ARTICLE DE RECHERCHE
Coherence Between Brain Activation and Speech
Envelope at Word and Sentence Levels
Showed Age-Related Differences
in Low Frequency Bands
Orsolya B. Kolozsvári1,2
, Weiyong Xu1,2
, Georgia Gerike1,2,3, Tiina Parviainen1,2
,
Lea Nieminen4
, Aude Noiray5
, and Jarmo A. Hämäläinen1,2
1Département de psychologie, University of Jyväskylä, Finlande
2Centre for Interdisciplinary Brain Research (CIBR), University of Jyväskylä, Finlande
3Niilo Mäki Institute, Jyväskylä, Finlande
4Centre for Applied Language Studies, University of Jyväskylä, Finlande
5Laboratory for Oral Language Acquisition (LOLA), University of Potsdam, Allemagne
Mots clés: speech perception, development, magnetoencephalography, speech tracking,
coherence, auditory responses
ABSTRAIT
Speech perception is dynamic and shows changes across development. In parallel, functional
differences in brain development over time have been well documented and these differences
may interact with changes in speech perception during infancy and childhood. Plus loin, là
is evidence that the two hemispheres contribute unequally to speech segmentation at the
sentence and phonemic levels. To disentangle those contributions, we studied the cortical
tracking of various sized units of speech that are crucial for spoken language processing in
enfants (4.7–9.3 years old, N = 34) and adults (N = 19). We measured participants’
magnetoencephalogram (MEG) responses to syllables, words, and sentences, calculated the
coherence between the speech signal and MEG responses at the level of words and sentences,
and further examined auditory evoked responses to syllables. Age-related differences were
found for coherence values at the delta and theta frequency bands. Both frequency bands
showed an effect of stimulus type, although this was attributed to the length of the stimulus and
not the linguistic unit size. There was no difference between hemispheres at the source level
either in coherence values for word or sentence processing or in evoked response to syllables.
Results highlight the importance of the lower frequencies for speech tracking in the brain
across different lexical units. Plus loin, stimulus length affects the speech–brain associations
suggesting methodological approaches should be selected carefully when studying speech
envelope processing at the neural level. Speech tracking in the brain seems decoupled from
more general maturation of the auditory cortex.
INTRODUCTION
Brain structure and function continue to develop into early adulthood, with some evidence for
different trajectories for the left and right hemispheres (Gogtay et al., 2004; Pang & Taylor,
2000; Parviainen et al., 2019). In adults, important functional differences between the left
un accès ouvert
journal
Citation: Kolozsvári, Ô. B., Xu, W.,
Gerike, G., Parviainen, T., Nieminen, L.,
Noiray, UN., & Hämäläinen, J.. UN. (2021).
Coherence between brain activation
and speech envelope at word and
sentence levels showed age-related
differences in low frequency bands.
Neurobiology of Language, 2(2),
226–253. https://est ce que je.org/10.1162
/nol_a_00033
EST CE QUE JE:
https://doi.org/10.1162/nol_a_00033
Informations complémentaires:
https://doi.org/10.1162/nol_a_00033
Reçu: 17 Juillet 2020
Accepté: 17 Février 2021
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Orsolya Beatrix Kolozsvári
orsolya.b.kolozsvari@jyu.fi
Éditeur de manipulation:
David Poeppel
droits d'auteur: © 2021
Massachusetts Institute of Technology.
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence.
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Functional near-infrared
spectroscopy (fNIRS):
Functional neuroimaging technique
using near-infrared spectroscopy
where cerebral hemodynamic
responses are measured.
Coherence:
A value that reflects how similar the
oscillatory activity present in two
signals is.
Event-related potential (ERP):
The brain response to a presented
stimulus, measured using
electroencephalography (EEG).
N1m:
Auditory evoked response related
to N1 response, measured using
magnetoencephalography
and right hemispheres have been demonstrated when processing syllable and phonemic in-
formation (par exemple., Poeppel, 2014). Cependant, little is known about the development of this func-
tional specialization in children. Functional near-infrared spectroscopy (fNIRS) and magnetic
resonance imaging (IRM) have provided evidence for a significant leftward asymmetry for
speech processing that is already present from birth (Dehaene-Lambertz et al., 2002; Pena
et coll., 2003). Drawing on these findings, we used magnetoencephalography (MEG) to examine
how hemispheric specialization is reflected in brain responses to various speech units (sen-
tence, words, syllables) and to uncover whether this specialization differs between children
and adults. To achieve those goals, we combined two experimental approaches: examining
general indices of auditory maturation as reflected in the age-related changes of onset-
réponses (event-related fields [ERF]) to simple speech sounds alongside examination of word
and sentence tracking in different frequency bands, as measured by coherence.
Previously, long lasting maturational effects have often been studied using the event-related
potentials (ERPs) and their magnetic equivalent ERFs to short sounds with EEG and MEG. Le
auditory ERPs in infancy and in the preschool age show prominent P1 and N2 responses,
which as children enter childhood start to become earlier in latency and decrease in ampli-
tude. En plus, P1 and N2 responses are separated by emerging N1 and P2 responses
around the age of 8 à 9 années (Albrecht et al., 2000; Ponton et al., 2000).
Differences in hemispheric maturation rates have also been observed using ERFs. The N1m
patterns measured with MEG were more adult-like in 7- to 8-year-olds in the right hemisphere
than in the left (Parviainen et al., 2019). This suggests fine-grained developmental trajectories
of the different auditory regions with clearly immature patterns of activation in the auditory
cortex around early school age (8 à 9 ans).
While studying the event-related potentials and fields in response to individual phonemes
and syllables is a useful method to investigate the well-known maturational effects of auditory
traitement, auditory information in speech spans across multiple timescales encompassing
phonemes, syllables, words, and phrases. Multi-time resolution models of speech processing
(Ghitza, 2011; Ghitza & Greenberg, 2009; Poeppel, 2003; Poeppel & Assaneo, 2020) propose
that speech information is processed and integrated in a hierarchical and interdependent man-
ner by phase alignment or neural entrainment of the involved oscillatory networks in the au-
ditory cortices with different specialization for the left and right auditory areas.
Coherence analysis can be used to study speech perception in these longer speech seg-
ments. Coherence is the computation of synchrony between two signals in the frequency do-
main. The coherence value reflects the consistency of phase difference between two signals
(here between the speech envelope and brain activity) at any given frequency. This technique
can be used to investigate tracking of the speech signal in the brain, which has been argued to
reflect relevant linguistic operations such as parsing and chunking of hierarchical linguistic
structures of speech (Bourguignon et al., 2013; Ding et al., 2016; Gross et al., 2013; Molinaro
& Lizarazu, 2018; Peelle & Davis, 2012).
Neuronal oscillations in frequency bands present in speech (delta, 1–3 Hz, theta, 4–8 Hz,
beta, 15–30 Hz, and low gamma, 30–50 Hz; Poeppel, 2014) have been theorised to provide a
basis for parsing the continuous speech signal into different linguistic units (par exemple., delta: syllable
stress patterns; theta: syllables; beta: onset-rime units; low gamma: phonetic information;
Ghitza et al., 2013; Leong & Goswami, 2014; Poeppel, 2014; Poeppel & Assaneo, 2020).
In this framework, the linguistic information associated with the different timescales would
be then integrated to give the final speech percept. Low frequency cortical activity appears
to synchronise to the rhythms of multiple linguistic units (Ding et al., 2016, 2017), while higher
Neurobiology of Language
227
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
frequencies (such as beta and gamma) may be more sensitive to syntactic and semantic infor-
mation (Ding et al., 2016). Ensemble, these results suggest that during listening to connected
speech, the brain synchronizes cortical rhythms to track the rhythm of the different linguistic
units (Ding et al., 2017).
Speech processing involves both left and right auditory cortices (Poeppel, 2003; Poeppel &
Assaneo, 2020). In its early stage the representation of the input speech signal has a bilateral
symmetry, which then branches out in subsequent processing steps. Left auditory areas have
been suggested to sample information from short (20–40 ms) integration windows (Giraud
et coll., 2007; Poeppel, 2003; Poeppel & Assaneo, 2020), and right areas to sample informa-
tion from longer (150–200 ms) integration windows (Giraud et al., 2007; Luo & Poeppel,
2007; Poeppel, 2003; Poeppel & Assaneo, 2020). These differences are reflected in oscillatory
neuronal activity in different bands (mostly in gamma and theta bands, respectivement).
Cependant, changes in brain activity have been reported during childhood with respect to
general auditory sound processing as well as more specific speech processing (par exemple., Ríos-
López et al., 2020; Uhlhaas et al., 2010). Developmental changes in neural synchrony have
been demonstrated (for a review, see Uhlhaas et al., 2010) using auditory stimulation (Müller
et coll., 2009), whereby young children showed reduced synchronisation in the delta and theta
(Müller et al., 2009) frequencies compared to adolescents and adults.
There is converging evidence that hemispheric specialisation to different windows of inte-
gration for auditory information and speech is present from the first year of life; cependant, résultats
differ as to which hemisphere shows the strongest response to long speech-like chunks
(Telkemeyer et al., 2009, 2011). The developmental pattern of hemispheric dominance for
processing spoken sentences seems to shift between brain hemispheres with age. Greater
entrainment to speech was found in the left hemisphere compared to right in the theta band with
7-month-old infants (Kalashnikova et al., 2018). Cependant, this specialization was not found in
young children between the ages of 4 et 7 années (Ríos-López et al., 2020) in the delta band.
Enfin, a higher correlation in the right as compared to the left hemisphere between the ampli-
tude envelope of sentences and their corresponding brain responses was found in older 9- à
13-year-old children (Abrams et al., 2008, 2009).
Building on those findings, the current study investigated (un) age-related differences and (b)
hemispheric balance in word and sentence tracking in low frequency bands to separate the
word to phrasal levels of processing. Based on previous studies on adults and older children,
we expected hemispheric differences to already be present in 5- to 9-year-olds in the delta
(1–4 Hz) and theta (4–8 Hz) bands with the right hemisphere showing higher coherence than
the left hemisphere.
To examine if and how the maturation of the synchrony measures is related to the estab-
lished maturation of the onset response (reflected in the changes in ERFs to syllables), nous
compared the coherence values for words and sentences with the age-related changes in
the N1m response to syllables. Evoked brain activity to sounds has been shown to change
from preschool to school age and to adulthood. While the specific N1m response is absent
in early childhood, it seems to emerge at around 8 à 9 years of age and only become fully
mature in adulthood (Albrecht et al., 2000; Ponton et al., 2000). If the N1m amplitude has a
common underlying maturational mechanism with the speech tracking index, our results
should show similar developmental effects. On the other hand, synchronization of brain ac-
tivity to speech could utilize partly separate brain mechanisms that follow a different devel-
opmental trajectory and are affected more by environmental input than by developmental
changes reflected by N1m.
Neurobiology of Language
228
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
We also examined correlations between the processing of speech envelopes and phono-
logical skills. Speech envelope processing has been related to segmentation into syllable and
phoneme level elements (Poeppel, 2014). As for phonological skills, broadly defined they in-
clude the awareness of various speech units (par exemple., phonemes, syllables, words), working mem-
ory operations for speech sounds, and access to phonological representations (par exemple., Fowler,
1991; Goswami & Bryant, 2016; for a review, see Noiray, Popescu et al., 2019). Ceux-ci sont
thought to be represented, Par exemple, by rapid naming, phoneme deletion, and speech rep-
etition tasks. Basé sur ceci, we hypothesized that speech envelope processing could be linked
to phonological skill development (Goswami, 2011).
MATERIALS AND METHODS
Participants
Two age groups participated in the study: typically developing children and young adults. Le
adults were studying at the University of Jyväskylä, Finlande. Tableau 1 shows the number of par-
ticipants, mean age and age-range, genre, handedness, and average hearing level for each
group. All participants were Finnish native speakers.
The children were recruited via the National Registry of Finland and the adults via email
lists of the university. Exclusion criteria at the time of recruitment were head injuries, ADHD or
learning difficulties, neurological diseases and medication affecting the central nervous sys-
tem, or any reported hearing deficits. Children recruited for the study were typically develop-
ing and did not present any neurological, cognitive, or language-related deficiency. Dans
addition, the hearing level of the participants was tested using audiometry, with most of them
performing at or below 25 dBs for 250 Hz, 500 Hz, 1000 Hz, et 2000 Hz sounds in the left
and right ears.
After data collection 13 participants were excluded overall, all of them from the child
group. Five were excluded based on the measurement because of too much movement and
inability to follow instructions during the recording, two because of noisy data, four because of
technical problems (instrumentation failure or software issues), one based on incidental find-
ings during the measurements (based on the neurologist’s report), and one because of high
amplitude fluctuations in the data.
Enrolment in the study was voluntary; all adults and children participants as well as their
parent/caregiver provided written informed consent prior to their participation in the study.
Subsequent to the MEG study, all participants received either a movie ticket or a gift card
Tableau 1. Description of participants
# of participants included in the analysis (measured in MEG)
Mean age (SD)
Age range (Minimum–Maximum; y = years, m = months)
Gender ratio (M.:F)
Handedness (gauche:les deux:droite)
Average hearing level in DBs (gauche:right ear)
Neurobiology of Language
Children
34 (47)
7.53 (1.34)
4y8m–9y4m
18:16
5:1:28
21.25:21.37
Adults
19 (19)
24.80 (3.73)
20y3m–35y2m
2:17
0:1:18
Self-report of normal
hearing level
229
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
as compensation for their participation. Individual structural MR images were acquired from a
private company offering MRI services (Synlab Jyväskylä). T1-weighted 3D-SE images were
collected on a GE 1.5 T (GoldSeal Signa HDxt) MRI scanner using a standard head coil and
with the following parameters: TR/TE = 540/10 ms, flip angle = 90, matrix size = 256 × 256,
slice thickness = 1.2 mm, sagittal orientation.
This study was carried out in accordance with the Declaration of Helsinki and approved by
the Ethical Committee of the University of Jyväskylä, Finlande.
Behavioural Test Battery
D'abord, we conducted a battery of behavioural tests assessing the children’s general cognitive
abilities, with an emphasis on language-related skills. For a description of the behavioural
tests, see Table 2.
Three different age-appropriate tests, WPPSI-III (Wechsler, 2003un), WISC-IV (Wechsler,
2003b), and WAIS-IV (Wechsler, 2008), were used to measure participants’ visuo-spatial rea-
soning and vocabulary, and two tests, WISC-IV and WAIS-IV, were used for working memory.
