RECHERCHE
The ascending arousal system promotes optimal
performance through mesoscale network
integration in a visuospatial attentional task
Gabriel Wainstein1
, Daniel Rojas-Líbano2
, Vicente Medel1,3, Dag Alnæs4,5
,
Knut K. Kolskår4,6,7
, Tor Endestad6,8,9
, Bruno Laeng6,8
, Tomas Ossandon3,10
,
Nicolás Crossley3, Elie Matar1
, and James M. Shine1,11
1Brain and Mind Centre, University of Sydney, Sydney, NSW, Australia
2Centro de Neurociencia Humana y Neuropsicología, Facultad de Psicología, Universidad Diego Portales, Santiago, Chili
3Département de psychiatrie, École de médecine, Pontificia Universidad Católica de Chile, Santiago, Chili
4NORMENT, Division of Mental Health and Addiction, University of Oslo, and Oslo University Hospital, Oslo, Norway
5Bjørnnes College, Oslo, Norway
6Département de psychologie, University of Oslo, Oslo, Norway
7Sunnaas Rehabilitation Hospital HT, Nesodden, Norway
8RITMO Centre for Interdisciplinary Studies in Rhythm, Time, and Motion, University of Oslo, Oslo, Norway
9Helgelandssykehuset Mosjøen, Helse Nord, Norway
10Institute for Biological and Medical Engineering, Schools of Engineering, Medicine and Biological Sciences,
Pontificia Universidad Católica de Chile, Santiago, Chili
11Centre for Complexity, University of Sydney, Sydney, NSW, Australia
Mots clés: Pupil diameter, Locus coeruleus, Network integration, Noradrenergic system,
Neuromodulation, Attention, Mental effort, IRMf
ABSTRAIT
Previous research has shown that the autonomic nervous system provides essential constraints
over ongoing cognitive function. Cependant, there is currently a relative lack of direct empirical
evidence for how this interaction manifests in the brain at the macroscale level. Ici, nous
examine the role of ascending arousal and attentional load on large-scale network dynamics
by combining pupillometry, functional MRI, and graph theoretical analysis to analyze data
from a visual motion-tracking task with a parametric load manipulation. We found that
attentional load effects were observable in measures of pupil diameter and in a set of brain
regions that parametrically modulated their BOLD activity and mesoscale network-level
l'intégration. En outre, the regional patterns of network reconfiguration were correlated
with the spatial distribution of the α2a adrenergic receptor. Our results further solidify the
relationship between ascending noradrenergic activity, large-scale network integration, et
cognitive task performance.
RÉSUMÉ DE L'AUTEUR
In our daily lives, it is usual to encounter highly demanding cognitive tasks. They have been
traditionally regarded as challenges that are solved mainly through cerebral activity, specifically
via information-processing steps carried by neurons in the cerebral cortex. Activity in cortical
networks thus constitutes a key factor for improving our understanding of cognitive processes.
Cependant, recent evidence has shown that evolutionary older players in the central nervous
système, such as brain stem’s ascending modulatory systems, might play an equally important
role in diverse cognitive mechanisms. Our article examines the role of the ascending arousal
un accès ouvert
journal
Citation: Wainstein, G., Rojas-Líbano,
D., Medel, V., Alnæs, D., Kolskår, K. K.,
Endestad, T., Laeng, B., Ossandon, T.,
Crossley, N., Matar, E., & Shine, J.. M..
(2021). The ascending arousal system
promotes optimal performance through
mesoscale network integration in a
visuospatial attentional task. Réseau
Neurosciences, 5(4), 890–910. https://est ce que je
.org/10.1162/netn_a_00205
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00205
Informations complémentaires:
https://doi.org/10.1162/netn_a_00205
https://github.com/gabwainstein/MOT
https://doi.org/10.6084/m9
.figshare.13244504.v1
Reçu: 15 Mars 2021
Accepté: 2 Août 2021
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
James M.. Shine
mac.shine@sydney.edu.au
Éditeur de manipulation:
Christopher Honey
droits d'auteur: © 2021
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
un
_
0
0
2
0
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
system on large-scale network dynamics by combining pupillometry, functional MRI, and graph
theoretical analysis.
Réseau:
Theoretical representation of a
système. Each part is represented as a
node and the connection between
nodes as edges.
L'intégration:
Number of connections of a region
or set of regions outside its own
module.
Locus coeruleus:
Principal noradrenergic
neuromodulatory nuclei located
in the brain stem. Projects towards
the central nervous system.
Noradrenaline:
A monoaminergic neurotransmitter
that modulates the neuronal activity
of the target populations in the
nervous system.
INTRODUCTION
Cognitive processes emerge from the dynamic interplay between diverse mesoscopic brain
systèmes (Shine, 2021; Shine et al., 2016). Ainsi, the neural activity supporting cognition does
not exist in a vacuum, but instead is deeply embedded within the ongoing dynamics of the
physiological networks of the body (Varela et al., 2001). En particulier, the neural processes
underlying cognition are shaped and constrained by the ascending arousal system, whose ac-
tivity acts to facilitate the integration between internal states and external contingencies
(Parvizi & Damasio, 2001). Timely and selective interactions between the ascending arousal
system and the network-level configuration of the brain are thus likely to represent crucial
constraints on cognitive and attentional processes. Encore, despite these links, we currently have
a relatively poor understanding of how the ascending arousal system helps the brain as a
whole to functionally reconfigure during cognitive processes, such as attention, pour
facilitate effective cognitive performance.
Recent evidence has linked higher order cognitive functions in the brain to the intersection
between whole-brain functional network architecture and the autonomic arousal system
(Alnæs et al., 2015; Alnæs et al., 2014; Munn et al., 2021; Shine et al., 2016; Shine, Hearne,
et coll., 2019). Central to these relationships is the unique neuroanatomy of the ascending
noradrenergic system. Par exemple, the pontine locus coeruleus, which is a major hub of the
ascending arousal system, sends widespread projections to the rest of the brain (Samuels &
Szabadi, 2008). Upon contact, adrenergic axons release noradrenaline, which acts as a ligand
on three types of post- and presynaptic adrenergic receptors (c'est à dire., α1, α2, and β). The functional
effects of each of these receptors depend on their differential sensitivities to noradrenaline
(affinities for the ligand differ across receptors: α2 > α1 > β) and intracellular cascades, aussi
as their neuronal and regional distributions (Aston-Jones & Waterhouse, 2016; Bouret & Sarah,
2005; Robbins & Arnsten, 2009; Samuels & Szabadi, 2008; Sarah, 2009; Shine, 2019). Par
modulating the excitability of targeted regions, the locus coeruleus can effectively coordinate
neural dynamics across large portions of the cerebral cortex (Shine et al., 2021; X. J.. Wang,
2020). Cependant, it is challenging to noninvasively track the engagement of the locus coeru-
leus during whole-brain neuroimaging and cognitive task performance.
Heureusement, it has been widely shown that the pupil diameter directly responds to changes
in the activity of the locus coeruleus, and thus serves as an indirect, noninvasive measure of
the noradrenergic system (Aston-Jones & Cohen, 2005; S. Joshi et al., 2016). Spécifiquement, pupil
diameter has been shown to indirectly monitor the neuromodulatory influences of the ascend-
ing arousal system on a variety of different brain regions (Alnæs et al., 2014; Liu et al., 2017;
Sarah, 2009; van den Brink et al., 2016). De plus, noradrenergic-mediated dilations in pupil
diameter have been shown to effectively track the allocation of attentional resources
(Gilzenrat et al., 2010; Kahneman & Beatty, 1966; Wainstein et al., 2017), in addition to both
physical and mentally effortful processes (Mulder, 2012; Varazzani et al., 2015). Fast, phasic
changes in pupil diameter have also been shown to directly relate to changes in the activity of
the locus coeruleus (S. Joshi et al., 2016; Murphy et al., 2016; Reimer et al., 2014). Pendant que là
Neurosciences en réseau
891
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
un
_
0
0
2
0
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
is some evidence that pupil diameter covaries with other subcortical systems (S. Joshi & Gold,
2020), such as the cholinergic and serotoninergic system (Cazettes et al., 2020), the physio-
logical mechanism for these effects is more opaque, and there is also clear causal evidence
linking stimulation of the locus coeruleus to dilation of the pupil (Liu et al., 2017; Zerbi et al.,
2019). Despite these insights, several questions remain unanswered regarding how these pro-
cesses are related to the complex architecture of the brain (Shenhav et al., 2017). Par exemple,
the processes by which the ascending arousal system modulates the functional dynamics of
brain networks to facilitate attention, decision-making, and optimal behavioral performance
have only begun to be explored (de Gee et al., 2017; Shine, Breakspear, et coll., 2019; Shine
et coll., 2018; Zerbi et al., 2019).
To examine these relationships in more detail, participants performed a motion-tracking
task (top panel of Figure 1A) involving four levels of increasing attentional load, ce qui était
modulated by manipulating the number of items required to covertly attend to over an 11-s
tracking period. Spécifiquement, subjects were instructed to covertly track the movement of sev-
eral preidentified targets (two to five) in a field of nontarget stimuli (10 in total, including tar-
gets; voir la figure 1). To investigate the network topological signatures of performing this task,
we collected concurrent BOLD fMRI and pupillometry data. We hypothesized that, if increas-
ing mental effort led to the reconfiguration of large-scale network architecture via the ascend-
ing arousal system, then the number of items required to be tracked over time (c'est à dire., le
attentional load) should relate to (un) increased pupil diameter; (b) heightened BOLD activity
within attentional networks; et (c) augmented topological integration. Aussi, we predicted
that individual differences in pupil diameter should track individual differences in effective
attentional performance and decision processes (de Gee et al., 2017; de Gee et al., 2014;
Donner et al., 2000). Enfin, we tested whether the regional patterns of network configuration
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
un
_
0
0
2
0
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
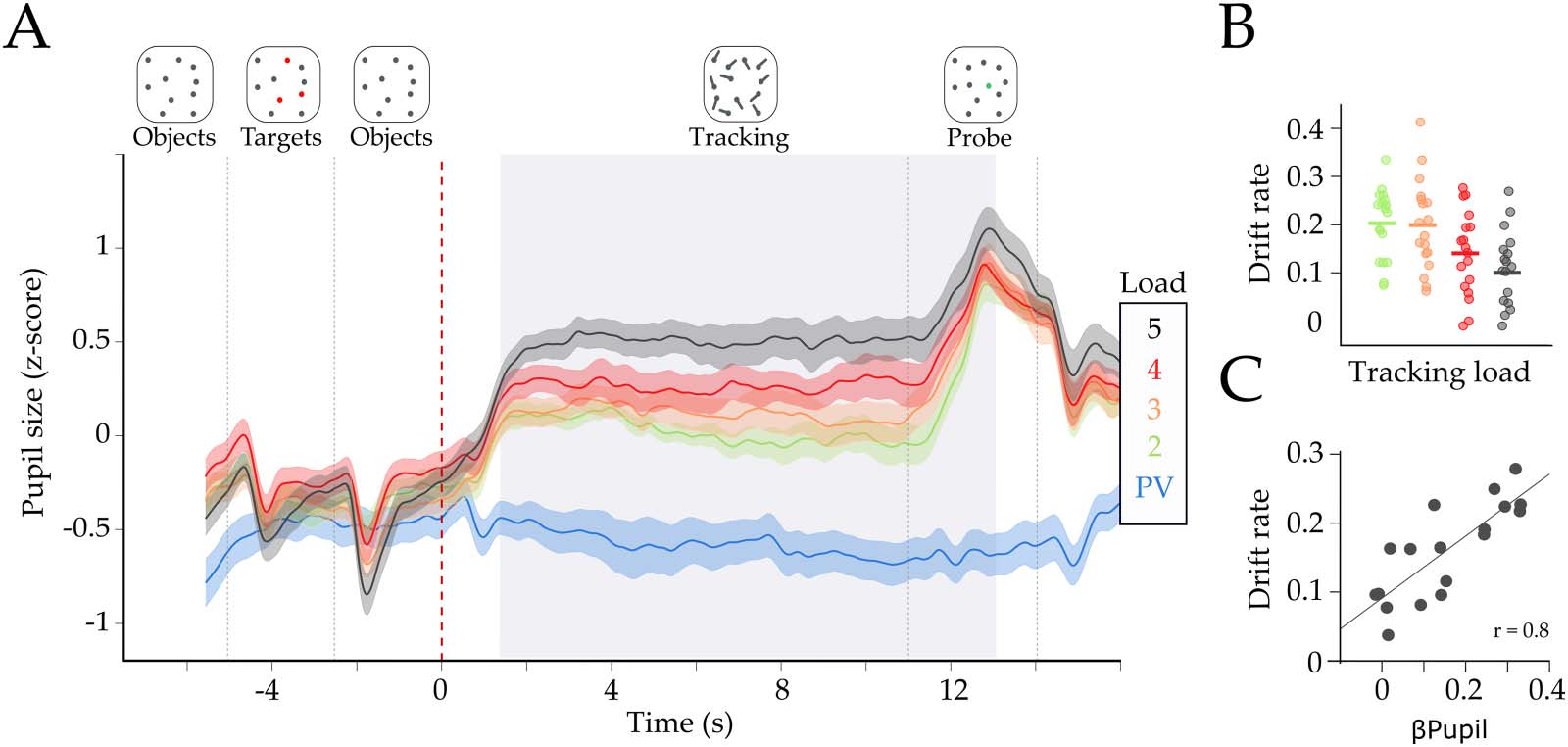
Chiffre 1. Effect of task difficulty on pupil diameter. (UN) Group average (z-score) pupil diameter time series for each load condition. Colors
represent passive viewing (PV) in blue, and Loads 2 à 5 in green, orange, red, and black, respectivement. The shaded area represents the standard
error of the mean. We observed an average increase in pupil diameter, during tracking, with each load condition. The light gray area repre-
sents time points with significant parametric effect (β
pupil > 0; FDR corrected at p < 0.01). Dotted lines represent the onset of each trial event
(shown in the top part of the figure). The red dotted line (time = 0) is the tracking onset period when the dots began to move. (B) Drift rate in
−7). (C) Pearson
each load condition. Each dot is the drift rate for each subject and load (mean β
−4). The
correlation between the pupil parametric effect of load (β
x-axis is the mean beta estimate of the pupillary load effect of the significative time window (β
pupil), and the y-axis represents the mean drift
rate across loads.
