Review
Intrinsic connectome organization across
temporal scales: New insights from
cross-modal approaches
Sepideh Sadaghiani
1,2 and Jonathan Wirsich 2
1Psychology Department, University of Illinois at Urbana-Champaign, Urbana, IL, Etats-Unis
2Beckman Institute for Advanced Science and Technology, University of Illinois at Urbana-Champaign, Urbana, IL, Etats-Unis
Mots clés: Connectome, Intrinsic, Multimodal, EEG, MEG, IRMf
un accès ouvert
journal
ABSTRAIT
The discovery of a stable, whole-brain functional connectivity organization that is largely
independent of external events has drastically extended our view of human brain function.
Cependant, this discovery has been primarily based on functional magnetic resonance imaging
(IRMf). The role of this whole-brain organization in fast oscillation-based connectivity as
measured, Par exemple, by electroencephalography (EEG) and magnetoencephalography
(MEG) is only beginning to emerge. Ici, we review studies of intrinsic connectivity and its
whole-brain organization in EEG, MEG, and intracranial electrophysiology with a particular
focus on direct comparisons to connectome studies in fMRI. Synthesizing this literature, nous
conclude that irrespective of temporal scale over four orders of magnitude, intrinsic
neurophysiological connectivity shows spatial similarity to the connectivity organization
commonly observed in fMRI. A shared structural connectivity basis and cross-frequency
coupling are possible mechanisms contributing to this similarity. Acknowledging that a stable
whole-brain organization governs long-range coupling across all timescales of neural
processing motivates researchers to take “baseline” intrinsic connectivity into account when
investigating brain-behavior associations, and further encourages more widespread
exploration of functional connectomics approaches beyond fMRI by using EEG and MEG
modalities.
INTRODUCTION
Although even the simplest behaviors and conscious percepts involve a distributed set of brain
régions, new empirical observations continue to challenge our understanding of such large-
scale neural connectivity. Until about the mid-2000s, cognitive neuroimaging studies using
imagerie par résonance magnétique fonctionnelle (IRMf) almost exclusively focused on the brain’s re-
sponse to experimentally controlled events. The discovery of a stable intrinsic functional con-
nectivity (FC) organization (Beckmann et al., 2005; Biswal et al., 1995; Greicius et al., 2003)
has drastically extended the focus of human functional neuroimaging (Raichle, 2009). Le
investigation of this stable organization was later expanded to whole-brain functional graphs
or “connectomes” (Achard et al., 2006), building on the notion of a structural connectome
(Sporns et al., 2005). This discovery revealed that the larger proportion of neural activity is
continuously ongoing irrespective of specific external events and cognitive challenges (hence
“intrinsic”), and is governed by FC across large-scale neurocognitive networks both during
task-free resting state and various tasks (Cole et al., 2014; Krienen et al., 2014).
Citation: Sadaghiani, S., & Wirsich,
J.. (2020). Intrinsic connectome
organization across temporal scales:
New insights from cross-modal
approaches. Neurosciences en réseau,
4(1), 1–29. https://est ce que je.org/10.1162/
netn_a_00114
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00114
Reçu: 18 Juillet 2019
Accepté: 11 Novembre 2019
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Sepideh Sadaghiani
sepideh.sadaghiani@gmail.com
Éditeur de manipulation:
Olaf Sporns
droits d'auteur: © 2019
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Functional connectivity (FC):
At the systems level denoting the
temporal dependency of signal time
courses (par exemple., from fMRI, EEG, ou
MEG) measured from distributed
brain regions.
Connectome:
A whole-brain map of structural or
functional neural connectivity. At the
systems level, connections are
typically established among brain
régions, par exemple., of a brain atlas.
Canonical oscillations:
Rhythmic variations of neural
population activity observable in
specific frequency bands thought to
represent different neural processes,
including δ (∼1–3Hz), je (∼4–7Hz), un
(∼8–13Hz), β (∼15–25Hz), and γ
(>30Hz).
Ici, we emphasize another major conceptual advance that is currently emerging through
direct comparison of whole-brain FC across data modalities that operate at different timescales
(Chiffre 1). Moving beyond fMRI, this advance suggests that a spatially similar connectome
organization governs long-range neural FC across connectivity measures and timescales. UN
comparable intrinsic large-scale network organization has been observed for infraslow fluc-
tuations around 0.1 Hz (IRMf: Beckmann et al., 2005; EEG recorded concurrently to fMRI:
Hiltunen et al., 2014) and for the full breadth of canonical oscillations up to the high γ fre-
quency range around 100 Hz (Deligianni et al., 2014; Finger et al., 2016; Hipp & Siegel, 2015;
Tewarie et al., 2018; Wirsich et al., 2017b).
The central goal of this review is to provide an overview of comparative approaches to in-
trinsic whole-brain connectome organization across data modalities. For the purpose of this
review we collectively refer to various electrophysiological and electromagnetic recordings
as neurophysiological methods. These methods comprise magnetoencephalography (MEG),
electroencephalography (EEG), and intracranial recordings in animals and humans electrocor-
ticography (ECoG). At the cost of limited spatial resolution or coverage, these data modalities
provide real-time temporal information that contrast the less direct and temporally blurred
measures of neural activity based on fMRI. Whereas the first investigations of whole-brain
connectivity organization in functional and structural MRI modalities date back to the mid-
2000s (Beckmann et al., 2005; Hagmann, 2005; Sporns et al., 2005), such investigations in
neurophysiological data are more recent (Brookes et al., 2011; Hillebrand et al., 2012; Hipp
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
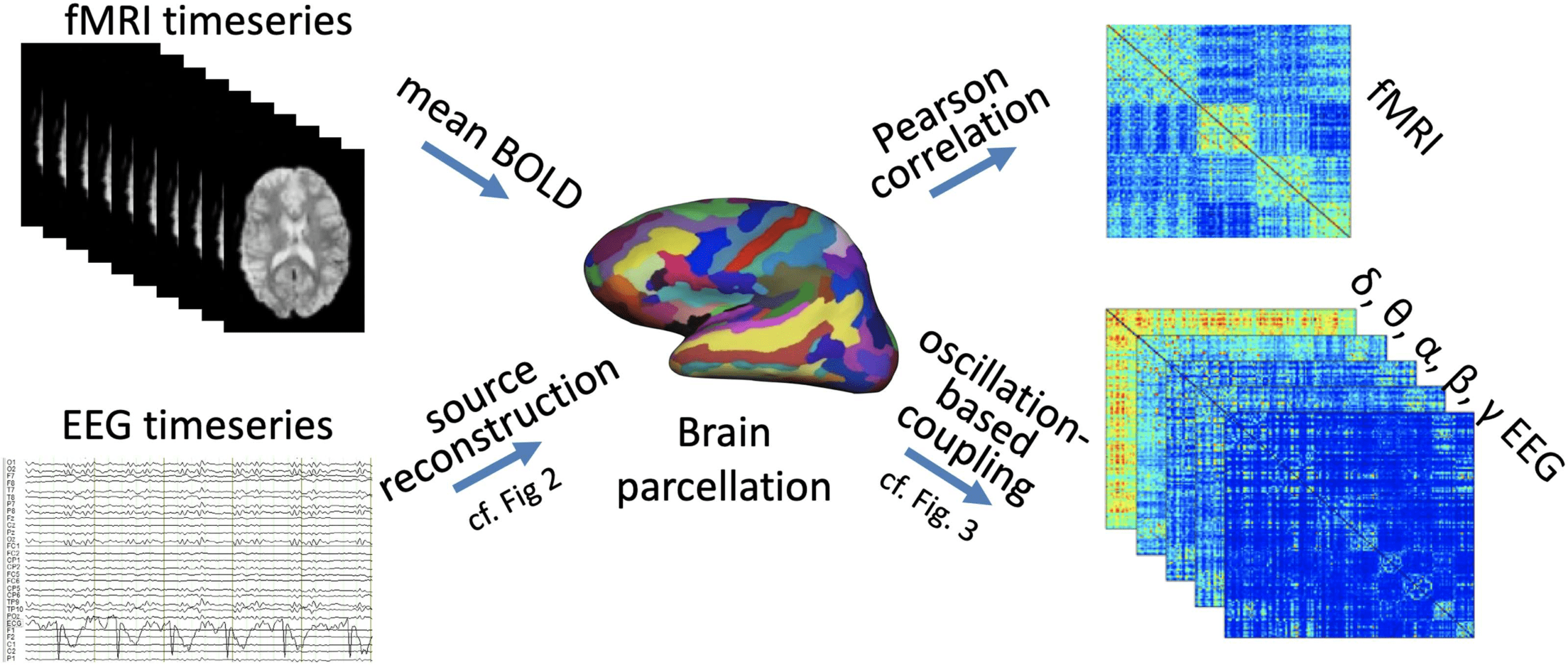
Chiffre 1. Whole-brain connectomes can be derived for various functional modalities covering
different temporal scales. (Top) fMRI BOLD signal time courses are averaged across all voxels falling
within each brain region of a whole-brain parcellation, such as an anatomically or functionally
defined parcellation atlas. Functional connectivity is then derived as the pairwise dependence of
signal fluctuations for each pair of brain regions of the atlas, most commonly by using Pearson
correlations. The ensuing connectivity matrix of all-by-all brain regions reflects the fMRI-derived
whole-brain functional connectome. (Bottom) EEG (or MEG) sensor space signal time courses are
source reconstructed onto a whole-brain parcellation (cf. Chiffre 2). This process can be performed
for various different frequency bands, often comprising canonical oscillation bands δ through γ.
Oscillation-based FC is then defined for each pair of brain regions, typically by using either phase
coupling or amplitude coupling (cf. Chiffre 3). The ensuing FC matrix of all-by-all brain regions (pour
each band) reflects the neurophysiologically derived whole-brain functional connectome. Note that
using the same parcellation for different data modalities permits direct comparison of the respective
connectomes.
Neurosciences en réseau
2
Cross-modal review of connectome organization across temporal scales
et coll., 2012). This delay is largely due to the methodological challenges of neurophysiological
data modalities with extensive cortical coverage, c'est, MEG and EEG, for which solutions
have been developed (Hassan & Wendling, 2018; O’Neill et al., 2017) (cf. Box 3).
The review begins by briefly introducing what is known about the functional connectome
from the fMRI literature (section 2). We then discuss the more recent advances in neurophysio-
logical whole-brain FC and the degree to which they converge with the fMRI-based (section 3)
and structural MRI-based connectome organization (section 4). Sections 3 et 4 will also
highlight studies in concurrently recorded EEG-fMRI. Unless otherwise stated, we will focus
on task-free resting-state studies, as most investigations of the intrinsic connectome have been
undertaken at rest. We will close by discussing possible neurobiological scenarios that may
explain the broad range of timescales governed by a universal connectome organization, et
the implications for our understanding of long-range communication in the brain.
2. FUNCTIONAL CONNECTOMES BASED ON THE HEMODYNAMIC SIGNAL
At the turn of the millennium, about a decade after the birth of fMRI, functional neuroimaging
began to substantially extend beyond investigations of task-related activation and FC changes
among small sets of task-relevant brain regions. This paradigm shift toward whole-brain FC
approaches first appeared in task-free resting-state studies and gradually extended to task
settings. As fMRI-based connectomics has been extensively covered in prior literature (par exemple.,
Buckner et al., 2013; Smith et al., 2013), the following section provides only a brief introduc-
tion. For a brief overview of core methodological considerations see Box 2.
2un. Discovery of Intrinsic Connectivity Networks
Only 3 years after the first human blood oxygen level dependent (AUDACIEUX) signal recordings, it
was discovered that spontaneous (c'est à dire., task-independent) BOLD signal fluctuations are tempo-
rally correlated across distant brain areas. This discovery initially comprised the somatomotor
(SM) réseau (Biswal et al., 1995) and later the default mode network (DMN) (Greicius et al.,
2003). It soon became evident that the observation applied to all major neurocognitive systems
from sensory to higher order control systems, leading to the notion of intrinsic connectivity net-
travaux (ICNs) (Damoiseaux et al., 2006; De Luca et al., 2006; Fox et al., 2005).
2b. Whole-Brain Connectivity Graphs
Initially driven by the fundamental goal to mathematically describe the emergence of con-
scious awareness (Edelman, 1990; Edelman & Tononi, 2001), Tononi, Sporns and Edelman in-
troduced an information-theoretic concept of whole-brain segregation and integration (Tononi
et coll., 1994). Plus tard, Sporns and (independently) Hagman coined the term “connectome” for a
whole-brain graph representation of anatomical connectivity based on fiber tracking of non-
invasive diffusion MRI (dMRI) (Hagmann, 2005; Sporns et al., 2005). This advance coincided
with the above-described discovery of a stable ICN architecture.
Par conséquent, whole-brain connectivity approaches were rapidly adopted to fMRI-derived
“functional connectomes.” In functional connectomes, the strongest dependencies emerge
across regions of the above-described ICNs (Chiffre 1, top row). Graph representations opened
the functional neuroimaging field to complex network tools well developed in other sciences
(Rubinov & Sporns, 2010). Functional connectome characteristics inform about differences in
behavior within subjects (Sadaghiani et al., 2015), across healthy individuals (Finn et al., 2015;
Nomi et al., 2017), and between clinical populations (Fornito & ullmore, 2010).
Default mode network (DMN):
A network or ICN of distributed brain
areas that show increased activation
as external cognitive demands
diminish.
Intrinsic connectivity networks
(ICNs):
Networks that spontaneously exhibit
temporal dependency among neural
activity time courses of their
distributed regions. Regions of a
given ICN also co-activate in
response to the same cognitive
demands.
Neurosciences en réseau
3
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
It is important to note that the spatial organization of the fMRI-derived connectome and
its ICNs is very stable, experiencing only minor task-related changes in FC (Cole et al., 2014;
Gratton et al., 2018; Krienen et al., 2014). Despite some degree of change, this organiza-
tion largely persists in the absence of consciousness such as during sleep or anesthesia (Amico
et coll., 2017; Hutchison et al., 2013; Picchioni et al., 2013; Wirsich et al., 2017un). The collective
spatial organization over all ICNs is thus considered the brain’s intrinsic functional architec-
ture (Petersen & Sporns, 2015). The functional raison-d’ˆetre of a relatively stable connectivity
pattern that comes at a high energy cost may be provided by predictive coding accounts. Tel
accounts view this architecture as a “memory system” modeling the statistical structure of the
monde (Sadaghiani et al., 2010un; Sadaghiani & Kleinschmidt, 2013).
2c. Dynamic Connectivity Reconfigurations
Beyond the above-described stability of the fMRI-derived FC organization, reconfigurations
are observed when the connectome is constructed from shorter periods (“dynamic” or “time-
varying” connectivity) rather than the full recording (“static” or “time-averaged” connectivity)
(Allen et al., 2014; Chang & Glover, 2010; Griffa et al., 2017; Liu & Duyn, 2013; Tagliazucchi
et coll., 2012b; Vidaurre et al., 2017). Considerable methodological debates notwithstanding
(Hindriks et al., 2016; Laumann et al., 2017), the interest in dynamic FC has grown tremen-
dously over the last few years (Cohen, 2017; Keilholz et al., 2017; Preti et al., 2017). The core
driver behind this interest is the fact that cognition is inherently dynamic. Donc, FC recon-
figurations are likely to be associated with cognitive processes, and by extension, differences
in cognitive abilities across individuals and clinical populations. This association of at least
portions of observed FC dynamics with cognitive processes finds support in their impact on
trial-by-trial perceptual outcomes (Sadaghiani et al., 2015), and in the cross-subject similarity
of FC dynamics while listening to an identical story (stimulus-induced intersubject correlations;
Simony et al., 2016). The neurophysiological counterparts of fMRI FC dynamics are discussed
at the end of section 3c.
3. FUNCTIONAL CONNECTOMES BASED ON NEUROPHYSIOLOGICAL SIGNALS AND
THEIR RELATIONSHIP TO FMRI-BASED CONNECTOMES
Compared with fMRI-based functional connectomes, estimating whole-brain FC patterns from
neurophysiological signals is a more recent development. There is a long-standing view that
oscillatory neural activity and its synchronization across brain regions facilitates long-range
communication. Encore, such communication has traditionally been evaluated across a small set of
sensors or reconstructed sources and temporally confined to relatively brief task-related processes
(Chanteur, 1999; Uhlhaas et al., 2009; Varela et al., 2001). There is a well-established under-
standing that even in the absence of task, neurophysiological signal power organizes in reoc-
curring coarse spatial patterns (“microstates”) (Koukkou-Lehmann et al., 1980; Lehmann et al.,
1987). Cependant, deriving neurophysiological whole-brain connectomes is a relatively recent con-
cept (Chiffre 2) (par exemple., Deligianni et al., 2014; Hipp & Siegel, 2015; Tewarie et al., 2016). Le
current section discusses this latter view of ongoing oscillatory processes as functional depen-
dencies unfolding continuously across distributed networks governing the whole brain.
Sources:
Plausible neural generators in the
brain hypothesized to cause the
signals observed at EEG or MEG
sensors over the scalp.
3un. Conceptual Considerations
Source leakage:
Smearing of the source signal over a
relatively large brain volume in the
reconstructed source space (cf.
Box 3).
