ARTICLE DE RECHERCHE
Combining Concepts Across Categorical
Domains: A Linking Role of the Precuneus
Giuseppe Rabini
, Silvia Ubaldi
, and Scott Fairhall
Centre for Mind/ Brain Sciences (CIMeC), University of Trento, Trento, Italy
un accès ouvert
journal
Mots clés: IRMf, combinatorial semantics, langue, phrases, precuneus, category selectivity
ABSTRAIT
The human capacity for semantic knowledge entails not only the representation of single
concepts but also the capacity to combine these concepts into the increasingly complex ideas
that underlie human thought. This process involves not only the combination of concepts
from within the same semantic category but also frequently the conceptual combination
across semantic domains. In this fMRI study (N = 24) we investigate the cortical mechanisms
underlying our ability to combine concepts across different semantic domains. Using five
different semantic domains (Personnes, Places, Food, Objects, and Animals), we present
sentences depicting concepts drawn from a single semantic domain as well as sentences that
combine concepts from two of these domains. Contrasting single-category and combined-
category sentences reveals that the precuneus is more active when concepts from different
domains have to be combined. En même temps, we observe that distributed category
selectivity representations persist when higher-order meaning involves the combination of
categories and that this category-selective response is captured by the combination of the
single categories composing the sentence. Collectively, these results suggest that the precuneus
plays a role in the combination of concepts across different semantic domains, potentiellement
functioning to link together category-selective representations distributed across the cortex.
Citation: Rabini, G., Ubaldi, S., &
Fairhall, S. (2021). Combining concepts
across categorical domains: A linking
role of the precuneus. Neurobiology of
Language, 2(3), 354–371. https://doi.org
/10.1162/nol_a_00039
EST CE QUE JE:
https://doi.org/10.1162/nol_a_00039
Informations complémentaires:
https://doi.org/10.1162/nol_a_00039
Reçu: 13 Décembre 2020
Accepté: 8 Avril 2021
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
INTRODUCTION
Auteur correspondant:
Scott Fairhall
scott.fairhall@unitn.it
Éditeur de manipulation:
Yanchao Bi
droits d'auteur: © 2021
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
The human capacity for semantic knowledge involves not only the representation of single
concepts but also the capacity to combine these concepts into the increasingly complex ideas
that underlie human thought. A wealth of research on single concepts has shown that the hu-
man brain implements semantic representation over a complex system involving regions that
are sensitive to specific semantic classes of objects, such as people, food, or places, in addition
to regions that are generally more active for semantically richer stimuli, regardless of category.
Accordingly, the functioning of the semantic system is reflected in a dynamic interplay be-
tween domain-specific and domain-general representations (Binder, 2016; Binder et al.,
2009; Binder & Desai, 2011; Chen et al., 2017; Kiefer & Pulvermüller, 2012; Martine, 2016;
Patterson et al., 2007). The distributed representation of semantic knowledge in the brain po-
tentially indicates a fundamental organisational principle, whereby basic object-related
knowledge extends to complex, multi-faceted units of meaning.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
un
_
0
0
0
3
9
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
The combination of concepts into higher-order representations not only involves the linking
of concepts from within the same semantic domain but also frequently entails the flexible
association of concepts spanning different domains. Par exemple, reading about a boy playing
with his dog in the garden requires the system to link concepts from distinct conceptual domains
(c'est à dire., people, animals, and places) to build a distinct and coherent representation. À ce jour, comment-
jamais, it is still unclear how category-selective brain regions interact when concepts from different
domains have to be combined into higher-order semantic units. En particulier, there remain two
open questions: (1) Do specific brain regions coordinate information contained in category-
selective cortical regions? Et (2) do complex ideas that combine category information from
multiple domains continue to utilise those domain-selective representations of single concepts,
or do derived multi-categorical concepts rely more on domain-general semantic mechanisms?
The general semantic processing of semantically richer stimuli recruits a left-lateralised cor-
tical network encompassing several heteromodal associative regions (the angular gyrus (AG),
lateral temporal cortex, ventral temporal cortex, dorso-medial and ventro-medial prefrontal cor-
tex, inferior frontal gyrus (IFG), and the precuneus) (Binder et al., 2009). Although modality-
specific activations emerge during modality-specific conceptual processing—for example,
perceptual/motor-related concepts activate the respective perceptual/motor brain areas (voir
Binder & Desai, 2011; Borghesani & Piazza, 2017; Kiefer & Pulvermüller, 2012)—the general
semantic network appears to be clearly distinguished from primary sensory and motor cortices
(Binder et al., 2009). While a strong embodied view of cognition states that conceptual knowl-
edge emerges exclusively from sensory and action/motor experience and is therefore grounded
and represented in the related cortical regions (Barsalou, 2010; Gallese, 2005; see also Mahon &
Caramazza, 2008, for a critical perspective), a softer version (“embodied abstraction”; Binder &
Desai, 2011) gives an alternative view. Spécifiquement, it states that different levels of abstraction starting
from sensory, moteur, and emotional experiences model our conceptual representation. Higher-level
concepts can be abstracted from the primary sensory meaning, and different levels of abstraction
can be selectively activated depending on several factors, such as context and task demand. Dans
this view, heteromodal cortices might be involved in the representation of more abstract—high-
level—concepts, which are not necessarily and directly linked to sensorimotor experience.
The position has found complementary support in neurocomputational models of neural
semantic representation (Chen et al., 2017) and in recent theoretical proposals (Lambon
Ralph et al., 2016). In the controlled semantic cognition framework, the representational sub-
system has been described as “the hub-and-spoke model” (Patterson et al., 2007; Rogers et al.,
2004). This model predicts that concepts emerge from both verbal and nonverbal experience
and that modality-selective cortices, distributed across the whole brain, represent this specific
information (“the spokes”). En outre, a unique transmodal core region (“the hub”), identi-
fied in the ventro-lateral anterior temporal lobe (ATL) (see also Visser et al., 2010), would be
engaged in modality-invariant representations, so that this region can represent object-
concepts in multimodal, more abstract, global units. Recent advances further elaborated on
this proposal by advocating a graded specialisation within the ATL, based on the different con-
nectivity patterns of its subparts (Lambon Ralph et al., 2016; Rice et al., 2015).
The presence of distributed cortical representations of semantic information, linked to the
notion of “spokes,” as previously defined, grounds its evidence in longstanding research
focusing on object–concept representation in the brain and how the semantic category to which
the object belongs can affect its cortical representation. There is now compelling evidence that
conceptual knowledge can be selectively impaired following focal brain lesions (Capitani
et coll., 2003; Caramazza & Mahon, 2003; Miceli et al., 2000; Warrington & Shallice, 1984)
and that the neural responses of discrete brain regions are more sensitive to specific semantic
Neurobiology of Language
355
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
un
_
0
0
0
3
9
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
catégories (Caramazza & Shelton, 1998; Kuhnke et al., 2020; Mahon et al., 2009; Mahon &
Caramazza, 2011; Martine, 2016; Noppeney et al., 2006). This cortical category selectivity, à
least for people and places, has been shown to persist, both considering specific, unique en-
tities (people: “Leonardo DiCaprio”; lieux: “The colosseum”; Fairhall et al., 2014) and gen-
eral semantic knowledge (“kind of”—people: “lawyer”; lieux: “courthouse”; Fairhall &
Caramazza, 2013un), thus demonstrating an additional category-specific sensitivity for concepts
abstracted from their principal sensorimotor counterparts. Like object category, accessed con-
tent also selectively recruits specialised conceptual representations. When atypical informa-
tion is accessed about people or food, such as the geographical provenance of the item,
regions classically associated with place selectivity are recruited (Fairhall, 2020).
Semantic category sensitivity in the human brain has also been highlighted through data-
driven approaches, by assessing brain responses to naturally spoken narrative stories (Deniz
et coll., 2019; Huth et al., 2016), further mapping single word-related semantic selectivity on the
whole brain surface (see also Pereira et al., 2018).
While crucial for the understanding of conceptual knowledge, single concepts do not
capture the complexities of the semantic contents we manage in everyday life (Frankland &
Vert, 2020). The capacity to flexibly combine multiple concepts from distinct categories
into unitary representations is a fundamental feature of human semantic cognition. The present
study focuses on the cortical mechanism underlying the formation of higher-order meaning
that necessitates the combination of concepts belonging to different semantic domains, spe-
cifically those concepts that are frequently represented across distinct category-selective brain
régions. To this end, we implemented an event-related fMRI paradigm presenting written sen-
tences regarding a single semantic category (c'est à dire., Personnes, Places, Food, Objects, and Animals)
or sentences encompassing two distinct conceptual domains (par exemple., People and Food). Our ob-
jective was to answer two main questions: (1) Are brain regions differentially activated when
information from different semantic categories has to be combined? (2) Do higher-order se-
mantic representations that combine concepts across category continue to rely on category-
specific representations, or do these more derived combinatorial semantic meanings rely more
heavily on general semantic representations?
