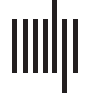ARTICLE DE RECHERCHE
Sources of Heterogeneity in Functional
Connectivity During English Word Processing
in Bilingual and Monolingual Children
un accès ouvert
journal
Nia Nickerson1
, Valeria Caruso1, Tai-Li Chou4
, Adriene M. Beltz1
James R. Booth5
, Xiao-Su Hu1
, and Ioulia Kovelman1
Xin Sun1,2
, Rebecca A. Marks3
, Rachel L. Eggleston1, Kehui Zhang1
, Chi-Lin Yu1
,
, Twila Tardif1
,
1Département de psychologie, Université du Michigan, Ann-Arbor, MI, Etats-Unis
2Département de psychologie, University of British Columbia, Vancouver, Canada
3Department of Brain and Cognitive Sciences, Massachusetts Institute of Technology, Cambridge, MA, Etats-Unis
4Département de psychologie, National Taiwan University, Taipei, Taiwan
5Department of Psychology and Human Development, Vanderbilt University, Nashville, TN, Etats-Unis
Mots clés: brain development, functional connectivity, bilingualism, enfants, fNIRS, individual
differences
ABSTRAIT
Diversity and variation in language experiences, such as bilingualism, contribute to
heterogeneity in children’s neural organization for language and brain development. À
uncover sources of such heterogeneity in children’s neural language networks, the present
study examined the effects of bilingual proficiency on children’s neural organization for
language function. To do so, we took an innovative person-specific analytical approach
to investigate young Chinese-English and Spanish-English bilingual learners of structurally
distinct languages. Bilingual and English monolingual children (N = 152, M.(SD)age =
7.71(1.32)) completed an English word recognition task during functional near-infrared
spectroscopy neuroimaging, along with language and literacy tasks in each of their languages.
Two key findings emerged. D'abord, bilinguals’ heritage language proficiency (Chinese or Spanish)
made a unique contribution to children’s language network density. Deuxième, the findings
reveal common and unique patterns in children’s patterns of task-related functional
connectivité. Common across all participants were short-distance neural connections within
left hemisphere regions associated with semantic processes (within middle temporal and
frontal regions). Unique to more proficient language users were additional long-distance
connections between frontal, temporal, and bilateral regions within the broader language
réseau. The study informs neurodevelopmental theories of language by revealing the effects
of heterogeneity in language proficiency and experiences on the structure and quality of
emerging language neural networks in linguistically diverse learners.
INTRODUCTION
Early language experiences shape a child’s mind and brain while also laying foundations for
reading (Werker & Hensch, 2015). Bilingualism offers enriched linguistic experiences that add
to the heterogeneity in children’s neural organization for language and reading acquisition
(Hernandez et al., 2019). To better capture the developing neural heterogeneity for language
traitement, the present study utilized a person-specific network mapping approach to
Citation: Sun, X., Marks, R.. UN.,
Eggleston, R.. L., Zhang, K., Yu, C.-L.,
Nickerson, N., Caruso, V., Chou, T.-L.,
Hu, X.-S., Tardif, T., Booth, J.. R., Beltz,
UN. M., & Kovelman, je. (2023). Sources
of heterogeneity in functional
connectivity during English word
processing in bilingual and
monolingual children. Neurobiology of
Language, 4(2), 198–220. https://doi.org
/10.1162/nol_a_00092
EST CE QUE JE:
https://doi.org/10.1162/nol_a_00092
Informations complémentaires:
https://doi.org/10.1162/nol_a_00092
Reçu: 1 Juin 2022
Accepté: 10 Novembre 2022
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Xin Sun
sunxin@umich.edu
Éditeur de manipulation:
Marcela Peña Garay
droits d'auteur: © 2023
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
un
_
0
0
0
9
2
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
characterize sources of heterogeneity in children’s emerging neural pathways for English and
to identify how these language neural networks are influenced by early bilingual experiences
with Spanish or Chinese.
According to the Neuroemergentist Framework, complex neurocognitive processes
develop out of interactions between an individual’s expertise and the environmental ecosys-
tem (Claussenius-Kalman et al., 2021). Bilingual development thus stems from dynamic com-
munications between individuals’ developing cognitive skills (par exemple., attention and working
mémoire) as well as their bilingual experiences and contexts (par exemple., linguistic and orthographic
features and contexts of use). Par conséquent, bilinguals may form different patterns of neural orga-
nization for language processing in relation to monolinguals and/or bilinguals of different lan-
guage groups. Par exemple, a meta-analysis found that bilinguals speaking a language with
more predictable sound-to-print mapping (par exemple., French, as compared to English) rely on
enhanced phonological networks (c'est à dire., bilateral temporal regions). In contrast, those speaking
a language with less predictable associations between language sounds and printed form (par exemple.,
Chinese, as compared to English) rely on enhanced networks for lexical integrations (c'est à dire., gauche
middle/inferior frontal regions; Liu & Cao, 2016). De plus, connectivity studies have found
que, due to the increased cognitive demands of bilingual coordination, compared with mono-
linguals, bilinguals form enhanced connectivity between bilateral inferior frontal regions, comme
well as between the basal ganglia and the frontal cortex, which guides language perception,
comprehension, and cognitive control (Berken et al., 2016; Marian et al., 2017).
Most prior research has approached bilingual brain development with group averages.
Cependant, variations in bilingual experiences may yield meaningful variability in the neural
networks within groups. To advance the understanding of heterogeneity in neural mechanisms
of spoken language processing, and how they are influenced by bilingualism, we examined
the functional connectivity of cortical networks for spoken word recognition. Prior work has
shown that children’s language and reading proficiency are positively associated with
strengthened neural connectivity along key neural pathways of language processing (Skeide
et coll., 2016; Yeatman et al., 2011). De plus, these neural networks develop as a function of
language experience, including bilingualism (Ip et al., 2017; Kovelman et al., 2008; Marian
et coll., 2017). Autrement dit, bilingual experiences contribute to the neural network heteroge-
neity of language development (Claussenius-Kalman et al., 2021; Hernandez et al., 2019).
Using a person-specific approach, we aimed to uncover sources of individual variation in
the development of neural networks that support language and literacy development.
The Developing Neural Basis for Spoken Word Processing
Spoken words are comprised of sound (phonological units) and meaning (semantic units). Pro-
ficient adult speakers typically engage two parallel processing streams that allow them to
simultaneously consider the multifaceted nature of phonological and lexico-semantic repre-
sentations during word recognition. In the adult brain, these are commonly represented as
dorsal and ventral neural streams (Hickok, 2022; Hickok & Poeppel, 2007). The dorsal or pho-
nological stream includes the dorsal aspect of the left inferior frontal gyrus (IFG) and superior
temporal gyrus (STG), as well as the arcuate fasciculus (AF) fiber tract that connects those
régions. The ventral or semantic stream includes the ventral aspect of the left IFG, the middle
temporal gyrus (MTG), and the inferior fronto-occipital fasciculus that connects them (Su et al.,
2018). These two parallel processing streams improve in their functionality over the course of
children’s language development, as children learn to efficiently access both lexical and sub-
lexical information. Children’s spoken language skills are linked to functional and anatomical
Neurobiology of Language
199
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
un
_
0
0
0
9
2
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
strengths within and between these networks (Cao et al., 2008; Friederici et al., 2011; Skeide
et coll., 2016; Yeatman et al., 2011; Yu et al., 2018).
Functional connectivity studies reveal how brain regions work together during a language
task and how these brain connections relate to developmental outcomes in language profi-
ciency (Friederici et al., 2011; Jasińska et al., 2020; Qi et al., 2021; Xiao et al., 2016; Yu
et coll., 2018, 2021). This research generally suggests a gradual shift in the development of
inter- (entre) and then intra- (within) hemisphere associations. Par exemple, Friederici
et autres. (2011) examined functional connectivity in 6-year-old children and adults who were
performing an auditory sentence comprehension task during functional magnetic resonance
imaging (IRMf). Findings revealed that younger children formed stronger functional connec-
tions between the left frontal regions and their right hemisphere homologs than adults. In con-
trast, adults showed stronger connectivity between the left frontotemporal regions. This and
similar findings (Enge et al., 2020; Weiss-Croft & Baldeweg, 2015) exemplify the merits of
functional connectivity research in revealing changes in language development, paving the
way for more nuanced inquiries into sources of heterogeneity of such change.
Connecting Spoken Language Networks to Reading
Learning to read requires children to connect their understanding of spoken words to orthog-
raphie, or written symbols. Donc, neural networks for spoken language are essential for
children’s behavioral outcomes such as emergent literacy (Jasińska et al., 2020; Yu et al.,
2018, 2021). Par exemple, Jasińska et al. (2020) examined the longitudinal effects of functional
connectivity in 4-year-old children who passively listened to words during functional near-
infrared spectroscopy (fNIRS). Findings revealed that children who exhibited stronger func-
tional connectivity during this auditory task between the left IFG and right STG regions at
âge 4 years had better reading proficiency a year later. Building upon this and similar prior
findings (Qi et al., 2021), we aimed to advance beyond the traditional functional correlation
methods that average across diverse speakers. Here we estimate individualized neural net-
works to better capture sources of heterogeneity in children’s emerging neural architecture
for language and how the neural networks speak to children’s developing language and liter-
acy skills (Arredondo et al., 2022; Beltz et al., 2016).
Individual Differences in the Neural Connectivity for Language in Bilingual Children
Neuroimaging research on bilingualism often finds connectivity differences between bilingual
and monolingual populations. These examinations include both anatomical connectivity as
studied through white matter tracts (García-Pentón et al., 2014; Mohades et al., 2012) et
resting-state functional connectivity (Berken et al., 2016; Sun et al., 2019; Thieba et al.,
2019). Par exemple, in an anatomical diffusion tensor imaging study, Gao et al. (2022) exam-
ined the relation between bilingual proficiency and white matter tracts in Chinese-English
bilingual children raised in China. Findings revealed that children with thicker AF tracts
around left STG regions had better word reading proficiency in both English and Chinese.
The AF is a tract that connects frontal and temporal language regions and generally increases
in its thickness over the course of language development (Skeide & Friederici, 2016). Tel
neuroanatomical findings support the idea that there is a relation between bilingualism factors
and neural connections critical for language processing (Bialystok et al., 2012).
Resting-state connectivity studies ask participants to stay awake while they are not engaged
in any given task to reveal a presumed default state of brain operations. A resting-state fMRI
study found that adults with early bilingual exposure (before age 5) showed stronger intrinsic
Neurobiology of Language
200
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
un
_
0
0
0
9
2
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
functional connectivity between the left and right IFG regions and between left IFG and pre-
frontal regions than later-exposed bilinguals (Berken et al., 2016). The findings suggest that
early bilingual exposure influences the neural organization of the frontal lobe network essen-
tial for language control (Berken et al., 2016; Bialystok et al., 2012). The advantage of resting-
state neuroimaging studies is that they can capture spontaneous signals that do not tie to a
specific mental state (c'est à dire., a task). Nevertheless, non-task resting-state paradigms may lack
empirical benefits such as sensitivity to brain-behavior associations (Finn, 2021). To the best
of our knowledge, no prior study has examined bilingual children’s functional connectivity
networks while participants engage in a language task, which is a knowledge gap we aim
to fill in the present work.
Another important but understudied issue is how to best depict the neural networks of
language processing for bilingual children. Much current knowledge about bilingualism
stems from analytical approaches that have dichotomized bilinguals versus monolinguals or
otherwise categorized different groups of bilinguals, such as splitting by age of exposure or
proficiency (par exemple., Liu & Cao, 2016; Sulpizio et al., 2020). Cependant, bilinguals can differ in
many ways. Newly emerging research thus advocates for approaches that leverage the hetero-
geneity of bilingual profiles to better understand bilingualism (Luk & Bialystok, 2013; Marian &
Hayakawa, 2021). The present work thus adopts a person-specific approach to examine
such heterogeneity of functional connectivity for language in relation to children’s bilingual
language and reading development.
