynamics may reflect
processes concerned with maintenance of the long-term stability of the brain’s functional
organization. Recent studies have further stratified samples at different stages of develop-
ment (children, adolescents, and adults) to investigate how diagnosis by age interactions affect
modularity, segregation, and integration in the functional brain networks. To address the sta-
bility and integrity of functional architecture, recent studies have further looked at the regional
flexibility, cohesion, and disjointness in key core brain regions comprising the DMN, execu-
tive, and salience networks (Fu et al., 2019; Guo et al., 2017; Guo et al., 2020; Harlalka et al.,
2018, 2019; He et al., 2018). These recent findings along with the previous findings using static
306
Node disjointness:
Node disjointness defines the
fraction of time a node changes its
community affiliation over time,
independent of the other nodes.
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
Table 2.
Sampling of fMRI studies capturing atypical core-periphery brain dynamics and relationships with symptom severity
ASD and TD samples
ASD: 79 (included 31 out of 79)
TD: 105 (included 44 out of 105)
(ABIDE I)
ASD: 24
TD: 26 (ABIDE Utah site primary,
Indiana and Zurich site replication)
Total 507 male subjects
ASD: 209
TD: 298 (all ABIDE sites)
TD and ASD children combined: 774;
560 with SRS ASD: 22 (ABIDE sites)
TD children: 28
ASD children: 29
ASD children: 26
TD children: 26
Adolescent ASD: 28
Adolescent TD: 28
Adult ASD: 18
Adult TD: 18
(ABIDE NYU site)
ASD and typical controls, with = 10
individuals/group ABIDE I children and adults
(i.e., PITT, NYU, USM) n = 211, ASD = 103,
TD = 108 from three sites: (a) NYU Langone
Medical Center (NYU, 35/51 ASD/controls);
(b) University of Utah, School of Medicine (USM,
49/37 ASD/controls); (c) University of Pittsburg,
School of Medicine (PITT, 19/20 ASD/controls)
Replication data are from ABIDE II subsample, 103
individuals: (a) Trinity Centre for Health Sciences,
Trinity College Dublin (TCD, 12/16 ASD/controls);
(b) NYU Langone Medical Center (NYU, 25/18
ASD/controls); (c) Institut Pasteur/Robert Debré
Hospital (IP, 11/21 ASD/controls)
TD: 195, ASD: 170 (all ABIDE sites)
TD: 26, ASD: 25 (all male adults)
ASD: 105, TD: 102 (all ABIDE sites)
Age range (mean, SD)
ASD: 7–18
(not including 18 years)
(mean: 12.46, SD: 3.1)
TD: 7–18
(mean: 11.51, SD: 2.64)
ASD: 18.4–38.9
(mean: 25.3, SD: 5.5)
TD: 18.2–39.3
(mean: 25.3, SD: 6.3)
ASD: 6–36
(mean: 16.5, SD: 6.2)
TD: 6–36
(mean: 16.8, SD: 6.2)
Combined: 6–10
(mean: 7.99, SD: 1)
ASD: 3–7
(mean: 4.99, SD: 1.32)
TD: 3–6
(mean: 4.99, SD: 1.01)
Child ASD: 7.15–10.06
(mean: 9.51, SD: 1.12)
Child TD: 6.47–10.86
(mean: 9.10, SD: 1.32)
Adolescent ASD:
11.01–17.88
(mean: 13.71, SD: 1.79)
Adolescent TD: 11.32–16.93
(mean: 14.01, SD: 1.74)
Adult ASD: 18.58–39.1
(mean: 24.13, SD: 3.92)
Adult TD: 18.59–31.78
(mean: 25.41, SD: 5.87)
Discovery Data
ASD: 12.7–28.9
(mean: 20.8, SD: 8.1)
TD: 12.1–26.3
(mean: 19.2, SD: 7.1)
Analysis pipeline
dFCVar estimate using time-varying functional connectivity
of seven subnetworks composed of subcortical (SC), auditory (AU),
visual (VIS), somatomotor (SM), cognitive control (CC), default
mode (DM), and cerebellar (CB) networks. To determine the
connectivity states, covariance matrices of ASD and TD were
clustered by k-means clustering algorithm based on Manhattan
distance. Clustered centroid matrices were covariance matrices
of connectivity states and their relationship with symptom severity.
Energy-landscape analysis across seven well-established
resting-state brain networks to characterize atypical neural state
transition probability between core DMN, CEN, VAN, DAN, and
peripheral sensory networks and to quantify relationship with
symptom severity.
Standard deviation (SD) of dFC (dFCVar) matrix to compute
hypervariant connections across whole-brain regions of
interest and relationship with symptom severity.
Sliding-window correlation to estimate dFC and estimation of
dwell time based on fractional occupancy (FO) index. Globally
disconnected vs. hyperconnected whole-brain networks and core
DMN hub network and relationship with symptom severity.
K-means cluster analysis was performed to identify distinct
temporal states based on the spatial similarity of each
functional connectivity pattern. Estimation of dynamic
functional connectivity variance (dFCVar) between the hub regions
of the core DMN and sensory-motor network and
relationship with symptom severity.
Sliding-window analysis to calculate variability of dFC
(dFCVar) in order to quantify proportion of short-range,
long-range hypo- and hyperconnectivity (in each age group) patterns
in core-periphery brain networks composed of
visual, sensorimotor, subcortical, DMN, attention (identified using
multilayer modularity detection algorithm).
Quantification of atypical flexibility, cohesiveness, and
disjointness of core hub regions and peripheral brain regions and
relationship with symptom severity.
Reference
Yao et al. (2016)
Watanabe and
Rees (2017)
H. Chen et al. (2017)
Rashid et al. (2018)
He et al. (2018)
Harlalka et al. (2019)
Functional gradient analysis between core DMN regions
and sensory regions (primary auditory, visual, and
sensory-motor). Altered macroscale gradients and stepwise
functional connectivity (SFC) and relationship with symptom severity.
Hong et al. (2019)
ASD: 8.22–22.92
(mean: 15.57, SD: 7.35)
TD: 10.12–21.92
(mean: 16.02, SD: 5.90)
TD: 18.1–39.4
(mean: 25.3, SD: 6.3)
ASD: 18.4–50
(mean: 27.3, SD: 7.9)
TD: 7–12
(mean: 10.02, SD: 1.38)
ASD: 7–12
(mean: 10.1, SD: 1.26)
Dynamic functional network connectivity (dFNC) between 51
intrinsic connectivity network controls using independent component
analysis and a sliding-window approach. A hard clustering state
analysis and a fuzzy metastate analysis were conducted, respectively,
for the exploration of local and global aberrant dynamic connectivity
patterns in ASD. dFNC between thalamic and sensory networks in
each functional state and group differences in four high-dimensional
dynamic measures and relationship with symptom severity.
Atypical intrinsic neural timescales estimated from sensory and core
hub brain regions frontoparietal control, DMN, using autocorrelation
function and related to underlying anatomical connectivity SC.
Areas with shorter and longer timescales in the cortical hierarchy and
relationship with symptom severity.
Intra- and interhemispheric functional connectivity dynamics (FCD)
mapping between core-peripheral brain regions and
relationship with symptom severity.
Fu et al. (2019)
Watanabe et al.
(2019)
Guo et al. (2020)
Network Neuroscience
307
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
Rand index:
A network measure reflecting how
similar network organization was to
a normative set of networks. This
measure compares similarity
between two clustering assignments
and can be used for comparison
against a normative set of labels.
brain network analysis quantifying normative network measures such as Rand index, density,
and cohesiveness of regions of interest from ASD and TD suggest reduction in cohesion in
somatosensory motor, auditory, subcortical, ventral attention, and memory retrieval regions in
individuals with ASD compared with typically developing participants (Kenny et al., 2019).
This was accounted for by globally reduced cohesion and density, but increased dispersion of
the above networks (Kenny et al., 2019). Recent findings further suggest that there were signif-
icant differences in rich-club connectivity (among the hubs), which was generally increased in
the ASD group. There are also hypervariant ASD connections in the dFCVar matrix (H. Chen
et al., 2017; Guo et al., 2020; Harlalka et al., 2019). These hypervariant connections com-
prise inter- and intrahemispheric connectivity patterns (atypical long-range and short-range
connection distributions). Recent evidence further suggests that a large majority of hypervari-
ant connections in children and adults are long-range, while adolescents are seen to have a
majority of short-range connections (Figure 2). In particular, it was reported that brain regions
exhibiting reduced flexibility in ASD compared with TD are superior temporal gyrus (STG),
putamen (PUT), amygdala (AMYG), cuneus (CUN), inferior occipital gyrus (IOG), left inferior
parietal (IPL), and angular gyrus (ANG; Delmonte et al., 2013; Guo et al., 2017; Harlalka et al.,
2018; Jao Keehn et al., 2019; Kenny et al., 2019). It was also reported that several regions that
show effect of age include superior frontal orbital, amygdala (AMYG), cuneus (CUN), infe-
rior occipital gyrus (IOG), left inferior parietal (IPL), angular gyrus (ANG), caudate nucleus
(CAU), putamen (PUT), thalamus (THAL), dorsal SFG, and left superior temporal gyrus (STG;
Harlalka et al., 2019). There were interesting group differences found between ASD and TD in
pallidus gyrus (periphery region), which further showed a significant increase in flexibility in
adults as compared with both adolescents and children, while the superior frontal orbital (pe-
riphery region) shows a significant increase of flexibility in adults compared with adolescents
(Guo et al., 2020; Harlalka et al., 2019; Figure 3). In a recent study, autistic traits were accu-
rately characterized using 774 children 6 to 10 years old based on the quantification of longer
dwell times (less frequent state transitions in ASD compared with TD) and by global discon-
nection using dynamic functional connectivity analysis (Rashid et al., 2018). These findings
suggest that globally atypical brain dynamics in ASD may be already present at the earliest
stages of development and could be due to hub architecture being partly altered in ASD (Gollo
et al., 2015; Keown et al., 2017). In summary, these findings support a core-periphery brain
network dynamic model of reduced network integration (i.e., connectivity within networks)
and differentiation (or segregation; based on connectivity outside network boundaries) in ASD
(Guo et al., 2020; Harlalka et al., 2018, 2019; Keown et al., 2017; Preti & Van De Ville, 2019;
Rashid et al., 2018).
Although reproducibility in findings based on static and dynamic functional connectivity
methods described above is still a major hurdle in autism research (King et al., 2019), dynamic
functional connectivity approaches are already uncovering atypical patterns of brain dynam-
ics that distinguish autistic from neurotypical individuals in various developmental stages of
relevance for sensory and cognitive behavioral deficits (H. Chen et al., 2017; Guo et al., 2020;
Harlalka et al., 2019; He et al., 2018; Hong et al., 2019; Nomi, Bolt, et al., 2017; Nomi, Vij,
et al., 2017; Rashid et al., 2018; Vidaurre et al., 2017; Watanabe & Rees, 2017; see Table 2
for a sample list of studies).