MATERIALS AND METHODS
Participants
Twenty-eight native Italian speakers were recruited for the study. Three participants were ex-
cluded due to head motion exceeding 2 mm during scanning. Another participant did not
perform the entire protocol and therefore was excluded. Ainsi, the final sample consisted of
24 participants (12 males, mean age 24.9 années). Before entering the scanner, participants
underwent a medical interview with a neurologist, and all of them reported no history of neu-
rological or psychiatric disease. Participants gave informed consent and were compensated
for participation (15 A/ hour). The study was conducted in line with the declaration of
Helsinki (1964, amended in 2013) and was approved by the Ethical Committee of the
University of Trento.
Experimental Design
Stimuli
The stimuli set was composed of 288 Italian written sentences formed by a subject, a verb, et
a complement. Sentences were of three types. (je) Single-category sentences (mean number of
Neurobiology of Language
356
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
un
_
0
0
0
3
9
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
words: 5.79 (0.67)): sentences in which both the subject and the complement belong to the
same semantic category [Personnes, Places, Food, Objects, Animals]. (ii) Combined-category sen-
tences (mean number of words: 5.59 (0.60)): sentences in which the subject and the comple-
ment belong to different semantic categories. Il y avait 10 combinations of the five main
conceptual categories [Personnes & Places, Personnes & Food, Personnes & Objects, Personnes &
Animals, Places & Food, Places & Objects, Places & Animals, Food & Objects, Food &
Animals, Objects & Animals]. (iii) Bizarre sentences (mean number of words: 5.6 (0.68)): gram-
matically correct sentences but with an anomalous semantic meaning (par exemple., “The window was
inside the tomato”). Representative sentences (and the English translation) for each sentence
type are presented in Table 1 (the full list can be found in Supplementary Table S3; supporting
information can be found online at https://www.mitpressjournals.org/doi/suppl/10.1162/nol_a
_00039). Il y avait au total 16 sentences for each of the single- and combined-category
conditions, et 48 sentences for the bizarre condition.
To assess the imageability of different sentences, 10 participants (who did not take part in
the main experiment) rated the imageability of each sentence on a 5-point Likert scale.
Globally, the sentences were perceived as highly imaginable (mean = 4.17). Imageability rat-
ings were averaged within each condition and statistical analysis performed across partici-
pants. A paired sample t test between the average ratings for single-category (4.105) et
combined-category sentences (4.197), revealed a subtle difference (0.092, t(9) = 2.02, p =
0.0371). Comparatively, variation across sentence types was more pronounced over condi-
tions within the 5 single-category conditions (range: 3.969–4.375) and within the 10
combined-category conditions (range: 3.769–4.456).
Sentences were matched on the proportion of action to state verbs in the single (71%) et
combined category sentences (80%, z = 1.6, p = 0.12; normal approximation to the binomial).
We additionally considered the sociality of verbs. Verbal phrases were presented in isolation
and labelled by two raters (S.U. and G.R.) according to the criteria: “likely to relate to an in-
teraction between two individuals.” Social verbs occurred more frequently in single-category
phrases (15%) compared to combined-category sentences (5.6%). This was primarily driven
by an over-representation of social verbs in the person–person condition, where they were
present in 12 out of 16 phrases.
fMRI experimental task
The fMRI session was divided into six experimental runs. In each run, there were 8 trials for
each single- and combined-category sentence (5 single and 10 combined), 24 trials with bi-
zarre sentences, et un supplémentaire 24 fixation-cross null-events. In an event-related paradigm,
sentences were pseudo-randomized across runs and randomly interleaved with fixation cross
events (sentences were repeated three times across the experiment). Using MATLAB (www
.mathworks.com) and Psychophysics Toolbox Version 3 (psychtoolbox.org), each sentence
was presented in black font against a gray background and presented consecutively in three
fragments (sujet, verb, complement). Each trial lasted 2.5 s. Each sentence-fragment was
presented consecutively on the centre of the screen for 400 ms. After the 1.2 s of stimulus
presentation, a black fixation cross appeared in the centre of the screen for the remainder
du procès. The participant’s task was to indicate via button-press if the sentence was seman-
tically meaningful (index finger) or was “bizarre” (right middle). Reaction times (RTs) were cal-
culated from the onset of the last sentence fragment, and responses faster than 400 ms or
slower than 1,700 ms were excluded. RT data for one participant was unavailable due to mea-
surement error.
Neurobiology of Language
357
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
un
_
0
0
0
3
9
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
Tableau 1.
Representative stimuli
Single
Sentence category
Personnes
Representative sentence
I poliziotti arrestano i ladri
Places
Food
The cops arrest the thieves
Le fabbriche sono fuori dalle cittá
Factories are outside cities
I bigné sono ripieni di cioccolata
Cream puffs are filled with chocolate
Objects
Le lampade illuminano i tavoli
Animals
I cani rincorrono le lepri
Lamps illuminate tables
Dogs chase hares
Combined
Personnes & Places
Gli studenti si trovano all’universitá
Personnes & Food
Le mamme cuociono le crostate
Students are at the university
Mothers bake pies
Personnes & Objects
I camerieri sistemano le forchette
Personnes & Animals
I cacciatori sparano ai cervi
Waiters arrange forks
Hunters shoot deer
Places & Food
Le pere crescono nel frutteto
Pears grow in the orchard
Places & Objects
Le forbici si vendono al supermercato
Scissors are sold at the supermarket
Places & Animals
I delfini saltano nel mare
Dolphins jump in the sea
Food & Objects
I caffé si bevono nelle tazzine
Coffees are drunk in cups
Food & Animals
I gatti bevono sempre il latte
Objects & Animals
I topi evitano le trappole
Cats always drink milk
Mice avoid traps
La finestra era dentro al pomodoro
Window was inside the tomato
Bizarre
Neurobiology of Language
358
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
un
_
0
0
0
3
9
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
Post-scanner test
After the fMRI session, participants were again presented the meaningful sentences they had
read in the fMRI session. In the task, a part of the sentence (subject/complement) was missing,
and participants were instructed to complete the missing part.
MRI Scanning Parameters
00
Functional and structural data were collected with a Prisma 3T scanner (Siemens AG,
Erlangen, Allemagne) at the Centre for Mind/ Brain Sciences (CIMeC) of the University of
Trento. Participants lay in the scanner and viewed the visual stimuli through a mirror system
connecté à un 42
, MR-compatible Nordic NeuroLab LCD monitor positioned at the back of
the magnet bore. Data collection was performed using a 64-channel head coil. Functional
images were acquired using echo planar imaging (EPI) T2*-weighted scans. Acquisition
parameters were: repetition time (TR) de 2 s, an echo time (TE) de 28 ms, a flip angle of 75°,
a field of view (FoV) de 100 mm, and a matrix size of 100 × 100. Total functional acquisition
consisted of 1,266 volumes for the six experimental runs, each of 78 tranches axiales (lequel
covered the whole brain) with a thickness of 2 mm and a gap of 2 mm, AC/PC aligned.
High-resolution (1 × 1 × 1 × mm) T1-weighted MPRAGE sequences were also collected (sagittal
slice orientation, centric phase encoding, image matrix = 288 × 288, FoV = 288 mm, 208 slices
with 1-mm thickness, TR = 2,290, LE = 2.74, inversion time (TI) = 950 ms, 12° flip angle).
fMRI Data Analysis
Data were analysed and preprocessed with SPM12 (https://www.fil.ion.ucl.ac.uk/spm/). Le
first four volumes of each run were dummy scans. All images were corrected for head move-
ment. Functional images were normalized to the Montreal Neurological Institute (MNI) T1
espace, resampled to a voxel size of 2 × 2 × 2 mm and spatially smoothed with 6-mm
FWHM kernel. Subject-specific parameter estimates (β weights) for each of the 16 conditions
(see Experimental Design section for details) were derived through a general linear model
(GLM) and a more lenient implicit mask for inclusion in the GLM (0.1 instead of the SPM de-
fault of 0.8). The control condition with a fixation cross formed the implicit baseline. The six
head-motion parameters were included as additional regressors of no interest.
Region of Interest Selection
Region of interest (ROI) analysis was performed within category-selective ROIs defined using
an Omnibus ANOVA to highlight cortical regions showing a differential response across
categories for the single category sentences only. ROIs were defined as the intersection
between a sphere of 5-mm radius around the group peak coordinates, and the activation map
for the Omnibus ANOVA thresholded at p < 0.001. The location of ROIs is indicated in
Supplementary Table S1.
Voxel:
A three-dimensional pixel produced
by volumetric imaging techniques
such as fMRI.
