ARTICLE DE RECHERCHE
Resting State Network Segregation Modulates
Age-Related Differences in Language Production
un accès ouvert
journal
Haoyun Zhang1,2
and Michele T. Diaz2
1Centre for Cognitive and Brain Sciences, University of Macau, Macau SAR, Chine
2Département de psychologie, Université d'État de Pennsylvanie, Parc universitaire, Pennsylvanie, Etats-Unis
Mots clés: cognitive aging, language production, network segregation, resting state functional
connectivité
ABSTRAIT
Older adults typically exhibit decline in language production. Cependant, how the brain supports
or fails to support these processes is unclear. De plus, there are competing hypotheses about
the nature of age-related neural changes and whether age-related increases in neural activity
reflect compensation or a decline in neural efficiency. In the current study, we investigated
the neural bases of language production focusing on resting state functional connectivity.
We hypothesized that language production performance, functional connectivity, and their
relationship would differ as a function of age. Consistent with prior work, older age was
associated with worse language production performance. Functional connectivity analyses
showed that network segregation within the left hemisphere language network was maintained
across adulthood. Cependant, increased age was associated with lower whole brain network
segregation. De plus, network segregation was related to language production ability. In both
network analyses, there were significant interactions with age—higher network segregation
was associated with better language production abilities for younger and middle-aged adults,
but not for older adults. Fait intéressant, there was a stronger relationship between language
production and the whole brain network segregation than between production and the
language network. These results highlight the utility of network segregation measures as an
index of brain function, with higher network segregation associated with better language
production ability. De plus, these results are consistent with stability in the left hemisphere
language network across adulthood and suggest that dedifferentiation among brain networks,
outside of the language network, is a hallmark of aging and may contribute to age-related
language production difficulties.
INTRODUCTION
Language is one of the most common and universal features of human society. Cependant, comme
people age, they often face some decline in certain aspects of language, particularly in spoken
language production (c'est à dire., speech; Burke & Shafto, 2008; Diaz et al., 2016). Par exemple, older
adults speak more slowly in a variety of situations (Duchin & Mysak, 1987; Mortensen et al.,
2006; Spieler & Griffin, 2006), have increased retrieval failures (par exemple., Burke et al., 1991; Zhang
et coll., 2019), produce more filler words (Horton et al., 2010), produce more omissions
(MacKay & James, 2004), produce more disfluent speech (Bortfeld et al., 2001; Obler & Albert,
1981), and produce less grammatically complex speech (par exemple., Kemper et al., 2003). Several
hypotheses have been proposed that these age differences in production may be due to
Citation: Zhang, H., & Diaz, M.. T. (2023).
Resting state network segregation
modulates age-related differences in
language production. Neurobiology of
Language, 4(2), 382–403. https://doi.org
/10.1162/nol_a_00106
EST CE QUE JE:
https://doi.org/10.1162/nol_a_00106
Informations complémentaires:
https://doi.org/10.1162/nol_a_00106
Reçu: 12 Juin 2022
Accepté: 28 Mars 2023
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Haoyun Zhang
haoyunzhang@um.edu.mo
Éditeur de manipulation:
Steven Small
droits d'auteur: © 2023
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
Resting state functional connectivity:
A method that examines how brain
regions work together and thus how
they form functional networks
without additional task demands.
Network segregation:
A measure that compares the
difference between within- et
between-network connectivities
divided by within-network
connectivité, providing a more
complete picture regarding the
network organization across the
whole brain.
declines in phonological processing (Burke et al., 1991; Burke & Shafto, 2008), aspects of
executive function (Hasher et al., 1991; Hoffman et al., 2018; Lustig et al., 2007), or process-
ing speed (Salthouse, 1996, 2000).
Although these behavioral age-related differences in language production have been com-
monly observed, how the brain supports or fails to support these processes is still not clear.
De plus, there are competing hypotheses about the nature of age-related neural changes.
One prominent account is the dedifferentiation hypothesis, which suggests that age-related
increases in neural activity reflect a decline in neural efficiency (Li et al., 2001). Others have
suggested that age-related increases in neural activity are compensatory (Cabeza et al., 2018).
One approach to investigating brain function is with resting state functional connectivity
(RSFC), which examines how brain regions work together and thus how they form functional
réseaux (Biswal et al., 1995; Harmelech & Malach, 2013). While RSFC can be used to inves-
tigate specific local networks, it can also measure many networks, thereby capturing a whole
brain profile that can be used to investigate how overall brain function relates to cognition.
Although studies have reported that more efficient brain networks are associated with
enhanced cognition, few studies have focused on language production. Donc, the current
study investigated the relationship between RSFC and age-related differences in spoken lan-
guage production, using network segregation, a measure that captures both within network
and between network connectivities. We hypothesized that more segregated brain networks
would be associated with enhanced language production and that this relationship would be
modulated by age such that older adults would show less differentiated networks and weaker
brain–behavior relationships.
Age-related behavioral differences in cognition and spoken language production are com-
monly observed, and this may be related to age-related differences in the brain. Even in the
absence of disease, some degree of neural decline is a natural aspect of aging. Par exemple,
as people age they often show structural changes in the brain, such as decreased gray matter
volume (Raz et al., 2005; Sowell et al., 2004) and decreased white matter integrity (Bartzokis
et coll., 2012; Imperati et al., 2011; Mwangi et al., 2013). In addition to structural brain changes,
older and younger adults also exhibit different patterns of task-based functional activation, avec
older adults often showing increases in brain activation (par exemple., greater bilateral prefrontal activation;
Cabeza & Dennis, 2013; Davis et al., 2008; Diaz et al., 2021; Grady et al., 2015; Wierenga
et coll., 2008; Zhang et al., 2019). Although age-related differences in functional activation are
commonly observed, the mechanism underlying such changes has been debated. Compensatory
accounts suggest that age-related increases in functional activation may reflect greater neural
fiançailles, which then translates to maintained or improved performance (Cabeza et al.,
2018). On the other hand, dedifferentiation accounts suggest that increased activation reflects
neural inefficiency and may lead to age-related declines in behavior (Li et al., 2001).
In addition to examining age-related differences in brain structure and task-based functional
activation, another way to explore age-related differences in the brain is to investigate func-
tional connectivity. Functional connectivity analyses use functional magnetic resonance imag-
ing (IRMf) data to examine how signals from different brain regions covary (c'est à dire., patterns of
correlated brain activity). These patterns of functional connectivity reflect how different brain
regions work together and interact with each other (Friston, 1994). When brain regions work
together or are functionally connected, they are said to form networks. Common functional
networks include the default mode network (DMN), the salience network (SN), et le
frontal-parietal network (FP; Power et al., 2011), with each network serving different cognitive
les fonctions (par exemple., DMN and monitoring, FP and attentional control). De plus, functional con-
nectivity analyses can be particularly useful in examining age-related differences as the
Neurobiology of Language
383
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
approach minimizes task demands which can affect older and younger adults differentially.
Data are often collected during rest where participants are asked to simply relax and look
at a fixation cross. These resting state data reflect spontaneous brain activity in the absence
of any explicit task.
Resting state data can be used to examine connectivity within a single network (c'est à dire., within
network connectivity), which reflects how activity is correlated among those regions and is
thought to reflect network integrity. Many previous studies on age-related differences in brain
activity have focused on specific networks such as the DMN or FP (Power et al., 2011). UN
common finding is that increased age is often associated with lower within-network functional
connectivité (Betzel et al., 2014; Cao et al., 2014; Geerligs et al., 2015; Onoda et al., 2012;
Siman-Tov et al., 2017; Song et al., 2014; Tomasi & Volkow, 2012; Zhang, Gertel, et coll., 2021).
De plus, these age-related differences in within-network connectivity have been associated
with worse behavioral performance across several aspects of cognition, including episodic
mémoire (Wang et al., 2010), fluid intelligence (Onoda et al., 2012), and other cognitive abilities
(for a review, see Sala-Llonch et al., 2015). De plus, several studies have investigated the rela-
tionships between aging, functional connectivity, and language (Antonenko et al., 2013; Ferré
et coll., 2019; Gertel et al., 2020; Krieger-Redwood et al., 2019; Pistono et al., 2021; Zhang, Bai,
et coll., 2021). Consistent with the broader literature, studies that have examined within-network
connectivity and language have found weaker within-network connectivity among older adults
compared to younger adults, and stronger within-network connectivity has been associated
with better language functions in, Par exemple, semantic control (Wawrzyniak et al., 2017),
verbal fluency (Miró-Padilla et al., 2017), and syntax (Antonenko et al., 2013).
While within-network activity reflects coherence or integrity of that network, réseaux
often interact with one another both beneficially (par exemple., coordinating the observation of an
event with action) and detrimentally (par exemple., failing to monitor because of distraction from a
salient event). Ainsi, examining connectivity across networks is also important in understand-
ing overall brain functioning. Between-network connectivity can be examined in two primary
ways. The first approach considers all or most of the brain’s networks, and examines how net-
works interact with one another to provide an overall picture of network interaction and brain
fonction (c'est à dire., whole brain between-network functional connectivity). Previous studies taking
this approach have found higher whole brain connectivity among older adults during language
tasks such as synonym/antonym decisions and picture naming (Ferré et al., 2019). Examining
the relationship of these findings to behavior revealed that higher whole brain between-
network resting state connectivity was associated with poorer language ability among older
adultes (Zhang, Gertel, et coll., 2021), suggesting that increased whole brain between-network
connectivity is related to worse cognitive performance. The second approach is more focal
and examines interactions between specific regions and/or networks to better understand
how two networks or regions might interact. Studies that have taken this more focal approach
suggest that some increases in connectivity may be compensatory (Gertel et al., 2020; Pistono
et coll., 2021). Gertel and colleagues found that although older adults had overall decreased
functional connectivity compared to younger adults, older adults who had stronger resting
state functional connectivity between left inferior frontal gyrus and right hemisphere executive
function regions performed better on the Stroop task. These findings suggest that the increased
RSFC between a core language region and executive function regions had a compensatory
fonction. De la même manière, Pistono et al. (2021) reported that stronger connections between the lan-
guage network (c'est à dire., a left lateralized frontal-temporal network involved in core language func-
tion) and the multiple demand network (c'est à dire., a bilateral frontal-parietal network involved in
domain general cognitive functions) were associated with better verbal fluency performance in
Neurobiology of Language
384
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
older adults. Ainsi, these two approaches to examining between-network connectivity provide
slightly different views of brain function. The whole brain approach reflects overall brain func-
tion and has generally shown higher between-network connectivity for older adults that is
associated with worse performance. The more focal approach suggests that higher between-
network connectivity, at least between language and executive regions, may have a compen-
satory function (par exemple., Cabeza et al., 2018; Reuter-Lorenz & Capelle, 2008).
Cependant, examining either within-network or between-network connectivities in isolation
does not illustrate the whole picture of aging and brain function. One way to integrate both
within- and between-network characteristics is to use a measurement of network segregation
(Chan et coll., 2014). Network segregation compares the difference between within- et
between-network connectivities divided by within-network connectivity. Ainsi, network segrega-
tion provides a more complete picture regarding the network organization across the whole
brain. Several studies have used network segregation to investigate how it relates to age-related
differences in cognition (Chan et coll., 2014; Chan et coll., 2018; King et al., 2018; Varangis et al.,
2019). The findings showed that increasing age was associated with lower network segregation,
consistent with previous findings looking at within- and between-network connectivity sepa-
rately. En plus, lower network segregation across the whole brain has been related to lower
cognitive performance in the domains of speed, episodic memory, and fluid intelligence (Chan
et coll., 2014; Chan et coll., 2018; King et al., 2018; Varangis et al., 2019). Cependant, these integrated
measures of brain function have not been used to examine language and aging to date.