Neural Substrates of Behavioral Variability in Autism: Prediction From Atypical
Core-Periphery Timescales
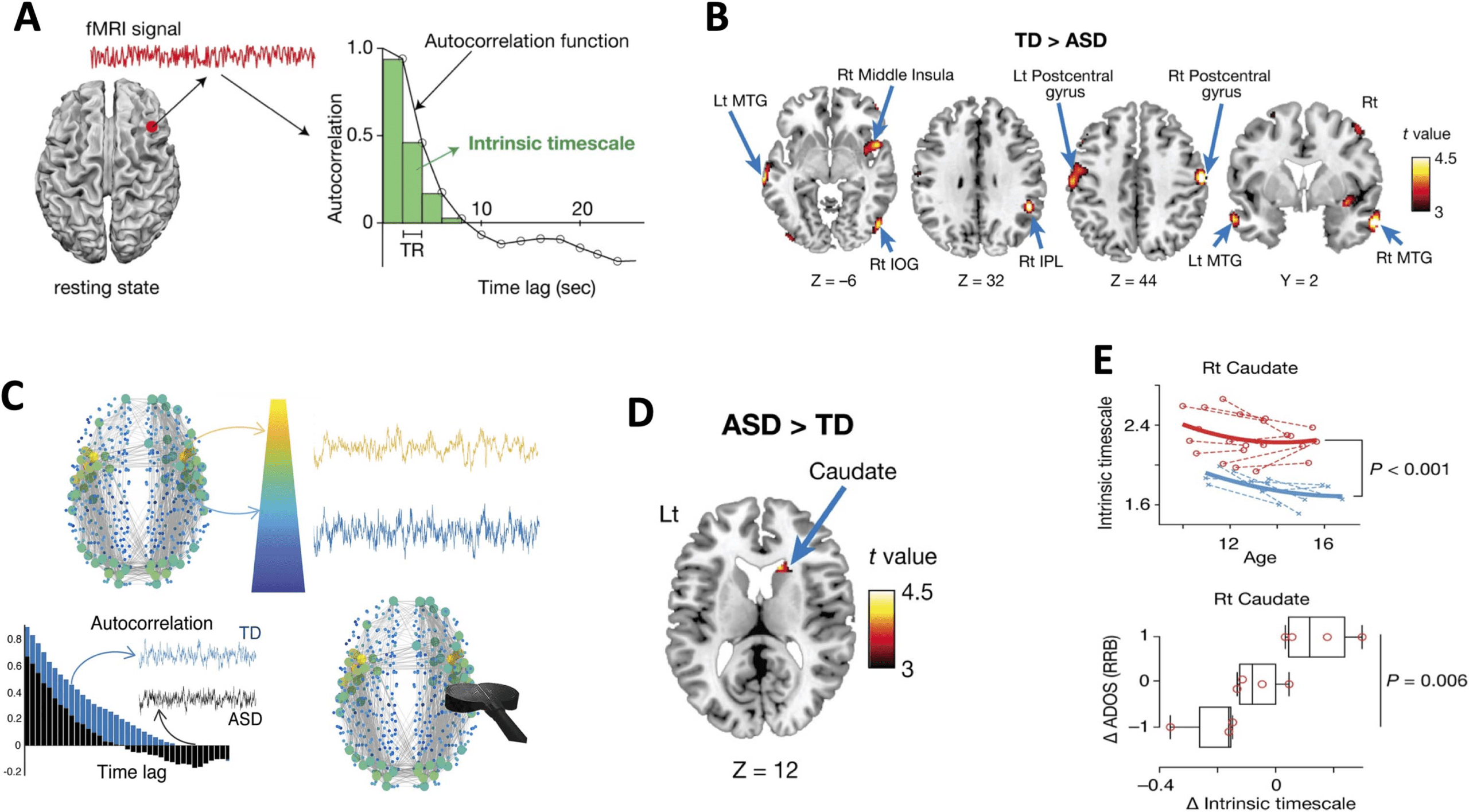
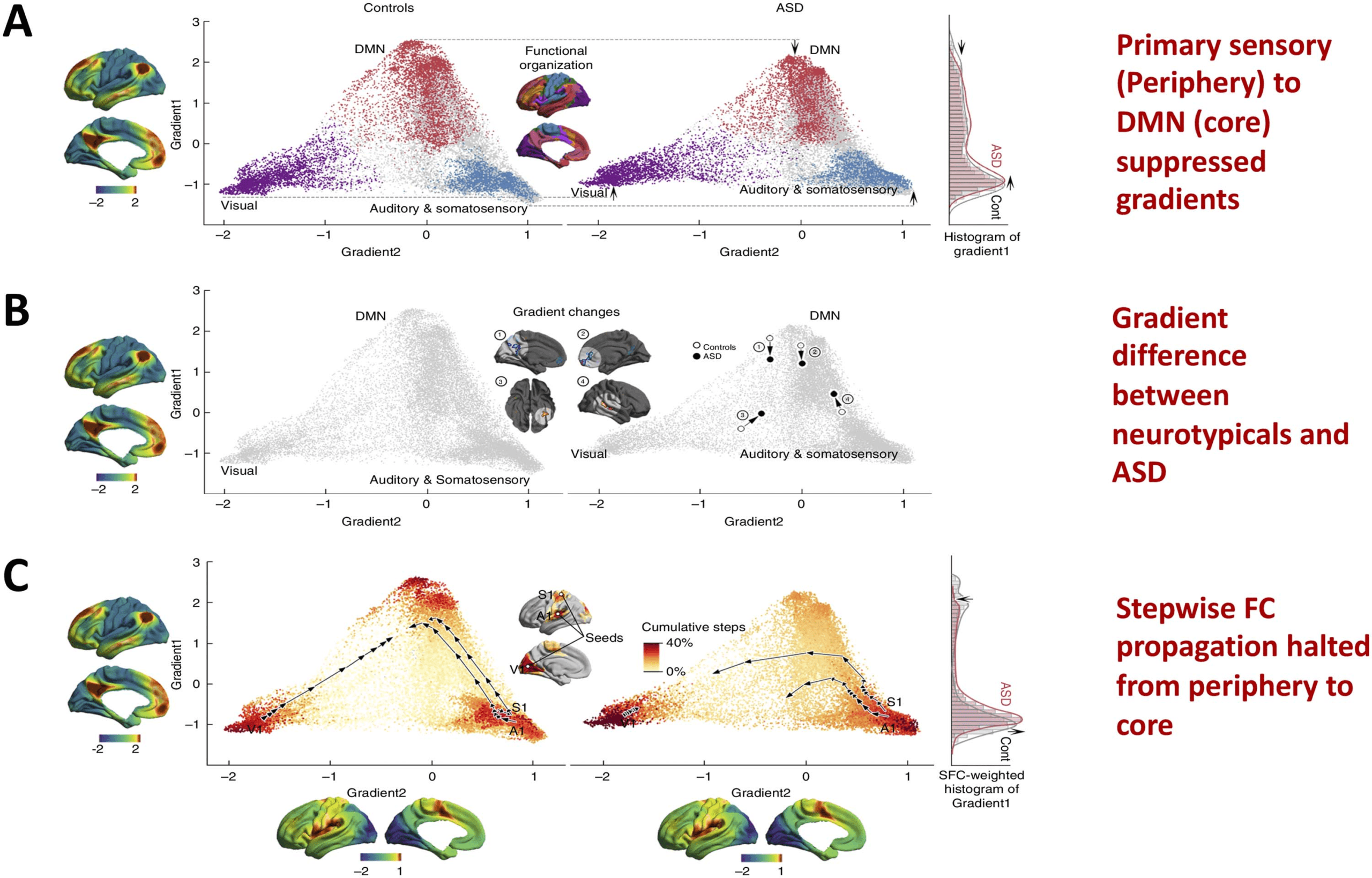
In the past few years, understanding cortical hierarchical processing and timescales associ-
ated with core-periphery brain network dynamics has rapidly accelerated from primate to hu-
man studies in the quest to understand unifying principles of brain dynamics, hierarchical
Network Neuroscience
308
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
organization, and cognition (Bolton et al., 2020; Gollo et al., 2017; Gollo et al., 2015; Hasson
et al., 2008; Hong et al., 2019; Khambhati et al., 2018; Kiebel et al., 2008; Liégeois et al.,
2019; Nomi, Vij, et al., 2017; Preti & Van De Ville, 2019; Vidaurre et al., 2017; Watanabe
et al., 2019). As such, the application of timescale separation and hierarchy of brain network
dynamics in autism is relatively new, but the findings overlap significantly with findings based
on other dynamical characterizations discussed here. Specifically, in autism, sensory stimuli
often elicit delayed evoked responses in the auditory domain, and integration of multiple local
stimuli into a global percept often requires a wider window of temporal binding (Robertson,
Martin, Baker, and Baron-Cohen, 2012).
In recent work using EEG- and fMRI-based neu-
roimaging evidence, it was found in healthy young individuals that the time differences of
cross-modal perception may particularly tax multisensory processing and temporal dichotomy
of dual-stream processing (streams for action versus stimulus awareness), in which local sen-
sory stimuli must be integrated from two modalities (audio and visual, for example). In particu-
lar during dynamic perception, the sensory signals are built up and integrated over time (Kumar
et al., 2016; Kumar, Dutta, Talwar, Roy, & Banerjee , 2012; Ray et al., 2020).
Hence, there is a growing concurrence among studies that in the autistic brain, the flexibility
of brain dynamics is lost partly because of the distortion of timescales of integration of sensory
input and routing and relaying of these sensory updates to higher order core brain regions for
further processing in a hierarchical manner (Hong et al., 2019; Vidaurre et al., 2017; Watanabe
& Rees, 2017; Watanabe et al., 2019).
including 18 years)
A recent study using children 7–18 years old (not
from ABIDE
(Yao et al., 2016) found that those with ASD showed overall imbalance of strong and weakly
connected brain regions, and decreased functional connectivity associated with DMN hub ar-
eas (precuneus/posterior cingulate gyrus with medial prefrontal gyrus). This study also found
that compared with TD children, the strong connectivity over the peripheral sensory brain
regions was maintained for a longer time (dwell time characterizing longer timescales of
brain state occupation) between brain areas of children with ASD (exhibiting overstability),
and ratios of weaker connectivity strength in brain states varied dramatically (exhibiting tran-
sient stability) in ASD (Yao et al., 2016). Hence, atypical connectivity strengths and states
may be a macroscopical reflection of the excitatory/inhibitory imbalance at the cellular level
(Yao et al., 2016).