Region of interest (ROI) analysis:
As an alternative to investigating
experimental voxels over the tens of
thousands of voxels in the brain, ROI
analysis is a more acute and focused
measure that designates a specific
region to investigate and in which to
perform analysis.
Additional Voxel-Wise Multivariate Pattern Analysis
Multivariate pattern analysis (MVPA):
An information-based measure that,
as applied to brain imaging
techniques, tests whether the spatial
pattern of brain activation reliably
distinguishes between two cognitive
states.
A supplementary voxel-wise multivariate pattern analysis (MVPA) was performed within a pre-
cuneus ROI showing a greater response to combined-category than single-category sentences.
In this analysis, the constituent single-category sentences were used to predict patterns pro-
duced by combined-category sentences. Specifically, correlation based MVPA analysis was
performed between pairs of combined category sentences (e.g., (A) people and food sentences
versus (B) place and object sentences) using the summed pattern of responses of the relevant
Neurobiology of Language
359
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
single category sentences (e.g., (C) people+food single category sentences and (D) place+
object single-category sentences). To assess whether category information present for single-
category sentences persisted in the combined-category sentences, correlations between unlike
sentence types (A & D; B & C) were subtracted from like sentence types (A & C; B & D). This
process was repeated 45 times for each of the possible pairwise combinations of the ten
combined-category conditions. One-sample t tests were performed on the resulting values
to allow inference.
RESULTS
Behavioural Results
Reaction times on the meaningfulness judgment did not differ between single-category (mean =
716 ms, SD = 120) and combined-category (mean = 723 ms, SD = 120) sentences (t < 1). A
repeated-measure ANOVA revealed that RTs differed among sentence-category for single-
category sentences (F(4, 92) = 17.2, p < 0.001), and combined-category sentences (F(5.3,
121) = 10.5, p < 0.001; Greenhouse-Geisser corrected). In this way, RT-related effects
will not influence comparisons between single-category and combined-category sentences
but may influence category-selective effects. This will be further discussed in the relevant
sections.
Task compliance was high, with meaningful sentences being judged meaningful 90.5% of
the time and bizarre sentences identified as bizarre 84.2% (16.51) of the time. In the post-
scanner test, participants were able to provide the missing sentence fragment with a high de-
gree of accuracy (mean = 68.56%, SD = 15), further indicating a high level of engagement in
the scanner task.
Combination of Concepts Across Semantic Domains
Our first goal was to determine which brain regions may coordinate the combination of con-
cepts across different semantic domains. To identify brain regions showing an increased fMRI
response when conceptual domains are combined, we compared sentences presenting a com-
bination of conceptual categories (e.g., Places & Animals: “The dog is in the kitchen”) to sen-
tences involving a single conceptual domain (e.g., Animals: “The cat is next to the dog”).
The weighted contrast [Combined-Category sentences > Single-Category sentences] iden-
tified a significant cluster in the precuneus (Extent 1,214 voxels, p < 0.001fwe-cluster, peak =
[−4 −50 28], Figure 1), indicating that the precuneus plays a role in the combination of con-
cepts across categorical domains. To exclude the possibility that a single category was driving
this effect, we repeated the analysis iteratively, excluding one category from both the single-
category and combined-category sentences. The increased response in the precuneus for sen-
tences that combine information across categories persisted when removing animals (Extent:
1,189 voxels, p < 0.001fwe-cluster, peak = [2 −54 18]), people (Extent: 210 voxels, p =
0.002fwe-cluster, peak = [−6 −48 28]), places (Extent: 415 voxels, p < 0.001fwe-cluster, peak =
[−12 −56 20]), food (Extent: 219 voxels, p = 0.002fwe-cluster, peak = [10 −54 40]) and objects
(Extent: 1,525 voxels, p < 0.001fwe-cluster, peak = [−4 −68 32]).
This iterative leave-one-category-out process was also used to assess the potential role of
imageability on activation within the precuneus by recalculating single-/combined-category
differences in imageability based only on the included conditions. On iterations where image-
ability was balanced between single-category and combined-category conditions, when re-
moving Places (difference = 0.02, t < 1) and Objects (difference = 0.04, t < 1), the greater
Neurobiology of Language
360
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
Figure 1. Selective activation related to combinations of semantic concepts compared to concepts
related to a single semantic category. The whole-brain univariate response for the contrast
[Combined-Category sentences > Single-Category sentences] is shown in the figure. A significant
cluster emerged in the precuneus (Extent 1,214 voxels, p < 0.001fwe-cluster, peak = [−4 −50 28]).
response in the precuneus for combined-category sentences is seen to persist, indicating that
imageability does not drive this effect. We performed a second analysis to assess which brain
regions are modulated by sentence imageability over our 15 stimulus types. Using a weighted
contrast, we identified voxels where the response amplitude was predicted by the average im-
ageability rating of the 15 experimental conditions. While we observed positive evidence for
response modulation by imageability in left ATL (−54x −8y −14z; extent: 148 voxels; p =
0.017, cluster-corrected) and vmPFC (−4x 58y −10z; extent: 171 voxels; p = 0.008, cluster-
corrected), evidence was not present in the precuneus. Collectively, these results suggest that,
in the present paradigm, imageability-related processes are not driving the increased fMRI re-
sponse in the precuneus for combined-category compared to single-category sentences. Like
nouns, the sociality of verbs may selectively influence cortical activation and is known to in-
crease the response in the precuneus (Lin et al., 2019). As social verbs are more prevalent in
the single-category condition (see Materials and Methods), such factors cannot account for the
increased response for combined-category sentences.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
/
.
l
Category Selective Semantic Representations
To characterise category-selective semantic responses, we contrasted each single category
with the remaining four single categories (e.g., People > [Places, Food, Objects, Animals],
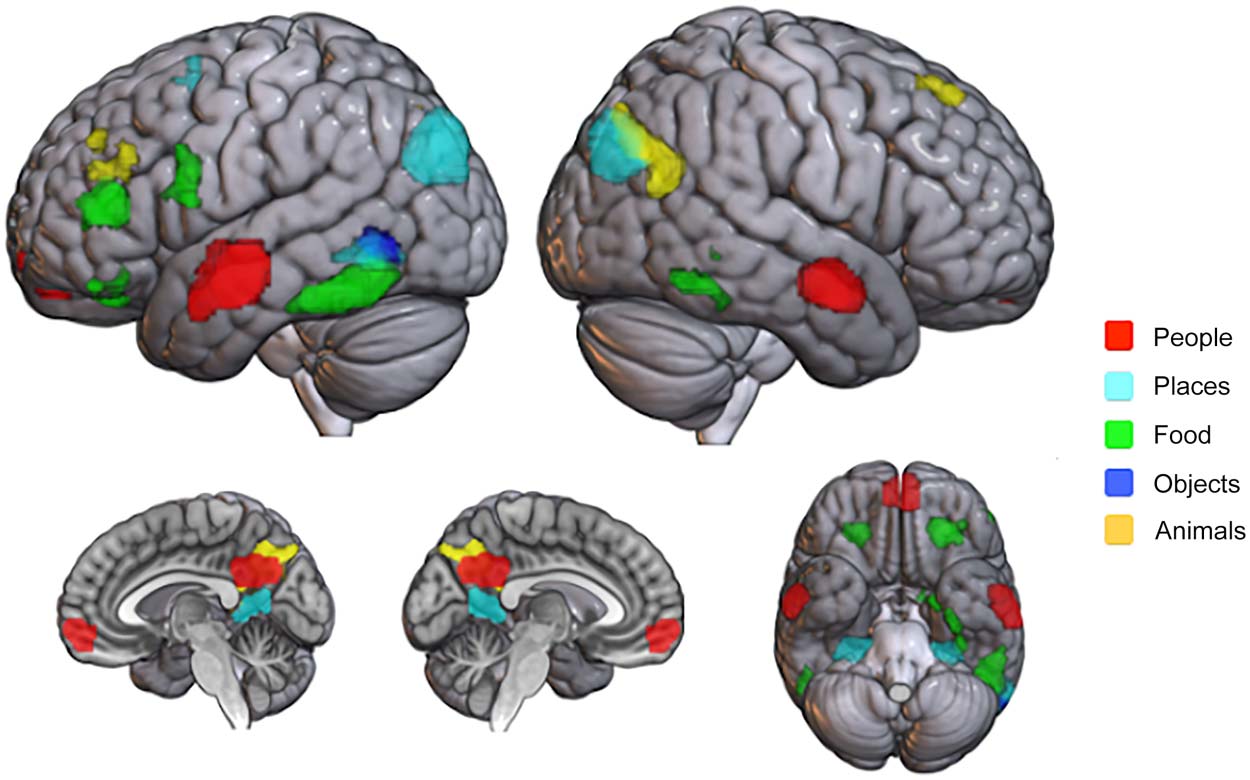
voir la figure 2). We observed strong category-selective fMRI responses for People in ventro-
medial prefrontal cortex (vmPFC), precuneus, and bilateral ATL; for Places in bilateral
para-hippocampal place area (APP), bilateral tranverse occipital sulcus (TOS), bilateral retro-
splenial complex (RSC), left middle temporal gyrus (pMTG), left anterior superior temporal
gyrus (aSTG), and left dorsal superior frontal gyrus (dSFG); for Food in the bilateral orbito-
frontal cortex (OFC), left IFG, left preMotor cortex, left and right posterior inferior temporal
gyrus (pITG), left amygdala, left ventro-temporal cortex ( VTC), and right pITG; for Objects in
the left inferior temporal gyrus (ITG); for Animals in the precuneus, right superior frontal gyrus
(SFG), left dorso-lateral prefrontal cortex (dlPFC), left inferior parietal sulcus (IPS), and right
temporo-parietal junction (TPJ). The results of the category-selective contrasts are reported in
Tableau 2.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The Role of Category-Selective Representations in Combined-Category Sentences
Having ascertained the presence of strong category-selective representation for single-category
phrases, we asked whether these regions make a comparable contribution when concepts
are combined across semantic domains. To this end, we assessed whether the activation
Neurobiology of Language
361
Combining concepts across categorical domains
FWE cluster-corrected:
A family-wise error correction to
overcome multiple comparisons
issues in fMRI using Random Field
Theory. This approach considers the
probability of observing a contiguous
cluster of significant voxels,
correcting for multiple comparisons
over the whole brain volume.