pupil) with the average drift rate across subjects (rdrift = 0.8, p = 1.0 × 10
Drift = −0.03, t(17) = −7.43, p = 9.7 × 10
Network Neuroscience
892
The ascending arousal system shapes mesoscale network integration
were predicted by the distribution of a predefined adrenergic receptor density atlas (Fornito
et al., 2019; Richiardi et al., 2015; Shine, Breakspear, et al., 2019; Zerbi et al., 2019). Our
results confirm these predictions, and hence provide a mechanistic link between network
topology, ascending noradrenergic arousal, and attentional load.
RESULTS
Drift rate:
A marker of the speed of the
accumulation of decision evidence
during the decision-making process.
RT = 0.06, t(17) = 5.10, p = 8.8 × 10
The Relationship Between Sympathetic Tone and Attentional Processing
Consistent with previous work (Alnæs et al., 2014), our two-level analysis—linear regression
within each subject, and a two-tailed t test between subjects—found that task performance
(i.e., correct responses) decreased with attentional load (mean β
Acc = −6.66; t(17) = −5.19,
−5; Supplementary Figure S1B) while the reaction time (RT) increased with atten-
p = 7.2 × 10
−5). We expanded on this result by
tional load (mean β
translating performance into EZ-diffusion model parameters. Roughly, this approach uses
the accuracy and reaction time distribution to estimate three latent parameters (de Gee et al.,
2014): drift rate, a marker of the accumulation of decision evidence (Equation 1); boundary
criteria, the amount of evidence required to make a decision (Equation 2); and non-decision
time, the epoch spent processing the tasks perceptually (Equation 3). The advantages of using
this model are twofold: first, there are well-known links between the parameters to decision-
making processes (Ratcliff et al., 2016; Ratcliff et al., 2015), pupil diameter (Murphy et al.,
2016; Murphy et al., 2014) and network reconfiguration (Shine et al., 2016); second, drift rate
accounts for the accuracy–reaction time trade-off, as it takes into consideration both accuracy
and the variability in reaction time into its calculation. In this way, our approach offers a better
approximation of the ongoing computational processing during the task than does accuracy and
RT (Ratcliff et al., 2015; Wagenmakers et al., 2007). Using this approach, we observed a
decrease in both the boundary criteria (β
Bound = −0.01, t(17) = −2.70, p = 0.015) and the drift
−7; Figure 1B), and an increase in the
rate (mean β
−5) with increasing attentional load.
non-decision time (mean β
Drift = −0.03, t(17 ) = −7.43, p = 9.7 × 10
nd = 0.07, t(17) = 5.32, p = 5.5 × 10
By calculating the linear effect of load on pupil size across a moving average window of
160 ms (see Methods), we observed a main effect of increased pupil diameter across both the
tracking and the probe epochs (β
pupil > 0, pFDR < 0.01; light gray area in Figure 1A depicts
significant epochs of time during the task; and in Supplementary Figure S1A shows the group
average β
pupil time series). We also observed a positive correlation between mean β
pupil during
the significant period (for simplicity we will refer to this value as βpupil) to the mean drift rate,
−4;
mean boundary criteria, and accuracy across all loads (Pearson’s rdrift = 0.8, p = 1.0 × 10
−4).
−3, Supplementary Figure S1C; rBound = 0.71, p = 9 × 10
Figure 1C; racc = 0.68, p = 1.5 × 10
The same relationships were not observed with non-decision time (Pearson’s rnd = −0.31, p =
0.19). Additionally, we analyzed whether this effect was present both within and between sub-
jects in a trial-by-trial manner. To this end, we created a logistic linear mixed model (Equation 6)
to test whether pupil diameter was a predictor of performance (i.e., correct or incorrect re-
sponse), as we would expect that incorrect responses should relate to decreased pupil diameter
in difficult trials. We used the average pupil diameter within each trial of Load 4 and 5 (to
account for the ceiling effect of Load 2 and 3) as regressors and subject as a grouping variable.
We found a statistically significant fixed effect of pupil diameter on performance within each trial
−4; t(286) = 2.48; p = 0.013). Furthermore, we analyzed the random-effect
(β = 0.0127 ± 5 × 10
coefficients, which are the dispersion of the regressor across the grouping variable from the
fixed regressor (in this case there is one value per subject), to assess the role of average across
task performance. We found that the random effect covaried with the average performance
−5; Drift: Pearson’s
and drift rate of each subject (Accuracy: Pearson’s r = 0.73, p = 8 × 10
Network Neuroscience
893
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
−5), suggesting that trial-by-trial pupil diameter was a better predictor of
r = 0.73, p = 5 × 10
performance (i.e., correct or incorrect) on subjects with higher average performance in com-
parison to subjects with lower performance across the task. In conclusion, these results suggest
that attentional load manipulation and pupil dilation covaried with performance on this atten-
tionally demanding task both within and between subjects.
Network Integration Increases as a Function of Attentional Load
Based on previous studies, we hypothesized that an increase in attentional load should recruit
a distributed functional network architecture (Alnæs et al., 2014), heightening network inte-
gration (Shine, 2019; Shine et al., 2016; Shine, Breakspear, et al., 2019). To test this hypoth-
esis, we implemented a hierarchical topological network analysis (Bassett et al., 2010;
Meunier et al., 2010; Meunier et al., 2009) on the average time-resolved functional connec-
tivity matrix calculated across the tracking period of the task. Our analysis identified a subnet-
work of tightly interconnected regions that were part of attentional, somatomotor, and
cerebellar network (red in Figure 2) that increased its BOLD activity after the tracking onset
(Figure 2F). The tightly integrated regions were diversely connected to a separate frontoparietal
submodule (blue in Figure 2) that was less active during the trial. Two remaining submodules
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
Module:
Group of nodes that have more
connection strength between them
than to the rest of the system.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
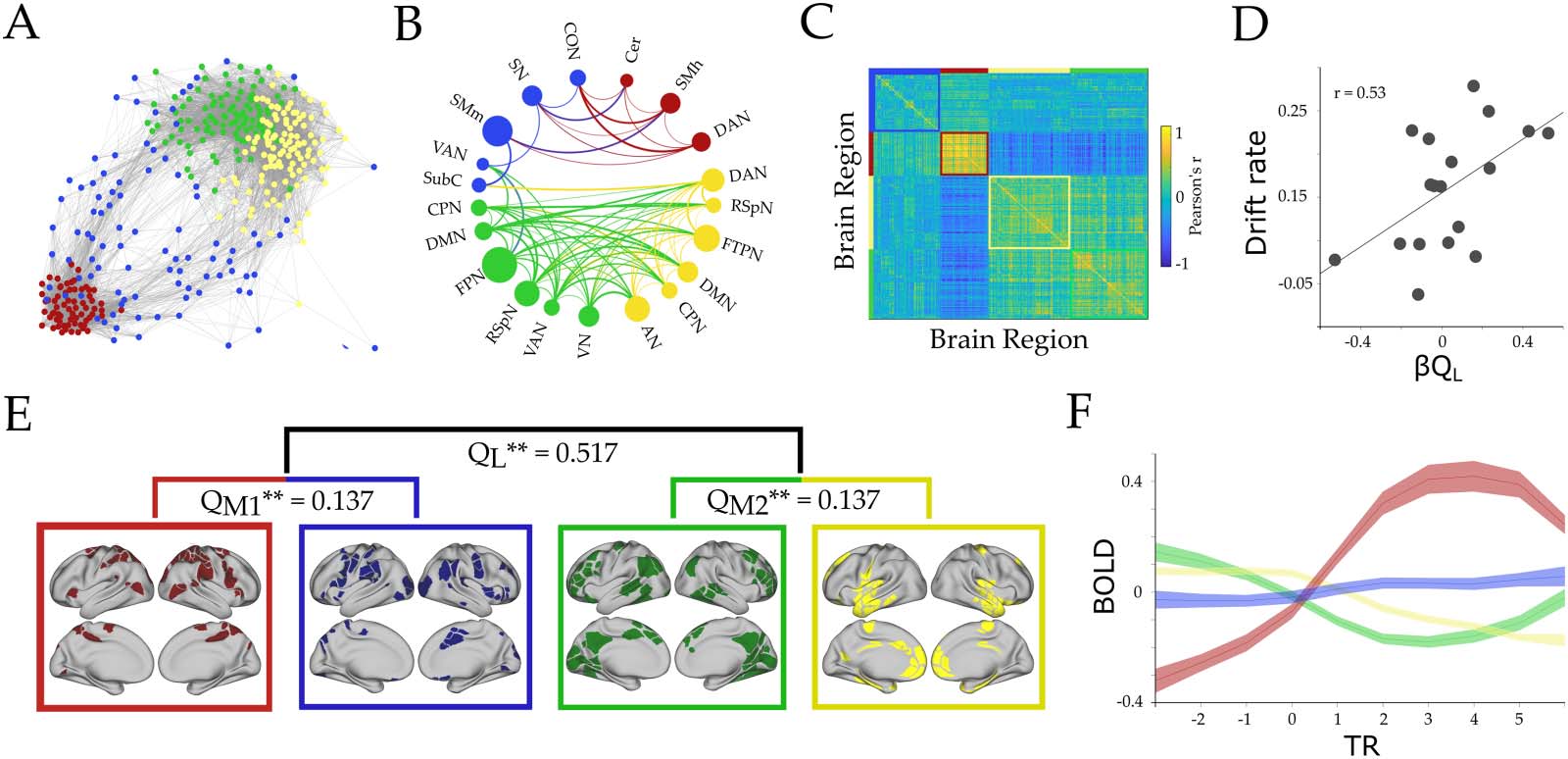
Figure 2. Hierarchical functional topology analysis of the brain during tracking across all loads. We observed two large-scale modules and
two mesoscale modules within each larger module (Module 1 [M1, red/blue] and Module 2 [M2, green/yellow], respectively): M1 corre-
sponded to predominantly attentional and somatomotor network, and M2 to frontoparietal network (FPN) and default mode network
(DMN), among others (B and E). (A) Forced directed plot representation of the average cluster across subjects. Edges stronger than 0.3 are
shown. Each color represents a unique submodule. (B) A circle plot representing the resting-state regions that were included within each
submodule, with networks with >30% of regions in each submodule shown in the plot. The diameter of the circles corresponds to the per-
centage of network regions that participated in that cluster. Connection width relates to average positive connection strength (functional con-
nectivity); cependant, only connections with r > 0.1 are shown. (C) Connectivity matrix (Pearson’s r) between all pairs of regions ordered by
module assignments—note the strong anticorrelation between the red and green/yellow submodules. (D) Correlation between parametric load
effect on large-scale modularity (β
Q value) and drift rate (Pearson’s r = 0.53; p = 0.022). (E) Hierarchical analysis representation: QL, QM1, et
QM2 represent the modularity value for each level (QL large-scale, and QM1–M2 mesoscale level), et ** represents the probability of finding
this value when running a null model ( p = 0 for all three modularity values). The brain maps correspond to the cortical regions associated with
each submodule. (F) BOLD mean effect for each subcluster. Each line represents the group average, and shaded areas are the standard error of
the mean. X-axis is repetition time (TR) centered around tracking onset (TR = 0). DAN, dorsal attention; VN, visuel; FPN, frontoparietal; SN,
salience; CO, cingulo-opercular; VAN, ventral attention; SMm, somatomotor mouth; SMh, somatomotor hand; RSpN, retrosplenial; FTP, fron-
totemporal; DMN, default mode; AN, auditory; CPN, cinguloparietal; SubC, subcortex; Cer, cerebellar.