Another recent study found reduced transitions between brain states in adults with autism
using energy landscape analysis applied to seven well-established resting-state brain networks
(Watanabe & Rees, 2017). Energy landscape and entropy analysis characterize atypical neural
state transition probabilities between core DMN, executive control network (ECN), ventral at-
tention network (VAN), dorsal attention network (DAN), and peripheral sensory networks and
correlate with symptom severity. To quantify altered brain dynamics and state transitions in
neurotypical individuals compared with ASD, the authors used an accurately fitted pairwise
maximum entropy model (MEM) to estimate the energy landscape (Watanabe & Rees, 2017).
They calculated so-called hypothetical energy values of all the possible brain activity patterns
(in total 27
patterns) among all the known seven resting-state networks (Watanabe & Rees,
2017). They examined hierarchal relationships between the 27
energy values and systemati-
cally searched for dominant brain activity patterns that showed locally minimum energy values
and that were more likely to be observed than similar activity patterns. They discovered that
the TD and ASD groups had energy landscapes with similar hierarchal structures in the above
networks. Their results suggest that direct transitions between major brain states are not differ-
ent among the two groups but indirect transitions are. In individuals with autism, these atypical
neural transitions are rare, which means that their brain dynamics in an energy landscape are
Network Neuroscience
309
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
Intrinsic timescales:
A neuroimaging analysis technique
that allows estimation of an intrinsic
neural timescale of an fMRI signal as
the sum of autocorrelation function
(ACF) values of the signals in the
initial positive period of the ACF. The
period is defined as the area under
the ACF up to the time lag value
just before the one where the ACF
becomes nonpositive for the first
time as the time lag increases.
Subsequently, multiplying the
obtained area under the ACF by the
repetition time (TR) formally defines
the index for the intrinsic timescale.
more stable than in a neurotypical. Thus, their brain activity tends to occupy major brain states
specifically in the visual, sensorimotor, and auditory networks for a sufficiently longer duration
(Watanabe & Rees, 2017). While neurotypical individuals frequently transition between two
major brain states via an intermediate state that has stability, high-functioning adults with ASD
show fewer neural transitions because of an unstable intermediate state, and these infrequent
transitions predict the severity of autism (Watanabe & Rees, 2017).
Another recent finding introduces a very interesting and elegant approach to estimate an
intrinsic neural timescale of an fMRI signal, based on the sum of autocorrelation function (ACF)
values of the signals in the initial positive period of the ACF (Raut et al., 2020; Watanabe et al.,
2019). The period is conceptually defined as an area under the ACF up to the time lag value
(TR = 2 s) just before the one where the ACF becomes nonpositive for the first time as the
time lag increases. The resulting sum when multiplied by the obtained area under the ACF by
the repetition time (TR) is defined as the index of intrinsic timescale associated with a brain
region of interest. Individuals with ASD had significantly shorter intrinsic timescales in bilat-
eral postcentral gyri, right inferior parietal lobule (IPL), right middle insula, bilateral middle
temporal gyri (MTG), and right inferior occipital gyrus (IOG), whereas the intrinsic timescale
in the right caudate was significantly longer in the ASD group compared with the TD group
(Figures 4A–E).The brain integrates incoming information over different timescales that are
characteristic for different regions. Such a hierarchy of timescales also mirrors a hierarchy in
brain structure (based on functional gradients in core-periphery interactions, see next section;
Gollo, 2019; Hong et al., 2019; Raut et al., 2020; Watanabe et al., 2019). Brain regions located
at the top of the hierarchy are represented as large (yellow) circles and have longer timescales.
They are located at the core and have strong connections to other brain regions (Gollo, 2019;
Hong et al., 2019; Raut et al., 2020; Watanabe et al., 2019). Brain regions located at the
periphery are represented by small (blue) circles and have shorter timescales (Gollo, 2019;
Raut et al., 2020; Watanabe et al., 2019). These differences correlate with the severity of
symptoms of ASD. The intrinsic neural timescale in the right caudate was consistently longer
in the ASD group during adolescence, which is interesting given that recent studies reported
that loss of cohesiveness and flexibility in adolescence from right caudate regions could pos-
sibly indicate an impairment in motor coordination and restrictive behavior. In general, the
overdevelopment of the intrinsic timescale was correlated with progression of RRB symptoms
and replicates earlier findings that functional coordination in the brains of adults with ASD un-
derpins overly stable neural dynamics, which supports both ASD core symptoms and cognitive
abilities (Robertson & Baron-Cohen, 2017).
Neural Substrates of Behavioral Variability in Autism: Prediction From Atypical Core-Periphery
Functional Gradients
Specific patterns of brain dynamics along the cortical hierarchy are associated with impair-
ment of sensory and higher order cognitive processing in children, adolescents, and adults.
Recent evidence suggests that mean variability of dFC between the attention and DMN net-
works is positively correlated with the Autism Diagnostic Observation Schedule (ADOS) scores
(Douw, Wakeman, Tanaka, Liu, & Stufflebeam, 2016; Harlalka et al., 2019). This further sug-
gests that intersubject variability is related to symptom severity and behavioral variability in
task performance (Seghier & Price, 2018). Previous findings suggest that higher dFCVar values
indicate better performance in task and poor performance in resting state (Douw et al., 2016).
Similarly, higher variability in the functional connectivity strength of PCC to other DMN areas
(within the same network) in the resting state is related to slower reaction times on a subsequent
attention task (Lin et al., 2016; Liu et al., 2018). The hypervariance in ASD is an interesting
Network Neuroscience
310
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Atypical intrinsic neural timescale in autism. (A) Estimated neural timescale from fMRI BOLD signal, based on sum of autocorre-
lation function. (B) Intrinsic neural timescales are plotted in bilateral middle insula, pre- and postcentral gyrus (exhibiting shorter timescales).
(C) Brain core regions located at the top of the hierarchy are shown in large (yellow) circles and have longer timescales. Brain regions located
at the periphery are represented by small (blue) circles and have shorter timescales. Individuals with autism spectrum disorders (ASD, black)
have different intrinsic timescales (quantified by the autocorrelation function) compared with typically developing individuals (TD, blue). A
schematic displays that noninvasive brain stimulation (black coil) may be used to selectively modulate atypical brain regions to restore their
intrinsic timescales. (D) Intrinsic neural timescales in the right caudate are longer in the ASD group compared with the TD group. (E) The
intrinsic neural timescale in the right caudate is plotted as a function of age in TD (blue) and ASD (red) during adolescence and the correlation
of intrinsic neural timescales with progression of RRB symptoms. Autocorrelation function, ACF; typically developing, TD; autism spectrum
disorder, ASD. Figure adapted with permission from Watanabe et al. (2019).
observation and could potentially lead to a globally disconnected state between sensory and
core brain areas (frontostriatal, fronto-occipital, DMN, SN, etc.). These results taken together
indicate that there could be a relation between the atypical hypervariance in ASD which leads
to an increase in ADOS scores and a decrease in cognitive performance. Recent studies also re-
ported a significant number of hypervariable small-, medium-, and long-range connections in
three groups (children, adolescents, adults) as shown in Figure 2. The long-range connections
define the backbone of the functional network and often connect the hub regions to minimize
wiring and energy costs (Deco et al., 2017; Gollo et al., 2015). In ASD, hypervariance in the
long-range connections could cause instability in information transmission between hubs. In-
terestingly, for adolescents, recent studies found a higher number of hypervariable short-range
connections (Harlalka et al., 2019). The hypervariance in short-range connections could in-
dicate instability of local-module connectivity (Gollo et al., 2015). Further, several nodes, in-
cluding orbitofrontal cortex and caudate, showed both hypervariability in connection strength
and altered modular organization (flexibility) in ASD (Harlalka et al., 2019). A very recent find-
ing further sought to resolve whether alterations in the macroscale hierarchy could provide
a parsimonious explanation of the diverse symptoms (SA, RRB, and sociocognitive deficits)
simultaneously (Hong et al., 2019). This study introduces a very elegant approach to quantify
Network Neuroscience
311
Core-periphery brain network dynamics in autism
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Atypical functional gradient and SFC from periphery to core brain regions in autism. (A) Scatterplot of the first two connectivity
embedding gradients in controls and ASD. Gradient 1 (y-axis) runs from primary sensorimotor (dark turquoise) to transmodal DMN (sienna).
Gradient 2 (x-axis) separates somatomotor and auditory cortex from visual cortex. Triangular scattered points are colored with respect to
established functional communities. Histograms on right show the point density in ASD (light red) and controls (gray), suggesting overall
compression of the first gradient in ASD. (B) Positional shifts of the four significant clusters from the surface-based analysis.
(1) posterior
cingulate cortex (PCC)/precuneus (PCU); (2) middle prefrontal cortex (mPFC); (3) occipito-temporal (OT); (4) posterior middle temporal gyrus
(pMTG). (C) Stepwise functional connectivity (SFC) is estimated in the gradient space. Points are colored with respect to cumulative steps when
simultaneously seeding from primary visual area (V1), somatosensory area (S1), and auditory area (A1). Trajectories (sampled every 20th step)
illustrate the direct SFC from the primary sensory (periphery) seeds to transmodal DMN (core) in controls (left). ASD show an initially more
rapid transition; however, trajectories deflect from a straight path and do not reach the DMN, even after 200 steps. Histograms on the right
show point densities, weighted by the cumulative SFC. Figure adapted with permission from Hong et al. (2019).
Connectome gradient:
A graph signal processing technique
that estimates a low-dimensional
embedding from a high-dimensional
functional connectivity matrix
derived from all brain regions of
interest. In this space, cortical
vertices that are strongly
interconnected by either many
connections or a few very strong
connections are closer together,
whereas vertices with only little or
no interconnectivity are farther apart,
giving a gradient of information flow
in the anatomical connectivity space.
macroscale hierarchy by introducing a novel combination of connectome gradient and step-
wise functional connectivity (SFC) analyses, which offer a complementary characterization of
hierarchical brain anomalies in ASD. The gradient analysis and SFC estimation in ASD allow
visualization of spatial trends in connectivity variations (as displayed in Figure 5) following the
putative cortical hierarchy, while SFC is initiated from a priori selected sensory regions of inter-
est to map stepwise connectivity transitions from peripheral nodes to core DMN nodes. Trans-
modal association cortices functionally shifted more toward peripheral sensory areas in ASD
(Figure 5), a pattern that makes sensory input harder to ignore for participants with ASD. This
may compromise higher order cognitive processing such as mentalization, communication,
and speech processing by preventing the segregation of internally driven cognitive processes
(Jasmin et al., 2019). These works are beginning to unravel the links between brain dynamics
and flexible cognition and core symptoms such as RRBs (Bolton et al., 2020; Sahoo, Pathak,
Deco, Banerjee, & Roy, 2020; Uddin, 2021).