The motor development of the participants was tested using subtests from the Developmental
Neuropsychological Assessment (NEPSY; Korkman et al., 1998), the oro-motor task, et le
Tableau 2. Description of behavioural test scores
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Behavioural measure
WPPSI-III, WISC-IV,
WAIS-IV
Subtest
Block design
Vocabulary
Digit span
NEPSY
Repetition of nonsense words
NEPSY-II
Oro-motor task
Visuo-motor task
(car and motorcycle)
Mean (SD)
score reported
sp
sp
sp
sp
sp
combined sp
Phonological processing
sp
Repetition of sentences
# correct
Rapid automatized
naming (RAN)
Letter knowledge task
Objects
Letters
Lukilasse
Word reading
Pseudoword list reading
Pseudoword text reading
Lukilasse
Dictation
temps (s)
temps (s)
total
percentile
total
fluency
percentile
Children
10.00 (3.82)
10.96 (3.15)
Adults
11.26 (3.21)
11.36 (3.01)
10.46 (3.07)
13.79 (2.02)
X
9.96 (2.15)
11.26 (2.96)
10.09 (2.43)
10.30 (2.81)
9.42 (3.02)
11.33 (2.33)
25.62 (3.08)
64.25 (13.66)
34.70 (10.64)
22.67 (8.27)
46.38 (40.32)
34.45 (13.98)
99.59 (0.22)
58.10 (39.06)
X
X
X
X
X
34.37 (7.78)
18.90 (4.62)
X
X
X
X
X
Note. sp: standard point; SD: standard deviation; WPPSI-III: Wechsler Preschool and Primary Scale of Intelligence (Wechsler, 2003un); WISC-IV: Wechsler
Intelligence Scale for Children (Wechsler, 2003b); WAIS-IV: Wechsler Adult Intelligence Scale (Wechsler, 2008); NEPSY: Neuropsychological Assessment test
battery I (Korkman et al., 1998); NEPSY II (Korkman et al., 2008); RAN: Rapid automatized naming (Denckla & Rudel, 1976); Lukilasse (Häyrinen et al., 1999).
Neurobiology of Language
230
Age-related differences in coherence
NEPSY II visuo-motor task (Korkman et al., 2008). Participants’ phonological processing was
tested using the NEPSY II subtest. To assess speed of lexical retrieval, the Rapid automatized
naming (RAN; Denckla & Rudel, 1976) Objects and Letters subtests were used. To measure
memory for sentences, the NEPSY II Sentence Repetition subtest was used.
Reading skills were tested using the word reading task from the Lukilasse test battery
(Häyrinen et al., 1999), the pseudoword reading task adapted from TOWRE (Torgesen et al.,
1999), and the pseudoword text reading task (Eklund et al., 2015).
For a detailed description of the behavioural tests, see Supplementary Material 1 (supporting in-
formation can be found online at https://www.mitpressjournals.org/doi/suppl/10.1162/nol_a_00033).
Stimuli
Three types of stimuli characterizing various temporal and linguistic structures were used for
the speech tracking task: syllables, words, and sentences. Syllables varied in consonants’ place
of articulation (moving from front to back: bilabial stop /p/, dental stop /t/, and palatal stop /k/),
while the vowel remained identical (/a/).
Each syllable was presented 18 times (total of 54 syllable presentations), and words starting
with the same syllables (18 words for each syllable, total of 54 words), ainsi que 54 phrases,
each starting with one of the word category stimuli. For a description and exemplars of the
stimuli, see Table 3.
All words were common, everyday nouns. The words were 2 à 3 syllables long. Sentences
were composed of 3 à 4 words and always started with a noun followed by a form of the verb
“to be” in the present tense. Stimuli were chosen with the help of an expert developmental
linguist. The stimuli were produced by a female native Finnish speaker. All stimuli were sep-
arate, unique tokens produced separately. Stimuli were recorded using a 44 kHz sampling
frequency, 32-bit quantisation in a professional recording studio. The sound files were cut into
individual segments using Praat (Boersma & Weenink, 2018).
The same syllables and words were used for each stimulus type to get comparable onset
evoked brain responses. To see the list of stimuli used, see Supplementary Material 2.
Procedure
Experimental design
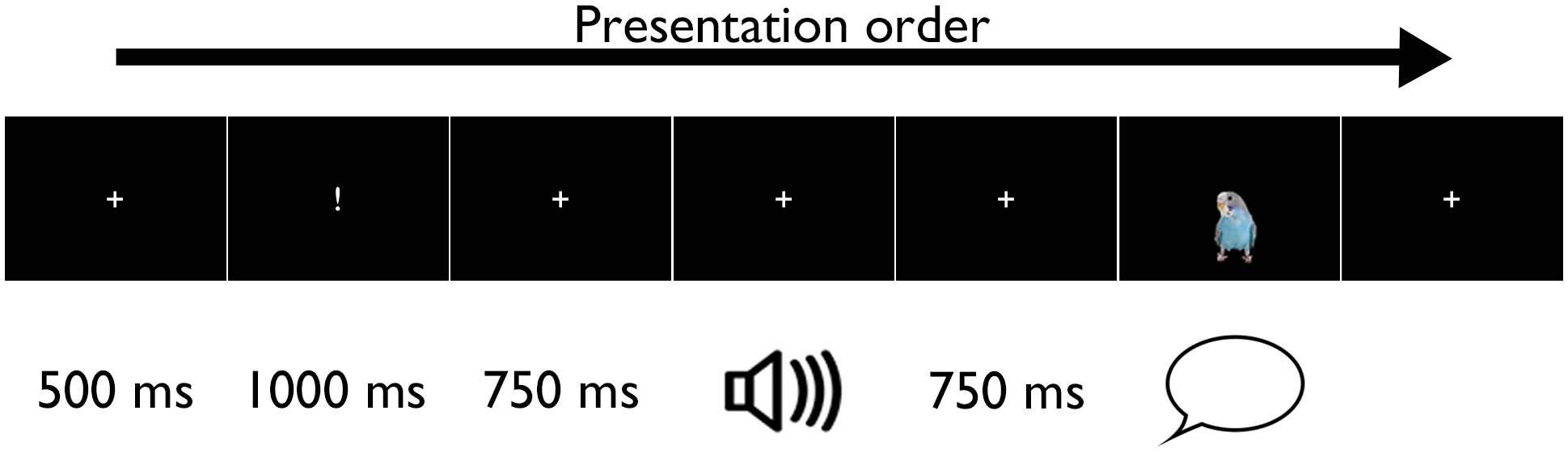
Each speech tracking trial consisted of a fixation cross in the middle of the screen for 500 ms,
then an exclamation mark appeared in the same space for 1,000 ms signalling that a sound is
going to come soon, followed by the fixation cross for 750 ms. The auditory stimuli were then
Tableau 3. Description of stimuli
Stimulus
type
Syllable
Word
Sentence
Average duration
(ms)
209.33
574.54
1,438.54
SD
25.58
103.22
240.49
Range (ms)
185–236
352–797
1,039–2,051
Exemplars: Finnish
English translation
ka, ta, Pennsylvanie
kala, paju, talo
fish, willow, maison
Kala on akvaariossa.
Paju on taipuisa puu.
Talo on aivan uusi.
The fish is in the aquarium.
A willow is a flexible tree.
The house is brand new.
Neurobiology of Language
231
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
presented via earphones, with the fixation cross on the screen. The fixation cross remained on
the screen for 750 ms after the end of sound. This was followed by a still image of a parrot
appearing for 1,250–4,250 ms (presentation duration depended on the type of stimuli heard)
which provided the cue for the participants to repeat the previously heard stimuli aloud (voir
Chiffre 1).
Participants were instructed to first listen to a speech sequence (c'est à dire., a syllable, a word, or a
sentence) and to repeat it after seeing the visual cue on the screen (a parakeet). The visual
stimuli were presented on a black background with white standard characters (a cross for fix-
ation and an exclamation mark alerting to the auditory stimuli) in Times New Roman font and
a font size 64. The bird stimuli were 9 × 15 cm in size on the projection screen. Here only the
time-window of the auditory stimulus presentation was analysed.
Participants were first given instructions and 6 practice trials (2 of each type of stimuli, pre-
sented in random order). In the actual experiment 162 stimuli (le 3 syllables repeated 18
times each, 54 words, et 54 phrases) were presented in random order.
Stimuli were presented in 9 blocks, avec 2 longer breaks after 3 blocks and shorter breaks
(duration determined by the participant) in between the blocks. Three blocks lasted approxi-
mately 8 min, and it took approximately 30 min to complete the task, instructions and practice
included.
The task was embedded in a child-friendly narrative to stimulate children’s attention and
motivation to complete the task. Participants were told they are teaching 3 parrots how to
“speak.” Their task was to wait for the parrot to start listening (when the cue appeared on
the screen) and their instructions included keeping eye-contact with the parrot to make sure
the bird is paying attention (to minimize movement-related artefacts in the recording).
En outre, they were asked to repeat what they heard at a normal speaking loudness
(c'est à dire., to not mumble the syllables, words, or sentences) since the parrots will “not be able to
learn if they don’t hear the speech properly” (to be able to record the production as clearly as
possible). This also ensured the children were fully engaged in the task. Correct production
was on average for children 88.41% and for adults 97.86%. At the end of each third block
(c'est à dire., before the longer breaks and the end of the test), the parrots “repeated” some of the heard
sounds, which were new sounds created by raising the pitch of the original stimuli, to give the
impression that the parrots were the ones repeating them. The first and second time it was the
syllables, while at the end of the MEG recording it was one sentence from each syllable type.
Participants sat in a magnetically shielded, sound attenuated room under the MEG helmet,
at a 68 degree position. The stimuli were presented through insert earphones (Rotel RA-1570
Chiffre 1. Schematic representation of one trial of the experimental paradigm. Data analysis was
focused on the time-window during the stimuli presentation, indicated by the picture of the
loudspeaker.
Neurobiology of Language
232
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
système; eartips were ER3-14B for children and ER3-14A for adults) at a comfortable loudness
level. The participants sat 1 m from the projection screen. During measurement, a research
assistant was also present in the room when necessary for the children. Presentation software
(version 18.1; Neurobehavioral Systems, Inc., Albany, Californie, Etats-Unis) was used to present the stim-
uli, running on a Microsoft Windows computer (sound card: Sound Blaster Audigy RX; video
card: NVIDIA Quadro K5200). Measurements were video monitored to make sure participants
were paying attention and doing the task.
MEG recording
306-channel (102 magnetometers and 102 planar gradiometer pairs) MEG data were recorded
in a magnetically shielded room using the Elekta Neuromag® TRIUX™ system (Elekta AB,
Stockholm, Sweden) at the Centre for Interdisciplinary Brain Research, at the University of
Jyväskylä, Finlande.
The head position in relation to the sensors in the helmet was monitored continuously with
five digitised head position indicator (HPI) coils attached to the scalp. Three HPI coils were
placed on the forehead and one behind each ear. The position of the HPI coils was determined
by three anatomic landmarks (nasion, left and right preauricular points) using the Polhemus
Isotrak digital tracker system (Polhemus, Colchester, VT) at the beginning of the recording. Un
additional set of points (>100) randomly distributed over the scalp was also digitised. Electro-
oculogram was recorded with two electrodes attached diagonally close to the left and right
eyes and one ground electrode attached to the collar bone.
The sampling rate of the recording was 1000 Hz and a 0.03–330 Hz online band-pass filter
was used.
Data Analysis
Pre-processing
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
All data were pre-processed using the temporal extension of the signal space separation method
with buffers of 30 s (Taulu & Kajola, 2005; Taulu et al., 2005) in Maxfilter 2.2™ (Elekta AB)
to remove external interference and correct for head movements. Bad channels were identified
and reconstructed by the Maxfilter program. Head position was estimated in 200 ms time-
windows and 10 ms steps for movement compensation. Data were saved in three separate
files containing three recording blocks. Initial head position of the first file was used for trans-
forming the head position to the same position across the files.
Data were pre-processed using independent component analysis (ICA) using fastICA algo-
rithm (Hyvärinen & Oja, 2000) to remove eye blinks, horizontal eye movements, and cardiac
artifacts in MNE Python (0.16.2; Gramfort et al., 2013), and the separate MEG recordings were
concatenated. The rest of the data analysis was done in the FieldTrip toolbox (Oostenveld et al.,
2011) in MATLAB R2016 (https://www.mathworks.com/).
The continuous MEG recording was epoched to 100 ms before and 1,000 ms after the onset
of sound in the syllable stimuli (for analysis of the evoked fields), et 100 ms before the onset of
sound and 100 ms after the end of the sound in the word and sentence stimuli (for analysis of
the frequency contents). Epochs were visually inspected and bad trials were rejected, with an
average of 2.18% of epochs rejected for the children and 0.78% of epochs rejected for the
adultes. Data were low-pass filtered at 45 Hz. The epoched data was baseline corrected using
le 100 ms preceding the onset of the stimuli.
Neurobiology of Language
233
Age-related differences in coherence
We examined the data using two approaches. D'abord, to examine how closely the brain fol-
lows the frequency contents of the speech signal, coherence was calculated between the MEG
signal and the speech signal. Deuxième, the evoked fields to the syllable stimuli were calculated
to examine possible associations between the relatively well-known developmental changes
of the evoked fields (particularly responses around 100 ms) and the coherence measures.
Coherence measures
We conducted coherence analysis at different frequency bands to investigate how brain activity
changes while tracking the speech envelope of stimuli with different durations at different ages.
The speech stimuli were downsampled to 1000 Hz from 44.1 kHz. The absolute hilbert
envelope was calculated for each stimulus separately in MATLAB (abs(hilbert(audiosignal))).
The envelope was then appended to the epoched MEG data as a 307th channel.
Earlier studies looking into cross-correlations between the speech envelope and brain ac-
tivity removed the first 250 ms of brain activity to avoid the onset evoked response (par exemple.,
Abrams et al., 2008). Cependant, the effects of the onset response on the coherence measures
have not been reported before. Donc, we performed the coherence analyses two times:
d'abord, for data without the evoked response, second, for the whole epoch length (voir
Supplementary Material 3). As shown in Supplementary Material 3, this did not have a large
effect on the results. The results reported in the main text are based on analysis conducted
using data where the evoked response was removed.