Chiffre 2. Category-sensitive activations associated with single-category (Personnes, red; Places, light
blue; Food, vert; Objects, blue; Animals, yellow) phrases. Significant clusters (voxel-wise p <
0.001, uncorrected, FWE cluster-corrected p < 0.05) are shown on the same brain surface map
(standard MNI152, MRIcroGL software, https://www.nitrc.org/projects/mricrogl). Transparency
has been applied to highlight the whole identified clusters. We reported significant clusters for
People in the vmPFC, precuneus, and bilateral ATL; for Places in bilateral PPA, bilateral TOS, bi-
lateral RSC, left MTG, left aSTG, and left dSFG; for Food in the bilateral OFC, left IFG, left preMotor,
left pITG, left amygdala, left VTC, and right pMTG; for Objects in the left MTG; and for Animals in
the precuneus, right SFG, left dlPFC, left IPS, and right TPJ.
patterns across this network produced by single-category sentences predict the neural re-
sponse of the related combined-category sentences, or in other words, whether the response
evoked by a sentence combining two categorical representations can be reconstructed from
the individual contribution of the two categories.
ROIs were defined via an Omnibus ANOVA as isolated brain regions within which activity
varies across single-category sentences without introducing bias towards a particular object
category. Resulting ROIs were consistent with category-selective regions identified in the pre-
ceding section (see Figure S1 and Table S2 for network visualisation and ROI data).
As noted earlier, behavioural difference existed between the categories, which may partially
account for the effects observed here. To ensure that category-selectivity effects were not due to
RT confounds, the persistence of category effects after controlling for RT differences was
assessed. Specifically, within ROIs described in the next section, for each subject, the beta
responses for each category were regressed against the mean RT for each single-category con-
dition. Then, the category-selective contrasts were recomputed on the residuals of this regression
(now with the linear effects of RT removed). Category-selective responses persisted in all regions
( p < 0.001) with the exception of left IPS, left lateral preMotor, vmPFC, and right lateral PFC. As
we cannot be sure of the veracity of the category-selective nature of the responses in these re-
gions, while we report them in Figure 2 and Figure S1, and Table 2 and Table S2, they have not
been included in subsequent analyses.
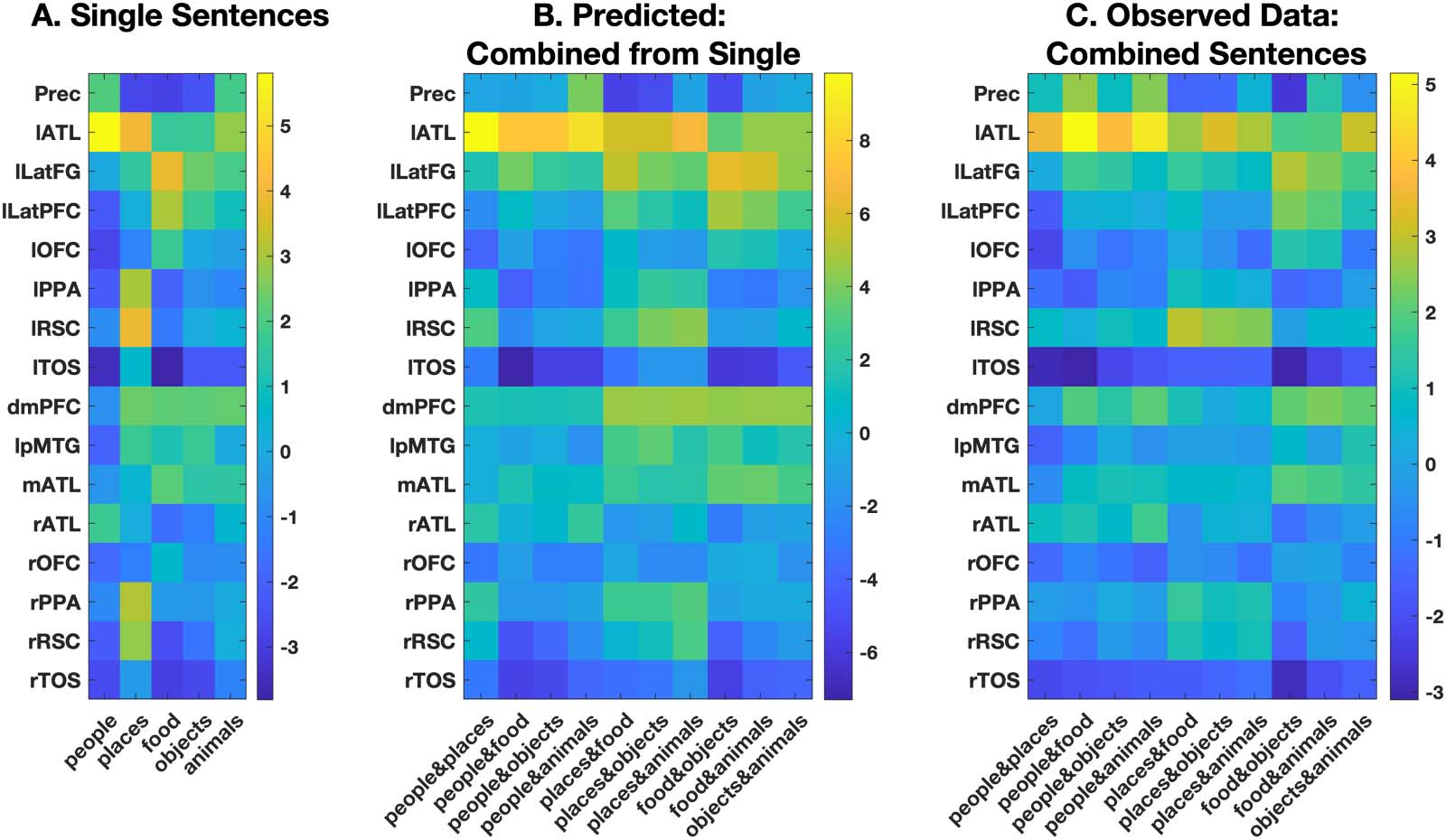
To construct the estimate of the combined-category sentences, within each ROI we took the
response of single-category sentences (Figure 3A) and averaged them to form a prediction of
the amplitude of the regional response for each combined-category sentence (Figure 3B). In
this way, the regional response to sentences involving a person and place (Figure 3B, column 1)
Neurobiology of Language
362
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
Category
People
Places
Food
Objects
Animals
Table 2.