Examining Person-Specific Neural Network With GIMME
Person-specific analytical approaches, such as group iterative multiple model estimation
(GIMME; Gates & Molenaar, 2012), advance upon conventional data analysis methods that
average across heterogenous individuals by instead identifying connections among a priori
regions of interest (ROI) that are shared across participants (group level), across a subgroup
of participants (subgroup level), as well as connections that are unique to one or some individ-
uals (individual level). In this way, GIMME networks capture both the broad homogeneity of the
group and the heterogeneity of individuals. Spécifiquement, GIMME uses a data-driven approach to
yield person-specific directed connectivity maps; GIMME begins with a null network and then
adds connections among ROIs that are meaningful (c'est à dire., significant) for at least 75% of partic-
ipants to all participants’ networks followed by adding connections that are meaningful for a
subgroup of participants. Enfin, GIMME adds connections that are meaningful just to an indi-
vidual. Connections are added until each person’s network represents their observed data well
and has person-specific weights. Simulation studies suggest that GIMME shows exceptional
robustness in modeling heterogeneous data compared to nearly 40 alternative methods, et
it has been applied to a wide range of psychological studies with neuroimaging data (Dotterer
et coll., 2020; Gates & Molenaar, 2012; Goetschius et al., 2020; Lane et al., 2019; Price et al.,
2020). Altogether, as a data-driven network mapping approach, GIMME addresses the limita-
tions of traditional group-oriented approaches that rely on averages while also allowing for both
group-level inferences and accurate reflections of individual-level heterogeneity.
GIMME has been used to examine the functional connectivity of the attention networks in bilin-
gual children. Arredondo et al. (2022) used GIMME to estimate bilingual and monolingual chil-
dren’s neural connectivity with the attention network task among six pre-specified left superior,
middle frontal, as well as parietal brain channels with fNIRS. GIMME identified two subgroups,
one that consisted of almost all monolinguals (92%) and half of the bilinguals (54%), and another
that consisted of a small portion of monolinguals (8%) and the other half of bilinguals (46%).
GIMME:
Group iterative multiple model
estimation; a network-mapping
method that identifies connections
of variables (c'est à dire., brain signals)
collected among multiple time points
pour chaque individu.
Neurobiology of Language
201
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
un
_
0
0
0
9
2
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
Heritage language:
Language learned by its speakers at
home as children, while they are
often exposed to a different language
outside home environments.
Notably, the bilinguals in the first group were more English-dominant (c'est à dire., “monolingual-like”),
whereas the bilinguals in the second group had more balanced proficiency across their two lan-
guages. Surtout, the second group also had significantly higher network density (c'est à dire., number
of connections) centered around the left frontal regions compared to the first group, which also
corresponded to higher attention task accuracy. These results suggest more complex attentional
neural networks for early bilingual children with more balanced dual language proficiency. Dans
sum, GIMME has been shown to be an effective approach for understanding sources of heteroge-
neity in the neural organization of cognitive functions in bilingual children, but many questions
remain unanswered, particularly regarding neural networks during a language task.
The Present Study
The current study employed GIMME analysis of fNIRS data to examine the effects of early and
systematic bilingual experiences on children’s emerging neural architecture for language pro-
cesses and their relation to literacy development. The participant groups included children
(ages 5–10 years) who were English monolinguals, Chinese-English bilinguals, or Spanish-
English bilinguals, all experiencing English-dominant education in the US. The bilinguals were
exposed to a heritage language (Chinese or Spanish) at home from birth, to English around age
2, and were capable of reading words/characters in their heritage languages. The study spe-
cifically asked participants to complete an auditory word-processing task during fNIRS neuro-
imaging. Children heard three words and were asked to identify the two words that shared a
unit of meaning (morpheme) while ignoring a phonological distractor (par exemple., bedroom, class-
room, mushroom). The task probed children’s ability to analyze words’ lexico-semantic and
phonological constituents necessary for successful word processing. The ability to operate
upon words’ sound and meaning units is thought to support children’s emergent literacy
(Kuo & Anderson, 2006; Sun, Zhang, Marks, Nickerson, et coll., 2022).
Functional connectivity analyses were performed with a priori brain regions of language
traitement, including bilateral frontal and left temporal areas. These regions have been iden-
tified as essential to spoken word recognition by previous research (Enge et al., 2020; Friederici
et coll., 2011; Jasińska et al., 2020) as well as for the current sample (see Sun et al., 2023, for the
functional activation patterns). We used GIMME to ask two experimental questions. D'abord, nous
asked: What is the relation between individual differences in functional connectivity for word
processing and children’s literacy skills in English? To answer this question, we applied
GIMME to identify potentially different groups of learners. We then examined the relationship
between children’s English proficiency and their network characteristics, focusing on network
density within the identified language regions, which is thought to reflect the quality of the
language network (Jasińska et al., 2020). Deuxième, we asked: How do bilinguals’ heritage lan-
guage skills contribute to the neural network quality of English word processing? We predicted
significant associations between children’s neural networks and behavioral profiles, and there-
fore examined the brain-behavioral associations between children’s connectivity network pat-
terns and their proficiency in English and their heritage language. Ensemble, the goal of the
study was to inform our understanding of the effects of bilingualism and sources of heteroge-
neity in children’s emergent language networks.
MATERIALS AND METHODS
Participants
Participants were 152 enfants (75 girls, Mage = 7.71 années, SDage = 1.32, age range = 5.12–
10.19) recruited from southeast Michigan, Etats-Unis. Participants were all typically developing
Neurobiology of Language
202
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
un
_
0
0
0
9
2
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
without a history of developmental delays in language or literacy, deficits in hearing, ou autre
neurological or physical disorders. All children grew up in the United States, attended English-
only schools, and were proficient English users, as determined by standard vocabulary scores
over 85 on the Peabody Picture Vocabulary Test 5 (PPVT-5; Dunn, 2019). All three groups
were matched on age, genre, grade distribution, maternal education, and non-verbal working
mémoire (see Table 1). Parents and children provided appropriate informed consent or assent
and received $40 for their participation. The study was approved by the Institutional Review
Board for research with human subjects.
Participants had diverse language experiences: 35.5% were English monolinguals (N = 54),
while the remaining 64.5% were either bilingual English-Chinese (N = 48) or English-Spanish
(N = 50) speakers. According to the parental reports, the bilinguals were exposed to their
heritage language (Chinese or Spanish) from birth at home and with at least one parent
Demographics
Age
Gradea
Tableau 1. Demographics and English task performance by participant group
English monolingual N = 54
M.(SD) or n
Spanish bilingual N = 50
M.(SD) or n
Chinese bilingual N = 48
M.(SD) or n
7.66 (1.32)
7.84 (1.22)
7.63 (1.44)
K
1
2
3
4
14
9
16
8
7
6
18
7
12
5
16
9
7
11
7
Working Memoryb
Maternal Educationc
7.15 (2.43)
88.9%
7.36 (2.15)
84.0%
7.76 (2.64)
95.8%
English task performance
Vocabulary
158.40 (26.84)
144.30 (28.14)
145.09 (32.95)
Phonological awareness
Morphological awareness
Word reading
Reading comprehension
Sentence reading fluency
21.42 (7.79)
25.31 (11.32)
46.75 (16.79)
26.12 (9.18)
37.55 (17.68)
23.62 (7.23)
24.42 (10.01)
48.54 (15.27)
24.64 (6.88)
34.78 (18.49)
22.63 (7.48)
24.54 (11.18)
50.47 (14.36)
27.22 (8.35)
40.71 (21.39)
p
0.921
0.051
0.443
–
0.022
0.401
0.712
0.234
0.540
0.335
fNIRS task accuracy (%)
79.6 (9.3)
77.2 (10.2)
81.1 (9.8)
0.336
a Grade distribution used a χ2 test with df = 8.
b Measured by a backward digit span task (Wechsler, 2014).
c % bachelor’s degree or above.
Neurobiology of Language
203
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
un
_
0
0
0
9
2
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
considering themselves to be a native speaker of the language. Bilingual children were also
systematically exposed to English before or beginning at age two (c'est à dire., used English regularly in
contexts such as daycare or preschool). Heritage language vocabulary was used to identify
children’s heritage language proficiency. Of note is that although they provide some informa-
tion about language proficiency, both standard scores of Chinese and Spanish should be inter-
preted cautiously, as the norm of the Chinese vocabulary task was based on children growing
up in Taiwan in 1988 (PPVT–Revised; Dunn & Dunn, 1998), and the Spanish norm was based
on children growing up in Mexico and Puerto Rico in 1986 (Test de Vocabulario en Imágenes
Peabody [TVIP]; Dunn et al., 1986). To account for the limitation from the norm and to capture
variations in the bilingual heritage speakers, no participants were excluded on account of low
heritage language vocabulary. Néanmoins, all Spanish bilingual participants had a Spanish
receptive vocabulary standard score above 70, et 93% of Chinese bilingual participants
passed this threshold in Chinese receptive vocabulary.
Measures and Procedure
All participants completed the full battery of behavioral and neuroimaging tests during a
single laboratory visit. Participants completed language and literacy measurements in each
of their languages including vocabulary, phonological awareness, word reading, reading
comprehension and fluency, and morphological awareness. Across languages, these tasks
were maximally matched by either using similar standardized assessments that are already
available (par exemple., vocabulary across languages) or building measures that were maximally similar
across language assessments (par exemple., an experimental elision task in Chinese to match the Span-
ish and English versions). In selecting language measures, we took into account the need to
make these measures maximally comparable and the fact that the measures need to capture
specific features of each language. We therefore acknowledge that the tests are maximally
comparable in capturing respective skills, but not identical across languages. All self-
developed tasks are openly available and can be found in Sun, Zhang, Marks, Karas, et autres.
(2022). For the current study, data and codes can be found at https://osf.io/uv3t6/?view_only
=46569a15ebd241808a01d51f550c65dd.
Vocabulary
Vocabulary was tested with the Peabody Picture Vocabulary Test in English (PPVT-5; Dunn,
2019); in Chinese (PPVT-Revised; Dunn & Dunn, 1998); and in Spanish (TVIP; Dunn et al.,
1986). Children saw four pictures, heard a word, and selected the picture that best describes
the word.
Phonological awareness
Phonological awareness was measured with a sound elision task in which children heard a
word and were asked to omit a phonetic unit from the word (par exemple., “Cat without /k/ is ___.”
[à]). The English task used the Elision subtest from the Comprehensive Test of Phonological
Processing (Wagner et al., 1999), the Spanish task used the Test of Phonological Processing in
Spanish (Francis et al., 2001), and the Chinese task was adapted from Newman et al.’s (2011)
measure with the same paradigm.
Morphological awareness
For this task, we aimed to tap into lexical morphological awareness across languages and cap-
ture morphological features of each language (c'est à dire., compound structures in Chinese and both
compound and derivational structures in English/Spanish). In English, we used the Early
Neurobiology of Language
204
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
un
_
0
0
0
9
2
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
Lexical Morphology Measure (Marks, Labotka, et coll., 2022), which includes compound and
derivational words. Children were asked to complete a sentence with part of a given word
(par exemple., “Football. Ouch! You stepped on my ____.” [foot]; “Friendly. She is my best ___.”
[friend]). A parallel task was used in Spanish (Marks, Sun, et coll., 2022). In Chinese, a morpho-
logical construction measure was used (modified from Song et al., 2015). Children were asked
to create a new word with a given word, Par exemple, “Apple trees grow apples. What trees
might grow bread? [bread trees].»