Frequency analysis of the data was done to compute the cross and power spectra of the
trials using a multitaper frequency transformation method, where the maximum trial length
was rounded up to the next power of 2 (cfg.pad = nextpow2) using FieldTrip’s ft_freqanalysis
fonction, entre 1 et 45 Hz with a 3 Hz smoothing and keeping the trials. This was
followed by coherence analysis between the sound envelope and the MEG data using the
ft_connectivityanalysis function.
Plus loin, to see if the coherence between the brain and speech signals was significant at the
individual level, we calculated 1,000 permutations of coherence, where the sound envelopes
were randomly paired with the brain activity of another sound envelope, then compared with
the original coherence value. For each participant at least one channel of the original speech–
brain pair showed a coherence value larger than 95% of the permuted values (for visualiza-
tion, see Supplementary Material 4).
To examine the effect of the stimulus length on the coherence values, we first checked the
lengths of trials for word stimuli. Deuxième, we cut out the end of the sentence stimuli to be of
equal length with the word stimuli (c'est à dire., the initial part of the sentence was used in the new
analyse). We then recalculated the coherence between these shortened sentence stimuli and
brain activity (see Supplementary Material 5). The results showed that shortened trials also had
larger coherence values in both frequencies.
For further analyses, channels were grouped together by hemispheres (see Supplementary
Material 6 for grouping of sensors across hemispheres). In the statistical analysis, data from
magnetometers were averaged based on hemispheres and separated into two frequency
bands: 1–3.5 Hz (delta), 4.5–8 Hz (theta).
For children, source reconstruction was based on their own T1 MRIs, while for adults the
fsaverage brain template from Freesurfer (RRID: SCR_001847; Martinos Center for Biomedical
Imagerie, Charlestown, MA, Etats-Unis) was used. Coregistration was done between the digitized
head points and the brain template with 3-parameter scaling.
Neurobiology of Language
234
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
.
/
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Global mean field power (GMFP):
A measure used to characterize
global MEG activity.
Source analysis was done using the ft_sourceanalysis, using the dynamic imaging of coher-
ent sources method (Gross et al., 2001) between 1–8 Hz for every 0.5 Hz. The resulting
coherence values were then averaged together according to the frequency band defined—
delta band: 1–3.5 Hz, theta band: 4.5–8 Hz. The coherence values were then extracted based
on the Desikan-Killiany Atlas (Desikan et al., 2006). Two regions of interest (ROI) were se-
lected a priori: the temporal area, including the superior temporal, transverse gyrus, and bank
of superior temporal sulcus areas; and the inferior frontal area, including the pars opercularis,
pars orbitalis, pars triangularis, and precentral areas (voir, par exemple., Molinaro et al., 2016).
Identification of responses around 100 ms to syllable stimuli and correlation with coherence values for
the word and sentence stimuli
Trials for syllables were averaged together for each participant separately. Global mean field
pouvoir (GMFP) was calculated for each group separately, and the time-window of auditory
response was identified. Based on the GMFP peaks, the time-windows were defined by auto-
matically finding the peak near 100 ms, and using a time-window of +/−25 ms for each hemi-
sphere and group. Ainsi, the time-windows used in further analyses were 94–144 ms in the left
hemisphere and 92–142 ms in the right hemisphere for adults, and 114–164 ms in the left and
113–163 ms in the right hemisphere for children. We averaged together the squared values
from the temporal channels from the two hemispheres separately. The values were then cor-
related with the coherence values in the left and right hemispheres.
Topography of the averages was visually inspected to confirm the correct N1m response pattern or
its equivalent in children. Earlier ERP/ERF research has shown that the N1m pattern reflects current
direction towards inferior-posterior direction, and the opposite direction was referred to as
P1m/P1m-like response. En effet, averaging or grouping together opposite field patterns would
obscure the outcome, and these patterns are likely to reflect distinct processes. Responses
were separated based on hemisphere, then squared. The squared amplitude of the response
was then correlated with the coherence values from the left and right hemispheres for the delta
and theta bands.
A missing response could be due to noisy ERF signal. Donc, signal-to-noise ratio was
calculated by averaging and squaring together the baseline periods of the ERFs (time-window:
−100–0 ms), and used as a covariate in separate ANOVAs to ensure that it was not the source
of the differences found at sensor level. We found that it did not affect the significant effects.
Source analysis of the ERFs was done using ft_sourceanalysis, using the minimum-norm
estimate (MNE) method (Hämäläinen & Ilmoniemi, 1994), and the power of each source com-
ponent was calculated using ft_sourcedescriptives and used in the statistical analyses.
MNE source estimates were calculated for ERFs, and source power waveforms were extracted
based on the Desikan-Killiany Atlas (Desikan et al., 2006). One ROI was selected a priori from the
temporal areas around the auditory cortex including the temporal area, including the superior
temporal, transverse gyrus, and bank of superior temporal sulcus areas, postcentral and supramar-
ginal areas. The same time-windows were used as in the sensor level analysis. The literature
clearly defines the sources of the N1m response near auditory cortex (Parviainen et al., 2019;
Ponton et al., 2002). The ROIs for the coherence value analysis and ERFs were therefore expected
to be slightly different with the former encompassing more frontal regions (Molinaro et al., 2016).
Statistical analyses
The age, hemisphere, and stimulus type effect on the coherence values for the different fre-
quency bands were analysed in SPSS (IBM SPSS Statistics v. 24) using a 2 (Type: Word,
Neurobiology of Language
235
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Sentence) × 2 (Hemisphere: Gauche, Droite) × 2 (Groupe: Children, Adults) repeated measures
mixed ANOVA at both sensor and source levels. Significant interactions were further exam-
ined using independent samples t tests, and paired samples t tests where groups were involved
in the interaction.
Pearson correlation was calculated between the coherence values at source level and the
children’s ages in years rounded to months.
The averaged and squared responses around N1m to syllables were compared in a 2
(Hemisphere: Gauche, Droite) × 2 (Groupe: Children, Adults) repeated measures mixed ANOVA.
Plus loin, Pearson correlation coefficients were calculated to examine the relationship between
the peak amplitudes of the auditory responses around 100 ms and coherence values.
Pearson correlation coefficients were calculated to examine the relationship between the
scores of three behavioural tests (RAN: objects subtests, NEPSY: Phonological processing and
Sentence repetition subtests) and coherence values at source level.
Alpha level was 0.05. False discovery rate (FDR) correction for multiple comparisons was
calculated for each analysis.
RÉSULTATS
Coherence Between Brain and Speech Signals for Words and Sentences
Sensor level
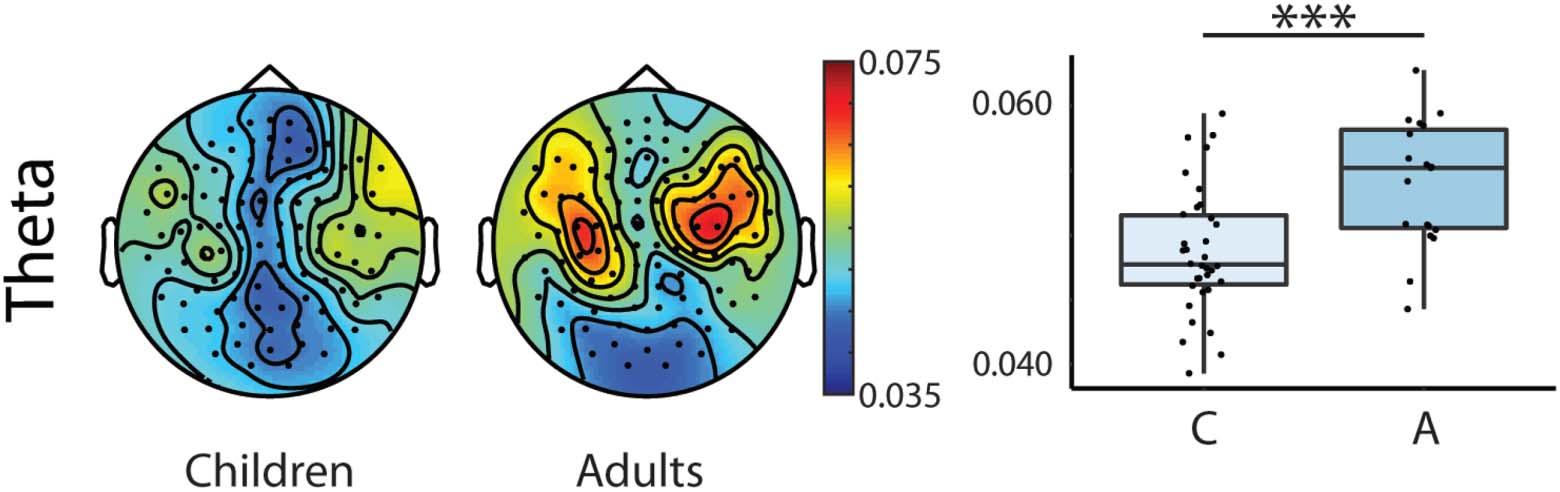
The results of the repeated measures ANOVA revealed first, that adults had the largest coher-
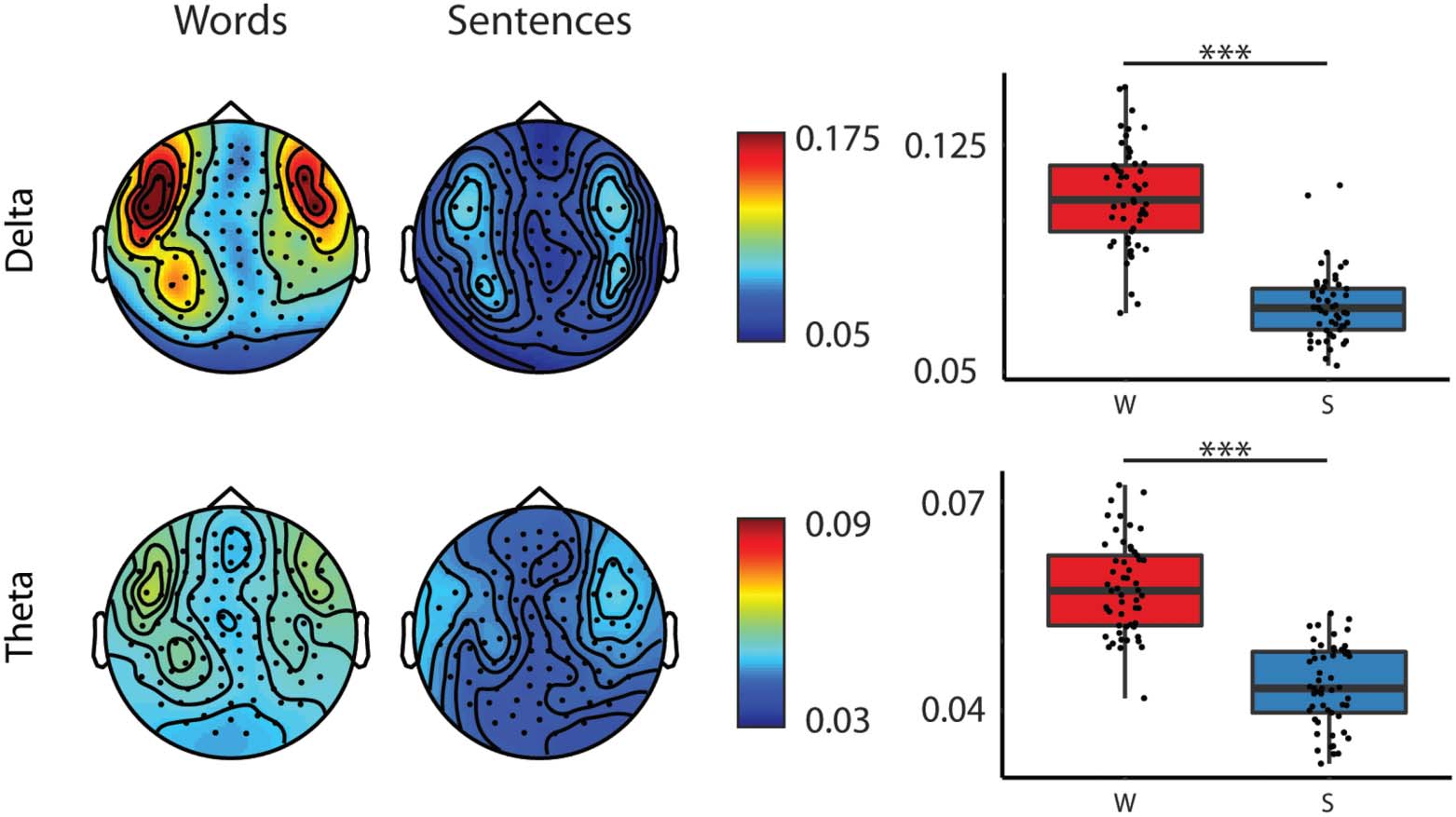
ence values (see Tables 4 et 5, Group main effect; and Figure 2). Deuxième, larger coherence
values were observed for words as compared to sentences for both delta and theta frequency
bands (see Tables 4 et 5, Type main effect; Chiffre 3). Plus loin, we found that coherence
values in the delta band were larger in the left compared to right hemispheres in adults’ brain
responses and that adults had larger coherence values in the left hemisphere than children (voir
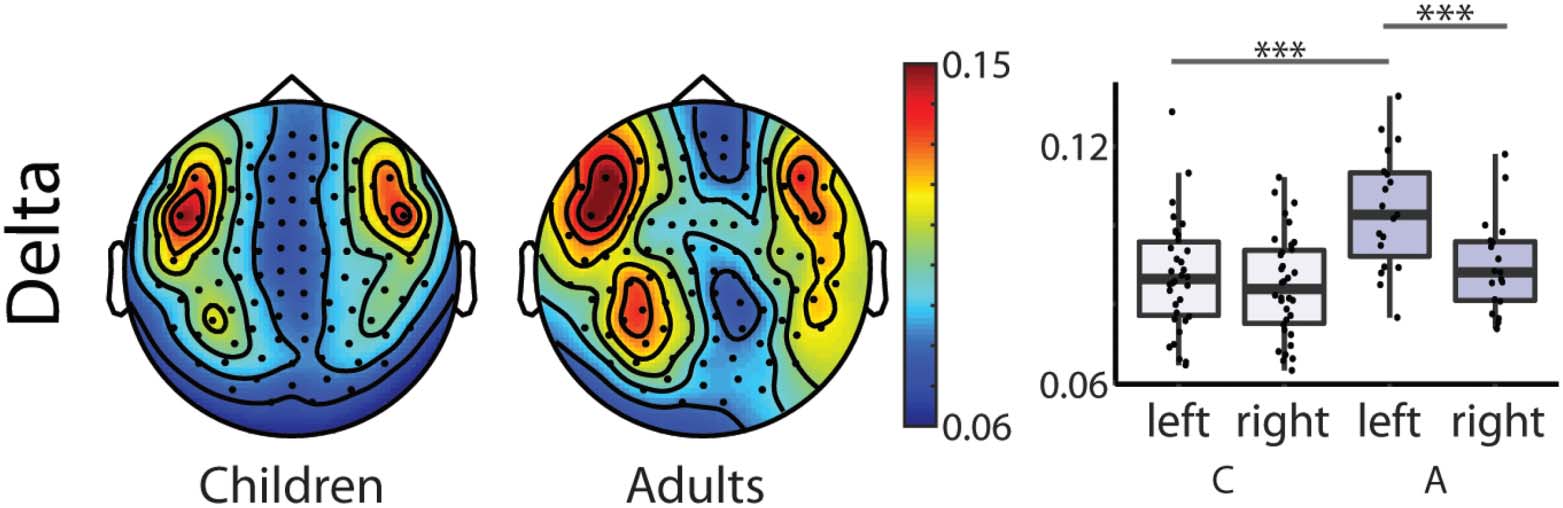
Tableau 4, Hemisphere × Group interaction; Chiffre 4).
