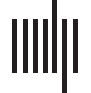RECHERCHE
Brain connectivity alterations during sleep by
closed-loop transcranial neurostimulation
predict metamemory sensitivity
Ryan J. Hubbard1,2*
, Iman Zadeh1,3*, Aaron P. Jones4, Bradley Robert4,
Natalie B. Bryant4, Vincent P. Clark4, and Praveen K. Pilly1
1Center for Human-Machine Collaboration, Information and Systems Sciences Laboratory,
HRL Laboratories, LLC, Malibu, Californie, Etats-Unis
2Beckman Institute for Advanced Science and Technology, University of Illinois Urbana-Champaign, IL, Etats-Unis
3Oracle Cloud Infrastructure, Oracle Corporation, 500 Oracle Parkway Redwood Shores, Californie, Etats-Unis
4Psychology Clinical Neuroscience Center, Département de psychologie, The University of New Mexico, Albuquerque, NM, Etats-Unis
*R.. J.. Hubbard and I. Zadeh contributed equally to this work.
Mots clés: Brain stimulation, EEG, Metamemory, Functional connectivity, Graph theory, Sleep
ABSTRAIT
Metamemory involves the ability to correctly judge the accuracy of our memories. The retrieval
of memories can be improved using transcranial electrical stimulation (tES) pendant
dormir, but evidence for improvements to metamemory sensitivity is limited. Applying tES can
enhance sleep-dependent memory consolidation, which along with metamemory requires
the coordination of activity across distributed neural systems, suggesting that examining
functional connectivity is important for understanding these processes. Nevertheless, little
research has examined how functional connectivity modulations relate to overnight changes
in metamemory sensitivity. Ici, we developed a closed-loop short-duration tES method,
time-locked to up-states of ongoing slow-wave oscillations, to cue specific memory replays
in humans. We measured electroencephalographic (EEG) coherence changes following
stimulation pulses, and characterized network alterations with graph theoretic metrics. Using
machine learning techniques, we show that pulsed tES elicited network changes in multiple
frequency bands, including increased connectivity in the theta band and increased efficiency
in the spindle band. En plus, stimulation-induced changes in beta-band path length were
predictive of overnight changes in metamemory sensitivity. These findings add new insights
into the growing literature investigating increases in memory performance through brain
stimulation during sleep, and highlight the importance of examining functional connectivity
to explain its effects.
RÉSUMÉ DE L'AUTEUR
Numerous studies have demonstrated a clear link between sleep and memory—namely,
memories are consolidated during sleep, leading to more stable and long-lasting representations.
We have previously shown that tagging episodes with specific patterns of brain stimulation during
encoding and replaying those patterns during sleep can enhance this consolidation process to
improve confidence and decision-making of memories (metamemory). Ici, we extend this work
to examine network-level brain changes that occur following stimulation during sleep that predict
metamemory improvements. Using graph theoretic and machine-learning methods, we found that
stimulation-induced changes in beta-band path length predicted overnight improvements in
un accès ouvert
journal
Citation: Hubbard, R.. J., Zadeh, JE.,
Jones, UN. P., Robert, B., Bryant, N. B.,
Clark, V. P., & Pilly, P.. K. (2021).
Brain connectivity alterations during
sleep by closed-loop transcranial
neurostimulation predict metamemory
sensitivity. Neurosciences en réseau, 5(3),
734–756. https://doi.org/10.1162/netn_a
_00201
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00201
Informations complémentaires:
https://doi.org/10.1162/netn_a_00201
Reçu: 6 Janvier 2021
Accepté: 15 May 2021
Corresponding Authors:
Praveen K. Pilly
pkpilly@hrl.com
Ryan J. Hubbard
rjhubbard@hrl.com
Éditeur de manipulation:
James Shine
droits d'auteur: © 2021
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
metamemory. This novel finding sheds new light on the neural mechanisms of memory
consolidation and suggests potential applications for improving metamemory.
INTRODUCTION
The brain has the remarkable ability to turn brief encounters and episodes, even “one-shot”
encounters, into long lasting memories. This occurs through a process known as memory con-
solidation, in which memories in a labile state are replayed during sleep and converted to
more stable representations. Cependant, successful retrieval of memories involves control pro-
cesses and decision-making, and even memories that are consolidated during sleep may be
difficult to recall, or retrieved with little confidence in their veracity (Koriat & Goldsmith,
1996). Metamemory sensitivity, or an individual’s ability to judge the accuracy of their mem-
ories with confidence, plays a critical role in the usage of our memories. Par exemple, an eye-
witness to a crime may have successfully encoded critical details of the episode, but may not
be confident in their memory, leading to incorrect decisions (Luus & Wells, 1994; Memon
et coll., 2003; Sporer et al., 1995). Ainsi, improving not only memory retrieval, but also meta-
memory sensitivity, is of critical importance. Ici, we investigated the improvement of sen-
sitivity with an intervention while individuals slept.
During sleep, neuronal ensembles representing previously encoded memories are reactivated
in both the hippocampus and neocortical areas (Euston et al., 2007; Ji & Wilson, 2007; Nádasdy
et coll., 1999; Sirota et al., 2003; Skaggs & McNaughton, 1996; Wilson & McNaughton, 1994).
Memory replays are predominantly observed during slow-wave sleep, particularly during the
positive phases, or up-states, of the ongoing 0.5–1.2 Hz oscillation (Lee & Wilson, 2002;
Mölle & Born, 2011). Reactivations of encoding-specific neuronal patterns are accompanied
by 12–15 Hz thalamo-cortical oscillatory activity known as spindles, as well as short-lived
high-frequency bursts in the hippocampus called ripples (De Gennaro & Ferrara, 2003; Mölle
et coll., 2006). The intricate coordination of replays, spindles, and ripples is essential to facilitate
the consolidation of memories into long-term storage, or to transfer memories from the hippocam-
pus to the neocortex (McClelland et al., 1995; McGaugh, 2000; Rasch & Born, 2013; Staresina
et coll., 2015). Consolidation of memories not only may facilitate their later retrieval, but also may
be related to the individual’s future confidence in them; namely, consolidation can strengthen
memories and improve learning (Walker & Stickgold, 2004), and confidence in memories is
related to memory fidelity (Dallenbach, 1913). Ainsi, targeting the consolidation process with
an intervention could benefit not only retrieval success, but metamemory sensitivity.
Over the past decade, researchers have increasingly investigated ways of boosting memory
consolidation processes through external manipulations. These intervention studies have
shown that sleep-dependent memory consolidation can be enhanced in two ways. D'abord,
memory reactivations can be triggered during sleep by reexposing individuals to external
sensory cues, such as odors or sounds that were present during encoding (Antony et al.,
2012; Oudiette & Paller, 2013; Schreiner & Rasch, 2014; Rasch et al., 2007; Rudoy et al.,
2009). This cued reactivation can lead to benefits in recall for the specific items previously
associated with the cues. Deuxième, multiple studies have shown that applying transcranial elec-
trical stimulation (tES) at particular frequencies to the brain during sleep can potentiate endog-
enous electrophysiological processes, leading to facilitation of memory consolidation and
subsequent recognition or recall (Ketz et al., 2018; Ladenbauer et al., 2016, 2017; Lustenberger
et coll., 2016; Marshall et al., 2006, 2004; Westerberg et al., 2015). These studies have demonstrated
735
Transcranial electrical stimulation (tES):
Noninvasive brain stimulation
technique in which electrical current
flows from an anodal electrode to a
cathode placed on the scalp. Petit
amounts of current pass through the
skin and skull, reaching neurons in
the brain and increasing membrane
excitability.
Neurosciences en réseau
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
a general increase in memory retrieval performance following tES during sleep. Surtout, ce
stimulation-related benefit to memory could potentially be due to frequency-specific alterations
in functional connectivity between brain regions (Krause et al., 2017).
To summarize, previous research indicates that retrieval of memories can be strengthened
through neurostimulation during sleep, and would potentially suggest that individuals would
also have greater metamemory sensitivity, or greater correlation in the accuracy and confi-
dence for their memory decisions, for these episodes. En effet, some research has demonstrated
a relationship between healthy uninterrupted sleep and intact metamemory judgments (Daurat
et coll., 2010). Cependant, this may not be the case, as other research has demonstrated dissoci-
ations between first-order decisions (recognition judgments) and second-order decisions (con-
fidence judgments; Del Cul et al., 2009; Hebart et al., 2016; Rounis et al., 2010). This is
important, as memory confidence can decrease over time (Shapira & Pansky, 2019), leading
to errors in memory reporting and poorer decision-making. Neural stimulation of the prefrontal
cortex has been shown to improve memory monitoring for general knowledge questions (Chua
& Ahmed, 2016; Chua et al., 2017), and theta-burst stimulation to depress activity of the
frontopolar cortex influenced metacognitive judgments (Ryals et al., 2016), suggesting tES
techniques could be effective for improving and maintaining memory sensitivity for newly
one-shot encoded episodes. En effet, recent work in our lab has shown that unique spatiotem-
poral amplitude-modulated patterns (STAMPs) of tES can be used to boost the sleep consoli-
dation and sensitivity of judgments of specific episodic memories acquired in immersive
virtual reality (Pilly et al., 2020).
In this paper, we extend previous work on cueing memory reactivation by investigating
changes in functional connectivity following short-duration tES patterns (namely, STAMPs)
during sleep. Functional connectivity has previously been shown to be affected by tES during
wake (Polanía et al., 2011, 2012), as well as by memory consolidation during sleep (Mölle
et coll., 2004). En plus, consolidation of memories in the brain is thought to be a
systems-level process, in that it is supported by a combination of short-range and long-range
communication across brain structures (Staresina et al., 2015). This is likely similar for meta-
mémoire, as research has shown connectivity between a distributed network of brain areas,
including the frontal cortex, precuneus, and hippocampus supports memory and metacogni-
tive judgments (Baird et al., 2013; Molenberghs et al., 2016; Morales et al., 2018; Ren et al.,
2018; Ye et al., 2019). Ainsi, understanding how changes in functional connectivity relate to
this process is critical; cependant, to our knowledge no study to date has examined how changes
in functional connectivity due to stimulation during sleep might affect or relate to memory
consolidation and decision-making processes.
To examine changes in functional connectivity, we used measures of EEG coherence (Nunez,
1995), specifically the imaginary part of coherency (Nolte et al., 2004), and extracted features of
connectivity from the coherence data with graph theoretical analyses (Bullmore & Sporns, 2009;
Sporns, 2003). This approach models the functional connectivity of the brain as an interconnected
graph, and allows for exploration of the relationship of network structure and function. While we
were interested in connectivity changes in the Spindle band, we expanded the analysis to include
several other frequency bands, as activity in many spectral bands have been related to memory
processes (Hanslmayr & Staudigl, 2014; Hanslmayr et al., 2012; Lisman & Jensen, 2013). We then
employed machine learning–based techniques to determine the important graph theoretic fea-
tures for discriminating between Active and Sham stimulation conditions, as well as predicting
overnight changes in episodic memory behavior. In this way, we provide novel insight into mod-
ulations in functional connectivity following pulsed tES that are related to changes in metamemory
sensitivity for specific episodic memories.
Spatiotemporal amplitude-
modulated pattern (STAMP):
tES in which the distributed pattern of
electrical stimulation across the scalp
changes rapidly through time.
Electroencephalography (EEG):
Brain imaging technique in which
electrodes are placed on an
individual’s scalp, allowing for
recording of electrical activity
propagated from the brain to the
surface of the head.
Neurosciences en réseau
736
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
MATERIALS AND METHODS
The participants reported in this paper are the same group of participants from Pilly et al.
(2020). They received unique brief spatiotemporal patterns of tES (namely, STAMPs) pendant
encoding of episodic information, half of which were reapplied during up-states of slow-wave
oscillations (SWOs) during subsequent nights to cue reactivation of the specific associated
memories (Active condition). At another point in time, the same individuals also performed
the memory task without brain stimulation (Sham condition). Donc, we were interested
not only in the changes in functional connectivity that differed between the Active and
Sham stimulation conditions, but also in the connectivity changes following STAMPs that were
related to changes in the recall of specific episodic memories from pre- to postsleep.