Donc, in the current study, we used a whole brain network approach to investigate the
relationship between functional connectivity and age-related differences in language produc-
tion. D'abord, we used network segregation as our measure to examine how language production
is related to language network integrity. Cependant, language production involves more than
just language ability per se; it also relies on perceptual regions for visual processes and object
recognition and motor processing for articulation (Guenther et al., 2006; Tremblay et al.,
2016), as well as other cognitive abilities such as monitoring, planning, and executive func-
tioning (for review, see Burke & Shafto, 2008; Diaz et al., 2016). C'est, language production
ability may involve not only the language network, but also other brain networks. Donc
our second goal was to investigate how language production is associated with overall, whole
brain network integrity, using network segregation calculated across the brain. Based on
previous studies, we predicted that there would be main effects of age on both language pro-
duction ability and network measures. Spécifiquement, older adults would show lower language
production ability and lower network segregation for both the language network and the
whole brain. En plus, we hypothesized that both network measures would be associated
with language production ability and that more segregated networks would be related to better
language production. De plus, the patterns in these relationships can inform our understand-
ing of cognitive aging. If increased age is related to weaker brain–behavior relationships, ce
would be consistent with a dedifferentiation account of aging (Ghisletta & Lindenberger, 2003;
Li et al., 2001). On the other hand, if high performing older adults rely on a broader neural
network to support language functions compared to younger adults, this would be consistent
with compensatory accounts of aging (Cabeza et al., 2018).
MATERIALS AND METHODS
Participants
Ninety adults (ages: 22–78 yr) participated in the experiment. All participants were
community-dwelling, right-handed, native English speakers who were not fluent in a second
Neurobiology of Language
385
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
langue. All participants had normal or corrected-to-normal vision and reported no history of
neurological, psychologique, or major medical conditions (Christensen et al., 1992). Eight
participants were removed from the analysis because of missing data points—seven did not
have picture naming data (5 did not finish the task and there were 2 recording failures) et
one participant did not complete the story elicitation task. Three more participants were
removed because of having outlier data points on behavioral measurements (see Data
Analyses for details), leaving 79 participants’ data in the final analyses (ages: 22–78 yr, mean
age = 48.2 yr, SD = 16.1 yr; 49 female).
Every participant first completed a behavioral testing session with a battery of psychometric
and neuropsychological tests to assess basic cognitive functions such as speed, executive
fonction, mémoire, and language. The psychometric tasks included the Mini-Mental State
Exam to screen for mild cognitive impairment or dementia (MMSE; Folstein et al., 1975),
and the Geriatric Depression Score (GDS) short version to screen for depression (Guerin
et coll., 2018; Sheikh & Yesavage, 1986). Participants also completed several standardized or
adapted neuropsychological assessments. For processing speed and general executive func-
tion, participants completed simple (c'est à dire., respond to a black square as quickly as possible)
and choice (c'est à dire., identify the direction of left/right arrows as quickly as possible) reaction time
tests; WAIS-III vocabulary assessed vocabulary size, forward and backward digit span assessed
working memory, and a computerized adaptation of the digit-symbol subtest assessed process-
ing speed (Wechsler, 1997); and a computerized color Stroop task assessed executive function
(c'est à dire., make a response to the color of the ink when it is consistent/inconsisent with the word
meaning (MacLeod, 1991; Stroop, 1935). Participants also completed a reading span task to
assess verbal working memory (Conway et al., 2005), and the California Verbal Learning Test
to assess immediate and delayed memory (c'est à dire., one learning trial, 16 word list in 4 catégories,
one immediate recall assessment, one delayed recall assessment; Woods et al., 2006). Pour
language assessments, participants completed the author recognition test and a comparative
reading habit questionnaire to assess reading habits (Acheson et al., 2008).
En plus, participants performed several tasks measuring different aspects of language
production. These tasks included a verbal fluency task (Patterson, 2011), a picture naming
task, and a speech elicitation task. During the verbal fluency task, participants were asked
to overtly generate as many words as possible in each phonemic (F, UN, S) and categorical
(animals, supermarkets) catégorie. Participants were given one minute per category to respond
and were asked to avoid saying proper names of people or places, and repetitions of words.
During the picture naming task, pictures of different objects were presented in the center of the
screen and participants were asked to overtly name each picture as quickly and accurately as
possible. During the speech elicitation task, participants were asked to generate a story from
the picture book Frog, Where Are You?, by Mercer Mayer. They were encouraged to tell the
story to the experimenter as if the experimenter had never heard the story before. Les participants
responses were recorded for offline analyses and there was no time limit for them to respond.
Demographic characteristics and assessment scores are reported in Table 1. All participants
gave written, informed consent, and were paid for their participation ($15–30/hr). All proce-
dures were approved by the Institutional Review Board at Pennsylvania State University.
Acquisition of MRI Data
All imaging data were acquired on a 3T Siemens Prisma Fit scanner using a 64-channel head
coil. Localizer images were collected and used to define a volume for data collection, higher-
order shimming, and alignment to the anterior commissure and posterior commissure (AC-PC).
Neurobiology of Language
386
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
Tableau 1.
Participant demographics, neuropsychological testing scores, and correlation with age
Demographic information
N
Age (années)
Gender (M/F)
Participant characteristics
Éducation (années)
MMSE (score out of 30)
Depression (GDS) (score out of 15)
Cognitive assessments
Simple RT (box, ms)
Choice RT (arrow, ms)
WAIS vocabulary (score out of 66)
Digit symbol RT (ms)
Digit span forward (score out of 16)
Digit span backward (score out of 16)
Stroop effect RT (Incongruent–Congruent, ms)
Verbal working memory (score out of 1)
Immediate recall (score out of 16)
Delayed recall (score out of 16)
Author Recognition Testa (score out of 76)
Comparative reading (score out of 35)
Language production measures
Verbal fluency (number of correct responses)
Phonemic fluency (F, UN, S)
Category fluency (animal and supermarket)
Picture naming RT (ms)
Story elicitation task
Mean length of utterance
Moving average type token ratiob
Mean (SD)
79
48.2 (16.1)
30/49
16.8 (3.1)
28.8 (1.1)
0.8 (1.0)
298.1 (52.7)
352.0 (75.6)
54.7 (5.2)
1574.0 (384.6)
11.0 (2.1)
7.3 (2.0)
58.8 (80.6)
0.4 (0.2)
10.6 (2.5)
8.9 (2.9)
24.9 (14.1)
26.0 (4.7)
88.1 (14.9)
41.3 (9.5)
46.8 (8.7)
Range
22–78
6–25
19–30
0–5
237.3–479.9
252.6–781.9
41–66
889.0–3001.0
6–16
4–14
−63.7–420.0
0.02–0.8
3–16
2–16
3–64
11–35
50–125
16–68
27–69
1046.7 (134.2)
741.3–1430.0
9.4 (2.1)
0.66 (0.04)
5.6–18.7
0.54–0.74
Age regression
−0.009
−0.004
−0.01
0.78*
2.48***
0.04
16.5***
−0.003
−0.007
1.90***
−0.002
−0.05**
−0.05*
0.50***
0.05
−0.24*
−0.06
−0.18**
1.24
−0.01
−0.0004
Note. The second column displays raw score means, with standard deviations (SD). The third column indicates the score range of each test. The fourth column
indicates its regression coefficient with age. GDS = Geriatric Depression Score; MMSE = Mini-Mental State Exam; RT = reaction time; WAIS = WAIS-III: Wechsler
Adult Intelligence Scale.
a Author Recognition Test scores are calculated as the number of correct identifications − the number of incorrect responses.
b MATTR used a moving window of 50 words.
* p < 0.05; ** p < 0.01; *** p < 0.001.
Neurobiology of Language
387
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
T1-weighted anatomical images were then collected using a magnetization-prepared rapid
acquisition gradient echo (MPRAGE) sequence (repetition time [TR] = 2,300 ms; echo time
[TE] = 2.28 ms; inversion time [TI] = 900 ms; flip angle = 8°; echo spacing = 7 ms; acceleration
factor = 2; field of view [FOV] = 256 mm2; voxel size = 1 × 1 × 1 mm; 160 contiguous slices).
After the structural scan, blood-oxygen level dependent (BOLD) resting state data were
acquired using an echoplanar imaging (EPI) sequence (TR = 2,000 ms; TE = 25.0 ms; flip
angle = 90°; echo spacing = 0.49 ms; FOV = 240 mm2; voxel size = 3 × 3 × 4 mm; 33
contiguous slices, parallel to the AC-PC; phase encoding = anterior to posterior, fat saturation =
on; slice acquisition = sequential, descending; volumes = 180; run duration = ~6 min). Two
additional volumes were acquired and deleted at the start of the scan to reach steady state
equilibrium. During the resting state run, participants were instructed to relax in the scanner
with their eyes open and to look at a fixation cross presented in the center of the screen with-
out falling asleep. Four task-based runs using the same parameters as the resting state run were
also collected after the resting state run (task run duration = ~5.6 minutes). During the task
runs, participants were presented with words and were asked to read aloud words as quickly
and accurately as possible. Results from the task will be reported elsewhere; here we focus
only on the resting state data.
Finally, a field map sequence was performed with a double-echo spoiled gradient echo
sequence (TR = 446 ms; TE = 4.92 ms; flip angle = 60°; FOV = 240 mm2; voxel size = 3 ×
3 × 4 mm; 33 contiguous slices; phase encoding = anterior to posterior, fat saturation = off;
duration = 1:12 min) that generated two magnitude images and one phase image that were
used for correcting susceptibility distortions in the functional data.
fMRI Data Preprocessing
Data quality was first assessed using the fBIRN QA tool (Glover et al., 2012; https://www.nitrc
.org/projects/bxh_xcede_tools/), measuring the number of potentially clipped voxels, mean
signal fluctuation-to-noise ratio (SFNR), and per-slice variation. Additionally, the anatomical
and functional images were visually inspected for artifacts and signal drop-out. Preprocessing
analyses were carried out using the CONN functional connectivity toolbox ( Version 18.a)
under the MATLAB environment (Whitfield-Gabrieli & Nieto-Castanon, 2012). Preprocessing
steps included functional realignment and unwarping to estimate and correct for participant
motion, distortion correction using a voxel-displacement map calculated based on the field
map, and a slice-timing correction which corrected for maturation of the BOLD signal over
time (Huettel et al., 2004). Additionally, functional outliers were detected with an ART (Artifact
Detection Tools)-based identification method (NITRC, 2015), in which outliers were defined
using a conservative threshold (i.e., 97th percentile), and subsequently removed. All anatom-
ical and functional images were normalized into standard Montreal Neurological Institute
(MNI) space. The anatomical images were segmented into gray matter, white matter, and cerebral
spinal fluid (CSF) tissue classes using SPM12 unified segmentation and normalization procedure,
then these masks were applied to the functional images (Ashburner & Friston, 2005). During reg-
istration, functional images were aligned to anatomical images and both were normalized to
standard space. A smoothing kernel of 6 mm was used to increase the signal-to-noise ratio, as
well as to reduce spurious activations of single voxels. During denoising, the representative
noise signal from white matter (5 components) and CSF (5 components) was extracted, and
any signal correlated with these components was removed from the BOLD signal. The noise
removal used the CompCor approach, which extracts multiple signals from CSF and white
matter areas to capture motion and physiological artifacts while excluding neural signals,
Neurobiology of Language
388
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
which avoids introducing artifactual negative correlations in the connectivity measures (Chai
et al., 2012; Liu et al., 2017; Liu et al., 2021). To eliminate frequencies of less interest, a
band-pass filter (0.008, 0.09) was used (Davey et al., 2013; Gohel & Biswal, 2015; Hallquist
et al., 2013). The effects of the following quality assurance parameters were controlled for dur-
ing data analysis: number of outlier and non-outlier scans (outlier threshold = 0.5 mm), max
and mean motion, and max and mean global BOLD signal changes (outlier threshold = global-
signal z-value of 3). The average number of invalid scans was 1.9 out of 180 scans/volumes
(1%, SD = 5.4), and it was not significantly affected by age (β = 0.71, SE = 0.60, p = 0.24).
The mean amount of motion was 0.20 mm (SD = 0.08 mm), and increased age was associated
with higher head motion (β = 0.03, SE = 0.01, p < 0.001). The analyses removing variance
associated with the variables described above occurred in a single linear regression step,
and the residualized BOLD signal was used for further statistical analyses.