Adults showed larger coherence values in the left hemisphere compared to the right hemi-
sphere in the delta band (t(18) = 5.437, p = 0.000) when compared in a paired samples t test.
Tableau 4.
Results of repeated measures mixed ANOVA for the delta frequency band at sensor level
Delta
Main effects and interactions
Type
Hemisphere
Groupe
Type × Group
Hemisphere × Group
Type × Hemisphere
Type × Hemisphere × Group
df
1,51
1,51
1,51
1,51
1,51
1,51
1,51
F value
227.754
11.631
12.739
0.295
5.822
3.670
0.996
p value
0.000
0.001
0.001
0.589
0.019
0.061
0.323
Note. Bold values remained significant after false discovery rate correction.
Neurobiology of Language
partial η2
0.817
0.186
0.200
0.006
0.102
0.067
0.019
236
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
e
d
toi
n
o
/
je
/
je
un
r
t
je
c
e
–
p
d
F
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
un
_
0
0
0
3
3
p
d
/
.
je
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Tableau 5.
Results of repeated measures mixed ANOVA for the theta frequency band at sensor level
Theta
Main effects and interactions
Type
Hemisphere
Groupe
Type × Group
Hemisphere × Group
Type × Hemisphere
Type × Hemisphere × Group
df
1,51
1,51
1,51
1,51
1,51
1,51
1,51
F value
259.307
0.171
14.089
0.836
0.132
0.017
0.051
p value
0.000
0.681
0.000
0.365
0.718
0.896
0.822
partial η2
0.836
0.003
0.216
0.016
0.003
0.000
0.001
Note. Bold values remained significant after false discovery rate correction.
Children’s coherence values did not differ significantly in the two hemispheres (t(33) = 0.730,
p = 0.470). Plus loin, independent samples t tests showed that adults had larger coherence
values in the delta band in the left hemisphere compared to children (t(51) = −4.044, p =
0.000), and the groups did not differ significantly in their coherence values in the right hemi-
sphère (t(51) = −1.386, p = 0.172).
Source level
Similar to the sensor level, the results of the repeated measures ANOVA revealed that adults
had the largest coherence values (see Tables 6 et 7, Group main effect; Chiffre 5) at source
level. Deuxième, larger coherence values were observed for words compared to sentences for
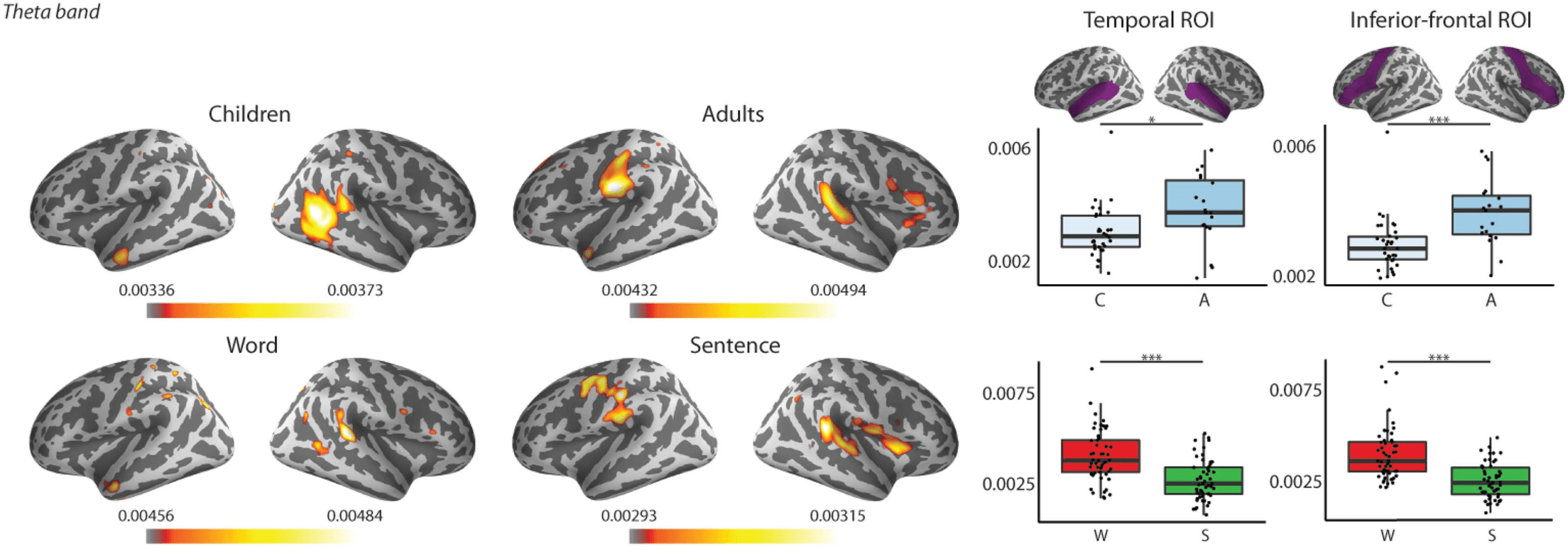
both delta and theta frequency bands (see Tables 6 et 7: Type main effect; Chiffre 6).
Troisième, we found that the adults had larger coherence values compared to children in the delta
Chiffre 2. Topographic distribution of the coherence values and box plots of the theta frequency
band showing Age main effect in the repeated measures mixed ANOVA for the two groups
(Children, N = 34; Adults, N = 19) collapsed across hemispheres and stimulus types.
Topographies: Warmer colours reflect higher coherence between the stimuli envelope and the
brain data. Right box plots: Bold lines denote the median of the coherence values; the bottom
and top edges of the box indicate the 25th and 75th percentiles, respectivement. Light blue boxes show
average coherence for children, dark blue boxes for adults (C = Children, A= Adults). (*** < 0.001)
Neurobiology of Language
237
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Figure 3. Topographic maps of coherence values of the two different frequency bands to word ( W)
and sentence (S) stimuli and box plots of averaged coherences for the delta and theta frequency
bands collapsed across hemispheres and ages. Topographies: Warmer colours reflect higher coher-
ence between the stimuli envelope and the brain data. Boxplots: Bold lines denote the median of
the coherence values; the bottom and top edges of the box indicate the 25th and 75th percentiles,
respectively. Red boxes represent average coherence values for words, and blue boxes for sen-
tences. (*** < 0.001)
band in the temporal region in case of both words and sentences, and that adults had larger
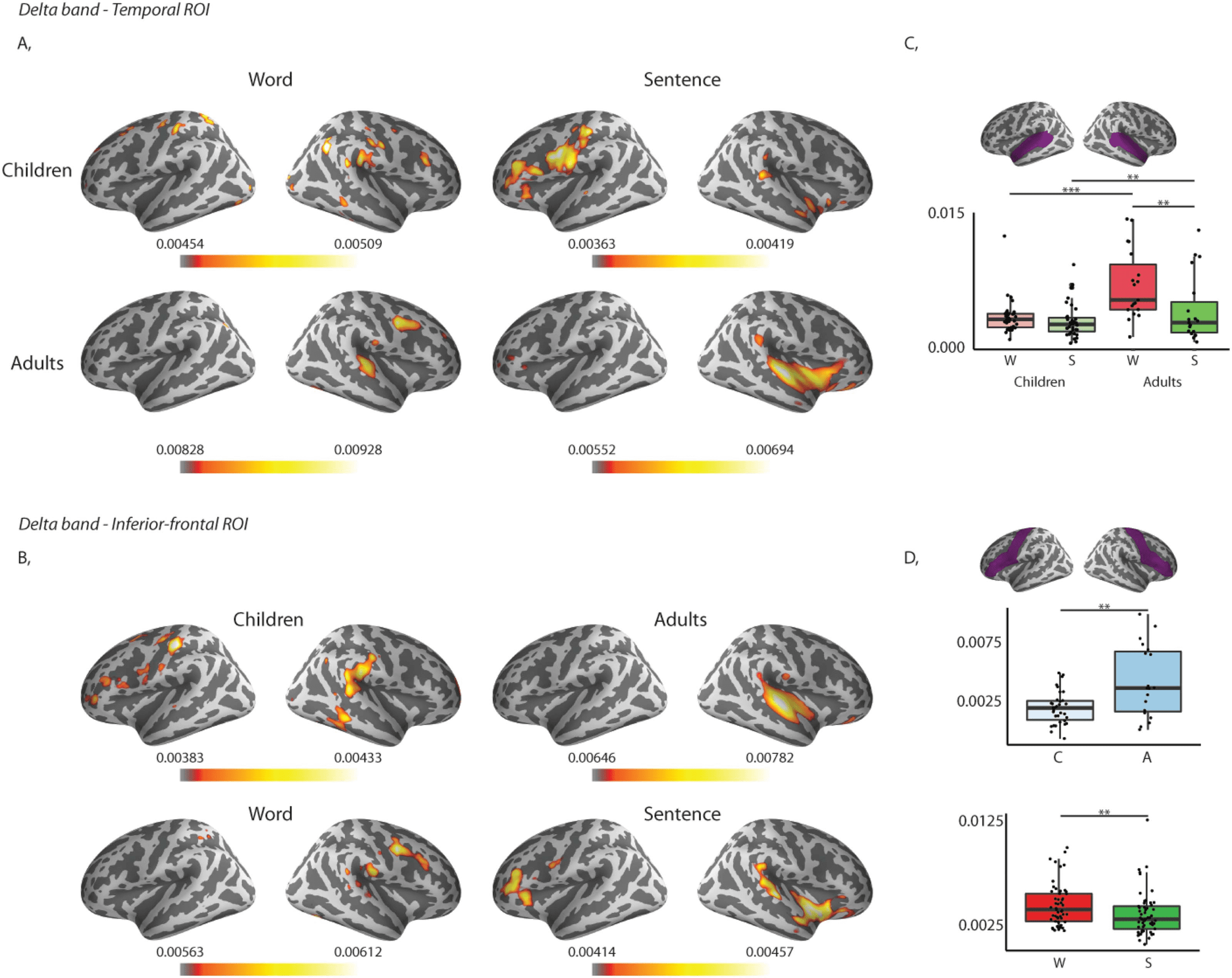
values for words than sentences (See Table 6, Type × Group interaction; Figure 5).
Post hoc independent samples t tests revealed that adults had significantly larger coherence
values for words (t(51) = −4.467, p = 0.000) and for sentences (t(51) = −1.598, p = 0.002)
Figure 4. Topographic distribution of the coherence values and box plots of the delta frequency
band showing a Hemisphere × Group interaction in the repeated measures mixed ANOVA
(Children, N = 34; Adults, N = 19) collapsed across stimulus types. Topographies: Warmer colours
reflect higher coherence between the stimuli envelope and the brain data. Right box plots: Bold
lines denote the median of the coherence values; the bottom and top edges of the box indicate
the 25th and 75th percentiles, respectively. Light purple boxes show average coherence for chil-
dren, dark purple boxes for adults (C = Children, A = Adults). (*** < 0.001)
Neurobiology of Language
238
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Table 6.
Results of repeated measures mixed ANOVA for the delta frequency band at source level in the two regions of interests
Delta – Temporal region
Main effects and interactions
Type
Hemisphere
Group
Type × Group
Hemisphere × Group
Type × Hemisphere
Type × Hemisphere × Group
Delta – Inferior-frontal region
Main effects and interactions
Type
Hemisphere
Group
Type × Group
Hemisphere × Group
Type × Hemisphere
Type × Hemisphere × Group
Note. Bold values remained significant after false discovery rate correction.
df
1,51
1,51
1,51
1,51
1,51
1,51
1,51
df
1,51
1,51
1,51
1,51
1,51
1,51
1,51
F value
12.939
5.266
13.897
6.519
1.727
0.182
0.996
p value
0.001
0.026
0.000
0.014
0.195
0.672
0.323
partial η2
0.202
0.094
0.214
0.113
0.033
0.004
0.019
F value
p value
partial η2
9.143
0.014
13.476
1.291
1.960
0.737
0.037
0.004
0.907
0.001
0.261
0.168
0.395
0.849
0.152
0.000
0.209
0.025
0.037
0.014
0.001
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
.
/
l
compared to children. Paired samples t tests revealed that adults also had significantly larger
coherence values for words compared to sentences (t(18) = 3.200, p = 0.005), and children’s
coherence values did not differ significantly between words and sentences (t(33) = 1.000, p =
0.325).
Because the child group spanned a relatively large age range (4.7–9.3 years), we examined
whether age was linearly related to changes in coherence values. We did not find any signif-
icant correlation between the observed coherence values and age (see Table 8).