Word/character reading
Word/Character reading was measured by presenting a list of words/characters and asking
children to read them aloud. The English task was the Letter-Word Identification subtest from
Woodcock-Johnson IV (Schrank et al., 2014); the Spanish task was the Word Identification
subtest from Batería III Woodcock-Muñoz (Muñoz-Sandoval et al., 2005); and the Chinese task
was a self-developed measure (Sun, Zhang, Marks, Karas, et coll., 2022).
Sentence reading fluency
Sentence reading fluency was measured using a 3-min timed task in which children read short
sentences and indicate whether each sentence is true or false (par exemple., “The sky is blue” is “True”;
“The milk is black” is “False”). English and Spanish tasks used the Sentence Reading Fluency
subtest from the Woodcock-Johnson IV (Schrank et al., 2014) and Woodcock-Muñoz (Muñoz-
Sandoval et al., 2005), respectivement, and the Chinese task was a self-developed measure (Sun,
Zhang, Marks, Karas, et coll., 2022).
Passage reading comprehension
Passage reading comprehension was tested in English and Spanish. They both used the Pas-
sage Comprehension Woodcock-Johnson IV (Schrank et al., 2014) and Woodcock-Muñoz
(Muñoz-Sandoval et al., 2005), respectivement. Passage-level reading comprehension was not
measured in Chinese because the Chinese-speaking children were generally not able to read
and comprehend passage-long texts in Chinese.
Tableau 1 displays children’s English task performance by bilingual group, and the three
groups were maximally matched in these tasks except for English vocabulary (Monolinguals >
Bilinguals, and the two bilingual groups did not differ). Tableau 2 displays all children’s task per-
formance on the behavioral tasks by language. Note that the current sample included early
exposed, simultaneous dual-language learners with relatively balanced bilingual proficiency,
and it is typical for these children to show positive associations between skills of their two
languages (Chung et al., 2019; Wagley et al., 2022).
Neuroimaging Word Processing Task
The neuroimaging word processing task assessed children’s morpho-semantic word knowl-
edge using a lexical decision task. During each task item, children heard three words, un
target word followed by two words of choice. Children were asked to select the word that
shared either a root or derivational morpheme with the target word. Example items are bed-
room, classroom, mushroom (shared root morpheme -room); disagree, dishonest, distance
(shared derivational morpheme dis-). In the control condition, one of the choice words
matched the target in its entirety (whole word match: country, country, dentist). La tâche
followed a block design with 12 four-trial blocks (48 items in total). During each trial, partici-
pants heard three words and were instructed to select which of the last two matches the first with
Neurobiology of Language
205
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
un
_
0
0
0
9
2
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
Tableau 2.
Behavioral and neuroimaging task performance by language (Ms and SDs)
English task
Score
Chinese task
Score
r
Partial r c
Score
Spanish task
r
Partial r c
Oral Language Measures
Vocabulary
149.6 (29.9)
54.8 (29.2)
0.15
0.03
67.7 (19.4)
0.70***
0.46***
Phonological awareness
22.6 (7.5)
22.0 (9.6)
0.84***
0.79***
13.6 (6.2)
0.80***
0.79***
Morphological awareness
24.2 (10.8)
13.5 (6.3)
0.52***
0.27****
27.9 (13.3)
0.66***
0.59***
Literacy Measures
Single word reading
48.6 (15.6)
17.44 (13.8)
0.44***
0.14
42.8 (20.1)
0.67***
0.47***
Reading comprehension
26.0 (8.3)
/
/
/
19.9 (7.9)
0.69***
0.44***
Sentence reading fluency
37.2 (18.6)
11.93 (7.7)
0.54***
0.36***
26.1 (15.7)
0.76***
0.69***
Note. r = the English-Spanish or English-Chinese bivariate correlation of the respective language and literacy measure. c = partial correlation controlling for age.
*p < 0.05, **p < 0.01, ***p < 0.001.
a keypress. Each trial took 7.5 s and the whole task took about 7.2 min. An example item is
shown in Figure 1. All task items are available in Table S1 in the Supporting Information available
at https://doi.org/10.1162.nol_a_00092.
fNIRS Data Acquisition
fNIRS data were collected using the TechEN-CW6 system (NIRSOptix, 2018) with 690 and
830 nm wavelengths and a 50 Hz sampling frequency. The fNIRS cap had 12 near-infrared
light sources and 24 detectors that were symmetrically located on both hemispheres, yielding
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1. An example trial of the fNIRS word-processing task. For each trial, participants first hear the target word (e.g., “bedroom”) and see a
white box on the top of the screen, then they hear two words of choice (e.g., “classroom,” “mushroom”) and simultaneously see a blue and a
yellow box, respectively.
Neurobiology of Language
206
Neural network heterogeneity in children
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
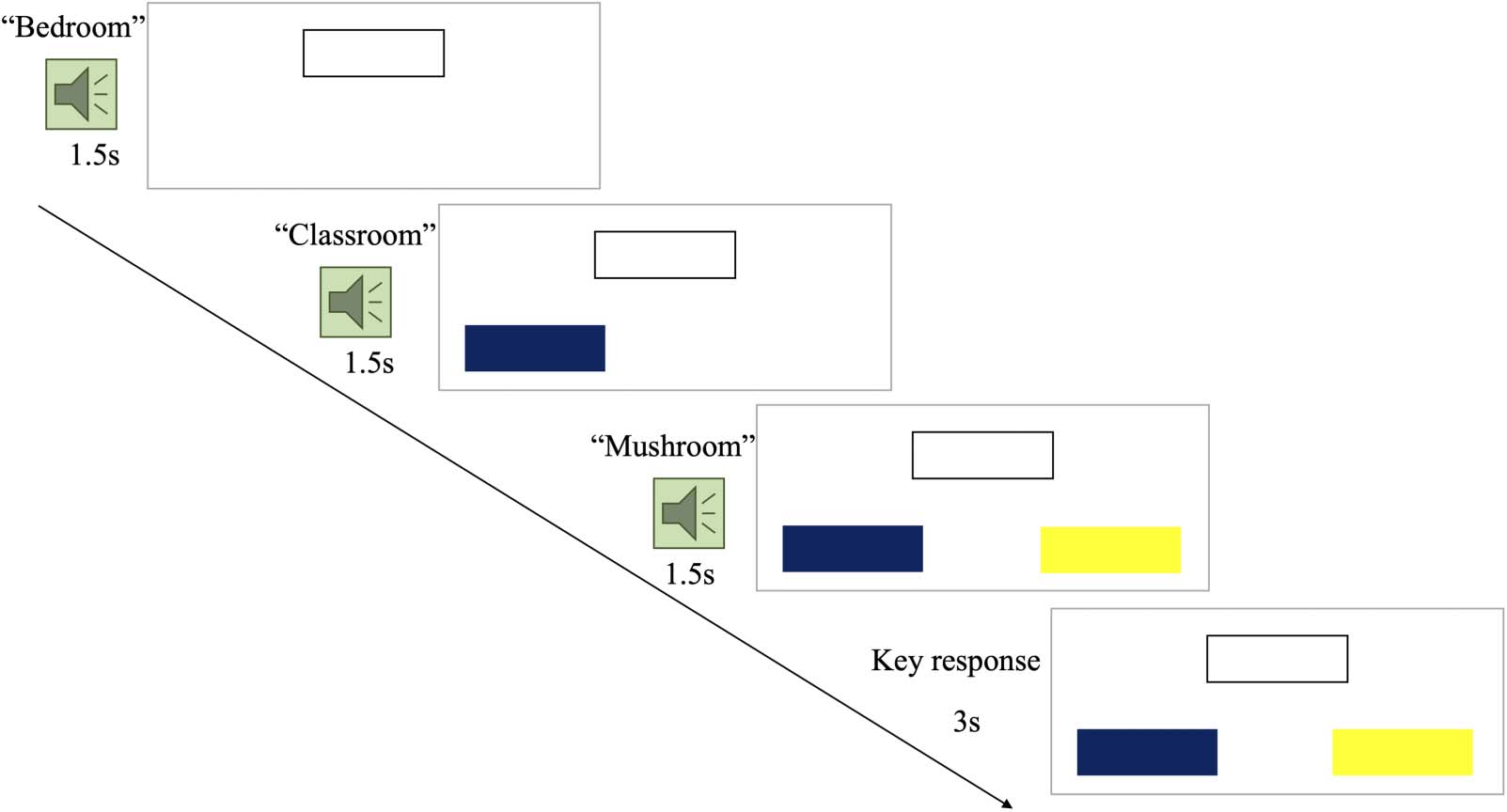
fNIRS probe setup and GIMME ROI location. Each ROI is formed by a source-detector
Figure 2.
pair. The 10 ROIs are bilateral C1, C2, C3, and left hemisphere C4, C5, C6, C7 (orange highlights).
Red squares: light sources; blue circles: light detectors. IPL: inferior parietal lobe; TTG: transverse
temporal gyri.
46 source-detector data channels (23 per hemisphere; see Figure 2). The fNIRS channels aimed
to capture key regions of language and reading networks, including frontal, temporal, and pari-
etal regions. Of important note is that fNIRS is a surface-based neuroimaging method that may
not provide the same level of precision as fMRI. Therefore, all references to anatomical loca-
tions are approximations of the neural regions maximally overlayed by specific channels. For
the current investigation, brain region localizations captured by the fNIRS channels were co-
registered using MRI as well as surface-based registration technologies. (For more information
about the channel MNI localization, see Figure S1 in the Supporting Information and Hu et al.,
2020). The depth of near-infrared light penetration was ∼3 cm, thus detecting cortical activities.
fNIRS data for the current project are openly available on the Deep Blue Data repository and
can be found in the data manuscript (Sun, Zhang, Marks, Karas, et al., 2022).
To ensure consistency in fNIRS cap placements across participants, trained experimenters
follow standardized study protocols as established in fNIRS and electroencephalography fields
to take head measurements and place caps. Specifically, experimenters first located partici-
pants’ nasion, inion, Fpz, and left and right pre-auricular points, and took the head circumfer-
ences. Next, F7, F8, T3, and T4 were anchored to their respective sources or detectors on the
fNIRS cap. Experimenters then attached the fNIRS cap to participants’ scalps and inserted the
optodes to their respective source or detector positions. Finally, experimenters conducted data
quality control by checking the participant’s cardiac signal components and the signal-to-
noise ratio among key channels of interest.
Neurobiology of Language
207
Neural network heterogeneity in children
Data Analysis
fNIRS data preprocessing
fNIRS data were analyzed with the NIRS brain AnalyzIR, a MATLAB-based toolbox (Santosa
et al., 2018), as well as self-developed scripts. Data were first downsampled from 50 Hz to
2 Hz to fit the standard analysis protocols of GIMME (as recommended by Beltz & Gates,
2017, and done in Arredondo et al., 2022). Specifically, because GIMME conducts network
mapping based on data temporal dynamics, data series with high frequency may exclusively
yield high autoregressions, making it harder to detect connections between ROIs, which are
often of primary interest (i.e., relationships between frontal and temporal regions; Beltz &
Molenaar, 2015). Next, applying the modified Beer-Lambert Law, the optical density data
was converted to hemoglobin concentration data. The data analysis focused on HbO signal
as it contributes to about 76% of the fNIRS signal and the TechEN CW6 system obtains the
HbO signal more reliably than HbR (Gagnon et al., 2012).
Regions of interest
We selected 10 ROIs with two steps. First, generally, ROIs should tap into key auditory word
and morpho-semantic processes according to prior literature (e.g., Bulut, 2022; Enge et al.,
2020; Ip et al., 2017). Thus, ROIs should include three main hubs, namely, frontal, superior
temporal, and middle temporal regions. Second, specifically, ROIs should stay engaged when
participants are working on the current task (for specific brain activation map, see Figure S2).