Node Definition and Network Measures
Power et al. (2011) identified 264 coordinates in the brain and created 5 mm fixed-radius
sphere nodes around these locations. We also used the same 264 locations and created the
5 mm radius non-overlapping nodes using the MNI152, 2 mm brain as the reference. Power
identified 12 networks (hand somatomotor, mouth somatomotor, visual, salience, auditory,
cingulo-opercular control, frontoparietal control, ventral attention, dorsal attention, default,
subcortical, and cerebellar) and divided nodes into these networks. Among all nodes, 33 were
excluded from the analysis due to poor classification fit with the Power networks. To further
identify nodes that belong to the language network, we used the language regions identified by
Fedorenko et al. (2010; left language network and right hemisphere homologues). We selected
the networks defined by Fedorenko and colleagues because the regions were identified func-
tionally (sentences > nonwords), were replicable within subjects, had a clear correspondence
across subjects and support broad language processes. Although it can be argued that this net-
work is a comprehension-based network, follow-up studies also suggested that it overlaps sig-
nificantly with the production network (Hu et al., 2023). The language network supports both
lexical access and sentence generation during language production. Donc, the left lan-
guage network they defined represents a broad language processing network that supports
both language comprehension and production. After defining these left language and right
hemisphere homologue networks, any nodes that overlapped with these two localizers were
categorized as the left language network and the right hemisphere homologue network. Le
remaining nodes were then binned across the 12 Power networks according to their location.
Nodes were double checked to ensure that no location belonged to more than one network.
The final set included 231 nonoverlapping nodes belonging to 14 réseaux. (See Table S1 in
the Supporting Information, available at https://doi.org/10.1162/nol_a_00106, for MNI coor-
dinates for node locations.)
For each participant, the resting state time series of each node was extracted, then a cross-
correlation of each node’s time course with every other node’s time course was calculated.
This was performed using the CONN functional connectivity toolbox ( Version 18.a) under the
MATLAB environment (Whitfield-Gabrieli & Nieto-Castanon, 2012). Correlation coefficients
were converted to Z-values using Fisher’s equation. Consistent with previous studies using
similar approaches (Chan et coll., 2014), negative correlations were not included in further anal-
ysis because of uncertainty regarding the meaning of negative correlations (Hallquist & Hillary,
2018). The final matrix for each participant was a 231 × 231 weighted Z-matrix with the diag-
onal and negative values set to zero. Correlation matrices were then imported in R for further
traitement (RStudio Team, 2022). The following R packages were used in the analyses: readxl
Neurobiology of Language
389
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
(Wickham & Bryan, 2019); tidyverse (Wickham et al., 2019); car (Fox & Weisberg, 2019);
interactions (Long, 2019); ggplot2 (Wickham, 2016); and ggpubr (Kassambara, 2020).
Using the same methods as Chan et al. (2014), for each participant we calculated network
segregation for the left language network and across the whole brain. D'abord, network definitions
were applied to each participant’s 231 × 231 weighted correlation matrix comprising 14
réseaux. Alors, for each network, within-network connectivity was calculated as the
mean node-to-node correlation of all nodes in that network (c'est à dire., diagonal blocks). Entre-
network connectivity was calculated as the mean correlation value between each node in
one network and the rest of the nodes outside of that network (c'est à dire., off diagonal blocks). Enfin,
network segregation was calculated as the difference between within-network connectivity and
between-network connectivity for a specific network divided by the within-network connec-
tivity of that network. The overall, whole brain network measures were calculated by averag-
ing the network segregation measures across all networks. All of the measures were calculated
at the subject level first before calculating group-level results (c'est à dire., each subject had one value
for left language network segregation, and one value for overall network segregation).
Data Analyses
As mentioned earlier, participants performed a series of neuropsychological tests to measure
cognitive functions across different domains (see Table 1 for the list of cognitive assessments).
For the purpose of the current study, we focused on variables measuring different aspects of
spoken language production (c'est à dire., verbal fluency, picture naming, and story elicitation). Pour
verbal fluency, repetitions of the same words, incorrect responses, and proper names were
excluded from the analysis. Responses were included if they matched the cue, and mythical
animals were counted as correct for the animals category (par exemple., unicorn, hippogriff ). The total
number of included responses for all categories were used as the verbal fluency score. For the
picture naming task, participants’ responses were coded offline and only responses that
matched the picture were coded as correct. The mean reaction time for correctly named
pictures was used as the picture naming performance score. For the story elicitation task,
participants’ stories were first transcribed using the CLAN software (MacWhinney & Wagner,
2010). For each participant, the mean length of utterances (MLU) measures utterance length
by calculating the ratio of total morphemes to the number of utterances, representing speech
complexity. An utterance is defined as a string of words that is followed by a pause of one
second or more, ends with a terminal intonation contour, or has a complete grammatical struc-
ture (not a necessary feature). Moving average type token ratio (MATTR) can capture lexical
diversity better than the traditional TTR measurement. TTR is obtained by dividing the number
of different words by the total number of words in the text, which is constrained by the overall
document length. MATTR, on the other hand, uses a fixed window length (50 words in the
current study) and computes the TTR for a moving window (1–50, 2–51, and so on to the
end of the text), and then the mean TTR is calculated for all windows. Donc, MATTR is
a length-invariant measurement of lexical diversity of spoken language production. All coding
was performed by trained research assistants and verified by a second research assistant.
To reflect the overall language production ability and different aspects of speech (par exemple.,
speed, complexity, and lexical diversity), a composite score was calculated for each partici-
pant by adding up the Z scores of the four production measurements (verbal fluency total,
picture naming reaction time, MLU, and MATTR; Tableau 1). To make the direction of effects
consistent across variables (c'est à dire., higher values reflecting better ability), picture naming reaction
times were reverse coded. To investigate age-related differences in language production, un
Neurobiology of Language
390
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
linear regression was conducted on the language production composite scores, including age
as the predictor. Three outliers were identified and removed from all analyses (Cook’s Distance
greater than 4/sample size; Cook, 1979). Donc, all analyses included a final set of 79 par-
ticipants. Enfin, all independent variables were standardized using the scale function in the R
environment ((score-mean)/SD) prior to conducting the linear regressions described below.
To investigate the relationships among age, brain network characteristics, and language
production, we first looked at age-related differences in language production by conducting
a linear regression of age on the language production composite scores. Suivant, because we were
interested in the language network, a simple linear regression was conducted on network
segregation within the left language network, including age as the predictor. To look at the over-
tous, whole brain structure, we also fit a linear regression on the whole brain network segregation
while including age as the predictor. En outre, to investigate how language production
relates to the language network segregation as well as the whole brain network structure, nous
conducted two sets of analyses on the language production composite scores. One set looked at
the contribution of the left language network to language production, and included age, gauche
language network segregation, and their interaction as independent variables. The other set
of analyses explored the contribution of the whole brain network structure to language pro-
duction, and independent variables included the main effects of age and whole brain network
segregation, as well as their interaction. For cases where the interactions were significant,
Johnson-Neyman tests were then conducted to identify the age ranges where the relationships
between network measures and the language production composite scores were significant
(Esarey & Sumner, 2018; Johnson & Fay, 1950). Spécifiquement, the Johnson-Neyman test reports
the value or values of the moderator (c'est à dire., âge) at which the effect of the predictor (c'est à dire., réseau
measures) on the dependent variable (c'est à dire., language production composite scores) was signif-
icant. En plus, given that head motion significantly differed across age, as indicated in the
fMRI Data Preprocessing section, all network analyses included the amount of motion as a
control variable to account for any remaining confounding effects of motion.
Enfin, although we focused on the relationships between a broad language production
measure and whole brain network segregation, we conducted additional analyses on each
language production variable and the relationships between the language network and each
other network to provide a more comprehensive picture. These analyses are touched upon
briefly in the results and reported in full in the Supporting Information.
RÉSULTATS
Age-Related Differences in Language Production
To examine age-related differences in language production, we conducted a linear regression
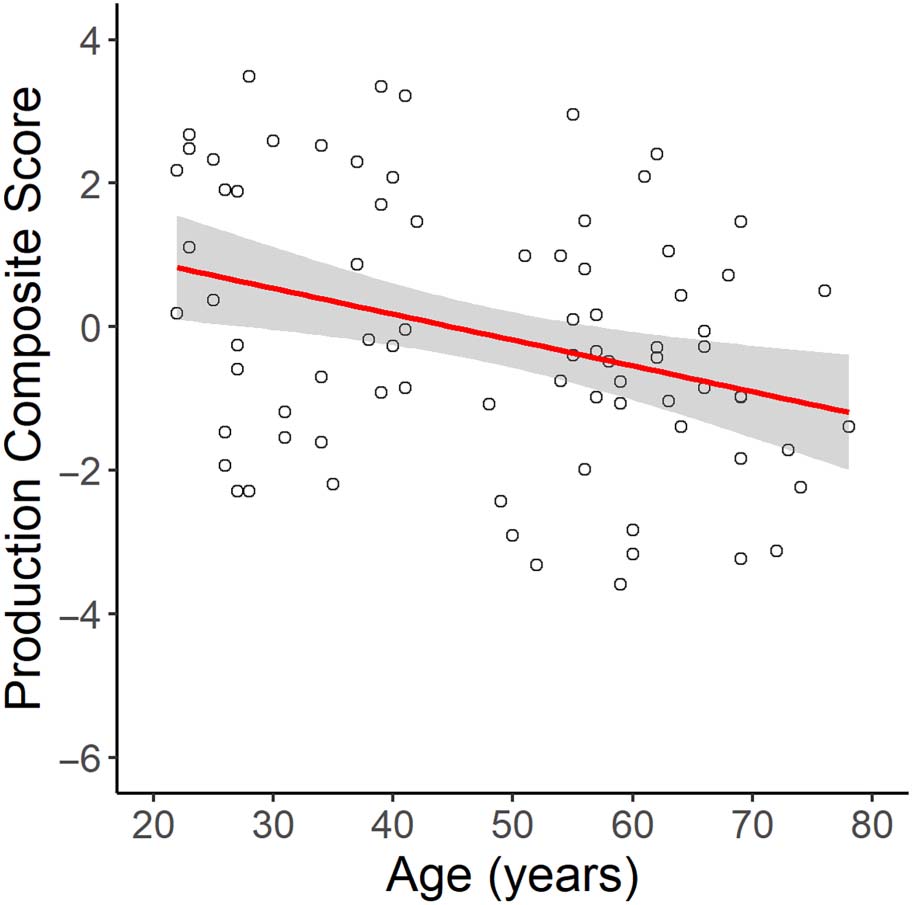
of age on the production composite scores. This showed that there was a significant effect of
age on language production (β = −0.59, SE = 0.20, p = 0.003). Spécifiquement, increasing age was
associated with lower production scores,
indicating lower language production ability
(Chiffre 1). As shown in Table 1 and Figure S2 in the Supporting Information, when looking
at the age effects on each language production measurement separately, there was only a sig-
nificant effect of age on the verbal fluency score (β = −0.24, SE = 0.12, p = 0.04).
Age-Related Differences in Network Structure
Linear regressions were conducted to investigate the effects of age on network segregation in
the language network and across the whole brain. Although there was no significant
Neurobiology of Language
391
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
Chiffre 1. The main effect of age on the language production composite score. Age was signifi-
cantly negatively correlated with production scores, where lower production scores indicate worse
performance.