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Evoked Responses to Syllables
Sensor level
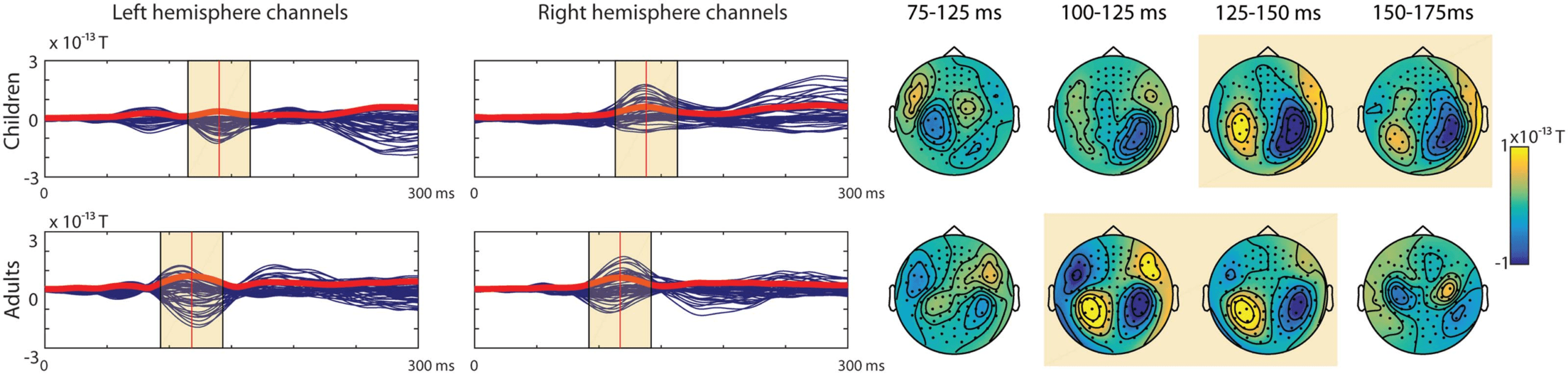
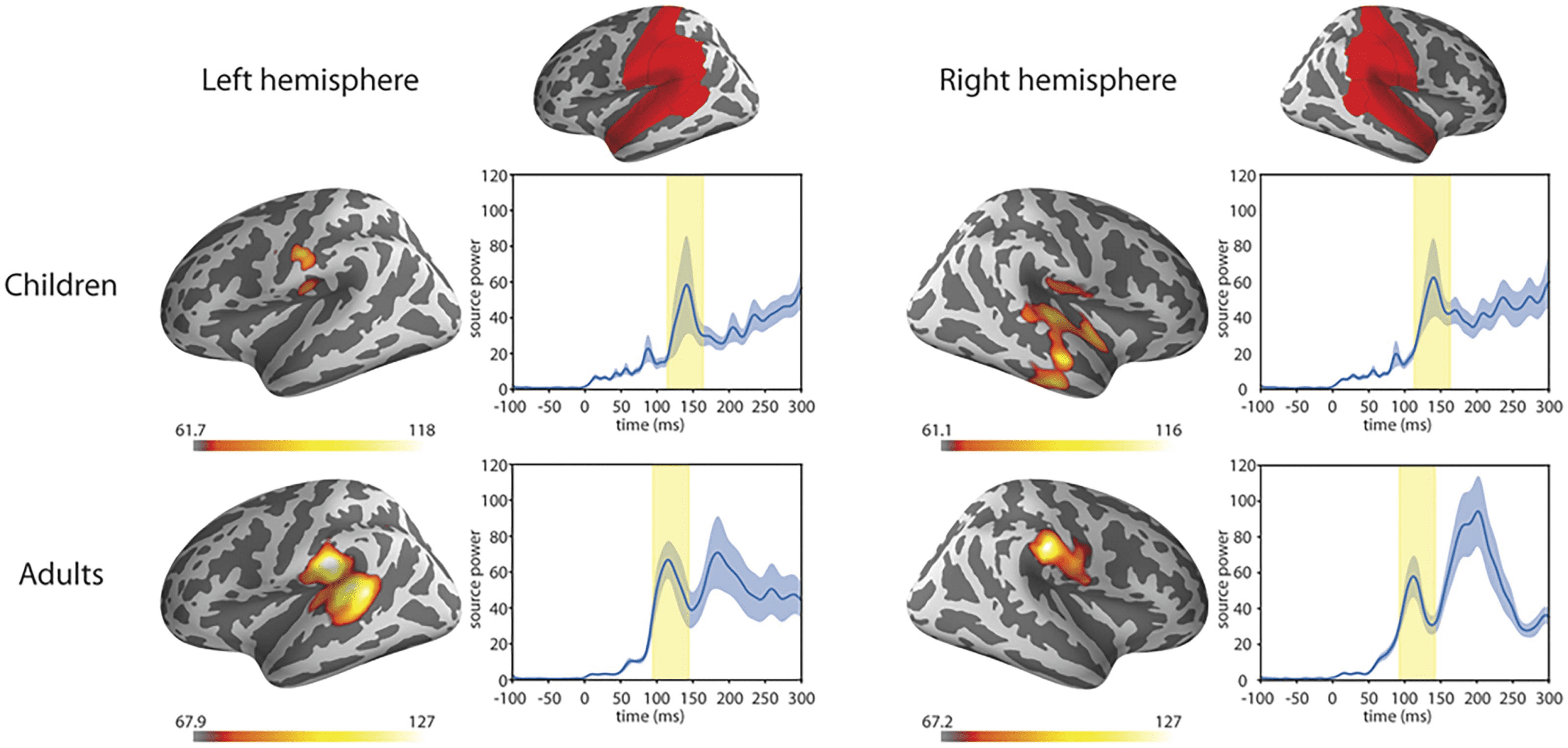
The averaged evoked responses’ topographies were typical of the N1m response in adults. In
children the topography reminiscent of the N1m was slightly later in time in the right hemi-
sphere. The left hemisphere showed a less clear pattern for children (see Figure 7). The topog-
raphies were also examined individually.
The averaged squared responses were compared in a 2 (Hemisphere: left, right) × 2 (Group:
Children, Adults) repeated measures mixed ANOVA (see Table 9). No significant differences
were found.
Neurobiology of Language
239
Age-related differences in coherence
Table 7.
Results of repeated measures mixed ANOVA for the theta frequency band at source level in the two regions of interests
Theta – Temporal region
Main effects and interactions
Type
Hemisphere
Group
Type × Group
Hemisphere × Group
Type × Hemisphere
Type × Hemisphere × Group
Theta – Inferior-frontal region
Main effects and interactions
Type
Hemisphere
Group
Type × Group
Hemisphere × Group
Type × Hemisphere
Type × Hemisphere × Group
Note. Bold values remained significant after false discovery rate correction.
df
1,51
1,51
1,51
1,51
1,51
1,51
1,51
df
1,51
1,51
1,51
1,51
1,51
1,51
1,51
F value
44.799
5.850
6.849
2.131
0.743
0.253
0.190
F value
50.638
0.540
14.688
0.001
0.865
0.398
0.003
p value
0.000
0.019
0.012
0.151
0.393
0.617
0.665
partial η2
0.468
0.103
0.118
0.040
0.014
0.005
0.004
p value
partial η2
0.000
0.465
0.000
0.977
0.357
0.531
0.960
0.498
0.011
0.224
0.000
0.017
0.008
0.000
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The averaged squared values were then correlated with the corresponding hemisphere’s
coherence values in the frequency bands. No significant correlations were found. (For the ta-
ble of correlation coefficients and p values, see Supplementary Material 7, Table 7.1.)
Topography of the averages was visually inspected then to confirm the correct N1m re-
sponse pattern in each participant, for left and right hemispheres separately.
The N1m response in the left hemisphere was observed in 4 (11.76%) children, with an
average latency of 130 ms, and 17 (89.47%) adults, with an average latency of 104 ms. Six
(17.65%) children with an average latency of 151 ms showed an activation pattern with an
opposite current direction to the adult-like N1m.
The N1m response in the right hemisphere was observed in 8 (23.53%) of the children’s
evoked responses, with an average latency of 141 ms, and in 17 (89.47%) of the adults’ evoked
responses, with an average latency of 105 ms. Five (14.71%) of the children with an average
latency of 143 ms showed an activation pattern with an opposite current direction to the
adult-like N1m.
To examine whether the N1m amplitude and coherence values in the delta and theta bands
would follow a similar developmental pattern, correlations were calculated to quantify the
possible developmental relationship between the measures. Coherence values were plotted
Neurobiology of Language
240
Age-related differences in coherence
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5.
Left panels: Grand average of source level coherence values of children and adults to words and sentences. (A) In the delta
frequency band in the temporal region of interest. (B) In the delta frequency band in the inferior-frontal region of interest; top row: grand
averages of children and adults; bottom row: grand averages to words and sentences. Warmer colours reflect higher coherence between
the stimuli envelope and the brain data. Right panels: Region of interest highlighted in purple (as defined in the Desikan-Killiany Atlas;
Desikan et al., 2006). (C) Box plots of averaged coherence values in the delta frequency band in the temporal region collapsed across hemi-
spheres. (D) Box plots of averaged coherence values in the delta frequency band in the inferior-frontal region collapsed across hemispheres
and ages (top) or stimulus types (bottom). Bold lines denote the median of the coherence values; the bottom and top edges of the box indicate
the 25th and 75th percentiles, respectively. W = words, S = sentences, C = children, A = adults. (*** < 0.001, ** < 0.01)
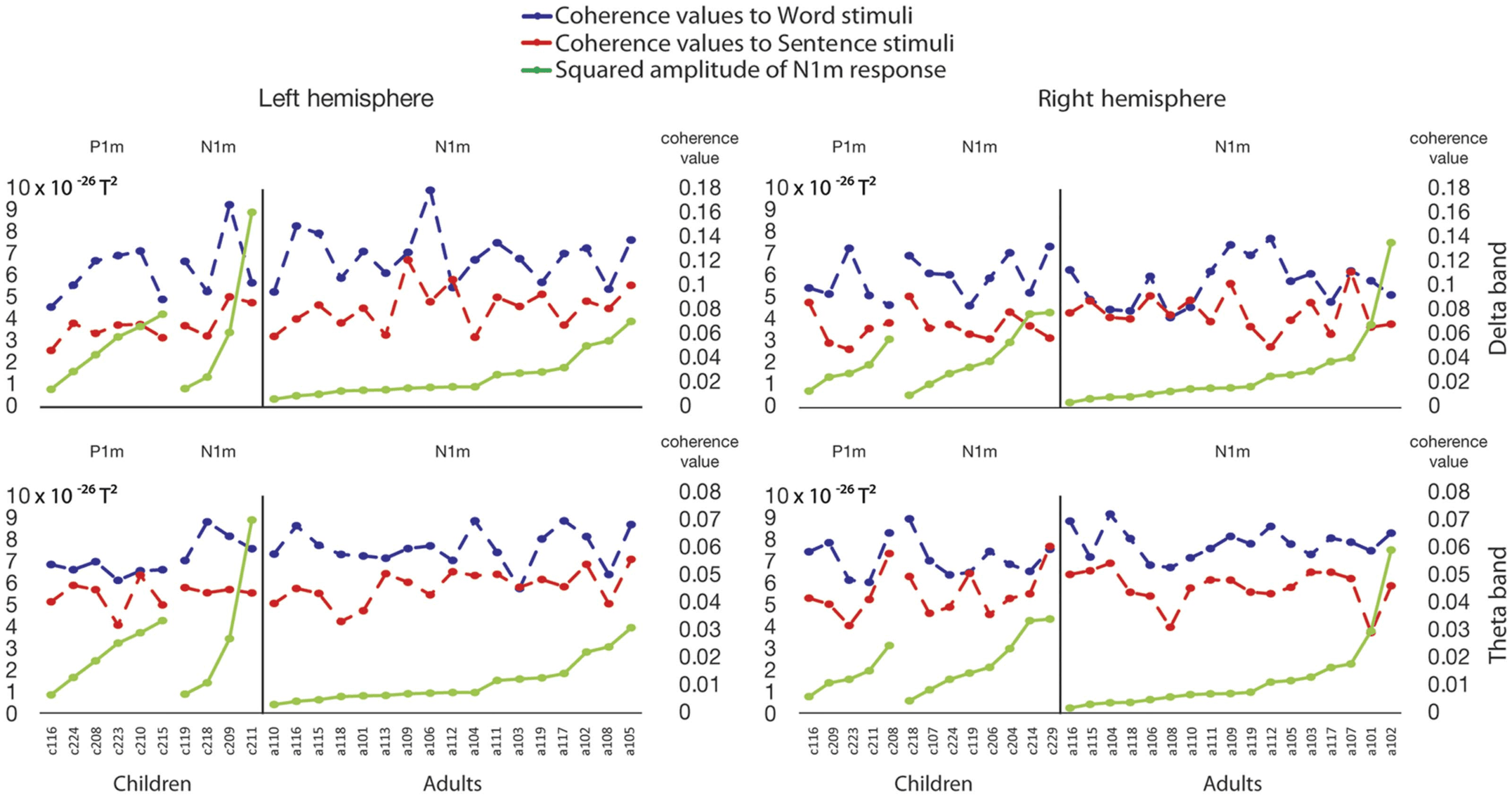
against the N1m responses in the child and adult groups (see Figure 8). No significant corre-
lations were found between N1m amplitude to syllables and delta and theta coherence values
to words and sentences in either the left or right hemispheres after correction for multiple com-
parisons. (For the table of correlation coefficients and p values, see Supplementary Material 7,
Table 7.2.)
Neurobiology of Language
241
Age-related differences in coherence
Figure 6. Left: Grand average of source level coherence values in the theta frequency band. Warmer colours reflect higher coherence be-
tween the stimuli envelope and the brain data. Top row: Grand averages of children and adults. Bottom row: Grand averages to words and
sentences. Right: Region of interest highlighted in purple (as defined in the Desikan-Killiany Atlas; Desikan et al., 2006) and box plots of
averaged coherences of the ROIs in the theta frequency band collapsed across hemispheres and ages (top) or stimulus types (bottom).
Bold lines denote the median of the coherence values; the bottom and top edges of the box indicate the 25th and 75th percentiles, respec-
tively. Top plot: Light blue boxes show average coherence for children (C), and dark blue boxes for the adults (A). Bottom plot: Red boxes
represent average coherence values for words ( W), and green boxes for sentences (S). (*** < 0.001, * < 0.05)
Table 8.
Results of correlations between the coherence values at source level and age in the
children group
Delta
Theta
Temp
Inf-front
Temp
Inf-front
Correlation coefficient
0.049
−0.036
0.165
0.056
Sig
0.785
0.840
0.352
0.752
N
34
34
34
34
Note. Sig = significance. Temp = Temporal. Inf-front = Inferior-frontal.