Despite methodological challenges, first and foremost source leakage (see Box 3; Palva et al.,
2018), connectomes can be successfully derived from EEG/MEG (as evidenced by the find-
ings detailed in sections 3b, 3c, et 4). Neurophysiological connectomes can be constructed
based on amplitude coupling or phase coupling (Chiffre 3), and further assessed across the full
Neurosciences en réseau
4
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
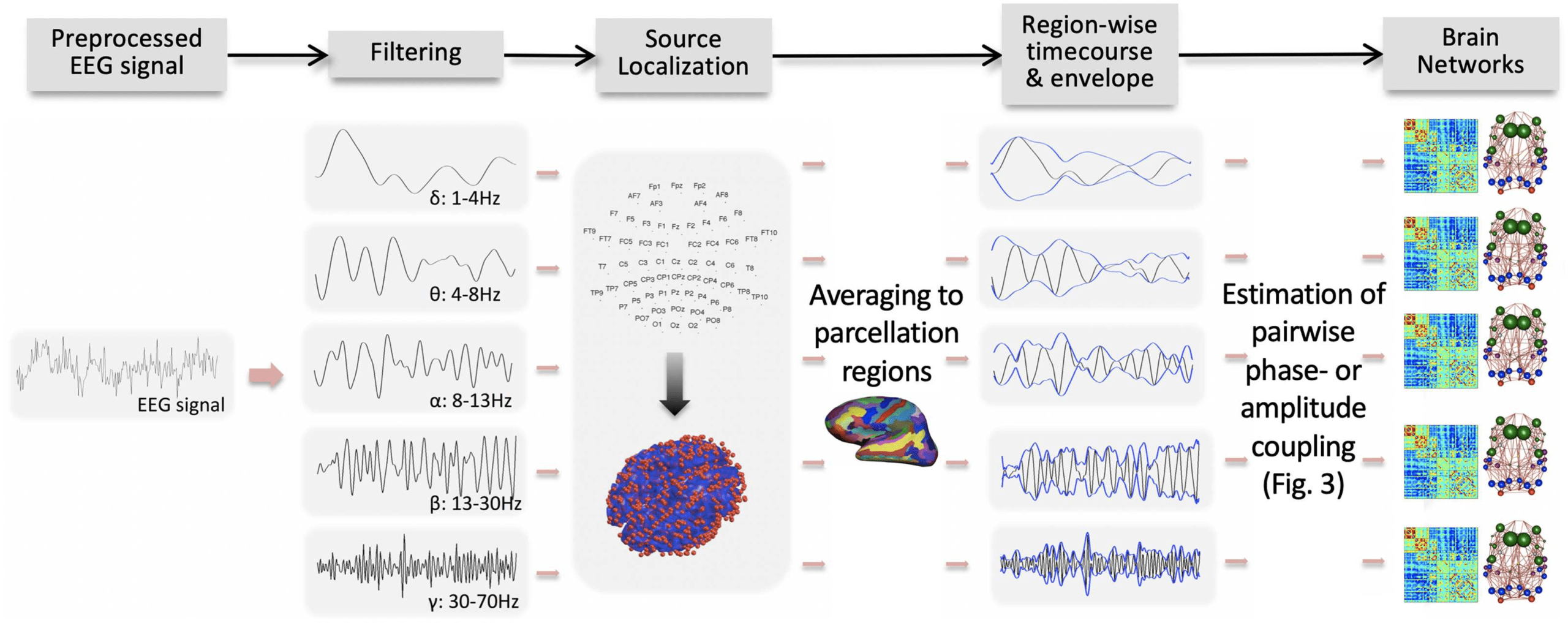
Chiffre 2. An example pipeline for constructing whole-brain connectomes from neurophysiolog-
ical signals. In this example, after preprocessing, scalp EEG or MEG data undergo filtering to fre-
quency bands of interest, source localization to regions of a brain parcellation, and estimation of
connectivity across region pairs. Although all these steps are typically considered important, ils
can occur in different orders and by using different algorithms. Source localization is crucial be-
cause the mapping of EEG/MEG sensor data to brain regions is not linear, and several sensors can
detect the activity of the same neuronal source (Farahibozorg et al., 2018). En effet, empirical com-
parisons show that the spatial topology of FC differs heavily between source and sensor space (Lai
et coll., 2018). Beyond source localization, another important step to avoid source leakage and false
positives in FC is to exclude any relationship at zero lag between two signal time courses, as leakage
is assumed to propagate instantaneously (Palva et al., 2018; Palva & Palva, 2012). Various methods
are available to this end (Brookes et al., 2011; Nolte et al., 2004; Stam et al., 2007). As further de-
tailed in Figure 3, the connectivity measure of interest can be based either on phase (par exemple., Wirsich
et coll., 2017b) or amplitude of the oscillatory signal (par exemple., Deligianni et al., 2014). Collectively, ces
steps result in a connectivity matrix for each oscillation band, which may be interrogated as a graph
(right column). Illustration modified with permission from Deligianni et al. (2014).
frequency range of neural population activity (infraslow to γ band). The rich information of
neurophysiological signals thus provides multiple concurrent connectomes (Figures 1 et 2).
Should we expect the neurophysiological connectomes at different temporal scales to differ
from each other and from the fMRI-derived connectome? In our opinion, the prior literature
implicitly reflects two conceptual viewpoints that lead to divergent predictions in regard to
cross-scale and cross-modality correspondence of FC organization.
The first viewpoint predicts that FC organization is sensitive to timescales. This view arises
on the basis of task-based neurophysiological experiments showing that both local power and
long-range coupling in different canonical oscillation bands have distinct functional roles.
Both local power and coupling are consequently more strongly tied to brain areas involved
in the respective cognitive functions. Par exemple, both amplitude (especially pronounced in
occipito-parietal areas; Gould et al., 2011; Haegens et al., 2011) and phase coupling of the
α-rhythm (Doesburg et al., 2009; Palva & Palva, 2007) are linked to selective prioritization
of specific processes and the allocation of attention, especially in the visuo-spatial domain
(Sadaghiani & Kleinschmidt, 2016). Another example of frequency-specific functional spe-
cialization is the role of the θ rhythm in navigation, memory encoding, and retrieval, especially
pronounced in the hippocampus but also relevant neocortex (local amplitude: Buzsáki, 2005;
Klimesch et al., 1996; amplitude coupling: Ekstrom et al., 2005; phase coupling: Backus et al.,
2016). Inversement, γ-oscillations are thought to generally reflect local representations of item
content (par exemple., individual stimuli; Jacobs & Kahana, 2009). Their amplitude and long-range cou-
pling are thus more widely observed in/across respective content-specific cortical areas (Fries,
2009; Rohenkohl et al., 2018). En résumé, oscillation amplitudes (Kahana, 2006), oscillation
Amplitude coupling:
The similarity of amplitude envelopes
of band-limited oscillations, c'est à dire., comment
the change in strength of a particular
oscillation is coupled across two
locations. Commonly quantified as
correlation of envelopes (depuis
Hilbert transform or power of the
spectrum at specific frequency).
Phase coupling:
The consistency of phase lag
between two oscillatory time courses
as quantified by various measures,
par exemple.,(imaginary) coherence, phase
locking value, and phase lag index.
Neurosciences en réseau
5
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
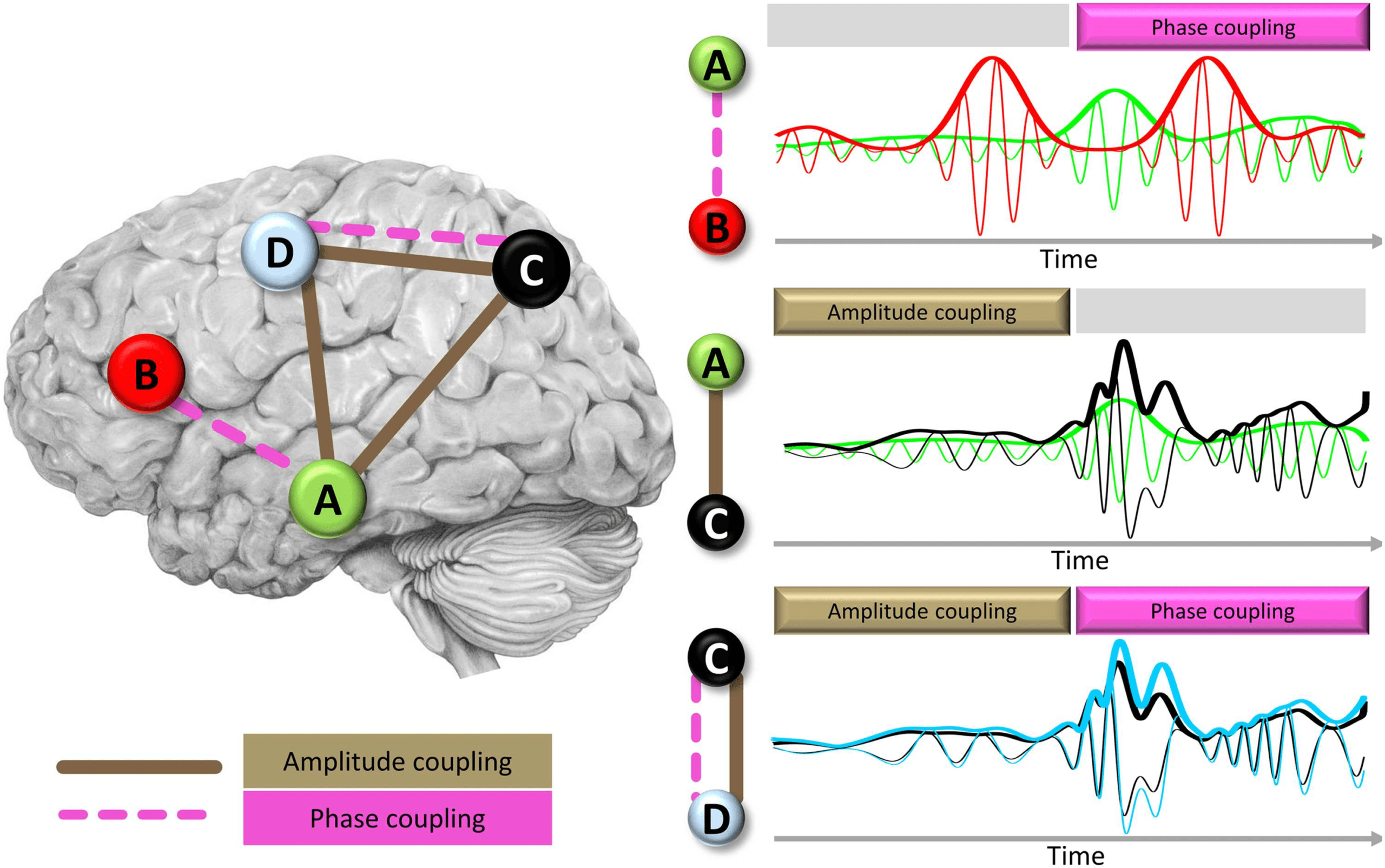
Chiffre 3. Schematic view of phase coupling and amplitude coupling as two different neurophysi-
ological mechanisms of functional connectivity. (Left panel) Graph view of FC organization between
four nodes and their connecting edges depicting either phase coupling (dashed pink lines) or am-
plitude coupling (solid brown lines). (Right panel) Narrow lines show time courses of neural activity
in a given oscillatory frequency band overlaid for the two nodes involved in a given connection.
Thicker lines in the plots show amplitude envelope of the signals with corresponding colors. Colors
of nodes (left panel) and time courses (right panel) correspond to each other. The top, middle, et
bottom right plots show connections with phase coupling only (A and B), amplitude coupling only
(A and C), or both types of coupling (C and D), respectivement.
phase cycle (VanRullen, 2016), et, importantly, oscillation-based FC (Palva & Palva, 2018)
correlate with behavioral outcomes on different cognitive processes in a frequency-dependent
manière.
This synthesis of prior literature (rather than any individual study) may lead to the predic-
tion that the spatial organization of oscillation-based FC differs heavily across frequencies, tel
that certain region pairs primarily couple in particular frequency bands. Specifically, a connec-
tome based on α-oscillations is likely not reflecting coupling of the same functional content as
a connectome based on γ-oscillations. This functional specialization of each frequency band
suggests that FC in that band would primarily occur across brain regions involved in the re-
spective function. Important for our discussion, this frequency specificity of FC distribution
would imply that the spatial topography differs substantially across connectomes derived from
different frequency bands. By extension, the spatial relationship across electrophysiological
and fMRI-derived connectomes would differ across different oscillation frequency bands.
Another viewpoint in favor of a timescale-invariant spatial organization emerges from com-
putational and modeling studies. In an early example, Honey et al. (2007) simulated neuronal
activity on different timescales by using a neural mass model (Larter et al., 1999; Morris &
Lecar, 1981). They showed that synchronization levels derived from faster timescales (10 Hz)
correlate with the infraslow fluctuations of the simulated BOLD signal (using the Balloon-
Windkessel model from Friston et al. (2003)). Deco et al. (2009) demonstrated that using
realistic time delays and coupling strengths in a neuronal model (Wilson-Cowan Oscillators)
leads to oscillators synchronous at 40 Hz (c), which in turn exhibit amplitude fluctuations
Neurosciences en réseau
6
Cross-modal review of connectome organization across temporal scales
Structural (or anatomical)
connectivité:
A representation of the physical
white-matter connections between
distant brain regions usually derived
from diffusion MRI.
in the infraslow range of the BOLD signal <0.01 Hz. Importantly, an ICN organization at a
slow timescale did not only emerge in this specific model but has similarly been observed
in other modeling approaches (FitzHugh-Nagumo oscillators; Ghosh et al., 2008), chaotic
fluctuations (Honey et al., 2007), and a reduced Wong-Wang model (Deco et al., 2013; Hansen
et al., 2015). Beyond the above-described models that fit fMRI from dMRI data, Schirner et al.
(2018) have recently shown that it is possible to fine-tune the fit between empirical and sim-
ulated fMRI (from dMRI using a Wong-Wang model) by injecting concurrently recorded source-
reconstructed EEG power. Cabral et al. (2014) demonstrated that FC modeled from structural
connectivity using a Kuramoto model of phase-coupled oscillators is related to empirical
MEG FC (envelope correlation) across all frequency bands. These observations suggest that
neurophysiological recordings either hold supplementary information to model the structure-
function relationship or help to improve SNR of the functional measures by providing a second
independent measure of FC.
In summary, the above-described models predict the emergence of an intrinsic FC organi-
zation as a result of cluster synchronization between nodes at faster timescales (for review see
Breakspear, 2017; Deco et al., 2011). According to these models, the function-structure cou-
pling is strong across long timescales, whereas dynamic subnetwork configurations arise from
shorter timescales (Deco et al., 2011). Collectively, the discussed models suggest a common
spatial organization across all timescales when averaged over sufficiently long periods.
However, the presented models assume homogenous circuit properties of the underlying
model. Conversely, it has recently been shown that relaxing those parameters (such as allowing
for recurrent connection strength and excitatory subcortical input to differ across cortical re-
gions) can improve the fit to empirical fMRI (Wang et al., 2019). While—as discussed above—
the global view puts forward a common spatial organization across all timescales, the relaxed
parameters proposed by Wang et al. implicate the possibility that individual regions oscillate
at different frequencies. Future work should explore whether such relaxed model parameters
that allow for frequency-sensitivity across space can improve the correspondence between
EEG/MEG and fMRI FC.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
.
t
To anticipate the studies covered in the following sections, empirical observations indeed
show that a unifying connectome organization is qualitatively present in neurophysiological
long-range FC of all oscillation frequencies, in line with the viewpoint of a timescale-invariant
spatial organization. Furthermore, one might find it surprising that anatomical connectivity
may explain a quite substantial proportion of the variance in a largely invariant neurophysio-
logical FC organization (e.g., cf. r2 > 0.5; Finger et al., 2016). Cependant, quantitative spectral
differences (see sections 3b and 4) support an additional frequency-specific contribution in
line with the viewpoint of a scale-sensitive FC organization.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
We also note another difference across the two viewpoints with respect to their implications
for an evoked versus intrinsic nature of FC (Raichle, 2009). When focusing on the functional
specialization of each frequency band, one may expect that coupling would be primarily con-
fined to time periods with particular processing demands directly associated with the given
frequency band. Inversement, considering the observations of the above-described modeling
approaches one would expect FC to occur in a largely ongoing and continuous manner. Le
neuroimaging field has recognized that the largest proportion of fMRI FC occurs in an intrinsic
manner rather than as a reaction to external events and demands. Although direct rest-task
comparison in neurophysiological connectomes are needed (see Box 1), we argue that the fol-
lowing sections are indicative of a similar scenario in neurophysiological FC in which a largely
stable intrinsic spatial organization governs the majority of FC with minor yet cognitively con-
sequential task-related changes.
Neurosciences en réseau
7
Cross-modal review of connectome organization across temporal scales
Box 1. Future Directions
We are likely to see the younger field of neurophysiological connectomes traverse equiva-
lent research trends and advances previously observed and currently ongoing in the fMRI
connectomics field. These trends include the following:
• Identification of connectome changes in aging and neurodevelopment (Brookes et al.,
2018).
• Identification of connectome features affected by psychiatric and neurological condi-
tion (Douw et al., 2019).
• The study of individual differences, paralleling the respective fMRI-based developments
(Finn et al., 2017; Kong et al., 2019; Mueller et al., 2013). The ability to identify
monozygotic twins in MEG-based connectomes indicates feasibility to harness geneti-
cally driven individual differences (Demuru et al., 2017).
• Time-varying FC dynamics and connectome state identification/clustering, extending
on recent source space examples (instantaneous coactivation, Baker et al., 2014; et
phase coupling, Vidaurre et al., 2018).
• Comparisons across different mental states. It is known that fMRI-derived functional
connectomes reconfigure only marginally during tasks compared with resting state
(Cole et al., 2014; Gratton et al., 2018; Krienen et al., 2014), and the characterization of
such subtle context-dependent reconfigurations is ongoing (Cohen & D'Esposito, 2016;
Hearne et al., 2017). It has been reported that connectivity is spatially highly similar
across levels of consciousness (Chu et al., 2012), and that topological graph properties
are largely conserved across rest and a simple motor task for MEG in all canonical fre-
quencies (Bassett et al., 2006), albeit in sensor space. It is important to perform such
comparisons across mental states in source space whole-brain FC, especially since the
possibility of a largely persistent intrinsic FC pattern is not commonly considered in
neurophysiological investigations of task-related cognitive processes.
En plus, direct cross-modal comparisons—including from concurrent multimodal
recordings—are an especially promising avenue to address the following key challenges:
• Dissociating the neurobiological scenarios that may give rise to the observed similarity
of FC organization across timescales and data modalities, as discussed in the conclu-
sions section. Integrating noninvasive modalities with intracranial electrophysiological
recordings will be especially helpful to aid in this endeavor.