The final ROIs included bilateral C1 (ventral IFG [vIFG]), bilateral C2 (middle frontal gyrus
[MFG], and IFG), bilateral C3 (vIFG), left C4, and C5 (STG), and left C6 and C7 (MTG).
GIMME model fitting
GIMME builds person-specific connectivity networks with group-level, subgroup-level, and
individual-level connections based on time-series data among a set of pre-determined ROIs
(Lane et al., 2019). The connections can be contemporaneous, which depicts directed asso-
ciations between ROIs at the same time points; and the connections can be lagged, which
shows directed associations from a time point to its next time point within the same ROI or
from one ROI to another (Beltz & Gates, 2017). For the current fNIRS data set, we focused on
contemporaneous associations to better describe the cross-ROI relationships (for similar appli-
cations, see Goetschius et al., 2020). fNIRS data has high autocorrelations within a channel,
which often yields lagged connections within each individual ROI and these connections typ-
ically do not provide much meaningful information but are important to model statistically
(Smith et al., 2011, 2012).
The fNIRS HbO time-series data for each participant were extracted and submitted to the
GIMME algorithm in R (Lane et al., 2017; https://cran.r-project.org/web/packages/gimme).
GIMME first estimates a null model and gradually adds group-level connections that would
significantly improve the model fit for 75% of the sample, according to Lagrange multiplier
tests (criterion supported by simulations in Gates & Molenaar, 2012; Lane et al., 2019). After
all group-level connections are added, GIMME then prunes connections that may no longer
meet the 75% criterion. Next, GIMME ide”tifi’s subgroups using the Walktrap community
detection algorithm and adds subgroup-level connections using a 50% criterion so that iden-
tification of a subgroup connection means significantly improving model fit for 50% of the
subgroup, according to Lagrange multiplier tests (criterion supported by simulations in Lane
et al., 2019). The last stage adds significant individual-level connections for a participant,
according to Lagrange multiplier tests, until the network fits well. According to Brown
Neurobiology of Language
208
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
(2014), models with excellent fit should have at least two out of four fit indices meet the
following criteria: standardized root mean residual (SRMR) ≤ 0.05, comparative fit index
(CFI) ≥ 0.95, root mean squared error of approximation (RMSEA) ≤ 0.05, and non-normed
fit index (NNFI) ≥ 0.95.
Group and subgroup neural connectivity
Group-level connections and subgroup-level connectivity patterns were described and com-
pared by the location of the connections and connection density by subgroup. To examine
how subgroups differ from each other, we further compared participants’ in-scanner task
accuracy as well as their language and literacy task performance across subgroups with
one-way analysis of variance (ANOVA).
Person-specific neural network density
For each participant, network density was calculated by the number of connections within their
neural network (Dotterer et al., 2020; Goetschius et al., 2020). To investigate how participants’
English language and literacy proficiency is associated with their brain networks, we ran bivar-
iate and partial correlation analyses correlating network density with task performance, includ-
ing neuroimaging task accuracy and individual standardized assessments of English (i.e.,
vocabulary, word reading, reading comprehension, sentence reading fluency, respectively),
partial correlations controlling for age. To investigate how bilingual children’s heritage lan-
guage proficiency is associated with their brain networks, for each bilingual group, we further
conducted separate multiple regression analyses using heritage language vocabulary and
word/character reading to predict children’s brain network density, controlling for age and
English proficiency. We chose these two measures as indicators of heritage oral and reading
proficiency, respectively. We excluded analyses with the sentence-level fluency reading task
because many children were not able to read and comprehend full sentences in their heritage
language (N = 15 Spanish and N = 22 Chinese children were not able to complete the task).
RESULTS
The current GIMME analysis yielded well-fitting models across participants, with an average
SRMR at 0.027, CFI at 0.962, RMSEA at 0.103, and NNFI at 0.940. We next report group-,
subgroup-, and person-specific results in greater detail.
Group-Level Neural Connections
GIMME identified two group-level connections that were shared by over 75% of participants.
One was located between two left frontal channels: left C1 (IFG) and left C2 (MFG/IFG). The
second group-level connection was located between the two left MTG channels (left C6 and
C7; see Figure 3, black connections).
Subgroup Neural Connectivity
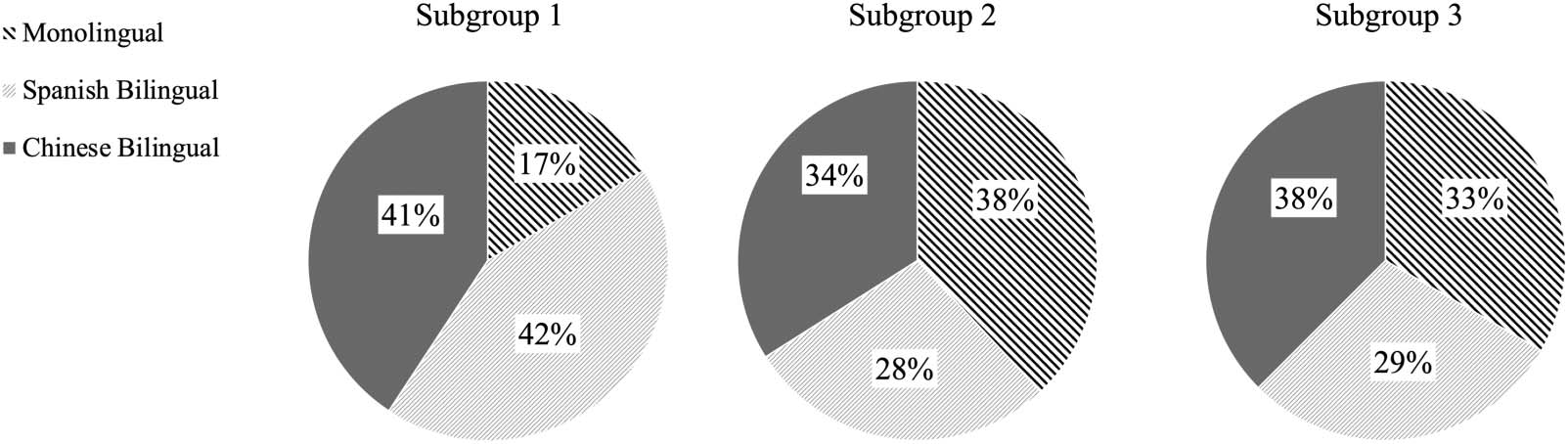
Three subgroups emerged from the data driven GIMME search. Subgroups 1, 2, and 3 had
approximately equivalent numbers of participants, N = 44, 51, and 47, respectively. Partici-
pants from the three language groups equally fell into the three subgroups, χ2(4) = 6.91, p =
0.141. Subgroup 1 had 9 monolinguals, 23 Spanish bilinguals, and 22 Chinese bilinguals. Sub-
group 2 had 19 monolinguals, 14 Spanish bilinguals, and 17 Chinese bilinguals. Subgroup 3
had 16 monolinguals, 14 Spanish bilinguals, and 18 Chinese bilinguals (see Figure 4 for a pie
chart display of the subgroup composition).
Neurobiology of Language
209
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
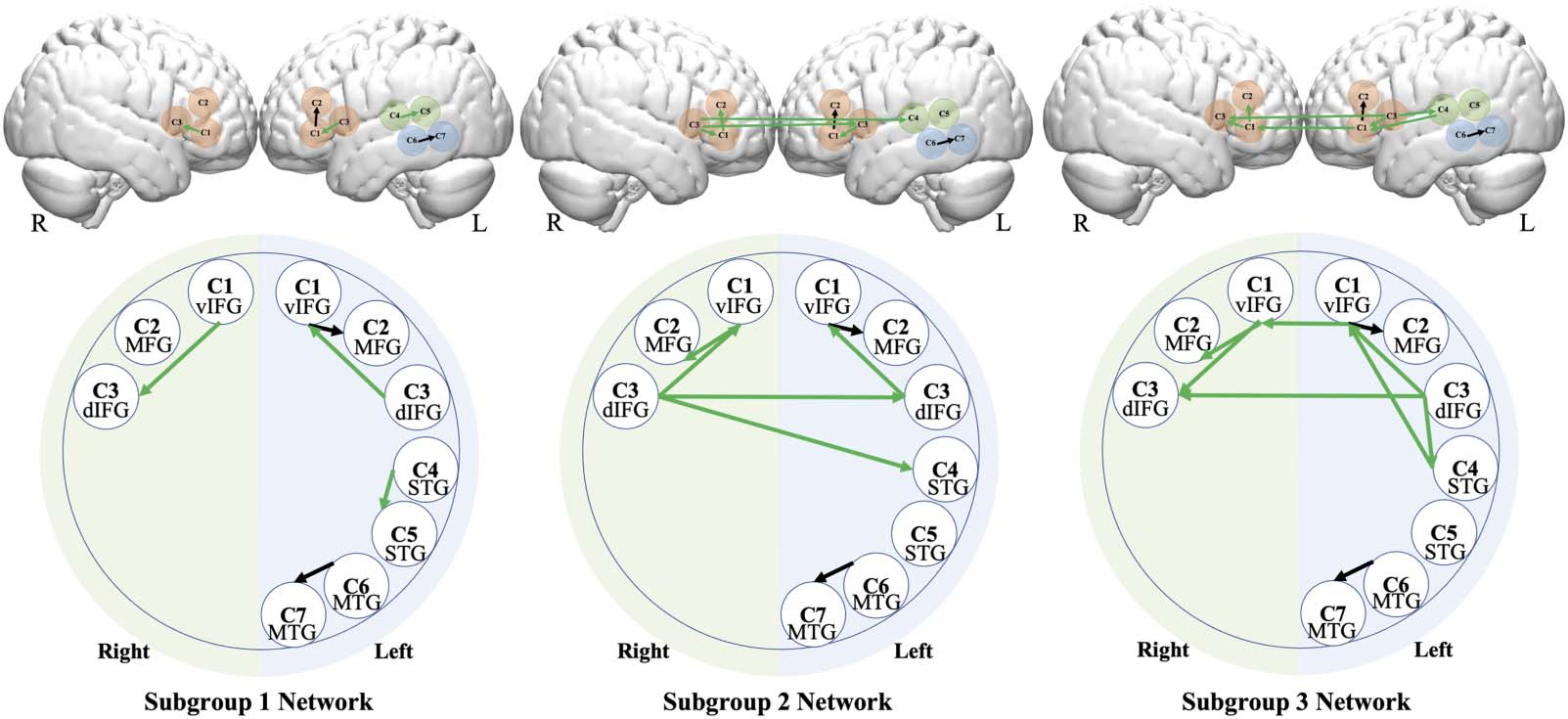
Figure 3. GIMME subgroup neural networks: The brain illustration (top row) and the map illustration (bottom row). Black lines indicate
group-level connections; green lines indicate subgroup-level connections. Each circle represents a channel. IFG: inferior frontal gyrus;
STG: superior temporal gyrus; MTG: middle temporal gyrus; d: dorsal; v: ventral.
The subgroup-level connections are shown as green connections in Figure 4. Subgroup 1
had three subgroup-level connections: within left IFG (left C3–C1); within right IFG (right C3–
C1); and between the two left STG channels (C4–C5). Subgroup 2 had five subgroup-level
connections: within left IFG (left C1–C3); left and right contralateral IFG (bilateral C3); within
right IFG (right C3–C1); right IFG and MFG (right C1–C2); and right IFG and contralateral left
STG (right C3–left C4). Subgroup 3 had eight subgroup-level connections: left and right
contralateral IFG (bilateral C1); left and right contralateral IFG (bilateral C3); within left IFG
(left C3–C1); within right IFG (right C1–C3); right IFG and MFG (right C1–C2); left STG and IFG
(left C5–C1); left IFG and STG (left C3–C5); the two left STG channels (left C4–C5).