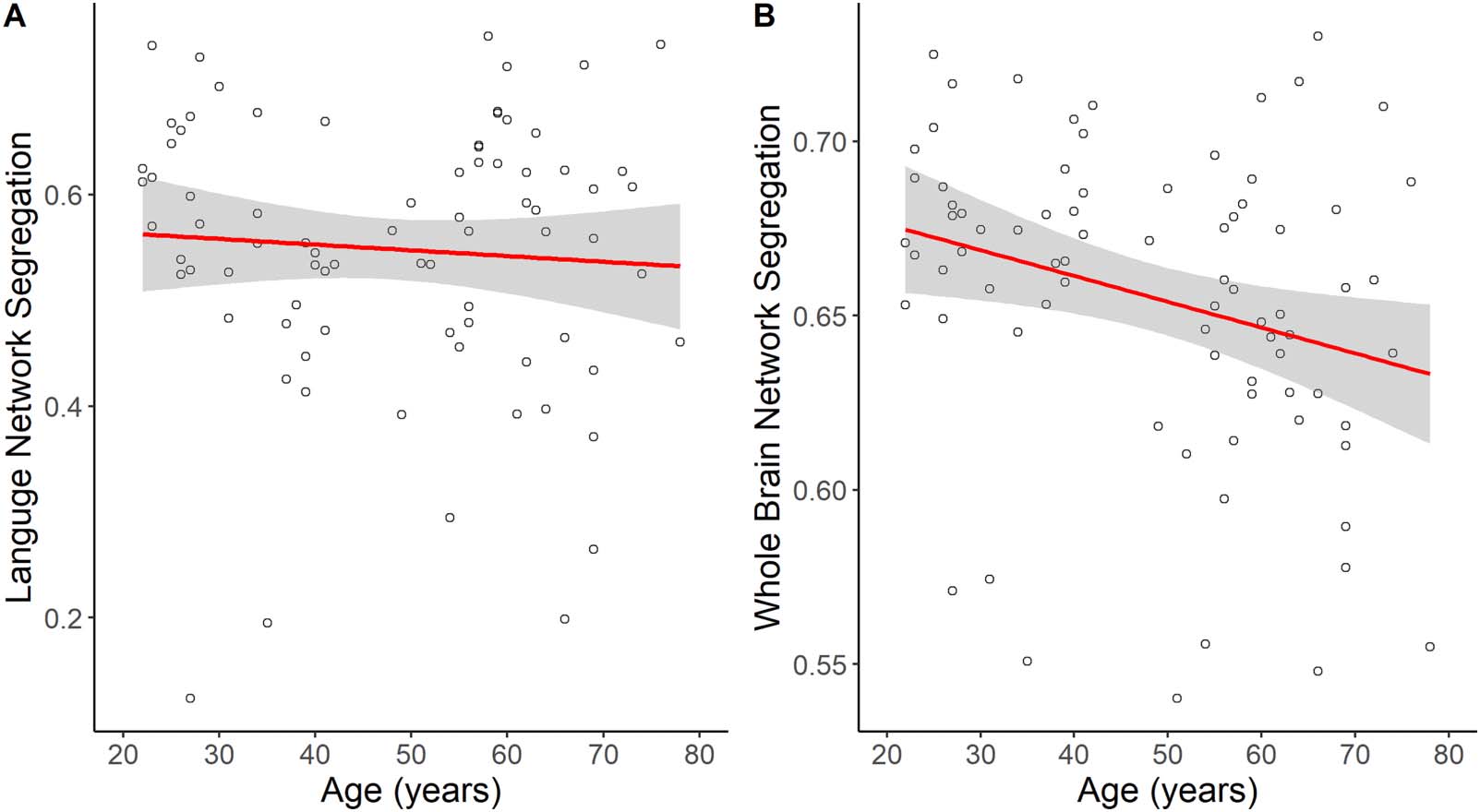
relationship between age and left language network segregation (β = −0.01, SE = 0.02, p =
0.47; Figure 2A), older age was significantly associated with lower whole brain network seg-
regation (β = −0.01, SE = 0.01, p = 0.02; Figure 2B).
Relationship Among Age, Network Structure, and Language Production
To look at how language production ability relates to the language network as well as the whole
brain network, we conducted two analyses, one using network segregation of the language
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 2. The main effects of age on language network segregation and whole brain network segregation. The effect of age on (UN) langue
network segregation was not significant, mais (B) whole brain network segregation was significantly lower with increasing age.
Neurobiology of Language
392
Network segregation supports language production across adulthood
network and one examining whole brain network segregation. For the first analysis, a regression
was conducted on the language production composite scores, including age, left language net-
work segregation, and their interaction as independent variables. Consistent with the results
from our previous regression, increasing age was significantly associated with lower language
production composite scores (β = −0.55, SE = 0.21, p = 0.01). Although the main effect of lan-
guage network segregation on the language production composite scores was not significant
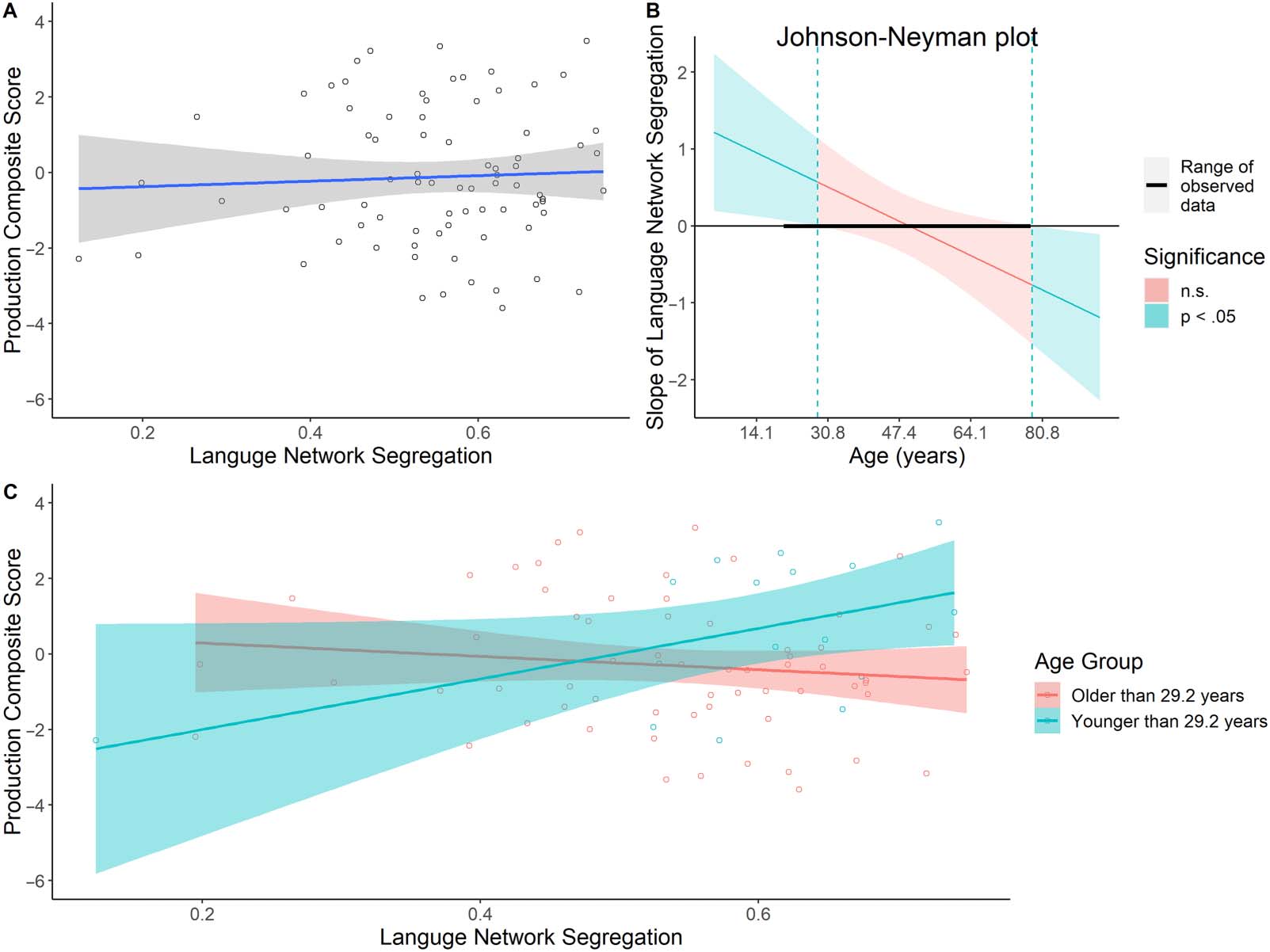
(β = 0.06, SE = 0.19, p = 0.75; Figure 3A), the interaction between age and language network
segregation was significant (β = −0.45, SE = 0.18, p = 0.02). To further specify the interaction,
we conducted a Johnson-Neyman test (Figure 3B). Results showed that the positive relationship
between language network segregation and language production scores was significant at ages
up to 29.2 années. These results suggest that higher language network segregation was associated
with better language production ability but only in relatively young adults (Figure 3C).
Deuxième, we investigated how language production was affected by the whole brain net-
work structure and age using a regression on the language production composite scores while
including age, whole brain network segregation, and their interaction in the model. Consistent
with prior models, age was significantly negatively associated with language production
composite scores (β = −0.44, SE = 0.21, p = 0.04). The main effect of whole brain network
segregation on the language production composite scores was significant (β = 0.42, SE = 0.20,
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
un
_
0
0
1
0
6
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 3. Age-related differences in the relationship between language network segregation and language production composite scores. (UN)
There was no significant relationship between language network segregation and language production. (B) The age ranges for which
interaction between language network segregation and language production ability
the Johnson-Neyman test
(22–29.2 yr old, for our sample). (C) Although there was no significant main effect of language network segregation on language production,
the interaction between these variables was significant. For younger adults (< 29.2 yr old), there was a significant positive relationship
between language network segregation and language production scores. This relationship was not significant in adults older than 29.2 years
old. n.s. = not signitficant.
identified a significant
Neurobiology of Language
393
Network segregation supports language production across adulthood
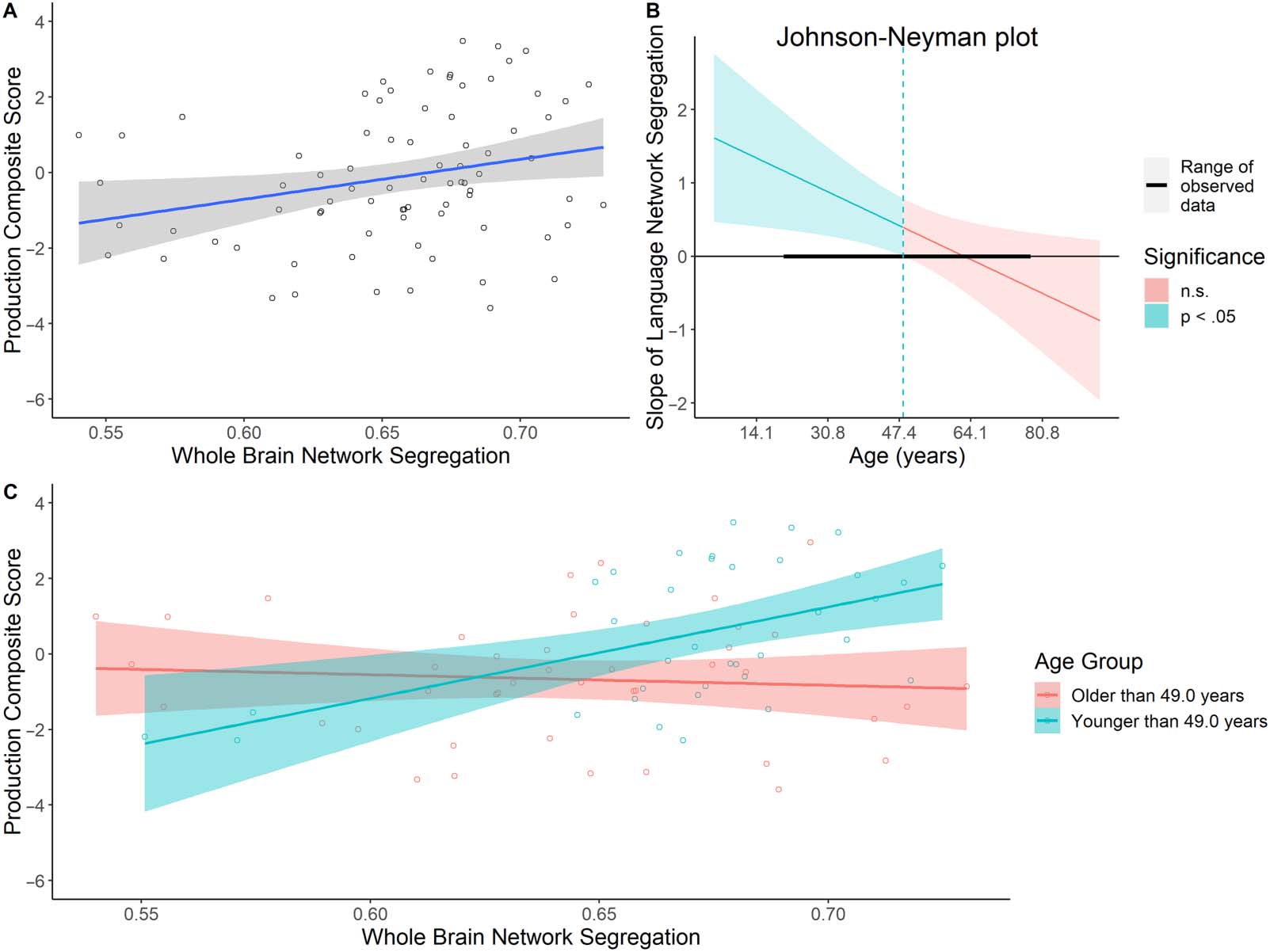
p = 0.04). Specifically, higher whole brain network segregation was associated with higher
production scores in general (Figure 4A). Moreover, the interaction between age and the
whole brain network segregation was significant (β = −0.46, SE = 0.19, p = 0.02). The
Johnson-Neyman test indicated that for individuals younger than 49.0 years of age, there
was a significant relationship between whole brain network segregation and language produc-
tion scores, with higher network segregation associated with higher language production
scores. This suggests that having more segregated networks in general was associated with
better language production in younger and middle-aged adults (Figure 4B and 4C).