• Identifying and quantifying the contribution of different factors leading to the remaining
dissimilarity of FC organization across timescales and data modalities. Specifically, it
is currently unclear to what degree such dissimilarity across frequency-specific neuro-
physiological connectomes and across neurophysiological and fMRI-derived connec-
tomes is of biological nature or driven by data quality issues (cf. Box 2).
3b. Neurophysiology-Based Connectomes Are Spatially Linked to fMRI-based Connectomes
This section begins with evidence that an ICN organization governs both amplitude and phase
coupling in various neurophysiological frequency bands. We then discuss direct comparisons
of edgewise FC strength of the whole-brain connectome across EEG/MEG and fMRI. Note
that most of the cross-modal comparisons discussed below (sections 3b, 3c, et 4) compare
group-averaged rather than individual connectomes across modalities. Autrement dit, ils
assess the similarity of the principal core of the connectome’s organization that is common to
all subjects.
Neurosciences en réseau
8
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Box 2. Methodological considerations: fMRI connectomes
Addressing nonneural sources of noise is crucial for fMRI-based FC.
Dealing with noise.
Such noise is considerably more likely to cause false positives/distortions in fMRI-based in-
trinsic FC than task-related fMRI activation, since the former lacks experimentally controlled
timing of events of interest. By its very nature, the BOLD signal is susceptible to respira-
tion and heart rate (Birn et al., 2006; Chang et al., 2013b). To reduce this impact, regression
of peripheral physiological recordings (Glover et al., 2000) and data-driven decomposition
approaches (par exemple., Behzadi et al., 2007) are being used. For a more detailed review see Power
et autres. (2017). Subsecond sampling rates of more recent accelerated fMRI sequences are ex-
pected to reduce aliasing of physiological noise and improve cleaning methods. De la même manière,
head motion can lead to systematic and widespread, but not uniformly distributed, changes
in fMRI-based FC (Van Dijk et al., 2012). Although remaining a key concern, the fidelity of
fMRI-based FC can improve considerably through removal of high-motion subjects and vol-
umes or interpolation of such volumes, regression of estimated head motion or global signal
shifts (but see Bright et al., 2017, for methodoloigcal challenges of nuisance regression), et
data-driven decomposition (Parkes et al., 2018; Siegel et al., 2017).
Although possible, fMRI voxel-wise FC estimation is computationally
Parcellation schemes.
expensive and may be conceptually suboptimal. Plutôt, the goal is typically to assess FC be-
tween regions that are functionally homogeneous. Homogeneity can be defined in numerous
ways, cependant, including uniformity of cytoarchitecture, task-evoked functional response, ou
FC at either the group or individual subject level1 (Arslan et al., 2017). It is worth noting
that for comparisons to other data modalities, atlases are often inevitably confined to delin-
eation of gross anatomical landmarks (cf. section 3a). Although dependence of findings on
the chosen parcellation scheme is inevitable, researchers can demonstrate the robustness of
their findings by replication in a second scheme.
Regarding measures of temporal dependency of fMRI timeseries,
Connectivity measures.
Pearson’s correlation is by far the most common approach. Cependant, fMRI-based FC can be
conceptualized in various other ways, including partial correlations (Marrelec et al., 2006),
measures of nonlinear dependencies (Hlinkaa et al., 2011), or effective connectivity (Frässle
et coll., 2018; Stephan & Friston, 2010). Coupling of the phase of BOLD fluctuations has also
been used (par exemple., Sun et al., 2004), although such fluctuations lack the oscillatory nature that is
a hallmark of ongoing neurophysiological activity. Additional measures have been proposed
specifically for time-varying changes in fMRI FC (par exemple., Shine et al., 2016). Although the choice
of FC measure depends on the question at hand, Pearson’s correlation has proven to be an
intuitively interpretable and informative metric in the broadest set of fMRI FC studies over the
dernière décennie.
Intrinsic FC is most commonly recorded in task-free resting state. Cependant,
Mental states.
subject instructions differ considerably across resting-state scans. Par exemple, subjects may
be asked to rest with eyes closed or fixate on a central visual marker. De la même manière, they may
or may not receive instructions to control their flow of thought (par exemple., avoid fixating on any
particular thought). Although some aspects of static fMRI FC (particularly ICNs) are extremely
robust to these choices, the choices may be of substantial consequence for cross-modal com-
parisons (par exemple., cf. sensitivity of α-band neurophysiological FC to eyes open/closed condition;
Gómez-Ramírez et al., 2017). Plus récemment, it has been suggested that dissociating indi-
viduals based on their functional connectome—useful for understanding interindividual dif-
ferences in behavior and clinical conditions/outcomes—can be improved by constraining
functional connectome states through stimulation with an identical continuous movie or story
for all subjects (Finn et al., 2017). Some investigations even apply connectome approaches to
fMRI data from more traditional task settings containing discrete events in the same way typi-
cally performed for continuous states (par exemple., Bassett et al., 2011). This may be especially useful
Neurosciences en réseau
9
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
for understanding distributed FC reconfigurations during performance of cognitive tasks (voir
2a section on dynamics, par exemple., Shine et al., 2016).
The core conceptual difficulty of applying connectomics to task data is that the stimuli will
increase temporal dependency across concurrently activated regions without necessarily re-
flecting information exchange or FC across those regions (Sadaghiani & Kleinschmidt, 2013).
On the other hand, the presence of any task or stimulation has only minimal impact, at least
on the time-averaged (c'est à dire., static) fMRI-derived connectome (Cole et al., 2014; Gratton et al.,
2018; Krienen et al., 2014), and task settings can be conceptualized as providing (minor)
modulation to the connectome’s dynamic trajectory (Bolt et al., 2017).
In the context of this cross-modal review it is important to recall the low-
Temporal scale.
pass characteristics of the BOLD signal; by nature, BOLD imaging is limited to the slow speed
of neural activity–induced change in the concentration of deoxy-hemoglobin irrespective of
speed of data acquisition. Donc, fMRI-based FC can only measure the temporal relation-
ship of very slow BOLD signal fluctuations, cutting off little above the range of the infraslow
frequencies. Cependant, distant neural populations exchange information at speeds of 10s of
milliseconds and employ cross-region coupling of oscillatory activity in frequencies up to
∼100 Hz (cf. section 3a). Only the slower fluctuations in the regional amplitude (envelope)
of such fast neural oscillations would result in fluctuations of metabolic demands measurable
in the BOLD signal. Par conséquent, many studies aiming at understanding the neurophysio-
logical counterpart of fMRI-based FC in multimodal recordings use amplitude coupling of
neurophysiological signals (par exemple., Nir et al., 2008; Thompson et al., 2013). Cependant, neuro-
physiological FC can be conceptualized in terms of different mechanisms (cf. section 3a and
Chiffre 3). Ainsi, how BOLD signal–derived FC relates to the FC of the underlying neurophys-
iological processes remains a complex issue (Schölvinck et al., 2013; Thompson, 2017).
Initial evidence for the existence of a neurophysiological FC organization
Presence of ICNs.
comparable to that observed in fMRI came from intracranial animal and human recordings
(He et al., 2008; Nir et al., 2008; Shmuel & Leopold, 2008). Although intracranial recordings
avoid source leakage issue, we limit our discussion of these studies as they typically lack
whole-brain coverage (but see Betzel et al., 2019, for pooling over subjects). Intracranially
recorded FC with spatial similarity to fMRI-derived ICNs spans all four orders of magnitude of
neurophysiological signals and is observable in multiple FC metrics. Such similarity has been
reported for cross-region correlations of the direct ECoG signal time courses (c'est à dire., without
spectral power or phase estimation) in the infraslow range (<0.5 Hz) (He et al., 2008) and
in canonical frequency bands (Betzel et al., 2019), and for correlations of the band-limited
amplitude envelope of high-γ (∼40–100 Hz) (Keller et al., 2013; Ko et al., 2013; Kucyi et al.,
2018; Nir et al., 2008) and slower canonical frequency bands (Hacker et al., 2017).This sim-
ilarity also holds for measures involving the phase of canonical oscillations (Betzel et al., 2019,
Supplementary Material; Weaver et al., 2016).
For neurophysiological whole-brain connectomes typically inaccessible in intracranial
recordings, we turn to MEG and EEG investigations. Although FC across the full MEG/EEG
sensor space can be informative (e.g., Bassett et al., 2006; Betzel et al., 2012; Chu et al., 2012;
Stam, 2004), we focus on source space connectomes to understand whole-brain FC organiza-
tion across brain regions.
1 For a diverse list of current atlases
see https://www.lead-dbs.org/helpsupport/knowledge-base/
atlasesresources/cortical-atlas-parcellations-mni-space/
Network Neuroscience
10
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Box 3. Methodological considerations: Neurophysiological connectomes
The study of whole-brain connectomes in MEG/EEG entails important methodological con-
siderations that have been covered in informative reviews (e.g., O’Neill et al., 2017) and
extended to network neuroscience (graph theory) approaches (Hassan & Wendling, 2018).
We therefore only briefly touch on core methodological issues.
First and foremost, MEG- and EEG-based whole-brain connectomics
Source reconstruction.
require source projection, and empirical comparisons show that the spatial topology of FC
differs heavily between source and sensor space (Lai et al., 2018). In EEG, electrical signals
spread over the head because of volume conductance, although high-density EEG recording
may increase the reliability of EEG FC to levels similar to MEG (Marquetand et al., 2019). For
both EEG and MEG FC, source localization is required because the mapping of EEG/MEG
sensor data to brain regions is not linear, and several sensors can detect the activity of the
same neuronal source. Consequently, source projection is an ill-posed problem with many
possible solutions, that is, numerous possible source constellations for relatively few sensors.
Because of spatial source reconstruction methods being subject to these spatial uncertainties,
the smearing of the source signal over a relatively large brain volume can cause source leakage
in the reconstructed source space (Farahibozorg et al., 2018).
This issue is commonly addressed by excluding any relationship at zero lag between two
signal time courses, as leakage is assumed to propagate instantaneously (Palva & Palva, 2012).
Common approaches include removing the linear dependencies by orthogonalization of the
signal (Brookes et al., 2011), or only considering FC that does not occur at zero phase lag,
for example, by using the imaginary part of the coherency (Nolte et al., 2004), the imaginary
part of the phase-locking value (Sadaghiani et al., 2012), or an index of asymmetry of the
phase-difference distribution (Stam et al., 2007). Unfortunately, this conservative approach
comes at the cost of removing real zero lag long-range FC whose existence (e.g., Gray et al.,
1989; Rodriguez et al., 1999; Roelfsema et al., 1997) and contribution to the whole-brain
connectome (e.g., Finger et al., 2016) are supported empirically and theoretically (Viriyopase
et al., 2012).
For estimation of region-wise time courses, densely positioned sources can be estimated and
then averaged within parcellation regions (e.g., Deligianni et al., 2014; Wirsich et al., 2017b)
with optional weighting by the distance to the center of mass of each parcellation region
(Brookes et al., 2016; Tewarie et al., 2016). Other options include singular-value decompo-
sition (Colclough et al., 2016; Rubega et al., 2019), or restricting the regions to sparsely and
homogenously distributed solution points (e.g., Hipp & Siegel, 2015).
It should also be noted that the parcellation atlases used in MEG/EEG studies are compara-
ble to those applied to fMRI but with two limitations. First, subcortical areas are typically
not considered estimable for surface recordings. Second, the spatial resolution or number of
parcels is limited by the number of recorded sensors. When averaging the source activity to
an anatomical atlas, the above-described spatial uncertainty of the true neuronal source can
lead to mixing one source into several parcellation regions. This issue can be ameliorated by
minimizing the cross-talk between regions (Farahibozorg et al., 2018). Parcellating the brain
according to the cross-talk or by bundling close nodes together can further minimize spurious
FC (Palva et al., 2018). The latter issue also means that studies performing connection-wise
comparisons across neurophysiological and MRI-based connectivity may be limited to rela-
tively low-resolution parcellations for MRI as well.
Another core methodological issue of importance for cross-modal
Connectivity measures.
comparisons concerns the definition of FC in neurophysiological signals (Schölvinck et al.,
2013). fMRI measures neural activity fluctuations (and consequently FC), most notably in the
infraslow (<0.1 Hz) range because of the low-pass characteristics of the hemodynamic signal.
Contrarily, neurophysiological methods have real-time resolution and can cover signal vari-
ations over ∼4 orders of magnitude from infraslow fluctuations (if permitted by appropriate
Network Neuroscience
11
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
recording hardware) to the range of “canonical” band-specific oscillations (δ through high
γ; 1 through ∼100 Hz). Consequently, whereas the vast majority of fMRI-based connectome
studies use some measure of statistical dependency of signal amplitudes (commonly Pearson’s
correlation), neurophysiology-based FC can be derived both on the basis of amplitude
coupling as well as phase coupling (for more complex cross-frequency dependencies see
Discussion section). As Figure 3 demonstrates, it is plausible to assume that the two types of
measures capture mechanistically different neurophysiological FC phenomena.
Indeed, on this theoretical basis it has been proposed that amplitude coupling regulates the
(co)activation of brain regions, while phase coupling regulates the integration and flow of cog-
nitive contents (Engel et al., 2013). Unfortunately, neurophysiological FC in cognitive tasks
is primarily studied using phase coupling (e.g., Hirvonen et al., 2018) (for an exception see
Luckhoo et al., 2012). Contrarily, the neurophysiological FC investigations at resting state use
both phase-coupling and amplitude-coupling measures, with increasingly high prevalence
of the latter (see sections 3b–3d; e.g., Brookes et al., 2011; de Pasquale et al., 2012; Hipp
et al., 2012). The difference in the respectively preferred FC metric has resulted in some dis-
connection between the task-based and resting-state neurophysiological literatures. In task-
free resting-state MEG, high similarity (r >= 0.7) was observed across the group-averaged
FC matrices from various amplitude- and phase-coupling measures (albeit amplitude cou-
pling yielded higher intersubject reliability) (Colclough et al., 2016). The spatial similarity of
amplitude and phase coupling–based neurophysiological connectomes is further supported
indirectly by the similarity of both to fMRI-based connectomes (see sections 3b and 3c).
De Pasquale et al. used seed-based correlations of broadband MEG power (1–150 Hz)
and reported evidence for the existence of the DMN, SM, dorsal attention network (DAT)
(de Pasquale et al., 2010), visuel (VIS), ventral attention (VAN), and language networks
(de Pasquale et al., 2012). Hipp et al. investigated MEG seed-based power correlations at mul-
tiple logarithmically placed frequency bands. They observed auditory (AUD), VIS, SM, et
DAT networks most dominantly carried by FC in the α/β range, and additionally the DMN as
a set of regions with particularly high “hubness” (Hipp et al., 2012). Brookes et al. appliqué
temporal ICA (rather than spatial ICA, as common in fMRI) to MEG amplitude envelopes of
canonical frequency bands (δ through γ). They found multiple ICNs, including SM, VIS, fronto-
pariétal (FP), and cerebellar networks peaking in the β-band, and DMN in the α-band (Brookes
et coll., 2011). De la même manière, studying broadband (4–30 Hz) instantaneous amplitude coactivation
states, they observed recurring spatial FC patterns resembling DMN, VIS, and SM networks (cf.
section Presence of “dynamic” connectivity reconfigurations below; Baker et al., 2014). Spatial
ICA of EEG oscillation power over all canonical frequency bands followed by fMRI-informed
clustering of the independent components confirms spatial similarity to fMRI-derived ICNs
(Sockeel et al., 2016). While all above-described studies focused on power-based FC, MEG phase
coupling shows a similar ICN-conform spatial distribution (Colclough et al., 2016; Hillebrand
et coll., 2012 albeit the authors of this study caution about methodological limitations).
A conceptual advance in more recent
Connection-wise correspondence to fMRI connectivity.
MEG/EEG studies is the shift toward studying FC patterns across whole-brain parcellations.
Encore une fois, this advance parallels the progression occurring earlier in the fMRI literature,
from a focus on characterizing ICNs to investigating whole-brain FC patterns. Using the same
brain parcellation atlas across modalities enables quantitative comparison of their FC patterns,
and we mention respective effect sizes where possible.
Hipp and Siegel (2015) performed connection-wise comparison of MEG FC (band-limited
amplitude envelope correlations) and fMRI FC recorded in the same subjects. The correlation,
c'est, spatial correspondence, between the full MEG and fMRI FC matrices was significant
Neurosciences en réseau
12
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
but modest (r = 0.12 (0.38) prior to (respectively after) SNR normalization procedure using
Spearman’s correction for attenuation). Although this spatial similarity to fMRI was present to
some degree in all canonical frequency bands, the cross-modal similarity varied by connection
and frequency. It is important to note that this study reported mean individual subject correla-
tion. Contrarily, the studies discussed below report findings for group-averaged connectomes
instead, which may explain their larger effect sizes.
Tewarie et al. (2016) directly addressed how FC in the numerous oscillation frequencies col-
lectively contributes to fMRI-derived FC. They found that single-frequency band MEG-derived
networks explain statistically significant but small variance in the whole-brain fMRI FC matrix
(r up to 0.35 for amplitude coupling and r up to 0.24 for phase coupling). Surtout, predic-
tion of the fMRI FC pattern substantially improved when jointly considering all canonical MEG
frequencies, and further improvement was observed by including linear, nonlinear, and cross-
frequency combinations of MEG FC values (r = 0.6 for amplitude coupling and r = 0.5 pour
phase coupling). This observation suggests that neurophysiological FC in different frequencies
constitute not only common but also unique contributions to FC in fMRI, and that the cross-
modal relationship contains nonlinear components. Fait intéressant, a comprehensive model that
included both amplitude coupling and phase coupling was the best predictor of fMRI-derived
FC (r = 0.73). This observation further suggests that beyond a common core, amplitude and
phase coupling may be associated with unique and complimentary components of FC in fMRI.