In sum, for subgroup 1, the subgroup-level connections were exclusively within the same
brain hub (i.e., within IFG or left STG); for subgroup 2, there were additional cross-lateral
connections, especially among the phonological areas, such as between IFG and STG; and
subgroup 3 had additional left-lateralized connections across brain hubs, such as between left
IFG and STG. One-way ANOVA showed that the three subgroups differed significantly in their
network density (i.e., number of connections), F(149, 2) = 138.6, p < 0.001, η2 = 0.65.
Pairwise comparisons revealed that the three groups all differed from one another: subgroup
3 had the densest network compared to subgroup 2, followed by subgroup 1 (all ps < 0.001,
Tukey-corrected).
Figure 4. GIMME subgroup composition by language group.
Neurobiology of Language
210
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
Table 3.
Language and reading proficiency by GIMME subgroup
Age
Subgroup 1
M(SD)
7.35 (1.24)
Subgroup 2
M(SD)
7.66 (1.25)
Subgroup 3
M(SD)
8.04 (1.39)
fNIRS task accuracy (%)
76.43 (8.03)
79.00 (9.77)
81.91 (10.59)
Vocabulary
139.16 (27.98)
150.86 (28.85)
156.50 (30.52)
Phonological awareness
20.73 (7.29)
22.61 (7.23)
23.95 (7.82)
Morphological awareness
21.58 (10.94)
25.00 (10.46)
25.50 (10.96)
Single word reading
44.44 (15.91)
47.49 (15.78)
52.70 (14.32)
Reading comprehension
23.56 (8.86)
25.96 (7.99)
27.84 (7.78)
Sentence reading fluency
30.68 (16.06)
36.63 (19.50)
42.83 (18.00)
ANOVA or
Age-controlled
ANCOVA
p
0.032
Pairwise
comparison
(Tukey-applied)
G3 > G1*
0.005
<0.001 0.064 0.045 0.002 0.003 <0.001 G3 > G1**
G3 > G1***
G2 > G1*
G3 > G1+
G3 > G1*
G3 > G1**
G3 > G2*
G3 > G1**
G3 > G1***
G3 > G2*
Note. The analysis of covariance (ANCOVA) tests were age controlled except for the Age test, which used an ANOVA. +p < 0.10, *p < 0.05, **p < 0.01,
***p < 0.001.
To examine how GIMME subgroups may differ in the behavioral English tasks, we com-
pared the English behavioral task proficiency among the three groups of participants con-
trolled for age (see Table 3). Group 3 outperformed group 1 in the raw performances for all
tasks except for phonological awareness (marginal insignificance, p = 0.050).
Person-Specific Neural Network Density
Neural network density and English proficiency
Across all participants, children’s performance on all English measures, as estimated in raw
scores, was significantly associated with children’s neural network density (rs = 0.21–0.32,
ps < 0.011; Table 4). Notably, controlled for age, network density was still significantly
Table 4.
Correlation of neural network density with English language and reading proficiency
English vocabulary
English word reading
Passage comprehension
Sentence reading fluency
Neuroimaging task accuracy
Neural network density
Bivariate r
0.25
0.21
0.23
0.32
0.28
p
0.004
0.011
0.005
<0.001
<0.001
Age-controlled r
0.08
0.01
0.04
0.16
0.14
p
0.322
0.875
0.632
0.045
0.086
211
Neurobiology of Language
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
Table 5. Multiple regression predicting neural network density with Chinese/Spanish language and reading proficiency
Neural network density
Chinese bilingual
B
t
p
R2
0.161
Spanish bilingual
B
t
p
Model 1 Vocabulary as the predictor
Age
English vocabulary
Heritage language vocabulary
0.67
−0.35
0.08
2.95
−1.54
0.56
0.005
0.132
0.577
Model 2 Word reading as the predictor
0.183
Age
English word reading
0.33
−0.20
1.54
−0.96
0.131
0.340
Heritage language word reading
0.36
2.30
0.026
R2
0.125
0.030
−0.10
−0.51
0.08
0.43
0.34
2.20
0.09
−0.04
0.44
−0.17
0.28
1.36
0.611
0.735
0.033
0.664
0.866
0.181
associated with the score of reading fluency (r = 0.16, p = 0.045); while the associations with
vocabulary, word reading, and reading comprehension did not reach significance (rs = 0.01–
0.14, ps = 0.086–0.875; Table 4). Note that due to the highly correlated nature of the behav-
ioral tasks, it may not be appropriate to apply a multiple comparison correction. However, if
applied, the bivariate associations will generally survive multiple comparison corrections,
while age-controlled associations may not.
Neural network density and heritage language proficiency
For Chinese bilingual children, Chinese word reading was significantly associated with chil-
dren’s neural network density, controlling for age and English reading (B = 0.36, p = 0.026),
whereas Chinese vocabulary was not a significant predictor of neural network density (B =
0.08, p = 0.577; Table 5). In contrast, as for Spanish bilingual children, Spanish vocabulary
significantly predicted children’s neural network density controlling for age and English vocab-
ulary (B = 0.43, p = 0.033), whereas Spanish reading was not a significant predictor (B = 0.28,
p = 0.181; Table 5).
DISCUSSION
Children’s unique language experiences lead to heterogeneous behavioral and neural profiles
of language. Such individual variation makes it difficult to interpret group-level neuroimaging
findings in child language, literacy, and bilingual development (Luk & Bialystok, 2013; Marian
& Hayakawa, 2021). To advance our understanding of such heterogeneity, we used an inno-
vative person-specific approach, GIMME, to identify variation in children’s neural networks for
spoken word processing. The findings revealed that all participants, bilingual and monolingual
children, formed short-distance neural connections within the left frontal and temporal
regions, which are traditionally associated with word meaning retrieval and processing.
Children who were older and more proficient in spoken and written English showed more
long-distance connections within the broader language network and across the two hemi-
spheres, suggesting that advancements in language skills are supported by more integrated
neural networks (Hwang et al., 2013). Among bilinguals, those with stronger bilingual
Neurobiology of Language
212
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
proficiency showed greater neural network density along the key regions of language process-
ing, as a neurodevelopmental index of greater efficiency in cognitive processing (Schedlbauer
et al., 2014). The findings inform theoretical perspectives aiming to link children’s cognitive
and brain development by contextualizing the effects of heterogeneity in language experi-
ences and proficiency on their emerging neural architecture for language and literacy.
Shared Effects in the Neurobiology of Word Meaning Processes
Auditory word recognition builds upon the successful recognition of word sound and meaning
constituents (Gwilliams, 2020; Perfetti & Hart, 2002). The present study employed a morpho-
semantic word processing task that required children to dissect polysyllabic words into lexical
morphemes and analyze the meanings of the morphemic units (i.e., bedroom, classroom, and
mushroom). We acknowledge that here and henceforth our discussion of the observed results
refers to maximal anatomical overlays of the fNIRS channels (Hu et al., 2020). The findings
revealed that >75% of all participants showed common short-distance connections linking left
MTG subregions as well as IFG/MFG regions. MTG and IFG regions are commonly associated
with semantic analysis and lexical retrieval (Binder, 2017; Fiorentino & Poeppel, 2007),
whereas MFG is often associated with verbal working memory (Fegen et al., 2015; Gwilliams,
2020; Hagoort, 2019). Our findings thus support the idea that short-distance connections
within left frontal and middle temporal regions play key roles in successful word processing
by supporting morpho-semantic analyses that underlie spoken and written language develop-
ment (Arredondo et al., 2015; Ip et al., 2017; Sun et al., 2023). These shared connections have
implications for understanding the universality of language processing in children growing up
in diverse linguistic contexts.
Developmental Effects in Age and Proficiency Subgroups
GIMME identified three subgroups of participants with shared subgroup-level connections.
Subgroup 1 exhibited the simplest network with three additional short-distance connections:
one within right IFG, one within left IFG, and one within left STG regions. Subgroups 2 et 3
exhibited progressively more complex patterns with short- and long-distance connections.
They were located between the right frontal and left temporal or between the left frontal
and temporal regions. GIMME subgrouping was not related to children’s bilingual status, likely
due to the fact that all participants in the current study were proficient English language users
and attended English-only schools.
The subgrouping divisions correspond to children’s language and reading proficiency:
Controlling for age, Subgroup 1 had the least advanced English language and reading ability
and Subgroup 3 had the strongest competence. These findings suggest that language develop-
ment is supported by both short- and long-distance connectivity in a child’s brain (Ouyang
et coll., 2017). De plus, long-distance connections are likely critical in integrating different
aspects of language processes such as phonological and morpho-semantic analyses (Li
et coll., 2014; Qi et al., 2019). Of special note, the left frontotemporal connection only existed
in the most proficient Subgroup 3. This left vIFG–STG connection links regions of morpho-
semantic and phonological analyses, likely reflecting the lexically abstract derivational mor-
phemes in the current task (par exemple., singer, dancer, and finger; Gwilliams, 2020; Sun et al., 2023).
In sum, our findings suggest that children’s progress in word processing is supported by
improvements in how the language network nodes integrate to support different elements of
language subprocessing.
Neurobiology of Language
213
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
un
_
0
0
0
9
2
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
Language Proficiency and Person-Specific Network Density
To understand how neural heterogeneity speaks to children’s behavioral profiles, we exam-
ined associations between neural network density and English language and reading profi-
ciency across all participants. Prior work has linked low-density levels with early-life adversity
and disease (par exemple., Goetschius et al., 2020) whereas higher network density has been associated
with greater efficiency in cognitive tasks (Arredondo et al., 2022; Schedlbauer et al., 2014).
Donc, we had expected that children with stronger language and reading competencies
should exhibit greater network density along the key regions of language processing. This pre-
diction was generally supported by the findings, especially when we looked at children’s raw
score performance, including the in-scanner task accuracy (r = 0.30, p < 0.001) as well as the
behavioral measures (rs = 0.21–0.32, ps ≤ 0.011). This brain–behavior association remained
significant for sentence fluency controlling for age (although it should be noted that this may
not survive multiple corrections due to the highly correlated nature among the behavioral
tasks). This task requires a well-coordinated concert of word decoding, sentence comprehen-
sion, and cognitive monitoring skills, thus corresponding to a need for a more holistic neuro-
cognitive network that the current channels have covered (Norton & Wolf, 2012). The findings
for age-controlled scores for other tasks did not reach significance, likely due to the tightly
interrelated nature of age and raw performance (Qi et al., 2021). Nevertheless, their validity
is supported by both the sentence fluency task and the prior findings of positive associations
between functional connectivity and language/reading proficiency (Finn et al., 2014; Qi et al.,
2021; Skeide et al., 2016; Yu et al., 2018).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bilingual Proficiency and Person-Specific Network Density
To identify potential bilingual effects in children’s emerging neural networks for language, we
examined the role of heritage language proficiency in their network density controlling for age
and English proficiency. As heritage language measures differed across the two languages, the
analyses were done for the Spanish- and Chinese-speaking groups separately. The analyses
revealed significant contributions of heritage language proficiency to bilinguals’ neural net-
work density, but in different aspects across the two bilingual groups. In Spanish bilinguals,
the network density was associated with Spanish vocabulary, whereas in Chinese bilinguals,
the network density was associated with Chinese character reading.