Moreover, since older age was significantly associated with lower gray matter volume (β =
−0.80, SE = 0.07, p < 0.001), one potential concern is that the relationships among age, net-
work segregation, and language production were confounded with age-related differences in
gray matter volume. To address this concern, we re-ran all analyses including gray matter vol-
ume as a control variable. Results showed that the interaction between age and language net-
work segregation (β = −0.41, SE = 0.19, p = 0.03), and the interaction between age and whole
brain network segregation (β = −0.44, SE = 0.20, p = 0.03) on language production were both
still significant. Lastly, further analyses on each language production variable separately,
which is reported in full in the Supporting Information, showed that only picture naming speed
was marginally predicted by interactions between age and network measures (with language
network segregation, β = −28.89, SE = 15.47, p = 0.07; with whole brain network segregation,
β = −30.18, SE = 16.82, p = 0.08).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Age-related differences in the relationship between whole brain network segregation and language production composite scores.
(A) There was a significant positive relationship between whole brain network segregation and language production. (B) The Johnson-Neyman
test identified that the interaction was significant for ages 22–49.0 in our sample. (C) The significant main effect and interaction between whole
brain network segregation and language production was driven by adults younger than 49.0 years old. This relationship was not significant in
adults older than 49.0 years old.
Neurobiology of Language
394
Network segregation supports language production across adulthood
DISCUSSION
Older adults often exhibit declines in spoken language production such as slower speech rates
(Duchin & Mysak, 1987; Mortensen et al., 2006; Spieler & Griffin, 2006), increased word find-
ing difficulties, increased pauses and fillers in speech (Burke & Shafto, 2008; Horton et al.,
2010; MacKay & James, 2004), and more dysfluent and less grammatically complex speech
(e.g., Bortfeld et al., 2001; Kemper et al., 2003; Obler & Albert, 1981). Studies have suggested
that these age-related differences in language production may be related to declines in pho-
nological processing (Burke et al., 1991; Burke & Shafto, 2008), aspects of executive function
(Hasher et al., 1991; Hoffman et al., 2018; Lustig et al., 2007), or processing speed (Salthouse,
1996, 2000). Despite the abundant discussion regarding potential behavioral mechanisms, the
neural bases underlying these age-related differences are not entirely clear. Moreover, there is
general disagreement in the aging literature about how commonly observed age-related neural
differences should be interpreted, that is, as compensatory (Cabeza et al., 2018) or neural
dedifferentiation (Li et al., 2001). Specific to neural activities underlying language production,
some task-based fMRI studies have reported patterns supporting compensation (e.g., additional
brain activation associated with maintained behavioral performance; Nagels et al., 2012;
Wierenga et al., 2008) while others have found evidence for dedifferentiation (e.g., increased
brain activation related to weakened behavioral performance; Diaz et al., 2014; Meinzer
et al., 2009). In the current study, we focused on resting state functional connectivity and
investigated how the network characteristics in the language network and across the whole
brain relate to language production across adulthood. We specifically focused on a network
measure called network segregation, which incorporates both within-network connectivity
and between-network connectivity and highlights the degree to which networks are differen-
tiated. We hypothesized that language production performance, functional connectivity, and
their relationship would differ as a function of age.
As predicted, behaviorally, we found that older adults showed worse performance in lan-
guage production tasks. This is consistent with previous studies that have reported age-related
declines in production (for review, see Burke & Shafto, 2008; Diaz et al., 2016). This study,
however, enhances previous findings by demonstrating this main effect of age on a composite
score of language production from several different tasks, reflecting an overall profile of
language production in a broad sample of adults (ages 22–78 yr). Although we focused on
an integrated measure of language production ability based on multiple tasks, we also ana-
lyzed the effect of age on each production task performance separately (verbal fluency total
score, picture naming speed, MLU, MATTR; reported in Supporting Information). Briefly,
although all production measures showed the same decreasing trend with age, the effect of
age was only significant on verbal fluency total score. The age effect on verbal fluency has
been commonly observed (Gonzalez-Burgos et al., 2019; Rodríguez-Aranda & Martinussen,
2006; Troyer, 2000; Troyer et al., 1998). Compared to other measures, verbal fluency perfor-
mance relies on a broader set of abilities reflecting language production, vocabulary, speed, as
well as executive function. Therefore, the age-related difference in verbal fluency scores is
likely influenced by age-related differences in general cognitive functions as well as language.
In short, we found that older age was associated with worse overall language production,
which might be driven by the age effect on lexical access and executive function.
In addition to the behavioral results, our network analyses showed that while network seg-
regation within the left hemisphere language network was maintained in adults of all ages,
increased age was associated with lower whole brain network segregation. Moreover, we
examined how these network segregation results related to language production ability. For
both network segregation within the left hemisphere language network and whole brain
Neurobiology of Language
395
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
network segregation, there were significant interactions with age, such that increases in net-
work segregation were associated with better language production abilities, but only for youn-
ger and middle-aged adults. Interestingly, the whole brain network segregation contributed to
language production ability over a broader age range (up to age 49 yr) compared to the lan-
guage network segregation (up to age 29.2 yr). This could reflect the reliance of language pro-
duction on other cognitive abilities and could also be influenced by the stability of the left
hemisphere language network segregation in adulthood. Below, we examine these findings
in more detail.
First, consistent with previous studies of network segregation, older age was associated with
lower whole brain network segregation (Chan et al., 2014; Chan et al., 2018; Geerligs et al.,
2015; Petrican et al., 2017). Network segregation was calculated based on both within- and
between-network connectivities such that higher within-network connectivity and/or lower
between-network connectivity could lead to higher network segregation. On the whole brain
level, network segregation reflects the degree to which different networks in the brain share
connections among one another, thus, indicating a change in system specialization. As people
age, brain regions, especially those within the same networks, may show lower connectivity
strength (see Figure S1 in the Supporting Information). These age-related differences may reflect
a loss of functional specificity given that lower whole brain network segregation was associated
with worse language production performance in the current study, and lower network segrega-
tion has also been associated with lower memory function in previous studies (Chan et al.,
2014). Therefore, lower whole brain network segregation in older adults reflects a reduction
in overall functional specificity of network-based processes, supporting the dedifferentiation
account of aging (Ghisletta & Lindenberger, 2003). Despite significant age-related differences
in the whole brain network segregation, there was no significant main effect of age on language
network segregation. To our knowledge, no study has specifically looked at age differences in
the resting state language network using the integrated network segregation measurement. The
maintenance of the language network segregation across adulthood suggests that the functional
specificity of the language network and its interactions with other networks is relatively stable
across the lifespan. Although we focused on spoken language production in the current study,
the stability in the language network is, in fact, consistent with behavioral findings showing that
aspects of language comprehension, semantic processing, and vocabulary are well maintained
and even improve with age (e.g., Balota & Duchek, 1988; Burke & Peters, 1986; Cohen-Shikora
& Balota, 2016; Howard et al., 1981; Madden et al., 1993; Verhaeghen, 2003). These findings
suggest that, unlike other types of cognitive functions or neural structures that are vulnerable to
aging, many basic language functions, except speech, and the core neural structures related to
language are relatively well preserved regardless of increasing age.
Moreover, language production was associated with network segregation and this relation-
ship was modulated by age, even after controlling for the age-related differences in gray matter
volume. Specifically, only in relatively younger adults, higher network segregation in both the
language network and the whole brain network was associated with better language produc-
tion. These results indicate that more differentiated resting state networks may contribute to
enhanced language production ability in younger and middle-aged adults, consistent with pre-
vious studies using measures similar to network segregation (e.g., Duncan & Small, 2016). For
instance, although focusing on individuals with aphasia, Duncan and Small (2016) also found
that increased resting state network modularity (a measure similar to network segregation) was
positively associated with better performance on a narrative task.
Yet, this positive relationship between network segregation and language production was
not significant in neurotypical older adults. This result is consistent with previous studies that
Neurobiology of Language
396
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
have examined the relationship between age-related differences in language function and rest-
ing state networks. For example, Antonenko et al. (2013) found that younger adults showed
superior syntactic performance compared to older adults. Critically, only in younger, but not
older adults, stronger connectivity within the language network was associated with enhanced
behavioral performance. The lack of a brain–behavior relationship among older adults is also
consistent with other studies on language and aging that focused on task-based functional acti-
vation. For example, Diaz et al. (2014) reported that younger adults exhibited significant cor-
relations between fMRI activation and language function, which was not present in older
adults. These age-related differences in brain–behavior relationships in general, support the
dedifferentiation account of aging, which suggests that as people get older, their functional
networks become less structured and the relationships between functional measures and
behavior weaken (Ghisletta & Lindenberger, 2003).
At first glance, these findings supporting the dedifferentiation account seem to be inconsis-
tent with some previous language studies that reported results supporting compensatory neural
accounts of aging (Gertel et al., 2020; Pistono et al., 2021). In these studies, they found that
increased functional connectivities between language nodes and other networks were associ-
ated with better language function in older adults. However, it is worthwhile to mention that
these studies and the current study differ in several aspects. Foremost, the previous studies
incorporated an extreme-groups design that included two distinct age groups and used age
as a categorical variable, while the current study investigated the effects of age across adulthood
and treated age as a continuous variable. Critically, earlier studies focused on local relationships
between specific language nodes and domain-general regions, while the current study focused
on system-wide structures (i.e., between the language network and the rest of the brain and across
the whole brain). Specifically, rather than looking at a language network, Gertel et al. (2020)
took a more focused approach and included only one language region (left inferior frontal
gyrus) as the seed and found that the connectivity between this seed and the right hemisphere
executive function regions led to better Stroop performance in older adults. Although the
Stroop task involves language processing, it also requires a high degree of cognitive control.
So, the strong relationship between a language seed and executive function regions was
expected. Although Pistono et al. (2021) examined networks rather than individual regions, they
focused on the local relationships between two networks—the language network and the
multiple-demand network—and did not examine the rest of the networks in the brain. It could
be the case that reorganization of individual regions or select networks can serve a beneficial
function or reveal less age-related decline (i.e., the nonsignificant effect of age on language net-
work segregation), while whole brain analyses show an overall pattern of dedifferetiation in aging.
There is another interesting observation regarding the relationships among network segre-
gation, age, and language production. Specifically, the whole brain network segregation seems
to contribute to language production to a greater extent than the language network segrega-
tion, as reflected by the fact that the relationship between the network segregation and
production was significant through middle age whereas the relationship between language
network segregation and production was only significant for younger adults (49 yr old for
whole brain network segregation vs. 29.2 yr old for language network segregation). Recall that
there was no significant main effect of age on language network segregation, suggesting that
the core language network remains stable with age. Additionally, the main effect of language
network segregation on language production was not significant. At first glance, these results
may be surprising because one would expect that language function should largely rely on the
language network. However, language production and age-related differences in language are
multifaceted. For example, studies have shown that core language functions such as
Neurobiology of Language
397
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
comprehension or vocabulary are largely intact across adulthood (Madden et al., 1993;
Verhaeghen, 2003; Waters & Caplan, 2001). On the other hand, spoken language production
often declines with age. According to behavioral hypotheses such as the inhibition deficit theory
(Lustig et al., 2007) or the processing speed theory (Salthouse, 2000), age-related differences in
production may be in part because it relies on other domains such as visual and motor processing,
planning, monitoring, memory, and control, which suffer from age-related decline (for review, see
Burke & Shafto, 2008; Diaz et al., 2016). Although a more thorough investigation is needed, a
quick exploration of our data showed that enhanced production was associated with better
performance on the digit symbol task (β = 0.57, SE = 0.20, p = 0.005), suggesting that language
production covaries with domain general cognitive abilities. Therefore, while language com-
prehension may largely rely on the core language network (Wehbe et al., 2021), language
production relies on not only the language network but also many other networks that support
other cognitive functions (e.g., mouth somatomotor network for phonological production,
auditory network for hearing, ventral and dorsal attention networks, frontal-parietal control
networks). Finally, the age-related stability in the left hemisphere language network segrega-
tion may itself contribute to the weaker relationship between language network segregation
and language production ability. That is, if there is reduced variability in one measure and
higher variability in another, a significant relationship between the two will be less likely math-
ematically. The results of the present study suggest network segregation across a broader set of
networks may be required to support superior language production performance.