Time-varying changes in FC deserve a ded-
Presence of “dynamic” connectivity reconfigurations.
icated discussion given the extensive current interest in dynamics in the field of fMRI con-
nectomics (cf. section 2). Ici, we discuss literature that assesses spontaneously occurring
time-varying FC dynamics in EEG/MEG. Early studies of dynamics in sensor space EEG have
indicated the presence of subsecond FC reconfigurations (Betzel et al., 2012; Chu et al., 2012),
and have been extended to source-reconstructed connectomes: in MEG source space, hidden
Markov models have been used to detect recurring spatial patterns of instantaneous coacti-
vation of broadband (4–30 Hz) amplitude (Baker et al., 2014). As discussed above, this study
found transient occurrences of several activity patterns, some of which resemble DMN, SM,
and VIS ICNs observed in fMRI, but exhibiting much faster reconfigurations (∼100−200 ms).
The same group extended the hidden Markov model approach to the combination of instan-
taneous amplitude and phase coupling (Vidaurre et al., 2018). This work confirmed rapid
(∼50−100 ms) transient activation patterns in DMN, SM, and VIS networks constructed from
broadband (1–45 Hz) amplitude. These coactivation patterns were accompanied by transient
coherence across the same brain areas that showed coactivation. Cependant, the DMN was
reported to occur in the form of two independent patterns, a posterior subdivision operating
primarily in the δ–θ range and an anterior subdivision employing the α-band.
Some parallels can be drawn between the observed MEG states and the states identified
in fMRI-derived connectome dynamics. Par exemple, some states are dominated by SM and
sensory FC while other states are not (Allen et al., 2014; Vidaurre et al., 2017). En outre, un
key dissociating factor between fMRI-based FC states is the connectivity profile of the DMN,
and certain states similarly exhibit a posterior-anterior split of the DMN (par exemple., Allen et al.,
2014). It is unclear, cependant, whether one should expect fast-switching neurophysiological
connectome states to directly correspond to or even “sum up” to the slower fMRI-derived
states. Note that direct spatiotemporal correspondence and co-occurrence of time-varying FC
changes across EEG/MEG, and fMRI can be assessed only when acquired concurrently (voir
section 3c on dynamics).
Neurosciences en réseau
13
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
En résumé, evidence for a reproducible ICN organization in neurophysiological signals
is converging, with VIS, SM, and DMN among the most robustly reported networks. Plus loin-
plus, the reviewed whole-brain investigations collectively suggest that the intrinsic connec-
tome organization known from fMRI is to some degree present in neurophysiological FC. Ce
observation holds true for both phase- and amplitude-coupling measures. Although different
frequencies may contribute to specific connections or networks to different degrees, the cross-
modal correspondence of the whole-brain FC pattern is not confined to any particular oscilla-
tion frequency. Enfin, dynamic neurophysiological FC investigations suggest that connectivity
in the different ICNs occur in succession over shorter epochs. This dynamic pattern gives rise
to the whole-brain connectome architecture when integrated over longer time periods.
In sum, oscillation-based networks are stable over long periods, and their organization is
largely invariant to changing cognitive demands. Ainsi, the data suggest that phase coupling
and amplitude coupling are primarily intrinsic processes. This conclusion extends the under-
stating of neurophysiological FC beyond prevalent expectations of timescale sensitivity and
mental-state dependence discussed in section 3a.
3c. Relation to fMRI Connectivity Recorded Concurrently
Research on the relationship of FC across fMRI and neurophysiological signals over the respec-
tive timescales benefits from concurrent measurements in two major ways. D'abord, concurrent
recordings ensure that recordings stem from the same mental state such as levels of vigilance
that affect both measures. Deuxième, concurrent recordings permit cross-modal comparison of
time-varying changes in FC.
Studies recording EEG and hemodynamic signals concurrently have investigated the re-
lationship between fluctuations of neurophysiological signal power with BOLD amplitude
(Hiltunen et al., 2014; Laufs et al., 2003; Mantini et al., 2007; Sadaghiani et al., 2010b), neuro-
physiological signal power with BOLD FC (Allen et al., 2017; Chang et al., 2013un; Tagliazucchi
et coll., 2012b), and FC of neurophysiological signals (in sensor space) with BOLD amplitude
(Jann et al., 2009; Sadaghiani et al., 2012). These observations span the full breadth of EEG
timescales from infraslow fluctuations in direct current recordings (Hiltunen et al., 2014) à
canonical oscillations (par exemple., Allen et al., 2017; Mantini et al., 2007; Sadaghiani et al., 2012;
Tagliazucchi et al., 2012b). Many of these studies have been covered elsewhere (for infor-
mative reviews see Keilholz, 2014; Schölvinck et al., 2013). Inversement, neurophysiological
whole-brain connectomes, c'est, source-reconstructed FC across whole-brain parcellations
as detailed in Figure 1B andFigure 2, have only recently been extended to concurrent EEG-
IRMf (Deligianni et al., 2014; Wirsich et al., 2017b). Note that as in section 3b, each of these
connectome studies use the same atlas parcellation across the different data modalities.
Methodological considerations for concurrent recordings. General considerations of concurrent
EEG-fMRI, especially MRI-induced gradient and cardioballistic artifacts in EEG, are covered
extensively elsewhere (par exemple., Abreu et al., 2018). An additional consideration particularly im-
portant to connectomics is that different frequency bands might be affected by MRI-related
artifacts to different degrees. Specifically, gradient artifacts are often stronger at high frequen-
cies (Ritter et al., 2010). En outre, the helium pump responsible for cooling the supercon-
ductive MRI coil causes a vibration artifact in the EEG that leads at a scanner-specific peak
frequency (in the γ range) and its harmonics, unless it can be turned off during data acqui-
sition (Nierhaus et al., 2013). Another frequency-specific signal deterioration is the residual
gradient artifact at a frequency defined by MRI excitation pulses per second, often remaining
Neurosciences en réseau
14
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
visible in the EEG spectrum after gradient artifact removal. Such frequency-specific disruptions
of EEG oscillations and, par conséquent, EEG-derived FC must be considered when comparing
EEG connectomes from different bands and may contribute to reduced effects in the high bands
décrit ci-dessous. Recent developments improving the coverage of electrodes on the scalp to
up to 256 electrodes inside the scanner (Iannotti et al., 2015), and acceleration of fMRI vol-
ume acquisition (Uji et al., 2018), may help increase SNR of EEG and fMRI to strengthen the
cross-modal relationship.
The few existing concurrent EEG-
Cross-modal relation of “static” connectivity organization.
fMRI papers show convincing correspondence to fMRI FC irrespective of whether amplitude
coupling or phase coupling is used. Deligianni et al. (2014) found significant spatial similarity
across fMRI-derived and EEG-derived connectomes by using band-limited amplitude coupling.
This similarity was stronger for lower frequency bands than for β- and γ-bands. Fait intéressant,
when using a statistical prediction approach, prediction of fMRI-derived from EEG-derived
connectomes performed substantially better than the other way around across all bands. Le
authors concluded that the EEG connectome irrespective of frequency band carries richer infor-
mation than the fMRI connectome (at least at the spatial resolution of typical atlas parcellations,
which may reduce fMRI resolution; cf. Box 3). Autrement dit, the EEG connectome may carry
additional information about neural FC not present in the fMRI-derived connectome, more so
than the other way around.
Wirsich et al. (2017b) used EEG phase coupling (imaginary part of the coherency) and sim-
ilarly observed a similarity between EEG and fMRI connectomes. The spatial correspondence
was r > 0.3 for all bands except γ (r = 0.16). Both Wirsich et al. (2017b) and Deligianni et al.
(2016) further assessed the relation to structural connectivity as discussed in section 4. Of spe-
cial importance, the only concurrent intracranial EEG and fMRI study on FC existing to date
has delivered evidence for a spatial correspondence of FC strength across modalities without
requiring source reconstruction. Specifically, Ridley et al. (2017) investigated FC in ECoG and
depth electrodes based on amplitude envelopes. In normal (nonepileptic) régions, they found
small but significant spatial correspondence to concurrent fMRI FC strength for all canonical
bands (δ r = 0.19, gradually decreasing through γ r = 0.05) and for broadband EEG (r = 0.09).
Evidence for electrophysio-
Cross-modal relation of ‘dynamic’ connectivity reconfigurations.
logical correlates of fMRI-based FC dynamics from concurrent multimodal studies has been
informatively reviewed elsewhere (Keilholz, 2014; Thompson, 2017). Par exemple, neuro-
physiological oscillations recorded invasively in the rat show interhemispheric FC across ho-
mologous somatosensory areas that cofluctuate with concurrent fMRI-derived dFC across the
same regions (Pan et al., 2011; Thompson et al., 2013). In the concurrent human intracranial
EEG and fMRI study mentioned above (Ridley et al., 2017), region pairs with higher variabil-
ity in fMRI-derived FC also had higher variability in EEG-derived FC of α-, β-, and γ-bands
(where variability was measured as standard deviation of FC over time). Malheureusement, le
limited spatial coverage of such invasive electrophysiology studies does not inform about the
cross-modal correspondence of dynamic changes in the functional connectome’s whole-brain
spatial topography.
Although scalp EEG and fMRI provide whole-brain coverage, the vast majority of exist-
ing time-varying investigations of concurrent EEG-fMRI do not assess EEG source space con-
nectivity. Several studies have instead focused on EEG power correlates of fMRI-derived FC
dynamics. A prominent example is the study of EEG microstates, quasi-stable topographies of
Neurosciences en réseau
15
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
momentary scalp distributions (in sensor space) typically extracted from broadband global field
pouvoir. Several such microstates have been identified whose occurrence coincides with activa
tion in well-known ICNs in concurrent fMRI (Britz et al., 2010; Musso et al., 2010; Van De Ville
et coll., 2010). Interestingly the puzzling correlation between brief microstates (∼ 50−100 ms)
and the slow dynamics of intrinsic BOLD signal fluctuations (∼ 5−10 s) may be explained
by scale-free, self-similar dynamics of microstates that span over several scales from 256 ms
à 16 s (Van De Ville et al., 2010). This observation bridges across fast neural dynamics and
slowly fluctuating ICN organization.
Beyond microstates, periods of high and low EEG power in gross electrode groups have
been associated with various fMRI FC features. Examples of these features include DMN-DAT
anticorrelation (Chang et al., 2013un) or the average path length in the fMRI graph (Tagliazucchi
et coll., 2012b). The fMRI FC to EEG power relationship can substantially differ across frequen-
cies. Par exemple, the latter study (Tagliazucchi et al., 2012b) found that fluctuations in
un- and β-power co-occurred with widespread decrease in fMRI-derived FC, while γ-power was
associated with increase in long-range fMRI FC. Extending below the typically recorded EEG
frequency range, Keinänen et al. (2018) found that infraslow (<0.1 Hz) EEG and BOLD signal
fluctuations are more strongly correlated during periods of high fMRI FC in the DMN. Another
study decomposed EEG spectral power over all sensors into spatiotemporal activity patterns
with different spectral fingerprints. The time course of three such patterns temporally correlated
with sliding window fMRI FC dynamics across specific ICN pairs (Lamoš et al., 2018). In line
with an aforementioned study (Tagliazucchi et al., 2012b), a spatiotemporal EEG pattern with
high α- and β-power dominated when between-ICN fMRI FC was low (Lamoš et al., 2018).
Following the reverse analysis direction, Allen et al. (2017) first dissociated dynamically recur-
ring fMRI connectome states by applying a clustering algorithm to sliding window fMRI FC.
They found that the EEG power spectrum co-occurring with these distinct fMRI connectome
states differed from one another in certain electrode groups.
Beyond these studies on EEG power correlates of fMRI FC dynamics, a recent study as-
sessed EEG FC albeit in sensor space. The study identified recurring EEG connectivity states
(4–30 Hz broadband amplitude envelope correlations) by using hidden Markov models, and
found that fMRI coactivation patterns co-occurring with these EEG states resembled traditional
ICNs (Hunyadi et al., 2018).
To summarize section 3c, the few existing studies comparing concurrently recorded fMRI
connectomes and source-reconstructed EEG connectomes have established a cross-modal sim-
ilarity of static FC organization. Concurrently recorded EEG and fMRI are particularly useful to
understand the cross-modal relationship of time-varying dynamics, and several dynamic inves-
tigations have provided insights into EEG power and EEG sensor-level FC. To allow for a direct
comparison of whole-brain connectome reconfigurations across modalities, investigations of
source-reconstructed concurrent EEG will be required in the future.
To conclude, temporal convergence of spontaneous time-varying changes across concurrent
neurophysiological and fMRI measures supports a neural origin of fMRI-derived connectomes
(Schölvinck et al., 2013). This implication is especially critical in light of the susceptibility of
fMRI-derived FC dynamics to contamination from noise and sampling error (Keilholz, 2014).
Although this methodological conclusion is important, another implication is more profound
in our opinion. Specifically, the cross-modal static and dynamic convergence provides support
for a neural FC organization that crosses timescales, as further discussed in section 5.
Network Neuroscience
16
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
4. THE RELATIONSHIP ACROSS FUNCTIONAL AND STRUCTURAL CONNECTOMES
In the following, we discuss to what degree fMRI-based and neurophysiological connectomes
can be predicted from the underlying neural fiber tracts derived from dMRI-based tractography.
We close with trimodal studies of dMRI-, fMRI-, and neurophysiology-derived connectomes
that allow researchers to directly compare the structure-function relationship of fMRI and neu-
rophysiological modalities.
4a. The fMRI-based Connectome Is Linked to the Structural Connectome
Moderate but significant correlation (r ∼0.3) has been reported between connection strength
of the structural and the fMRI-derived connectomes (Honey et al., 2009; Skudlarski et al.,
2008). Furthermore, simulations discussed in section 3a suggest that structural connectivity
mechanistically contributes to fMRI-derived FC organization (Deco et al., 2011). As functional
connections can be realized without a direct structural connection (Damoiseaux & Greicius,
2009), several studies have focused on better modeling the function-structure relationship,
for example, by using network communication theory (Goñi et al., 2014), generative models
(Betzel et al., 2016), or partial least squares (Miši´c et al., 2016).
When modeling BOLD FC from the anatomical structure, the model predicts a closer cross-
modal relationship when averaging simulated FC across longer time periods (Honey et al.,
2007). However, the inherently flexible nature of cognition and the observation of FC dynamics
(cf. section 2) lead to the question of when/how the functional connectome undergoes flexible
excursions away from the structural core that provides its stable foundation. By applying a point
process analysis that can identify discrete spatiotemporal events in fMRI (Tagliazucchi et al.,
2012a), it has been shown that networks of transient spatiotemporal FC selectively propagate
along structural connections in a complex wave-like pattern (Griffa et al., 2017). Fukushima et al.
found that the fMRI-derived connectome is most similar to the structural connectome when
the former is in a highly integrated state. In other words, increased modular segregation of
the functional connectome reflects flexibility away from the structural connectome (Fukushima
et al., 2017). Empirically observed dynamics of fMRI-derived FC can be simulated by combin-
ing structural connectivity with stochastic processes (Hansen et al., 2015). However, how FC
dynamics are shaped by structural connectivity varies over network nodes (Shen et al., 2015),
and an exact explanation of this relationship is still outstanding.
4b. Neurophysiology-based Connectomes Are Linked to the Structural Connectome
A close link to anatomical connectivity would provide convincing evidence for the true nature
of source space neurophysiological connectomes, despite their methodological limitations (cf.
Box 2). Although dMRI-derived tractography has its own methodological issues (Maier-Hein
et al., 2017), spatial convergence across such reconstructed white-matter connectivity and
neurophysiological connectomes would speak to reliability of both modalities. Once again,
this conceptual evolution parallels that in fMRI in which the existence of intrinsic FC was
similarly validated by its spatial relationship to structural connectivity (Honey et al., 2009;
Skudlarski et al., 2008; van den Heuvel et al., 2009).
Furthermore, it is important to emphasize the relation of neurophysiological FC to the stable
anatomical structure from a conceptual point of view, since neurophysiological FC has tradi-
tionally been thought of as rapidly forming and dissolving circuits, depending on cognitive
demands (Kahana, 2006; Singer, 1999; Varela et al., 2001). Even in relatively speaking more
recent conceptual frameworks on the role of oscillation-based FC in cognition, an intrinsic
Network Neuroscience
17
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
and largely persistent whole-brain spatial organization is not central (Buzsáki & Watson, 2012;
Gratton, 2018) or only starting to be integrated (Singer, 2013).
Specifically, EEG source-reconstructed FC in all canonical frequency bands (δ through γ)
is higher between nodes with direct and indirect dMRI-derived connections compared with
those without structural connections (Chu et al., 2015, in epileptic children). This structure-
function relationship persisted even after accounting for the contribution of spatial proximity to
connectivity strength in both modalities, and was observed for both amplitude correlations and
phase coupling. To mechanistically establish a contribution of structural to neurophysiological
connectome organization, Cabral et al. set up a simple model (Kuramoto, two parameters only)
of simulated phase-coupled oscillators based on real dMRI-derived FC. As discussed in section
3a, the simulated neurophysiological data spontaneously exhibited amplitude coupling. This
coupling showed considerable spatial similarity to real MEG amplitude coupling in various
frequency bands (r = 0.41 for the optimal model parameters) (Cabral et al., 2014). A study of
EEG a-band phase coupling confirmed a close structure-function relationship (r = 0.48) (Finger
et al., 2016). Compared with using structural connectivity as a direct predictor, the association
substantially increased (r = 0.74) when dMRI connectivity was entered into a (Kuramoto)
model of phase-coupled oscillators to simulate EEG FC.