Participants
A total of 30 healthy participants completed the experiment, who were recruited using flyers
placed around campus of the University of New Mexico and surrounding community, et
received monetary compensation upon completion of the study. Of these, six participants
were excluded from the analyses due to either equipment failure to stimulate during an
Active night, or noncompliance in following task instructions. The sleep EEG data from addi-
tional six participants could not be used to calculate functional connectivity measures due to
excessive artifacts, leading to the inclusion of N = 18 participants in the final analysis and
reporting. All participants provided signed informed consent to participate in the study, lequel
was approved by the Chesapeake Institutional Review Board. All participants were native
English speakers, had normal or corrected-to-normal hearing and vision, and had no history
of neurological or psychiatric disorder, or drug abuse.
Behavioral Paradigm and Procedure
An outline of the experimental procedure is presented in Figure 1. The experiment was made up
of an acclimation period to train participants and let them sleep in the lab, followed by two
experimental nights involving learning and testing. The acclimation night was only to allow
participants to become accustomed to sleeping in the lab, and EEG data was not recorded or
analyzed from this period. Participants encoded information on the first experimental night. Le
memory task consisted of viewing virtual reality episodes administered with an HTC Vive VR
headset, followed by several memory recall tests on details from the episodes. Participants
encoded 14 virtual reality vignettes, each lasting about a minute, depicting a series of events
with two or more characters committing some action around an apartment complex. An addi-
tional vignette that was longer in length was used to train participants during the acclimation
period. Participants were tested on their memory for the vignettes across five test sessions
administered over a period of 48 hr with a non-VR computerized task built in MATLAB. Pour
each vignette, 10 test items were constructed that consisted of True/False statements on specific
aspects of the vignette. Each of the five test lists contained 28 items, 2 for each vignette.
Participants reported whether the test statements were True/False, as well as the confidence
of their recall on a scale of 1–10. Participants slept in the lab for each of the three nights.
The procedure consisted of four experimental sessions and one acclimation session. In the
acclimation session, participants viewed a long practice vignette and answered practice test
questions, and afterwards slept in the lab. In the following first experimental session (Session 1),
participants encoded the experimental stimuli. Half of the participants received unique tES
with unique STAMPs during viewing of vignettes (Active condition), whereas the other half did
not receive any stimulation (Sham condition). The participants received the opposite
Neurosciences en réseau
737
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 1. Outline of experimental procedure, reprinted with permission from Pilly et al. (2020). (UN) After an acclimation night, participants
viewed episodes accompanied by either Active or Sham spatiotemporal amplitude-modulated patterns (STAMPs) stimulation. After a gap of
temps, participants viewed more episodes accompanied by the other stimulation condition. (B) Encoding of episodes occurred in a virtual
environment. STAMP stimulation occurred during episode viewing. Following encoding, memory recall tests were given over a period of
2 days on details of the episodes. (C) During sleep, STAMPs were delivered during up-states of slow-wave oscillations (SWOs) to cue reac-
tivation of specific episodes. Scalp topography shows the amplitudes of current for an illustrative STAMP (red signifies positive and blue sig-
nifies negative currents).
stimulation condition over two additional experimental nights separated by about 1 week.
After completing the vignette viewing procedure, participants were given their first memory
recall test, and then slept overnight in the lab. This night, and not the acclimation night,
was considered “Night 1.” The participants in the Active condition received half of the
STAMPs during the night to cue consolidation of specific memories during the night. Dans
Neurosciences en réseau
738
Connectivity changes by tES during sleep predict meta-memory
contraste, participants in the Sham condition did not receive stimulation during the night. Note
that this study design allowed us to compare memory performance for episodes that received
STAMP stimulation during encoding and sleep (Tag & Cue) to episodes that only received
stimulation at encoding (Tag & No Cue). For the connectivity analyses, we focused on the
Tag & Cue and Sham (no encoding or sleep stimulation) conditions. After the participant
awoke, they were given a second memory test, and the experimental session concluded.
The second experimental session (Session 2) occurred in the evening after the first session.
Participants were given a third memory test in the evening, went to sleep, and received
another memory test after waking. This second period of sleeping in the lab was considered
“Night 2.” Active group participants once again received STAMPs during the night. The final
memory test was administered later in the evening. For the opposite stimulation condition
(Sessions 3 et 4), the participants viewed a new set of 14 vignettes and were administered
corresponding memory recall tests over 2 jours. In this way, the experiment was a within-
subjects manipulation, with the Active and Sham conditions occurring approximately 1 week
à part. The assignment of the stimulation condition order (Active first vs. Sham first), ainsi que
the assignment of vignettes, were counterbalanced across participants. The analyses reported
in this paper focus on Night 2, where the largest behavioral effects of STAMP stimulation were
trouvé (see Pilly et al., 2020, for further details).
Electrophysiological Recordings and Transcranial Stimulation
EEG data was collected and stimulation was applied simultaneously with a prototype StarStim64
from Neuroelectrics, Inc., with a 64-channel neoprene head cap including 32 EEG channels and
32 stimulation channels. EEG channels were placed in a montage according to the 10-10
système. EEG data was collected from 23 of the 32 sites sampled at 500 Hz and referenced to
channel Fz. The additional 9 electrodes recorded electrooculogram, electromyogram, and elec-
trocardiogram to allow for detection of sleep stages. No online hardware filtering, except line
bruit (60 Hz) filtering, was applied during collection.
Active transcranial stimulation was delivered in the form of unique arrays of spatiotemporal
currents across the 32 stimulation electrodes. The total positive electrical current was set to be
sous 2.5 mA, with each individual channel’s current under 1.5 mA and over 150 uA. Le
patterns of stimulation were created through gradient descent optimization to maximize orthog-
onality across the induced electric fields in the 3-D volume of an adult human head template by
penalizing correlations and anticorrelations. During the experiment, for the Active condition,
each of the 14 vignettes were randomly associated or “tagged” with a different pattern of
stimulation, and custom templates for each pattern were programmed in the Neuroelectrics
stimulation control software, CoreGUI. Only half of these tES patterns were reapplied during
the night to cue memory reactivation (Tag & Cue), and behavioral analyses focused on memory
recall test responses for the targeted vignettes. The unique patterns were primarily designed to
cue specific memories, not to alter neural activity in specific ways; thus, brain responses and
connectivity changes to individual STAMPs were not analyzed, and instead were combined
to increase trial counts.
Slow-Wave Oscillation Detection
Stimulation was automatically delivered time-locked to up-states of SWOs with a closed-loop
detection algorithm, adapted from a previously developed algorithm (Cox et al., 2014). During
dormir, incoming EEG data from 13 fronto-parieto-central channels (Cz, FC1, FC2, CP1, CP2,
Fz, C4, Pz, C3, F3, F4, P3, and P4) were stored in a running 5-s buffer that was continuously
Neurosciences en réseau
739
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
updated. To clean the data, moving average subtraction with a 1-s window was applied, et
any channels exceeding 500 μV min-to-max amplitude across the buffer period were rejected.
The remaining channels were averaged together to create a virtual channel for robust SWO
detection. Power within the slow-wave band (0.5–1.2 Hz) was compared to total broadband
pouvoir (0.1–250 Hz), and if this ratio was greater than 0.3, then the precise peak of power in the
slow-wave band (the center frequency) was computed. The virtual channel was then filtered
with a second-order Butterworth bandpass filter with a 1-Hz bandwidth around the center
frequency (with a minimum lower cutoff of 0.1 Hz), and a Hilbert transform was applied. Le
phase was shifted back 90 degrees, and the imaginary component was extracted, yielding the
instantaneous phase. A sine wave at the center frequency with optimized phase, offset, and am-
plitude of the filtered signal was then generated. The sine wave was projected forward in time to
determine the next point of zero phase (c'est à dire., the start of the up-state), and stimulation was started
at this time point and lasted for the predicted up-state with ramp up and ramp down times of
100 ms. During Sham nights, stimulation time points were recorded, but no stimulation was
delivered. Ainsi, the comparison of Active and Sham nights were at the same time points of the
SWO, with the only difference being the delivery of STAMP stimulation.
Validation of up-state detection was performed on data from Sham nights to avoid artifacts
produced by STAMPs in Active nights. Markers were extracted from the time points of up-states
predicted by the sine wave fit. Dans certains cas, due to hardware delay, timed stimulation was
moved to the up-state following the immediately detected up-state, which may have increased
the variability of phases to some degree. Epochs time-locked to stimulation onset markers (−5 to
+5 s) were extracted from the raw EEG data and bandpass filtered in the 0.5–1.2 Hz range. Phase
values at each onset marker time point were calculated using the Hilbert transform. Mean phase
values across trials were calculated for each participant, and these mean phases were submitted
to a v-test testing for a difference from 0 degrees (Berens, 2009). Figures and statistics on the
validation of the closed-loop algorithm are presented in the Supporting Information.
EEG Data Processing and Graph Theoretic Analysis
EEG signals were analyzed off-line with EEGLAB and custom MATLAB scripts. To clean the
data and separate sources of noise from brain signal, the EEG recordings were down-sampled
à 250 Hz, rereferenced to the average of all electrodes, and bandpass filtered using a
Butterworth filter (0.1–70 Hz, 24 dB/oct). Ocular and nonneural artifacts were identified
and manually removed using the independent component analysis (ICA) method in EEGLAB
and its built-in infomax algorithm (Delorme & Makeig, 2004). The artifact-free data were
segmented in epochs containing −6.4 to −2.4 s prior to the onset of stimulation as a baseline
window, et 3 à 7 s following the offset of stimulation as an analysis window. For each
epoch, the power spectrum at each channel was calculated by applying the Welch spectral
estimation method on 2048-ms segments with a 1,024-ms (50%) overlap, with each segment
tapered by a Hamming window.
Functional connectivity and graph theoretic analyses were performed with the BioNeCt
Toolbox, a custom MATLAB toolbox for brain connectivity analysis (Casanova, El-Baz, &
Suri, 2017). Functional brain connectivity between brain regions were estimated by computing
the cross-correlation in the frequency domain between EEG signals (Bowyer, 2016;
Mohammad-Rezazadeh et al., 2016; Nolte et al., 2004). Among the various metrics of func-
tional connectivity, imaginary coherence (iCoh) exclusively detects “true” interactions
between EEG signals occurring within a certain time delay, thus ignoring instantaneous inter-
action between neighboring electrodes likely produced by volume conduction of electrical
740
Imaginary coherence:
A measure of functional connectivity
used in analysis of EEG data which
uses the phase information of
coherency. This method avoids the
issue of volume conduction, lequel
can highly inflate measures of
connectivité, but also leads to a more
conservative estimate of
connectivité.
Neurosciences en réseau
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
activity from a common brain source (Bullmore & Sporns, 2009; Fallani et al., 2014;
Mohammad-Rezazadeh et al., 2016). iCoh values were computed for all possible electrode
pairs in the following frequency bands: delta (1–4 Hz), theta (4–8 Hz), alpha (8–12 Hz), spin-
dle (12–15 Hz), beta (16–30 Hz), and low gamma (40–50 Hz). These analyses were performed
on both the Active and Sham data. En plus, iCoh values were computed for both the post-
stimulation and prestimulation time windows, and the prestimulation window values were
subtracted from the poststimulation windows values for baseline correction.
The high multidimensionality of the iCoh measures was disentangled with a graph theoretic
approche. Graph theoretic metrics provide information on the degree of network segregation
(c'est à dire., the tendency of brain regions to form local clusters with dense functional interconnec-
tion) and network integration and efficiency (c'est à dire., the capacity of the network to become in-
terconnected and efficiently exchange information between brain regions; Bullmore & Sporns,
2009; Fallani et al., 2014; Mohammad-Rezazadeh et al., 2016). The following commonly
used graph measures were calculated for all of the above mentioned frequency bands in
the pre- and poststimulation periods: average clustering coefficient (the probability of neigh-
boring nodes being connected to each other, reflecting local connectedness); global efficiency
(how efficient the network is in transferring information); characteristic path length, radius, et
diameter (the average number of edges along the shortest paths, the minimum possible dis-
tance, and the largest possible distance, between all possible pairs of nodes, respectivement);
modularity (the degree to which the brain network is segregated into subnetworks or modules);
assortativity (the proportion of nodes that are attached to other nodes with similar degrees vs.
dissimilar degrees); density (the number of edges divided by the number of nodes in the graph);
and mean coherence (the average coherence between all pairs of nodes).
Classification and Regression Analyses
The resulting data consisted of 48 functional connectivity features (8 graph theoretic metrics ×
6 frequency bands) for both the Active and Sham stimulation conditions for each participant.