To summarize, language network segregation was stable across adulthood, highlighting the
neural stability of left hemisphere language regions and their interactions with other brain
regions. However, we found that increased age was associated with worse language produc-
tion performance and less segregated whole brain networks. Critically, whole brain network
segregation was associated with enhanced language production, but only among younger and
middle-aged adults. In contrast, older adults exhibited weaker relationships between network
segregation and language production. These results are consistent with the dedifferentiation
account of aging. Findings from the current study also indicate the contribution of the whole
brain network to production, implying that the age-related differences in language production
may be related to age-related differences in general cognitive abilities.
ACKNOWLEDGMENTS
This project was funded by a National Institutes of Health (NIH) National Institute on Aging
(NIA) grant. The writing of this paper was supported by the National Natural Science Founda-
tion of China and by the Start-up Research Grant (SRG2022–00003-ICI) and Multi-Year
Research Grant (MYRG2022–00148-ICI) from the University of Macau. We thank the staff
and scientists at the Social, Life, and Engineering Sciences Imaging Center (SLEIC) and the
Center for Language Science (CLS), where the study was conducted, for their support.
FUNDING INFORMATION
Michele T. Diaz, National Institute on Aging (https://dx.doi.org/10.13039/100000049), Award
ID: R01 AG034138. Haoyun Zhang, National Natural Science Foundation of China (https://dx
.doi.org/10.13039/501100001809), Award ID: 32200845.
AUTHOR CONTRIBUTIONS
Haoyun Zhang: Conceptualization, Formal analysis, Investigation, Methodology, Visualiza-
tion, Writing – original draft. Michele T. Diaz: Conceptualization, Funding acquisition,
Methodology, Project administration, Supervision, Writing – review & editing.
Neurobiology of Language
398
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
DATA AND CODE AVAILABILITY STATEMENTS
Data, analysis scripts, and supplementary materials are available at https://osf.io/3u9rw/?view
_only=4ba25920ed834408bda0fccd96f4cc2e.
REFERENCES
Acheson, D. J., Wells, J. B., & MacDonald, M. C. (2008). New and
updated tests of print exposure and reading abilities in college
students. Behavior Research Methods, 40(1), 278–289. https://
doi.org/10.3758/BRM.40.1.278, PubMed: 18411551
Antonenko, D., Brauer, J., Meinzer, M., Fengler, A., Kerti, L.,
Friederici, A. D., & Flöel, A. (2013). Functional and structural
syntax networks in aging. NeuroImage, 83, 513–523. https://doi
.org/10.1016/j.neuroimage.2013.07.018, PubMed: 23867559
Ashburner, J., & Friston, K. J. (2005). Unified segmentation. Neuro-
Image, 26(3), 839–851. https://doi.org/10.1016/j.neuroimage
.2005.02.018, PubMed: 15955494
Balota, D. A., & Duchek, J. M. (1988). Age-related differences in
lexical access, spreading activation, and simple pronunciation.
Psychology and Aging, 3(1), 84–93. https://doi.org/10.1037
/0882-7974.3.1.84, PubMed: 3268246
Bartzokis, G., Lu, P. H., Heydari, P., Couvrette, A., Lee, G. J.,
Kalashyan, G., Freeman, F., Grinstead, J. W., Villablanca, P.,
Finn, J. P., Mintz, J., Alger, J. R., & Altshuler, L. L. (2012). Multi-
modal magnetic resonance imaging assessment of white matter
aging trajectories over the lifespan of healthy individuals. Biolog-
ical Psychiatry, 72(12), 1026–1034. https://doi.org/10.1016/j
.biopsych.2012.07.010, PubMed: 23017471
Betzel, R. F., Byrge, L., He, Y., Goñi, J., Zuo, X.-N., & Sporns, O.
(2014). Changes in structural and functional connectivity among
resting-state networks across the human lifespan. NeuroImage,
102(Pt. 2), 345–357. https://doi.org/10.1016/j.neuroimage.2014
.07.067, PubMed: 25109530
Biswal, B., Zerrin Yetkin, F., Haughton, V. M., & Hyde, J. S. (1995).
Functional connectivity in the motor cortex of resting human
brain using echo-planar MRI. Magnetic Resonance in Medicine,
34(4), 537–541. https://doi.org/10.1002/mrm.1910340409,
PubMed: 8524021
Bortfeld, H., Leon, S. D., Bloom, J. E., Schober, M. F., & Brennan,
S. E. (2001). Disfluency rates in conversation: Effects of age, rela-
tionship, topic, role, and gender. Language and Speech, 44(2),
123–147. https://doi.org/10.1177/00238309010440020101,
PubMed: 11575901
Burke, D. M., MacKay, D. G., Worthley, J. S., & Wade, E. (1991).
On the tip of the tongue: What causes word finding failures in
young and older adults? Journal of Memory and Language,
30(5), 542–579. https://doi.org/10.1016/0749-596X(91)90026-G
Burke, D. M., & Peters, L. (1986). Word associations in old age:
Evidence for consistency in semantic encoding during adult-
hood. Psychology and Aging, 1(4), 283–292. https://doi.org/10
.1037/0882-7974.1.4.283, PubMed: 3267408
Burke, D. M., & Shafto, M. A. (2008). Language and aging. In F.
Craik & T. Salthouse (Eds.), The handbook of aging and cognition
(pp. 373–443). Psychology Press.
Cabeza, R., Albert, M., Belleville, S., Craik, F. I., Duarte, A., Grady,
C. L., Lindenberger, U., Nyberg, L., Park, D. C., Reuter-Lorenz,
P. A., Rugg, M. D., Steffener, J., & Rajah, M. N. (2018). Mainte-
nance, reserve and compensation: The cognitive neuroscience of
healthy ageing. Nature Reviews Neuroscience, 19(11), 701–710.
https://doi.org/10.1038/s41583-018-0068-2, PubMed: 30305711
Cabeza, R., & Dennis, N. A. (2013). Frontal lobes and aging: Dete-
rioration and compensation. In D. T. Stuss & R. T. Knight (Eds.),
Principles of frontal lobe function (pp. 628–652). Oxford Univer-
sity Press. https://doi.org/10.1093/med/9780199837755.003
.0044
Cao, M., Wang, J.-H., Dai, Z.-J., Cao, X.-Y., Jiang, L.-L., Fan, F.-M.,
Song, X.-W., Xia, M.-R., Shu, N., Dong, Q., Milham, M. P.,
Castellanos, F. X., Zuo, X.-N., & He, Y. (2014). Topological orga-
nization of the human brain functional connectome across the
lifespan. Developmental Cognitive Neuroscience, 7, 76–93.
https://doi.org/10.1016/j.dcn.2013.11.004, PubMed: 24333927
Chai, X. J., Castañón, A. N., Öngür, D., & Whitfield-Gabrieli, S.
(2012). Anticorrelations in resting state networks without global
signal regression. NeuroImage, 59(2), 1420–1428. https://doi.org
/10.1016/j.neuroimage.2011.08.048, PubMed: 21889994
Chan, M. Y., Na, J., Agres, P. F., Savalia, N. K., Park, D. C., & Wig,
G. S. (2018). Socioeconomic status moderates age-related differ-
ences in the brain’s functional network organization and anatomy
across the adult lifespan. Proceedings of the National Academy of
Sciences, 115(22), E5144–E5153. https://doi.org/10.1073/pnas
.1714021115, PubMed: 29760066
Chan, M. Y., Park, D. C., Savalia, N. K., Petersen, S. E., & Wig, G. S.
(2014). Decreased segregation of brain systems across the
healthy adult lifespan. Proceedings of the National Academy of
Sciences, 111(46), E4997–E5006. https://doi.org/10.1073/pnas
.1415122111, PubMed: 25368199
Christensen, K. J., Moye, J., Armson, R. R., & Kern, T. M. (1992).
Health screening and random recruitment for cognitive aging
research. Psychology and Aging, 7(2), 204–208. https://doi.org
/10.1037/0882-7974.7.2.204, PubMed: 1610509
Cohen-Shikora, E. R., & Balota, D. A. (2016). Visual word recognition
across the adult lifespan. Psychology and Aging, 31(5), 488–502.
https://doi.org/10.1037/pag0000100, PubMed: 27336629
Conway, A. R., Kane, M. J., Bunting, M. F., Hambrick, D. Z.,
Wilhelm, O., & Engle, R. W. (2005). Working memory span tasks:
A methodological review and user’s guide. Psychonomic Bulletin
& Review, 12(5), 769–786. https://doi.org/10.3758/BF03196772,
PubMed: 16523997
Cook, R. D. (1979). Influential observations in linear regression.
Journal of the American Statistical Association, 74(365), 169–174.
https://doi.org/10.1080/01621459.1979.10481634
Davey, C. E., Grayden, D. B., Egan, G. F., & Johnston, L. A. (2013).
Filtering induces correlation in fMRI resting state data. Neuro-
Image, 64, 728–740. https://doi.org/10.1016/j.neuroimage.2012
.08.022, PubMed: 22939874
Davis, S. W., Dennis, N. A., Daselaar, S. M., Fleck, M. S., &
Cabeza, R. (2008). Que PASA? The posterior–anterior shift in
aging. Cerebral Cortex, 18(5), 1201–1209. https://doi.org/10
.1093/cercor/bhm155, PubMed: 17925295
Diaz, M. T., Johnson, M. A., Burke, D. M., & Madden, D. J. (2014).
Age-related differences in the neural bases of phonological and
semantic processes. Journal of Cognitive Neuroscience, 26(12),
2798–2811. https://doi.org/10.1162/jocn_a_00665, PubMed:
24893737
Neurobiology of Language
399
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
Diaz, M. T., Karimi, H., Troutman, S. B. W., Gertel, V. H., Cosgrove,
A. L., & Zhang, H. (2021). Neural sensitivity to phonological
characteristics is stable across the lifespan. NeuroImage, 225,
Article 117511. https://doi.org/10.1016/j.neuroimage.2020
.117511, PubMed: 33129928
Diaz, M. T., Rizio, A. A., & Zhuang, J. (2016). The neural language
systems that support healthy aging: Integrating function, struc-
ture, and behavior. Language and Linguistics Compass, 10(7),
314–334. https://doi.org/10.1111/ lnc3.12199, PubMed:
28210287
Duchin, S. W., & Mysak, E. D. (1987). Disfluency and rate charac-
teristics of young adult, middle-aged, and older males. Journal of
Communication Disorders, 20(3), 245–257. https://doi.org/10
.1016/0021-9924(87)90022-0, PubMed: 3597824
Duncan, E. S., & Small, S. L. (2016). Increased modularity of resting
state networks supports improved narrative production in apha-
sia recovery. Brain Connectivity, 6(7), 524–529. https://doi.org/10
.1089/brain.2016.0437, PubMed: 27345466
Esarey, J., & Sumner, J. L. (2018). Marginal effects in interaction
models: Determining and controlling the false positive rate.
Comparative Political Studies, 51(9), 1144–1176. https://doi.org
/10.1177/0010414017730080
Fedorenko, E., Hsieh, P.-J., Nieto-Castañón, A., Whitfield-Gabrieli,
S., & Kanwisher, N. (2010). New method for fMRI investigations
of language: Defining ROIs functionally in individual subjects.
Journal of Neurophysiology, 104(2), 1177–1194. https://doi.org
/10.1152/jn.00032.2010, PubMed: 20410363
Ferré, P., Benhajali, Y., Steffener, J., Stern, Y., Joanette, Y., & Bellec,
P. (2019). Resting-state and vocabulary tasks distinctively inform
on age-related differences in the functional brain connectome.