Neurosciences en réseau
894
The ascending arousal system shapes mesoscale network integration
Modularity:
Quality function that represents the
amount of connection of nodes
within each module to nodes in
the same module.
Load effect:
Statistical linear effect of the task load
(c'est à dire., difficulty) on a given dependent
variable.
(yellow and green in Figure 2) showed a negative BOLD response during the tracking period
and were part of a diverse set of networks. Fait intéressant, 81% of the frontoparietal network
(FPN) and all the default mode network (DMN) were found to be within this less active group
(see Supplementary Table S2 for the complete list of regions and submodule assignments).
Contrary to expectations, we did not observe significant parametric topological change
(c'est à dire., modularity, Q) at the macroscopic level as a function of attentional load ( p > 0.05 pour
all TRs, Supplementary Figure S2A). Cependant, when analyzing the correlation between mod-
ularity and performance measures (c'est à dire., accuracy, drift rate, and pupil diameter), we observed
that an increase in the large-scale modularity load effect (c'est à dire., higher modularity with load, β
QL)
positively correlated with higher mean drift rate (Pearson’s r = 0.53; p = 0.022; Figure 2D),
mean accuracy (Pearson’s r = 0.61; p = 0.007; Supplementary Figure S3A), but was indepen-
dent from βpupil (Pearson’s r = 0.43; p = 0.073). These results suggested that the system
reconfigured during tracking towards increasing modularity, which in turn affected the effi-
cient encoding of the ongoing task during tracking and hence, the decision-making process
during the task probe.
Upon closer inspection of the data (Figure 2C), we observed a substantial number of nodes
that were playing an integrative role during task performance, albeit at a finer resolution than
the initial analysis suggested. We performed the modularity assignment within each large-
scale module. The hierarchical analysis resulted in two pairs of submodules at the mesoscale
level with a significant modularity (compared with 100 random graphs with preserved signed
degree distribution; QM1 = 0.137, p = 0; QM2 = 0.137, p = 0; Figure 2E). Spécifiquement, the red
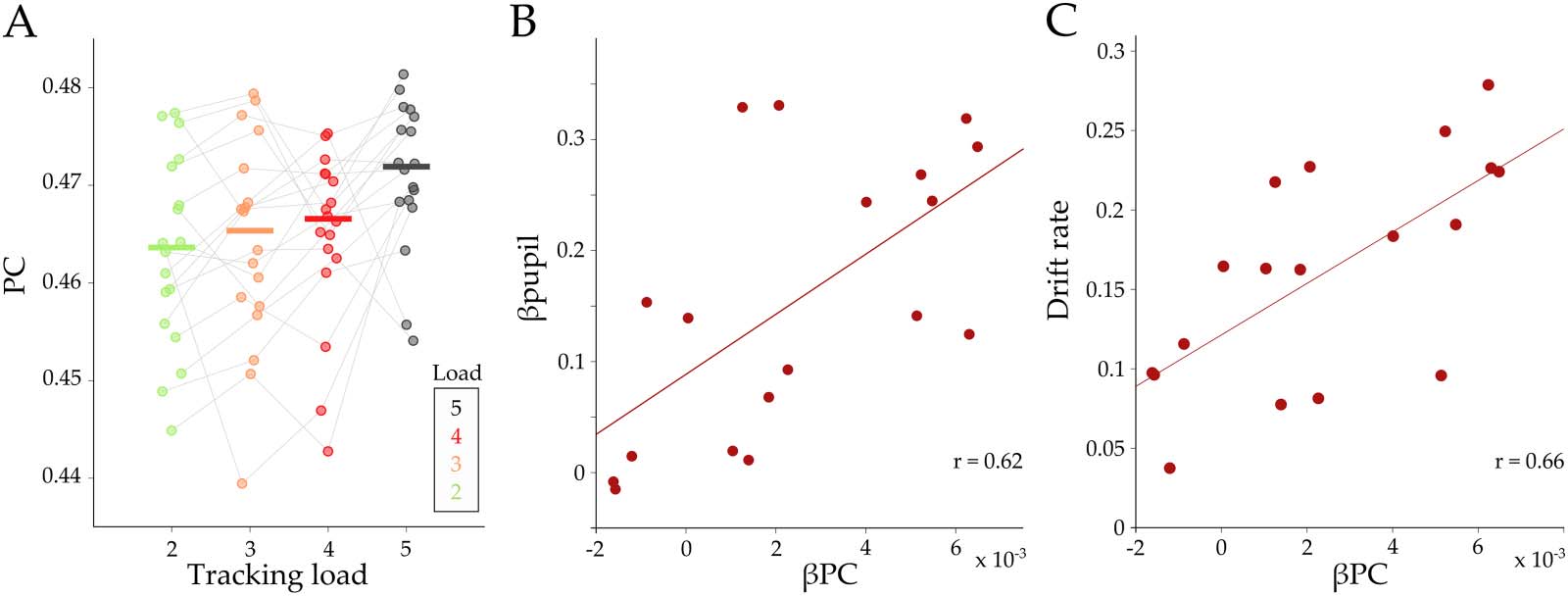
submodule was found to selectively increase its participation coefficient (PC) at the mesoscale
level (c'est à dire., by increasing the connection weights to the blue submodule in comparison with
intramodular connections; Équation 5) as a function of increasing attentional load (β
PC =
−3, t(17) = 3.57; p = 0.002; Figure 3A). En plus, the extent of integration in the
2.4 × 10
red submodule was positively correlated across subjects with βpupil (Pearson’s r = 0.62, p =
0.006; Figure 3B), drift rate (Pearson’s r = 0.66, p = 0.002; Figure 3C), and accuracy (Pearson’s
r = 0.57, p = 0.012, Supplementary Figure S3B). Surtout, these relationships were found
to be specific to the red submodule (blue: Pearson’s r = −0.02, p = 0.936; yellow: Pearson’s
r = −0.011, p = 0.965; vert: Pearson’s r = −0.12, p = 0.617).
Chiffre 3. Relationships between load effect on participation, pupil load effect, and drift rate. (UN) Average participation coefficient (PC) pour
each load, for the red module, during tracking. Each color represents the corresponding tracking load (depuis 2 à 5). Gray lines correspond to
chaque sujet. (B–C) A regression parameter (βPC) was calculated for each subject and then correlated to βpupil (B; r = 0.62; p = 0.006) and drift
rate (C; r = 0.66; p = 2.4 × 10
−3). Each circle corresponds to the mean value per subject.
Neurosciences en réseau
895
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
un
_
0
0
2
0
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
−3 ± 3 × 10
Based on these results, we implemented a linear mixed model (Équation 7), using the sub-
jects’ average pupil response within each load as a regressor and the average participation of
the red submodule as the dependent variable, with grouping by subject. Using this approach,
−3, t(70) =
we observed a significant fixed effect of pupil diameter on PC (β = 7.6 × 10
2.60, p = 0.011). En outre, the random-effect coefficients (c'est à dire., the between-subject varia-
tion of the regressor value) correlated positively with accuracy (Pearson’s r = 0.47, p = 0.048)
and drift rate (Pearson’s r = 0.62, p = 0.005), suggesting that subjects with a strong relationship
between red module integration and pupil diameter have better behavioral outcomes. Nous
then correlated the red βPC to the load effect on large-scale modularity (βQL, Figure 2D)
and observed a significant positive correlation (Pearson’s r = 0.59, p = 0.009). Enfin, given
that both topological parameters were correlated with drift rate and also with each other, nous
performed a partial correlation between drift rate and βPC controlling by βQL (r = 0.51, p =
0.034), and the partial correlation between drift rate and βQL controlling by βPC (r = 0.36, p =
0.145). This suggests that drift rate is correlated with the mesoscale integration of the red sub-
module, but less so with increases in large-scale modularity. Ainsi, although the macroscale
network did not demonstrate increased integration per se, the relative amount of mesoscale
integration within the red community was associated with increased performance (c'est à dire., drift
rate) and sympathetic arousal (c'est à dire., pupil diameter), both between and within subjects. Dans ce
chemin, these results provide a direct relationship between the effect of attention load on pupil-
lometry, drift rate, and a trade-off between large-scale segregation and mesoscale network
l'intégration.
Network Mesoscale Integration and Adrenergic Receptor Density
Given the relationship between mental effort, noradrenergic tone, and pupil dilation (Alnæs
et coll., 2014; S. Joshi et al., 2016; McGinley et al., 2015; Reimer et al., 2014; Varazzani et al.,
2015), the results of our analyses strongly suggested that the adrenergic system is involved in
the mesoscale network reconfiguration observed during attentional tracking. The locus coer-
uleus can impact the cortical system in multiple ways, both through direct release of noradren-
aline onto cortical neurons, and through the modulation of subcortical regions (such as the
thalamic nuclei) with concurrent impact on the cortical dynamic. Surtout, in either case,
the modulation is dependent on the noradrenergic receptors subtypes, which have different
sensitivities to noradrenaline (Robbins & Arnsten, 2009; M.. Wang et al., 2007) and variable
expression in the cerebral cortex (Santana & Artigas, 2017; Zilles & Palomero-Gallagher,
2017), and also belong to distinct classes (c'est à dire., α1, α2, and β receptors). En particulier, le
α2a has been previously associated with working memory, adaptive gain, and effective atten-
tion (Arnsten et al., 2012; Robbins & Arnsten, 2009; M.. Wang et al., 2007). To gain a deeper
insight into the role of α2a receptors in mesoscale integration during attentional tracking, nous
extracted the regional expression of the ADRA2A gene (which codes for α2a adrenoceptors)
from the Allen Human Brain Atlas repository (Gryglewski et al., 2018; Hawrylycz et al., 2012),
and compared the cortical regional expression of this gene with the brain activity patterns
identified in our network analysis (Figure 2E).
Based on the relationships between pupil diameter (Chiffre 1), topological signatures
(Chiffre 2), and task performance (Chiffre 3), and the known link between these variables
and engagement of the noradrenergic system, we hypothesized that the different modules
and submodules that we observed should have different densities of neuromodulatory recep-
tors to account for the differential patterns across the network. To test this hypothesis, we con-
ducted a two-tailed t test in each hierarchical level comparing the density of the ADRA2A
expression between modules. To account for spatial autocorrelation, we generated 5,000
α2a:
Noradrenergic G-protein coupled
receptor. It is the main adrenergic
presynaptic and postsynaptic
receptor subtype in the human brain.
ADRA2A atlas:
RNA expression atlas of the α2a. Il
is used as an indirect marker of
the density of the α2a receptor.
Neurosciences en réseau
896
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
un
_
0
0
2
0
5
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
surrogate maps with the same spatial autocorrelation of the ADRA2A map, calculated a t sta-
tistic for each surrogate, and evaluated the probability of finding the observed t statistic against
the null distribution (Burt et al., 2020; Markello & Misic, 2021). We indeed observed signifi-
cant differences between modules at the mesoscale level. Spécifiquement, we found significant
−4, pSA =
differences between the blue and yellow submodules (t(194) = 3.82, p = 2 × 10
0.02) and the differences between green and yellow submodules (t(177) = −4.47, p = 1.3 ×
−5, pSA = 0.004), while the other differences did not survive the spatial autocorrelation test
10
(green-red: t(152) = 0.47; p = 0.635, pSA = 0.590; yellow-red: t(156) = −3.02, p = 0.003, pSA =
0.121; green-blue: t(173) = −0.68, p = 0.496, pSA = 0.324; red-blue: t(135) = −1.30, p = 0.195,
pSA = 0.237; Supplementary Figure S5A).