Figure 7. Blue butterfly plots of the group-averaged magnetometers with the global mean field power (GMFP) (red line) and topographic
maps for the evoked responses to the syllable stimuli for the two age groups (Children, N = 34; Adults, N = 19). The yellow boxes highlight
where the auditory response was expected in the groups based on the GMFP.
Neurobiology of Language
242
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Table 9.
on the GMFP peaks at sensor level
Results of repeated measures mixed ANOVA for the averaged squared responses based
Main effects and interactions
Hemisphere
Group
Hemisphere × Group
df
1,51
1,51
1,51
F value
0.013
3.761
0.414
p value
0.531
0.058
0.523
partial η2
0.000
0.069
0.008
Source level
The responses were compared in a 2 (Hemisphere: left, right) × 2 (Group: Children, Adults)
repeated measures mixed ANOVA (see Table 10 and Figure 9). No significant differences were
found.
The averaged power was then correlated with the corresponding hemisphere’s coherence
values in the frequency bands. No significant correlations were found. (For the table of corre-
lation coefficients and p values, see Supplementary Material 7, Table 7.3.)
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 8. Line plots for coherence values in the left and right hemispheres to word and sentence stimuli in comparison to the squared am-
plitude of the N1m (T2) in the participants who showed a P1m or N1m pattern in their ERF responses. In each plot, the scale on the left side
shows squared amplitude of N1m response, and the scale on the right side shows coherence values. Blue dashed line: coherence values to
word stimuli; red dashed line: coherence values to sentence stimuli; green solid line: the squared N1m amplitude. Top row plots show
coherence values for the delta band, left and right hemispheres respectively; bottom row plots show coherence values for the theta band,
left and right hemispheres respectively. Values are organized in order of the squared amplitudes.
Neurobiology of Language
243
Age-related differences in coherence
Table 10.
on the GMFP peaks at source level
Results of repeated measures mixed ANOVA for the averaged squared responses based
Main effects and interactions
Hemisphere
Group
Hemisphere × Group
df
1,51
1,51
1,51
F value
0.3976
0.105
3.469
p value
0.531
0.747
0.068
partial η2
0.008
0.002
0.064
Correlations of Source Level Coherence with Behavioural Scores
Behavioural scores in the Phonological processing and Sentence repetition tasks did not cor-
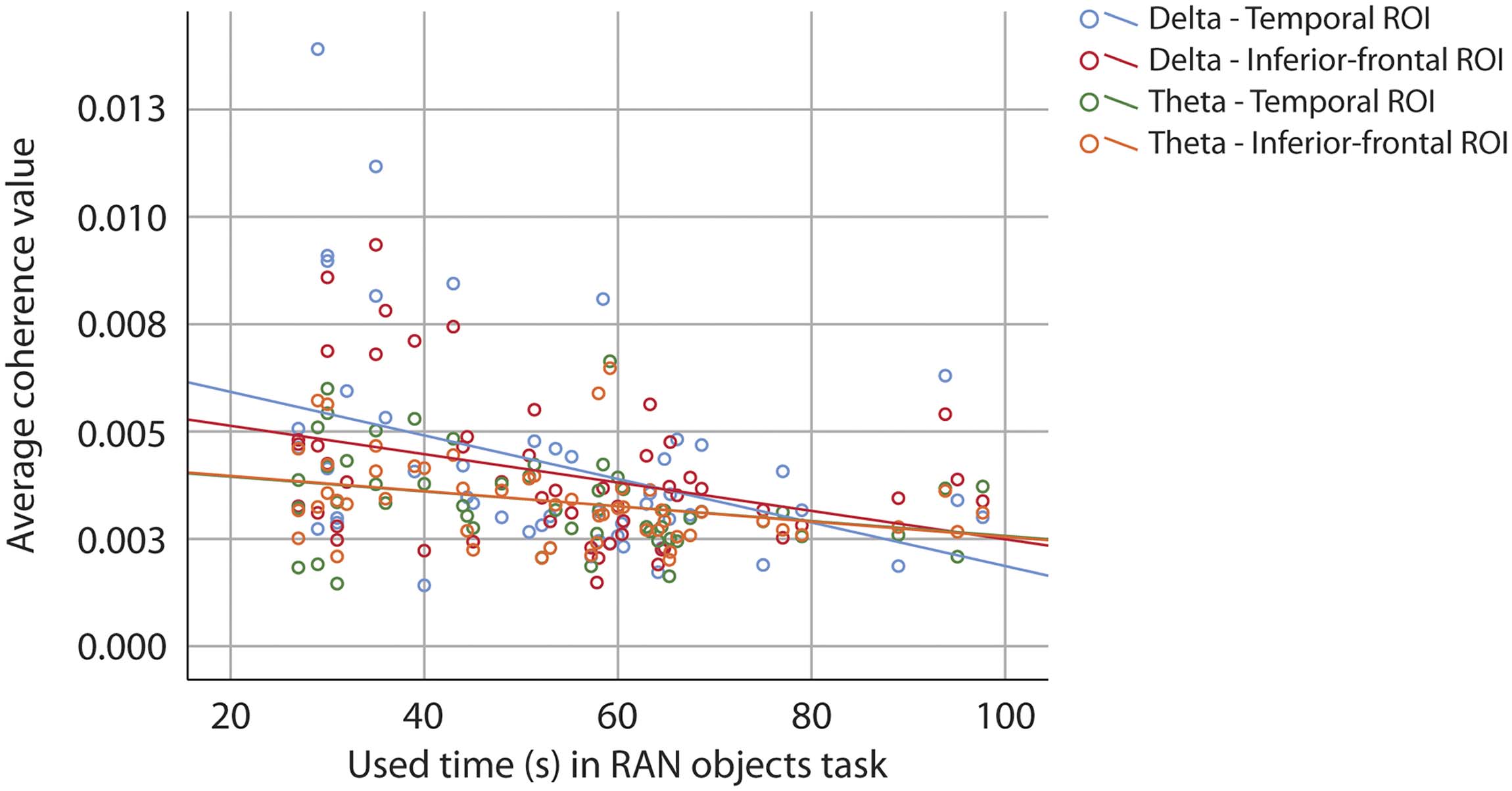
relate with coherence values from the delta and theta bands. RAN objects did correlate in-
versely with both frequency bands and both ROIs (see Table 11 and Figure 10), but when
age was controlled for, the correlation was no longer significant (see Table 12).
Coherence values were correlated with the child groups scores on NEPSY’s phonological
processing task and sentence repetition task (see Tables 13 and 14). No significant correlations
were found.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 9. Grand averaged source level ERFs to syllables. Warmer colours reflect higher source power of the event-related field. The red areas
highlighted were included in the region of interest (as defined in the Desikan-Killiany Atlas; Desikan et al., 2006). Right panels show the
average source waveform (MNE estimate) extracted from the brain regions. The blue shading represents the standard error of the mean,
and the yellow shading shows the time-windows used for the N1m response.
Neurobiology of Language
244
Age-related differences in coherence
Table 11. Correlations between performance on RAN of objects (time in seconds) and the coherence
values from the two regions of interest in the delta and theta frequency bands at source level
Delta
Theta
Temp
Inf-front
Temp
Inf-front
Correlation coefficient
−0.377
−0.350
−0.292
−0.330
Sig
0.005
0.010
0.034
0.016
N
53
53
53
53
Note. Sig = significance. Temp = Temporal. Inf-front = Inferior-frontal.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
Figure 10.
Scatter plot of correlation between performance on RAN objects and coherence values
at source level. Blue dots and line represent coherence values from the temporal region of interest,
and red dots and line represent values from the inferior-frontal region of interest in the delta frequency
band. Green dots and line represent values from the temporal region of interest, and orange dots and
line represent values from the inferior-frontal region of interest in the theta frequency band.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Table 12.
source level controlled for age
Correlations between performance on RAN of objects and the coherence values at
Delta
Theta
Temp
Inf-front
Temp
Inf-front
Correlation coefficient
−0.070
0.020
−0.005
0.055
Sig
0.624
0.888
0.971
0.699
Note. Sig = significance. Temp = Temporal. Inf-front = Inferior-frontal.
N
53
53
53
53
245
Neurobiology of Language
Age-related differences in coherence
Table 13. Correlations between performance on the phonological processing task and the
coherence values at source level
Delta
Theta
Temp
Inf-front
Temp
Inf-front
Correlation coefficient
0.037
−0.058
0.234
0.131
Sig
0.836
0.743
0.182
0.462
Note. Sig = significance. Temp = Temporal. Inf-front = Inferior-frontal.
Table 14.
Correlations between performance on the repetition of sentences task and the
coherence values at source level
Delta
Theta
Temp
Inf-front
Temp
Inf-front
Correlation coefficient
0.097
0.200
0.076
−0.055
Sig
0.585
0.256
0.670
0.757
Note. Sig = significance. Temp = Temporal. Inf-front = Inferior-frontal.
N
34
34
34
34
N
34
34
34
34
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
DISCUSSION
This study investigated whether children and adults differ in overall brain activity as well as in
left and right auditory cortex activity while listening to various speech units that are essential
for spoken language processing. More specifically, we examined how auditory processing of
words and sentences is reflected in the level of coherence and hemispheric lateralization
across development, and how these are related to the processing of syllables, at both the sen-
sor and source levels. To this end, two age groups were tested for comparison: children be-
tween the ages of 4.7 and 9.3 years and adults. Coherence is an interesting and useful measure
of the brain’s ability to track the speech envelope across different frequency bands by quan-
tifying the similarity in frequency content between brain activity and the speech envelope. The
higher the coherence between brain activity and the speech envelope, the better the speech
tracking.
First, we found an improvement with age in the brain’s ability to track speech evidenced as
increased coherence values in the delta and theta frequency bands between brain signal and
the speech envelope. Second, at the sensor level, where the whole hemispheres were exam-
ined, we found an interaction between hemispheres and age groups in the delta band with
adults showing larger values at the left than the right hemisphere. However, at the source level,
hemispheric differences in coherence values did not interact with age, which suggests no dif-
ferences in maturation rates for the left and right auditory and frontal cortices in the degree to
which the brain can synchronize to the speech envelope. Third, we also found differences in
the coherence values observed for the word and sentence stimuli independent of age, al-
though this was attributed to physical stimulus length rather than linguistic unit size, suggesting
that the methodological approach should be taken into account when interpreting findings
Neurobiology of Language
246
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
about speech perception. Last, we found no relationship between the general maturation of
auditory processing and speech tracking as indicated by early ERFs to syllables.
Developmental differences were found as an overall increase in the coherence values in
both the delta and theta frequency bands between adults and children, with adults showing
largest values compared to children. Further, the topography of the coherence for these fre-
quencies exhibited a clear pattern of auditory cortex activation at the sensor level. This was
mostly confirmed by the source level analysis. The coherence values reflect how similar the
frequency contents between the brain signal and the speech envelope are; therefore, our find-
ings could be interpreted as increased precision of the auditory system to track the speech from
childhood to adulthood. However, when examining the coherence values as a continuous
variable within the child groups, we did not find any correlation between age and coherence
values. There may be several reasons for the observed differences between adults and
children.
First, basic auditory processing matures slowly with major changes in, for example, ERP
responses noted at around ages 8–9 years with further changes until late adolescence
(Ponton et al., 2000). This slow maturation of basic processing could affect the precision of
speech processing in a bottom-up manner.
Second, the bottom-up process could be affected by genetically driven maturation or con-
tinued exposure to speech that refines the bottom-up pathway of the auditory system (Kuhl,
2000; Ponton et al., 2000). At the same time, continuous exposure to speech refines and
changes the brain’s ability to perceive speech in a top-down manner as well (Kuhl, 2000).
This environmental input would shape long-term memory representations, therefore affecting
speech processing.
Last, the development of speech tracking may interact with other co-developing cognitive
and language-related abilities (e.g., receptive and expressive vocabulary, speech motor and
phonological developments) in addition to maturational factors such as age. There is for in-
stance evidence that children initially process large units that are lexically based (e.g., words)
before developing representations for smaller units (syllables, individual phonemes; for a re-
view, see Vihman, 2017). This process may also be affected by reading acquisition that places
emphasis on phonemes (e.g., Brennan et al., 2013; Popescu & Noiray, 2019; Ziegler &
Goswami, 2005). Further, in speech production research investigating the size of coarticula-
tory units across age, Noiray and colleagues (Noiray et al., 2018; Noiray, Wieling et al., 2019)
noted that children do not mature their coarticulatory patterns in a linear fashion. Instead, they
found that preschoolers at the age of 3, 4, and 5 organised their speech in larger chunks com-
pared to primary school children at the age of 7 and adults (Noiray et al., 2018; Noiray,
Wieling et al., 2019).
In a subsequent study, Noiray and colleagues further demonstrated that the development of
children’s phonological awareness, that is, the awareness that the native language, is composed
of various size compounds (e.g., syllables, rhyme, and individual phonemes) and the ability to
manipulate those units interacts with children’s speech motor organisation (Noiray, Popescu
et al., 2019). Greater awareness of individual phonemes was associated with greater phonemic
differentiation of articulatory gestures. To summarise, relationships between several cognitive
and language-related abilities occur in the course of language acquisition, and they seem to
evolve dynamically over time (e.g., Noiray, Popescu et al., 2019; Noiray, Wieling et al.,
2019; Vihman, 2017). In future research, it will be important to investigate larger samples of
children spanning kindergarten to primary school to better understand the dynamics of these
relationships and how they contribute to the development of speech tracking specifically.
Neurobiology of Language
247
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Our research provides supplementary information about the processing of various speech-
sized units. More specifically, we confirmed the role of the lower frequency bands for sentence
processing (Molinaro & Lizarazu, 2018; Ríos-López et al., 2020) and extended this finding to
the processing of words. Indeed, there is evidence that theta and delta bands play a main role
for parsing the continuous speech signal into linguistic and prosodic units (Poeppel, 2014;
Poeppel & Assaneo, 2020). Thus, a developmental increase in brain coherence in these
frequency bands could be associated with a development in the processing and awareness
of those distinct speech units. While we did not find any significant correlation between chil-
dren’s coherence values and their performance on phonological processing or sentence rep-
etition, future studies should further examine the relationship among phonological awareness,
reading, and speech tracking in the brain with larger samples of children and longitudinally.