4c. Trimodal Comparisons
A few studies have assessed the connection-wise relationship across dMRI, fMRI, and neuro-
physiological connectomes. Amplitude coupling in MEG was spatially well correlated with
= 0.33 − 0.45; weaker for δ:
dMRI-derived connectivity for θ through γ bands (rSpearman
= 0.28)
= 0.14), outperforming the association across dMRI and fMRI (rSpearman
rSpearman
(Garcés et al., 2016). These dMRI-MEG and dMRI-fMRI associations weakened but persisted
after accounting for contribution of physical distance. A dMRI model exploration found that
Euclidian distance combined with a structural hub-to-hub connectivity measure in a temporo-
= 0.12, compared with
parietal network is linked to α-band MEG phase coupling at r2
= 0.33 for fMRI FC (nonoverlapping populations for functional and structural data;
r2
adjusted
Tewarie et al., 2014). Their follow-up study additionally confirmed a role of multinode (indi-
rect) structural connections in predicting FC, and this dMRI-based prediction was more accu-
rate for α-band MEG FC than it was for fMRI (Meier et al., 2016).
adjusted
Three of the trimodal studies recorded EEG and fMRI concurrently (cf. section 3c). Schirner
et al. (2018) showed that EEG power injection improves the fit of empirical resting-state fMRI
FC to simulated data modeled from dMRI connectivity. Deligianni et al. (2016) investigated
which connections of EEG-derived (amplitude coupling) and fMRI-derived connectomes pre-
dict various structural indices of anatomical connections. They observed that all EEG bands
and fMRI mapped onto structural indices in a similar set of connections. Wirsich et al. (2017b)
found that a measure of structural path length (weighted by the number of tracks between a
given region-pair) is linked to EEG phase coupling with r = 0.34 (γ) to r = 0.54 (β), compared
with r = 0.41 for the relation to fMRI-derived FC.
In summary, the partial spatial correspondence between neurophysiological and structural
connectivity organization, albeit often weak in effect size, is reassuring. This is especially im-
portant for EEG since spatial localizability of neural activity is more limited than for MEG
(although ameliorated by high-density EEG recording; Marquetand et al., 2019). The ability to
study the whole-brain connectome in EEG is especially useful for concurrent acquisition with
fMRI currently not possible for MEG (see section 3c), adding to EEG’s advantage in terms of
low cost.
Network Neuroscience
18
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
4d. Dissimilarities Across fMRI-based and Neurophysiological Connectomes and the Role of Structural
Connectivity Therein
What causes the correspondence between neurophysiological and fMRI-derived connectomes
(section 3) to be imperfect? It is generally difficult to determine which part of neurophysiolog-
ical FC organization is genuinely different from fMRI FC and which part is due to noise in
either modality (cf. Box 2 and Box 3). One approach to address this issue is to compare both
FC modalities to structural (dMRI) connectivity as “ground truth.” In this approach, any FC in-
formation that explains dMRI is considered true FC as opposed to noise. Despite dMRI’s own
methodological issues, this approach has proven informative.
For example, neurophysiological connectomes may reflect structural connectivity slightly
better than fMRI connectomes do (at least in relatively low-resolution parcellations typically
used for multimodal studies; Garcés et al., 2016; Wirsich et al., 2017b). One likely contrib-
utor to this observation is that in the absence of direct structural connections, EEG-based FC
is small (Chu et al., 2015), paralleling dMRI connectivity. Conversely, fMRI-based FC may be
strong even in the absence of direct structural connections (Damoiseaux & Greicius, 2009),
which may weaken dMRI-fMRI associations compared with dMRI-EEG. Indeed, in their tri-
modal studies, both Wirsich et al. (2017b) and (Deligianni et al., 2016) report converging
evidence that homologous connections are stronger in fMRI, whereas EEG is more strongly
characterized by intrahemispheric connections. This convergence was observed despite very
different EEG FC metrics used in the two studies (phase vs. amplitude coupling, and correcting
for vs. not excluding zero lag FC).
Wirsich et al. (2017b) also demonstrated some degree of frequency specificity in the above-
described complimentary information that EEG contributes to the structure-function rela-
tionship. While δ contributed globally to predicting structural connectivity, γ contributed local
information in the visual network. This observation suggests a common core shared across EEG,
fMRI, and dMRI connectivity, with additional smaller modulations on different timescales.
Beyond insights from comparisons to structure, causes of the divergence between neurophys-
iologcial and fMRI-derived connectomes remain open for future investigations (cf. Box 1).
To conclude section 4, the structural backbone mechanistically contributes to both fMRI-
derived and neurophysiological connectomes in various frequency bands (Cabral et al., 2014;
Honey et al., 2009), despite some modality- and frequency-specific connectivity differences.
Thus, a common structural basis provides a foundation for spatial similarity across fMRI-derived
and neurophysiological connectomes.
CONCLUSIONS
Based on fMRI observations, it has been widely accepted that infraslow neural activity
and its cross-region temporal dependencies are governed by a stable spatial organization
that is intrinsic in nature, that is, largely independent of mental states or external tasks (Petersen
& Sporns, 2015). The reviewed literature suggests that this intrinsic functional architecture
is not unique to the timescales of the hemodynamic signal. Rather, intrinsic FC is present
in neurophysiological data independent of particular cognitive processes and irrespective of
temporal scale (frequency band) or coupling mode (phase and amplitude coupling). Neuro-
physiological FC further shows small to moderate but significant spatial similarity to the FC
organization commonly observed in fMRI. We focus the following discussion on neurobiolog-
ical scenarios that can explain these similarities.
Network Neuroscience
19
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Regarding the relationship across neurophysiological and hemodynamic FC patterns, the
reviewed work is in line with the notion that numerous neurophysiological FC phenomena
(phase and amplitude coupling in various bands) contribute to a unitary hemodynamic signal
(Schölvinck et al., 2013). As discussed in section 3a, the more intriguing question is how
a fairly stable and reproducible FC pattern, or intrinsic “cognitive architecture” (Petersen &
Sporns, 2015), can to some degree be shared among this large range of neurophysiological FC
measures. Several scenarios are conceivable.
On the one hand, it is important to remember that spectral power at a given frequency does
not necessarily imply existence of oscillations at that center frequency. For example, bursts
of activity with sharp on- and offsets (Gratton, 2018) will have a broad spectral fingerprint,
that is, power across frequencies covering all canonical oscillation bands. On the other hand,
comparisons in the time domain have been able to dissociate broadband events such as bursts
from continuous oscillations, and one such investigation speaks against the broadband ac-
count (Deligianni et al., 2014). This study demonstrated low correlation between the dif-
ferent bands’ region-wise time series. This observation suggests that the whole-brain EEG
connectomes in the different canonical frequency bands are not driven by broadband signal
changes. Rather, EEG signals at different frequencies seem to operate within the same networks
(Deligianni et al., 2014).
Multiple scenarios are in line with the possibility that a given pair of regions may indeed
connect to each other using the full breadth of oscillatory frequencies. Since intrinsic FC is often
derived over extended periods (in the order of minutes), different neurophysiological FC pheno-
mena could dynamically emerge and fade in succession. The low correlation between individ-
ual bands’ FC time series discussed above speaks to a contribution of this scenario (Deligianni
et al., 2014). This possibility is further supported by observation of frequency specificity of
dynamic neurophysiological FC states (cf. dynamics section in 3b; Vidaurre et al., 2018).
Additionally, FC in different frequency bands could occur in different neural populations
within the same regions. A prominent example of this possibility suggests that γ oscillations
primarily occur in feed-forward connections originating in superficial cortical layers, while
α- and/or β-oscillations occur in feedback connections originating in deep layers, supported
by the observation that at least in sensory cortices γ and α/β are expressed to different degrees
in different layers (Fries, 2015; Scheeringa & Fries, 2019; Siegel et al., 2012).
Yet another possibility is that different frequency bands reflect temporally interdependent
activity patterns across and within neural populations. Periodic interdependencies across fre-
quencies are referred to as cross-frequency coupling (CFC; Canolty & Knight, 2010). Both at
rest and during task, this scenario finds support in empirical observations of phase-amplitude
CFC and phase-phase CFC binding (Siebenhuehner et al., 2019; Siebenhuehner et al., 2016).
In phase-amplitude CFC the phase of a relatively slow frequency modulates the amplitude of
faster frequencies, whereas phase-phase CFC binds the phases across different frequencies. The
likely most common example of CFC is neural activity oscillating in the γ range within time
segments rhythmically intermitted at α-, θ-, or δ-frequency. This phenomenon is observed in
representations of items in working memory (Axmacher et al., 2010; Siegel et al., 2009), rhyth-
mic sampling of sensory information (Lakatos et al., 2005), and their attentional modulation
(Jensen et al., 2014; Lakatos et al., 2008).
An interesting implication of a hierarchy of phase-amplitude CFC is that it may explain the
puzzling observation of a comparable spatial functional connectome pattern not only across
Network Neuroscience
20
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
temporal scales but also across FC measures, that is, both phase and amplitude coupling. Im-
portantly, CFC may hold across a series of frequencies in a hierarchical manner (Lakatos et al.,
2005). Mechanistically, such a hierarchy could allow large-scale and distributed processes in
relatively slower frequencies to modulate more local and faster processes (Canolty & Knight,
2010), with the consequence that even fast processes can show coupling across large distances.
As discussed in section 3a this multiscale structure is backed by brain modeling approaches
showing a common scale-free pattern supported by the anatomical backbone of the brain.
To consider an analogy, rather than thinking of FC in different frequency bands as a means
of protecting parallel and largely independent “conversations” from each other, they would
correspond to parallel melody lines of a polyphonic choir that hold an overarching temporal
relationship to each other for a holistic outcome.
An additional observation indicates that hierarchical CFC “nesting” may hold across the
full frequency spectrum from infraslow to high γ. Specifically, the power density of neuro-
physiological data is inversely proportional to oscillation frequency. This 1/frequency power
relationship may imply that perturbations occurring at slow frequencies cause a cascade of en-
ergy dissipation at higher frequencies. Consequently, widespread slow oscillations modulate
faster and more local events (Buzsáki & Draguhn, 2004; Zhigalov et al., 2017).
To conclude, rapid temporal switches across FC in different frequency bands within the
same spatial organization (shaped by anatomical structure), spatial overlap of neural popula-
tions with different frequency preferences, and CFC are plausible explanations for an intrinsic
FC organization that comprehensively covers mental states, timescales, and FC measures.
AUTHOR CONTRIBUTIONS
Sepideh Sadaghiani: Conceptualization; Writing - Original Draft; Writing - Review & Editing.
Jonathan Wirsich: Writing - Original Draft; Writing - Review & Editing.
FUNDING INFORMATION
Sepideh Sadaghiani, National
100000025), Award ID: 1R01MH116226-01A1.
Institute of Mental Health (http://dx.doi.org/10.13039/
REFERENCES
Abreu, R., Leal, A., & Figueiredo, P. (2018). EEG-informed fMRI:
A review of data analysis methods. Frontiers in Human Neuro-
science, 12. https://doi.org/10.3389/fnhum.2018.00029
Achard, S., Salvador, R., Whitcher, B., Suckling, J., & Bullmore,
E. (2006). A resilient, low-frequency, small-world human brain
functional network with highly connected association cortical
hubs. Journal of Neuroscience, 26(1), 63–72. https://doi.org/10.
1523/JNEUROSCI.3874-05.2006
Allen, E. A., Damaraju, E., Eichele, T., Wu, L., & Calhoun, V. D.
(2017). EEG signatures of dynamic functional network connec-
tivity states. Brain Topography, 1–16. https://doi.org/10.1007/
s10548-017-0546-2
Allen, E. A., Damaraju, E., Plis, S. M., Erhardt, E. B., Eichele, T.,
& Calhoun, V. D. (2014). Tracking whole-brain connectivity dy-
namics in the resting state. Cerebral Cortex, 24(3), 663–676.
https://doi.org/10.1093/cercor/bhs352
Amico, E., Marinazzo, D., Di Perri, C., Heine, L., Annen, J., Martial,
C., . . . Goñi, J. (2017). Mapping the functional connectome traits
of levels of consciousness. NeuroImage, 148, 201–211. https://
doi.org/10.1016/j.neuroimage.2017.01.020
Arslan, S., Ktena, S. I., Makropoulos, A., Robinson, E. C., Rueckert,
D., & Parisot, S. (2017). Human brain mapping: A systematic
comparison of parcellation methods for the human cerebral cor-
tex. 30. https://dx.doi.org/10.1016/j.neuroimage.2017.04.014
Axmacher, N., Henseler, M. M., Jensen, O., Weinreich, I., Elger, C.
E., & Fell, J. (2010). Cross-frequency coupling supports multi-item
working memory in the human hippocampus. Proceedings of the
National Academy of Sciences of the United States of America.
https://doi.org/10.1073/pnas.0911531107
Backus, A. R., Schoffelen, J.-M., Szebényi, S., Hanslmayr, S., &
Doeller, C. F. (2016). Hippocampal-prefrontal theta oscillations
support memory integration. Current Biology, 26(4), 450–457.
https://doi.org/10.1016/j.cub.2015.12.048
Network Neuroscience
21
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Baker, A. P., Brookes, M. J., Rezek, I. A., Smith, S. M., Behrens, T.,
Probert Smith, P. J., & Woolrich, M. (2014). Fast transient net-
works in spontaneous human brain activity. ELife, 3, e01867.
https://doi.org/10.7554/eLife.01867
Bassett, D. S., Meyer-Lindenberg, A., Achard, S., Duke, T., &
Bullmore, E. (2006). Adaptive reconfiguration of fractal small-
world human brain functional networks. Proceedings of the
National Academy of Sciences, 103(51), 19518–19523. https://
doi.org/10.1073/pnas.0606005103
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T. (2011). Dynamic reconfiguration of hu-
man brain networks during learning. Proceedings of the National
Academy of Sciences of the United States of America, 108(18),
7641–7646. https://doi.org/10.1073/pnas.1018985108
Beckmann, C. F., DeLuca, M., Devlin, J. T., & Smith, S. M. (2005).
Investigations into resting-state connectivity using independent
component analysis. Philosophical Transactions of the Royal So-
ciety B: Biological Sciences, 360(1457), 1001–1013. https://doi.
org/10.1098/rstb.2005.1634
Behzadi, Y., Restom, K., Liau, J., & Liu, T. T. (2007). A component
based noise correction method (CompCor) for BOLD and perfu-
sion based fMRI. NeuroImage, 37(1), 90–101. https://doi.org/10.
1016/j.neuroimage.2007.04.042
Betzel, R. F., Abell, M., O’Donnell, B. F., Hetrick, W. P., & Sporns,
O. (2012). Synchronization dynamics and evidence for a reper-
toire of network states in resting EEG. Frontiers in Computa-
tional Neuroscience, 6, 74. https://doi.org/10.3389/fncom.2012.
00074
Betzel, R. F., Avena-Koenigsberger, A., Goñi, J., He, Y., de Reus,
M. A., Griffa, A., . . . Sporns, O. (2016). Generative models of
the human connectome. NeuroImage, 124, 1054–1064. https://
doi.org/10.1016/j.neuroimage.2015.09.041
Betzel, R. F., Medaglia, J. D., Kahn, A. E., Soffer, J., Schonhaut, D. R.,
& Bassett, D. S. (2019). Structural, geometric and genetic factors
predict interregional brain connectivity patterns probed by elec-
trocorticography. Nature Biomedical Engineering, 1. https://doi.
org/10.1038/s41551-019-0404-5
Birn, R. M., Diamond, J. B., Smith, M. A., & Bandettini, P. A.
(2006). Separating respiratory-variation-related fluctuations from
neuronal-activity-related fluctuations in fMRI. NeuroImage, 31(4),
1536–1548. https://doi.org/10.1016/j.neuroimage.2006.02.048
Biswal, B., Yetkin, F. Z., Haughton, V. M., & Hyde, J. S. (1995).
Functional connectivity in the motor cortex of resting human
brain using echo-planar MRI. Magnetic Resonance in Medicine:
Official Journal of the Society of Magnetic Resonance in Medicine /
Society of Magnetic Resonance in Medicine, 34(4), 537–541.
Bolt, T., Anderson, M. L., & Uddin, L. Q. (2017). Beyond the
evoked/intrinsic neural process dichotomy. Network Neuro-
science, 2(1), 1–22. https://doi.org/10.1162/NETN_a_00028
Breakspear, M.
(2017). Dynamic models of large-scale brain ac-
tivity. Nature Neuroscience, 20(3), 340–352. https://doi.org/10.
1038/nn.4497
Bright, M. G., Tench, C. R., & Murphy, K. (2017). Potential pit-
falls when denoising resting state fMRI data using nuisance re-
gression. NeuroImage, 154, 159–168. https://doi.org/10.1016/j.
neuroimage.2016.12.027
Britz, J., Van De Ville, D., & Michel, C. M. (2010). BOLD corre-
lates of EEG topography reveal rapid resting-state network dy-
namics. NeuroImage, 52(4), 1162–1170. https://doi.org/10.1016/
j.neuroimage.2010.02.052
Brookes, M. J., Groom, M. J., Liuzzi, L., Hill, R. M., Smith, H. J. F.,
Briley, P. M., . . . Liddle, E. B. (2018). Altered temporal stability in
dynamic neural networks underlies connectivity changes in neu-
rodevelopment. NeuroImage, 174, 563–575. https://doi.org/10.
1016/j.neuroimage.2018.03.008
Brookes, M. J., Tewarie, P. K., Hunt, B. A. E., Robson, S. E.,
Gascoyne, L. E., Liddle, E. B., . . . Morris, P. G. (2016). A multi-
layer network approach to MEG connectivity analysis. Neuro-
Image, 132, 425–438. https://doi.org/10.1016/j.neuroimage.2016.
02.045
Brookes, M. J., Woolrich, M., Luckhoo, H., Price, D., Hale, J. R.,
Stephenson, M. C., . . . Morris, P. G. (2011). Investigating the
electrophysiological basis of resting state networks using mag-
netoencephalography. Proceedings of the National Academy of
Sciences, 108(40), 16783–16788. https://doi.org/10.1073/pnas.