Network Neuroscience
312
Core-periphery brain network dynamics in autism
Stepwise functional connectivity:
Stepwise functional connectivity
(SFC) analysis estimates Pearson
correlation strength between seed
(e.g., sensory-motor) and target
(DMN areas) brain regions as a
sequence of steps in connectivity
space. Usually, for information
reaching from sensory to higher
order cortical areas, seed-to-target
functional connectivity estimation in
principle should converge in fewer
steps (e.g., 20–30 steps).
To date, very few studies have explicitly explored the link between atypical flexibility, func-
tional gradients, and timescales, and their relation to core sensory and higher order cognitive
deficits. The hope is that with development of newer methods in the fields of brain dynam-
ics and unsupervised and supervised machine learning, these types of early neuroimaging
biomarkers may eventually pave the way to move from bench to bedside, and may predict id-
iosyncratic responses to interventions, as well as identify targeted treatment options for ASD.
OUTSTANDING ISSUES AND FUTURE DIRECTIONS
Neuroimaging of Individual Differences to Quantify Brain Dynamics of Atypical
Core-Periphery Interactions
Based on the work we review here, it is quite apparent that understanding of large-scale
brain network dynamics of core and periphery brain regions may provide critical insight into
fundamental cognitive functions and flexibility associated with atypical neurodevelopment
(Lynch et al., 2013; Gollo, 2019; Harlalka et al., 2019; Hong et al., 2019; Lin et al., 2016;
Nomi & Uddin, 2015; Nomi, Vij, et al., 2017; Watanabe et al., 2019). For example, one of the
key questions for future research is why individuals with autism display simultaneous impair-
ment of sensory and higher order cognitive processing (altered local–global processing). We
opine that this question is difficult to address without a proper dynamical framework to study
neuroimaging data from individuals with autism. Past studies were largely unable to provide a
detailed and satisfactory answer to reconcile these empirical observations (Lynch et al., 2013;
Robertson, Martin, Baker, and Baron-Cohen , 2012). One may ask whether much of the higher
order contextual cognitive deficits originate from a temporal processing problem. Altered tem-
poral processing of sensory stimuli is observed in multiple early sensory modalities in autism
(Ashwin, Ashwin, Rhydderch, Howells, & Baron-Cohen, 2009; Baron-Cohen et al., 2009;
Robertson & Baron-Cohen, 2017).
Characterization of individual differences in brain state transitions (Rashid et al., 2018;
Surampudi et al., 2019; Vidaurre et al., 2017; Watanabe & Rees, 2017) and core-periphery
interactions in the brain across timescales and spatial hierarchies (sensory to transmodal cortex)
may provide crucial insight to understand the dual impairments in autistic children, adoles-
cents, and adults (Hahamy et al., 2015; He et al., 2018; Lin et al., 2016; Mottron et al., 2014;
Pelphrey, Morris, and McCarthy, 2005; Watanabe & Rees, 2017; Watanabe et al., 2019). Inter-
estingly, a recent approach going beyond identifying presence of community structure in brain
networks but carrying out topological analysis of core-periphery structures has suggested the
overall organization of whole-brain activity mapped at a single-participant level. This method,
unlike most previous work, was able to successfully track both within- and between-task
transitions from one task block to the next using topological analysis of core-periphery brain
networks at the single-subject level (Saggar et al., 2018). Without the need to collapse neural
data in space or time, one may gain useful information about the brain’s dynamical organi-
zation. Hence, combining topological analysis with atypical sensory timescales may present
a promising avenue to investigate partial and full hub reorganization and contribute to the
neural basis of individual differences and idiosyncrasy in ASD that remains an open question
(Hahamy et al., 2015; Keown et al., 2017).
Recent work further suggests that autistic individuals with greater social impairment would
require greater between-network integration to perform social tasks similarly to TD individuals
(Jasmin et al., 2019). This compensatory neural strategy could be deployed in social situations
outside the laboratory, too. It would be highly interesting to see whether core-periphery brain
dynamics based on flexibility, functional gradients, and atypical timescales of sensory-motor
Network Neuroscience
313
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
areas may provide insights about whether autistic individuals using compensatory neural strate-
gies show similar brain dynamics to neurotypical individuals.
Future studies are needed to systematically address several issues to make valuable infer-
ences and predictions about the early onset of symptoms. First, there is limited generalizability
and replicability across connectivity studies carried out in small and nonoverlapping samples,
as seen from Table 2, and very little inclusion of females (Keown et al., 2017). Also, stud-
ies do not necessarily comprise individuals of varied socioeconomic status, who are largely
underrepresented to date in neuroimaging research (Uddin, 2021).
Consideration of Diagnosis by Age During Maturation of Core-Periphery Brain Networks
There are very few neuroimaging studies to date that have focused on age-wise stratifications
and diagnosis by age while reporting brain connectivity and dynamic alterations observed
in ASD. Additionally, it is completely unknown how the transitions from childhood to the
chronological time that marks the beginning of adolescence influence the development of
brain systems underlying core-periphery dynamics in autism and whether one can entirely
rely on chronological timescales of development (Andreae, 2019; Baron-Cohen et al., 2009;
Cheng et al., 2015; Cohen, 2018; Collignon et al., 2013; Di Martino et al., 2014; Uddin,
2021). Dissociable effects of hormonal changes and age on the adolescent brain have been
well documented, suggesting that pubertal stage may be a better predictor of development
of sensory and cognitive abilities and diversity of behavioral patterns than chronological age
(Baum et al., 2017; Uddin, 2021).
In this context, cognitive neuroscience work on ASD of
children, adolescents, and adults needs to consider both hormonal changes during maturation
as well as precise neuromodulatory effects on brain, behavior, and cognition (Liu et al., 2018;
Reijmer et al., 2015; Shine, 2019). The findings of atypical network hierarchy, flexibility, and
timescales of local sensory areas offer a novel and parsimonious account of the range of symp-
toms observed in ASD that encompasses multiple domains across sensory-motor, cognitive,
and social-communicative functioning (Harlalka et al., 2019; Hong et al., 2019; Watanabe
& Rees, 2017; Watanabe et al., 2019).
In addition, the shorter intrinsic timescale found in
the primary sensory/visual areas in autism was correlated with the overall severity of RRB in
autism (Watanabe et al., 2019). In fact, the signal variability and the longer intrinsic timescale
observed in the caudate nucleus were associated with the severity of RRBs in autistic indi-
viduals (Watanabe & Rees, 2017; Watanabe et al., 2019). In addition, a significant diagnosis
by age interaction effect was observed in cohesion strength primarily in right caudate regions
in autism but not in the left caudate (Figure 3). Taking together longer timescales and re-
duced cohesion strength particularly in the right caudate regions in ASD suggests that local
processing in subcortical areas is significantly altered (Harlalka et al., 2019; Watanabe & Rees,
2017; Watanabe et al., 2019). Hence, future task-based neuroimaging studies need to focus on
properly characterizing neural substrates of local subcortical regions and their role in atypical
cognitive impairment. However, temporal properties of local neural signals have already
been linked to local gray matter volumes (Watanabe et al., 2019). These crucial findings
indicate the possibility that functional and structural properties in local brain areas and their
strong coupling in sensory areas as opposed to divergence from tighter coupling in higher
order transmodal areas (Baum et al., 2017; Murphy et al., 2018; Preti & Van De Ville, 2019;
Reijmer et al., 2015; Vázquez-Rodríguez et al., 2019) could have a critical influence on higher
order cognitive symptoms in autism (Cerliani et al., 2015; B. Chen et al., 2020; Kana et al.,
2007). Furthermore, this possibility could be realized using gradient and SFC feature estima-
tion techniques that could guide a supervised learning algorithm to predict symptom severity
in individuals with ASD subtypes (Hong et al., 2019; Jao Keehn et al., 2019; Keown et al.,
Network Neuroscience
314
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
2017; Reiter et al., 2019; Uddin et al., 2013). These predictive features may for the first time
allow us to understand the role of DMN, language areas in STG, and multisensory areas for
processing dynamic stimuli in STS (transmodal cortex) in conjunction with peripheral sensory
(auditory, visual, sensory-motor) and subcortical systems (thalamus, caudate, putamen) during
maturation. This could resolve the puzzle of how both sensory and higher order aspects of the
cortical hierarchy underpin ASD symptomatology.
CONCLUSIONS
Core-periphery brain dynamics and atypical timescales of processing may facilitate optimal
systems-level functioning in ASD. Studies of the neural mechanisms during various devel-
opment stages are yet to be characterized comprehensively to fully understand atypical de-
velopment and behavioral variability. To this end, one of the key challenges that remains is
to understand how sensory deficits lead to higher order cognitive deficits in the domain of
language, memory, attention, self-related processing, and executive functioning in ASD. This
review provides an integrative view to explain simultaneity of sensory and cognitive deficits in
autism, parsing through evidence from recent topological network-analysis-based approaches.
More specifically, it highlights evidence that sensory and higher order cognitive deficits could
be explained by atypical network flexibility, functional gradients, and timescale hierarchy in
the cortex.
Moving forward, intrinsic timescales can be estimated using simple autocorrelations, which
may be used to identify biomarkers and to improve our understanding of disease subtypes
and treatment plans (Gollo, 2019). However, more research is necessary to fully compre-
hend and uncover the causes and implications of atypical intrinsic timescales. In individuals
with autism, shorter timescales in unimodal sensory areas could relate to a heightened sen-
sory perception and finer sensory discrimination, which is in line with excessive expectation
of changes from the sensory environment. Moreover, longer timescales and reduced cohe-
siveness found in the caudate nucleus might also indicate a neural compensation strategy to
deal with loaded sensory input due to heightened sensory perception and discrimination. In
addition, not just sensory perception tasks but behaviorally and cognitively relevant social
tasks can be carefully constructed to deal with varying cognitive demands in the service of
attention and executive functions to allow atypical groups to carry out flexible task switch-
ing. The quantification of state-invariant core processing and state-dependent contextual pro-
cessing can serve as a starting point for the characterization of a broader set of behavioral
subtypes in autism. Translating these recent results into clinical practice will involve many
practical challenges, but will be highly beneficial for furthering the neuroscience of autism.
AUTHOR CONTRIBUTIONS
Dipanjan Roy: Conceptualization;
Investigation; Methodology; Project administration;
Visualization; Writing – original draft; Writing – review & editing. Lucina Q. Uddin: Con-
ceptualization; Investigation; Methodology; Visualization; Writing – original draft; Writing –
review & editing.