More specifically, connectivity features were extracted following stimulation on Active nights
in which STAMP stimulation occurred, as well as during Sham nights in which no stimulation
occurred. To investigate what connectivity changes are predictive of stimulation condition, comme
well as relate to behavior, we implemented a participant-level classification and regression
cross-validation analysis, in which data from a subset of participants were left out as test data
to predict. To reduce variance, we ran 35 folds in which one to three participants were left out
at a time. This repeated random split method of sampling is preferred over the commonly im-
plemented “leave one out” method, which can lead to unstable and biased estimates of var-
iance (Scheinost et al., 2019; Varoquaux et al., 2017). We chose to take this data-driven
approach not only due to the large feature space of the data, but also due to the novelty of
the investigation. Few studies to date have examined relationships between graph theoretic
metrics of brain connectivity and metamemory sensitivity, and none have examined how this
changes with brain stimulation. Ainsi, we would potentially miss important relationships be-
tween functional connectivity and metamemory by only examining certain frequency bands
(par exemple., theta) or measures (par exemple., mean coherence) based on previous findings in the literature.
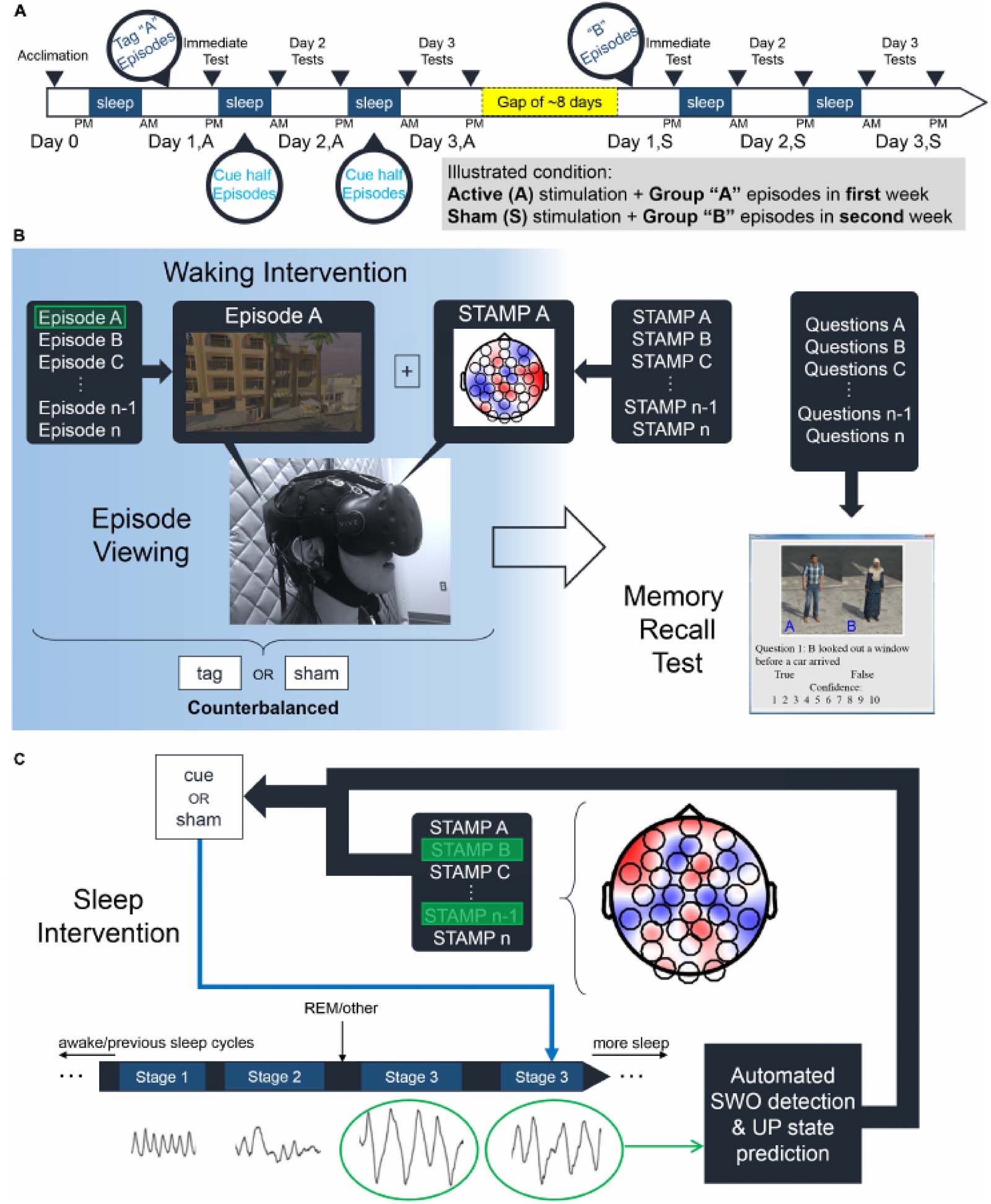
A diagram outlining the analysis pipeline is presented in Figure 2. For each fold, a feature
selection method was run on the training data to find graph theoretic features (poststimulation
baseline corrected with prestimulation) that discriminated between Active and Sham stimulation
conditions. Many features were highly correlated with one another, which can be seen in
Supporting Information Figure S2. Ainsi, first highly collinear variables were removed by finding
Neurosciences en réseau
741
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
Chiffre 2. Outline of analysis pipeline. Feature selection is performed for both the classification of Active vs. Sham condition, as well as the
prediction of overnight behavioral changes in the Active condition. The top features from each selection process are also combined and tested
for the classification and regression analyses.
pairs of features with high correlations (threshold r value of 0.8) and removing the variable with the
higher mean absolute correlation with the other variables. Additional information on removed
variables due to high collinearity can be found in the Supporting Information. Alors, a dependent
t test (as the study was a within-subjects design) was run on each feature, testing the difference
between Active and Sham conditions. Variables with low absolute t values (threshold of 1) étaient
removed, with the rationale that the difference between these variables was minimal.
The remaining features were entered into the Boruta feature selection algorithm (Kursa &
Rudnicki, 2010). Boruta is a permutation-based selection method, in which each feature, comme
well as a copy of the feature with its values shuffled across participants (called a “shadow
feature”), is entered into a random forest classifier, and the importance of each feature is deter-
mined as a Z score. The importance of the true features must be higher than the max Z score of
the shadow features to be considered significantly important. This procedure is repeated several
times with different permutations of the shadow features to obtain a distribution of values to
which the importance values of the actual features can be compared. Ainsi, Boruta is an
all-relevant feature selection method—essentially, all or none of the variables could be selected
by the process. Only features that had higher Z scores than the shadow features were selected.
This algorithm has been shown to improve performance in finding meaningful features com-
pared to other feature selection algorithms (Degenhardt et al., 2017; Kursa, 2017).
The features selected by Boruta were then entered into a logistic regression model predicting
Active versus Sham condition. We also ran a regularized logistic regression with L2 regularization
(ridge regression), in which the optimal λ parameter was determined through cross-validation
within the training data. Dernièrement, we also ran a logistic regression classification using only the most
important variable selected by the Boruta algorithm. After fitting the model parameters on the train-
ing data, the models were used to predict the left-out test data, yielding estimates of probability for
each test example either belonging to the Active or Sham condition. These probabilities were
concatenated across folds, and used to calculate relative operating characteristic curves, depuis
Neurosciences en réseau
742
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
which we derived area under the curve (AUC) valeurs (Fawcett, 2006). AUC is a more sensitive
measure of classifier performance than simply raw accuracy, and thus is reported here.
Within the same cross-validation fold, the training data was used to predict changes in
memory sensitivity for the targeted vignettes (namely, Tag & Cue) from the Night 2 presleep
test to the postsleep test in the Active condition. The measure of behavioral performance was
AUC of the type-2 relative operating characteristic curve, a bias-free nonparametric measure
of metacognitive sensitivity, with greater AUC reflecting greater memory sensitivity (Fleming &
Lau, 2014; Galvin et al., 2003). Ici, the models predicted the overnight change in AUC in
order to tie changes in connectivity with changes in memory sensitivity. All of the connectivity
features were entered into a similar feature selection process as the classification analysis.
Highly collinear features were removed as before. Instead of a t test, the correlation between
each feature and the overnight AUC change was computed, and features with low absolute r
valeurs (threshold of 0.1) were removed. The remaining features were run through the Boruta
algorithme, this time predicting overnight AUC changes. These features, as well as just the top
feature, were entered into linear regression and L2 regularized regression models predicting
left-out test data. Root-mean-square error (RMSE) values were calculated, and average
RMSE values across folds were compared to RMSE values from predictions from a linear re-
gression with only the intercept.
As a final analysis, we took Boruta-selected features from the classification analysis, ainsi que
the Boruta-selected features from the regression analysis, and reran classification and regression
with the combined feature set, as well as the top feature from each set. AUC values from clas-
sification analyses and RMSE values from regression analyses are reported for each feature set.
RÉSULTATS
Behavioral Effects
As reported in Pilly et al. (2020), STAMP stimulation did not lead to a significant improvement
of overall memory recall accuracy. Cependant, STAMP stimulation led to improved metamem-
ory sensitivity on Day 3 of the experiment (following Night 2). Spécifiquement, dependent t tests
comparing metamemory sensitivity across conditions revealed that sensitivity in the Tag & Cue
condition was significantly greater than both the Tag & No Cue condition [t(23) = 3.51,
adjusted p < 0.01, Holm–Bonferroni correction for two comparisons; paired-sample Cohen’s
d = 0.72] and Sham condition [t(23) = 2.089, adjusted p = 0.048, Holm–Bonferroni correction
for two comparisons; paired-sample Cohen’s d = 0.43]. A significant improvement in meta-
memory sensitivity was only found on Day 3 of the experiment. Thus, the application of
STAMPs during SWOs in the two nights following one-shot viewing led to specific enhance-
ment of metamemory for the episodes that were both tagged and cued.
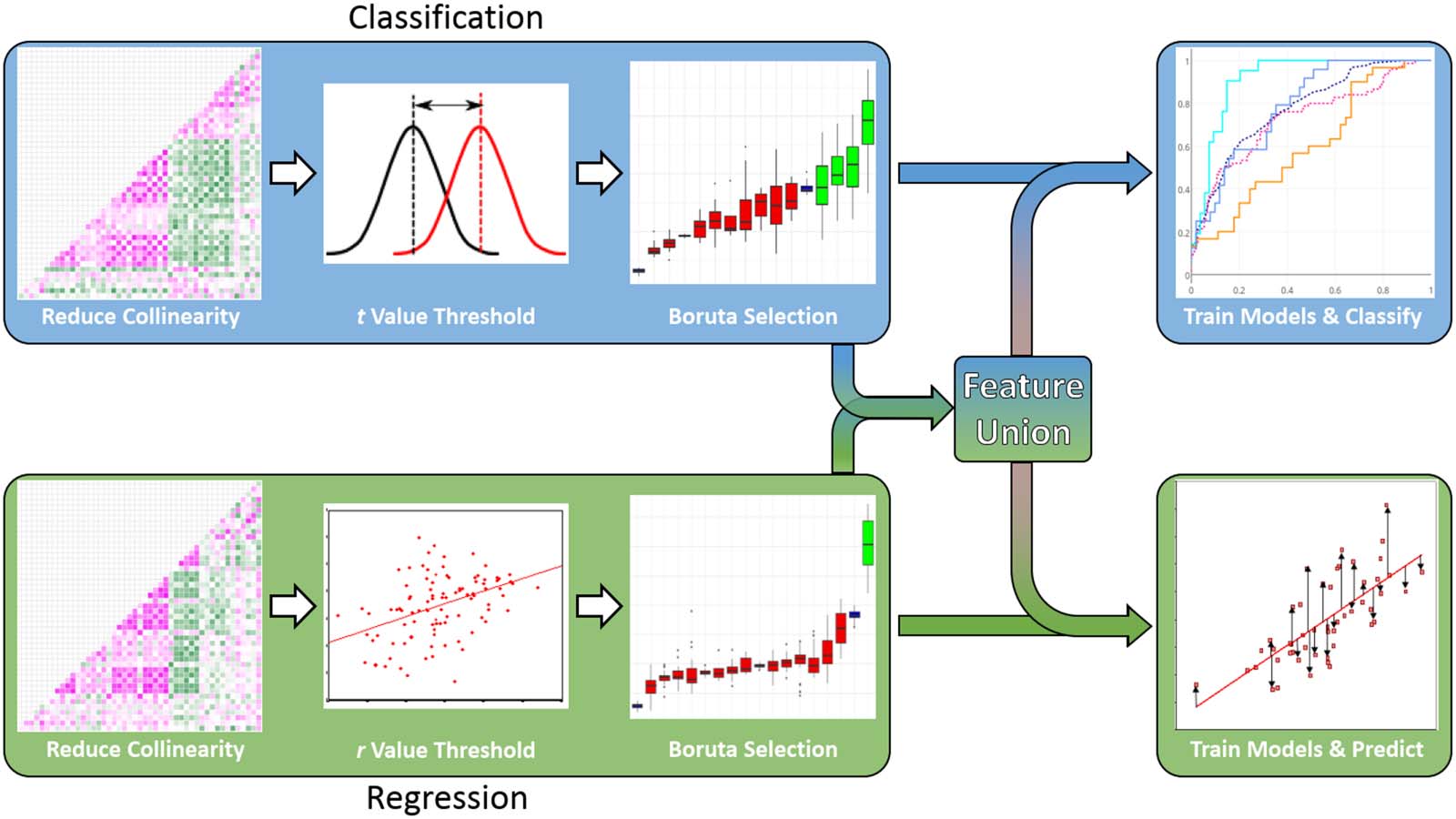
Global Coherence Changes
As a first pass, we examined overall changes in mean coherence across the scalp due to
STAMP stimulation to determine if stimulation led to any measurable changes in connectivity.