Language, Cognition and Neuroscience, 34(8), 949–972.
https://doi.org/10.1080/23273798.2019.1608072, PubMed:
31457069
Folstein, M. F., Folstein, S. E., & McHugh, P. R. (1975). “Mini-
mental state”: A practical method for grading the cognitive state of
patients for the clinician. Journal of Psychiatric Research, 12(3),
189–198. https://doi.org/10.1016/0022-3956(75)90026-6,
PubMed: 1202204
Fox, J., & Weisberg, S. (2019). An R companion to applied regres-
sion (3rd ed.). Sage. https://socialsciences.mcmaster.ca/jfox
/Books/Companion/
Friston, K. J. (1994). Functional and effective connectivity in neuro-
imaging: A synthesis. Human Brain Mapping, 2(1–2), 56–78.
https://doi.org/10.1002/hbm.460020107
Geerligs, L., Renken, R. J., Saliasi, E., Maurits, N. M., & Lorist,
M. M. (2015). A brain-wide study of age-related changes in func-
tional connectivity. Cerebral Cortex, 25(7), 1987–1999. https://
doi.org/10.1093/cercor/bhu012, PubMed: 24532319
Gertel, V. H., Zhang, H., & Diaz, M. T. (2020). Stronger right
hemisphere functional connectivity supports executive aspects
of language in older adults. Brain and Language, 206, Article
104771. https://doi.org/10.1016/j.bandl.2020.104771, PubMed:
32289553
Ghisletta, P., & Lindenberger, U. (2003). Age-based structural
dynamics between perceptual speed and knowledge in the
Berlin Aging Study: Direct evidence for ability dedifferentiation
in old age. Psychology and Aging, 18(4), 696–713. https://doi
.org/10.1037/0882-7974.18.4.696, PubMed: 14692858
Glover, G. H., Mueller, B. A., Turner, J. A., van Erp, T. G. M., Liu,
T. T., Greve, D. N., Voyvodic, J. T., Rasmussen, J., Brown, G. G.,
Keator, D. B., Calhoun, V. D., Lee, H. J., Ford, J. M., Mathalon,
D. H., Diaz, M., O’Leary, D. S., Gadde, S., Preda, A., Lim, K. O.,
… Potkin, S. G. (2012). Function biomedical informatics research
network recommendations for prospective multicenter func-
tional MRI studies. Journal of Magnetic Resonance Imaging,
36(1), 39–54. https://doi.org/10.1002/jmri.23572, PubMed:
22314879
Gohel, S. R., & Biswal, B. B. (2015). Functional integration between
brain regions at rest occurs in multiple-frequency bands. Brain
Connectivity, 5(1), 23–34. https://doi.org/10.1089/ brain.2013
.0210, PubMed: 24702246
Gonzalez-Burgos, L., Hernández-Cabrera, J. A., Westman, E.,
Barroso, J., & Ferreira, D. (2019). Cognitive compensatory
mechanisms in normal aging: A study on verbal fluency and
the contribution of other cognitive functions. Aging, 11(12),
4090–4106. https://doi.org/10.18632/aging.102040, PubMed:
31232698
Grady, C. L., Luk, G., Craik, F. I., & Bialystok, E. (2015). Brain
network activity in monolingual and bilingual older adults.
Neuropsychologia, 66, 170–181. https://doi.org/10.1016/j
.neuropsychologia.2014.10.042, PubMed: 25445783
Guenther, F. H., Ghosh, S. S., & Tourville, J. A. (2006). Neural
modeling and imaging of the cortical interactions underlying
syllable production. Brain and Language, 96(3), 280–301.
https://doi.org/10.1016/j.bandl.2005.06.001, PubMed:
16040108
Guerin, J. M., Copersino, M. L., & Schretlen, D. J. (2018). Clinical
utility of the 15-item Geriatric Depression Scale (GDS-15) for use
with young and middle-aged adults. Journal of Affective Disor-
ders, 241, 59–62. https://doi.org/10.1016/j.jad.2018.07.038,
PubMed: 30096593
Hallquist, M. N., & Hillary, F. G. (2018). Graph theory approaches
to functional network organization in brain disorders: A
critique for a brave new small-world. Network Neuroscience,
3(1), 1–26. https://doi.org/10.1162/netn_a_00054, PubMed:
30793071
Hallquist, M. N., Hwang, K., & Luna, B. (2013). The nuisance of
nuisance regression: Spectral misspecification in a common
approach to resting-state fMRI preprocessing reintroduces noise
and obscures functional connectivity. NeuroImage, 82, 208–225.
https://doi.org/10.1016/j.neuroimage.2013.05.116, PubMed:
23747457
Harmelech, T., & Malach, R. (2013). Neurocognitive biases and the
patterns of spontaneous correlations in the human cortex. Trends
in Cognitive Sciences, 17(12), 606–615. https://doi.org/10.1016/j
.tics.2013.09.014, PubMed: 24182697
Hasher, L., Stoltzfus, E. R., Zacks, R. T., & Rypma, B. (1991).
Journal of Experimental Psychology:
Age and inhibition.
Learning, Memory, and Cognition, 17(1), 163–169. https://doi
.org/10.1037/0278-7393.17.1.163, PubMed: 1826730
Hoffman, P., Loginova, E., & Russell, A. (2018). Poor coherence in
older people’s speech is explained by impaired semantic and
executive processes. ELife, 7, Article e38907. https://doi.org/10
.7554/eLife.38907, PubMed: 30179156
Horton, W. S., Spieler, D. H., & Shriberg, E. (2010). A corpus anal-
ysis of patterns of age-related change in conversational speech.
Psychology and Aging, 25(3), 708–713. https://doi.org/10.1037
/a0019424, PubMed: 20677883
Howard, D. V., McAndrews, M. P., & Lasaga, M. I. (1981). Semantic
priming of lexical decisions in young and old adults. Journal of
Gerontology, 36(6), 707–714. https://doi.org/10.1093/geronj/36
.6.707, PubMed: 7299087
Hu, J., Small, H., Kean, H., Takahashi, A., Zekelman, L., Kleinman,
D., Ryan, E., Nieto-Castañón, A., Ferreira, V., & Fedorenko, E.
(2023). Precision fMRI reveals that
the language-selective
network supports both phrase-structure building and lexical
Neurobiology of Language
400
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
access during language production. Cerebral Cortex, 33(8),
4384–4404. https://doi.org/10.1093/cercor/bhac350, PubMed:
36130104
Huettel, S. A., Song, A. W., & McCarthy, G. (2004). Functional mag-
netic resonance imaging (1st ed.). Sinauer.
Imperati, D., Colcombe, S., Kelly, C., Di Martino, A., Zhou, J.,
Castellanos, F. X., & Milham, M. P. (2011). Differential develop-
ment of human brain white matter tracts. PLOS ONE, 6(8), Article
e23437. https://doi.org/10.1371/journal.pone.0023437,
PubMed: 21909351
Johnson, P. O., & Fay, L. C. (1950). The Johnson-Neyman technique,
its theory and application. Psychometrika, 15(4), 349–367.
https://doi.org/10.1007/BF02288864, PubMed: 14797902
Kassambara, A. (2020). ggpubr: ‘ggplot2’ based publication ready
plots ( Version 0.4.0) [Software]. https://CRAN.R-project.org
/package=ggpubr
Kemper, S., Herman, R. E., & Lian, C. H. (2003). The costs of doing
two things at once for young and older adults: Talking while
walking, finger tapping, and ignoring speech of noise. Psychol-
ogy and Aging, 18(2), 181–192. https://doi.org/10.1037/0882
-7974.18.2.181, PubMed: 12825768
King, B., van Ruitenbeek, P., Leunissen, I., Cuypers, K., Heise, K.-F.,
Santos Monteiro, T., Hermans, L., Levin, O., Albouy, G.,
Mantini, D., & Swinnen, S. P.
(2018). Age-related declines
in motor performance are associated with decreased segre-
gation of
large-scale resting state brain networks. Cerebral
Cortex, 28(12), 4390–4402. https://doi.org/10.1093/cercor
/bhx297, PubMed: 29136114
Krieger-Redwood, K., Wang, H.-T., Poerio, G., Martinon, L. M.,
Riby, L. M., Smallwood, J., & Jefferies, E. (2019). Reduced seman-
tic control in older adults is linked to intrinsic DMN connectivity.
Neuropsychologia, 132, Article 107133. https://doi.org/10.1016/j
.neuropsychologia.2019.107133, PubMed: 31278908
Li, S. C., Lindenberger, U., & Sikström, S. (2001). Aging cognition:
From neuromodulation to representation. Trends in Cognitive
Sciences, 5(11), 479–486. https://doi.org/10.1016/S1364
-6613(00)01769-1, PubMed: 11684480
Liu, T. T., Nalci, A., & Falahpour, M. (2017). The global signal in
fMRI: Nuisance or information? NeuroImage, 150, 213–229.
https://doi.org/10.1016/j.neuroimage.2017.02.036, PubMed:
28213118
Liu, X., Leopold, D. A., & Yang, Y. (2021). Single-neuron firing cas-
cades underlie global spontaneous brain events. Proceedings of
t h e N a t i o n a l A c a d e m y o f S c i e n c e s, 11 8 ( 4 7 ) , A r t i c l e
e2105395118. https://doi.org/10.1073/pnas.2105395118,
PubMed: 34795053
Long, J. A. (2019). interactions: Comprehensive, user-friendly toolkit
for probing interactions ( Version 1.1.0) [Software]. https://cran.r
-project.org/package=interactions
Lustig, C., Hasher, L., & Zacks, R. T. (2007). Inhibitory deficit the-
ory: Recent developments in a “new view.” In D. S. Gorfein &
C. M. MacLeod (Eds.), Inhibition in cognition (pp. 145–162).
American Psychological Association. https://doi.org/10.1037
/11587-008
MacKay, D. G., & James, L. E. (2004). Sequencing, speech pro-
duction, and selective effects of aging on phonological and
morphological speech errors. Psychology and Aging, 19(1),
93–107. https://doi.org/10.1037/0882-7974.19.1.93, PubMed:
15065934
MacLeod, C. M. (1991). Half a century of research on the Stroop
effect: An integrative review. Psychological Bulletin, 109(2),
163–203. https://doi.org/10.1037/0033-2909.109.2.163,
PubMed: 2034749
MacWhinney, B., & Wagner, J. (2010). Transcribing, searching and
data sharing: The CLAN software and the TalkBank data reposi-
tory. Gesprachsforschung: Online-Zeitschrift zur verbalen Inter-
aktion, 11, 154–173. PubMed: 25484851
Madden, D. J., Pierce, T. W., & Allen, P. A. (1993). Age-related
slowing and the time course of semantic priming in visual word
identification. Psychology and Aging, 8(4), 490–507. https://doi
.org/10.1037/0882-7974.8.4.490, PubMed: 8292278
Meinzer, M., Flaisch, T., Wilser, L., Eulitz, C., Rockstroh, B.,
Conway, T., Gonzalez-Rothi, L., & Crosson, B. (2009). Neural
signatures of semantic and phonemic fluency in young and
old adults.
Journal of Cognitive Neuroscience, 21(10),
2007–2018. https://doi.org/10.1162/jocn.2009.21219,
PubMed: 19296728
Miró-Padilla, A., Bueichekú, E., Ventura-Campos, N., Palomar-
García, M.-Á., & Ávila, C. (2017). Functional connectivity in
resting state as a phonemic fluency ability measure. Neuropsycho-
logia, 97, 98–103. https://doi.org/10.1016/j.neuropsychologia
.2017.02.009, PubMed: 28202336
Mortensen, L., Meyer, A. S., & Humphreys, G. W. (2006). Age-
related effects on speech production: A review. Language and
Cognitive Processes, 21(1–3), 238–290. https://doi.org/10.1080
/01690960444000278
Mwangi, B., Hasan, K. M., & Soares, J. C. (2013). Prediction of indi-
vidual subject’s age across the human lifespan using diffusion
tensor imaging: A machine learning approach. NeuroImage,
75, 58–67. https://doi.org/10.1016/j.neuroimage.2013.02.055,
PubMed: 23501046
Nagels, A., Kircher, T., Dietsche, B., Backes, H., Marquetand, J., &
Krug, A. (2012). Neural processing of overt word generation in
healthy individuals: The effect of age and word knowledge. Neu-
roImage, 61(4), 832–840. https://doi.org/10.1016/j.neuroimage
.2012.04.019, PubMed: 22521476
NITRC. (2015). Artifact Detection Tools (ART) [Software]. https://
www.nitrc.org/projects/artifact_detect/
Obler, L. K., & Albert, M. L. (1981). Language and aging: A neuro-
behavioral analysis. In D. S. Beasley & G. A. Davis (Eds.), Aging:
Communication processes and disorders (pp. 107–121). Grune &
Stratton.