FUNDING INFORMATION
Dipanjan Roy, Department of Biotechnology, Government of India, Award ID: BT/RLF/
Re-entry/07/2014. Dipanjan Roy, Department of Science and Technology (DST) Ministry of
Science and Technology, Government of India, Award ID: SR/CSRI/21/2016. Dipanjan Roy,
BT/MED-III/NBRC/Flagship/Program/2019. Lucina Q. Uddin, National Institute of Mental
Network Neuroscience
315
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
Health, Award ID: R01MH107549. Lucina Q. Uddin, Canadian Institute for Advanced Re-
search. Lucina Q. Uddin, Gabelli Senior Scholar Award, University of Miami.
REFERENCES
Abbott, A. E., Linke, A. C., Nair, A.,
Jahedi, A., Alba, L. A.,
(2018). Repetitive behaviors
Keown, C. L., . . . Müller, R. A.
in autism are linked to imbalance of corticostriatal connectivity:
A functional connectivity MRI study. Social Cognitive and Affec-
tive Neuroscience, 13(1), 32–42. DOI: https://doi.org/10.1093
/scan/nsx129, PMID: 29177509, PMCID: PMC5793718
Alaerts, K., Nayar, K., Kelly, C., Raithel,
J., Milham, M. P., &
Di Martino, A. (2015). Age-related changes in intrinsic function
of the superior temporal sulcus in autism spectrum disorders. So-
cial Cognitive and Affective Neuroscience, 10(10), 1413–1423.
DOI: https://doi.org/10.1093/scan/nsv029, PMID: 25809403,
PMCID: PMC4590540
Alaerts, K., Swinnen, S. P., & Wenderoth, N. (2016). Sex differ-
ences in autism: A resting-state fMRI investigation of functional
brain connectivity in males and females. Social Cognitive and Af-
fective Neuroscience, 11(6), 1002–1016. DOI: https://doi.org/10
.1093/scan/nsw027, PMID: 26989195, PMCID: PMC4884321
Alaerts, K., Woolley, D. G., Steyaert, J., Di Martino, A., Swinnen,
S. P., & Wenderoth, N. (2014). Underconnectivity of the su-
perior temporal sulcus predicts emotion recognition deficits
in autism. Social Cognitive and Affective Neuroscience, 9(10),
1589–1600. DOI: https://doi.org/10.1093/scan/nst156, PMID:
24078018, PMCID: PMC4187281
Anderson, J. S., Druzgal, T. J., Froehlich, A., DuBray, M. B., Lange,
N., Alexander, A. L., . . . Bigler, E. D. (2011). Decreased inter-
hemispheric functional connectivity in autism. Cerebral Cortex,
21(5), 1134–1146. DOI: https://doi.org/10.1093/cercor/bhq190,
PMID: 20943668, PMCID: PMC3077433
Andreae, L. C.
(2019). Brain development
in autism: Timing
is everything. Science Translational Medicine, 11(476). DOI:
https://doi.org/10.1126/scitranslmed.aaw5314
Ashwin, E., Ashwin, C., Rhydderch, D., Howells, J., & Baron-
Cohen, S. (2009). Eagle-eyed visual acuity: An experimental
investigation of enhanced perception in autism. Biological Psy-
chiatry, 65(1), 17–21. DOI: https://doi.org/10.1016/j.biopsych
.2008.06.012, PMID: 18649873
Atasoy, S., Donnelly, I., & Pearson, J. (2016). Human brain networks
function in connectome-specific harmonic waves. Nature Com-
munications, 7, 10340. DOI: https://doi.org/10.1038/ncomms10340,
PMID: 26792267, PMCID: PMC4735826
Baron-Cohen, S., Ashwin, E., Ashwin, C., Tavassoli, T., &
(2009). Talent in autism: Hyper-systemizing,
Chakrabarti, B.
hyper-attention to detail and sensory hypersensitivity. Philoso-
phical Transactions of the Royal Society B: Biological Sciences,
364(1522), 1377–1383. DOI: https://doi.org/10.1098/rstb.2008
.0337, PMID: 19528020, PMCID: PMC2677592
Baum, G. L., Ciric, R., Roalf, D. R., Betzel, R. F., Moore, T. M.,
Shinohara, R. T., . . . Cook, P. A. (2017). Modular segregation of
structural brain networks supports the development of executive
function in youth. Current Biology, 27(11), 1561–1572. DOI:
https://doi.org/10.1016/j.cub.2017.04.051, PMID: 28552358,
PMCID: PMC5491213
Blakemore, S. J., Burnett, S., & Dahl, R. E. (2010). The role of pu-
berty in the developing adolescent brain. Human Brain Map-
ping, 31(6), 926–933. DOI: https://doi.org/10.1002/hbm.21052,
PMID: 20496383, PMCID: PMC3410522
Bolton, T. A., Morgenroth, E., Preti, M. G., & Van De Ville, D.
(2020). Tapping into multi-faceted human behavior and psycho-
pathology using fMRI brain dynamics. Trends in Neurosciences,
43(9), 667–680. DOI: https://doi.org/10.1016/j.tins.2020.06.005,
PMID: 32682563
Cerliani, L., Mennes, M., Thomas, R. M., Di Martino, A., Thioux,
(2015). Increased functional connectivity
M., & Keysers, C.
between subcortical and cortical
resting-state networks in
autism spectrum disorder. JAMA Psychiatry, 72(8), 767–777.
DOI: https://doi.org/10.1001/jamapsychiatry.2015.0101, PMID:
26061743, PMCID: PMC5008437
Chang, C., & Glover, G. H. (2010). Time-frequency dynamics of
resting-state brain connectivity measured with fMRI. Neuro-
Image, 50(1), 81–98. DOI: https://doi.org/10.1016/j.neuroimage
.2009.12.011, PMID: 20006716, PMCID: PMC2827259
Chaudhuri, R., Knoblauch, K., Gariel, M. A., Kennedy, H., &
Wang, X. J. (2015). A large-scale circuit mechanism for hierarchi-
cal dynamical processing in the primate cortex. Neuron, 88(2),
419–431. DOI: https://doi.org/10.1016/j.neuron.2015.09.008,
PMID: 26439530, PMCID: PMC4630024
Chen, B., Linke, A., Olson, L., Ibarra, C., Reynolds, S., Müller, R. A.,
. . . Fishman, I.
(2020). Greater functional connectivity be-
tween sensory networks is related to symptom severity in tod-
dlers with autism spectrum disorder. Journal of Child Psychology
and Psychiatry. DOI: https://doi.org/10.1111/jcpp.13268, PMID:
PMC7688487
Chen, H., Nomi, J. S., Uddin, L. Q., Duan, X., & Chen, H. (2017).
Intrinsic functional connectivity variance and state-specific
under-connectivity in autism. Human Brain Mapping, 38(11),
5740–5755. DOI: https://doi.org/10.1002/hbm.23764, PMID:
28792117, PMCID: PMC5783325
Cheng, W., Rolls, E. T., Gu, H., Zhang, J., & Feng, J. (2015). Autism:
Reduced connectivity between cortical areas involved in face ex-
pression, theory of mind, and the sense of self. Brain, 138(5),
1382–1393. DOI: https://doi.org/10.1093/brain/awv051, PMID:
25795704, PMCID: PMC4407191
Ciarrusta,
J., O’Muircheartaigh,
J., Dimitrova, R., Batalle, D.,
Cordero-Grande, L., Price, A., . . . Javed, A. (2019). Social brain
functional maturation in newborn infants with and without a
family history of autism spectrum disorder. JAMA Network Open,
2(4), e191868. DOI: https://doi.org/10.1001/jamanetworkopen
.2019.1868, PMID: 30951164, PMCID: PMC6450332
Cohen, J. R. (2018). The behavioral and cognitive relevance of time-
varying, dynamic changes in functional connectivity. NeuroImage,
Network Neuroscience
316
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
180, 515–525. DOI: https://doi.org/10.1016/j.neuroimage.2017
.09.036, PMID: 28942061, PMCID: PMC6056319
roImage, 190, 191–204. DOI: https://doi.org/10.1016/j.neuroimage
.2018.06.003, PMID: 29883735, PMCID: PMC6281849
Collignon, O., Charbonneau, G., Peters, F., Nassim, M., Lassonde,
(2013). Reduced multisensory
M., Lepore, F., . . . Bertone, A.
facilitation in persons with autism. Cortex, 49(6), 1704–1710.
DOI:
PMID:
https://doi.org/10.1016/j.cortex.2012.06.001,
22818902
Courchesne, E., Campbell, K., & Solso, S.
(2011). Brain growth
across the life span in autism: Age-specific changes in anatomi-
cal pathology. Brain Research, 1380, 138–145. DOI: https://doi
.org/10.1016/j.brainres.2010.09.101, PMID: 20920490, PMCID:
PMC4500507
Dajani, D. R., Odriozola, P., Winters, M., Voorhies, W., Marcano,
S., Baez, A., . . . Uddin, L. Q.
(2020). Measuring cognitive
flexibility with the flexible item selection task: From fMRI adap-
Journal of Cognitive
tation to individual connectome mapping.
Neuroscience, 32(6), 1026–1045. DOI: https://doi.org/10.1162
/jocn_a_01536, PMID: 32013686
Deco, G., Kringelbach, M. L., Jirsa, V. K., & Ritter, P. (2017).
The dynamics of resting fluctuations in the brain: Metastabil-
ity and its dynamical cortical core. Scientific Reports, 7(1), 1–14.
PMID:
https://doi.org/10.1038/s41598-017-03073-5,
DOI:
28596608, PMCID: PMC5465179
Delmonte, S., Gallagher, L., O’Hanlon, E., McGrath,
J., &
Balsters, J. H. (2013). Functional and structural connectivity of
frontostriatal circuitry in autism spectrum disorder. Frontiers in
Human Neuroscience, 7, 430. DOI: https://doi.org/10.3389
/fnhum.2013.00430, PMID: 23964221, PMCID: PMC3734372
Di Martino, A., O’Connor, D., Chen, B., Alaerts, K., Anderson, J. S.,
Assaf, M., . . . Blanken, L. M. (2017). Enhancing studies of the
connectome in autism using the Autism Brain Imaging Data
Exchange II. Scientific Data, 4(1), 1–15. DOI: https://doi.org/10
.1038/sdata.2017.10, PMID: 28291247, PMCID: PMC5349246
Di Martino, A., Yan, C. G., Li, Q., Denio, E., Castellanos, F. X.,
Alaerts, K., . . . Deen, B. (2014). The Autism Brain Imaging Data
Exchange: Towards a large-scale evaluation of the intrinsic brain
architecture in autism. Molecular Psychiatry, 19(6), 659–667.