1112685108
Buckner, R. L., Krienen, F. M., & Yeo, B. T. T. (2013). Opportunities
and limitations of intrinsic functional connectivity MRI. Nature
Neuroscience, 16(7), 832–837. https://doi.org/10.1038/nn.3423
Buzsáki, G. (2005). Theta rhythm of navigation: Link between path
integration and landmark navigation, episodic and semantic
memory. Hippocampus, 15(7), 827–840. https://doi.org/10.1002/
hipo.20113
Buzsáki, G., & Draguhn, A. (2004). Neuronal Oscillations in Cor-
tical Networks. Science, 304(5679), 1926–1929. https://doi.org/
10.1126/science.1099745
Buzsáki, G., & Watson, B. O. (2012). Brain rhythms and neural syn-
tax: Implications for efficient coding of cognitive content and
neuropsychiatric disease. Dialogues in Clinical Neuroscience,
14(4), 345–367.
Cabral, J., Luckhoo, H., Woolrich, M., Joensson, M., Mohseni, H.,
Baker, A., . . . Deco, G. (2014). Exploring mechanisms of spon-
taneous functional connectivity in MEG: How delayed network
interactions lead to structured amplitude envelopes of band-pass
filtered oscillations. NeuroImage, 90, 423–435. https://doi.org/
10.1016/j.neuroimage.2013.11.047
Canolty, R. T., & Knight, R. T.
(2010). The functional role of
cross-frequency coupling. Trends in Cognitive Sciences, 14(11),
506–515. https://doi.org/10.1016/j.tics.2010.09.001
Chang, C., & Glover, G. H.
(2010). Time-frequency dynamics
of resting-state brain connectivity measured with fMRI. Neuro-
Image, 50, 81–98. https://doi.org/10.1016/j.neuroimage.2009.
12.011
Chang, C., Liu, Z., Chen, M. C., Liu, X., & Duyn, J. H. (2013a).
EEG correlates of time-varying BOLD functional connectivity.
NeuroImage, 72, 227–236. https://doi.org/10.1016/j.neuroimage.
2013.01.049
Chang, C., Metzger, C. D., Glover, G. H., Duyn, J. H., Heinze,
H.-J., & Walter, M. (2013b). Association between heart rate
variability and fluctuations in resting-state functional connec-
tivity. NeuroImage, 68(0), 93–104. https://doi.org/10.1016/j.
neuroimage.2012.11.038
Network Neuroscience
22
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Chu, C.
J., Kramer, M. A., Pathmanathan,
J., Bianchi, M. T.,
Westover, M. B., Wizon, L., . . . Cash, S. S. (2012). Emergence
of stable functional networks in long-term human electro-
encephalography. Journal of Neuroscience, 32(8), 2703–2713.
https://doi.org/10.1523/JNEUROSCI.5669-11.2012
Chu, C., Tanaka, N., Diaz, J., Edlow, B., Wu, O., Hämäläinen, M.,
. . . Kramer, M. A. (2015). EEG functional connectivity is partially
predicted by underlying white matter connectivity. NeuroImage,
108, 23–33. https://doi.org/10.1016/j.neuroimage.2014.12.033
Cohen, J. R. (2017). The behavioral and cognitive relevance of time-
varying, dynamic changes in functional connectivity. NeuroImage.
https://doi.org/10.1016/j.neuroimage.2017.09.036
Cohen, J. R., & D’Esposito, M. (2016). The Segregation and Integra-
tion of Distinct Brain Networks and Their Relationship to Cogni-
tion. Journal of Neuroscience, 36(48), 12083–12094. https://doi.
org/10.1523/JNEUROSCI.2965-15.2016
Colclough, G. L., Woolrich, M. W., Tewarie, P. K., Brookes, M. J.,
Quinn, A. J., & Smith, S. M. (2016). How reliable are MEG
resting-state connectivity metrics? NeuroImage, 138, 284–293.
https://doi.org/10.1016/j.neuroimage.2016.05.070
Cole, M. W., Bassett, D. S., Power, J. D., Braver, T. S., & Petersen,
S. E. (2014). Intrinsic and Task-Evoked Network Architectures of
the Human Brain. Neuron, 83(1), 238–251. https://doi.org/10.
1016/j.neuron.2014.05.014
Damoiseaux, J. S., Rombouts, S. A. R. B., Barkhof, F., Scheltens, P.,
Stam, C. J., Smith, S. M., & Beckmann, C. F. (2006). Consistent
resting-state networks across healthy subjects. Proc Natl Acad
Sci U S A, 103(37), 13848–13853. https://doi.org/10.1073/pnas.
0601417103
Damoiseaux, Jessica S., & Greicius, M. D. (2009). Greater than the
sum of its parts: A review of studies combining structural con-
nectivity and resting-state functional connectivity. Brain Structure
and Function, 213(6), 525–533. https://doi.org/10.1007/s00429-
009-0208-6
De Luca, M., Beckmann, C. F., De Stefano, N., Matthews, P. M., &
Smith, S. M. (2006). FMRI resting state networks define distinct
modes of long-distance interactions in the human brain. Neuro-
Image, 29(4), 1359–1367. https://doi.org/10.1016/j.neuroimage.
2005.08.035
de Pasquale, F., Penna, S. D., Snyder, A. Z., Lewis, C., Mantini, D.,
Marzetti, L., . . . Corbetta, M. (2010). Temporal dynamics of
spontaneous MEG activity in brain networks. Proceedings of the
National Academy of Sciences, 107(13), 6040–6045. https://doi.
org/10.1073/pnas.0913863107
Deco, G., Jirsa, V. K., & McIntosh, A. R. (2011). Emerging concepts
for the dynamical organization of resting-state activity in the
brain. Nat Rev Neurosci, 12(1), 43–56. https://doi.org/10.1038/
nrn2961
Deco, G., Jirsa, V., McIntosh, A. R., Sporns, O., & Kötter, R. (2009).
Key role of coupling, delay, and noise in resting brain fluctua-
tions. Proceedings of the National Academy of Sciences, 106(25),
10302–10307. https://doi.org/10.1073/pnas.0901831106
Deco, G., Ponce-Alvarez, A., Mantini, D., Romani, G. L., Hagmann,
P., & Corbetta, M. (2013). Resting-State Functional Connectiv-
ity Emerges from Structurally and Dynamically Shaped Slow Lin-
ear Fluctuations. Journal of Neuroscience, 33(27), 11239–11252.
https://doi.org/10.1523/JNEUROSCI.1091-13.2013
Deligianni, F., Carmichael, D. W., Zhang, G. H., Clark, C. A., &
Clayden, J. D. (2016). NODDI and Tensor-Based Microstruc-
Indices as Predictors of Functional Connectivity. PLOS
tural
ONE, 11(4), e0153404. https://doi.org/10.1371/journal.pone.
0153404
Deligianni, F., Centeno, M., Carmichael, D. W., & Clayden, J. D.
(2014). Relating resting-state fMRI and EEG whole-brain connec-
tomes across frequency bands. Brain Imaging Methods, 8, 258.
https://doi.org/10.3389/fnins.2014.00258
Demuru, M., Gouw, A. A., Hillebrand, A., Stam, C. J., Dijk, B. W.,
van, Scheltens, P., . . . Visser, P. J. (2017). Functional and effec-
tive whole brain connectivity using magnetoencephalography to
identify monozygotic twin pairs. Scientific Reports, 7(1), 9685.
https://doi.org/10.1038/s41598-017-10235-y
de Pasquale, F., Della Penna, S., Snyder, A. Z., Marzetti, L., Pizzella,
V., Romani, G. L., & Corbetta, M. (2012). A Cortical Core for
Dynamic Integration of Functional Networks in the Resting Hu-
man Brain. Neuron, 74(4), 753–764. https://doi.org/10.1016/j.
neuron.2012.03.031
Doesburg, S. M., Green, J. J., McDonald, J. J., & Ward, L. M. (2009).
From local inhibition to long-range integration: A functional dis-
sociation of alpha-band synchronization across cortical scales in
visuospatial attention. Brain Research, 1303, 97–110. https://doi.
org/10.1016/j.brainres.2009.09.069
Douw, L., van Dellen, E., Gouw, A. A., Griffa, A., de Haan, W.,
van den Heuvel, M., . . . Stam, C. J. (2019). The road ahead
in clinical network neuroscience. Network Neuroscience, 3(4),
969–993. https://doi.org/10.1162/netn_a_00103
Edelman, G. (1990). The Remembered Present: A Biological Theory
of Consciousness (1st ed.). New York: Basic Books.
Edelman, G., & Tononi, G. (2001). A Universe of Consciousness:
How Matter Becomes Imagination. New York, NY: Basic Books.
Ekstrom, A. D., Caplan, J. B., Ho, E., Shattuck, K., Fried, I., &
Kahana, M. J. (2005). Human hippocampal theta activity dur-
ing virtual navigation. Hippocampus, 15(7), 881–889. https://
doi.org/10.1002/hipo.20109
Engel, A. K., Gerloff, C., Hilgetag, C. C., & Nolte, G. (2013). Intrin-
sic coupling modes: Multiscale interactions in ongoing brain ac-
tivity. Neuron, 80(4), 867–886. https://doi.org/10.1016/j.neuron.
2013.09.038
Farahibozorg, S.-R., Henson, R. N., & Hauk, O. (2018). Adaptive
cortical parcellations for source reconstructed EEG/MEG con-
nectomes. NeuroImage, 169, 23–45. https://doi.org/10.1016/j.
neuroimage.2017.09.009
Finger, H., Bönstrup, M., Cheng, B., Messé, A., Hilgetag, C., Thomalla,
G., . . . König, P. (2016). Modeling of Large-Scale Functional
Brain Networks Based on Structural Connectivity from DTI:
Comparison with EEG Derived Phase Coupling Networks and
Evaluation of Alternative Methods along the Modeling Path.
PLOS Computational Biology, 12(8), e1005025. https://doi.org/
10.1371/journal.pcbi.1005025
Finn, E. S., Scheinost, D., Finn, D. M., Shen, X., Papademetris, X.,
& Constable, R. T. (2017). Can brain state be manipulated to
emphasize individual differences in functional connectivity?
NeuroImage, 160, 140–151. https://doi.org/10.1016/j.neuroimage.
2017.03.064
Network Neuroscience
23
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
a
_
0
0
1
1
4
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Finn, E. S., Shen, X., Scheinost, D., Rosenberg, M. D., Huang,
J., Chun, M. M., . . . Constable, R. T. (2015). Functional con-
nectome fingerprinting: Identifying individuals using patterns of
brain connectivity. Nature Neuroscience, advance online publi-
cation. https://doi.org/10.1038/nn.4135
Fornito, A., & Bullmore, E. T. (2010). What can spontaneous fluc-
tuations of the blood oxygenation-level-dependent signal tell
us about psychiatric disorders? Current Opinion in Psychiatry.
https://doi.org/10.1097/YCO.0b013e328337d78d
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks. Pro-
ceedings of the National Academy of Sciences of the United
States of America, 102(27), 9673–9678. https://doi.org/10.1073/
pnas.0504136102
Frässle, S., Lomakina, E. I., Kasper, L., Manjaly, Z. M., Leff, A.,
Pruessmann, K. P., . . . Stephan, K. E. (2018). A generative model of
whole-brain effective connectivity. NeuroImage, 179, 505–529.
https://doi.org/10.1016/j.neuroimage.2018.05.058
Fries, P. (2009). Neuronal Gamma-Band Synchronization as a Fun-
damental Process in Cortical Computation. Annual Review of
Neuroscience, 32(1), 209–224. https://doi.org/10.1146/annurev.
neuro.051508.135603
Fries, P. (2015). Rhythms for cognition: Communication through
Coherence. Neuron, 88(1), 220–235. https://doi.org/10.1016/j.
neuron.2015.09.034
Friston, K. J., Harrison, L., & Penny, W. (2003). Dynamic causal
modelling. NeuroImage, 19(4), 1273–1302. https://doi.org/10.
1016/S1053-8119(03)00202-7
Fukushima, M., Betzel, R. F., He, Y., Heuvel, M. P., van den Zuo,
(2017). Structure–function relationships
X.-N., & Sporns, O.
during segregated and integrated network states of human brain
functional connectivity. Brain Structure and Function, 1–16.
https://doi.org/10.1007/s00429-017-1539-3
Garcés, P., Pereda, E., Hernández-Tamames, J. A., Del-Pozo, F.,
Maestú, F., & Pineda-Pardo, J. A. (2016). Multimodal description
of whole brain connectivity: A comparison of resting state MEG,
fMRI, and DWI. Human Brain Mapping, 37(1), 20–34. https://
doi.org/10.1002/hbm.22995
Ghosh, A., Rho, Y., McIntosh, A. R., Kötter, R., & Jirsa, V. K. (2008).
Noise during Rest Enables the Exploration of the Brain’s Dynamic
Repertoire. PLoS Comput Biol, 4(10), e1000196. https://doi.org/
10.1371/journal.pcbi.1000196
Glover, G. H., Li, T.-Q., & Ress, D. (2000). Image-based method for
retrospective correction of physiological motion effects in fmri:
RETROICOR. Magnetic Resonance in Medicine, 44(1), 162–167.
https://doi.org/10.1002/1522-2594(200007)44:1<162::AID-
MRM23>3.0.CO;2-E
Gómez-Ramírez, J., Freedman, S., Mateos, D., Velázquez, J.. L. P.,
& Valiante, T. UN. (2017). Exploring the alpha desynchronization
hypothesis in resting state networks with intracranial electroen-
cephalography and wiring cost estimates. Scientific Reports, 7.
https://doi.org/10.1038/s41598-017-15659-0
Goñi,
J., van den Heuvel, M.. P., Avena-Koenigsberger, UN.,
Mendizabal, N. V., de Betzel, R.. F., Griffa, UN., . . . Sporns, Ô.
(2014). Resting-brain functional connectivity predicted by ana-
lytic measures of network communication. Actes du
Académie nationale des sciences, 111(2), 833–838. https://est ce que je.org/
10.1073/pnas.1315529111
Gould, je. C., Rushworth, M.. F., & Nobre, UN. C. (2011). Indexing the
graded allocation of visuospatial attention using anticipatory alpha
oscillations. Journal de neurophysiologie, 105(3), 1318–1326.
https://est ce que je.org/10.1152/jn.00653.2010
Gratton, C., Laumann, T. O., Nielsen, UN. N., Vert, D.
J.,
Gordon, E. M., Gilmore, UN. W., . . . Petersen, S. E. (2018). Func-
tional Brain Networks Are Dominated by Stable Group and Indi-
vidual Factors, Not Cognitive or Daily Variation. Neurone, 98(2),
439–452.e5. https://doi.org/10.1016/j.neuron.2018.03.035
Gratton, G. (2018). Brain reflections: A circuit-based framework
for understanding information processing and cognitive control.
Psychophysiology, 55(3), e13038. https://doi.org/10.1111/psyp.
13038
Gray, C. M., Konig, P., Ange, UN. K., & Chanteur, W. (1989). Oscilla-
tory responses in cat visual cortex exhibit inter-columnar syn-
chronization which reflects global stimulus properties. Nature,
338(6213), 334–337. https://doi.org/10.1038/338334a0
Greicius, M.. D., Krasnow, B., Reiss, UN. L., & Menon, V. (2003).
Functional connectivity in the resting brain: A network analysis
of the default mode hypothesis. Proc Natl Acad Sci U S A, 100(1),
253–258. https://doi.org/10.1073/pnas.0135058100
Griffa, UN., Ricaud, B., Benzi, K., Bresson, X., Daducci, UN.,
Vandergheynst, P., . . . Hagmann, P.. (2017). Transient networks
of spatio-temporal connectivity map communication pathways
in brain functional systems. NeuroImage, 155, 490–502. https://
doi.org/10.1016/j.neuroimage.2017.04.015
Hacker, C. D., Snyder, UN. Z., Pahwa, M., Corbetta, M., & Leuthardt,
E. C. (2017). Frequency-specific electrophysiologic correlates of
resting state fMRI networks. NeuroImage, 149, 446–457. https://
doi.org/10.1016/j.neuroimage.2017.01.054
Haegens, S., Händel, B. F., & Jensen, Ô. (2011). Top-down Con-
trolled Alpha Band Activity in Somatosensory Areas Determines
Behavioral Performance in a Discrimination Task. The Journal
of Neuroscience, 31(14), 5197–5204. https://doi.org/10.1523/
JNEUROSCI.5199-10.2011
Hagmann, P.. (2005). From diffusion MRI to brain connectomics.
Thesis. https://doi.org/10.5075/epfl-thesis-3230
Hansen, E. C. UN., Battaglia, D., Spiegler, UN., Deco, G., & Jirsa, V. K.
(2015). Functional connectivity dynamics: Modeling the switch-
ing behavior of the resting state. NeuroImage, 105, 525–535.
https://doi.org/10.1016/j.neuroimage.2014.11.001
Hassan, M., & Wendling, F. (2018). Electroencephalography Source
Connectivity: Aiming for High Resolution of Brain Networks in
Time and Space. IEEE Signal Processing Magazine, 35(3), 81–96.
https://doi.org/10.1109/MSP.2017.2777518
Il, B. J., Snyder, UN. Z., Zempel, J.. M., Smyth, M.. D., & Raichle, M.. E.
(2008). Electrophysiological correlates of the brain’s intrinsic
large-scale functional architecture. Actes de la Nationale
Academy of Sciences of the United States of America, 105(41),
16039–16044. https://doi.org/10.1073/pnas.0807010105
Hearne, L. J., Cocchi, L., Zalesky, UN., & Mattingley, J.. B. (2017). Re-
configuration of Brain Network Architectures between Resting-
State and Complexity-Dependent Cognitive Reasoning. Journal
of Neuroscience, 37(35), 8399–8411. https://doi.org/10.1523/
JNEUROSCI.0485-17.2017
Neurosciences en réseau
24
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Hillebrand, UN., Barnes, G. R., Bosboom, J.. L., Berendse, H. W.,
& Stam, C. J.. (2012). Frequency-dependent functional connec-
tivity within resting-state networks: An atlas-based MEG beam-
former solution. NeuroImage, 59(4), 3909–3921. https://est ce que je.org/
10.1016/j.neuroimage.2011.11.005
Hiltunen, T., Kantola, J., Elseoud, UN. UN., Lepola, P., Suominen, K.,
Starck, T., . . . Palva, J.. M.. (2014). Infra-slow EEG Fluctuations
Are Correlated with Resting-State Network Dynamics in fMRI.