We also found that coherence was higher for words than sentences for all frequency bands.
This was somewhat surprising given longer stimuli should provide opportunities for brain activity
to lock to the ongoing auditory signal. It is important, however, to note that after checking the
coherence at the beginning of sentences trials (with the same length as used for words), we noted
that coherence increased compared to the original values for sentences. This suggests that the
higher coherence for words than sentences does not reflect differences that would be directly
relevant for neural computation of linguistic units, but more the characteristics of calculating the
coherence measure for short versus long stimuli. For example, longer stimuli provide greater
chances for brain activity unrelated to stimulation to occur, with higher likelihood of this noise
in the brain activity interacting with the coherence measure. Therefore, comparison of coher-
ence measures across different length stimuli should be done with care, as pure physical length
of the stimulus might have an effect on the results. In general, our findings confirm that speech
tracking can happen at a shorter length, such as words, as well as at sentence level.
While we expected to find a significant interaction between the coherence values in the left
and right hemispheres and the two age groups in the delta band, this difference was in the
opposite direction from our predictions, where we expected larger values in the right hemi-
sphere compared to the left, particularly in the adults (Luo & Poeppel, 2007). We observed
significantly larger coherence values in the left hemisphere than right for adults in the delta
band only at the sensor level—this was not observed at the source level. One possible reason
for this difference between sensor and source level findings could be the selection of channels
or regions used in the analysis. The sensor level comparison used an overall average of the
hemispheres, while the source level focused on the temporal regions. At the source level,
when focusing on the temporal regions, a hemispheric difference was found in the expected
direction of larger right side activation compared to the left; however, the difference was no
longer significant after FDR correction.
Finally, we investigated the overlap of different maturational processes across linguistic
units by examining the age-related changes in brain activity around 125 ms (the time-window
of the N1m response in adults to the syllables) and compared those to the coherence values for
words and sentences. The amplitudes of evoked responses and the coherence values likely
represent different neuronal mechanisms. The first presumably represents a more general mat-
uration of the auditory and speech perception system, and the latter is likely linked to top-
down processes such as comprehension of speech (Luo & Poeppel, 2007; Peelle et al.,
2013; Ponton et al., 2000, 2002). We compared the development of the evoked responses
and coherence using two approaches. In our first approach using GMFP, which included both
the P1m and N1m responses, we found no significant difference between the groups.
However, the P1m and N1m likely represent different computational processes of the sounds.
In early childhood the auditory ERPs show prominent P1 and N2 peaks. During development,
Neurobiology of Language
248
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
the P1 response shifts to earlier latencies accompanied by a decrease in amplitudes, and the
prominent N1 response emerges to the waveform at around early school years (Albrecht et al.,
2000; Ponton et al., 2000). Importantly, P1m of young children and N1m of older children and
adults show very similar timing, obscuring the interpretation of purely GMFP-based interpre-
tations (Parviainen et al., 2019).
Therefore, as a next step, we checked whether the spatial patterns and timings of responses
in the left and right hemispheres for each individual matched with the expected N1m pattern
based on the current direction in the magnetometer topography. We found no systematic cor-
relation between the responses in the time-window of the first prominent evoked field (the
N1m or P1m) and coherence between the speech envelope in the delta and theta bands.
Although the correlations were not significant it should be noted that several factors might
affect the result, such as sample size and methodology used. The ERFs and coherence were
examined using different approaches (ERFs in the time domain and coherence in the frequency
domain). It is possible that the use of these different approaches makes the measures difficult to
compare directly. Taking this into consideration, our results suggest that the evoked response
to syllables and the speech tracking might develop independently of each other and not share
robust maturational mechanisms. If this is the case, the ERF amplitudes could reflect more
bottom-up processes while the coherence values more top-down processes. Previous literature
shows that ERFs are clearly modulated by the physical features of the sounds (Näätänen &
Picton, 1986; Näätänen et al., 1997), and the speech envelope following seems to be linked
to speech intelligibility and attention (Peelle et al., 2013).
Furthermore, especially in the case of younger children, while the GMFP did show a re-
sponse around 100 ms, individual inspection of the responses showed that the response at the
time was not actually an N1m response, but rather P1m. Taking this into account, it could
perhaps explain why we found no differences between the groups when comparing the re-
sponses based on time-windows defined by only the peak in the GMFP.
Likewise, no hemispheric differences were observed in any of the age groups for the GMFP-
based values of the evoked response, in contrast to a previous MEG study (Parviainen et al.,
2019). However, the difference between these studies most likely reflects the chosen analysis
approach. While the GMFP reflects the overall response strength at the sensor level, at the
source level, measures of equivalent current dipoles depict the spatially specific amplitude
values at different time points. Indeed, our data demonstrated a similarly delayed pattern of
N1m topography in children, with more clear response in the right than left hemisphere, as
was implied by Parviainen and colleagues (2011, 2019). Inspection of the responses them-
selves revealed that only about one third of the children actually had the N1m evoked
response. Due to our sample size, comparison using brain responses of children who indeed
produced the N1m response would be unbalanced, and future research should look into this
comparison with a larger sample size for both groups.
One of the limitations of our study is related to the type of stimulus we used, as words ut-
tered in isolation are more pronounced than those in a sentence. However, our post hoc com-
parison of the coherence values to sentences at word-length trials vs. sentence-length trials
revealed that higher coherence was found at the beginning of the sentence regardless of stim-
ulus type. It is possible the word level stimuli could be affected by their short length in the
estimation of low frequencies, and therefore the results for the word stimuli should be regarded
with caution. Another potential limitation is the number of participants limiting the power of
the study to detect more subtle differences related to development or hemispheric processing
of the different stimuli.
Neurobiology of Language
249
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
In summary, we investigated developmental differences in speech processing of various
speech sized units that are linguistically relevant for spoken language processing, the syllable,
word, and sentence, using MEG. We also examined how the hemispheric specialization is
represented in brain responses and whether this specialization varies as a function of age.
Overall, we found that both delta and theta frequencies show coherence with speech and
seem to be important for speech processing. We also found developmental changes in the
coherence values, which could reflect both bottom-up maturation and top-down refinement
caused, for example, by continuous refinement of speech sound representations. Our data also
suggest that the general functional maturation of the auditory cortices follows a different tra-
jectory to that of the brain activity tracking the speech envelope.
ACKNOWLEDGMENTS
The authors would like to thank Katja Koskialho, Sonja Tiri, Ainomaija Laitinen, Annamaria
Vesterinen, Aino Sorsa, Maija Koskio, and Cherie Jenkins for their help with data collection.
This work has been supported by the European Union projects Predictable (Marie Curie
Innovative Training Networks, # 641858), and ChildBrain (Marie Curie Innovative Training
Networks, #641652).
FUNDING INFORMATION
Barbara Höhle, Horizon 2020 Framework Programme (https://dx.doi.org/10.13039/100010661),
Award ID: 641858. Paavo Leppänen, Horizon 2020 Framework Programme (https://dx.doi.org
/10.13039/100010661), Award ID: 641652.
AUTHOR CONTRIBUTIONS
Orsolya Beatrix Kolozsvári: Conceptualization: Equal; Data curation: Equal; Investigation:
Lead; Formal analysis: Equal; Methodology: Equal; Project administration: Equal; Software:
Equal; Visualisation: Lead; Writing–Original Draft: Equal; Writing–Review & Editing: Equal.
Weiyong Xu: Conceptualization: Equal; Data curation: Equal; Formal analysis: Equal;
Methodology: Equal; Software: Equal; Visualisation: Supporting; Writing–Original Draft:
Equal; Writing–Review & Editing: Equal. Georgia Gerike: Formal analysis: Equal;
Methodology: Equal; Visualisation: Supporting; Writing–Review & Editing: Equal. Tiina
Parviainen: Conceptualization: Equal; Methodology: Supporting; Supervision: Supporting;
Writing–Review & Editing: Equal. Lea Nieminen: Conceptualization: Equal; Methodology:
Supporting; Writing–Review & Editing: Equal. Aude Noiray: Supervision: Supporting;
Writing–Review & Editing: Equal. Jarmo Hämäläinen: Conceptualization: Equal; Formal anal-
ysis: Equal; Funding acquisition: Lead; Methodology: Equal; Project administration: Equal;
Supervision: Lead; Writing–Original Draft: Equal; Writing–Review & Editing: Equal.
REFERENCES
Abrams, D. A., Nicol, T., Zecker, S., & Kraus, N. (2008). Right-
hemisphere auditory cortex is dominant for coding syllable pat-
terns in speech. Journal of Neuroscience, 28(15), 3958–3965.
DOI: https://doi.org/10.1523/JNEUROSCI.0187-08.2008, PMID:
18400895, PMCID: PMC2713056
Abrams, D. A., Nicol, T., Zecker, S., & Kraus, N. (2009). Abnormal
cortical processing of the syllable rate of speech in poor readers.
Journal of Neuroscience, 29(24), 7686–7693. DOI: https://doi
.org/10.1523/ JNEUROSCI.5242-08.2009, PMID: 19535580,
PMCID: PMC2763585
Albrecht, R., Suchodoletz, W., & Uwer, R. (2000). The development
of auditory evoked dipole source activity from childhood to adult-
hood. Clinical Neurophysiology, 111(12), 2268–2276. DOI: https://
doi.org/10.1016/S1388-2457(00)00464-8, PMID: 11090781
Boersma, P., & Weenink, D. (2018). Praat: Doing phonetics by
computer [Computer program] ( Version 6.0.37). Retrieved from
https://www.praat.org/
Bourguignon, M., De Tiege, X., de Beeck, M. O., Ligot, N., Paquier,
P., Van Bogaert, P., Goldman, S., Hari, R. & Jousmäki, V. (2013).
The pace of prosodic phrasing couples the listener’s cortex to the
Neurobiology of Language
250
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
reader’s voice. Human Brain Mapping, 34(2), 314–326. DOI:
https://doi.org/10.1002/hbm.21442, PMID: 22392861, PMCID:
PMC6869855
Brennan, C., Cao, F., Pedroarena-Leal, N., McNorgan, C., & Booth,
J. R. (2013). Reading acquisition reorganizes the phonological
awareness network only in alphabetic writing systems:
Learning to read reorganizes language network. Human Brain
Mapping, 34(12), 3354–3368. DOI: https://doi.org/10.1002
/hbm.22147, PMID: 22815229, PMCID: PMC3537870
Dehaene-Lambertz, G., Dehaene, S., & Hertz-Pannier, L. (2002).
Functional neuroimaging of speech perception in infants.
Science, 298(5600), 2013–2015. DOI: https://doi.org/10.1126
/science.1077066, PMID: 12471265
Denckla, M. B., & Rudel, R. G. (1976). Naming of object-drawings
by dyslexic and other learning disabled children. Brain and
Language, 3(1), 1–15. DOI: https://doi.org/10.1016/0093-934X
(76)90001-8, PMID: 773516
Desikan, R. S., Ségonne, F., Fischl, B., Quinn, B. T., Dickerson, B. C.,
Blacker, D., Buckner, R. L., Dale, A. M., Maguire, R. P., Hyman,
B. T., & Albert, M. S. (2006). An automated labeling system for
subdividing the human cerebral cortex on MRI scans into gyral
based regions of interest. Neuroimage, 31(3), 968–980. DOI:
https://doi.org/10.1016/j.neuroimage.2006.01.021, PMID:
16530430
Ding, N., Melloni, L., Yang, A., Wang, Y., Zhang, W., & Poeppel,
D. (2017). Characterizing neural entrainment to hierarchical lin-
guistic units using electroencephalography (EEG). Frontiers in
Human Neuroscience, 11, 481. DOI: https://doi.org/10.3389
/fnhum.2017.00481, PMID: 29033809, PMCID: PMC5624994
Din, N., Melloni, L., Zhang, H., Tian, X., & Poeppel, D. (2016).
Cortical tracking of hierarchical linguistic structures in connected
speech. Nature Neuroscience. Nature Neuroscience, 19(1),
158–164. DOI: https://doi.org/10.1038/nn.4186, PMID:
26642090, PMCID: PMC4809195
Eklund, K., Torppa, M., Aro, M., Leppänen, P. H. T., & Lyytinen, H.
(2015). Literacy skill development of children with familial risk
for dyslexia through grades 2, 3, and 8. Journal of Educational
Psychology, 107(1), 126–140. DOI: https://doi.org/10.1037
/a0037121
Fowler, A. E. (1991). How early phonological development might
set the stage for phoneme awareness. In S. A. Brady & D. P.
Shankweiler (Eds.), Phonological processes in literacy: A tribute
to Isabelle Y. Liberman (pp. 97–117). Lawrence Erlbaum.
Ghitza, O. (2011). Linking speech perception and neurophysiology:
Speech decoding guided by cascaded oscillators locked to the
input rhythm. Frontiers in Psychology, 2. DOI: https://doi.org
/10.3389/fpsyg.2011.00130, PMID: 21743809, PMCID:
PMC3127251
Ghitza, O., Giraud, A.-L., & Poeppel, D. (2013). Neuronal oscilla-
tions and speech perception: Critical-band temporal envelopes
are the essence. Frontiers in Human Neuroscience, 6. DOI:
https://doi.org/10.3389/fnhum.2012.00340, PMID: 23316150,
PMCID: PMC3539830
Ghitza, O., & Greenberg, S. (2009). On the possible role of brain
rhythms in speech perception: Intelligibility of time-compressed
speech with periodic and aperiodic insertions of silence.