The modulatory effects of noradrenaline have been argued to depend directly on ongoing
glutamatergic activity in target regions (Mather et al., 2016; Shine, 2021). De plus, it has
been shown that the main source of the BOLD activity is the neurovascular response caused
by pyramidal neurons containing cyclo-oxygenase-2 (Lecrux & Hamel, 2016). Surtout,
this evoked response following noradrenergic activation is dependent on the ongoing activity
of the pyramidal neurons (Bekar et al., 2012). Ainsi, the role of noradrenaline on brain dynam-
ics and BOLD response depends critically on ongoing glutamatergic activity, which putatively
represents pooled neural spiking activity (Logothetis, 2003). Given the differential task-related
BOLD activity of the different submodules (c'est à dire., Figure 2F, Supplementary Figure S4, et
Figure 4A), and the observed regional variability and specificity of integration across the net-
travail, we hypothesized that network-level integration would be explained by the combined
effect of ongoing BOLD activity and the distribution of the adrenergic receptor expression.
Enfin, we predicted that the role of the α2a receptor atlas in shaping brain activity and to-
pology should be dependent of the subjects’ pupil diameter, such that higher βpupil should
rely on a stronger relationship between network topology and α2a receptor expression.
To evaluate between these different hypotheses, we created three linear mixed models in
order to better disentangle the different plausible interactions between the variables (voir
Methods), while still controlling for between-subject variability as a grouping variable.
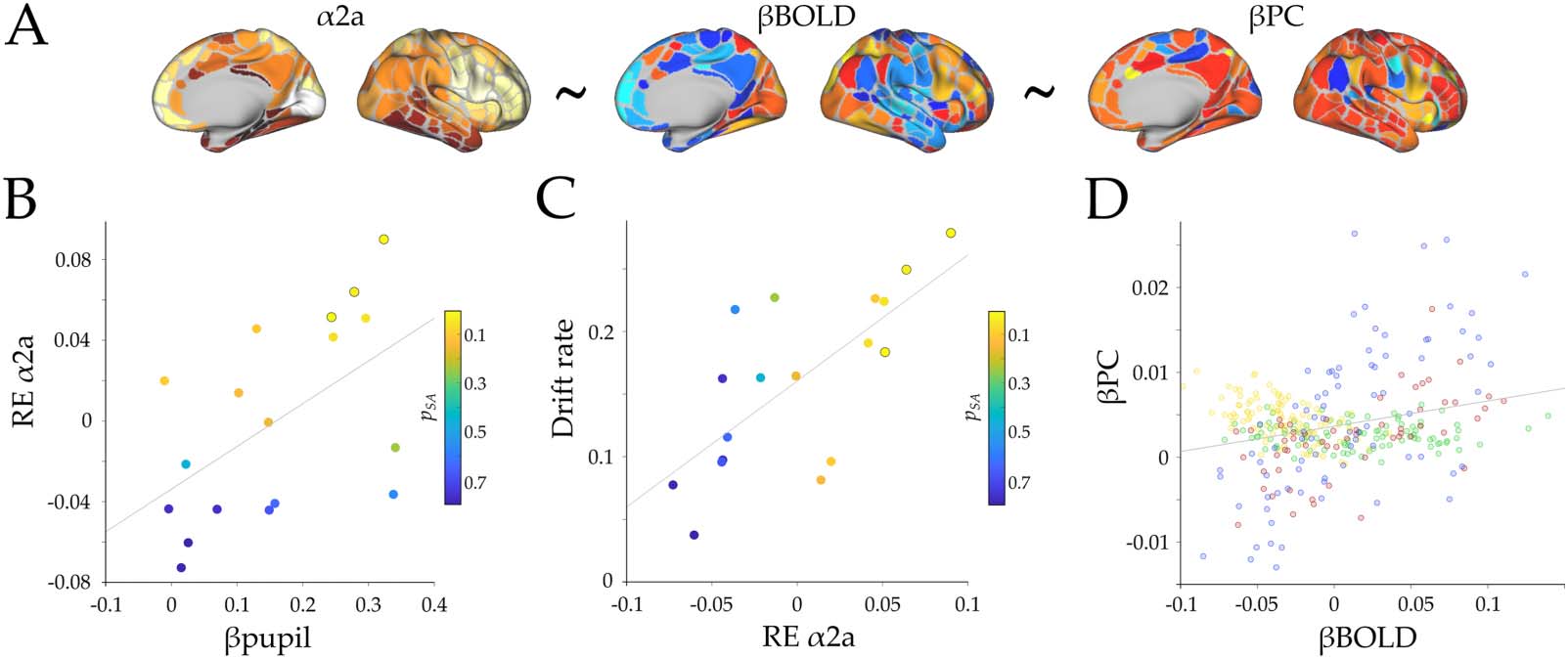
Chiffre 4. Receptor density analysis. (UN) Spatial maps of α2a density (gauche), BOLD parametric effect (middle), and participation coefficient
parametric effect (droite). The ~ symbol represents the linear model tested in the analysis. (B) Scatterplot depicting the relationship between
β
Pupil and the random effect of α2a (RE α2a; r = 0.54, p = 0.02). (C) Scatterplot depicting the relationship between the random effect of α2a and
drift rate (r = 0.70, p = 0.001); the colors of the dots represent the pSA value from the linear effect of α2a on βBOLD within each subject, et
the marked circles correspond to subjects with pSA < 0.05. (D) Pearson correlation of the group average BOLD parametric effect (β
BOLD)
and participation coefficient (β
−7). Colors represent each module assignment as in Figure 2.
PC; r = 0.26, p = 7 × 10
Network Neuroscience
897
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
Additionally, to control for spatial autocorrelation, we used 5,000 surrogate maps that main-
tained the spatial autocorrelation of the α2a while permuting the density values. In the first
model (Equation 8), we tested the hypothesis that the parametric BOLD effect (i.e., βBOLD,
Supplementary Figure S4) is shaped by the distribution of α2a receptors. We found significant
evidence for a positive fixed effect of α2a on βBOLD activity; however, this effect did not
survive correction for spatial autocorrelation (βα2a = 0.037 ± 0.016; t(5992) = 2.29; p =
0.022; pSA = 0.106; Supplementary Table S1). Furthermore, we correlated the random-effect
coefficients (from the original and the surrogate maps) to both βPC and βpupil and observed a
significant positive correlation between the participation coefficient and both pupils (Pearson’s
r = 0.54, p = 0.02, pSA = 0.036; Figure 4B) and mean drift rate (Pearson’s r = 0.70, p = 0.001,
pSA = 0.001; Figure 4C). This result shows the manner in which pupil diameter linearly shapes
βBOLD cortical map through the engagement of the α2a receptor expression map. Importantly,
although the fixed effect of α2a on βBOLD didn’t survive the spatial autocorrelation correction,
the linear correlation of this effect with both βpupil and drift rate (between subjects) did survive
the correction.
To further analyze the between-subject differences in the role of α2a receptor atlas in shap-
ing the βBOLD map, we ran a separate linear model within each subject with α2a as a regres-
sor and βBOLD of each region as the dependent variable (while also correcting for spatial
autocorrelation using 5,000 surrogate maps). As can be seen in Figure 4B–C, we observed
a dependency between the pSA value, βpupil, and drift rate, in which the respective within-
subject effects that survived the spatial autocorrelation correction are shown ( pSA < 0.05;
marked circles in Figure 4B–C). Despite these results, there was no significant effect of α2a
on βPC (Equation 9; βα2a = 0.001 ± 0.003; t(5992) = −0.51; p = 0.6), and no significant
Pearson’s correlations were found between the random effects and both βpupil or drift rate
(r = −0.24, p = 0.33; and r = −0.23, p = 0.341, respectively). However, we did find a signif-
−5;
icant effect of βBOLD on βPC (Equation 10; β = 0.0259 ± 0.006; t(5992) = 3.96; p = 7.55 × 10
Supplementary Table S1 and Figure 4D). Together these results propose a closer link between
pupil diameter, ascending neuromodulation, and the cortical neuromodulation dependent
on α2a receptor density.
Finally, we observed a differential relationship between βPC and βBOLD depending on the
large-scale module to which the regions were assigned. We expanded the former result by mea-
suring, within each subject, the Pearson correlation between the βBOLD and βPC separately in
each large-scale module (M1 being the modules assigned as red and blue, and M2 assigned as
yellow and green; Figure 2). The results demonstrated a significant difference between modules,
meaning that M1 has a higher correlation with βPC, in comparison with M2 (t(17) = −12.99, p =
−10, Supplementary Figure S5C). These results provided evidence that the adrenergic
2.93 × 10
receptor distribution of α2a shapes the βBOLD activation map in proportion to the subject’s
pupil diameter. Additionally, βBOLD activation map modulates (i.e., was related to) mesoscale
integration, and mesoscale integration is related to pupil diameter. Based on these results, we
hypothesize that the adrenergic system shapes the BOLD activity, which in turns shapes the
topology of the network towards integration. However, future work is required in order to test
this hypothesis more directly, for instance by combining optogenetic approaches with neuronal
recordings in awake animals.
DISCUSSION
Here, we leveraged a unique dataset to simultaneously track pupil diameter and network
topology during an attentional demanding task with increasing attentional load. Our results
Network Neuroscience
898
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
provide integrative evidence that links the ascending arousal system to the mesoscale topolog-
ical signature of the functional brain network during the processing of an attentionally de-
manding cognitive task. Pupil diameter was tracked with attentional load (Figure 1A) and
was related to the speed of information accumulation as estimated by a drift diffusion model
(Figure 1B–C). Additionally, we observed concurrent pupil dilations and adaptive mesoscale
parametric topological changes as a function of task demands (Figures 2 and 3). Finally, we
found evidence that topological reconfiguration was dependent on the regional activity and
the genetic expression of the adrenergic receptors in the brain (Figure 4). Together, these
results provide evidence for the manner in which the ascending arousal noradrenergic system
reconfigures brain network topology so as to promote attentional performance according to
task demands.
The relationship between performance and pupil diameter is consistent with the predictions
of adaptive gain theory (Aston-Jones & Cohen, 2005). Within this framework, the locus coer-
uleus is proposed to adaptively alter its activity according to the demands imposed on the
system. More specifically, the theory proposes that performance follows an inverted U-shaped
relationship with arousal, such that maximal operational flexibility in the noradrenergic system
is associated with optimal task performance (Arnsten et al., 2012; Robbins & Arnsten, 2009).
We observed that load-related increases in pupil diameter, presumably due to increased
activity in the ascending arousal system (Aston-Jones & Cohen, 2005; S. Joshi et al., 2016;
Liu et al., 2017), relates closely with the activity and topology of the broader brain network
(Figure 2), in a manner that is reflective of effective task performance (Figure 3). Similar effects
have been described in animal models after a chemogenetic activation of the locus coeruleus,
which strongly alters the large-scale network structure towards large-scale integration, specif-
ically in regions with heightened adrenergic receptor expression (Zerbi et al., 2019). How
these changes, which are likely related to the modulation of the neural gain that mediates
effective connections between distributed regions of the brain (Shine et al., 2021; Shine et al.,
2018), are traded off against requirements for specificity and flexibility remains an important
open question for future research.
The addition of attentional load was found to alter the integration of mesoscale submodules,
but not the higher level modular organization. This topological result is somewhat more
targeted than those described in previous work (Shine, Breakspear, et al., 2019; Shine et al.,
2016). While these differences may be related to disparities in the way that the data were
analyzed, the results of our study do demonstrate that alterations in the cerebral network
topology at a relatively local (i.e., submodular) level are crucial for effective task performance
(Akiki & Abdallah, 2019). Additionally, our results replicate and expand upon a previous study
(Mohr et al., 2016), in which the authors found that short-term practice on an attentional task
was related to increased coupling between attentional networks and segregation among task-
negative (DMN) and frontoparietal network (FPN). Our study replicates the graph theoretical
results of that study, while also directly relating the findings to the architecture of the ascending
neuromodulatory system. One potential explanation for these results comes from animal
studies, in which rapid changes in pupil diameter have been compared with changes in neural
population activity at the microscale (S. Joshi et al., 2016; McGinley et al., 2015; Reimer et al.,
2014). These studies suggest that the ascending arousal system may be able to alter the topology
of the network in a hierarchical manner that is commensurate with the spatiotemporal scale of
the arousal systems’ capacity (Shine et al., 2016). Future work that integrates results across
spatiotemporal scales is required to appropriately adjudicate the implications of this hypothesis.