DOI: https://doi.org/10.1038/mp.2013.78, PMID: 23774715,
PMCID: PMC4162310
Douw, L., Wakeman, D. G., Tanaka, N., Liu, H., & Stufflebeam,
S. M. (2016). State-dependent variability of dynamic functional
connectivity between frontoparietal and default networks relates
to cognitive flexibility. Neuroscience, 339, 12–21. DOI: https://
doi.org/10.1016/j.neuroscience.2016.09.034, PMID: 27687802,
PMCID: PMC5635855
Foxe, J. J., Molholm, S., Del Bene, V. A., Frey, H. P., Russo, N. N.,
Blanco, D., . . . Ross, L. A. (2015). Severe multisensory speech in-
tegration deficits in high-functioning school-aged children with
autism spectrum disorder (ASD) and their resolution during early
adolescence. Cerebral Cortex, 25(2), 298–312. DOI: https://doi.org
/10.1093/cercor/bht213, PMID: 23985136, PMCID: PMC4303800
Friston, K.
(2009). The free-energy principle: A rough guide to
the brain? Trends in Cognitive Sciences, 13(7), 293–301. DOI:
https://doi.org/10.1016/j.tics.2009.04.005, PMID: 19559644
.
.
Calhoun, V. D. (2019). Transient increased thalamic-sensory con-
nectivity and decreased whole-brain dynamism in autism. Neu-
Fu, Z., Tu, Y., Di, X., Du, Y., Sui,
J., Biswal, B. B.,
.
Gollo, L. L. (2019). Computational psychiatry: Exploring atypical
timescales in the brain. eLife, 8, e45089. DOI: https://doi.org/10
.7554/eLife.45089, PMID: 30717825, PMCID: PMC6363382
Gollo, L. L., Roberts, J. A., & Cocchi, L. (2017). Mapping how local
perturbations influence systems-level brain dynamics. Neuro-
Image, 160, 97–112. DOI: https://doi.org/10.1016/j.neuroimage
.2017.01.057, PMID: 28126550
Gollo, L. L., Zalesky, A., Hutchison, R. M., van den Heuvel, M.,
& Breakspear, M. (2015). Dwelling quietly in the rich club: Brain
network determinants of slow cortical fluctuations. Philosophi-
cal Transactions of the Royal Society B: Biological Sciences,
370(1668), 20140165. DOI: https://doi.org/10.1098/rstb.2014
.0165, PMID: 25823864, PMCID: PMC4387508
Guo, X., Chen, H., Long, Z., Duan, X., Zhang, Y., & Chen, H.
(2017). Atypical developmental trajectory of local spontaneous
brain activity in autism spectrum disorder. Scientific Reports,
7(1), 1–10. DOI: https://doi.org/10.1038/srep39822, PMID:
28057930, PMCID: PMC5216408
Guo, X., Duan, X., Chen, H., He, C., Xiao, J., Han, S., . . .
Chen, H. (2020). Altered inter-and intrahemispheric functional
connectivity dynamics in autistic children. Human Brain Map-
ping, 41(2), 419–428. DOI: https://doi.org/10.1002/hbm.24812,
PMID: 31600014, PMCID: PMC7268059
Guo, X., Duan, X., Suckling, J., Chen, H., Liao, W., Cui, Q.,
& Chen, H. (2019). Partially impaired functional connectiv-
ity states between right anterior insula and default mode net-
work in autism spectrum disorder. Human Brain Mapping, 40(4),
1264–1275. DOI: https://doi.org/10.1002/hbm.24447, PMID:
30367744, PMCID: PMC6865537
Griffa, A., & van den Heuvel, M. P.
(2018). Rich-club neurocir-
cuitry: Function, evolution, and vulnerability. Dialogues in Clini-
cal Neuroscience, 20(2), 121. DOI: https://doi.org/10.31887/DCNS
.2018.20.2/agriffa, PMID: 30250389, PMCID: PMC6136122
Hahamy, A., Behrmann, M., & Malach, R. (2015). The idiosyn-
cratic brain: Distortion of spontaneous connectivity patterns
in autism spectrum disorder. Nature Neuroscience, 18(2), 302.
DOI: https://doi.org/10.1038/nn.3919, PMID: 25599222
Harlalka, V., Bapi, R. S., Vinod, P. K., & Roy, D.
(2018). Age,
disease, and their interaction effects on intrinsic connectivity of
children and adolescents in autism spectrum disorder using func-
tional connectomics. Brain Connectivity, 8(7), 407–419. DOI:
https://doi.org/10.1089/brain.2018.0616, PMID: 30009617
Harlalka, V., Bapi, R. S., Vinod, P. K., & Roy, D.
(2019). Atypi-
cal flexibility in dynamic functional connectivity quantifies the
severity in autism spectrum disorder. Frontiers in Human Neuro-
science, 13, 6. DOI: https://doi.org/10.3389/fnhum.2019.00006,
PMID: 30774589, PMCID: PMC6367662
Hasson, U., Yang, E., Vallines, I., Heeger, D. J., & Rubin, N. (2008).
A hierarchy of temporal receptive windows in human cortex.
Journal of Neuroscience, 28(10), 2539–2550. DOI: https://doi
.org/10.1523/JNEUROSCI.5487-07.2008, PMID: 18322098,
PMCID: PMC2556707
He, C., Chen, Y., Jian, T., Chen, H., Guo, X., Wang, J., . . . Duan,
X. (2018). Dynamic functional connectivity analysis reveals de-
creased variability of the default-mode network in developing
Network Neuroscience
317
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
autistic brain. Autism Research, 11(11), 1479–1493. DOI:
https://doi.org/10.1002/aur.2020, PMID: 30270547
Henry, T. R., Dichter, G. S., & Gates, K.
(2018). Age and
gender effects on intrinsic connectivity in autism using func-
tional integration and segregation. Biological Psychiatry: Cog-
nitive Neuroscience and Neuroimaging, 3(5), 414–422. DOI:
https://doi.org/10.1016/j.bpsc.2017.10.006, PMID: 29735152
Hilgetag, C. C., & Goulas, A. (2020). ‘Hierarchy’ in the organization
of brain networks. Philosophical Transactions of the Royal Society
B, 375(1796), 20190319. DOI: https://doi.org/10.1098/rstb.2019
.0319, PMID: 32089116, PMCID: PMC7061955
Hong, S-J., Vos De Wael, R., Bethlehem, R. A. I., Lariviere, S.,
Paquola, C., Valk, S. L., . . . Bernhardt, B. C. (2019). Atypical func-
tional connectome hierarchy in autism. Nature Communications,
10(1), 1–13. DOI: https://doi.org/10.1038/s41467-019-08944-1,
PMID: 30833582, PMCID: PMC6399265
Huang, H., Shu, N., Mishra, V., Jeon, T., Chalak, L., Wang, Z. J.,
(2015). Development of human brain structural
. . . Dong, Q.
networks through infancy and childhood. Cerebral Cortex, 25(5),
1389–1404. DOI: https://doi.org/10.1093/cercor/bht335, PMID:
24335033, PMCID: PMC4397575
Jao Keehn, R.
J., Nair, S., Pueschel, E. B., Linke, A. C.,
Fishman, I., & Müller, R. A. (2019). Atypical local and distal
patterns of occipito-frontal functional connectivity are related to
symptom severity in autism. Cerebral Cortex, 29(8), 3319–3330.
DOI: https://doi.org/10.1093/cercor/bhy201, PMID: 30137241,
PMCID: PMC7342606
Jao Keehn, R. J., Sanchez, S. S., Stewart, C. R., Zhao, W., Grenesko-
Stevens, E. L., Keehn, B., & Müller, R. A. (2017). Impaired
downregulation of visual cortex during auditory processing is
associated with autism symptomatology in children and ado-
lescents with autism spectrum disorder. Autism Research, 10(1),
130–143. DOI: https://doi.org/10.1002/aur.1636, PMID: 27205875,
PMCID: PMC5892834
Jasmin, K., Gotts, S. J., Xu, Y., Liu, S., Riddell, C. D., Ingeholm, J. E.,
. . . Martin, A. (2019). Overt social interaction and resting
state in young adult males with autism: Core and contextual
neural features. Brain, 142(3), 808–822. DOI: https://doi.org/10
.1093/brain/awz003, PMID: 30698656, PMCID: PMC6391610
Kana, R. K., Keller, T. A., Minshew, N. J., & Just, M. A. (2007).
Inhibitory control in high-functioning autism: Decreased activa-
tion and underconnectivity in inhibition networks. Biological Psy-
chiatry, 62(3), 198–206. DOI: https://doi.org/10.1016/j.biopsych
.2006.08.004, PMID: 17137558, PMCID: PMC4492460
Kenny, L., Cribb, S. J., & Pellicano, E.
(2019). Childhood exec-
utive function predicts later autistic features and adaptive be-
havior in young autistic people: A 12-year prospective study.
Journal of Abnormal Child Psychology, 47(6), 1089–1099. DOI:
https://doi.org/10.1007/s10802-018-0493-8, PMID: 30421376
Keown, C. L., Datko, M. C., Chen, C. P., Maximo, J. O., Jahedi, A.,
& Müller, R. A. (2017). Network organization is globally atypical
in autism: A graph theory study of intrinsic functional connect-
ivity. Biological Psychiatry: Cognitive Neuroscience and Neu-
roimaging, 2(1), 66–75. DOI: https://doi.org/10.1016/j.bpsc.2016
.07.008, PMID: 28944305, PMCID: PMC5607014
Khambhati, A. N., Medaglia,
J. D., Karuza, E. A., Thompson-
Schill, S. L., & Bassett, D. S. (2018). Subgraphs of functional
brain networks identify dynamical constraints of cognitive control.
PLoS Computational Biology, 14(7), e1006234. DOI: https://doi
.org/10.1371/journal.pcbi.1006234, PMID: 29979673, PMCID:
PMC6056061
Kiebel, S. J., Daunizeau, J., & Friston, K. J. (2008). A hierarchy of
timescales and the brain. PLoS Computational Biology, 4(11),
e1000209. DOI: https://doi.org/10.1371/journal.pcbi.1000209,
PMID: 19008936, PMCID: PMC2568860
King, J. B., Prigge, M. B., King, C. K., Morgan, J., Weathersby, F., Fox,
J. C., . . . Bigler, E. D. (2019). Generalizability and reproducibil-
ity of functional connectivity in autism. Molecular Autism, 10(1),
27. DOI: https://doi.org/10.1186/s13229-019-0273-5, PMID:
31285817, PMCID: PMC6591952
Kumar, V. G., Halder, T., Jaiswal, A. K., Mukherjee, A., Roy, D.,
& Banerjee, A. (2016). Large scale functional brain networks
underlying temporal integration of audio-visual speech percep-
tion: An EEG study. Frontiers in Psychology, 7, 1558. DOI:
https://doi.org/10.3389/fpsyg.2016.01558, PMID: 27790169,
PMCID: PMC5062921
Kumar, V. G., Dutta, S., Talwar, S., Roy, D., & Banerjee, A. (2020).