Additionally, we sought to replicate previous work and assess whether stimulation would pro-
duce similar changes in coherence as learning during sleep (Mölle et al., 2004). Previous work
(Mölle et al., 2004) reported modulation of delta, spindle, and gamma band coherence during
sleep following learning; in contrast, other work (Polanía et al., 2011) demonstrated increased
connectivity across all frequency bands following tDCS stimulation. Therefore, we hypothe-
sized that STAMP stimulation would increase mean coherence compared to the Sham condi-
tion in in the delta, spindle, and gamma bands, but examined differences in other bands as
Network Neuroscience
743
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
a
_
0
0
2
0
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
well with conservative statistical testing. Mean iCoh values in all frequency bands were
computed for Active and Sham baseline-corrected data in order to investigate global
connectivity changes, and thus one-tailed t tests were conducted on mean coherence
values in each frequency band, and were corrected for using a false discovery rate of p <
0.05 (Benjamini & Hochberg, 1995). Coherence maps showing greater connectivity in the
Active condition across the scalp were visualized by setting the visualization threshold to
the maximum of the Sham condition for each frequency band, essentially subtracting the
Sham condition.
Stimulation-induced mean coherence changes in different frequency bands are plotted in
Figure 3. The topography plots display the coherence between pairs of electrodes in the Active
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
a
_
0
0
2
0
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 3. Overall changes in mean coherence across the scalp induced by STAMPs. For each frequency band, the scalp plot shows the active
mean coherence data thresholded by the maximum coherence of the Sham data. The boxplot shows the mean coherence change from the
baseline period to the poststimulation period across the entire scalp across participants for both the Active and Sham conditions, and the *
symbol designates a significant difference ( p < 0.05).
Network Neuroscience
744
Connectivity changes by tES during sleep predict meta-memory
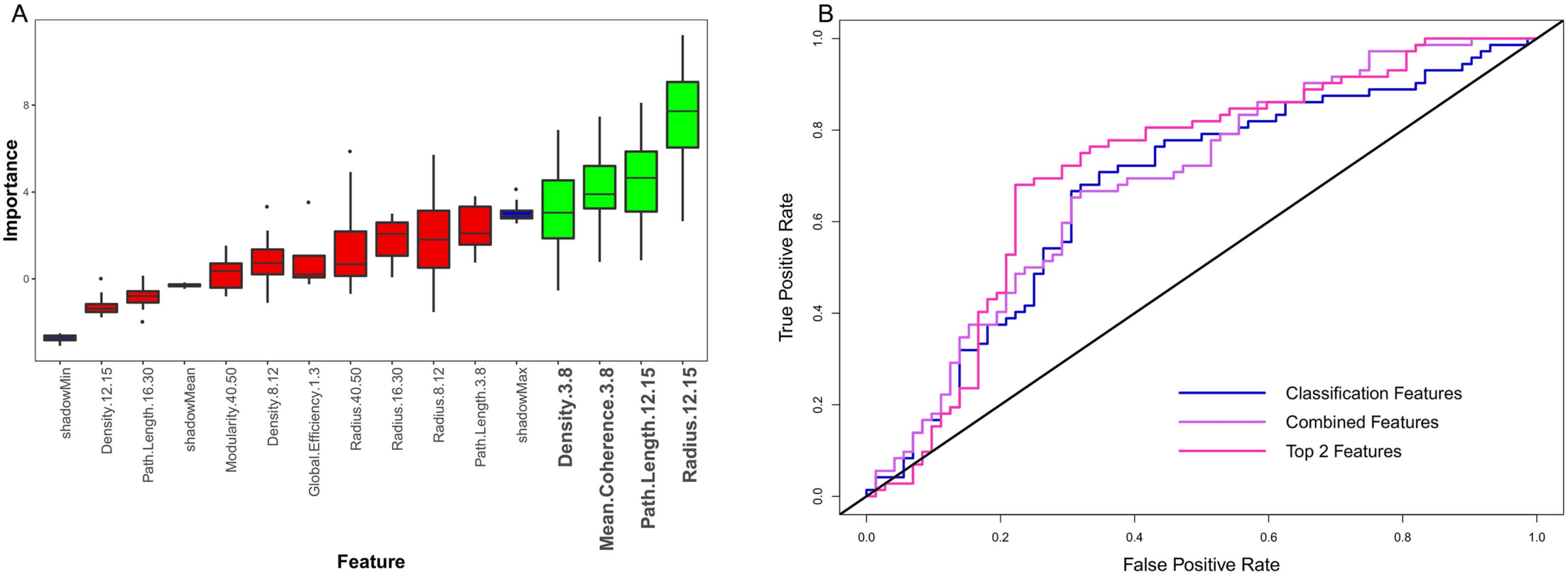
Figure 4. Classification results. (A) Boruta feature selection results, with red boxes showing rejected features, green boxes showing selected
features, and blue boxes showing shadow feature statistics. The selected features are in the theta and spindle bands. (B) Relative operating
characteristic curves for Active vs. Sham classification with different feature sets. All feature sets performed above chance accuracy, but the top
classification and regression features performed the best.
condition, thresholded by the coherence in the Sham condition, whereas the boxplots show
nonthresholded mean coherence values across the entire scalp. The analysis of overall changes
in coherence across the scalp revealed that STAMPs produced significant increases in mean
coherence in multiple frequency bands. EEG coherence was significantly increased following
STAMPs in the theta, alpha, and spindle bands ( p < 0.05) compared to the Sham condition.
Mean coherences in the delta, beta, and gamma bands did not significantly differ between
the stimulation conditions. Thus, STAMP stimulation induced changes in brain connectivity
in specific frequency bands.
Classification and Behavioral Prediction
Results from the classification analysis are plotted in Figure 4 and outlined in Table 1. Average
importance values across folds for Boruta-selected features are plotted on the left, with features
consistently considered important highlighted in green (significantly above the average shadow
max). Across folds, radius in the spindle band (12–15 Hz) was consistently selected as the most
important feature. Other important features included: path length (12–15 Hz) and mean coherence
and density (3–8 Hz). ROC curves of classifier performance are plotted on the right. As outlined in
Mason and Graham (2002), Mann–Whitney U tests were conducted on each classifier to determine
Table 1. AUC values and 95% bootstrap confidence intervals for classification using different
feature sets
Feature Set
Selected classification features
Combined classification and regression features
Top classification and regression features
AUC
0.671 (0.581–0.756)
0.689 (0.601–0.774)
0.715 (0.625–0.800)
745
Network Neuroscience
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
a
_
0
0
2
0
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
if classifier performance was statistically significant above chance (AUC = 0.5). All tests reported
significant p values ( p < 0.001). We additionally calculated 95% confidence intervals using a
stratified bootstrap resampling approach with 10,000 bootstrap replications, which further demon-
strated above chance classification performance (Carpenter & Bithell, 2000). Thus, classification
accuracy was above chance, and taking the top variables from classification and regression anal-
yses (path length in 16–30 Hz and radius in 12–15 Hz) led to the best classification performance.
As a follow-up analysis, the four features selected by the Boruta algorithm for classification were
submitted to t tests, testing the difference in connectivity between the Active and Sham conditions.
The resultant p values were corrected with a false discovery rate set at 0.05. The corrected p values
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
a
_
0
0
2
0
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
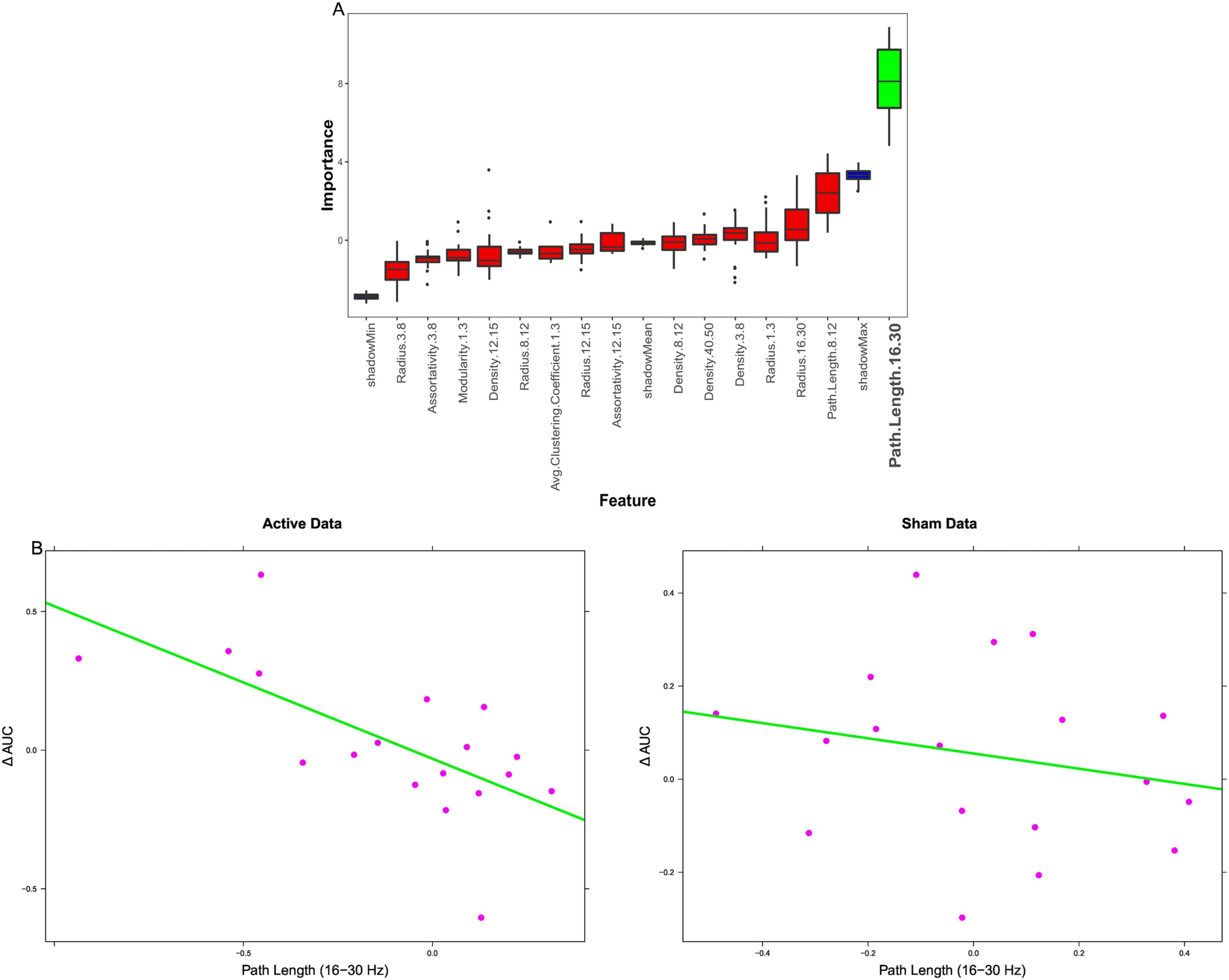
Figure 5. Regression results. (A) Boruta feature selection results, with red boxes showing rejected features, green boxes showing selected
features, and blue boxes showing shadow feature statistics. Beta path length is consistently selected across folds. (B) Relationship between beta
path length (x-axis) and overnight AUC change (y-axis) in the Active (left) and Sham (right) conditions. The relationship is significant in the
Active condition, not in Sham condition.