The Journal of Neuroscience, 34(2), 356–362. https://est ce que je.org/10.
1523/JNEUROSCI.0276-13.2014
Hindriks, R., Adhikari, M.. H., Murayama, Y., Ganzetti, M., Mantini,
D., Logothetis, N. K., & Deco, G. (2016). Can sliding-window
correlations reveal dynamic functional connectivity in resting-
state fMRI? NeuroImage, 127, 242–256. https://est ce que je.org/10.1016/
j.neuroimage.2015.11.055
Hipp, J.. F., Hawellek, D. J., Corbetta, M., Siegel, M., & Ange,
UN. K. (2012). Large-scale cortical correlation structure of sponta-
neous oscillatory activity. Neurosciences naturelles, 15(6), 884–890.
https://doi.org/10.1038/nn.3101
Hipp, J.. F., & Siegel, M..
(2015). BOLD fMRI Correlation Re-
flects Frequency-Specific Neuronal Correlation. Biologie actuelle,
25(10), 1368–1374. https://doi.org/10.1016/j.cub.2015.03.049
Hirvonen, J., Monto, S., Wang, S. H., Palva, J.. M., & Palva, S. (2018).
Dynamic large-scale network synchronization from perception
to action. Neurosciences en réseau, 2(4), 442–463. https://est ce que je.org/
10.1162/netn_a_00039
Hlinkaa, J., Paluša, M., Vejmelkaa, M., Mantini, D., & Corbetta, M..
(2011). Functional connectivity in resting-state fMRI: Is linear cor-
relation sufficient? NeuroImage, 54(3), 2218–2225. https://est ce que je.
org/10.1016/j.neuroimage.2010.08.042
Honey, C. J., Sporns, O., Cammoun, L., Gigandet, X., Thiran, J.. P.,
Meuli, R., & Hagmann, P.. (2009). Predicting human resting-
state functional connectivity from structural connectivity. Pro-
ceedings of the National Academy of Sciences of the United
States of America, 106(6), 2035–2040. https://doi.org/10.1073/
pnas.0811168106
Honey, Christopher J., Kötter, R., Breakspear, M., & Sporns, Ô.
(2007). Network structure of cerebral cortex shapes functional
connectivity on multiple time scales. Actes de la Nationale
Académie des Sciences, 104(24), 10240–10245. https://est ce que je.org/10.
1073/pnas.0701519104
Hunyadi, B., Woolrich, M., Quinn, UN., Vidaurre, D., & De Vos, M..
(2018). A dynamic system of brain networks revealed by fast tran-
sient EEG fluctuations and their fMRI correlates. NeuroImage.
https://doi.org/10.1016/j.neuroimage.2018.09.082
Hutchison, R.. M., Womelsdorf, T., Gati, J.. S., Everling, S., & Menon,
R.. S. (2013). Resting-state networks show dynamic functional
connectivity in awake humans and anesthetized macaques. Hu-
man Brain Mapping, 34(9), 2154–2177. https://doi.org/10.1002/
hbm.22058
Iannotti, G. R., Pittau, F., Michel, C. M., Vulliemoz, S., & Grouiller,
F. (2015). Pulse Artifact Detection in Simultaneous EEG–fMRI
Recording Based on EEG Map Topography. Brain Topography,
28(1), 21–32. https://doi.org/10.1007/s10548-014-0409-z
Jacobs, J., & Kahana, M.. J.. (2009). Neural representations of indi-
vidual stimuli in humans revealed by gamma-band electrocor-
ticographic activity. The Journal of Neuroscience: The Official
Journal of the Society for Neuroscience, 29(33), 10203–10214.
https://doi.org/10.1523/JNEUROSCI.2187-09.2009
Jann, K., Dierks, T., Boesch, C., Kottlow, M., Strik, W., & Koenig,
T. (2009). BOLD correlates of EEG alpha phase-locking and the
fMRI default mode network. NeuroImage, 45(3), 903–916.
Jensen, O., Gips, B., Bergmann, T. O., & Bonnefond, M.. (2014).
Temporal coding organized by coupled alpha and gamma os-
cillations prioritize visual processing. Trends in Neurosciences,
37(7), 357–369. https://doi.org/10.1016/j.tins.2014.04.001
Kahana, M.. J.. (2006). The Cognitive Correlates of Human Brain Os-
cillations. Journal des neurosciences, 26(6), 1669–1672. https://
doi.org/10.1523/JNEUROSCI.3737-05c.2006
Keilholz, S. D. (2014). The neural basis of Time-Varying Resting-State
functional connectivity. Brain Connectivity, 4(10), 769–779.
https://doi.org/10.1089/brain.2014.0250
Keilholz, S. D., Caballero-Gaudes, C., Bandettini, P., Deco, G., &
Calhoun, V. D. (2017). Time-resolved resting state fMRI analysis:
Current status, challenges, and new directions. Brain Connectiv-
ville. https://doi.org/10.1089/brain.2017.0543
Keinänen, T., Rytky, S., Korhonen, V., Huotari, N., Nikkinen, J.,
Tervonen, O., . . . Kiviniemi, V. (2018). Fluctuations of the EEG-
fMRI correlation reflect intrinsic strength of functional connectiv-
ity in default mode network. Journal of Neuroscience Research,
96(10), 1689–1698. https://doi.org/10.1002/jnr.24257
Keller, C. J., Bickel, S., Honey, C. J., Groppe, D. M., Entz, L.,
Craddock, R.. C., . . . Mehta, UN. D. (2013). Neurophysiological
Investigation of Spontaneous Correlated and Anticorrelated Fluc-
tuations of the BOLD Signal. Journal des neurosciences, 33(15),
6333–6342. https://doi.org/10.1523/JNEUROSCI.4837-12.2013
Klimesch, W., Doppelmayr, M., Russegger, H., & Pachinger, T.
(1996). Theta band power in the human scalp EEG and the en-
coding of new information. Neuroreport, 7(7), 1235–1240.
Ko, UN. L., Weaver, K. E., Hakimian, S., & Ojemann, J.. G. (2013).
Identifying Functional Networks Using Endogenous Connectivity
in Gamma Band Electrocorticography. Brain Connectivity, 3(5),
491–502. https://doi.org/10.1089/brain.2013.0157
Kong, R., Li, J., Orban, C., Sabuncu, M.. R., Liu, H., Schaefer, UN.,
. . . Yeo, B. T. T. (2019). Spatial Topography of Individual-Specific
Cortical Networks Predicts Human Cognition, Personality, et
Emotion. Cortex cérébral, 29(6), 2533–2551. https://est ce que je.org/10.
1093/cercor/bhy123
Koukkou-Lehmann, M., Lehmann, D., & Angst, J.. (1980). Functional
states of the brain: Their determinants. Elsevier/North-Holland
Biomedical Press.
Krienen, F. M., Yeo, B. T., Thomas, & Buckner Randy, L. (2014).
Reconfigurable task-dependent functional coupling modes clus-
ter around a core functional architecture. Philosophical Trans-
actions of the Royal Society B: Sciences biologiques, 369(1653),
20130526. https://doi.org/10.1098/rstb.2013.0526
Kucyi, UN., Schrouff, J., Bickel, S., Foster, B. L., Shine, J.. M., &
Parvizi, J.. (2018). Intracranial electrophysiology reveals repro-
ducible intrinsic functional connectivity within human brain net-
travaux. Journal des neurosciences, 0217–0218. https://est ce que je.org/10.
1523/JNEUROSCI.0217-18.2018
Lai, M., Demuru, M., Hillebrand, UN., & Fraschini, M.. (2018). A com-
parison between scalp- and source-reconstructed EEG networks.
Neurosciences en réseau
25
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Scientific Reports, 8(1), 12269. https://doi.org/10.1038/s41598-
018-30869-w
Resting-State Functional Connectivity Metrics. Brain Connectiv-
ville. https://doi.org/10.1089/brain.2019.0662
Lakatos, P., Karmos, G., Mehta, UN. D., Ulbert, JE., & Schroeder, C. E.
(2008). Entrainment of Neuronal Oscillations as a Mechanism of
Attentional Selection. Science, 320(5872), 110–113. https://est ce que je.
org/10.1126/science.1154735
Lakatos, P., Shah, UN. S., Knuth, K. H., Ulbert, JE., Karmos, G., &
Schroeder, C. E. (2005). An Oscillatory Hierarchy Controlling
Neuronal Excitability and Stimulus Processing in the Auditory Cor-
tex. J Neurophysiol, 94(3), 1904–1911. https://doi.org/10.1152/
jn.00263.2005
Lamoš, M., Mareˇcek, R., Slavíˇcek, T., Mikl, M., Rektor,
JE., &
Jan, J.. (2018). Spatial-temporal-spectral EEG patterns of BOLD
functional network connectivity dynamics. Journal of Neural
Engineering, 15(3), 036025. https://doi.org/10.1088/1741-2552/
aab66b
Larter, R., Speelman, B., & Worth, R.. M.. (1999). A coupled ordinary
differential equation lattice model for the simulation of epilep-
tic seizures. Chaos (Woodbury, N.Y.), 9(3), 795–804. https://est ce que je.
org/10.1063/1.166453
Laufs, H., Krakow, K., Sterzer, P., Eger, E., Beyerle, UN., Salek-
(2003). Electroencephalo-
Haddadi, UN., & Kleinschmidt, UN.
graphic signatures of attentional and cognitive default modes
in spontaneous brain activity fluctuations at rest. Procédure
of the National Academy of Sciences of the United States of
America, 100(19), 11053–11058. https://doi.org/10.1073/pnas.
1831638100
Laumann, T. O., Snyder, UN. Z., Mitra, UN., Gordon, E. M., Gratton,
C., Adeyemo, B., . . . Petersen, S. E. (2017). On the Stability of
BOLD fMRI Correlations. Cortex cérébral, 27(10), 4719–4732.
https://doi.org/10.1093/cercor/bhw265
Lehmann, D., Ozaki, H., & Pal, je. (1987). EEG alpha map se-
ries: Brain micro-states by space-oriented adaptive segmentation.
Électroencéphalographie et neurophysiologie clinique, 67(3),
271–288. https://est ce que je.org/10.1016/0013-4694(87)90025-3
Liu, X., & Duyn, J.. H. (2013). Time-varying functional network infor-
mation extracted from brief instances of spontaneous brain activ-
ville. Actes de l'Académie nationale des sciences, 110(11),
4392–4397. https://doi.org/10.1073/pnas.1216856110
Luckhoo, H., Hale, J.. R., Stokes, M.. G., Nobre, UN. C., Morris, P.. G.,
Brookes, M.. J., & Woolrich, M.. W. (2012). Inferring task-related
networks using independent component analysis in magnetoen-
cephalography. NeuroImage, 62(1), 530–541. https://est ce que je.org/10.
1016/j.neuroimage.2012.04.046
Maier-Hein, K. H., Neher, P.. F., Houde,
J.-C., Côté, M.-A.,
Garyfallidis, E., Zhong, J., . . . Descoteaux, M.. (2017). The chal-
lenge of mapping the human connectome based on diffusion
tractography. Communications naturelles, 8(1), 1349. https://est ce que je.
org/10.1038/s41467-017-01285-x
Mantini, D., Perrucci, M.. G., Gratta, C. D., Romani, G. L., &
Corbetta, M.. (2007). Electrophysiological signatures of resting
state networks in the human brain. Actes de la Nationale
Academy of Sciences of the United States of America, 104(32),
13170–13175. https://doi.org/10.1073/pnas.0700668104
Marquetand, J., Vannoni, S., Carboni, M., Li Hegner, Y., Stier,
C., Brun, C., & Focke, N. K. (2019). Reliability of Magne-
toencephalography and High-Density Electroencephalography
J., & Benali, H.
Marrelec, G., Krainik, UN., Duffau, H., Pélégrini-Issac, M., Lehéricy,
(2006). Partial correlation for
S., Doyon,
functional brain interactivity investigation in functional MRI.
NeuroImage,32(1),228–237. https://doi.org/10.1016/j.neuroimage.
2005.12.057
Meier, J., Tewarie, P., Hillebrand, UN., Douw, L., van Dijk, B. W.,
Stufflebeam, S. M., & Van Mieghem, P.. (2016). A Mapping Be-
tween Structural and Functional Brain Networks. Brain Connec-
tivité, 6(4), 298–311. https://doi.org/10.1089/brain.2015.0408
Miši´c, B., Betzel, R.. F., de Reus, M.. UN., van den Heuvel, M.. P.,
Berman, M.. G., McIntosh, UN. R., & Sporns, Ô. (2016). Réseau-
Level Structure-Function Relationships in Human Neocortex.
Cérébral, Cortex (New York NY), 26(7), 3285–3296. https://est ce que je.
org/10.1093/cercor/bhw089
Morris, C., & Lecar, H. (1981). Voltage oscillations in the barnacle
giant muscle fiber. Biophysical Journal, 35(1), 193–213. https://
doi.org/10.1016/S0006-3495(81)84782-0
Mueller, S., Wang, D., Fox, M.. D., Yeo, B. T. T., Sepulcre, J.,
Sabuncu, M.. R., . . . Liu, H. (2013). Individual Variability in Func-
tional Connectivity Architecture of the Human Brain. Neurone,
77(3), 586–595. https://doi.org/10.1016/j.neuron.2012.12.028
Musso, F., Brinkmeyer, J., Mobascher, UN., Warbrick, T., & Winterer,
G. (2010). Spontaneous brain activity and EEG microstates. UN
novel EEG/fMRI analysis approach to explore resting-state net-
travaux. NeuroImage, 52(4), 1149–1161. https://est ce que je.org/10.1016/
j.neuroimage.2010.01.093
Nierhaus, T., Gundlach, C., Goltz, D., Thiel, S. D., Pleger, B., &
Villringer, UN. (2013). Internal ventilation system of MR scan-
ners induces specific EEG artifact during simultaneous EEG-fMRI.
NeuroImage, 74, 70–76. https://doi.org/10.1016/j.neuroimage.
2013.02.016
Nir, Y., Mukamel, R., Dinstein, JE., Privman, E., Harel, M., Fisch,
L., . . . Malach, R.. (2008). Interhemispheric correlations of slow
spontaneous neuronal fluctuations revealed in human sensory
cortex. Neurosciences naturelles, 11(9), 1100–1108. https://est ce que je.org/
10.1038/nn.2177
Nolte, G., Bai, O., Wheaton, L., Mari, Z., Vorbach, S., & Hallett, M..
(2004). Identifying true brain interaction from EEG data using the
imaginary part of coherency. Neurophysiologie clinique, 115(10),
2292–2307. https://doi.org/10.1016/j.clinph.2004.04.029
Nomi, J.. S., Vij, S. G., Dajani, D. R., Steimke, R., Damaraju, E.,
Rachakonda, S., . . . Uddin, L. Q. (2017). Chronnectomic pat-
terns and neural flexibility underlie executive function. Neuro-
Image, 147, 861–871. https://doi.org/10.1016/j.neuroimage.2016.
10.026
O’Neill, G. C., Tewarie, P., Vidaurre, D., Liuzzi, L., Woolrich, M.. W.,
& Brookes, M.. J.. (2017). Dynamics of large-scale electrophysio-
logical networks: A technical review. https://est ce que je.org/10.1016/j.
neuroimage.2017.10.003
Palva, J.. M., Wang, S. H., Palva, S., Zhigalov, UN., Monto, S., Brookes,
M.. J., . . . Jerbi, K. (2018). Ghost interactions in MEG/EEG
source space: A note of caution on inter-areal coupling measures.
NeuroImage, 173, 632–643. https://doi.org/10.1016/j.neuroimage.
2018.02.032
Neurosciences en réseau
26
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Palva, S., & Palva, J.. M.. (2007). New vistas for [alpha]-frequency
band oscillations. Trends in Neurosciences, 30(4), 150–158.
https://doi.org/10.1016/j.tins.2007.02.001
Palva, S., & Palva, J.. M.. (2012). Discovering oscillatory interaction
networks with M/EEG: Challenges and breakthroughs. Trends
in Cognitive Sciences, 16(4), 219–230. https://est ce que je.org/10.1016/
j.tics.2012.02.004
Palva, S., & Palva, J.. M.. (2018). Roles of Brain Criticality and Multi-
scale Oscillations in Temporal Predictions for Sensorimotor Pro-
cessation. Trends in Neurosciences, 41(10), 729–743. https://est ce que je.
org/10.1016/j.tins.2018.08.008
Pan, W.-J., Thompson, G., Magnuson, M., Majeed, W., Jaeger, D.,
& Keilholz, S. (2011). Broadband local field potentials correlate
with spontaneous fluctuations in functional magnetic resonance
imaging signals in the rat somatosensory cortex under isoflurane
anesthesia. Brain Connectivity, 1(2), 119–131. https://est ce que je.org/10.
1089/brain.2011.0014
Parkes, L., Fulcher, B., Yücel, M., & Fornito, UN.
(2018). Un
evaluation of the efficacy, reliability, and sensitivity of motion
correction strategies for resting-state functional MRI. Neuro-
Image, 171, 415–436. https://doi.org/10.1016/j.neuroimage.
2017.12.073
Petersen, S. E., & Sporns, Ô. (2015). Brain networks and cognitive
architectures. Neurone, 88(1), 207–219. https://est ce que je.org/10.1016/
j.neuron.2015.09.027
Picchioni, D., Duyn, J.. H., & Horovitz, S. G. (2013). Sleep and the
functional connectome. NeuroImage, 80, 387–396. https://est ce que je.
org/10.1016/j.neuroimage.2013.05.067
Power, J.. D., Plitt, M., Laumann, T. O., & Martine, UN. (2017). Sources
and implications of whole-brain fMRI signals in humans. Neuro-
Image, 146, 609–625. https://doi.org/10.1016/j.neuroimage.2016.