Biophysical mechanisms governing large-scale brain network
dynamics underlying individual-specific variability of percep-
tion. European Journal of Neuroscience, 52(7), 3746–3762. DOI:
https://doi.org/10.1111/ejn.14747, PMID: 32304122
Lai, M. C., Lerch,
J. P., Floris, D. L., Ruigrok, A. N., Pohl,
Imaging
A., Lombardo, M. V., & Baron-Cohen, S.
sex/gender and autism in the brain: Etiological implications.
Journal of Neuroscience Research, 95(1–2), 380–397. DOI:
https://doi.org/10.1002/jnr.23948, PMID: 27870420
(2017).
Laumann, T. O., Snyder, A. Z., Mitra, A., Gordon, E. M., Gratton,
C., Adeyemo, B., . . . McCarthy, J. E. (2017). On the stability of
BOLD fMRI correlations. Cerebral Cortex, 27(10), 4719–4732.
Lawson, R. A., Papadakis, A. A., Higginson, C.
I., Barnett,
J. E., Wills, M. C., Strang,
. . . Kenworthy, L. (2015).
J. F.,
Everyday executive function impairments predict comorbid
psychopathology in autism spectrum and attention deficit
hyperactivity disorders. Neuropsychology, 29(3), 445. DOI:
https://doi.org/10.1037/neu0000145, PMID: 25313979
Liégeois, R., Li, J., Kong, R., Orban, C., Van De Ville, D., Ge, T.,
. . . Yeo, B. T. (2019). Resting brain dynamics at different time-
scales capture distinct aspects of human behavior. Nature Com-
munications, 10(1), 1–9. DOI: https://doi.org/10.1038/s41467
-019-10317-7, PMID: 31127095, PMCID: PMC6534566
Lin, P., Yang, Y., Jovicich, J., De Pisapia, N., Wang, X., Zuo, C. S., &
Levitt, J. J. (2016). Static and dynamic posterior cingulate cortex
nodal topology of default mode network predicts attention task
performance. Brain Imaging and Behavior, 10, 212–225. DOI:
https://doi.org/10.1007/s11682-015-9384-6, PMID: 25904156
Liu,
J., Liao, X., Xia, M., & He, Y.
(2018). Chronnectome
fingerprinting:
Identifying individuals and predicting higher
cognitive functions using dynamic brain connectivity patterns.
Human Brain Mapping, 39(2), 902–915. DOI: https://doi.org/10
.1002/hbm.23890, PMID: 29143409, PMCID: PMC6866558
London, E. B. (2018). Neuromodulation and a reconceptualization
of autism spectrum disorders: Using the locus coeruleus func-
tioning as an exemplar. Frontiers in Neurology, 9, 1120. DOI:
Network Neuroscience
318
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
https://doi.org/10.3389/fneur.2018.01120, PMID: 30619071,
PMCID: PMC6305710
136(6), 1942–1955. DOI: https://doi.org/10.1093/brain/awt079,
PMID: 23739917, PMCID: PMC3673456
Lynch, C. J., Uddin, L. Q., Supekar, K., Khouzam, A., Phillips, J.,
& Menon, V. (2013). Default mode network in childhood autism:
posteromedial cortex heterogeneity and relationship with social
deficits. Biological Psychiatry, 74(3), 212–219. DOI: https://doi
.org/10.1016/j.biopsych.2012.12.013, PMID: 23375976, PMCID:
PMC3710546
Manning, C., Tibber, M. S., Charman, T., Dakin, S. C., & Pellicano,
E. (2015). Enhanced integration of motion information in child-
Journal of Neuroscience, 35(18), 6979–6986.
ren with autism.
DOI: https://doi.org/10.1523/JNEUROSCI.4645-14.2015, PMID:
25948250, PMCID: PMC4420775
Mash, L. E., Reiter, M. A., Linke, A. C., Townsend, J., & Müller,
R. A.
(2018). Multimodal approaches to functional connec-
tivity in autism spectrum disorders: An integrative perspective.
Developmental Neurobiology, 78(5), 456–473. DOI: https://
https://doi.org/10.1002/dneu.22570, PMID: 29266810, PMCID:
PMC5897150
McKinnon, C. J., Eggebrecht, A. T., Todorov, A., Wolff, J. J., Elison,
J. T., Adams, C. M., . . . McKinstry, R. C.
(2019). Restricted
and repetitive behavior and brain functional connectivity in in-
fants at risk for developing autism spectrum disorder. Biologi-
cal Psychiatry: Cognitive Neuroscience and Neuroimaging, 4(1),
50–61. DOI: https://doi.org/10.1016/j.bpsc.2018.09.008, PMID:
30446435, PMCID: PMC6557405
Mottron, L., Belleville, S., Rouleau, G. A., & Collignon, O. (2014).
Linking neocortical, cognitive, and genetic variability in autism
with alterations of brain plasticity: The Trigger-Threshold-Target
model. Neuroscience and Biobehavioral Reviews, 47, 735–752.
DOI: https://doi.org/10.1016/j.neubiorev.2014.07.012, PMID:
25155242
Moul, C., Cauchi, A., Hawes, D. J., Brennan, J., & Dadds, M. R.
(2015). Differentiating autism spectrum disorder and overlapping
psychopathology with a brief version of the social responsive-
ness scale. Child Psychiatry and Human Development, 46(1),
108–117. DOI: https://doi.org/10.1007/s10578-014-0456-4,
PMID: 24604214
Murphy, C.,
from input:
Jefferies, E., Rueschemeyer, S. A., Sormaz, M.,
(2018).
Wang, H. T., Margulies, D. S., & Smallwood,
Distant
regions within the de-
fault mode network supporting perceptually-decoupled and
conceptually-guided cognition. NeuroImage, 171, 393–401.
DOI: https://doi.org/10.1016/j.neuroimage.2018.01.017, PMID:
29339310, PMCID: PMC5883322
Evidence of
J.
Naik, S., Banerjee, A., Bapi, R. S., Deco, G., & Roy, D.
(2017).
Metastability in senescence. Trends in Cognitive Sciences, 21(7),
509–521. DOI: https://doi.org/10.1016/j.tics.2017.04.007, PMID:
28499740
Naik, S., Subbareddy, O., Banerjee, A., Roy, D., & Bapi, R. S.
(2017). Metastability of cortical BOLD signals in maturation and
senescence. In 2017 International Joint Conference on Neural
Networks (IJCNN) (pp. 4564–4570). IEEE. DOI: https://doi.org/10
.1109/IJCNN.2017.7966435
Nair, A., Treiber, J. M., Shukla, D. K., Shih, P., & Müller, R. A. (2013).
Impaired thalamocortical connectivity in autism spectrum dis-
order: A study of functional and anatomical connectivity. Brain,
Noel, J. P., De Niear, M. A., Stevenson, R., Alais, D., & Wallace,
M. T. (2017). Atypical rapid audio-visual temporal recalibration
in autism spectrum disorders. Autism Research, 10(1), 121–129.
DOI: https://doi.org/10.1002/aur.1633, PMID: 27156926
Nomi, J. S., Bolt, T. S., Ezie, C. C., Uddin, L. Q., & Heller, A. S.
(2017). Moment-to-moment BOLD signal variability reflects
regional changes in neural flexibility across the lifespan. Journal
of Neuroscience, 37(22), 5539–5548. DOI: https://doi.org/10
.1523/JNEUROSCI.3408-16.2017, PMID: 28473644, PMCID:
PMC5452342
Nomi, J. S., & Uddin, L. Q. (2015). Developmental changes in large-
scale network connectivity in autism. NeuroImage: Clinical,
7, 732–741. DOI: https://doi.org/10.1016/j.nicl.2015.02.024,
PMID: 25844325, PMCID: PMC4375789
Nomi, J. S., Vij, S. G., Dajani, D. R., Steimke, R., Damaraju, E.,
Rachakonda, S., . . . Uddin, L. Q. (2017). Chronnectomic pat-
terns and neural flexibility underlie executive function. Neuro-
Image, 147, 861–871. DOI: https://doi.org/10.1016/j.neuro
image.2016.10.026, PMID: 27777174, PMCID: PMC5303676
Oldham, S., & Fornito, A.
(2019). The development of brain
network hubs. Developmental Cognitive Neuroscience, 36,
100607. DOI: https://doi.org/10.1016/j.dcn.2018.12.005, PMID:
30579789, PMCID: PMC6969262
Olson, L. A., Mash, L. E., Linke, A., Fong, C. H., Müller, R. A.,
& Fishman, I.
(2020). Sex-related patterns of intrinsic func-
tional connectivity in children and adolescents with autism spec-
trum disorders. Autism, 1362361320938194. DOI: https://doi
.org/10.1177/1362361320938194, PMID: 32689820, PMCID:
PMC7541740
Ostrolenk, A., Bao, V. A., Mottron, L., Collignon, O., & Bertone,
A.
(2019). Reduced multisensory facilitation in adolescents
and adults on the autism spectrum. Scientific Reports, 9(1),
1–9. DOI: https://doi.org/10.1038/s41598-019-48413-9, PMID:
31427634, PMCID: PMC6700191
Padmanabhan, A., Lynch, C. J., Schaer, M., & Menon, V.
(2017).
The default mode network in autism. Biological Psychiatry: Cog-
nitive Neuroscience and Neuroimaging, 2(6), 476–486. DOI:
https://doi.org/10.1016/j.bpsc.2017.04.004, PMID: 29034353,
PMCID: PMC5635856
Pelphrey, K. A., Morris, J. P., & McCarthy, G. (2005). Neural basis of
eye gaze processing deficits in autism. Brain, 128(5), 1038–1048.
DOI: https://doi.org/10.1093/brain/awh404, PMID: 15758039
Pillai, A. S., & Jirsa, V. K.
(2017). Symmetry breaking in space-
time hierarchies shapes brain dynamics and behavior. Neuron,
94(5), 1010–1026. DOI: https://doi.org/10.1016/j.neuron.2017
.05.013, PMID: 28595045
Plaisted, K., O’Riordan, M., & Baron-Cohen, S. (1998). Enhanced
visual search for a conjunctive target in autism: A research note.
Journal of Child Psychology and Psychiatry, 39(5), 777–783.