Importantly, our approach is not without limitations. For one, the participation measures
used in our linear mixed model were estimated at the mesoscale level, and hence derived from
Network Neuroscience
899
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
different modular partitions. Furthermore, the specificity of the pupillary response as a corre-
late of locus coeruleus (LC) activity is currently under active debate. For instance, in addition
to the strong empirical links between the noradrenergic system and pupil dilation, there is also
evidence that the pupil is dilated in concert with activity in the basal forebrain cholinergic
system (Reimer et al., 2016), however it bears mention that both peripheral (Kaymak et al.,
2018) and central cholinergic tone (Yüzgeç et al., 2018) are associated with pupillary constric-
tion. There are more plausible physiological routes for the serotonergic system to dilate the
pupil (via the excitation of the intermediolateral cell column), and in keeping with this, there
is evidence that the serotonergic system is linked with pupil dilation (Cazettes et al., 2020).
Nevertheless, it is important to take into account that the neuromodulatory arousal system is
replete with complex interconnections (Avery & Krichmar, 2017; Briand et al., 2007; A. Joshi
et al., 2017; Smiley et al., 1999). In addition, based on the current lack of a specific mecha-
nism involving pupillary changes through the cholinergic system, it is highly probable that
those correlations are due to indirect modulation of pupillary responses (e.g., via indirect neu-
romodulation mediated by the LC system). On the other hand, we acknowledge the limitations
of the atlas receptor analysis and the linear model used in our study. More specific neurobi-
ological properties of the receptor distributions are needed to make better inferences, and
hence provide more accurate answers of their role in brain dynamics. For instance, it would
be ideal to compare receptor distributions that incorporated layer-specific expression, as there
are well-known cellular and circuit differences across layers in the cerebral cortex (Douglas &
Martin, 2004; Palomero-Gallagher & Zilles, 2019). Importantly, taking into consideration the
strong correlation between different genetic expression maps (Fornito et al., 2019), it is possi-
ble that the current correlation between ADRA2A expression and brain activity is a false pos-
itive caused by another neuroanatomical gradient strongly correlated to the ADRA2A.
Therefore, future work studying the interaction between genetic expression of the neuromo-
dulatory receptors, pupil diameter, and brain activity is needed. In spite of this limitation, we
believe in the importance of integrating pupil diameter and receptor distribution in the analysis
as the relationships between noradrenergic tone, brain activity, and network topology will
help us to disentangle the mechanistic steps connecting the locus coeruleus system to both
pupil diameter and brain dynamics.
In summary, we provide evidence linking mesoscale topological network integration, hier-
archical organization, and BOLD dynamics in the human brain that increases in attentional
load, thus providing further mechanistic clarity over the processes that underpin the adaptive
gain model of noradrenergic function in the central nervous system.
METHODS
Participants
Eighteen right-handed individuals (age 19–26 years; five male) were included in this study.
Exclusion criteria included standard contraindications for MRI; neurological disorders; and
mental disorders or drug abuse. All participants gave written informed consent before the
experiment.
Parametric Motion Tracking Task
Each trial of the task involved the same basic pattern (Figure 1A): The task begins with a dis-
play presenting the objects (i.e., blue colored disks); after a 2.5-s delay, a subset of the disks
turn red for another 2.5 s; all of the disks then return to blue (2.5 s) before they start moving
randomly inside the tracking area. The participants’ job is to track the “target” dots on the
Network Neuroscience
900
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
screen while visually fixating at the cross located at the center of the screen. After a tracking
period of ~11 s, one of the disks is highlighted in green (a “probe”) and the subject is then
asked to respond, as quickly as possible, as to whether the green probe object was one of
the original target objects. The number of objects that subjects were required to attend to
across the tracking period varied across trials. There were five trial types: passive viewing
(PV), in which no target is assigned; and four load conditions, in which two to five targets were
assigned for tracking. We operationalized attentional load as the linear effect of increasing task
difficulty (i.e., the number of targets to be tracked).
The experiment was conducted using a blocked design, in which each block included the
following: instruction (1 s); fixation (0.3 s, present throughout the rest of trial); object presen-
tation (all objects were blue; 2.5 s); target assignment (i.e., the targets changed color from blue
to red; 2.5 s); object representation (objects back to the original blue color; 2.5 s); object
movement/attentional tracking (moving blue dots; 11 s); object movement cessation (0.5 s);
and a final probe (color change to green and response; 2.5 s). The total duration of each trial
was 22.8 s. Each condition was repeated four times in one fMRI run, which also included four
separate fixation periods of 11 s each between five consecutive trials. All participants completed
four separate runs of the experiment, each of which comprised 267 volumes. The order of the
conditions was pseudorandom, such that the different conditions were grouped in sub-runs of
triplets: PV, pseudorandom blocks of Loads 2 through 5, and a fixation trial. All objects were
identical during the tracking interval and standard object colors were isoluminant (to minimize
incidental pupillary responses during the task).
Behavior and EZ-Diffusion Model
The EZ-diffusion model was used to interpret the performance measures from the task (Ratcliff
& Rouder, 1998; Wagenmakers et al., 2007). This model considers the mean RT of correct
trials, the standard deviation of the reaction time (SD-RT) across correct trials, and mean ac-
curacy across the task, and computes from these a value for drift rate (v, Equation 1), boundary
separation (a, Equation 2) , and non-decision time (Equation 3)—the three main parameters for
the drift diffusion model (Ratcliff et al., 2016; Ratcliff & Rouder, 1998).
(cid:3)
(cid:1)
v ¼ sign P − 1
2
: 0:1 :
4
s
(cid:5)
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
(cid:8)
(cid:6)
log
(cid:7)
(cid:5)
: P2 : log
(cid:5)
− P : log
(cid:6)
(cid:6)
þ P − 1
2
P
1 − P
P
1 − P
P
1 − P
;
VRT
a ¼ 0:01 :
log
(cid:6)
(cid:5)
P
1 − P
v
;
Ter ¼ MRT −
a
2 (cid:2) v
(cid:2) 1 − e−100
ð
1 þ e−100
ð
:
:
:
:
v
v
a
a
Þ
Þ
;
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
(1)
(2)
(3)
in which P is the average performance (range between 0 and 1); sign is an operator that will be
−1 if P < 0.5 or +1 if P > 0.5; VRT is the standard deviation of reaction time (in seconds); et
MRT is the mean reaction time (in seconds).
Pupillometry
Fluctuations in pupil diameter of the left eye were collected using an MR-compatible coil-
mounted infrared EyeTracking system (NNL EyeTracking camera, NordicNeuroLab, Bergen,
Norway), at a sampling rate of 60 Hz and recorded using the iView X Software (SensoMotoric
Instruments, SMI GmbH, Allemagne). Blinks, artifacts, and outliers were removed and linearly
interpolé (Wainstein et al., 2017). High-frequency noise was smoothed using a second-order
Neurosciences en réseau
901
The ascending arousal system shapes mesoscale network integration
2.5-Hz low-pass Butterworth filter. To obtain the pupil diameter average profile for each level of
attentional load (Figure 1B), data from each participant were normalized across each task block
(corresponding to the five consecutive trials between fixations). This allowed us to correct for
low-frequency baseline changes without eliminating the load effect and baseline differences
due to load manipulations (Campos-Arteaga et al., 2020; Rojas-Líbano et al., 2019).
Following this, a linear regression was performed in each time point using the task load as re-
gressor and resulting in a “load effect” time series for each subject.
MRI Data
Imaging data were collected on a Philips Achieva 3 Tesla MR-scanner, equipped with an
eight-channel Philips SENSE head coil (Philips Medical Systems, Best, Netherlands) à la
Intervention Centre, Oslo University Hospital, Norway. Functional data were collected using
a BOLD-sensitive T2*-weighted echo-planar imaging sequence (36 slices, no gap; repetition
temps (TR), 2,2 s; echo time (TE), 30 ms; flip angle, 80°; voxel size, 3 × 3 × 3; field of view
(FOV), 240 × 240 mm; interleaved acquisition). Anatomical T1-weighted images consisting
de 180 sagittal-oriented slices were obtained using a turbo field echo pulse sequence (TR,
6.7 ms; TE, 3.1 ms; flip angle 8°; voxel size 1 × 1.2 × 1.2 mm; FOV, 256 × 256 mm).
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
un
_
0
0
2
0
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
fMRI Data Preprocessing
After realignment (using FSL’s MCFLIRT), we used FEAT to unwarp the EPI images in the y-
direction with a 10% signal loss threshold and an effective echo spacing of 0.333. Following
noise-cleaning with FIX (custom training set for scanner, threshold 20, included regression of
estimated motion parameters), the unwarped EPI images were then smoothed at 6-mm FWHM,
and nonlinearly coregistered with the anatomical T1 to 2-mm isotropic MNI space. Temporal
artifacts were identified in each dataset by calculating framewise displacement (FD) from the
derivatives of the six rigid-body realignment parameters estimated during standard volume re-
alignment (Power et al., 2014), as well as the root mean square change in BOLD signal from
volume to volume (DVARS). Frames associated with FD > 0.25 mm or DVARS > 2.5% étaient
identified; cependant, as no participants were identified with greater than 10% of the resting
time points exceeding these values, no trials were excluded from further analysis. Il y avait
no differences in head motion parameters between the four sessions ( p > 0.500). Following
artifact detection, nuisance covariates associated with the six linear head movement parame-
ters (and their temporal derivatives), DVARS, physiological regressors (created using the
RETROICOR method), and anatomical masks from the cerebrospinal fluid and deep cerebral
white matter were regressed from the data using the CompCor strategy (Behzadi et al., 2007).
Enfin, in keeping with previous time-resolved connectivity experiments (Gu et al., 2015), un
temporal band pass filter (0.0071 < f < 0.125 Hz) was applied to the data.
Brain Parcellation
Following preprocessing, the mean time series was extracted from 375 predefined regions of
interest (ROIs). To ensure whole-brain coverage, we extracted the following: (a) 333 cortical
parcels (161 and 162 regions from the left and right hemispheres, respectively) using the
Gordon atlas (Gordon et al., 2016); (b) 14 subcortical regions from the Harvard-Oxford sub-
cortical atlas (bilateral thalamus, caudate, putamen, ventral striatum, globus pallidus, amyg-
dala, and hippocampus; https://fsl.fmrib.ox.ac.uk/); and (c) 28 cerebellar regions from the SUIT
atlas (Diedrichsen et al., 2009) for each participant in the study.
Network Neuroscience
902
The ascending arousal system shapes mesoscale network integration
Time-Resolved Functional Connectivity and Network Analysis
To estimate functional connectivity between the 375 ROIs, we used the jackknife correlation
( JC) approach (Thompson et al., 2018). Briefly, this approach estimates the static correlations
between each pair of regions, and then recalculates the correlation between each pair after
systematically removing each temporal “slice” of data (i.e., each TR). By subtracting the jack-
knifed correlation matrix from the original “static” matrix, the difference in connectivity at
each slice from the static connectivity value can be used as an estimate of time-resolved func-
tional connectivity between each pair of regions at each TR in a way that does not require
windowing.
Community Structure
The Louvain modularity algorithm from the Brain Connectivity Toolbox (Rubinov & Sporns,
2010) was used in combination with the JC to estimate both time-averaged and time-resolved
community structure. The Louvain algorithm iteratively maximizes the modularity statistic, Q,
for different community assignments until the maximum possible score of Q has been obtained
(Equation 4).
QT ¼ 1
vþ
X
(cid:9)
w
ij
(cid:10)
δMiMj
−
þ
ij
− e
þ
ij
1
vþ þ v−
X
(cid:9)
w
ij
(cid:10)
δMiMj
:
−
ij
−
− e
ij
(4)
Equation 4: Louvain modularity algorithm, where v is the total weight of the network (sum of
all negative and positive connections); wij is the weighted and signed connection between
regions i and j; eij is the strength of a connection divided by the total weight of the network;
and δ
Mi Mj is set to 1 when regions are in the same community and 0 otherwise. The + and −
superscripts denote all positive and negative connections, respectively.
For each subject, we calculated the mean adjacency matrix from 1 TR before tracking until
the end of the tracking period. Afterwards, a consensus partition was estimated across subjects.
Finally, to identify multilevel structure in our data, we repeated the modularity analysis for
each of the modules identified in the first step (Meunier et al., 2010; Meunier et al., 2009).
With this final module assignment, we were afforded an estimate of the time-resolved, multi-
level modularity (QT) within each temporal window for each participant in the study.
Regional Integration
Based on the group consensus community assignments, we estimated between-module con-
nectivity using the participation coefficient, BT, which quantifies the extent to which a region
connects across all modules (i.e., between-module strength; Equation 5). In our experiment,
we used two separate community assignments, one for each of the modularity levels. In this
manner we measure (a) how the first hierarchical-level (i.e., large-scale) topology changed
during tracking across the complete brain; and (b) how the topology of the submodules chan-
ged across the task. These values were calculated in each time point using the time-resolved
adjacency matrix across each load condition.