Network Neuroscience
746
Connectivity changes by tES during sleep predict meta-memory
Table 2.
RMSE values for regression using different feature sets
Feature Set
Base model (intercept only)
Selected regression features
Top regression feature
Combined classification and regression features
Top classification and regression features
RMSE
0.249
0.226
0.208
0.240
0.215
for 12–15 Hz path length and radius, and 3–8 Hz mean coherence were significant ( p = 0.029);
3–8 Hz density was also significant at alpha level of significance ( p = 0.05). Thus, connectivity
features that were significantly altered following Active stimulation compared to Sham stimulation
were found by the feature selection pipeline. Path length and radius features from pre- to post-
stimulation were significantly decreased in the Active condition compared to Sham, whereas
mean coherence and density features were significantly increased compared to Sham. Boxplots
showing these features for both conditions are presented in Supporting Information Figure S3.
Results from the behavioral prediction analysis are plotted in Figure 5 and outlined in
Table 2. Here, changes in neural measures following stimulation were used as features to pre-
dict change in recall test metamemory sensitivity (AUC) for both the Tag & Cue and Sham
conditions from Day 2 to Day 3 in order to identify brain-behavior relationships. The impor-
tance values from Boruta for graph theoretic features are plotted on the left—clearly, path
length in the beta band (16–30 Hz) was considered highly important for predicting changes
in metamemory sensitivity, and the only feature selected. The relationship between
stimulation-induced change in 16–30 Hz path length and overnight change in AUC perfor-
mance for both the Active (Tag & Cue) data and Sham conditions are plotted on the right.
The correlation between change in beta-band path length and change in AUC was significant
for the Active condition (r = −0.66, p = 0.003), indicating that individuals with decreased path
length following STAMPs tended to show a positive change in memory performance. This
relationship was not significant for the Sham condition (r = −0.22, p = 0.39).
Based on the outcome of these analyses, follow-up linear regressions were ran predicting
overnight change in metacognitive sensitivity using path length in the 16–30 Hz band for both
the Active data and the Sham data. For the Active data, 16–30 Hz path length significantly
predicted AUC change (t = −3.56, p = 0.003, adj. R2 = 0.41). For the Sham data, 16–30 Hz path
length was not a significant predictor (t = −0.88, p = 0.39, adj. R2 = −0.01). Additional analyses
to compare the Active and Sham conditions were also performed and are outlined further in the
Supporting Information.
DISCUSSION
In the present experiment, participants engaged in an episodic memory task while receiving
unique spatiotemporal patterns of transcranial electrical stimulation, and a subset of the patterns
were reapplied during up-states of SWOs in subsequent nights to cue reactivation of specific
associated memories (see Pilly et al., 2020). Here, we report changes in functional brain
connectivity induced by these tES patterns during sleep that were predictive of changes in
metamemory sensitivity from pre- to postsleep. Using machine learning techniques to identify
important features for classifying between Active and Sham stimulation, we found that
Network Neuroscience
747
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
a
_
0
0
2
0
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
connectivity features in the theta and spindle bands significantly differed between the two con-
ditions. Additionally, we found that changes in path length in the beta band predicted memory
sensitivity changes, in that performance increases across the night were related to beta path
length decreases following stimulation. Taken together, these results contribute to a growing
body of research investigating the neural processes underlying memory reactivations and con-
solidation during sleep, and suggest that modulation of path length at specific frequencies during
the consolidation process may lead to higher confidence and better decision-making for newly
consolidated memories.
The short bursts of tES, or STAMPs, utilized in the current experiment increased overall
scalp coherence in lower frequency bands—namely, in the theta, alpha, and spindle bands,
or essentially from 3–15 Hz. Interestingly, changes in spindle-band connectivity during sleep
was also found following learning; however, changes in connectivity were not found in the
delta or gamma bands following STAMP stimulation, which is somewhat at odds with pre-
vious experimental work (Mölle et al., 2004). However, the stimulation protocol used here
largely differed from previous studies—namely, we used short bursts of spatiotemporally dis-
tributed electrical currents that were primarily designed to cue specific memories due to
their uniqueness and association with the encoding period, not modulate or entrain specific
oscillations. Importantly, activity in the reported frequency bands, particularly the theta and
spindle bands, has been shown to be related to the memory consolidation process (Staresina
et al., 2015). Thus, STAMPs may have modulated the ongoing processes of memory reacti-
vation or consolidation during sleep, leading to the changes in connectivity in the observed
frequency bands.
Recent work has questioned the effectiveness of tES in significantly affecting behavior
(Horvath et al., 2015), as well as in its ability to modulate neural firing patterns or oscillatory
activity (Lafon et al., 2017; Vöröslakos et al., 2018). However, other studies have demonstrated
measurable neural changes in humans and nonhuman primates following tES, even at lower
intensity levels (Huang et al., 2017; Krause et al., 2017; Opitz et al., 2016). A particular con-
cern is the ability of tES, specifically alternating current stimulation, to entrain neural oscilla-
tions to a specific frequency. Here, we do not claim that the reported electrophysiological
changes are due to oscillatory entrainment to the short bursts of stimulation, or that we are
inducing these changes de novo, but rather that the STAMPs act to boost ongoing memory
consolidation processes, which occurs through subtle modulations of network-level connec-
tivity. We would argue that the observed connectivity patterns during sleep are natural phe-
nomena that occur and lead to memory consolidation, and STAMP stimulation acted upon
these preexisting processes to boost metamemory sensitivity.
The reported modulation of graph theoretic metrics in the theta and spindle bands following
STAMPs is in line with previous research implicating these oscillations in the processes of
memory reactivation and consolidation. Targeted memory reactivation during sleep with au-
ditory cues elicits increases in theta and spindle power (Ong et al., 2016; Schreiner et al.,
2015; Schreiner & Rasch, 2014). Numerous studies have also shown the importance of
theta-band activity for memory encoding and retrieval during wake (Backus et al., 2016;
Lega et al., 2012; Nyhus & Curran, 2010); in particular, theta oscillations in the hippocampus
are critical for episodic memory encoding (Hasselmo et al., 2002; Lin et al., 2017). Similarly,
thalamocortical spindles are related to episodic memory (Van Der Helm et al., 2011) and play
an important role in the process of memory consolidation, potentially facilitating the transfer of
newly encoded information in the medial temporal lobes to stable representations in the
cerebral cortex (Sirota et al., 2003; Staresina et al., 2015). During sleep, Theta activity may
be indicative of the memory reactivation process, whereas spindle activity may reflect the
Network Neuroscience
748
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
a
_
0
0
2
0
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
active consolidation of the reactivated memories. Here, STAMPs may have cued reactivation
of the associated memories, leading to boosted consolidation of this information. This consol-
idation led to improved confidence for memories downstream.
The theta-band graph theoretic measures that were significantly modulated by STAMPs were
mean coherence and density. Mean coherence refers to the average coherence across all ver-
tices, while density is calculated as the number of connections in the graph, normalized by the
total possible number of connections (Sporns, 2003). Changes in these measures suggest that
STAMPs increased overall coherence in the theta band between channels, leading to a greater
number of edges above the cutoff threshold and thus a higher average connection density. This
greater coherence and density may reflect an increase in the exchange of information in the theta
band across neural areas following STAMPs. This is in line with previous findings implicating
increases in theta coherence between the hippocampus and rhinal cortex (Fell et al., 2003), as
well as frontal cortex (Anderson et al., 2009; Benchenane et al., 2010), as important for learning
and memory, and potentially critical for memory reactivations (Carr et al., 2011).
The spindle-band graph theoretic measures that significantly differed following STAMP stim-
ulation were path length and radius. These measures are related—radius refers to the minimum
eccentricity of the graph and path length refers to the graph’s average eccentricity, where the
eccentricity of a node is defined as the maximum distance to any other node (Sporns, 2003).
Path length is often considered a measure of network integration or efficiency of information
transfer, with smaller path lengths reflecting greater efficiency (Bullmore & Sporns, 2009;
Reijneveld et al., 2007). Here, STAMPs led to an average decrease in path length and radius
in the spindle band, meaning the average and minimum eccentricity were decreased, and
efficiency of information transfer was increased. This may reflect a reorganization of the net-
work toward a small-world network (Watts & Strogatz, 1998), a near optimal structure between
a perfectly ordered and randomly organized system, allowing for greater flexibility and syn-
chronization of neural activity (Barahona & Pecora, 2002; Bassett & Bullmore, 2006;
Masuda & Aihara, 2004). Changes in “small-worldness” are often measured as changes in path
length without concomitant changes in local clustering, or cluster coefficient. Thus, the
observed change in path length may suggest boosted synchronization between brain areas,
leading to more efficient information transfer by spindles. Note that effects in the theta band
were more closely related to overall changes in mean coherence—essentially, changes in
the magnitude of connectivity as opposed to the efficiency. While the data here is unable to
establish a causal link between these changes, a hypothetical relationship is that STAMPs
caused greater connectivity in the theta band (Fell et al., 2003), and Spindle information transfer
efficiency, measured as path length, was ramped up to compensate for this increase, leading
to increased memory reactivation.
Across studies, there is high variability in the frequency bands chosen to delineate slow and
fast spindles (Ayoub et al., 2013; Holz et al., 2012; Schabus et al., 2007), and recent evidence
suggests these delineations may in fact vary in each individual (Cox et al., 2017). We selected
8–12 Hz and 12–15 Hz as bands to separately investigate slow versus fast spindles, and our
results suggest that connectivity in the fast spindle band was specifically modulated by
STAMPs. However, there may be variability in spindle-band delineations across the partici-
pants in the experiment, leading to overlap of slow and fast spindle activity in these bands.
In our data, path lengths in the 8–12 Hz and 12–15 Hz bands were highly correlated (r = 0.88),
suggesting a high degree of overlap between these frequency bands. Thus, we claim here that
spindle activity was modulated by STAMPs, but further analysis will be required to specifically
delineate between fast and slow spindles. Additionally, we used a frequency band–based ap-
proach to accommodate the connectivity and machine learning analysis we implemented;
Network Neuroscience
749
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
a
_
0
0
2
0
1
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
however, spindles are transient events that occur briefly in time. While the approach here
likely captured activity related to spindles, a more focused event-based approach in which
specific spindle events are identified may further elucidate their role in metacognition.
The results of the regression analysis showed that changes in path length in the beta band
were related to changes in memory sensitivity from presleep to postsleep in the Active condi-
tion. The reported relationship was in a sensible direction, with lower path length (greater
efficiency) related to improvements in metacognitive performance. Note that beta path length
did not, however, significantly differ between the Active and Sham conditions, suggesting the
effect captures more fine-grained individual differences, in which individuals that exhibit
network reorganization toward greater efficiency following STAMP stimulation show greater
overnight metamemory improvement than those that do not. This is discussed further in the
Supporting Information. This finding is fairly novel, as no prior study to our knowledge has
examined changes in functional connectivity following stimulation during sleep that relate
to behavioral changes. Studies investigating visuomotor learning and performance, such as
with mirror tracing tasks, have reported changes in beta-band synchronization and connectiv-
ity during and following the task (Classen et al., 1998; Kilner et al., 2004; Piantoni et al., 2015);
however, studies examining graph theoretic connectivity relationships to episodic memory
performance, which was tested in the current experiment, have generally not examined or
reported changes in beta-band connectivity (Burke et al., 2013). However, changes in beta
path length have been related to the progression of Alzheimer’s disease—namely,
Alzheimer’s disease patients exhibit higher beta path lengths than healthy controls, and beta
path length is negatively correlated to minimental state examination (MMSE) score (Stam et al.,
2006). Additionally, research in the oscillatory neurophysiology of sleep has demonstrated
that faster rhythmic activity (i.e., in the beta/gamma range) is present and sustained during
the depolarization phase of the slow wave and largely absent during the hyperpolarization
phase (Compte et al., 2008; Steriade, 2006; Steriade et al., 1996). This increased activity
may be specifically linked to noradrenergic neurons in the locus coeruleus system
(Eschenko et al., 2012), which may promote cortical plasticity, an important component
of memory consolidation. Thus, it is a possibility that beta-band network organization and
activity during SWOs may generally be an important aspect of episodic memory functioning
and consolidation processes, which can be targeted and enhanced through specific STAMP
stimulation.