Category-sensitive brain regions
Cluster
Peak
Region
Left ATL
Precuneus
Right ATL
vmPFC
Left PPA
Right RSC
Left RSC
Left TOS
Right PPA
Right TOS
Left pMTG
Left aSTG
Left dSFG
Left OFC
Left pITG
Left IFG
Right OFC
Left preMotor
Left VTC
Left Amygdala
Right pMTG
Left pMTG
Precuneus
Right TPJ
Left IPS
Right SFG
Left dlPFC
p(FWE-cor)
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
0.036
0.002
<0.001
<0.001
<0.001
0.014
<0.001
0.011
0.018
0.014
0.026
<0.001
<0.001
0.001
0.014
0.005
Extent
699
873
297
276
2597
1315
617
872
283
126
222
420
940
495
154
331
161
146
155
136
p(FEW-cor)
<0.001
<0.001
0.001
0.002
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
0.047
0.095
0.141
<0.001
<0.001
<0.001
<0.001
0.010
0.012
0.623
0.742
0.685
1685
<0.001
705
233
155
185
0.020
0.0.381
0.0386
0.0901
t
9.01
7.48
5.87
5.59
15.16
11.15
11.09
12.59
11.69
8.30
5.04
4.86
4.26
8.47
7.75
7.20
6.80
5.39
5.34
4.28
4.18
4.23
7.36
5.24
4.47
4.47
4.03
p(unc)
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
<0.001
Note. Significant clusters are reported separately for each semantic category ( p < 0.05, cluster-corrected).
x, y, z (MNI)
−54, −10, −14
−4, −50, 28
58, −6, −14
−4, 58, −12
−30, −40, −12
10, −54, 10
−10, −54, 8
−38, −84, 32
28, −36, −16
40, −84, 34
−60, −58, −4
−52, −6, −18
−20, 22, 46
−22, 38, −16
−50, −52, −18
−42, 36, 10
22, 38, −18
−44, 6, 24
−36, −28, −24
−20, −2, −24
52, −48, −16
−54, −58, −2
4, −58, 30
48, −62, 20
−26, −56, 38
28, 30, 56
−38, 42, 38
Neurobiology of Language
363
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
Figure 3. Consistency of regional response from single-category to combined-category sentences. Average beta values are shown for each
selected ROI and sentence type. Values obtained from the single-category sentences (A) were averaged to create a prediction vector for the
response to the sentence that combined those categories (B) for comparison against the veridical response to the combined-category sen-
tences (C).
is predicted by the combination of the regional response of sentences about people and the
regional response of sentences about places (Figure 3A, columns 1 and 2).
Overall, the response to the 10 combined-category sentences was closely predicted by
their constituents. This is evident in the high congruence between the patterns evident in
Figure 3B and Figure 3C. While the combined response to single-category sentences did
not predict the response to combined-category sentences in dmPFC (r = −0.008, p = 0.98),
pMTG (r = 0.50, p = 0.14), or left TOS (r = 0.59, p = 0.07), the combined response to
single-category sentences did predict combined-category sentences in the remaining 13 re-
gions. Prediction ability was high, with a mean r value across regions of 0.81 (min, 0.70,
max, 0.92, all p values < 0.05). Thus, the combination of single-category sentences explained,
on average, 66.0% of the observed response to combined-category sentences in these regions.
Importantly, RT differences in the single-category sentences did not predict response pat-
terns in the combined-category sentences. Average RTs from the single categories used to form
the predicted response (i.e., Figure 3B) were correlated with the RTs from the 10 combined-
category sentences. Correlation was non-significant (r = −0.067, p = 0.86), indicating that this
analysis is not contaminated by RT confounds.
To reconcile whole brain analysis showing a greater response for combined-category than
single-category sentences in the precuneus and ROI analysis showing a category-selective
effect in the precuneus, we performed two supplementary analyses. Firstly, we assessed the
relationship between single-category and combined-category sentences in the category-
selective precuneus ROI. The response in the category-selective precuneus ROI for
combined-category sentences is super-additive. Specifically, the response for combined-
category sentences is greater than the summed response to the composite single-category sen-
tences (t(23) = 2.38, p = 0.026). This demonstrates that the category-selective defined
Neurobiology of Language
364
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
precuneus ROI exhibited a greater response for combined- than single-category sentences,
consistent with the whole brain contrast (c.f. Figure 1).
Next, within the precuneus cluster identified in the Combined-Category > Single-Category
whole-brain analysis, we performed a correlation-based MVPA across voxels (see Materials
and Methods). Spécifiquement, the multivoxel pattern for each combined-category condition
(par exemple., People and Places) was reconstructed by combining the single-category sentences of
its constituent categories (par exemple., Single-Category People + Single-Category Places). Alors, recon-
structed patterns were used to distinguish between pairs of combined-category conditions
(par exemple., People and Places vs. Food and Places). The combination of single-category sentences
could accurately predict the combined-category sentences in 36 out of 45 pairwise combina-
tion ( p < 0.05). Of the 9 cases where prediction failed, these were equally likely to occur
when one of the two categories was present in both elements of the pair (6/30) as it was to
occur when no categories were shared across the pair (3/15). These results indicate both the
presence of category-sensitive neurons within this ROI and the consistency of category-
sensitive patterns from single to combined sentences.
DISCUSSION
Semantic knowledge involves not just the representation of single concepts but also the com-
bination of these singular concepts into complex ideas. The construction of these higher-order
units of meaning often requires the combination of concepts arising from different conceptual
domains, which are differentially represented across the cortex. In this work, we asked two
related questions: (1) Do specific brain regions play a particular role in combining concepts
from different domains? (2) Do units of meaning that combine object categories continue to
show a decentralised cortical representation, or are they represented in more centralised
domain-general semantic regions?
To address these questions, we presented participants with meaningful sentences comprising
concepts referring to a single semantic category (e.g., People: “The doctors treat the patients”) or
different semantic categories (e.g., People and Places: “The employees go to the office”). We
found that the precuneus showed an increased response when meaning had to be constructed
across distinct semantic domains and that the distributed representation of conceptual contents
across category-selective regions persists when multidomain, higher-order meaning is
constructed.
Precuneus Activity Increases When Concepts Are Combined Across Domains
The precuneus responded more strongly when sentences involved the combination of concep-
tual domains compared to when sentences involved a single semantic category. This activa-
tion remained significant when individual categories were iteratively removed from the
contrast, indicating that the response is not driven by a specific category-selective response
(Binder et al., 1999; Fairhall & Caramazza, 2013a). Furthermore, using the same univariate
contrast, we did not observe any cortical modulation in other “language” regions associated
with linguistic processing or complexity (Fedorenko, 2014; Santi & Grodzinsky, 2010), indi-
cating that our strategy of matching sentence structure between single-category and
combined-category items was successful and did not strongly influence brain regions associ-
ated with linguistic demand. Taken together, these findings suggest that the precuneus is a
component of the neuronal circuitry involved in the flexible construction of unitary meaning
originating from distinct conceptual domains.
Neurobiology of Language
365
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
Contemporary research has shown that the precuneus plays a dominant role in the general
semantic system, being one of the most widely reported regions responding to semantically
richer stimuli (Binder et al., 2009 for a review). The precuneus is a key element of the default
mode network, an interconnected set of regions involved in internalised cognitive processes
(Buckner et al., 2008; Buckner & DiNicola, 2019; Raichle et al., 2001). In this framework, the
precuneus and posterior cingulate cortex have been described as a central hub of the network,
acting as a convergence zone of distinct functional subsystems (Buckner et al., 2008; Xu et al.,
2016). Concurrently, research has suggested that the precuneus plays a pivotal role in the core
network enabling episodic memory retrieval and prospective imagining (Schacter et al., 2007;
Schacter et al., 2012). It is interesting to note that within episodic memory retrieval the pre-
cuneus appears to play a linking role, flexibly binding together disparate conceptual informa-
tion into meaningful units, a functional role that is notably consistent with the linking of
concepts across domains to form transitory, higher-order semantic representations (for a related
discussion, see Frankland & Greene, 2020).
These observations are not incompatible with previously proposed models, such as the
“convergence zone hypothesis” (Damasio et al., 2004), the “distributed-plus-hub view”
(Patterson et al., 2007), or the classical “hub-and-spoke” model (Rogers et al., 2004).
According to the latter theories, a single hub in the ATL supports transmodal conceptual
representation (Lambon Ralph et al., 2016). In this view, the function of the ATL is to link
modality-specific information into a unitary representation related to a stable, singular,
conceptual representation. The precuneus may play an analogous role, forming transitory
links between discrete concepts from different domains into a higher-order unit of semantic
meaning.
This interpretation is based on the assumption that concepts from two domains are com-
bined when presented within these simple sentences, which is consistent with the automatic
processioning of language (Kutas & Federmeier, 2011). However, it is possible that the precu-
neus response may reflect the presence of two domains not their combination. To exclude this
possibility, it would be necessary to formulate a condition where concepts from different
domains are present but are not integrated, which may not be possible due to the mind’s
tendency to impose sense by linking even inexplicably connected concepts (e.g., “aardvark,”
“cannon”). A potential indicator that concepts are being combined across domains is the
superadditive nature of the precuneus response evident in the independently defined
category-selective ROI—the response to combined sentences is greater than that of the
summed response of the constituent single-category sentences. Such non-linearity of response
has long been seen as a marker for integration in the multisensory literature (Holmes & Spence,
2005) and suggests some interaction beyond the representation of the two concepts in isola-
tion in the present study.