BiT ¼ 1 −
X
nM
s¼1
(cid:1) (cid:3)
2
κisT
κiT
:
(5)
Equation 5: Participation coefficient BiT, where κ
isT is the strength of the positive connections
of region i to regions in module s at time T, and κ
iT is the sum of strengths of all positive con-
nections of region i at time T. The participation coefficient of a region is therefore close to 1 if
its connections are uniformly distributed among all the modules and 0 if all of its links are
within its own module.
Network Neuroscience
903
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
Neurotransmitter Receptor Mapping
To investigate the potential correlates of mesoscale integration, we interrogated the neuro-
transmitter receptor signature of each region of the brain. We used the Allen Brain Atlas
microarray atlas dataset (https://human.brain-map.org/; Hawrylycz et al., 2012) to identify
the regional signature of genetic expression of the α2a subtype of the adrenergic receptor
(ADRA2A). This receptor has been a priori related to cognitive function and attention
(Arnsten & Haven, 2013), and is one of the most abundant adrenergic subtypes expressed
in the cerebral cortex (Perez, 2020). This atlas contains postmortem samples of six donors
that underwent microarray transcriptional characterization. The spatial map of α2a mRNA
expression was obtained in volumetric 2-mm isotropic MNI space, following improved
nonlinear registration and whole-brain prediction using variogram modeling (Gryglewski
et al., 2018). We used these data instead of the native sample-wise values in the AHBA
database to prevent bias that could occur because of spatial inhomogeneity of the sampled
locations. We projected the volumetric α2a expression data onto the Gordon atlas with
linear interpolation and calculated the mean value within each parcel using custom MATLAB
codes.
STATISTICAL ANALYSIS
The Relationship Between Sympathetic Tone and Attentional Processing
We analyzed the between-subject effect of load on the behavioral, pupillometric, and fMRI-
related variables by performing a two-level linear model analysis. In the first level, we used
attentional load as a regressor (2 to 5) and—in independent models—the mean accuracy, re-
action time, standard deviation of reaction time, drift rate, boundary criteria, and non-decision
time as dependent variables (i.e., four values per subject). From this, we ran a two-tailed t test
on the statistical effects (i.e., the β value from the regression, one for each subject; N = 18).
Similarly, to calculate the load effect on pupil diameter, we calculated the average pupil di-
ameter on each load condition within each subject. Then, we performed a first-level analysis
in which we ran a linear regression in each time frame (1600 frames in total, corresponding to
26.6 seconds). This procedure resulted in one β timeseries (i.e., the statistical load effect on
pupil diameter) for each subject across the trial (Supplementary Figure S1A). After this, we
performed a right tailed t test in each frame across subjects (n = 18 in each frame) to find
the periods of time where the β values where higher than 0. Finally, we corrected by false
discovery rate (FDR; Benjamini & Yekutieli, 2001) for multiple testing, which resulted in a
period of time in which the load effect was higher than 0 (light gray area in Figure 1A). The
mean β values during this section was calculated in each subject and defined as “βpupil.”
Finally, following the same pipeline, we calculated the effect of attentional load on the
brain-related signals (i.e., BOLD, participation coefficient [PC], and modularity [Q]). The effect
of load on BOLD was calculated running a separate linear model in each subject and region
within each TR (18 subjects; 375 regions; 10 TRs; 4 load conditions), resulting in a matrix of
β values of 18 × 375 × 10.
To evaluate the statistical effect of pupil diameter on accuracy, we performed a logistic
linear mixed-effects model. We used the mean pupil diameter of the significant time period
(Figure 1A) of the high load trials (Loads 4 and 5) and the accuracy (i.e., correct or incorrect) as
the predictor variable of each trial, grouping by subject as the random effect. The statistical
model is described in the following equation:
Accuracy e Pupil þ 1 þ Pupil þ 1jSubject
ð
Þ:
(6)
904
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
Network Integration Increases as a Function of Attentional Load
To evaluate whether the modularity of the network we observed was higher than chance, we
generated 100 random networks in each hierarchical level (300 random networks in total),
with a preserved degree distribution (using the MATLAB randmio_und_signed function
from the Brain Connectivity Toolbox; Rubinov & Sporns, 2010). We calculated the modularity
value of each random network and used the resultant values to populate a null distribution
(Figure 2D).
We analyzed the statistical effect of pupil diameter on the participation coefficient both
within and between subjects by performing a linear mixed model using the time-varying PC
of the red submodule (Figure 3A) of each load as a dependent variable (N = 72), and the
respective pupil diameter as a regressor, with grouping by subject. The statistical model is
described in the following equation:
ð
PC e Pupil þ 1 þ Pupil þ 1jSubject
Þ:
(7)
Network Mesoscale Integration and Adrenergic Receptor Density
Expression of brain genetic atlas varies smoothly across the surface and thus is associated with
nontrivial spatial autocorrelation that in turn violates the assumption of independence be-
tween samples (Burt et al., 2020; Markello & Misic, 2021; Vos de Wael et al., 2020). To ac-
count for the spatial autocorrelation in these brain maps, we used spatial autocorrelation null
maps as implemented in Brain Surrogate Maps with Autocorrelated Spatial Heterogeneity
(BrainSMASH) Python toolbox (Burt et al., 2020). A geodesic distance matrix of the atlas par-
cels using the surface of the Gordon atlas was obtained to build the surrogates using
BrainSMASH functions. We generated 5,000 null maps that were used to generate null distri-
bution of the different statistics corrected by spatial autocorrelation.
We measure the statistical difference in the receptor density between submodules by a two-
tailed t test between each pair of modules. The same procedure was performed using the sur-
rogate maps to generate a null distribution of t statistics. To evaluate the effect of the density of
each adrenergic receptor on the neural activity in the attentional task, we built a linear mixed
model aimed at predicting regional differences in BOLD activity and participation coefficient.
We created a model using the receptor density atlas of α2a receptor to predict parametric
BOLD activity (i.e., linear increase of BOLD activity with task load) during tracking
(Equation 8). To evaluate the relationship between BOLD activity, adrenergic receptor expres-
sion, and changes in participation coefficient as a function of attentional load, we tested two
models: one using the adrenergic receptor density as independent factor (Equation 9), and an-
other using the parametric BOLD effect as an independent factor (Equation 10). Additionally,
we assessed the across-subject variability using the subjects’ ID as a grouping variable in order
to evaluate the random effects on the independent factor. We corrected the spatial autocorre-
lation by running the same model using 5,000 surrogate maps. Then we used the fixed-effect
null distribution to calculate the pSA (i.e., the probability of finding the fixed effect within the
95th percentile of the null distribution). The deterministic part of the model is expressed in the
following equations (Wilkinson & Rogers, 1973):
βBOLD e α2a þ 1 þ α2a þ 1jSubject
ð
Þ;
Þ;
ð
βPC e α2a þ 1 þ α2a þ 1jSubject
ð
βPC e BOLD þ 1 þ BOLD þ 1jSubject
Þ;
(8)
(9)
(10)
905
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
where PC is the parametric effect of mesoscale participation coefficient (i.e., βPC), BOLD is the
parametric effect of load on BOLD activity during tracking for each region, and α2a are the
regional densities of the respective adrenergic receptor atlas. We then correlated the random-
effects parameters to pupil diameter responses and behavior and then compared these with the
Pearson’s correlation of the null distribution using the random effect of the surrogate maps.
Finally, we performed a linear model within each subject with α2a as a regressor and
βBOLD as dependent variable. Again, the statistical effect (i.e., β value) was compared against
the null distribution when performing the regression using the surrogate maps (Figure 4B–C).
ACKNOWLEDGMENTS
We thank P. Billeke for his thoughtful comments on our manuscript.
DATA AND CODE AVAILABILITY
The anonymized preprocessed fMRI and pupillometry data can be found at https://figshare.com
/articles/dataset/MOT_data_mat/13244504 (Wainstein et al., 2020). The ADRA2A expression
atlas can be downloaded from https://www.meduniwien.ac.at/neuroimaging/mRNA.html.
All analysis of the fMRI and pupil diameter data were performed on MATLAB 2020a. The sur-
rogate maps of the ADRA2A atlas were generated on Python. Documented code for repro-
ducing the analyses is provided in https://github.com/gabwainstein/MOT (Wainstein, 2021).
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00205.
AUTHOR CONTRIBUTIONS
Gabriel Wainstein: Conceptualization; Formal analysis; Investigation; Methodology;
Visualization; Writing – original draft; Writing – review & editing. Daniel Rojas-Líbano:
Visualization; Writing – original draft; Writing – review & editing. Vicente Medel: Formal
analysis; Writing – review & editing. Dag Alnæs: Conceptualization; Data curation; Funding
acquisition; Investigation; Methodology; Writing – review & editing. Knut K. Kolskår:
Conceptualization; Investigation; Validation; Writing – review & editing. Tor Endestad:
Conceptualization; Investigation; Methodology; Writing – review & editing. Bruno Laeng:
Conceptualization; Investigation; Writing – review & editing. Tomas Ossandon: Supervision;
Writing – review & editing. Nicolás Crossley: Validation; Writing – review & editing. Elie
Matar: Conceptualization; Writing – review & editing. James M. Shine: Conceptualization;
Data curation; Formal analysis; Investigation; Methodology; Project administration;
Resources; Supervision; Validation; Writing – original draft; Writing – review & editing.
FUNDING INFORMATION
James M. Shine, the University of Sydney Robinson Fellowship. James M. Shine, National
Health and Medical Research Council (https://dx.doi.org/10.13039/501100000925), Award
ID: GNT1156536. Gabriel Wainstein, Becas Chile.
REFERENCES
Akiki, T. J., & Abdallah, C. G. (2019). Determining the hierarchical
architecture of the human brain using subject-level clustering of
functional networks. Scientific Reports, 9(1), 1–15. https://doi.org
/10.1038/s41598-019-55738-y, PubMed: 31848397
Alnæs, D., Kaufmann, T., Richard, G., Duff, E. P., Sneve, M. H.,
Endestad, T., Nordvik, J. E., Andreassen, O. A., Smith, S. M., &
Westlye, L. T. (2015). Attentional load modulates large-scale
functional brain connectivity beyond the core attention net-
works. NeuroImage, 109, 260–272. https://doi.org/10.1016/j
.neuroimage.2015.01.026, PubMed: 25595500
Alnæs, D., Sneve, M. H., Espeseth, T., Endestad, T., van de Pavert,
S. H. P., & Laeng, B. (2014). Pupil size signals mental effort
Network Neuroscience
906
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
deployed during multiple object tracking and predicts brain
activity in the dorsal attention network and the locus coeruleus.
Journal of Vision, 14(2014), 1–20. https://doi.org/10.1167/14.4.1,
PubMed: 24692319
Arnsten, A. F. T., & Haven, N. (2013). The neurobiology of thought:
The groundbreaking discoveries of Patricia Goldman-Rakic
1937–2003. Cerebral Cortex, 23(10), 2269–2281. https://doi
.org/10.1093/cercor/bht195, PubMed: 23926115
Arnsten, A. F. T., Wang, M. J., & Paspalas, C. D. (2012).
Neuromodulation of thought: Flexibilities and vulnerabilities in
prefrontal cortical network synapses. Neuron, 76(1), 223–239.
https://doi.org/10.1016/j.neuron.2012.08.038, PubMed:
23040817
Aston-Jones, G., & Cohen, J. D. (2005). An integrative theory of
locus coeruleus-norepinephrine function: Adaptive gain and
optimal performance. Annual Review of Neuroscience, 28(1),
403–450. https://doi.org/10.1146/annurev.neuro.28.061604
.135709, PubMed: 16022602
Aston-Jones, G., & Waterhouse, B. (2016). Locus coeruleus: From
global projection system to adaptive regulation of behavior.
Brain Research, 1645, 75–78. https://doi.org/10.1016/j.brainres
.2016.03.001, PubMed: 26969408
Avery, M. C., & Krichmar, J. L. (2017). Neuromodulatory systems
and their interactions: A review of models, theories, and experi-
ments. Frontiers in Neural Circuits, 11(December), 1–18. https://
doi.org/10.3389/fncir.2017.00108, PubMed: 29311844
Bassett, D. S., Greenfield, D. L., Meyer-Lindenberg, A.,
Weinberger, D. R., Moore, S. W., & Bullmore, E. T. (2010).