One potential limitation of the current study is the lack of a control condition to rule out the
overall effect of stimulation on connectivity. Namely, STAMPs could have been applied during
sleep that were not tied to any episode during encoding to compare connectivity changes fol-
lowing tagged STAMPs to changes following untagged STAMPs. Given the limited number of
STAMPs and complexity of the experimental design, we chose to maximize our statistical
power by using all of the STAMPs to cue memory during sleep. However, this condition would
have been particularly useful for understanding our classification results. Currently, we can say
that the selected features can delineate the Active and Sham conditions, and this may be due
to consolidation processes in the Active condition compared to the Sham condition; however,
these may also be general changes following stimulation. In contrast, we are confident that the
beta path length relationship to metamemory sensitivity changes is related to sleep-dependent
consolidation processes. Our behavioral results show that STAMP stimulation led to benefits
for the Tag & Cue condition compared to the other conditions, demonstrating a specific benefit
as opposed to a more generic stimulation-induced benefit to metamemory. In line with this,
changes in beta path length specifically predicted changes in Tag & Cue AUC changes, and
was not predictive of sensitivity in the Sham condition.
Network Neuroscience
750
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
a
_
0
0
2
0
1
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
Previous work examining brain connectivity with graph theoretic measures has often taken
a more limited approach by examining specific measures or specific frequency bands of
interest (Sadaghiani et al., 2015; Song et al., 2014). We chose to utilize a machine learning
approach of searching through a large collection of measures in different frequency bands to
identify significant changes following stimulation, and to use cross-validation and comparison
to base models to validate our findings. However, this method is not without its limitations.
The features selected may vary based on the algorithm or the set of processing steps chosen.
Additionally, to greatly improve performance, we removed highly collinear features—
however, these removed features could potentially still be of theoretical interest. We
performed follow-up statistical tests to assure that our final set of selected features did statisti-
cally differ between the stimulation conditions and/or predict performance; nevertheless, there
may be other connectivity features in certain bands that are also significantly modulated by
STAMPs and/or are predictive of behavior, but simply were not selected by the algorithm.
However, we believe that the beta path length relationship to behavior is robust, as it was
consistently considered highly significant by the Boruta algorithm and significantly predicted
the behavioral data, and this effect likely would not have been found had we only examined
more standard measures and frequency bands. Future studies may identify the specific role
beta network connectivity plays in episodic memory and decision-making processes.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
ACKNOWLEDGMENTS
This work was supported by the DARPA and the Army Research Office. The views, opinions
and/or findings expressed are those of the author and should not be interpreted as representing
the official views or policies of the Department of Defense or the United States Government.
SUPPORTING INFORMATION
The connectivity features, as well as metamemory sensitivity values, are included in the
Supporting Information for this article available at https://doi.org/10.1162/netn_a_00201.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
a
_
0
0
2
0
1
p
d
.
t
AUTHOR CONTRIBUTIONS
Ryan Hubbard: Conceptualization; Formal analysis; Methodology; Writing – original draft;
Writing – review & editing. Iman Zadeh: Conceptualization; Formal analysis; Methodology;
Resources; Software; Writing – original draft. Aaron Jones: Data curation; Resources. Bradley
Robert: Data curation; Resources. Natalie Bryant: Data curation; Resources; Software.
Vincent Clark: Data curation; Resources. Praveen Pilly: Conceptualization; Formal Analysis;
Funding acquisition; Investigation; Project administration; Supervision; Writing – original draft;
Writing - review & editing.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
FUNDING INFORMATION
Praveen Pilly, Defense Advanced Research Projects Agency (https://dx.doi.org/10.13039
/100000185), Award ID: W911NF-16-C-0018.
CONFLICT OF INTEREST STATEMENT
The last author, Praveen K. Pilly, has a patent on using transcranial electrical stimulation for
targeted memory reactivation (US Patent No. 10,307,592). Vincent P. Clark is a scientific
advisor with NeuroGeneces, LLC. All the authors declare that the research was conducted
in the absence of any other competing financial or nonfinancial interests.
Network Neuroscience
751
Connectivity changes by tES during sleep predict meta-memory
REFERENCES
Anderson, K. L., Rajagovindan, R., Ghacibeh, G. A., Meador, K. J.,
& Ding, M. (2009). Theta oscillations mediate interaction be-
tween prefrontal cortex and medial temporal lobe in human
memory. Cerebral Cortex, 20(7), 1604–1612. https://doi.org/10
.1093/cercor/bhp223, PubMed: 19861635
Antony, J. W., Gobel, E. W., O’Hare, J. K., Reber, P. J. & Paller,
K. A. (2012). Cued memory reactivation during sleep influences
skill learning. Nature Neuroscience, 15, 1114–1116. https://doi
.org/10.1038/nn.3152, PubMed: 22751035
Ayoub, A., Aumann, D., Hörschelmann, A., Kouchekmanesch, A.,
Paul, P., Born, J., & Marshall, L. (2013). Differential effects on fast
and slow spindle activity, and the sleep slow oscillation in humans
with carbamazepine and flunarizine to antagonize voltage-
dependent Na+ and Ca2+ channel activity. Sleep, 36(6),
905–911. https://doi.org/10.5665/sleep.2722, PubMed: 23729934
Backus, A. R., Schoffelen, J. M., Szebényi, S., Hanslmayr, S., &
Doeller, C. F. (2016). Hippocampal-prefrontal theta oscillations
support memory integration. Current Biology, 26(4), 450–457.
https://doi.org/10.1016/j.cub.2015.12.048, PubMed: 26832442
Baird, B., Smallwood, J., Gorgolewski, K. J., & Margulies, D. S.
(2013). Medial and lateral networks in anterior prefrontal cortex
support metacognitive ability for memory and perception.
Journal of Neuroscience, 33(42), 16657–16665. https://doi.org
/10.1523/JNEUROSCI.0786-13.2013, PubMed: 24133268
Barahona, M., & Pecora, L. M. (2002). Synchronization in small-
world systems. Physical Review Letters, 89(5), 054101. https://
doi.org/10.1103/PhysRevLett.89.054101, PubMed: 12144443
Bassett, D. S., & Bullmore, E. D. (2006). Small-world brain net-
works. The Neuroscientist, 12(6), 512–523. https://doi.org/10
.1177/1073858406293182, PubMed: 17079517
Benchenane, K., Peyrache, A., Khamassi, M., Tierney, P. L., Gioanni,
Y., Battaglia, F. P., & Wiener, S. I. (2010). Coherent theta oscillations
and reorganization of spike timing in the hippocampal-prefrontal
network upon learning. Neuron, 66(6), 921–936. https://doi.org
/10.1016/j.neuron.2010.05.013, PubMed: 20620877
Benjamini, Y., & Hochberg, Y. (1995). Controlling the false discovery
rate: A practical and powerful approach to multiple testing. Journal
of the Royal Statistical Society, 57, 289–300. https://doi.org/10
.1111/j.2517-6161.1995.tb02031.x
Berens, P. (2009). CircStat: A MATLAB toolbox for circular statis-
tics. Journal of Statistical Software, 31(10), 1–21. https://doi.org
/10.18637/jss.v031.i10
Bowyer, S. M. (2016). Coherence a measure of the brain networks:
past and present. Neuropsychiatric Electrophysiology, 2(1), 1.
https://doi.org/10.1186/s40810-015-0015-7
Bullmore, E., & Sporns, O. (2009). Complex brain networks: graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10(3), 186. https://doi.org/10.1038
/nrn2575, PubMed: 19190637
Burke, J. F., Zaghloul, K. A., Jacobs, J., Williams, R. B., Sperling,
M. R., Sharan, A. D., & Kahana, M. J. (2013). Synchronous and
asynchronous theta and gamma activity during episodic memory
formation. Journal of Neuroscience, 33(1), 292–304. https://doi
.org/10.1523/JNEUROSCI.2057-12.2013, PubMed: 23283342
Carpenter, J., & Bithell, J. (2000). Bootstrap confidence intervals:
when, which, what? A practical guide for medical statisticians.
Statistics in Medicine, 19(9), 1141–1164. https://doi.org/10
.1002/(SICI)1097-0258(20000515)19:9<1141::AID-SIM479>3.0
.CO;2-F, PubMed: 10797513
Carr, M.. F., Jadhav, S. P., & Frank, L. M.. (2011). Hippocampal replay
in the awake state: A potential substrate for memory consolidation
and retrieval. Neurosciences naturelles, 14(2), 147. https://est ce que je.org/10
.1038/nn.2732, PubMed: 21270783
Casanova, M.. F., El-Baz, UN., & Suri, J.. S. (Éd.). (2017). Autism imaging
and devices. CRC Press. https://doi.org/10.1201/9781315371375
Chua, E. F., & Ahmed, R.. (2016). Electrical stimulation of the dor-
solateral prefrontal cortex improves memory monitoring.
Neuropsychologie, 85, 74–79. https://est ce que je.org/10.1016/j
.neuropsychologia.2016.03.008, PubMed: 26970142
Chua, E. F., Ahmed, R., & Garcia, S. M.. (2017). Effects of HD-tDCS
on memory and metamemory for general knowledge questions
that vary by difficulty. Brain Stimulation, 10(2), 231–241.
https://doi.org/10.1016/j.brs.2016.10.013, PubMed: 27876306
Classen, J., Gerloff, C., Honda, M., & Hallett, M.. (1998). Integrative
visuomotor behavior is associated with interregionally coherent
oscillations in the human brain. Journal de neurophysiologie, 79(3),
1567–1573. https://doi.org/10.1152/jn.1998.79.3.1567, PubMed:
9497432
Compte, UN., Reig, R., Descalzo, V. F., Harvey, M.. UN., Puccini, G. D.,
& Sanchez-Vives, M.. V. (2008). Spontaneous high-frequency
(10–80 Hz) oscillations during up states in the cerebral cortex
in vitro. Journal des neurosciences, 28(51), 13828–13844. https://
doi.org/10.1523/JNEUROSCI.2684-08.2008, PubMed: 19091973
Cox, R., Korjoukov, JE., de Boer, M., & Talamini, L. M.. (2014).
Sound asleep: Processing and retention of slow oscillation
phase-targeted stimuli. PLoS One, 9(7), e101567. https://doi.org
/10.1371/journal.pone.0101567, PubMed: 24999803
Cox, R., Schapiro, UN. C., Manoach, D. S., & Stickgold, R.. (2017).
Individual differences in frequency and topography of slow and
fast sleep spindles. Frontiers in Human Neuroscience, 11, 433.
https://doi.org/10.3389/fnhum.2017.00433, PubMed: 28928647
Dallenbach, K. M.. (1913). The relation of memory error to time in-
terval. Psychological Review, 20(4), 323–337. https://est ce que je.org/10
.1037/h0076103
Daurat, UN., Huet, N., & Tiberge, M.. (2010). Metamemory beliefs
and episodic memory in obstructive sleep apnea syndrome.
Psychological Reports, 107(1), 289–302. https://doi.org/10.2466
/10.13.20.22.PR0.107.4.289-302, PubMed: 20923074
De Gennaro, L., & Ferrara, M.. (2003). Sleep spindles: An overview.
Sleep Medicine Reviews, 7(5), 423. https://doi.org/10.1053/smrv
.2002.0252
Degenhardt, F., Seifert, S., & Szymczak, S. (2017). Evaluation of
variable selection methods for random forests and omics data
sets. Briefings in Bioinformatics, 20, 492–503. https://est ce que je.org/10
.1093/bib/bbx124
Del Cul, UN., Dehaene, S., Reyes, P., Bravo, E., & Slachevsky, UN.
(2009). Causal role of prefrontal cortex in the threshold for access
to consciousness. Cerveau, 132(9), 2531–2540. https://est ce que je.org/10
.1093/brain/awp111, PubMed: 19433438
Delorme, UN., & Makeig, S. (2004). EEGLAB: An open source toolbox
for analysis of single-trial EEG dynamics including independent
component analysis. Journal of Neuroscience Methods, 134(1),
Neurosciences en réseau
752
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
9–21. https://doi.org/10.1016/j.jneumeth.2003.10.009, PubMed:
15102499
Eschenko, O., Magri, C., Panzeri, S., & Sarah, S. J.. (2012). Noradrenergic
neurons of the locus coeruleus are phase locked to cortical up-down
states during sleep. Cortex cérébral, 22(2), 426–435. https://doi.org
/10.1093/cercor/bhr121, PubMed: 21670101
Euston, D. R., Tatsuno, M., & McNaughton, B. L. (2007). Fast-
forward playback of recent memory sequences in prefrontal cortex
during sleep. Science, 318(5853), 1147–1150. https://est ce que je.org/10
.1126/science.1148979, PubMed: 18006749
Fallani, F. D. V., Richiardi, J., Chavez, M., & Achard, S. (2014).