There are several possible explanations for these findings. Our English word processing
functional task used in this study involves recognizing multimorphemic word units and the
ability to dissect and comprehend words is critical for literacy success (Ehri, 1998; Goodwin
et al., 2012); for bilingual learners, the properties of their home language may interact differ-
ently with English to influence this mechanism. The Spanish language contributes to English
morpho-semantic skills through a cross-linguistic transfer at points of shared morphemic units
including roots and affixes (Hernández et al., 2016). Prior behavioral data has shown that bilin-
guals with better Spanish vocabulary knowledge have better morphological literacy skills than
English monolinguals and bilinguals who are less proficient in Spanish (Kuo et al., 2017). Our
new neuroimaging findings suggest that children’s proficiency with Spanish vocabulary may
facilitate their neural efficiency for processing morphologically complex English words, poten-
tially via cross-linguistic transfer of shared morpho-semantic competencies.
In Chinese bilinguals, network density was positively associated with Chinese reading pro-
ficiency. Unlike Spanish-English bilingualism where speakers can enjoy the knowledge of
cross-linguistically shared morphemic units, there are very few shared words between Chinese
and English, as manifested by the null-to-small associations of vocabulary skills across the two
Neurobiology of Language
214
Neural network heterogeneity in children
languages (r = 0.10 according to a meta-analysis by Yang et al., 2017). Nevertheless, a critical
element of Chinese literacy is that it is monosyllabic and Chinese characters reflect mor-
phemes at the lexical level (McBride et al., 2022). Prior work has shown that Chinese-English
bilinguals place greater reliance on morpho-semantic literacy skills and show enhanced neu-
ral activations of semantic processing during morpho-semantic tasks in English, relative to
English monolinguals (Dong et al., 2020; Ip et al., 2017, 2019; Ruan et al., 2018; Sun
et al., 2023; Sun, Zhang, Marks, Nickerson, et al., 2022). Structural neuroimaging research
has found that Chinese-English bilinguals with better reading skills in both of their languages
also had thicker left AF white matter tracts linking left IFG and STG regions (Gao et al., 2022).
It is therefore possible that Chinese reading proficiency contributes to children’s neural effi-
ciency for morphologically complex words in English, potentially via cross-linguistic transfer
of morpho-syllabic literacy skills that are shared across bilinguals’ two languages.
Theoretical Contributions and Inferences
Successful word recognition builds upon neurocognitive processes and integrations of word
sound and meaning constituents. Therefore, neurodevelopmental frameworks pose that
advancements in language faculty are supported by the emergence of networks that serve both
specific and integrative language functions (Hickok & Poeppel, 2007; Werker & Hensch,
2015). Our findings advance these theoretical perspectives by demonstrating that school-
age children have developed short-range neural connections that are specific to the word
task at hand. More specifically, for our meaning-based task, most children demonstrated
short-distance functional connectivity within the left MTG regions known for their key role
in lexico-semantic processes, as well as within left IFG/ MFG regions known to support
analytical and cognitive demands for lexical tasks (Hagoort, 2019). Advancing beyond these
short-distance connections, older and more proficient language learners built long-distance
connections linking the critical regions of language functions, reflecting more integrated
neural processes (Schedlbauer et al., 2014). In other words, our findings advance theories of
language, cognition, and brain development by revealing the neurodevelopmental differences
in language network quality and its association with literacy during elementary school years.
Language experiences differ across individuals. Bilingualism adds to the variability as chil-
dren grow up with dual language experiences. Variations in bilingual experiences have long
puzzled researchers who aim to identify core features of the elusive “bilingual brain” and its
development (Claussenius-Kalman et al., 2021; Marian & Hayakawa, 2021). The present work
leveraged this variability to better understand how individual differences contribute to bilin-
gual language development and processing. Remarkably, the findings converged across
two linguistically different bilingual groups: Spanish-English and Chinese-English bilingual
children. Both groups showed greater network density in English in relation to their heritage
language skills. The findings demonstrate that heritage language skills, even in languages as
distinct as Spanish and Chinese, are related to children’s neural integration for language pro-
cessing, a core characteristic of efficient language processes.
Limitations
Several limitations should be considered when interpreting the current results. First, the sample
included a wide age range, making it somewhat difficult to dissect the impacts of developmen-
tal maturity and skill proficiency. However, our analysis was able to parse out age, and the
results, in general, revealed that both age and bilingual proficiency play significant roles in
children’s neural network connectivity for English word processing. Future studies could
Neurobiology of Language
215
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
recruit children at similar developmental stages to better obviate the effects of age. It is likely
that for children of the same ages, those with higher language and reading proficiency also
have a higher neural density within the broad language networks. Second, although the cur-
rent study was able to recruit children with heterogeneous language experiences, the sample is
still homogenous in many other aspects. For example, children were mostly from middle-class
families and attended schools in southeast Michigan. Future studies could look to dissect neural
network variation in groups that are diverse in these aspects such as socioeconomic
backgrounds. Prior resting-state research has found that adolescents with childhood adverse
experiences had sparser neural networks within the salience and default mode networks
(Goetschius et al., 2020). It is therefore likely that lower-income socioeconomic backgrounds
are associated with network sparsity within the brain regions for language. Third, the two
bilingual groups were not fully equivalent in their heritage language reading proficiency, as
the Spanish bilinguals on average had higher reading skills in Spanish than the Chinese
bilinguals in Chinese. This is likely due to their English-dominant educational context,
making it easier to transfer English literacy to Spanish than to Chinese. However, both groups
were indeed competent in reading single words/characters in their heritage language, and their
spoken language environments and proficiency were maximally equivalent.
Conclusion
The study investigated sources of heterogeneity in children’s neural organization for spoken
language skills that underlie both spoken and written language development. The findings
revealed that, across participants, children’s English language proficiency was associated with
their neural network characteristics, as manifested by the connectivity density within key brain
regions of language processes. A more focal examination of the bilingual participants in the
study further revealed that children’s dual-language proficiency was associated with their
neural network characteristics, a finding that advances our understanding of the benefits of
heritage language exposure and literacy instruction for children who speak a home language
that is different from the society’s dominant languages. The findings thus highlight the impor-
tance of understanding not only group-level but also individual effects of language experience
on the neural organization for cognitive function.
ACKNOWLEDGMENTS
We are grateful to the families in Ann Arbor, Michigan, and the surrounding neighborhoods for
their participation in our study. We also thank the research assistant team for their help with
data collection.
FUNDING INFORMATION
Ioulia Kovelman, National Institutes of Health (https://dx.doi.org/10.13039/100000002),
Award ID: R01HD092498.
AUTHOR CONTRIBUTIONS
Xin Sun: Conceptualization; Formal analysis; Investigation; Visualization; Writing – original
draft; Writing – review & editing. Rebecca A. Marks: Conceptualization; Writing – review &
editing. Rachel L. Eggleston: Investigation; Writing – review & editing. Kehui Zhang: Investi-
gation; Writing – review & editing. Chi-Lin Yu: Investigation; Writing – review & editing. Nia
Neurobiology of Language
216
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
Nickerson: Investigation; Writing – review & editing. Valeria Caruso: Investigation; Writing –
review & editing. Tai-Li Chou: Writing – review & editing. Xiao-Su Hu: Investigation; Writing –
review & editing. Twila Tardif: Writing – review & editing. James R. Booth: Writing – review &
editing. Adriene M. Beltz: Methodology; Writing – review & editing. Ioulia Kovelman:
Conceptualization; Funding acquisition; Methodology; Resources; Writing – review & editing.
DATA AVAILABILITY STATEMENT
Data for the broader project are available on Deep Blue; see Sun, Zhang, Marks, Karas, et al.
(2022, https://doi.org/10.1016/j.dib.2022.108048). For the current study, data and codes can
be found at https://osf.io/uv3t6/?view_only=46569a15ebd241808a01d51f550c65dd.
REFERENCES
Arredondo, M. M., Ip, K. I., Shih Ju Hsu, L., Tardif, T., & Kovelman,
I. (2015). Brain bases of morphological processing in young chil-
dren. Human Brain Mapping, 36(8), 2890–2900. https://doi.org
/10.1002/hbm.22815, PubMed: 25930011
Arredondo, M. M., Kovelman, I., Satterfield, T., Hu, X., Stojanov, L., &
Beltz, A. M. (2022). Person-specific connectivity mapping uncovers
differences of bilingual language experience on brain bases of
attention in children. Brain and Language, 227, Article 105084.
https://doi.org/10.1016/j.bandl.2022.105084, PubMed: 35176615
Beltz, A. M., & Gates, K. M. (2017). Network mapping with GIMME.
Multivariate Behavioral Research, 52(6), 789–804. https://doi.org
/10.1080/00273171.2017.1373014, PubMed: 29161187
Beltz, A. M., & Molenaar, P. C. (2015). A posteriori model valida-
tion for the temporal order of directed functional connectivity
maps. Frontiers in Neuroscience, 9, Article 304. https://doi.org
/10.3389/fnins.2015.00304, PubMed: 26379489
Beltz, A. M., Wright, A. G., Sprague, B. N., & Molenaar, P. C.
(2016). Bridging the nomothetic and idiographic approaches to
the analysis of clinical data. Assessment, 23(4), 447–458.
https://doi.org/10.1177/1073191116648209, PubMed:
27165092
Berken, J. A., Chai, X., Chen, J. K., Gracco, V. L., & Klein, D. (2016).
Effects of early and late bilingualism on resting-state functional
connectivity. Journal of Neuroscience, 36(4), 1165–1172.
https://doi.org/10.1523/ JNEUROSCI.1960-15.2016, PubMed:
26818505
Bialystok, E., Craik, F. I., & Luk, G. (2012). Bilingualism: Conse-
quences for mind and brain. Trends in Cognitive Sciences,
16(4), 240–250. https://doi.org/10.1016/j.tics.2012.03.001,
PubMed: 22464592
Binder, J. R. (2017). Current controversies on Wernicke’s area and
its role in language. Current Neurology and Neuroscience
Reports, 17, 58. https://doi.org/10.1007/s11910-017-0764-8,
PubMed: 28656532
Brown, T. A. (2014). Confirmatory factor analysis for applied
research. Guilford Press.
Bulut, T. (2022). Neural correlates of morphological processing:
An activation likelihood estimation meta-analysis. Cortex, 151,
49–69. https://doi.org/10.1016/j.cortex.2022.02.010, PubMed:
35397379
Cao, F., Bitan, T., & Booth, J. R. (2008). Effective brain connectivity
in children with reading difficulties during phonological process-
ing. Brain and Language, 107(2), 91–101. https://doi.org/10.1016
/j.bandl.2007.12.009, PubMed: 18226833
Chung, S. C., Chen, X., & Geva, E. (2019). Deconstructing and
reconstructing cross-language transfer in bilingual reading
development: An interactive framework. Journal of Neurolinguis-
tics, 50, 149–161. https://doi.org/10.1016/j.jneuroling.2018.01
.003
Claussenius-Kalman, H., Hernandez, A. E., & Li, P. (2021).
Expertise, ecosystem, and emergentism: Dynamic developmental
bilingualism. Brain and Language, 222, Article 105013. https://
doi.org/10.1016/j.bandl.2021.105013, PubMed: 34520977
Dong, Y., Tang, Y., Chow, B. W.-Y., Wang, W., & Dong, W.-Y.
(2020). Contribution of vocabulary knowledge to reading com-
prehension among Chinese students: A meta-analysis. Frontiers
in Psychology, 11, Article 525369. https://doi.org/10.3389/fpsyg
.2020.525369, PubMed: 33132948
Dotterer, H. L., Hyde, L. W., Shaw, D. S., Rodgers, E. L., Forbes,
E. E., & Beltz, A. M. (2020). Connections that characterize
callousness: Affective features of psychopathy are associated
with personalized patterns of resting-state network connectivity.
NeuroImage: Clinical, 28, Article 102402. https://doi.org/10
.1016/j.nicl.2020.102402, PubMed: 32891038
Dunn, D. M. (2019). Peabody Picture Vocabulary Test (5th ed.).