Efficient physical embedding of topologically complex informa-
tion processing networks in brains and computer circuits. PLoS
Computational Biology, 6(4), e1000748. https://doi.org/10.1371
/journal.pcbi.1000748, PubMed: 20421990
Behzadi, Y., Restom, K., Liau, J., & Liu, T. T. (2007). A component
based noise correction method (CompCor) for BOLD and perfu-
sion based fMRI. NeuroImage, 37(1), 90–101. https://doi.org/10
.1016/j.neuroimage.2007.04.042, PubMed: 17560126
Bekar, L. K., Wei, H. S., & Nedergaard, M. (2012). The locus
coeruleus-norepinephrine network optimizes coupling of cere-
bral blood volume with oxygen demand. Journal of Cerebral
Blood Flow and Metabolism, 32(12), 2135–2145. https://doi
.org/10.1038/jcbfm.2012.115, PubMed: 22872230
Benjamini, Y., & Yekutieli, D. (2001). The control of the false dis-
covery rate in multiple testing under dependency. Annals of
Statistics, 29(4), 1165–1188. https://doi.org/10.1214/aos
/1013699998
Bouret, S., & Sara, S. J. (2005). Network reset: A simplified over-
arching theory of locus coeruleus noradrenaline function.
Trends in Neurosciences, 28(11), 574–582. https://doi.org/10
.1016/j.tins.2005.09.002, PubMed: 16165227
Briand, L. A., Gritton, H., Howe, W. M., Young, D. A., & Sarter, M.
(2007). Modulators in concert for cognition: Modulator interac-
tions in the prefrontal cortex. Progress in Neurobiology, 83(2),
69–91. https://doi.org/10.1016/j.pneurobio.2007.06.007,
PubMed: 17681661
Burt, J. B., Helmer, M., Shinn, M., Anticevic, A., & Murray, J. D.
(2020). Generative modeling of brain maps with spatial autocor-
relation. NeuroImage, 220(February), 117038. https://doi.org/10
.1016/j.neuroimage.2020.117038, PubMed: 32585343
Campos-Arteaga, G., Forcato, C., Wainstein, G., Lagos, R.,
Palacios-García, I., Artigas, C., Morales, R., Pedreira, M. E. E.,
& Rodríguez, E. (2020). Differential neurophysiological corre-
lates of retrieval of consolidated and reconsolidated memories
in humans: An ERP and pupillometry study. Neurobiology of
Learning and Memory, 107279. https://doi.org/10.1016/j.nlm
.2020.107279, PubMed: 32710932
Cazettes, F., Reato, D., Morais, J. P., Renart, A., & Mainen, Z. F.
(2020). Phasic activation of dorsal raphe serotonergic neurons in-
creases pupil size. Current Biology, 192–197. https://doi.org/10
.1016/j.cub.2020.09.090, PubMed: 33186549
de Gee, J. W., Colizoli, O., Kloosterman, N. A., Knapen, T.,
Nieuwenhuis, S., & Donner, T. H. (2017). Dynamic modulation
of decision biases by brainstem arousal systems. eLife, 6, e23232.
https://doi.org/10.7554/eLife.23232, PubMed: 28383284
de Gee, J. W., Knapen, T., & Donner, T. H. (2014). Decision-related
pupil dilation reflects upcoming choice and individual bias.
Proceedings of the National Academy of Sciences, 111(5),
E618–E625. https://doi.org/10.1073/pnas.1317557111,
PubMed: 24449874
Diedrichsen, J., Balsters, J. H., Flavell, J., Cussans, E., & Ramnani,
N. (2009). A probabilistic MR atlas of the human cerebellum.
N e u r o I m a g e , 4 6 ( 1 ) , 3 9 – 4 6 . h t t p s : / / d o i . o r g / 1 0 . 1 0 1 6 / j
.neuroimage.2009.01.045, PubMed: 19457380
Donner, T., Kettermann, A., Diesch, E., Ostendorf, F., Villringer, A.,
& Brandt, S. A. (2000). Involvement of the human frontal eye
field and multiple parietal areas in covert visual selection during
conjunction search. European Journal of Neuroscience, 12(9),
3407–3414. https://doi.org/10.1046/j.1460-9568.2000.00223.x,
PubMed: 10998123
Douglas, R. J., & Martin, K. A. C. (2004). Neuronal circuits of the
neocortex. Annual Review of Neuroscience, 27(1), 419–451.
https://doi.org/10.1146/annurev.neuro.27.070203.144152,
PubMed: 15217339
Fornito, A., Arnatkevic(cid:1)iu(cid:3)te_, A., & Fulcher, B. D. (2019). Bridging the
gap between connectome and transcriptome. Trends in
Cognitive Sciences, 23(1), 34–50. https://doi.org/10.1016/j.tics
.2018.10.005, PubMed: 30455082
Gilzenrat, M. S., Nieuwenhuis, S., Jepma, M., & Cohen, J. D.
(2010). Pupil diameter tracks changes in control state predicted by
the adaptive gain theory of locus coeruleus function. Cognitive,
Affective and Behavioral Neuroscience, 10(2), 252–269. https://doi
.org/10.3758/CABN.10.2.252, PubMed: 20498349
Gordon, E. M., Laumann, T. O., Adeyemo, B., Huckins, J. F.,
Kelley, W. M., & Petersen, S. E. (2016). Generation and evalua-
tion of a cortical area parcellation from resting-state correlations.
Cerebral Cortex, 26(1), 288–303. https://doi.org/10.1093/cercor
/bhu239, PubMed: 25316338
Gryglewski, G., Seiger, R., James, G. M., Godbersen, G. M.,
Komorowski, A., Unterholzner, J., Michenthaler, P., Hahn, A.,
Wadsak, W., Mitterhauser, M., Kasper, S., & Lanzenberger, R.
(2018). Spatial analysis and high resolution mapping of the hu-
man whole-brain transcriptome for integrative analysis in neuro-
imaging. NeuroImage, 176(January), 259–267. https://doi.org/10
.1016/j.neuroimage.2018.04.068, PubMed: 29723639
Gu, S., Pasqualetti, F., Cieslak, M., Telesford, Q. K., Yu, A. B.,
Kahn, A. E., Medaglia, J. D., Vettel, J. M., Miller, M. B., Grafton,
S. T., & Bassett, D. S. (2015). Controllability of structural brain
Network Neuroscience
907
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
networks. Nature Communications, 6, 1–10. https://doi.org/10
.1038/ncomms9414, PubMed: 26423222
Hawrylycz, M. J., Lein, E. S., Guillozet-bongaarts, A. L., Shen, E. H.,
Ng, L., Miller, J. A., van de Lagemaat, L. N., Smith, K. A., Ebbert,
A., Riley, Z. L., Abajian, C., Beckmann, C. F., Bernard, A.,
Bertagnolli, D., Boe, A. F., Cartagena, P. M., Chakravarty,
M. M., Chapin, M., Chong, J., … Zielke, H. R. (2012). An ana-
tomically comprehensive atlas of the adult human brain tran-
scriptome. Nature, 489(7416), 391–399. https://doi.org/10.1038
/nature11405, PubMed: 22996553
Joshi, A., Youssofzadeh, V., Vemana, V., McGinnity, T. M., Prasad,
G., & Wong-Lin, K. F. (2017). An integrated modelling frame-
work for neural circuits with multiple neuromodulators. Journal
of the Royal Society Interface, 14(126). https://doi.org/10.1098
/rsif.2016.0902, PubMed: 28100828
Joshi, S., & Gold, J. I. (2020). Pupil size as a window on neural substrates
of cognition. Trends in Cognitive Sciences, 24(6), 466–480.
https://doi.org/10.1016/j.tics.2020.03.005, PubMed: 32331857
Joshi, S., Li, Y., Kalwani, R. M., & Gold, J. I. (2016). Relationships
between pupil diameter and neuronal activity in the locus coer-
uleus, colliculi, and cingulate cortex. Neuron, 89(1), 221–234.
https://doi.org/10.1016/j.neuron.2015.11.028, PubMed:
26711118
Kahneman, D., & Beatty, J. (1966). Pupil diameter and load on
memory. Science, 154(3756), 1583–1585. https://doi.org/10
.1126/science.154.3756.1583, PubMed: 5924930
Kaymak, H., Fricke, A., Mauritz, Y., Löwinger, A., Klabe, K., Breyer,
D., Lagenbucher, A., Seitz, B., & Schaeffel, F. (2018). Short-term
effects of low-concentration atropine eye drops on pupil size and
accommodation in young adult subjects. Graefe’s Archive for
Clinical and Experimental Ophthalmology, 2211–2217. https://
doi.org/10.1007/s00417-018-4112-8, PubMed: 30145612
Lecrux, C., & Hamel, E. (2016). Neuronal networks and mediators
of cortical neurovascular coupling responses in normal and
altered brain states. Philosophical Transactions of the Royal
Society B: Biological Sciences, 371(1705), 20150350. https://
doi.org/10.1098/rstb.2015.0350, PubMed: 27574304
Liu, Y., Rodenkirch, C., Moskowitz, N., Schriver, B., & Wang, Q.
(2017). Dynamic lateralization of pupil dilation evoked by
locus coeruleus activation results from sympathetic, not para-
sympathetic, contributions. Cell Reports, 20(13), 3099–3112.
https://doi.org/10.1016/j.celrep.2017.08.094, PubMed:
28954227
Logothetis, N. K. (2003). The underpinnings of the BOLD functional
magnetic resonance imaging signal. Journal of Neuroscience,
23(10), 3963–3971. https://doi.org/10.1523/JNEUROSCI.23-10
-03963.2003, PubMed: 12764080
Markello, R. D., & Misic, B. (2021). Comparing spatial null models
for brain maps. NeuroImage, 236(April), 118052. https://doi.org
/10.1016/j.neuroimage.2021.118052, PubMed: 33857618
Mather, M., Clewett, D., Sakaki, M., & Harley, C. W. (2016).
Norepinephrine ignites local hotspots of neuronal excitation:
How arousal amplifies selectivity in perception and memory.
Behavioral and Brain Sciences, 39. https://doi.org/10.1017
/S0140525X15000667, PubMed: 26126507
McGinley, M. J., Vinck, M., Reimer, J., Batista-Brito, R., Zagha, E.,
Cadwell, C. R., Tolias, A. S., Cardin, J. A., & McCormick, D. A.
(2015). Waking state: Rapid variations modulate neural and
behavioral responses. Neuron, 87(6), 1143–1161. https://doi
.org/10.1016/j.neuron.2015.09.012, PubMed: 26402600
Meunier, D., Lambiotte, R., & Bullmore, E. T. (2010). Modular and
hierarchically modular organization of brain networks. Frontiers
in Neuroscience, 4(200), 1–11. https://doi.org/10.3389/fnins
.2010.00200, PubMed: 21151783
Meunier, D., Lambiotte, R., Fornito, A., Ersche, K. D., & Bullmore,
E. T. (2009). Hierarchical modularity in human brain functional
networks. Frontiers in Neuroinformatics, 3(37), 1–12. https://doi
.org/10.3389/neuro.11.037.2009, PubMed: 19949480
Mohr, H., Wolfensteller, U., Betzel, R. F., Mišic(cid:4), B., Sporns, O.,
Richiardi, J., Ruge, H., Mohr, H., Wolfensteller, U., Betzel,
R. F., & Mis, B. (2016). Integration and segregation of large-
scale brain networks during short-term task automatization.
Nature Communications, 7(1), 1–12. https://doi.org/10.1038
/ncomms13217, PubMed: 27808095
Mulder, G. (2012). The concept and measurement of mental effort.
Energetics and Human Information Processing, 175–198. https://
doi.org/10.1007/978-94-009-4448-0_12
Munn, B., Müller, E. J., Wainstein, G., & Shine, J. M. (2021). The
ascending arousal system shapes low-dimensional brain dynamics
to mediate awareness of changes in intrinsic cognitive states.
bioRxiv. https://doi.org/10.1101/2021.03.30.437635
Murphy, P. R., Boonstra, E., & Nieuwenhuis, S. (2016). Global gain
modulation generates time-dependent urgency during perceptual
choice in humans. Nature Communications, 7(1), 1–15. https://
doi.org/10.1038/ncomms13526, PubMed: 27882927
Murphy, P. R., Vandekerckhove, J., & Nieuwenhuis, S. (2014).
Pupil-linked arousal determines variability in perceptual decision
making. PLoS Computational Biology, 10(9). https://doi.org/10
.1371/journal.pcbi.1003854, PubMed: 25232732
Palomero-Gallagher, N., & Zilles, K. (2019). Cortical layers: Cyto-,
myelo-, receptor- and synaptic architecture in human cortical
areas. NeuroImage, 197, 716–741. https://doi.org/10.1016/j
.neuroimage.2017.08.035, PubMed: 28811255
Parvizi, J., & Damasio, A. (2001). Consciousness and the brainstem.