The activation of the precuneus, together with the right AG, has been previously reported
during the combination of noun–noun pairs (Graves et al., 2010). Converging findings from
healthy adults and patients with neurodegenerative diseases (Price et al., 2015; Price et al.,
2016) also support the role of the left AG in the integration of different concepts units
(adjective–noun pairs) into meaningful combinations. While the general aim of these studies
is consistent with the present work, here we adopted a paradigm with richer sentence stimuli
(subjective, verb, complement) that was grounded in the combination of domain-specific
concepts embedded in coherent meanings at the sentence level. Thus, while the AG may play
a role in combining words to form more specific meaning (“lake house” or “red ball”), the
precuneus is a potential mechanism by which concepts from different domains are genera-
tively combined into higher-order meaning.
Neurobiology of Language
366
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
Category-Selective Conceptual Representations for Single-Category Sentences
While often reported for image processing in ventral and dorsal streams, category-selective
responses are less common for word stimuli (Bi et al., 2016). Here, using sentences depicting
specific semantic categories, we observe robust category-selective responses for People,
Places, Food and Animals.
Consistent with previous research, person-selective representations were seen in the precu-
neus and vmPFC, (Fairhall et al., 2014; Fairhall & Caramazza, 2013a; Leibenluft et al., 2004;
Wang et al., 2016) as well as lateral ATL (Fairhall & Caramazza, 2013a; Grabowski et al.,
2001; Tempini et al., 1998; Wang et al., 2016). Likewise, selective activation of bilateral
PPA, TOS, and RSC for places is highly consistent not only with selectivity during the percep-
tion of places and scenes (Dilks et al., 2011; Epstein et al., 2007), but also in word-meaning
related to places (Bi et al., 2016; Binder et al., 2009; Fairhall et al., 2014; Fairhall &
Caramazza, 2013a), as well as spatially relevant geographic information about non-place ob-
jects such as food or people (Fairhall, 2020). Food-selective responses were found in bilateral
OFC, consistent with previous literature investigating neural responses to food-related pictures
(García-García et al., 2013; Killgore et al., 2003; Simmons et al., 2005), potentially reflecting
the role of this region in processing the reward value (Mainen & Kepecs, 2009). Selective re-
sponses in VTC and amygdala have been previously reported in response to food pictures in
relation to different motivational contexts (pre-/post-meal) in children and adolescents (Holsen
et al., 2005). We did not observe a significant response in the left insula, which is frequently
reported with studies using pictures (van der Laan et al., 2011, for a meta analysis). A food-
selective response in the insula has been reported for word stimuli when participants access
taste knowledge but the generalisation of this response to non-taste-related conceptual infor-
mation is subtle and persists only at the voxel-level pattern (Fairhall, 2020). Animals selectively
activated the precuneus, consistent with previous studies presenting participants with spoken
names of animals (Binder et al., 1999). Responses in left IPS and right TPJ have been similarly
reported by eliciting mental pictures of animals through spoken words (Lambert et al., 2002).
We saw category selectivity for objects (here as human-made concrete items including manip-
ulable objects) restricted to the pMTG, a region previously known to exhibit tool selectivity for
word stimuli (Noppeney et al., 2006; Peelen et al., 2013).
Combined-Category Representations Continue to Rely on Category-Selective Representations
Previous work investigating the combination of words has emphasised centralised, default
mode, semantic systems (Graves et al., 2010; Palliera et al., 2011; for a discussion, see
Frankland & Greene, 2020), elements of which are known to contain representations of dif-
ferent categories of objects (Bruffaerts et al., 2013; Devereux et al., 2013; Fairhall &
Caramazza, 2013b; Liuzzi et al., 2020). To address whether the combination of concepts
across domains continues to rely on distributed category-selective regions or is rather centra-
lised into domain-general semantic systems, we compared the representation based upon
single-category sentences to those of combined-category sentences across domain-sensitive
ROIs. Specifically, we used the response evoked by the single-category sentences (e.g.,
People or Food) to predict the response to the relevant combined-category sentence (e.g.,
People and Food). The high consistency between the observed regional pattern of combined-
category sentences and that predicted by combining the patterns of the relevant single-category
sentences indicates that category-sensitive regions respond similarly when single-domain-
specific information is processed and when a combination of domain-specific concepts
are processed. Thus, both during the formation of complex ideas from single categories
Neurobiology of Language
367
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
and in the combination of concepts across domains, distributed category-selective semantic
representations continue to play a role in concept representation. This underscores the impor-
tance of distributed category-selective semantic representations both during the formation of
complex ideas from single categories and in the combination of concepts across domains.
Conclusions
Domain-specific concepts are a fundamental building block of our semantic cognition. At the
same time, our cognitive system is constantly faced with the challenge of binding together
distinct, category-selective semantic information in order to create the higher-order unitary
meanings that allow flexible knowledge manipulation. In this work, we provide partial insight
into how the human brain combines concepts into complex ideas. Our results suggest that the
precuneus plays an important role in this regard, acting on diverse domain-specific semantic
concepts across their respective neural representations and thus representing an important
functional node of the human semantic system. Concurrently, the present findings showing
highly comparable responses in category-sensitive regions, when both single and multiple
domain-specific concepts are processed, indicates the persistence of decentralised represen-
tations of conceptual knowledge when derived information combining concepts from multiple
categories are formed. Collectively, these results show the importance of category-selective
representations in the formation of higher-order semantic representations and the potential role
of the precuneus in binding these together.
FUNDING INFORMATION
Scott Fairhall, H2020 European Research Council (StG), Award ID: 640594.
AUTHOR CONTRIBUTIONS
Silvia Ubaldi*: Investigation: Lead; Project administration: Lead; Data curation: Equal; Formal
analysis: Equal; Project administration: Lead. Giuseppe Rabini*: Data curation: Equal; Formal
analysis: Equal; Writing – original draft: Equal; Writing – review & editing: Equal. Scott L.
Fairhall: Conceptualization: Lead; Formal analysis: Equal; Writing – original draft: Equal;
Writing – review & editing: Equal; Supervision: Lead; Funding acquisition: Lead. [*These
authors contributed equally to the research.]
REFERENCES
Barsalou, L. W. (2010). Grounded cognition: Past, present, and
future. Topics in Cognitive Science, 2, 716–724. https://doi
.org/10.1111/j.1756-8765.2010.01115.x, PubMed: 25164052
Bi, Y., Wang, X., & Caramazza, A. (2016). Object domain and mo-
dality in the ventral visual pathway. Trends in Cognitive
Sciences, 20, 282–290. https://doi.org/10.1016/j.tics.2016.02
.002, PubMed: 26944219
Binder, J. R. (2016). In defense of abstract conceptual representa-
tions. Psychonomic Bulletin & Review, 23, 1096–1108. https://
doi.org/10.3758/s13423-015-0909-1, PubMed: 27294428
Binder, J. R., & Desai, R. H. (2011). The neurobiology of semantic
memory. Trends in Cognitive Sciences, 11, 527–536. https://doi
.org/10.1016/j.tics.2011.10.001, PubMed: 22001867
Binder, J. R., Desai, R. H., Graves, W. W., & Conant, L. L. (2009).
Where is the semantic system? A critical review and meta-
analysis of 120 functional neuroimaging studies. Cerebral
Cortex, 19, 2767–2796. https://doi.org/10.1093/cercor/bhp055,
PubMed: 19329570
Binder, J. R., Frost, J. A., Hammeke, T. A., Bellgowan, P. S. F., Rao,
S. M., & Cox, R. W. (1999). Conceptual processing during the
conscious resting state: A functional MRI study. Journal of
Cognitive Neuroscience, 11, 80–93. https://doi.org/10.1162
/089892999563265, PubMed: 9950716
Borghesani, V., & Piazza, M. (2017). The neuro-cognitive representa-
tions of symbols: The case of concrete words. Neuropsychologia,
105, 4–17. https://doi.org/10.1016/j.neuropsychologia.2017.06
.026, PubMed: 28648571
Bruffaerts, R., Dupont, P., Peeters, R., De Deyne, S., Storms, G., &
Vandenberghe, R. (2013). Similarity of fMRI activity patterns in
left perirhinal cortex reflects semantic similarity between words.
Journal of Neuroscience, 33, 18597–18607. https://doi.org/10
.1523/JNEUROSCI.1548-13.2013, PubMed: 24259581
Neurobiology of Language
368
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
Buckner, R. L., Andrews-Hanna, J. R., & Schacter, D. L., (2008).
The brain’s default network: Anatomy, function, and relevance
to disease. Annals of the New York Academy of Sciences,
11(24), 1–38. https://doi.org/10.1196/annals.1440.011,
PubMed: 18400922
Buckner, R. L., & DiNicola, L. M. (2019). The brain’s default net-
work: Updated anatomy, physiology and evolving insights.