Pearson.
Dunn, L. M., & Dunn, L. M. (1998). The Peabody Picture Vocabu-
lary Test—Revised (L. Lu & H. Liu, Trans.; Chinese ed.). Taipei:
Psychology Publisher. (Original work published in 1997)
Dunn, L., Padilla, E., Lugo, D., & Dunn, L. (1986). TVIP: Test Voca-
bolario Imágenes Peabody. Pearson.
Ehri, L. C. (1998). Grapheme-phoneme knowledge is essential
for learning to read words in English. In J. L. Metsala & L. C. Ehri
(Eds.), Word recognition in beginning literacy (pp. 3–40).
Routledge. https://doi.org/10.4324/9781410602718-6
Enge, A., Friederici, A. D., & Skeide, M. A. (2020). A meta-analysis of
fMRI studies of language comprehension in children. NeuroImage,
215, Article 116858. https://doi.org/10.1016/j.neuroimage.2020
.116858, PubMed: 32304886
Fegen, D., Buchsbaum, B. R., & D’Esposito, M. (2015). The effect of
rehearsal rate and memory load on verbal working memory.
NeuroImage, 105, 120–131. https://doi.org/10.1016/j.neuroimage
.2014.10.034, PubMed: 25467303
Finn, E. S. (2021). Is it time to put rest to rest? Trends in Cognitive
Sciences, 25(12), 1021–1032. https://doi.org/10.1016/j.tics.2021
.09.005, PubMed: 34625348
Finn, E. S., Shen, X., Holahan, J. M., Scheinost, D., Lacadie, C.,
Papademetris, X., Shaywitz, S. S., Shaywitz, B. A., & Constable,
R. T. (2014). Disruption of functional networks in dyslexia: A
whole-brain, data-driven analysis of connectivity. Biological Psy-
chiatry, 76(5), 397–404. https://doi.org/10.1016/j.biopsych.2013
.08.031, PubMed: 24124929
Neurobiology of Language
217
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
Fiorentino, R., & Poeppel, D. (2007). Compound words and struc-
ture in the lexicon. Language and Cognitive Processes, 22(7),
953–1000. https://doi.org/10.1080/01690960701190215
Francis, D., Carlo, M., August, D., Kenyon, D., Malabonga, V.,
Caglarcan, S., & Louguit, M. (2001). Test of Phonological Pro-
cessing in Spanish. Center for Applied Linguistics.
Friederici, A. D., Brauer, J., & Lohmann, G. (2011). Maturation of
the language network: From inter- to intrahemispheric connectiv-
ities. PLOS ONE, 6(6), Article e20726. https://doi.org/10.1371
/journal.pone.0020726, PubMed: 21695183
Gagnon, L., Yücel, M. A., Dehaes, M., Cooper, R. J., Perdue, K. L.,
Selb, J., Huppert, T. J., Hoge, R. D., & Boas, D. A. (2012). Quan-
tification of the cortical contribution to the NIRS signal over the
motor cortex using concurrent NIRS-fMRI measurements. Neuro-
Image, 59(4), 3933–3940. https://doi.org/10.1016/j.neuroimage
.2011.10.054, PubMed: 22036999
Gao, Y., Meng, X., Bai, Z., Liu, X., Zhang, M., Li, H., Ding, G., Liu,
L., & Booth, J. R. (2022). Left and right arcuate fasciculi are
uniquely related to word reading skills in Chinese-English bilin-
gual children. Neurobiology of Language, 3(1), 109–131. https://
doi.org/10.1162/nol_a_00051
García-Pentón, L., Pérez Fernández, A., Iturria-Medina, Y., Gillon-
Dowens, M., & Carreiras, M. (2014). Anatomical connectivity
changes in the bilingual brain. NeuroImage, 84, 495–504.
https://doi.org/10.1016/j.neuroimage.2013.08.064, PubMed:
24018306
Gates, K. M., & Molenaar, P. C. (2012). Group search algorithm
recovers effective connectivity maps for individuals in homoge-
neous and heterogeneous samples. NeuroImage, 63(1), 310–319.
https://doi.org/10.1016/j.neuroimage.2012.06.026, PubMed:
22732562
Goetschius, L. G., Hein, T. C., McLanahan, S. S., Brooks-Gunn, J.,
McLoyd, V. C., Dotterer, H. L., Lopez-Duran, N., Mitchell, C.,
Hyde, L. W., Monk, C. S., & Beltz, A. M. (2020). Association
of childhood violence exposure with adolescent neural network
density. JAMA Network Open, 3(9), Article e2017850. https://doi
.org/10.1001/jamanetworkopen.2020.17850, PubMed:
32965498
Goodwin, A., Lipsky, M., & Ahn, S. (2012). Word detectives: Using
units of meaning to support literacy. The Reading Teacher, 65(7),
461–470. https://doi.org/10.1002/TRTR.01069
Gwilliams, L. (2020). How the brain composes morphemes into
meaning. Philosophical Transactions of the Royal Society B,
375(1791), Article 20190311. https://doi.org/10.1098/rstb.2019
.0311, PubMed: 31840591
Hagoort, P. (2019). The neurobiology of language beyond
single-word processing. Science, 366(6461), 55–58. https://doi
.org/10.1126/science.aax0289, PubMed: 31604301
Hernández, A. C., Montelongo, J. A., & Herter, R. J. (2016). Using
Spanish-English cognates in children’s choices picture books to
develop Latino English learners’ linguistic knowledge. The Read-
ing Teacher, 70(2), 233–239. https://doi.org/10.1002/trtr.1511
Hernandez, A. E., Claussenius-Kalman, H. L., Ronderos, J., Castilla-
Earls, A. P., Sun, L., Weiss, S. D., & Young, D. R. (2019). Neuro-
emergentism: A framework for studying cognition and the brain.
Journal of Neurolinguistics, 49, 214–223. https://doi.org/10.1016
/j.jneuroling.2017.12.010, PubMed: 30636843
Hickok, G. (2022). The dual stream model of speech and language
processing. Handbook of Clinical Neurology, 185, 57–69. https://
doi.org/10.1016/ B978-0-12-823384-9.00003-7, PubMed:
35078610
Hickok, G., & Poeppel, D. (2007). The cortical organization of speech
processing. Nature Reviews Neuroscience, 8(5), 393–402. https://
doi.org/10.1038/nrn2113, PubMed: 17431404
Hu, X.-S., Wagley, N., Rioboo, A. T., DaSilva, A. F., & Kovelman, I.
(2020). Photogrammetry-based stereoscopic optode registration
method for functional near-infrared spectroscopy. Journal of
Biomedical Optics, 25(9), Article 095001. https://doi.org/10
.1117/1.JBO.25.9.095001, PubMed: 32880124
Hwang, K., Hallquist, M. N., & Luna, B. (2013). The development
of hub architecture in the human functional brain network. Cere-
bral Cortex, 23(10), 2380–2393. https://doi.org/10.1093/cercor
/bhs227, PubMed: 22875861
Ip, K. I., Hsu, L. S.-J., Arredondo, M. M., Tardif, T., & Kovelman, I.
(2017). Brain bases of morphological processing in Chinese-
English bilingual children. Developmental Science, 20(5), Article
e12449. https://doi.org/10.1111/desc.12449, PubMed:
27523024
Ip, K. I., Marks, R. A., Hsu, L. S.-J., Desai, N., Kuan, J. L., & Tardif, T.
(2019). Morphological processing in Chinese engages left tempo-
ral regions. Brain and Language, 199, Article 104696. https://doi
.org/10.1016/j.bandl.2019.104696, PubMed: 31655417
Jasińska, K. K., Shuai, L., Lau, A. N. L., Frost, S., Landi, N., & Pugh,
K. R. (2020). Functional connectivity in the developing language
network in 4-year-old children predicts future reading ability.
Developmental Science, 24(2), Article e13041. https://doi.org
/10.1111/desc.13041, PubMed: 33032375
Kovelman, I., Baker, S. A., & Petitto, L. A. (2008). Age of first
bilingual language exposure as a new window into bilingual
reading development. Bilingualism: Language and Cognition,
11(2), 203–223. https://doi.org/10.1017/S1366728908003386,
PubMed: 19823598
Kuo, L.-J., & Anderson, R. C. (2006). Morphological awareness
and learning to read: A cross-language perspective. Educational
Psychologist, 41(3), 161–180. https://doi.org/10.1207
/s15326985ep4103_3
Kuo, L.-J., Ramirez, G., de Marin, S., Kim, T.-J., & Unal-Gezer, M.
(2017). Bilingualism and morphological awareness: A study with
children from general education and Spanish-English dual lan-
guage programs. Educational Psychology, 37(2), 94–111.
https://doi.org/10.1080/01443410.2015.1049586
Lane, S. [T.], Gates, K. [M.], Molenaar, P., Hallquist, M., & Pike, H.
(2017). GIMME: Group iterative multiple model estimation
[Computer software manual]. https://cran.r-project.org/web
/packages/gimme
Lane, S. T., Gates, K. M., Pike, H. K., Beltz, A. M., & Wright, A. G. C.
(2019). Uncovering general, shared, and unique temporal patterns
in ambulatory assessment data. Psychological Methods, 24(1),
54–69. https://doi.org/10.1037/met0000192, PubMed: 30124300
Li, H., Xue, Z., Ellmore, T. M., Frye, R. E., & Wong, S. T. (2014).
Network-based analysis reveals stronger local diffusion-based
connectivity and different correlations with oral language skills
in brains of children with high functioning autism spectrum dis-
orders. Human Brain Mapping, 35(2), 396–413. https://doi.org
/10.1002/hbm.22185, PubMed: 23008187
Liu, H., & Cao, F. (2016). L1 and L2 processing in the bilingual
brain: A meta-analysis of neuroimaging studies. Brain and Lan-
guage, 159, 60–73. https://doi.org/10.1016/j.bandl.2016.05
.013, PubMed: 27295606
Luk, G., & Bialystok, E. (2013). Bilingualism is not a categorical
variable: Interaction between language proficiency and usage.
Journal of Cognitive Psychology, 25(5), 605–621. https://doi.org
/10.1080/20445911.2013.795574, PubMed: 24073327
Marian, V., Bartolotti, J., Rochanavibhata, S., Bradley, K., & Hernandez,
A. E. (2017). Bilingual cortical control of between-and within-
language competition. Scientific Reports, 7(1), Article 11763.
https://doi.org/10.1038/s41598-017-12116-w, PubMed:
28924215
Neurobiology of Language
218
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
Marian, V., & Hayakawa, S. (2021). Measuring bilingualism: The
quest for a “bilingualism quotient.” Applied Psycholinguistics,
42(S2), 527–548. https://doi.org/10.1017/S0142716420000533,
PubMed: 34054162
Marks, R. A., Labotka, D., Sun, X., Nickerson, N., Zhang, K., Eggleston,
R. L., Yu, C.-L., Uchikoshi, Y., Hoeft, F., & Kovelman, I. (2022).
Morphological awareness and its role in early word reading in
English monolinguals, Spanish–English, and Chinese–English
simultaneous bilinguals. Bilingualism: Language and Cognition,
26(2), 268–283. https://doi.org/10.1017/S1366728922000517
Marks, R. A., Sun, X., McAlister López, E., Nickerson, N.,
Hernandez, I., Caruso, V. C., Satterfiled, T., & Kovelman, I.