09.038
Preti, M.. G., Bolton, T. UN., & Van De Ville, D. (2017). The dynamic
functional connectome: State-of-the-art and perspectives. Neu-
roImage. https://doi.org/10.1016/j.neuroimage.2016.12.061
Raichle, M.. E. (2009). A Paradigm Shift in Functional Brain Imag-
ing. J.. Neurosci., 29(41), 12729–12734. https://doi.org/10.1523/
JNEUROSCI.4366-09.2009
Ridley, B., Wirsich,
J., Bettus, G., Rodionov, R., Murta, T.,
Chaudhary, U., . . . Guye, M.. (2017). Simultaneous Intracranial
EEG-fMRI Shows Inter-Modality Correlation in Time-Resolved
Connectivity Within Normal Areas but Not Within Epileptic
Regions. Brain Topography, 30(5), 639–655. https://est ce que je.org/10.
1007/s10548-017-0551-5
Ritter, P., Becker, R., Freyer, F., & Villringer, UN. (2010). EEG Quality:
The Image Acquisition Artefact. In C. Mulert & L. Lemieux (Éd.),
EEG – IRMf: Physiological Basis, Technique, and Applications
(pp. 153–171). https://doi.org/10.1007/978-3-540-87919-0_9
Rodriguez, E., George, N., Lachaux, J.-P., Martinerie, J., Renault,
B., & Varela, F. J.. (1999). Perception’s shadow: Long-distance
synchronization of human brain activity. Nature, 397(6718),
430–433. https://doi.org/10.1038/17120
Roelfsema, P.. R., Ange, UN. K., Konig, P., & Chanteur, W. (1997). Visuo-
motor integration is associated with zero time-lag synchroniza-
tion among cortical areas. Nature, 385(6612), 157–161. https://
doi.org/10.1038/385157a0
Rohenkohl, G., Bosman, C. UN., & Fries, P.. (2018). Gamma Synchro-
nization between V1 and V4 Improves Behavioral Performance.
Neurone, 100(4), 953–963.e3. https://est ce que je.org/10.1016/j.neuron.
2018.09.019
Rubega, M., Carboni, M., Seeber, M., Pascucci, D., Tourbier,
S., Toscano, G., . . . Michel, C. M.. (2019). Estimating EEG
Source Dipole Orientation Based on Singular-value Decomposi-
tion for Connectivity Analysis. Brain Topography, 32(4), 704–719.
https://doi.org/10.1007/s10548-018-0691-2
Rubinov, M., & Sporns, Ô. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003
Sadaghiani, S., Hesselmann, G., Friston, K. J., & Kleinschmidt, UN.
(2010un). The relation of ongoing brain activity, evoked neural re-
sponses, and cognition. Frontiers in Systems Neuroscience, 4(20).
https://doi.org/10.3389/fnsys.2010.00020
Sadaghiani, S., & Kleinschmidt, UN. (2013). Functional interactions
between intrinsic brain activity and behavior. NeuroImage, 80,
379–386. https://doi.org/10.1016/j.neuroimage.2013.04.100
Sadaghiani, S., & Kleinschmidt, UN. (2016). Brain networks and α-
oscillations: Structural and Functional Foundations of Cognitive
Contrôle. Tendances des sciences cognitives, 20(11), 805–817. https://
doi.org/10.1016/j.tics.2016.09.004
Sadaghiani, S., Poline, J.-B., Kleinschmidt, UN., & D'Esposito, M..
(2015). Ongoing dynamics in large-scale functional connectiv-
ity predict perception. Proceedings of the National Academy
of Sciences, 112(27), 8463–8468. https://doi.org/10.1073/pnas.
1420687112
Sadaghiani, S., Scheeringa, R., Lehongre, K., Morillon, B.,
Giraud, A.-L., D'Esposito, M., & Kleinschmidt, UN. (2012). Alpha-
Band Phase Synchrony is Related to Activity in the Fronto-
Parietal Adaptive Control Network. The Journal of Neuroscience,
32(41), 14305–14310. https://doi.org/10.1523/JNEUROSCI.1358-
12.2012
Sadaghiani, S., Scheeringa, R., Lehongre, K., Morillon, B., Giraud,
A.-L., & Kleinschmidt, UN. (2010b). Intrinsic Connectivity Net-
travaux, Alpha Oscillations, and Tonic Alertness: A Simultaneous
Electroencephalography/Functional Magnetic Resonance Imag-
ing Study. J.. Neurosci., 30(30), 10243–10250. https://est ce que je.org/10.
1523/JNEUROSCI.1004-10.2010
Scheeringa, R., & Fries, P.. (2019). Cortical layers, rhythms and
BOLD signals. NeuroImage, 197, 689–698. https://est ce que je.org/10.
1016/j.neuroimage.2017.11.002
Schirner, M., McIntosh, UN. R., Jirsa, V., Deco, G., & Ritter, P..
(2018). Inferring multi-scale neural mechanisms with brain net-
work modelling. ELife, 7, e28927. https://doi.org/10.7554/eLife.
28927
Schölvinck, M.. L., Leopold, D. UN., Brookes, M..
J., & Khader,
P.. H. (2013). The contribution of electrophysiology to functional
connectivity mapping. NeuroImage. https://est ce que je.org/10.1016/j.
neuroimage.2013.04.010
Shen, K., Hutchison, R.. M., Bezgin, G., Everling, S., & McIntosh,
UN. R.. (2015).Network Structure Shapes Spontaneous Functional Con-
nectivity Dynamics. The Journal of Neuroscience, 35(14), 5579–5588.
https://doi.org/10.1523/JNEUROSCI.4903-14.2015
Shine, J.. M., Bissett, P.. G., Cloche, P.. T., Koyejo, O., Balsters, J.. H.,
Gorgolewski, K. J., . . . Poldrack, R.. UN. (2016). The Dynamics
Neurosciences en réseau
27
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
of Functional Brain Networks: Integrated Network States during
Cognitive Task Performance. Neurone, 92(2), 544–554. https://
doi.org/10.1016/j.neuron.2016.09.018
Shmuel, UN., & Leopold, D. UN. (2008). Neuronal correlates of spon-
taneous fluctuations in fMRI signals in monkey visual cortex: Im-
plications for functional connectivity at rest. Hum Brain Mapp.
https://doi.org/10.1002/hbm.20580
Siebenhuehner, F., Wang, S. H., Arnulfo, G., Nobili, L., Palva,
J.. M., & Palva, S. (2019). Resting-state cross-frequency coupling
networks in human electrophysiological recordings. BioRxiv,
547638. https://doi.org/10.1101/547638
Siebenhühner, F., Wang, S. H., Palva, J.. M., & Palva, S. (2016). Cross-
frequency synchronization connects networks of fast and slow
oscillations during visual working memory maintenance. ELife,
5, e13451. https://doi.org/10.7554/eLife.13451
Siegel, J.. S., Mitra, UN., Laumann, T. O., Seitzman, B. UN., Raichle, M.,
Corbetta, M., & Snyder, UN. Z. (2017). Data Quality Influences
Observed Links Between Functional Connectivity and Behav-
ior. Cortex cérébral, 27(9), 4492–4502. https://est ce que je.org/10.1093/
cercor/bhw253
Siegel, M., Donner, T. H., & Ange, UN. K. (2012). Spectral fingerprints
of large-scale neuronal interactions. Nat Rev Neurosci, 13(2),
121–134. https://doi.org/10.1038/nrn3137
Siegel, M., Warden, M.. R., & Miller, E. K. (2009). Phase-dependent
neuronal coding of objects in short-term memory. Procédure
of the National Academy of Sciences, 106(50), 21341–21346.
https://doi.org/10.1073/pnas.0908193106
Simony, E., Honey, C. J., Chen, J., Lositsky, O., Yeshurun, Y., Wiesel,
UN., & Hasson, U. (2016). Dynamic reconfiguration of the default
mode network during narrative comprehension. Nature Commu-
nications, 7(1), 1–13. https://doi.org/10.1038/ncomms12141
Chanteur, W. (1999). Neuronal Synchrony: A Versatile Code for the
Definition of Relations? Neurone, 24(1), 49–65. https://est ce que je.org/10.
1016/S0896-6273(00)80821-1
Chanteur, W. (2013). Cortical dynamics revisited. Tendances cognitives
les sciences, 17(12), 616–626. https://doi.org/10.1016/j.tics.2013.
09.006
Skudlarski, P., Jagannathan, K., Calhoun, V. D., Hampson, M.,
Skudlarska, B. UN., & Pearlson, G. (2008). Measuring brain con-
nectivity: Diffusion tensor imaging validates resting state tempo-
ral correlations. NeuroImage, 43(3), 554–561. https://est ce que je.org/10.
1016/j.neuroimage.2008.07.063
Forgeron, S. M., Vidaurre, D., Beckmann, C. F., Glasser, M.. F.,
Jenkinson, M., Miller, K. L., . . . Van Essen, D. C. (2013). Func-
tional connectomics from resting-state fMRI. Tendances cognitives
les sciences, 17(12), 666–682. https://doi.org/10.1016/j.tics.2013.
09.016
Sockeel, S., Schwartz, D., Pélégrini-Issac, M., & Benali, H. (2016).
Large-Scale Functional Networks Identified from Resting-State
EEG Using Spatial ICA. PLOS ONE, 11(1), e0146845. https://est ce que je.
org/10.1371/journal.pone.0146845
Sporns, O., Tononi, G., & Kötter, R.. (2005). The Human Connec-
tome: A Structural Description of the Human Brain. PLOS Com-
putational Biology, 1(4), e42. https://doi.org/10.1371/journal.pcbi.
0010042
Stam, C. J.. (2004). Functional connectivity patterns of human mag-
netoencephalographic recordings: A “small-world” network?
Neuroscience Letters, 355(1–2), 25–28. https://est ce que je.org/10.1016/
j.neulet.2003.10.063
Stam, Cornelis J., Nolte, G., & Daffertshofer, UN. (2007). Phase lag
index: Assessment of functional connectivity from multi chan-
nel EEG and MEG with diminished bias from common sources.
Human Brain Mapping, 28(11), 1178–1193. https://est ce que je.org/10.
1002/hbm.20346
Stephan, K. E., & Friston, K. J.. (2010). Analyzing effective connectiv-
ity with fMRI. Wiley Interdisciplinary Reviews. Sciences cognitives,
1(3), 446–459. https://doi.org/10.1002/wcs.58
Sun, F. T., Miller, L. M., & D'Esposito, M.. (2004). Measuring in-
terregional functional connectivity using coherence and partial
coherence analyses of fMRI data. NeuroImage, 21(2), 647–658.
https://doi.org/10.1016/j.neuroimage.2003.09.056
Tagliazucchi, E., Balenzuela, P., Fraiman, D., & Chialvo, D. R..
(2012un). Criticality in large-scale brain fMRI dynamics unveiled
by a novel point process analysis. Fractal Physiology, 3, 15.
https://doi.org/10.3389/fphys.2012.00015
Tagliazucchi, E., von Wegner, F., Morzelewski, UN., Brodbeck, V., &
Laufs, H. (2012b). Dynamic BOLD functional connectivity in hu-
mans and its electrophysiological correlates. Frontiers in Human
Neurosciences, 6. https://doi.org/10.3389/fnhum.2012.00339
Tewarie, P., Bright, M.. G., Hillebrand, UN., Robson, S. E.,
Gascoyne, L. E., Morris, P.. G., . . . Brookes, M.. J.. (2016). Predict-
ing haemodynamic networks using electrophysiology: The role of
non-linear and cross-frequency interactions. NeuroImage, 130,
273–292. https://doi.org/10.1016/j.neuroimage.2016.01.053
Tewarie, P., Hillebrand, UN., van Dellen, E., Schoonheim, M.. M.,
Barkhof, F., Polman, C. H., . . . Stam, C. J.. (2014). Structural
degree predicts functional network connectivity: A multimodal
resting-state fMRI and MEG study. NeuroImage, 97, 296–307.
https://doi.org/10.1016/j.neuroimage.2014.04.038
Tewarie, Prejaas, Hunt, B. UN. E., O’Neill, G. C., Byrne, UN., Aquino,
K., Bauer, M., . . . Brookes, M.. J.. (2018). Relationships Between
Neuronal Oscillatory Amplitude and Dynamic Functional Connec-
tivité. Cortex cérébral. https://doi.org/10.1093/cercor/bhy136
Thompson, Garth J.
(2017). Neural and metabolic basis of dy-
namic resting state fMRI. NeuroImage. https://est ce que je.org/10.1016/
j.neuroimage.2017.09.010
Thompson, Garth John, Merritt, M.. D., Pan, W.-J., Magnuson, M.. E.,
Grooms, J.. K., Jaeger, D., & Keilholz, S. D. (2013). Neural cor-
relates of time-varying functional connectivity in the rat. Neuro-
Image, 83. https://doi.org/10.1016/j.neuroimage.2013.07.036
Tononi, G., Sporns, O., & Edelman, G. M.. (1994). A measure for
brain complexity: Relating functional segregation and integration
in the nervous system. Proceedings of the National Academy of
Sciences of the United States of America, 91(11), 5033–5037.
Uhlhaas, P.. J., Pipa, G., Lima, B., Melloni, L., Neuenschwander,
S., Nikoli´c, D., & Chanteur, W. (2009). Neural synchrony in cor-
tical networks: Histoire, concept and current status. Frontiers in
Integrative Neuroscience, 3, 17. https://doi.org/10.3389/neuro.
07.017.2009
Uji, M., Wilson, R., Francis, S. T., Mullinger, K. J., & Mayhew,
S. D.
(2018). Exploring the advantages of multiband fMRI
with simultaneous EEG to investigate coupling between gamma
frequency neural activity and the BOLD response in humans.
Neurosciences en réseau
28
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Cross-modal review of connectome organization across temporal scales
Human Brain Mapping, 39(4), 1673–1687. https://est ce que je.org/10.
1002/hbm.23943
Académie nationale des sciences, 114(48), 12827–12832. https://
doi.org/10.1073/pnas.1705120114
Van De Ville, D., Britz, J., & Michel, C. M.. (2010). EEG microstate
sequences in healthy humans at rest reveal scale-free dynam-
ics. Actes de l'Académie nationale des sciences, 107(42),
18179–18184. https://doi.org/10.1073/pnas.1007841107
van den Heuvel, M.. P., Mandl, R.. C. W., Kahn, R.. S., & Hulshoff Pol,
H. E. (2009). Functionally linked resting-state networks reflect
the underlying structural connectivity architecture of the human
brain. Human Brain Mapping, 30(10), 3127–3141. https://est ce que je.
org/10.1002/hbm.20737
Van Dijk, K. R.. UN., Sabuncu, M.. R., & Buckner, R.. L. (2012).
The influence of head motion on intrinsic functional connectiv-
ity MRI. NeuroImage, 59(1), 431–438. https://est ce que je.org/10.1016/j.
neuroimage.2011.07.044
VanRullen, R.. (2016). Perceptual cycles. Trends in Cognitive Sci-
ences, 20(10). https://doi.org/10.1016/j.tics.2016.07.006
Varela, F., Lachaux, J.-P., Rodriguez, E., & Martinerie, J.. (2001).
The brainweb: Phase synchronization and large-scale integra-
tion. Nat Rev Neurosci, 2(4), 229–239. https://doi.org/10.1038/
35067550
Vidaurre, D., Hunt, L. T., Quinn, UN. J., Hunt, B. UN. E., Brookes,
M.. J., Nobre, UN. C., & Woolrich, M.. W. (2018). Spontaneous cor-
tical activity transiently organises into frequency specific phase-
coupling networks. Communications naturelles, 9(1), 2987. https://
doi.org/10.1038/s41467-018-05316-z
Vidaurre, D., Forgeron, S. M., & Woolrich, M.. W. (2017). Brain network
dynamics are hierarchically organized in time. Actes du
Viriyopase, UN., Bojak, JE., Zeitler, M., & Gielen, S. (2012). Quand
Long-Range Zero-Lag Synchronization is Feasible in Cortical
Networks. Frontiers in Computational Neuroscience, 6. https://
doi.org/10.3389/fncom.2012.00049
Wang, P., Kong, R., Kong, X., Liégeois, R., Orban, C., Deco, G., . . .
Yeo, B. T. T. (2019). Inversion of a large-scale circuit model reveals
a cortical hierarchy in the dynamic resting human brain. Science
Advances, 5(1), eaat7854. https://doi.org/10.1126/sciadv.aat7854
Weaver, K. E., Wander, J.. D., Ko, UN. L., Casimo, K., Grabowski,
T. J., Ojemann, J.. G., & Darvas, F. (2016). Directional patterns of
cross frequency phase and amplitude coupling within the rest-
ing state mimic patterns of fMRI functional connectivity. Neuro-
Image,128,238–251. https://doi.org/10.1016/j.neuroimage.2015.
12.043
Wirsich, J., Rey, M., Guye, M., Bénar, C., Lanteaume, L., Ridley, B.,
. . . Consortium, T. P.. (2017un). Brain Networks are Independently
Modulated by Donepezil, Sleep, and Sleep Deprivation. Cerveau
Topography, 1–12. https://doi.org/10.1007/s10548-017-0608-5
Wirsich, J., Ridley, B., Besson, P., Jirsa, V., Bénar, C., Ranjeva, J.-P.,
& Guye, M.. (2017b). Complementary contributions of concur-
rent EEG and fMRI connectivity for predicting structural con-
nectivity. NeuroImage, 161, 251–260. https://est ce que je.org/10.1016/j.
neuroimage.2017.08.055
Zhigalov, UN., Arnulfo, G., Nobili, L., Palva, S., & Palva, J.. M.. (2017).
Modular co-organization of functional connectivity and scale-
free dynamics in the human brain. Neurosciences en réseau, 1(2),
143–165. https://doi.org/10.1162/NETN_a_00008
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
1
1
1
8
6
6
6
8
7
n
e
n
_
un
_
0
0
1
1
4
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurosciences en réseau
29