Phonetica, 66(1–2), 113–126. DOI: https://doi.org/10.1159
/000208934, PMID: 19390234
Giraud, A.-L., Kleinschmidt, A., Poeppel, D., Lund, T. E.,
Frackowiak, R. S. J., & Laufs, H. (2007). Endogenous cortical
rhythms determine cerebral specialization for speech perception
and production. Neuron, 56(6), 1127–1134. DOI: https://doi.org
/10.1016/j.neuron.2007.09.038, PMID: 18093532
Gogtay, N., Giedd, J. N., Lusk, L., Hayashi, K. M., Greenstein, D.,
Vaituzis, A. C., Nugent, T. F., Herman, D. H., Clasen, L. S., &
Toga, A. W. (2004). Dynamic mapping of human cortical devel-
opment during childhood through early adulthood. Proceedings
of the National Academy of Sciences, 101(21), 8174–8179. DOI:
https://doi.org/10.1073/pnas.0402680101, PMID: 15148381,
PMCID: PMC419576
Goswami, U. (2011). A temporal sampling framework for develop-
mental dyslexia. Trends in Cognitive Sciences, 15(1), 3–10. DOI:
https://doi.org/10.1016/j.tics.2010.10.001, PMID: 21093350
Goswami, U., & Bryant, P. (2016). Phonological skills and learning
to read. Psychology Press. DOI: https://doi.org/10.4324
/9781315695068
Gramfort, A., Luessi, M., Larson, E., Engemann, D. A., Strohmeier,
D., Brodbeck, C., Goj, R., Jas, M., Brooks, T., Parkkonen, L., &
Hämäläinen, M. (2013). MEG and EEG data analysis with MNE-
Python. Frontiers in Neuroscience, 7. DOI: https://doi.org/10
. 3 3 8 9 / f n i n s . 2 0 1 3 . 0 0 2 6 7 , P M I D : 2 4 4 3 1 9 8 6 , P M C I D :
PMC3872725
Gross, J., Hoogenboom, N., Thut, G., Schyns, P., Panzeri, S., Belin,
P., & Garrod, S. (2013). Speech rhythms and multiplexed oscil-
latory sensory coding in the human brain. PLoS Biology, 11(12),
e1001752. DOI: https://doi.org/10.1371/journal.pbio.1001752,
PMID: 24391472, PMCID: PMC3876971
Gross, J., Kujala, J., Hämäläinen, M., Timmermann, L., Schnitzler,
A., & Salmelin, R. (2001). Dynamic imaging of coherent sources:
Studying neural interactions in the human brain. Proceedings of
the National Academy of Sciences, 98(2), 694–699. DOI: https://
doi.org/10.1073/pnas.98.2.694, PMID: 11209067, PMCID:
PMC14650
Hämäläinen, M. S., & Ilmoniemi, R. J. (1994). Interpreting magnetic
fields of the brain: Minimum norm estimates. Medical &
Biological Engineering & Computing, 32(1), 35–42. DOI:
https://doi.org/10.1007/BF02512476, PMID: 8182960
Häyrinen, T., Serenius-Sirve, S., & Korkman, M. (1999). Lukilasse.
Lukemisen, Kirjoittamisen Ja Laskemisen Seulontatestistö
Peruskoulun Ala-Asteen Luokille. Helsinki: Psykologien
Kustannus Oy.
Hyvärinen, A., & Oja, E. (2000). Independent component analysis:
Algorithms and applications. Neural Networks, 13(4–5), 411–430.
DOI: https://doi.org/10.1016/S0893-6080(00)00026-5, PMID:
10946390
Kalashnikova, M., Peter, V., Di Liberto, G. M., Lalor, E. C., &
Burnham, D. (2018). Infant-directed speech facilitates seven-
month-old infants’ cortical tracking of speech. Scientific
Reports, 8(1). DOI: https://doi.org/10.1038/s41598-018-32150-6,
PMID: 30214000, PMCID: PMC6137049
Korkman, M., Kirk, U., & Kemp, S. L. (1998). NEPSY: A developmental
neuropsychological assessment. Psychological Corporation.
Korkman, M., Kirk, U., & Kemp, S. L. (2008). NEPSY-II: Lasten neu-
ropsykologinen tutkimus [NEPSY-II: A developmental neuropsy-
chological assessment]. Psykologien Kustannus Oy.
Kuhl, P. K. (2000). A new view of language acquisition.
Proceedings of the National Academy of Sciences, 97(22),
11850–11857. DOI: https://doi.org/10.1073/pnas.97.22.11850,
PMID: 11050219, PMCID: PMC34178
Leong, V., & Goswami, U. (2014). Assessment of rhythmic entrain-
ment at multiple timescales in dyslexia: Evidence for disruption
to syllable timing. Hearing Research, 308, 141–161. DOI: https://
doi.org/10.1016/j.heares.2013.07.015, PMID: 23916752,
PMCID: PMC3969307
Luo, H., & Poeppel, D. (2007). Phase patterns of neuronal re-
sponses reliably discriminate speech in human auditory cortex.
Neurobiology of Language
251
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Neuron, 54(6), 1001–1010. DOI: https://doi.org/10.1016/j
.neuron.2007.06.004, PMID: 17582338, PMCID: PMC2703451
Molinaro, N., & Lizarazu, M. (2018). Delta(but not theta)-band cor-
tical entrainment involves speech-specific processing. European
Journal of Neuroscience, 48(7), 2642–2650. DOI: https://doi.org
/10.1111/ejn.13811, PMID: 29283465
Molinaro, N., Lizarazu, M., Lallier, M., Bourguignon, M., &
Carreiras, M. (2016). Out-of-synchrony speech entrainment in de-
velopmental dyslexia: Altered cortical speech tracking in dyslexia.
Human Brain Mapping, 37(8), 2767–2783. DOI: https://doi.org
/10.1002/hbm.23206, PMID: 27061643, PMCID: PMC6867425
Müller, V., Gruber, W., Klimesch, W., & Lindenberger, U. (2009).
Lifespan differences in cortical dynamics of auditory perception.
Developmental Science, 12(6), 839–853. DOI: https://doi.org/10
.1111/j.1467-7687.2009.00834.x, PMID: 19840040
Näätänen, R., Lehtokoski, A., Lennes, M., Cheour, M., Huotilainen,
M., Iivonen, A., Vainio, M., Alku, P., Ilmoniemi, R. J., Luuk, A.,
& Allik, J., (1997). Language-specific phoneme representations
revealed by electric and magnetic brain responses. Nature,
385(6615), 432–434. DOI: https://doi.org/10.1038/385432a0,
PMID: 9009189
Näätänen, R., & Picton, T. W. (1986). N2 and automatic versus
controlled processes. Electroencephalography and Clinical
Neurophysiology Supplement, 38, 169–186. PMID: 3466775
Noiray, A., Abakarova, D., Rubertus, E., Krüger, S., & Tiede, M.
(2018). How do children organize their speech in the first years
of life? Insight from ultrasound imaging. Journal of Speech,
Language, and Hearing Research, 61(6), 1355–1368. DOI:
https://doi.org/10.1044/2018_JSLHR-S-17-0148, PMID: 29799996
Noiray, A., Popescu, A., Killmer, H., Rubertus, E., Krüger, S., &
Hintermeier, L. (2019). Spoken language development and the
challenge of skill integration. Frontiers in Psychology, 10. DOI:
https://doi.org/10.3389/fpsyg.2019.02777, PMID: 31920826,
PMCID: PMC6938249
Noiray, A., Wieling, M., Abakarova, D., Rubertus, E., & Tiede, M.
(2019). Back from the future: Nonlinear anticipation in adults’
and children’s speech. Journal of Speech, Language, and
Hearing Research, 62(8S), 3033–3054. DOI: https://doi.org/10
.1044/2019_JSLHR-S-CSMC7-18-0208, PMID: 31465705
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011).
FieldTrip: Open source software for advanced analysis of MEG,
EEG, and invasive electrophysiological data. Computational
Intelligence and Neuroscience, 1–9. DOI: https://doi.org/10
.1155/2011/156869, PMID: 21253357, PMCID: PMC3021840
Pang, E., & Taylor, M. (2000). Tracking the development of the N1
from age 3 to adulthood: An examination of speech and non-
speech stimuli. Clinical Neurophysiology, 111(3), 388–397. DOI:
https://doi.org/10.1016/S1388-2457(99)00259-X
Parviainen, T., Helenius, P., Poskiparta, E., Niemi, P., & Salmelin,
R. (2011). Speech perception in the child brain: Cortical timing
and its relevance to literacy acquisition. Human Brain Mapping,
32(12), 2193–2206. DOI: https://doi.org/10.1002/ hbm.21181,
PMID: 21391257, PMCID: PMC6870499
Parviainen, T., Helenius, P., & Salmelin, R. (2019). Children show
hemispheric differences in the basic auditory response properties.
Human Brain Mapping, 40(9), 2699–2710. DOI: https://doi.org/10
.1002/hbm.24553, PMID: 30779260, PMCID: PMC6865417
Peelle, J. E., & Davis, M. H. (2012). Neural oscillations carry speech
rhythm through to comprehension. Frontiers in Psychology, 3,
320. DOI: https://doi.org/10.3389/fpsyg.2012.00320, PMID:
22973251, PMCID: PMC3434440
Peelle, J. E., Gross, J., & Davis, M. H. (2013). Phase-locked re-
sponses to speech in human auditory cortex are enhanced during
comprehension. Cerebral Cortex, 23(6), 1378–1387. DOI:
https://doi.org/10.1093/cercor/ bhs118, PMID: 22610394,
PMCID: PMC3643716
Pena, M., Maki, A., Kovacic, D., Dehaene-Lambertz, G., Koizumi,
H., Bouquet, F., & Mehler, J. (2003). Sounds and silence: An op-
tical topography study of language recognition at birth.
Proceedings of the National Academy of Sciences, 100(20),
11702–11705. DOI: https://doi.org/10.1073/pnas.1934290100,
PMID: 14500906, PMCID: PMC208821
Poeppel, D. (2003). The analysis of speech in different temporal
integration windows: Cerebral lateralization as ‘asymmetric
sampling in time’. Speech Communication, 41(1), 245–255.
DOI: https://doi.org/10.1016/S0167-6393(02)00107-3
Poeppel, D. (2014). The neuroanatomic and neurophysiological
infrastructure for speech and language. Current Opinion in
Neurobiology, 28, 142–149. DOI: https://doi.org/10.1016/j
.conb.2014.07.005, PMID: 25064048, PMCID: PMC4177440
Poeppel, D., & Assaneo, M. F. (2020). Speech rhythms and their
neural foundations. Nature Reviews Neuroscience, 21(6), 322–334.
DOI: https://doi.org/10.1038/s41583-020-0304-4, PMID: 32376899
Ponton, C. W., Eggermont, J. J., Khosla, D., Kwong, B., & Don, M.
(2002). Maturation of human central auditory system activity:
Separating auditory evoked potentials by dipole source model-
ing. Clinical Neurophysiology, 113(3), 407–420. DOI: https://
doi.org/10.1016/S1388-2457(01)00733-7
Ponton, C. W., Eggermont, J. J., Kwong, B., & Don, M. (2000).
Maturation of human central auditory system activity: Evidence
from multi-channel evoked potentials. Clinical Neurophysiology,
111(2), 220–236. DOI: https://doi.org/10.1016/S1388-2457(99)
00236-9, PMID: 10680557
Popescu, A., & Noiray, A. (2019, November 7–10). Reading profi-
ciency and phonemic awareness as predictors for coarticulatory
gradients in children. In Proceedings of the 44th Boston
University Conference on Language Development, Boston, MA.
Ríos-López, P., Molinaro, N., Bourguignon, M., & Lallier, M.
(2020). Development of neural oscillatory activity in response
to speech in children from 4 to 6 years old. Developmental
Science 23(6), e12947. DOI: https://doi.org/10.1111/desc
.12947, PMID: 32043677, PMCID: PMC7685108
Taulu, S., & Kajola, M. (2005). Presentation of electromagnetic
multichannel data: The signal space separation method. Journal
of Applied Physics, 97(12), 124905. DOI: https://doi.org/10.1063
/1.1935742
Taulu, S., Simola, J., & Kajola, M. (2005). Applications of the sig-
nal space separation method. IEEE Transactions on Signal
Processing, 53(9), 3359–3372. DOI: https://doi.org/10.1109/TSP
.2005.853302
Telkemeyer, S., Rossi, S., Koch, S. P., Nierhaus, T., Steinbrink, J.,
Poeppel, D., Obrig, H., & Wartenburger, I. (2009). Sensitivity
of newborn auditory cortex to the temporal structure of sounds.
Journal of Neuroscience, 29(47), 14726–14733. DOI: https://doi
.org/10.1523/ JNEUROSCI.1246-09.2009, PMID: 19940167,
PMCID: PMC6666009
Telkemeyer, S., Rossi, S., Nierhaus, T., Steinbrink, J., Obrig, H., &
Wartenburger, I. (2011). Acoustic processing of temporally mod-
ulated sounds in infants: Evidence from a combined near-infrared
spectroscopy and EEG study. Frontiers in Psychology, 1. DOI:
https://doi.org/10.3389/fpsyg.2011.00062, PMID: 21716574,
PMCID: PMC3110620
Torgesen, J. K., Wagner, R. K., Rashotte, C. A., Rose, E.,
Lindamood, P., Conway, T., & Garvan, C. (1999). Preventing
reading failure in young children with phonological processing
disabilities: Group and individual responses to instruction.
Neurobiology of Language
252
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
/
.
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Age-related differences in coherence
Journal of Educational Psychology, 91(4), 579. DOI: https://doi
.org/10.1037/0022-0663.91.4.579
Uhlhaas, P. J., Roux, F., Rodriguez, E., Rotarska-Jagiela, A., & Singer,
W. (2010). Neural synchrony and the development of cortical net-
works. Trends in Cognitive Sciences, 14(2), 72–80. DOI: https://
doi.org/10.1016/j.tics.2009.12.002, PMID: 20080054
Vihman, M. M. (2017). Learning words and learning sounds: Advances
in language development. British Journal of Psychology, 108(1),
1–27. DOI: https://doi.org/10.1111/bjop.12207, PMID: 27449816
Wechsler, D. (2003a). Wechsler preschool and primary scale of in-
telligence – Third Edition ( WPPSI-III). NCS Pearson, Inc., USA.
Psykologien Kustannus Oy, Helsinki. DOI: https://doi.org/10
.1037/t15177-000
Wechsler, D. (2003b). WISC-IV: Administration and scoring manual.
Psychological Corporation.
Wechsler, D. (2008). Wechsler adult intelligence scale – Fourth Edition
(WAIS–IV). NCS Pearson. DOI: https://doi.org/10.1037/t15169-000
Ziegler, J. C., & Goswami, U. (2005). Reading acquisition, develop-
mental dyslexia, and skilled reading across languages: A psycho-
linguistic grain size theory. Psychological Bulletin, 131(1), 3–29.
DOI: https://doi.org/10.1037/0033-2909.131.1.3, PMID:
15631549
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
e
d
u
n
o
/
l
/
l
a
r
t
i
c
e
-
p
d
f
/
/
/
/
2
2
2
2
6
1
9
1
5
9
4
3
n
o
_
a
_
0
0
0
3
3
p
d
.
/
l
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurobiology of Language
253