Cognition, 79(1–2), 135–160. https://doi.org/10.1016/S0010
-0277(00)00127-X, PubMed: 11164026
Perez, D. M. (2020). A1-adrenergic receptors in neurotransmission,
synaptic plasticity, and cognition. Frontiers in Pharmacology,
11(September), 1–22. https://doi.org/10.3389/fphar.2020.581098,
PubMed: 33117176
Power, J. D., Mitra, A., Laumann, T. O., Snyder, A. Z., Schlaggar,
B. L., & Petersen, S. E. (2014). Methods to detect, characterize,
and remove motion artifact in resting state fMRI. NeuroImage,
84, 320–341. https://doi.org/10.1016/j.neuroimage.2013.08.048,
PubMed: 23994314
Ratcliff, R., & Rouder, J. (1998). Modeling response times for two-
choice decisions. Psychological Science, 9(5), 347–356. https://
doi.org/10.1111/1467-9280.00067
Ratcliff, R., Smith, P. L., Brown, S. D., & Mckoon, G. (2016).
Diffusion decision model: Current issues and history. Trends in
Cognitive Sciences, 20(4), 260–281. https://doi.org/10.1016/j
.tics.2016.01.007, PubMed: 26952739
Ratcliff, R., Thompson, C. A., & McKoon, G. (2015). Modeling in-
dividual differences in response time and accuracy in numeracy.
Cognition, 137, 115–136. https://doi.org/10.1016/j.cognition
.2014.12.004, PubMed: 25637690
Network Neuroscience
908
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
Reimer, J., Froudarakis, E., Cadwell, C. R., Yatsenko, D., Denfield,
G. H., & Tolias, A. S. (2014). Pupil fluctuations track fast
switching of cortical states during quiet wakefulness. Neuron,
84(2), 355–362. https://doi.org/10.1016/j.neuron.2014.09.033,
PubMed: 25374359
Reimer, J., McGinley, M. J., Liu, Y., Rodenkirch, C., Wang, Q.,
McCormick, D. A., & Tolias, A. S. (2016). Pupil fluctuations track
rapid changes in adrenergic and cholinergic activity in cortex.
Nature Communications, 7(May), 1–7. https://doi.org/10.1038
/ncomms13289, PubMed: 27824036
Richiardi, J., Altmann, A., Milazzo, A.-C., Chang, C., Chakravarty,
M. M., Banaschewski, T., Barker, G. J., Bokde, A. L. W.,
Bromberg, U., Büchel, C., Conrod, P., Fauth-Bühler, M., Flor,
H., Frouin, V., Gallinat, J., Garavan, H., Gowland, P., Heinz,
A., Lemaître, H., … IMAGEN Consortium. (2015). Correlated
gene expression supports synchronous activity in brain networks.
Science, 348(6240), 1241–1244. https://doi.org/10.1126/science
.1255905, PubMed: 26068849
Robbins, T. W., & Arnsten, A. F. T. (2009). The neuropsychopharma-
cology of fronto-executive function: Monoaminergic modulation.
Annual Review of Neuroscience, 32, 267–289. https://doi.org/10
.1146/annurev.neuro.051508.135535, PubMed: 19555290
Rojas-Líbano, D., Wainstein, G., Carrasco, X., Aboitiz, F., Crossley,
N., & Ossandón, T. (2019). A pupil size, eye-tracking and neu-
ropsychological dataset from ADHD children during a cognitive
task. Scientific Data, 6(1), 25. https://doi.org/10.1038/s41597
-019-0037-2, PubMed: 30975993
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Samuels, E., & Szabadi, E. (2008). Functional neuroanatomy of the
noradrenergic locus coeruleus: Its roles in the regulation of
arousal and autonomic function part II: Physiological and phar-
macological manipulations and pathological alterations of locus
coeruleus activity in humans. Current Neuropharmacology, 6(3),
254–285. https://doi.org/10.2174/157015908785777193,
PubMed: 19506724
Santana, N., & Artigas, F. (2017). Laminar and cellular distribution
of monoamine receptors in rat medial prefrontal cortex. Frontiers
in Neuroanatomy, 11(87), 1–13. https://doi.org/10.3389/fnana
.2017.00087, PubMed: 29033796
Sara, S. J. (2009). The locus coeruleus and noradrenergic modulation
of cognition. Nature Reviews Neuroscience, 10(3), 211–223.
https://doi.org/10.1038/nrn2573, PubMed: 19190638
Shenhav, A., Musslick, S., Lieder, F., Kool, W., Griffiths, T. L.,
Cohen, J. D., & Botvinick, M. M. (2017). Toward a rational
and mechanistic account of mental effort. Annual Review of
Neuroscience, 40(1), 99–124. https://doi.org/10.1146/annurev
-neuro-072116-031526, PubMed: 28375769
Shine, J. M. (2019). Neuromodulatory influences on integration and
segregation in the brain. Trends in Cognitive Sciences, 23(7),
572–583. https://doi.org/10.1016/j.tics.2019.04.002, PubMed:
31076192
Shine, J. M. (2021). The thalamus integrates the macrosystems of
the brain to facilitate complex, adaptive brain network dynamics.
Progress in Neurobiology, 199, 101951. https://doi.org/10.1016/j
.pneurobio.2020.101951, PubMed: 33189781
Shine, J. M., Bissett, P. G., Bell, P. T., Koyejo, O., Balsters, J. H.,
Gorgolewski, K. J., Moodie, C. A., & Poldrack, R. A. (2016).
The dynamics of functional brain networks: Integrated network
states during cognitive task performance. Neuron, 92(2), 544–554.
https://doi.org/10.1016/j.neuron.2016.09.018, PubMed: 27693256
Shine, J. M., Breakspear, M., Bell, P. T., Ehgoetz Martens, K., Shine,
R., Koyejo, O., Sporns, O., & Poldrack, R. A. (2019). Human cog-
nition involves the dynamic integration of neural activity and neu-
romodulatory systems. Nature Neuroscience, 22(2), 289–296.
https://doi.org/10.1038/s41593-018-0312-0, PubMed: 30664771
Shine, J. M., Hearne, L. J., Breakspear, M., Hwang, K., Müller, E. J.,
Sporns, O., Poldrack, R. A., Mattingley, J. B., & Cocchi, L. (2019).
The low-dimensional neural architecture of cognitive complexity
is related to activity in medial thalamic nuclei. Neuron, 104(5),
849–855. https://doi.org/10.1016/j.neuron.2019.09.002,
PubMed: 31653463
Shine, J. M., Müller, E. J., Munn, B., Cabral, J., Moran, R. J., &
Breakspear, M. (2021). Computational models link cellular
mechanisms of neuromodulation to large-scale neural dynamics.
Nature Neuroscience, 24(6), 765–776. https://doi.org/10.1038
/s41593-021-00824-6, PubMed: 33958801
Shine, J. M., van den Brink, R. L., Hernaus, D., Nieuwenhuis, S., &
Poldrack, R. A. (2018). Catecholaminergic manipulation alters
dynamic network topology across cognitive states. Network
Neuroscience, 2(3), 381–396. https://doi.org/10.1162/netn_a
_00042, PubMed: 30294705
Smiley, J. F., Subramanian, M., & Mesulam, M. M. (1999).
Monoaminergic-cholinergic interactions in the primate basal
forebrain. Neuroscience, 93(3), 817–829. https://doi.org/10
.1016/S0306-4522(99)00116-5, PubMed: 10473248
Thompson, W. H., Richter, C. G., Plavén-Sigray, P., & Fransson, P.
(2018). Simulations to benchmark time-varying connectivity
methods for fMRI. PLoS Computational Biology, 14(5), 1–23.
https://doi.org/10.1371/journal.pcbi.1006196, PubMed:
29813064
van den Brink, R. L., Pfeffer, T., Warren, C. M., Murphy, P. R.,
Tona, K.-D., van der Wee, N. J. A., Giltay, E., van Noorden,
M. S., Rombouts, S. A. R. B., & Donner, T. H. (2016).
Catecholaminergic neuromodulation shapes intrinsic MRI func-
tional connectivity in the human brain. Journal of Neuroscience,
36(30), 7865–7876. https://doi.org/10.1523/ JNEUROSCI.0744
-16.2016, PubMed: 27466332
Varazzani, C., San-Galli, A., Gilardeau, S., & Bouret, S. (2015).
Noradrenaline and dopamine neurons in the reward/effort
trade-off: A direct electrophysiological comparison in behaving
monkeys. Journal of Neuroscience, 35(20), 7866–7877. https://
doi.org/10.1523/jneurosci.0454-15.2015, PubMed: 25995472
Varela, F., Lachaux, J.-P., Rodriguez, E., & Martinerie, J. (2001). The
brainweb: Phase synchronization and large-scale integration.
Nature Reviews Neuroscience, 2(4), 229–239. https://doi.org/10
.1038/35067550, PubMed: 11283746
Vos de Wael, R., Benkarim, O., Paquola, C., Lariviere, S., Royer, J.,
Tavakol, S., Xu, T., Hong, S. J., Langs, G., Valk, S., Misic, B.,
Milham, M., Margulies, D., Smallwood, J., & Bernhardt, B. C.
(2020). BrainSpace: A toolbox for the analysis of macroscale
gradients in neuroimaging and connectomics datasets.
Communications Biology, 3(1). https://doi.org/10.1038/s42003
-020-0794-7, PubMed: 32139786
Network Neuroscience
909
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
The ascending arousal system shapes mesoscale network integration
Wagenmakers, E. J., Van Der Maas, H. L. J., & Grasman, R. P. P. P.
(2007). An EZ-diffusion model for response time and accuracy.
Psychonomic Bulletin and Review, 14(1), 3–22. https://doi.org
/10.3758/BF03194023, PubMed: 17546727
Wainstein, G. (2021). MOT, GitHub, https://github.com
/gabwainstein/MOT
Wainstein, G., Rojas-Líbano, D., Alnæs, D., Kolskår, K., Endestad,
T., Laeng, B., Ossandon, T., Crossley, N., Matar, E., & Shine,
J. M. (2020). MOT_data.mat ( Version 1), Figshare. https://doi
.org/10.6084/m9.figshare.13244504.v1
Wainstein, G., Rojas-Líbano, D., Crossley, N. A., Carrasco, X.,
Aboitiz, F., & Ossandón, T. (2017). Pupil size tracks attentional
performance in attention-deficit/hyperactivity disorder. Scientific
Reports, 7(1), 1–9. https://doi.org/10.1038/s41598-017-08246-w,
PubMed: 28811624
Wang, M., Ramos, B. P., Paspalas, C. D., Shu, Y., Simen, A.,
Duque, A., Vijayraghavan, S., Brennan, A., Dudley, A., & Nou,
E. (2007). A2A-adrenoceptors strengthen working memory net-
works by inhibiting cAMP-HCN channel signaling in prefrontal
cortex. Cell, 129(2), 397–410. https://doi.org/10.1016/j.cell
.2007.03.015, PubMed: 17448997
Wang, X. J. (2020). Macroscopic gradients of synaptic excitation
and inhibition in the neocortex. Nature Reviews Neuroscience,
21(3), 169–178. https://doi.org/10.1038/s41583-020-0262-x,
PubMed: 32029928
Wilkinson, G. N., & Rogers, C. E. (1973). Symbolic description of
factorial models for analysis of variance. Journal of Applied
Statistics, 22(3), 392–399. https://doi.org/10.2307/2346786
Yüzgeç, Ö., Prsa, M., Zimmermann, R., & Huber, D. (2018). Pupil size
coupling to cortical states protects the stability of deep sleep via
parasympathetic modulation. Current Biology, 28(3), 392–400.
https://doi.org/10.1016/j.cub.2017.12.049, PubMed: 29358069
Zerbi, V., Floriou-Servou, A., Markicevic, M., Vermeiren, Y.,
Sturman, O., Privitera, M., Von Ziegler, L., Ferrari, K. D., Weber,
B., De Deyn, P. P., Wenderoth, N., & Bohacek, J. (2019). Rapid
reconfiguration of the functional connectome after chemogenetic
locus coeruleus activation. Neuron, 103(4), 702–718. https://doi
.org/10.2139/ssrn.3334983
Zilles, K., & Palomero-Gallagher, N. (2017). Multiple transmitter re-
ceptors in regions and layers of the human cerebral cortex.
Frontiers in Neuroanatomy, 11, 1–26. https://doi.org/10.3389
/fnana.2017.00078, PubMed: 28970785
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
4
8
9
0
1
9
7
5
1
7
4
n
e
n
_
a
_
0
0
2
0
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
910