(2022). Cross-linguistic differences in the associations between
morphological awareness and reading in Spanish and English
in young simultaneous bilinguals. International Journal of Bilin-
gual Education and Bilingualism, 25(10), 3907–3923. https://doi
.org/10.1080/13670050.2022.2090226
McBride, C., Pan, D. J., & Mohseni, F. (2022). Reading and writing
words: A cross-linguistic perspective. Scientific Studies of
Reading, 26(2), 125–138. https://doi.org/10.1080/10888438
.2021.1920595
Mohades, S. G., Struys, E., Van Schuerbeek, P., Mondt, K., Van De
Craen, P., & Luypaert, R. (2012). DTI reveals structural differ-
ences in white matter tracts between bilingual and monolingual
children. Brain Research, 1435, 72–80. https://doi.org/10.1016/j
.brainres.2011.12.005, PubMed: 22197702
Muñoz-Sandoval, A. F., Woodcock, R. W., McGrew, K. S., &
Mather, N. (2005). Batería III Woodcock-Muñoz. Riverside
Publishing.
Newman, E. H., Tardif, T., Huang, J., & Shu, H. (2011). Phonemes
matter: The role of phoneme-level awareness in emergent
Chinese readers. Journal of Experimental Child Psychology,
108(2), 242–259. https://doi.org/10.1016/j.jecp.2010.09.001,
PubMed: 20980019
NIRSOptix. 2018. CW6 system [Apparatus]. https://nirsoptix.com
/CW6.html
Norton, E. S., & Wolf, M. (2012). Rapid automatized naming (RAN)
and reading fluency: Implications for understanding and
treatment of reading disabilities. Annual Review of Psychology,
63(1), 427–452. https://doi.org/10.1146/annurev-psych-120710
-100431, PubMed: 21838545.
Ouyang, M., Kang, H., Detre, J. A., Roberts, T. P., & Huang, H.
(2017). Short-range connections in the developmental connec-
tome during typical and atypical brain maturation. Neuroscience
& Biobehavioral Reviews, 83, 109–122. https://doi.org/10.1016/j
.neubiorev.2017.10.007, PubMed: 29024679
Perfetti, C. A., & Hart, L. (2002). The lexical quality hypothesis. In
L. Verhoeven, C. Elbro, & P. Reitsma (Eds.), Precursors of func-
tional literacy (pp. 189–213). John Benjamins. https://doi.org/10
.1075/swll.11.14per
Price, R. B., Beltz, A. M., Woody, M. L., Cummings, L., Gilchrist,
D., & Siegle, G. J. (2020). Neural connectivity subtypes predict
discrete attentional-bias profiles among heterogeneous anxiety
patients. Clinical Psychological Science, 8(3), 491–505. https://
doi.org/10.1177/2167702620906149, PubMed: 33758682
Qi, T., Schaadt, G., Cafiero, R., Brauer, J., Skeide, M. A., & Friederici,
A. D. (2019). The emergence of long-range language network
structural covariance and language abilities. NeuroImage, 191,
36–48. https://doi.org/10.1016/j.neuroimage.2019.02.014,
PubMed: 30738206
Qi, T., Schaadt, G., & Friederici, A. D. (2021). Associated func-
tional network development and language abilities in children.
NeuroImage, 242, Article 118452. https://doi.org/10.1016/j
.neuroimage.2021.118452, PubMed: 34358655
Ruan, Y., Georgiou, G. K., Song, S., Li, Y., & Shu, H. (2018). Does
writing system influence the associations between phonological
awareness, morphological awareness, and reading? A meta-
analysis. Journal of Educational Psychology, 110(2), 180–202.
https://doi.org/10.1037/edu0000216
Santosa, H., Zhai, X., Fishburn, F., & Huppert, T. (2018). The NIRS
brain AnalyzIR toolbox. Algorithms, 11(5), 73. https://doi.org/10
.3390/a11050073
Schedlbauer, A. M., Copara, M. S., Watrous, A. J., & Ekstrom, A. D.
(2014). Multiple interacting brain areas underlie successful
spatiotemporal memory retrieval in humans. Scientific Reports,
4(1), Article 6431. https://doi.org/10.1038/srep06431, PubMed:
25234342
Schrank, F. A., McGrew, K. S., & Mather, N. (2014). Woodcock-
Johnson IV Tests of Cognitive Abilities. Riverside.
Skeide, M. A., Brauer, J., & Friederici, A. D. (2016). Brain functional
and structural predictors of language performance. Cerebral
Cortex, 26(5), 2127–2139. https://doi.org/10.1093/cercor
/bhv042, PubMed: 25770126
Skeide, M. A., & Friederici, A. D. (2016). The ontogeny of the
cortical language network. Nature Reviews Neuroscience,
17(5), 323–332. https://doi.org/10.1038/nrn.2016.23, PubMed:
27040907
Smith, S. M., Bandettini, P. A., Miller, K. L., Behrens, T. E. J., Friston,
K. J., David, O., Liu, T., Woolrich, M. W., & Nichols, T. E. (2012).
The danger of systematic bias in group-level FMRI-lag-based
causality estimation. NeuroImage, 59(2), 1228–1229. https://doi
.org/10.1016/j.neuroimage.2011.08.015, PubMed: 21867760
Smith, S. M., Miller, K. L., Salimi-Khorshidi, G., Webster, M.,
Beckmann, C. F., Nichols, T. E., Ramsey, J. D., & Woolrich,
M. W. (2011). Network modelling methods for FMRI. Neuro-
Image, 54(2), 875–891. https://doi.org/10.1016/j.neuroimage
.2010.08.063, PubMed: 20817103
Song, S., Su, M., Kang, C., Liu, H., Zhang, Y., McBride-Chang, C.,
Tardif, T., Li, H., Liang, W., Zhang, Z., & Shu, H. (2015). Tracing
children’s vocabulary development from preschool through the
school-age years: An 8-year longitudinal study. Developmental
Science, 18(1), 119–131. https://doi.org/10.1111/desc.12190,
PubMed: 24962559
Su, M., Zhao, J., de Schotten, M. T., Zhou, W., Gong, G., Ramus, F.,
& Shu, H. (2018). Alterations in white matter pathways underly-
ing phonological and morphological processing in Chinese
developmental dyslexia. Developmental Cognitive Neurosci-
ence, 31, 11–19. https://doi.org/10.1016/j.dcn.2018.04.002,
PubMed: 29727819
Sulpizio, S., Del Maschio, N., Fedeli, D., & Abutalebi, J. (2020).
Bilingual language processing: A meta-analysis of functional
neuroimaging studies. Neuroscience & Biobehavioral Reviews,
108, 834–853. https://doi.org/10.1016/j.neubiorev.2019.12.014,
PubMed: 31838193
Sun, X., Li, L., Ding, G., Wang, R., & Li, P. (2019). Effects of
language proficiency on cognitive control: Evidence from
resting-state functional connectivity. Neuropsychologia, 129,
263–275. https://doi.org/10.1016/j.neuropsychologia.2019.03
.020, PubMed: 30951741
Sun, X., Marks, R. A., Zhang, K., Yu, C.-L., Eggleston, R. L.,
Nickerson, N., Chou, T.-L., Hu, X.-S., Tardif, T., Satterfield, T.,
& Kovelman, I. (2023). Brain bases of English morphological
p r o c e s s i n g : A c o m p a r i s o n b e t w e e n C h i n e s e - E n g l i s h ,
Spanish-English bilingual, and English monolingual children.
Developmental Science, 26(1), Article e13251. https://doi.org
/10.1111/desc.13251, PubMed: 35188687
Sun, X., Zhang, K., Marks, R. [A.], Karas, Z., Eggleston, R.,
Nickerson, N., Yu, C.-L., Wagley, N., Hu, X., Caruso, V., Chou,
Neurobiology of Language
219
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neural network heterogeneity in children
T.-L., Satterfield, T., Tardif, T., & Kovelman, I. (2022). Morphological
and phonological processing in English monolingual, Chinese–
English bilingual, and Spanish–English bilingual children: An fNIRS
neuroimaging dataset. Data in Brief, 42, Article 108048. https://doi
.org/10.1016/j.dib.2022.108048, PubMed: 35313503
Sun, X., Zhang, K., Marks, R. A., Nickerson, N., Eggleston, R. L., Yu,
C.-L., Chou, T.-L., Tardif, T., & Kovelman, I. (2022). What’s in a
word? Cross-linguistic influences on Spanish-English and
Chinese-English bilingual children’s word reading development.
Child Development, 93(1), 84–100. https://doi.org/10.1111/cdev
.13666, PubMed: 34570366
Thieba, C., Long, X., Dewey, D., & Lebel, C. (2019). Young children
in different linguistic environments: A multimodal neuroimaging
study of the inferior frontal gyrus. Brain and Cognition, 134, 71–79.
https://doi.org/10.1016/j.bandc.2018.05.009, PubMed: 30007529
Wagley, N., Marks, R. A., Bedore, L. M., & Kovelman, I. (2022).
Contributions of bilingual home environment and language pro-
ficiency on children’s Spanish–English reading outcomes. Child
Development, 93(4), 881–899. https://doi.org/10.1111/cdev
.13748, PubMed: 35289947
Wagner, R. K., Torgesen, J. K., Rashotte, C. A., & Pearson, N. A. (1999).
Comprehensive Test of Phonological Processing: CTOPP. Pro-ed.
Wechsler, D. (2014). Wechsler Intelligence Scale for Children—Fifth
Edition ( WISC-V). The Psychological Corporation.
Weiss-Croft, L. J., & Baldeweg, T. (2015). Maturation of language
networks in children: A systematic review of 22 years of func-
tional MRI. NeuroImage, 123, 269–281. https://doi.org/10.1016
/j.neuroimage.2015.07.046, PubMed: 26213350
Werker, J. F., & Hensch, T. K. (2015). Critical periods in speech per-
ception: New directions. Annual Review of Psychology, 66(1),
173–196. https://doi.org/10.1146/annurev-psych-010814
-015104, PubMed: 25251488
Xiao, Y., Friederici, A. D., Margulies, D. S., & Brauer, J. (2016). Lon-
gitudinal changes in resting-state fMRI from age 5 to age 6 years
covary with language development. NeuroImage, 128, 116–124.
https://doi.org/10.1016/j.neuroimage.2015.12.008, PubMed:
26690809
Yang, M., Cooc, N., & Sheng, L. (2017). An investigation of
cross-linguistic transfer between Chinese and English: A meta-
analysis. Asian-Pacific Journal of Second and Foreign Language
Education, 2(1), Article 15. https://doi.org/10.1186/s40862-017
-0036-9
Yeatman, J. D., Dougherty, R. F., Rykhlevskaia, E., Sherbondy, A. J.,
Deutsch, G. K., Wandell, B. A., & Ben-Shachar, M. (2011). Ana-
tomical properties of the arcuate fasciculus predict phonological
and reading skills in children. Journal of Cognitive Neuroscience,
23(11), 3304–3317. https://doi.org/10.1162/jocn_a_00061,
PubMed: 21568636
Yu, X., Ferradal, S. L., Sliva, D. D., Dunstan, J., Carruthers, C.,
Sanfilippo, J., Zuk, J., Zöllei, L., Boyd, E., Gagoski, B., Ou, Y.,
Grant, P. E., & Gaab, N. (2021). Functional connectivity in
infancy and toddlerhood predicts long-term language and preli-
teracy outcomes. Cerebral Cortex, Article bhab230. https://doi
.org/10.1093/cercor/bhab230, PubMed: 34347052
Yu, X., Raney, T., Perdue, M. V., Zuk, J., Ozernov-Palchik, O.,
Becker, B. L., Raschle, N. M., & Gaab, N. (2018). Emergence
of the neural network underlying phonological processing from
the prereading to the emergent reading stage: A longitudinal
study. Human Brain Mapping, 39(5), 2047–2063. https://doi.org
/10.1002/hbm.23985, PubMed: 29380469
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
4
2
1
9
8
2
0
7
9
0
1
9
n
o
_
a
_
0
0
0
9
2
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
220