Onoda, K., Ishihara, M., & Yamaguchi, S. (2012). Decreased func-
tional connectivity by aging is associated with cognitive decline.
Journal of Cognitive Neuroscience, 24(11), 2186–2198. https://
doi.org/10.1162/jocn_a_00269, PubMed: 22784277
Patterson, J. (2011). Controlled oral word association test. In J. S.
Kreutzer, J. DeLuca, & B. Caplan (Eds.), Encyclopedia of clinical
neuropsychology (pp. 703–706). Springer. https://doi.org/10
.1007/978-0-387-79948-3_876
Petrican, R., Taylor, M. J., & Grady, C. L. (2017). Trajectories of
brain system maturation from childhood to older adulthood:
Implications for lifespan cognitive functioning. NeuroImage,
163, 125–149. https://doi.org/10.1016/j.neuroimage.2017.09
.025, PubMed: 28917697
Pistono, A., Guerrier, L., Péran, P., Rafiq, M., Giméno, M., Bézy, C.,
Pariente, J., & Jucla, M. (2021). Increased functional connec-
tivity supports language performance in healthy aging despite
gray matter loss. Neurobiology of Aging, 98, 52–62. https://doi
.org/10.1016/j.neurobiolaging.2020.09.015, PubMed:
33246137
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes, K. A.,
Church, J. A., Vogel, A. C., Laumann, T. O., Miezin, F. M.,
Schlaggar, B. L., & Petersen, S. E. (2011). Functional network
organization of the human brain. Neuron, 72(4), 665–678. https://
doi.org/10.1016/j.neuron.2011.09.006, PubMed: 22099467
Neurobiology of Language
401
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
Raz, N., Lindenberger, U., Rodrigue, K. M., Kennedy, K. M., Head,
D., Williamson, A., Dahle, C., Gerstorf, D., & Acker, J. D. (2005).
Regional brain changes in aging healthy adults: General trends,
individual differences and modifiers. Cerebral Cortex, 15(11),
1676–1689. https://doi.org/10.1093/cercor/ bhi044, PubMed:
15703252
Reuter-Lorenz, P. A., & Cappell, K. A. (2008). Neurocognitive aging
and the compensation hypothesis. Current Directions in Psycho-
logical Science, 17(3), 177–182. https://doi.org/10.1111/j.1467
-8721.2008.00570.x
Rodríguez-Aranda, C., & Martinussen, M. (2006). Age-related dif-
ferences in performance of phonemic verbal fluency measured
by Controlled Oral Word Association Task (COWAT ): A
meta-analytic study. Developmental Neuropsychology, 30(2),
697–717. https://doi.org/10.1207/s15326942dn3002_3,
PubMed: 16995832
RStudio Team. (2022). RStudio: Integrated development environ-
ment for R. RStudio. https://www.rstudio.com/
Sala-Llonch, R., Bartrés-Faz, D., & Junqué, C. (2015). Reorganiza-
tion of brain networks in aging: A review of functional connec-
tivity studies. Frontiers in Psychology, 6, 663. https://doi.org/10
.3389/fpsyg.2015.00663, PubMed: 26052298
Salthouse, T. A. (1996). The processing-speed theory of adult age
differences in cognition. Psychological Review, 103(3),
403–428. https://doi.org/10.1037/0033-295X.103.3.403,
PubMed: 8759042
Salthouse, T. (2000). A theory of cognitive aging. North Holland.
Sheikh, J. I., & Yesavage, J. A. (1986). Geriatric Depression Scale
(GDS): Recent evidence and development of a shorter version.
Clinical Gerontologist: The Journal of Aging Mental Health,
5(1–2), 165–173. https://doi.org/10.1300/J018v05n01_09
Siman-Tov, T., Bosak, N., Sprecher, E., Paz, R., Eran, A., Aharon-
Peretz, J., & Kahn, I. (2017). Early age-related functional connec-
tivity decline in high-order cognitive networks. Frontiers in Aging
Neuroscience, 8, 330. https://doi.org/10.3389/fnagi.2016.00330,
PubMed: 28119599
Song, J., Birn, R. M., Boly, M., Meier, T. B., Nair, V. A., Meyerand,
M. E., & Prabhakaran, V. (2014). Age-related reorganizational
changes in modularity and functional connectivity of human
brain networks. Brain Connectivity, 4(9), 662–676. https://doi
.org/10.1089/brain.2014.0286, PubMed: 25183440
Sowell, E. R., Thompson, P. M., & Toga, A. W. (2004). Mapping
changes in the human cortex throughout the span of life. The
Neuroscientist, 10(4), 372–392. https://doi.org/10.1177
/1073858404263960, PubMed: 15271264
Spieler, D. H., & Griffin, Z. M. (2006). The influence of age on the
time course of word preparation in multiword utterances. Lan-
guage and Cognitive Processes, 21(1–3), 291–321. https://doi
.org/10.1080/01690960400002133
Stroop, J. R. (1935). Studies of interference in serial verbal reac-
tions. Journal of Experimental Psychology, 18(6), 643–662.
https://doi.org/10.1037/h0054651
Tomasi, D., & Volkow, N. D. (2012). Aging and functional brain
networks. Molecular Psychiatry, 17(5), 549–558. https://doi.org
/10.1038/mp.2011.81, PubMed: 21727896
Tremblay, P., Deschamps, I., & Gracco, V. L. (2016). Neurobiology
of speech production: A motor control perspective. In Neurobi-
ology of language (pp. 741–750). Elsevier. https://doi.org/10
.1016/B978-0-12-407794-2.00059-6
Troyer, A. K. (2000). Normative data for clustering and switching on
verbal fluency tasks. Journal of Clinical and Experimental Neuro-
psychology, 22(3), 370–378. https://doi.org/10.1076/1380
-3395(200006)22:3;1-V;FT370, PubMed: 10855044
Troyer, A. K., Moscovitch, M., Winocur, G., Alexander, M. P., &
Stuss, D. (1998). Clustering and switching on verbal fluency:
The effects of focal frontal-and temporal-lobe lesions. Neuropsy-
chologia, 36(6), 499–504. https://doi.org/10.1016/S0028
-3932(97)00152-8, PubMed: 9705059
Varangis, E., Habeck, C., Razlighi, Q., & Stern, Y. (2019). The effect
of aging on resting state connectivity of predefined networks in
the brain. Frontiers in Aging Neuroscience, 11, 234. https://doi
.org/10.3389/fnagi.2019.00234, PubMed: 31555124
Verhaeghen, P. (2003). Aging and vocabulary score: A meta-
analysis. Psychology and Aging, 18(2), 332–339. https://doi.org
/10.1037/0882-7974.18.2.332, PubMed: 12825780
Wang, L., LaViolette, P., O’Keefe, K., Putcha, D., Bakkour, A., Van
Dijk, K. R. A., Pihlajamäki, M., Dickerson, B. C., & Sperling, R. A.
(2010). Intrinsic connectivity between the hippocampus and
posteromedial cortex predicts memory performance in cogni-
tively intact older individuals. NeuroImage, 51(2), 910–917.
https://doi.org/10.1016/j.neuroimage.2010.02.046, PubMed:
20188183
Waters, G. S., & Caplan, D. (2001). Age, working memory, and
on-line syntactic processing in sentence comprehension. Psy-
chology and Aging, 16(1), 128–144. https://doi.org/10.1037
/0882-7974.16.1.128, PubMed: 11302362
Wawrzyniak, M., Hoffstaedter, F., Klingbeil, J., Stockert, A., Wrede,
K., Hartwigsen, G., Eickhoff, S. B., Classen, J., & Saur, D. (2017).
Fronto-temporal interactions are functionally relevant for seman-
tic control in language processing. PLOS ONE, 12(5), Article
e0177753. https://doi.org/10.1371/journal.pone.0177753,
PubMed: 28505211
Wechsler, D.
Intelligence
(1997). WAIS-III: Wechsler Adult
Scale. Psychological Corporation. https://doi.org/10.1037/t49755
-000
Wehbe, L., Blank, I. A., Shain, C., Futrell, R., Levy, R., von der
Malsburg, T., Smith, N., Gibson, E., & Fedorenko, E. (2021).
Incremental language comprehension difficulty predicts activity
in the language network but not the multiple demand network.
Cerebral Cortex, 31(9), 4006–4023. https://doi.org/10.1093
/cercor/bhab065, PubMed: 33895807
Whitfield-Gabrieli, S., & Nieto-Castanon, A. (2012). CONN: A
functional connectivity toolbox for correlated and anticorrelated
brain networks. Brain Connectivity, 2(3), 125–141. https://doi.org
/10.1089/brain.2012.0073, PubMed: 22642651
Wickham, H. (2016). ggplot2: Elegant graphics for data analysis.
Springer-Verlag. https://doi.org/10.1007/978-3-319-24277-4
Wickham, H., Averick, M., Bryan, J., Chang, W., D’Agostino
McGowan, L., François, R., Grolemund, G., Hayes, A., Henry,
L., Hester, J., Kuhn, M., Pedersen, T. L., Miller, E., Bache,
S. M., Müller, K., Ooms, J., Robinson, D., Seidel, D. P., Spinu,
V., … Yutani, H. (2019). Welcome to the Tidyverse. Journal of
Open Source Software, 4(43), 1686. https://doi.org/10.21105
/joss.01686
Wickham, H., & Bryan, J. (2019). readxl: Read Excel files ( Version
1.3) [Software]. https://cran.r-project.org/package=readxl
Wierenga, C. E., Benjamin, M., Gopinath, K., Perlstein, W. M.,
Leonard, C. M., Rothi, L. J. G., Conway, T., Cato, M. A., Briggs,
R., & Crosson, B. (2008). Age-related changes in word retrieval:
Role of bilateral frontal and subcortical networks. Neurobiology
o f A g i n g , 2 9 ( 3 ) , 4 3 6 – 4 5 1 . h t t p s : / / d o i . o rg / 1 0 . 1 0 1 6 / j
.neurobiolaging.2006.10.024, PubMed: 17147975
Woods, S. P., Delis, D. C., Scott, J. C., Kramer, J. H., & Holdnack,
J. A. (2006). The California Verbal Learning Test–second edition:
Test–retest reliability, practice effects, and reliable change indi-
ces for the standard and alternate forms. Archives of Clinical
Neurobiology of Language
402
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network segregation supports language production across adulthood
Neuropsychology, 21(5), 413–420. https://doi.org/10.1016/j.acn
.2006.06.002, PubMed: 16843636
Zhang, H., Bai, X., & Diaz, M. T. (2021). The intensity and connectivity
of spontaneous brain activity in a language network relate to aging
and language. Neuropsychologia, 154, Article 107784. https://doi
.org/10.1016/j.neuropsychologia.2021.107784, PubMed: 33571489
Zhang, H., Eppes, A., & Diaz, M. T. (2019). Task difficulty modu-
lates age-related differences in the behavioral and neural bases of
language production. Neuropsychologia, 124, 254–273. https://
doi.org/10.1016/j.neuropsychologia.2018.11.017, PubMed:
30513288
Zhang, H., Gertel, V. H., Cosgrove, A. L., & Diaz, M. T. (2021).
Age-related differences in resting-state and task-based network
characteristics and cognition: A lifespan sample. Neurobiology
o f A g i n g , 1 0 1 , 2 6 2 – 2 7 2 . h t t p s : / / d o i . o r g / 1 0 . 1 0 1 6 / j
.neurobiolaging.2020.10.025, PubMed: 33602583
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
3
8
2
2
1
3
1
6
0
3
n
o
_
a
_
0
0
1
0
6
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
403