Graph analysis of functional brain networks: Practical issues in
translational neuroscience. Philosophical Transactions of the
Royal Society of London B, 369(1653), 20130521. https://est ce que je
.org/10.1098/rstb.2013.0521, PubMed: 25180301
Fawcett, T. (2006). An introduction to ROC analysis. Pattern
Recognition Letters, 27(8), 861–874. https://est ce que je.org/10.1016/j
.patrec.2005.10.010
Fell, J., Klaver, P., Elfadil, H., Schaller, C., Élan, C. E., & Fernández,
G. (2003). Rhinal–hippocampal theta coherence during declara-
tive memory formation: Interaction with gamma synchronization?
European Journal of Neuroscience, 17(5), 1082–1088. https://est ce que je
.org/10.1046/j.1460-9568.2003.02522.x, PubMed: 12653984
Fleming, S. M., & Lau, H. C. (2014). How to measure metacogni-
tion. Frontiers in Neuroscience, 8, 443. https://doi.org/10.3389
/fnhum.2014.00443, PubMed: 25076880
Galvin, S. J., Podd, J.. V., Drga, V., & Whitmore, J.. (2003). Type 2
tasks in the theory of signal detectability: Discrimination between
correct and incorrect decisions. Psychonomic Bulletin and
Review, 10(4), 843–876. https://doi.org/10.3758/BF03196546,
PubMed: 15000533
Hanslmayr, S., & Staudigl, T. (2014). How brain oscillations form
memories—A processing based perspective on oscillatory subse-
quent memory effects. NeuroImage, 85, 648–655. https://doi.org
/10.1016/j.neuroimage.2013.05.121, PubMed: 23769913
Hanslmayr, S., Staudigl, T., & Fellner, M.. C. (2012). Oscillatory
power decreases and long-term memory: The information via de-
synchronization hypothesis. Frontiers in Human Neuroscience,
6, 74. https://doi.org/10.3389/fnhum.2012.00074, PubMed:
22514527
Hasselmo, M.. E., Bodelón, C., & Wyble, B. P.. (2002). A proposed
function for hippocampal theta rhythm: separate phases of en-
coding and retrieval enhance reversal of prior learning. Neural
Computation, 14(4), 793–817. https://est ce que je.org/10.1162
/089976602317318965, PubMed: 11936962
Hebart, M.. N., Schriever, Y., Donner, T. H., & Haynes, J.. D. (2016).
The relationship between perceptual decision variables and con-
fidence in the human brain. Cortex cérébral, 26(1), 118–130.
https://doi.org/10.1093/cercor/bhu181, PubMed: 25112281
Holz, J., Piosczyk, H., Feige, B., Spiegelhalder, K., Baglioni, C.,
Riemann, D., & Nissen, C. (2012). EEG sigma and slow-wave ac-
tivity during NREM sleep correlate with overnight declarative
and procedural memory consolidation. Journal of Sleep
Research, 21(6), 612–619. https://doi.org/10.1111/j.1365-2869
.2012.01017.X, PubMed: 22591117
Horvath, J.. C., Forte, J.. D., & Carter, Ô. (2015). Quantitative review
finds no evidence of cognitive effects in healthy populations from
single-session transcranial direct current stimulation (tDCS).
Brain Stimulation, 8(3), 535–550. https://doi.org/10.1016/j.brs
.2015.01.400, PubMed: 25701175
Huang, Y., Liu, UN. UN., Lafon, B., Friedman, D., Dayan, M., Wang, X.,
… Parra, L. C. (2017). Measurements and models of electric fields
in the in vivo human brain during transcranial electric stimulation.
Elife, 6, e18834. https://doi.org/10.7554/eLife.18834, PubMed:
28169833
Ji, D., & Wilson, M.. UN. (2007). Coordinated memory replay in the
visual cortex and hippocampus during sleep. Neurosciences naturelles,
10(1), 100–107. https://doi.org/10.1038/nn1825, PubMed:
17173043
Ketz, N., Jones, A.P., Bryant, N.B., Clark, V.P., & Pilly, P.K. (2018).
Closed-loop slow-wave tACS improves sleep-dependent long-
term memory generalization by modulating endogenous oscilla-
tion. Journal des neurosciences, 38(33), 7314–7326. https://est ce que je
.org/10.1523/JNEUROSCI.0273-18.2018, PubMed: 30037830
Kilner, J.. M., Paulignan, Y., & Boussaoud, D. (2004). Functional
connectivity during real vs. imagined visuomotor tasks: An EEG
étude. Neuroreport, 15(4), 637–642. https://doi.org/10.1097
/00001756-200403220-00013, PubMed: 15094467
Koriat, UN., & Goldsmith, M.. (1996). Monitoring and control processes
in the strategic regulation of memory accuracy. Psychological
Review, 103(3), 490. https://doi.org/10.1037/0033-295X.103.3
.490, PubMed: 8759045
Krause, M.. R., Zanos, T. P., Csorba, B. UN., Pilly, P.. K., Choe, J., Phillips,
M.. E., Datta, UN., & Pack, C. C. (2017). Transcranial direct current
stimulation facilitates associative learning and alters functional con-
nectivity in the primate brain. Biologie actuelle, 27, 3086–3096.
https://doi.org/10.1016/j.cub.2017.09.020, PubMed: 29033331
Kursa, M.. B. (2017). Efficient all relevant feature selection with ran-
dom ferns. In International Symposium on Methodologies for
Intelligent Systems (pp. 302–311). Cham: Springer. https://est ce que je
.org/10.1007/978-3-319-60438-1_30
Kursa, M.. B., & Rudnicki, W. R.. (2010). Feature selection with the
Boruta package. Journal of Statistical Software, 36(11), 1–13.
https://doi.org/10.18637/jss.v036.i11
Ladenbauer, J., Külzow, N., Passmann, S., Antonenko, D., Grittner,
U., Tamm, S., & Flöel, UN. (2016). Brain stimulation during an
afternoon nap boosts slow oscillatory activity and memory con-
solidation in older adults. NeuroImage, 142, 311–323. https://est ce que je
.org/10.1016/j.neuroimage.2016.06.057, PubMed: 27381076
Ladenbauer, J., Ladenbauer, J., Külzow, N., de Boor, R., Avramova,
E., Grittner, U., & Flöel, UN. (2017). Promoting sleep oscillations
and their functional coupling by transcranial stimulation en-
hances memory consolidation in mild cognitive impairment.
Journal des neurosciences, 37(30), 7111–7124. https://est ce que je.org/10
.1523/JNEUROSCI.0260-17.2017, PubMed: 28637840
Lafon, B., Henin, S., Huang, Y., Friedman, D., Melloni, L., Thesen, T.,
… Liu, UN. (2017). Low frequency transcranial electrical stimulation
does not entrain sleep rhythms measured by human intracranial
recordings. Communications naturelles, 8(1), 1199. https://doi.org
/10.1038/s41467-017-01045-x, PubMed: 29084960
Lee, UN. K., & Wilson, M.. UN. (2002). Memory of sequential experience
in the hippocampus during slow wave sleep. Neurone, 36(6),
1183–1194. https://doi.org/10.1016/S0896-6273(02)01096-6,
PubMed: 12495631
Lega, B. C., Jacobs, J., & Kahana, M.. (2012). Human hippocampal
theta oscillations and the formation of episodic memories.
Neurosciences en réseau
753
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
Hippocampus, 22(4), 748–761. https://doi.org/10.1002/ hipo
.20937, PubMed: 21538660
Lin, J.. J., Rugg, M.. D., Le, S., Stein, J., Rizzuto, D. S., Kahana, M.. J.,
& Lega, B. C. (2017). Theta band power increases in the posterior
hippocampus predict successful episodic memory encoding in
humans. Hippocampus, 27(10), 1040–1053. https://est ce que je.org/10
.1002/hipo.22751, PubMed: 28608960
Lisman, J.. E., & Jensen, Ô. (2013). The theta-gamma neural code.
Neurone, 77(6), 1002–1016. https://est ce que je.org/10.1016/j.neuron
.2013.03.007, PubMed: 23522038
Lustenberger, C., Boyle, M.. R., Alagapan, S., Mellin, J.. M., Vaughn, B. V.,
& Fröhlich, F. (2016). Feedback-controlled transcranial alternating
current stimulation reveals a functional role of sleep spindles in motor
memory consolidation. Biologie actuelle, 26(16), 2127–2136.
https://doi.org/10.1016/j.cub.2016.06.044, PubMed: 27476602
Luus, C. UN., & Wells, G. L. (1994). The malleability of eyewitness
confidence: Co-witness and perseverance effects. Journal de
Applied Psychology, 79(5), 714–723. https://doi.org/10.1037
/0021-9010.79.5.714
Maréchal, L., Helgadóttir, H., Mölle, M., & Born, J.. (2006). Boosting
slow oscillations during sleep potentiates memory. Nature,
444(7119), 610–613. https://doi.org/10.1038/nature05278,
PubMed: 17086200
Maréchal, L., Mölle, M., Hallschmid, M., & Born, J.. (2004).
Transcranial direct current stimulation during sleep improves
declarative memory. Journal des neurosciences, 24(44), 9985–9992.
https://doi.org/10.1523/ JNEUROSCI.2725-04.2004, PubMed:
15525784
Mason, S. J., & Graham, N. E. (2002). Areas beneath the relative
operating characteristics (ROC) and relative operating levels
(ROL) curves: Statistical significance and interpretation.
Quarterly Journal of the Royal Meteorological Society, 128(584),
2145–2166. https://doi.org/10.1256/003590002320603584
Masuda, N., & Aihara, K. (2004). Global and local synchrony of
coupled neurons in small-world networks. Biological Cybernetics,
90(4), 302–309. https://doi.org/10.1007/s00422-004-0471-9,
PubMed: 15085349
McClelland, J.. L., McNaughton, B. L., & O'Reilly, R.. C. (1995).
Why there are complementary learning systems in the hippo-
campus and neocortex: insights from the successes and failures
of connectionist models of learning and memory. Psychological
Review, 102(3), 419–457. https://doi.org/10.1037/0033-295X
.102.3.419, PubMed: 7624455
McGaugh, J.. L. (2000). Memory—A century of consolidation.
Science, 287(5451), 248–251. https://doi.org/10.1126/science
.287.5451.248, PubMed: 10634773
Memon, UN., Hope, L., & Bull, R.. (2003). Exposure duration: Effects on
eyewitness accuracy and confidence. British Journal of Psychology,
94(3), 339–354. https://doi.org/10.1348/000712603767876262
Mohammad-Rezazadeh, JE., Frohlich, J., Loo, S. K., & Jeste, S. S.
(2016). Brain connectivity in autism spectrum disorder. Actuel
Opinion in Neurology, 29(2), 137–147. https://doi.org/10.1097
/ WCO.0000000000000301, PubMed: 26910484
Molenberghs, P., Trautwein, F. M., Böckler, UN., Chanteur, T., &
Kanske, P.. (2016). Neural correlates of metacognitive ability and
of feeling confident: A large-scale fMRI study. Social Cognitive and
Affective Neuroscience, 11(12), 1942–1951. https://est ce que je.org/10
.1093/scan/nsw093, PubMed: 27445213
Mölle, M., & Born, J.. (2011). Slow oscillations orchestrating fast
oscillations and memory consolidation. Progress in Brain Research,
193, 93–110. https://doi.org/10.1016/B978-0-444-53839-0.00007-7,
PubMed: 21854958
Mölle, M., Maréchal, L., Gais, S., & Born, J.. (2004). Learning increases
human electroencephalographic coherence during subsequent
slow sleep oscillations. Proceedings of the National Academy of
Sciences of the United States of America, 101(38), 13963–13968.
https://doi.org/10.1073/pnas.0402820101, PubMed: 15356341
Mölle, M., Yeshenko, O., Maréchal, L., Sarah, S. J., & Born, J.. (2006).