DOI: https://doi.org/10.1111/1469-7610.00376, https://doi.org
/10.1017/S0021963098002613, PMID: 9690940
Preti, M. G., Bolton, T. A., & Van De Ville, D. (2017). The dynamic
functional connectome: State-of-the-art and perspectives. Neu-
roImage, 160, 41–54. DOI: https://doi.org/10.1016/j.neuroimage
.2016.12.061, PMID: 28034766
Network Neuroscience
319
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
Preti, M. G., & Van De Ville, D. (2019). Decoupling of brain
function from structure reveals regional behavioral specialization
in humans. Nature Communications, 10(1), 1–7. DOI: https://doi
.org/10.1038/s41467-019-12765-7, PMID: 31628329, PMCID:
PMC6800438
Rashid, B., Blanken, L. M., Muetzel, R. L., Miller, R., Damaraju,
E., Arbabshirani, M. R., . . . Tiemeier, H.
(2018). Connectivity
dynamics in typical development and its relationship to autis-
tic traits and autism spectrum disorder. Human Brain Mapping,
39(8), 3127–3142. DOI: https://doi.org/10.1002/hbm.24064,
PMID: 29602272, PMCID: PMC6045960
Raut, R. V., Snyder, A. Z., & Raichle, M. E. (2020). Hierarchi-
cal dynamics as a macroscopic organizing principle of the hu-
man brain. Proceedings of the National Academy of Sciences,
117(34),
20890–20897. DOI: https://doi.org/10.1073/pnas
.2003383117, PMID: 32817467
Ray, D., Hajare, N., Roy, D., & Banerjee, A.
(2020). Large-scale
functional integration, rather than functional dissociation along
dorsal and ventral streams, underlies visual perception and ac-
tion. Journal of Cognitive Neuroscience, 32(5), 847–861. DOI:
https://doi.org/10.1162/jocn_a_01527, PMID: 31933430
Reijmer, Y. D., Schultz, A. P., Leemans, A., O’Sullivan, M. J.,
Gurol, M. E., Sperling, R., . . . Hedden, T. (2015). Decoupling
of structural and functional brain connectivity in older adults
with white matter hyperintensities. NeuroImage, 117, 222–229.
DOI: https://doi.org/10.1016/j.neuroimage.2015.05.054, PMID:
26025290, PMCID: PMC4511724
Reiter, M. A., Mash, L. E., Linke, A. C., Fong, C. H., Fishman, I.,
& Müller, R. A.
(2019). Distinct patterns of atypical functional
connectivity in lower-functioning autism. Biological Psychiatry:
Cognitive Neuroscience and Neuroimaging, 4(3), 251–259. DOI:
https://doi.org/10.1016/j.bpsc.2018.08.009, PMID: 30343132,
PMCID: PMC7202917
Robertson, C. E., & Baron-Cohen, S. (2017). Sensory perception in
autism. Nature Reviews Neuroscience, 18(11), 671–684. DOI:
https://doi.org/10.1038/nrn.2017.112, PMID: 28951611
Robertson, C. E., Martin, A., Baker, C. I., & Baron-Cohen, S. (2012).
Atypical integration of motion signals in autism spectrum con-
ditions. PLoS ONE, 7(11), e48173. DOI: https://doi.org/10
PMCID:
.1371/journal.pone.0048173,
PMC3502435
PMID: 23185249,
Rosenthal, M., Wallace, G. L., Lawson, R., Wills, M. C., Dixon, E.,
Yerys, B. E., & Kenworthy, L. (2013). Impairments in real-world
executive function increase from childhood to adolescence in
autism spectrum disorders. Neuropsychology, 27(1), 13–18.
DOI: https://doi.org/10.1037/a0031299, PMID: 23356593,
PMCID: PMC4747021
Saggar, M., Sporns, O., Gonzalez-Castillo, J., Bandettini, P. A.,
Carlsson, G., Glover, G., & Reiss, A. L. (2018). Towards a new
approach to reveal dynamical organization of the brain using
topological data analysis. Nature Communications, 9(1), 1–14.
DOI:
PMID:
https://doi.org/10.1038/s41467-018-03664-4,
29643350, PMCID: PMC5895632
Sahoo, B., Pathak, A., Deco, G., Banerjee, A., & Roy, D. (2020).
Lifespan associated global patterns of coherent neural commu-
nication. NeuroImage, 216, 116824. DOI: https://doi.org/10
.1016/j.neuroimage.2020.116824, PMID: 32289459
Schmitz, N., Rubia, K., Daly, E., Smith, A., Williams, S., &
Murphy, D. G. (2006). Neural correlates of executive function in
autistic spectrum disorders. Biological Psychiatry, 59(1), 7–16.
DOI: https://doi.org/10.1016/j.biopsych.2005.06.007, PMID:
16140278
Seghier, M. L., & Price, C. J. (2018). Interpreting and utilising
intersubject variability in brain function. Trends in Cognitive
Sciences, 22(6), 517–530. DOI: https://doi.org/10.1016/j.tics
.2018.03.003, PMID: 29609894, PMCID: PMC5962820
Shafritz, K. M., Dichter, G. S., Baranek, G. T., & Belger, A.
(2008). The neural circuitry mediating shifts in behavioral re-
sponse and cognitive set in autism. Biological Psychiatry, 63(10),
974–980. DOI: https://doi.org/10.1016/j.biopsych.2007.06.028,
PMID: 17916328, PMCID: PMC2599927
Shine, J. M. (2019). Neuromodulatory influences on integration
and segregation in the brain. Trends in Cognitive Sciences,
23(7), 572–583. DOI: https://doi.org/10.1016/j.tics.2019.04.002,
PMID: 31076192
Supekar, K., Uddin, L. Q., Khouzam, A., Phillips, J., Gaillard,
W. D., Kenworthy, L. E., . . . Menon, V. (2013). Brain hyper-
connectivity in children with autism and its links to social
deficits. Cell Reports, 5(3), 738–747. DOI: https://doi.org/10
.1016/j.celrep.2013.10.001,
PMCID:
PMC3894787
24210821,
PMID:
Surampudi, S. G., Misra, J., Deco, G., Bapi, R. S., Sharma, A., &
Roy, D. (2019). Resting state dynamics meets anatomical struc-
ture: Temporal multiple kernel learning (tMKL) model. NeuroIm-
age, 184, 609–620. DOI: https://doi.org/10.1016/j.neuroimage
.2018.09.054, PMID: 30267857
Uddin, L. Q.
(2021). Brain mechanisms supporting flexible cog-
nition and behaviour in adolescents with autism spectrum
disorder. Biological Psychiatry, 89(2), 172–183. DOI: https://doi
.org/10.1016/j.biopsych.2020.05.010, PMID: 32709415
Uddin, L. Q., Supekar, K., Lynch, C. J., Khouzam, A., Phillips, J.,
(2013). Salience network–based
Feinstein, C., . . . Menon, V.
classification and prediction of symptom severity in children with
JAMA Psychiatry, 70(8), 869–879. DOI: https://doi.org
autism.
/10.1001/jamapsychiatry.2013.104, PMID: 23803651, PMCID:
PMC3951904
Uddin, L. Q., Supekar, K., Lynch, C. J., Cheng, K. M., Odriozola,
P., Maria E., Barth, M. E. , . . . Menon, V.
(2015). Brain state
differentiation and behavioral inflexibility in autism. Cerebral
Cortex, 25(12), 4740–4747. DOI: https://doi.org/10.1093/cercor
/bhu161, PMID: 25073720, PMCID: PMC4635916
van den Heuvel, M. P., Kahn, R. S., Goñi, J., & Sporns, O. (2012).
High-cost, high-capacity backbone for global brain communica-
tion. Proceedings of the National Academy of Sciences, 109(28),
11372–11377. DOI: https://doi.org/10.1073/pnas.1203593109,
PMID: 22711833, PMCID: PMC3396547
Vázquez-Rodríguez, B., Suárez, L. E., Markello, R. D., Shafiei, G.,
Paquola, C., Hagmann, P., . . . Misic, B. (2019). Gradients of
structure–function tethering across neocortex. Proceedings of the
National Academy of Sciences, 116(42), 21219–21227. DOI:
https://doi.org/10.1073/pnas.1903403116, PMID: 31570622,
PMCID: PMC6800358
Vidaurre, D., Smith, S. M., & Woolrich, M. W. (2017). Brain net-
work dynamics are hierarchically organized in time. Proceedings
Network Neuroscience
320
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Core-periphery brain network dynamics in autism
of the National Academy of Sciences, 114(48), 12827–12832.
DOI: https://doi.org/10.1073/pnas.1705120114, PMID: 29087305,
PMCID: PMC5715736
Wang, P., Kong, R., Kong, X., Liégeois, R., Orban, C., Deco, G.,
. . . Yeo, B. T. (2019). Inversion of a large-scale circuit model
reveals a cortical hierarchy in the dynamic resting human
brain. Science Advances, 5(1), eaat7854. DOI: https://doi.org/10
.1126/sciadv.aat7854, PMID: 30662942, PMCID: PMC6326747
Wang, S., Jiang, M., Duchesne, X. M., Laugeson, E. A., Kennedy,
D. P., Adolphs, R., & Zhao, Q. (2015). Atypical visual saliency
in autism spectrum disorder quantified through model-based eye
tracking. Neuron, 88(3), 604–616. DOI: https://doi.org/10.1016
/j.neuron.2015.09.042, PMID: 26593094, PMCID: PMC4662072
Watanabe, T., & Rees, G. (2017). Brain network dynamics in high-
functioning individuals with autism. Nature Communications,
8(1), 1–14. DOI: https://doi.org/10.1038/ncomms16048, PMID:
28677689, PMCID: PMC5504272
Watanabe, T., Rees, G., & Masuda, N.
(2019). Atypical intrinsic
neural timescale in autism. eLife, 8, e42256. DOI: https://doi.org
/10.7554/eLife.42256, PMID: 30717827, PMCID: PMC6363380
Xia, C. H., Ma, Z., Ciric, R., Gu, S., Betzel, R. F., Kaczkurkin,
A. N., . . . Cui, Z.
(2018). Linked dimensions of psychopatho-
logy and connectivity in functional brain networks. Nature Com-
munications, 9(1), 1–14. DOI: https://doi.org/10.1038/s41467
-018-05317-y, PMID: 30068943, PMCID: PMC6070480
Yao, Z., Hu, B., Xie, Y., Zheng, F., Liu, G., Chen, X.,
. .
Zheng, W. (2016). Resting-state time-varying analysis reveals
aberrant variations of functional connectivity in autism. Frontiers
in Human Neuroscience, 10, 463. DOI: https://doi.org/10.3389
/fnhum.2016.00463, PMID: 27695408, PMCID: PMC5025431
.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
2
2
9
5
1
9
1
3
5
1
8
n
e
n
_
a
_
0
0
1
8
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
321