Nature Reviews Neuroscience, 10, 593–608. https://doi.org/10
.1038/s41583-019-0212-7, PubMed: 31492945
Capitani, E., Laiacona, M., Mahon, B., & Caramazza, A. (2003).
What are the facts of semantic category-specific deficits? A crit-
ical review of the clinical evidence. Cognitive Neuropsychology,
20(3), 213–261. https://doi.org/10.1080/02643290244000266,
PubMed: 20957571
Caramazza, A., & Mahon, B. Z. (2003). The organization of con-
ceptual knowledge: The evidence from category-specific seman-
tic deficits. Trends in Cognitive Sciences, 7, 354–361. https://doi
.org/10.1016/S1364-6613(03)00159-1, PubMed: 12907231
Caramazza, A., & Shelton, J. R. (1998). Domain-specific knowledge
systems in the brain: The animate-inanimate distinction. Journal
of Cognitive Neuroscience, 10, 1–34. https://doi.org/10.1162
/089892998563752, PubMed: 9526080
Chen, L., Lambon Ralph, M. A., & Rogers, T. T. (2017). A unified
model of human semantic knowledge and its disorders. Nature
Human Behaviour, 1, 1–24. https://doi.org/10.1038/s41562-016
-0039, PubMed: 28480333
Damasio, H., Tranel, D., Grabowski, T., Adolphs, R., & Damasio,
A. (2004). Neural systems behind word and concept retrieval.
Cognition, 92, 179–229. https://doi.org/10.1016/j.cognition
.2002.07.001, PubMed: 15037130
Deniz, F., Nunez-Elizalde, A. O., Huth, A. G., & Gallant, J. L.
(2019). The representation of semantic information across human
cerebral cortex during listening versus reading is invariant to
stimulus modality. Journal of Neuroscience, 39, 7722–7736.
https://doi.org/10.1523/ JNEUROSCI.0675-19.2019, PubMed:
31427396
Devereux, B. J., Clarke, A., Marouchos, A., & Tyler, L. K. (2013).
Representational similarity analysis reveals commonalities and
differences in the semantic processing of words and objects.
Journal of Neuroscience, 33, 18906–18916. https://doi.org/10
.1523/JNEUROSCI.3809-13.2013, PubMed: 24285896
Dilks, D. D., Julian, J. B., Kubilius, J., Spelke, E. S., & Kanwisher, N.
(2011). Mirror-image sensitivity and invariance in object and
scene processing pathways. Journal of Neuroscience, 31,
11305–11312. https://doi.org/10.1523/ JNEUROSCI.1935-11
.2011, PubMed: 21813690
Epstein, R. A., Parker, W. E., & Feiler, A. M. (2007). Where am I
now? Distinct roles for parahippocampal and retrosplenial cortices
in place recognition. Journal of Neuroscience, 27, 6141–6149.
https://doi.org/10.1523/ JNEUROSCI.0799-07.2007, PubMed:
17553986
Fairhall, S. L. (2020). Cross recruitment of domain-selective cortical
representations enables flexible semantic knowledge. Journal of
Neuroscience, 40(15), 3096–3103. https://doi.org/10.1523
/JNEUROSCI.2224-19.2020, PubMed: 32152199
Fairhall, S. L., Anzellotti, S., Ubaldi, S., & Caramazza, A. (2014).
Person- and place-selective neural substrates for entity-specific
semantic access. Cerebral Cortex, 24, 1687–1696. https://doi
.org/10.1093/cercor/bht039, PubMed: 23425892
Fairhall, S. L., & Caramazza, A. (2013a). Category-selective neural
substrates for person- and place-related concepts. Cortex, 49,
2748–2757. https://doi.org/10.1016/j.cortex.2013.05.010,
PubMed: 23831433
Fairhall, S. L., & Caramazza, A. (2013b). Brain regions that repre-
sent amodal conceptual knowledge. Journal of Neuroscience,
33(25), 10552–10558. https://doi.org/10.1523/ JNEUROSCI
.0051-13.2013, PubMed: 23785167
Fedorenko, E. (2014). The role of domain-general cognitive control
in language comprehension. Frontiers in Psychology, 5, 335.
https://doi.org/10.3389/fpsyg.2014.00335, PubMed: 24803909
Frankland, S. M., & Greene, J. D. (2020). Concepts and composi-
tionality: In search of the brain’s language of thought. Annual
Review of Psychology, 71, 273–303. https://doi.org/10.1146
/annurev-psych-122216-011829, PubMed: 31550985
Gallese, V. (2005). Embodied simulation: From neurons to phe-
nomenal experience. Phenomenology and the Cognitive
Sciences, 4, 23–48. https://doi.org/10.1007/s11097-005-4737-z
García-García, I., Narberhaus, A., Marqués-Iturria, I., Garolera, M.,
Ra(cid:1)doi, A., Segura, B., Pueyo, R., Ariza, M., & Jurado, M. A.
(2013). Neural responses to visual food cues: Insights from func-
tional magnetic resonance imaging. European Eating Disorders
Review, 21(2), 89–98. https://doi.org/10.1002/erv.2216,
PubMed: 23348964
Grabowski, T. J., Damasio, H., Tranel, D., Ponto, L. L. B., Hichwa,
R. D., & Damasio, A. R. (2001). A role for left temporal pole in
the retrieval of words for unique entities. Human Brain Mapping,
13(4), 199–212. https://doi.org/10.1002/ hbm.1033, PubMed:
11410949
Graves, W. W., Binder, J. R., Desai, R. H., Conant, L. L., &
Seidenberg, M. S. (2010). Neural correlates of implicit and ex-
plicit combinatorial semantic processing. Neuroimage, 53(2),
638–646. https://doi.org/10.1016/j.neuroimage.2010.06.055,
PubMed: 20600969
Holmes, N. P., & Spence, C. (2005). Multisensory integration:
Space, time and superadditivity. Current Biololgy, 15(18),
R762–R764. https://doi.org/10.1016/j.cub.2005.08.058,
PubMed: 16169476
Holsen, L. M., Zarcone, J. R., Thompson, T. I., Brooks, W. M.,
Anderson, M. F., Ahluwalia, J. S., Nollen, N. L., & Savage, C. R.
(2005). Neural mechanisms underlying food motivation in chil-
dren and adolescents. Neuroimage, 7(3), 669–676. https://doi
.org/10.1016/j.neuroimage.2005.04.043, PubMed: 15993629
Huth, A. G., De Heer, W. A., Griffiths, T. L., Theunissen, F. E., &
Jack, L. (2016). Natural speech reveals the semantic maps that
tile human cerebral cortex. Nature, 532, 453–458. https://doi
.org/10.1038/nature17637, PubMed: 27121839
Kiefer, M., & Pulvermüller, F. (2012). Conceptual representations in
mind and brain: Theoretical developments, current evidence and
future directions. Cortex, 48, 805–825. https://doi.org/10.1016/j
.cortex.2011.04.006, PubMed: 21621764
Killgore, W. D. S., Young, A. D., Femia, L. A., Bogorodzki, P.,
Rogowska, J., & Yurgelun-Todd, D. A. (2003). Cortical and lim-
bic activation during viewing of high- versus low-calorie foods.
Neuroimage, 19, 1381–1394. https://doi.org/10.1016/S1053
-8119(03)00191-5, PubMed: 12948696
Kuhnke, P., Kiefer, M., & Hartwigsen, G. (2020). Task-dependent
recruitment of modality-specific and multimodal regions during
conceptual processing. Cerebral Cortex, 30, 3938–3959. https://
doi.org/10.1093/cercor/bhaa010, PubMed: 32219378
Kutas, M., & Federmeier, K. D. (2011). Thirty years and counting:
Finding meaning in the N400 component of the event-related
brain potential (ERP). Annual Review of Psychology, 62, 621–647.
https://doi.org/10.1146/annurev.psych.093008.131123, PubMed:
20809790
Lambert, S., Sampaio, E., Scheiber, C., & Mauss, Y. (2002). Neural
substrates of animal mental imagery: Calcarine sulcus and dorsal
Neurobiology of Language
369
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
pathway involvement—An fMRI study. Brain Research, 924(2),
176–183. https://doi.org/10.1016/S0006-8993(01)03232-2,
PubMed: 11750903
Lambon Ralph, M. A., Jefferies, E., Patterson, K., & Rogers, T. T.
(2016). The neural and computational bases of semantic cogni-
tion. Nature Reviews Neuroscience, 18, 42–55. https://doi.org/10
.1038/nrn.2016.150, PubMed: 27881854
Leibenluft, E., Gobbini, M. I., Harrison, T., & Haxby, J. V. (2004).
Mothers’ neural activation in response to pictures of their chil-
dren and other children. Biological Psychiatry, 56(4), 225–32.
https://doi.org/10.1016/j.biopsych.2004.05.017, PubMed:
15312809
Lin, N., Xu, Y., Wang, X., Yang, H., Du, M., Hua, H., & Li, X.
(2019). Coin, telephone, and handcuffs: Neural correlates of so-
cial knowledge of inanimate objects. Neuropsychologia, 133,
107187. https://doi.org/10.1016/j.neuropsychologia.2019
.107187, PubMed: 31499047
Liuzzi, A. G., Aglinskas, A., & Fairhall, S. L. (2020). General and
feature-based semantic representations in the semantic network.
Scientific Reports, 10, 8931. https://doi.org/10.1038/s41598-020
-65906-0, PubMed: 32488152
Mahon, B. Z., Anzellotti, S., Schwarzbach, J., Zampini, M., &
Caramazza, A. (2009). Category-specific organization in the
human brain does not require visual experience. Neuron, 63,
397–405. https://doi.org/10.1016/j.neuron.2009.07.012,
PubMed: 19679078
Mahon, B. Z., & Caramazza, A. (2008). A critical look at the em-
bodied cognition hypothesis and a new proposal for grounding
conceptual content. Journal of Physiology Paris, 102(1–3), 59–70.
https://doi.org/10.1016/j.jphysparis.2008.03.004, PubMed:
18448316
Mahon, B. Z., & Caramazza, A. (2011). What drives the organiza-
tion of object knowledge in the brain? Trends in Cognitive
Sciences, 15, 97–103. https://doi.org/10.1016/j.tics.2011.01
.004, PubMed: 21317022
Mainen, Z. F., & Kepecs, A. (2009). Neural representation of behav-
ioral outcomes in the orbitofrontal cortex. Current Opinion in
Neurobiology, 19(1), 84–91. https://doi.org/10.1016/j.conb
.2009.03.010, PubMed: 19427193
Martin, A. (2016). GRAPES—Grounding representations in action,
perception, and emotion systems: How object properties and
categories are represented in the human brain. Psychonomic
Bulletin & Review, 23, 979–990. https://doi.org/10.3758
/s13423-015-0842-3, PubMed: 25968087
Miceli, G., Capasso, R., Daniele, A., Esposito, T., Magarelli, M., &
Tomaiuolo, F. (2000). Selective deficit for people’s names follow-
ing left temporal damage: An impairment of domain-specific
conceptual knowledge. Cognitive Neuropsychology, 17, 489–516.
https://doi.org/10.1080/02643290050110629, PubMed:
20945192
Noppeney, U., Price, C. J., Penny, W. D., & Friston, K. J. (2006).
Two distinct neural mechanisms for category-selective re-
sponses. Cerebral Cortex, 16, 437–445. https://doi.org/10.1093
/cercor/bhi123, PubMed: 15944370
Palliera, C., Devauchellea, A. D., & Dehaenea, S. (2011). Cortical
representation of the constituent structure of sentences.
Proceedings of the National Academy of Sciences of the USA,
108(6), 2522–2527. https://doi.org/10.1073/pnas.1018711108,
PubMed: 21224415
Patterson, K., Nestor, P. J., & Rogers, T. T. (2007). Where do you
know what you know? The representation of semantic knowl-
edge in the human brain. Nature Reviews Neuroscience, 8,
976–987. https://doi.org/10.1038/nrn2277, PubMed: 18026167
Peelen, M. V., Bracci, S., Lu, X., He, C., Caramazza, A., & Bi, Y.
(2013). Tool selectivity in left occipitotemporal cortex develops
without vision. Journal of Cognitive Neuroscience, 23(8), 1225–1234.
https://doi.org/10.1162/jocn_a_00411, PubMed: 23647514
Pereira, F., Lou, B., Pritchett, B., Ritter, S., Gershman, S. J.,
Kanwisher, N., Botvinick, M., & Fedorenko, E. (2018). Toward
a universal decoder of linguistic meaning from brain activation.
Nature Communications, 9(1), 963. https://doi.org/10.1038
/s41467-018-03068-4, PubMed: 29511192
Price, A. R., Bonner, M. F., Peelle, J. E., & Grossman, M. (2015).
Converging evidence for the neuroanatomic basis of combinato-
rial semantics in the angular gyrus. Journal of Neuroscience, 35(7),
3276–3284. https://doi.org/10.1523/JNEUROSCI.3446-14.2015,
PubMed: 25698762
Price, A. R., Peelle, J. E., Bonner, M. F., Grossman, M., & Hamilton,
R. H. (2016). Causal evidence for a mechanism of semantic in-
tegration in the angular gyrus as revealed by high-definition
transcranial direct current stimulation. Journal of Neuroscience,
36(13), 3829–3838. https://doi.org/10.1523/ JNEUROSCI.3120
-15.2016, PubMed: 27030767
Raichle, M. E., MacLeod, A. M., Snyder, A. Z., Powers, W. J.,
Gusnard, D. A., & Shulman, G. L. (2001). A default mode of
brain function. Proceedings of the National Academy of
Sciences of the USA, 98(2), 676–682. https://doi.org/10.1073
/pnas.98.2.676, PubMed: 11209064
Rice, G. E., Lambon Ralph, M. A., & Hoffman, P. (2015). The roles
of left versus right anterior temporal lobes in conceptual knowl-
edge: An ALE meta-analysis of 97 functional neuroimaging stud-
ies. Cerebral Cortex, 25, 4374–4391. https://doi.org/10.1093
/cercor/bhv024, PubMed: 25771223
Rogers, T. T., Lambon Ralph, M. A., Garrard, P., Bozeat, S.,
McClelland, J. L., Hodges, J. R., & Patterson, K. (2004).
Structure and deterioration of semantic memory: A neuropsycho-
logical and computational investigation. Psychological Review,
111, 205–235. https://doi.org/10.1037/0033-295X.111.1.205,
PubMed: 14756594
Santi, A., & Grodzinsky, Y. (2010). fMRI adaptation dissociates syntac-
tic complexity dimensions. Neuroimage, 51(4), 1285–93. https://
doi.org/10.1016/j.neuroimage.2010.03.034, PubMed: 20338244
(2007).
Remembering the past to imagine the future: The prospective
brain. Nature Reviews Neuroscience, 8, 657–661. https://doi
.org/10.1038/nrn2213, PubMed: 17700624
Schacter, D. L., Addis, D. R., & Buckner, R. L.
Schacter, D. L., Addis, D. R., Hassabis, D., Martin, V. C., Spreng, R. N.,
& Szpunar, K. K. (2012). The future of memory: Remembering,
imagining, and the brain. Neuron, 76(4), 677–694. https://doi
.org/10.1016/j.neuron.2012.11.001, PubMed: 23177955
Simmons, W. K., Martin, A., & Barsalou, L.W. (2005). Pictures of
appetizing foods activate gustatory cortices for taste and reward.
Cerebral Cortex, 15, 1602–1608. https://doi.org/10.1093/cercor
/bhi038, PubMed: 15703257
Tempini, M. L. G., Price, C. J., Josephs, O., Vandenberghe, R.,
Cappa, S. F., Kapur, N., & Frackowiak, R. S. J. (1998). The neural
systems sustaining face and proper-name processing. Brain, 121,
2103–2118. https://doi.org/10.1093/ brain/121.11.2103,
PubMed: 9827770
van der Laan, L. N., de Ridder, D. T. D., Viergever, M. A., &
Smeets, P. A. M. (2011). The first taste is always with the eyes:
A meta-analysis on the neural correlates of processing visual
food cues. Neuroimage, 55, 296–303. https://doi.org/10.1016/j
.neuroimage.2010.11.055, PubMed: 21111829
Visser, M., Jefferies, E., & Lambon Ralph, M. A. (2010). Semantic
processing in the anterior temporal lobes: A meta-analysis of
Neurobiology of Language
370
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Combining concepts across categorical domains
the functional neuroimaging literature. Journal of Cognitive
Neuroscience, 22, 1083–1094. https://doi.org/10.1162/jocn
.2009.21309, PubMed: 19583477
Wang, X., Peelen, M. V., & Han, Z., Caramazza, A., & Bi Y. (2016).
The role of vision in the neural representation of unique entities.
Neuropsychologia, 87, 144–156. https://doi.org/10.1016/j
.neuropsychologia.2016.05.007, PubMed: 27174518
Warrington, E. K., & Shallice, T. (1984). Category specific semantic
impairments. Brain, 107(Pt 3), 829–854. https://doi.org/10.1093
/brain/107.3.829, PubMed: 6206910
Xu, Y., Lin, Q., Han, Z., He, Y., & Bi, Y. (2016). Intrinsic functional
network architecture of human semantic processing: Modules
and hubs. Neuroimage, 132, 542–555. https://doi.org/10.1016/j
.neuroimage.2016.03.004, PubMed: 26973170
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
3
3
5
4
1
9
2
8
8
9
3
n
o
_
a
_
0
0
0
3
9
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
371