Hippocampal sharp wave-ripples linked to slow oscillations in
rat slow-wave sleep. Journal de neurophysiologie, 96(1), 62–70.
https://est ce que je.org/10.1152/jn.00014.2006, PubMed: 16611848
Morales, J., Lau, H., & Fleming, S. M.. (2018). Domain-general and
domain-specific patterns of activity supporting metacognition in
human prefrontal cortex. Journal des neurosciences, 38(14),
3534–3546. https://doi.org/10.1523/JNEUROSCI.2360-17.2018,
PubMed: 29519851
Nádasdy, Z., Hirase, H., Czurkó, UN., Csicsvari, J., & Buzsáki, G.
(1999). Replay and time compression of recurring spike
sequences in the hippocampus. Journal des neurosciences, 19(21),
9497–9507. https://doi.org/10.1523/ JNEUROSCI.19-21-09497
.1999, PubMed: 10531452
Nolte, G., Bai, O., Wheaton, L., Mari, Z., Vorbach, S., & Hallett, M..
(2004). Identifying true brain interaction from EEG data using the
imaginary part of coherency. Neurophysiologie clinique, 115(10),
2292–2307. https://doi.org/10.1016/j.clinph.2004.04.029,
PubMed: 15351371
Nunez, P.. L. (1995). Neuromodulation of neocortical dynamics. Dans
P.. L. Nunez (Éd.), Neocortical dynamics and human EEG
rhythms (pp. 591–627). New York: Presse universitaire d'Oxford.
Nyhus, E., & Curran, T. (2010). Functional role of gamma and theta
oscillations in episodic memory. Neuroscience and Biobehavioral
Reviews, 34(7), 1023–1035. https://doi.org/10.1016/j.neubiorev
.2009.12.014, PubMed: 20060015
Ong, J.. L., Lo, J.. C., Chee, N. JE., Santostasi, G., Paller, K. UN., Zee,
P.. C., & Chee, M.. W. (2016). Effects of phase-locked acoustic
stimulation during a nap on EEG spectra and declarative memory
consolidation. Sleep Medicine, 20, 88–97. https://est ce que je.org/10
.1016/j.sleep.2015.10.016, PubMed: 27318231
Opitz, UN., Falchier, UN., Yan, C. G., Yeagle, E. M., Linn, G. S.,
Megevand, P., … Schroeder, C. E. (2016). Spatiotemporal structure
of intracranial electric fields induced by transcranial electric stimu-
lation in humans and nonhuman primates. Rapports scientifiques, 6,
31236. https://doi.org/10.1038/srep31236, PubMed: 27535462
Oudiette, D., & Paller, K. UN. (2013). Upgrading the sleeping brain
with targeted memory reactivation. Tendances des sciences cognitives,
17(3), 142–149. https://doi.org/10.1016/j.tics.2013.01.006,
PubMed: 23433937
Piantoni, G., Van Der Werf, Oui. D., Jensen, O., & Van Someren, E. J..
(2015). Memory traces of long-range coordinated oscillations in
the sleeping human brain. Human Brain Mapping, 36(1), 67–84.
https://doi.org/10.1002/hbm.22613, PubMed: 25139521
Pilly, P.. K., Skorheim, S. W., Hubbard, R.. J., Jones, UN. P., Bryant,
N. B., Robert B, Roach, S. M., Hartholt, UN., Lerner, JE., Ketz, N.,
Choe, J., Rizzo, S., Clark, V. P., & Howard, M.. D. (2020). Un-
shot tagging during wake and cueing during sleep with spatio-
temporal patterns of transcranial electrical stimulation can boost
Neurosciences en réseau
754
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
long-term metamemory of individual episodes in humans.
Frontiers in Neuroscience, 13, 1416. https://doi.org/10.3389
/fnins.2019.01416, PubMed: 31998067
Polanía, R., Nitsche, M.. UN., & Paulus, W. (2011). Modulating func-
tional connectivity patterns and topological functional organiza-
tion of the human brain with transcranial direct current
stimulation. Human Brain Mapping, 32(8), 1236–1249. https://
doi.org/10.1002/hbm.21104, PubMed: 20607750
Polanía, R., Paulus, W., & Nitsche, M.. UN. (2012). Modulating
cortico-striatal and thalamo-cortical functional connectivity with
transcranial direct current stimulation. Human Brain Mapping,
33(10), 2499–2508. https://doi.org/10.1002/ hbm.21380,
PubMed: 21922602
Rasch, B., & Born, J.. (2013). About sleep’s role in memory.
Physiological Reviews, 93(2), 681–766. https://doi.org/10.1152
/physrev.00032.2012, PubMed: 23589831
Rasch, B., Büchel, C., Gais, S., & Born, J.. (2007). Odor cues during
slow-wave sleep prompt declarative memory consolidation.
Science, 315, 1426–1429. https://doi.org/10.1126/science
.1138581, PubMed: 17347444
Reijneveld, J.. C., Ponten, S. C., Berendse, H. W., & Stam, C. J..
(2007). The application of graph theoretical analysis to complex
networks in the brain. Neurophysiologie clinique, 118(11),
2317–2331. https://doi.org/10.1016/j.clinph.2007.08.010,
PubMed: 17900977
Ren, Y., Nguyen, V. T., Sonkusare, S., Lv, J., Pang, T., Guo, L., …
Guo, C. C. (2018). Effective connectivity of the anterior hippo-
campus predicts recollection confidence during natural memory
retrieval. Communications naturelles, 9(1), 1-dix. https://est ce que je.org/10
.1038/s41467-018-07325-4, PubMed: 30451864
Rounis, E., Maniscalco, B., Rothwell, J.. C., Passingham, R.. E., &
Lau, H. (2010). Theta-burst transcranial magnetic stimulation to
the prefrontal cortex impairs metacognitive visual awareness.
Neurosciences cognitives, 1(3), 165–175. https://est ce que je.org/10.1080
/17588921003632529, PubMed: 24168333
Rudoy, J.. D., Voss, J.. L., Westerberg, C. E. & Paller, K. UN. (2009).
Strengthening individual memories by reactivating them during
dormir. Science, 326, 1079. https://doi.org/10.1126/science
.1179013, PubMed: 19965421
Ryals, UN. J., Rogers, L. M., Gross, E. Z., Polnaszek, K. L., & Voss, J.. L.
(2016). Associative recognition memory awareness improved by
theta-burst stimulation of frontopolar cortex. Cortex cérébral,
26(3), 1200–1210. https://doi.org/10.1093/cercor/ bhu311,
PubMed: 25577574
Sadaghiani, S., Poline, J.. B., Kleinschmidt, UN., & D'Esposito, M..
(2015). Ongoing dynamics in large-scale functional connectivity
predict perception. Proceedings of the National Academy of
Sciences of the United States of America, 112(27), 8463–8468.
https://doi.org/10.1073/pnas.1420687112, PubMed: 26106164
Schabus, M., Dang-Vu, T. T., Albouy, G., Balteau, E., Boly, M.,
Carrier, J., … Phillips, C. (2007). Hemodynamic cerebral corre-
lates of sleep spindles during human non-rapid eye movement
dormir. Proceedings of the National Academy of Sciences of the
United States of America, 104(32), 13164–13169. https://doi.org
.org/10.1073/pnas.0703084104, PubMed: 17670944
Scheinost, D., Noble, S., Horien, C., Vert, UN. S., Lake, E. M.,
Salehi, M., … Constable, R.. T. (2019). Ten simple rules for pre-
dictive modeling of individual differences in neuroimaging.
NeuroImage, 193, 35–45. https://doi.org/10.1016/j.neuroimage
.2019.02.057, PubMed: 30831310
Schreiner, T., Lehmann, M., & Rasch, B. (2015). Auditory feedback
blocks memory benefits of cueing during sleep. Nature
Communications, 6, 8729. https://doi.org/10.1038/ncomms9729,
PubMed: 26507814
Schreiner, T., & Rasch, B. (2014). Boosting vocabulary learning by
verbal cueing during sleep. Cortex cérébral, 25(11), 4169–4179.
https://doi.org/10.1093/cercor/bhu139, PubMed: 24962994
Shapira, UN. UN., & Pansky, UN. (2019). Cognitive and metacognitive
determinants of eyewitness memory accuracy over time.
Metacognition and Learning, 14(3), 437–461. https://est ce que je.org/10
.1007/s11409-019-09206-7
Sirota, UN., Csicsvari, J., Buhl, D., & Buzsáki, G. (2003).
Communication between neocortex and hippocampus during
sleep in rodents. Proceedings of the National Academy of
Sciences of the United States of America, 100(4), 2065–2069.
https://doi.org/10.1073/pnas.0437938100, PubMed: 12576550
Skaggs, W. E., & McNaughton, B. L. (1996). Replay of neuronal
firing sequences in rat hippocampus during sleep following spa-
tial experience. Science, 271(5257), 1870–1873. https://doi.org
/10.1126/science.271.5257.1870, PubMed: 8596957
Song, J., Birn, R.. M., Boly, M., Meier, T. B., Naïr, V. UN., Meyerand,
M.. E., & Prabhakaran, V. (2014). Age-related reorganizational
changes in modularity and functional connectivity of human
brain networks. Brain Connectivity, 4(9), 662–676. https://est ce que je
.org/10.1089/brain.2014.0286, PubMed: 25183440
Sporer, S. L., Penrod, S., Read, D., & Cutler, B. (1995). Choosing, con-
fidence, and accuracy: A meta-analysis of the confidence-accuracy
relation in eyewitness identification studies. Psychological Bulletin,
118(3), 315–327. https://doi.org/10.1037/0033-2909.118.3.315
Sporns, Ô. (2003). Graph theory methods for the analysis of neural
connectivity patterns. In Neuroscience databases (pp. 171–185).
Boston, MA: Springer. https://doi.org/10.1007/978-1-4615-1079
-6_12
Stam, C. J., Jones, B. F., Nolte, G., Breakspear, M., & Scheltens, P..
(2006). Small-world networks and functional connectivity in
Alzheimer’s disease. Cortex cérébral, 17(1), 92–99. https://est ce que je
.org/10.1093/cercor/bhj127, PubMed: 16452642
Staresina, B. P., Bergmann, T. O., Bonnefond, M., Van Der Meij, R.,
Jensen, O., Tremper, L., … Fell, J.. (2015). Hierarchical nesting of
slow oscillations, spindles and ripples in the human hippocam-
pus during sleep. Neurosciences naturelles, 18(11), 1679–1686.
https://doi.org/10.1038/nn.4119, PubMed: 26389842
Steriade, M.. (2006). Grouping of brain rhythms in corticothalamic
systèmes. Neurosciences, 137(4), 1087–1106. https://est ce que je.org/10
.1016/j.neuroscience.2005.10.029, PubMed: 16343791
Steriade, M., Amzica, F., & Contreras, D. (1996). Synchronization
of fast (30–40 Hz) spontaneous cortical rhythms during brain
activation. Journal des neurosciences, 16(1), 392–417. https://
doi.org/10.1523/ JNEUROSCI.16-01-00392.1996, PubMed:
8613806
Van Der Helm, E., Gujar, N., Nishida, M., & Walker, M.. P.. (2011).
Sleep-dependent facilitation of episodic memory details. PloS
Un, 6(11), e27421. https://doi.org/10.1371/journal.pone
.0027421, PubMed: 22114672
Varoquaux, G., Raamana, P.. R., Engemann, D. UN., Hoyos-Idrobo, UN.,
Schwartz, Y., & Thirion, B. (2017). Assessing and tuning brain
Neurosciences en réseau
755
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Connectivity changes by tES during sleep predict meta-memory
decoders: Cross-validation, caveats, and guidelines. NeuroImage,
145, 166–179. https://doi.org/10.1016/j.neuroimage.2016.10
.038, PubMed: 27989847
Vöröslakos, M., Takeuchi, Y., Brinyiczki, K., Zombori, T., Oliva, UN.,
Fernández-Ruiz, UN., … Berényi, UN. (2018). Direct effects of trans-
cranial electric stimulation on brain circuits in rats and humans.
Communications naturelles, 9(1), 483. https://est ce que je.org/10.1038
/s41467-018-02928-3, PubMed: 29396478
Walker, M.. P., & Stickgold, R.. (2004). Sleep-dependent learning
and memory consolidation. Neurone, 44(1), 121–133. https://est ce que je
.org/10.1016/j.neuron.2004.08.031, PubMed: 15450165
Watts, D. J., & Strogatz, S. H. (1998). Collective dynamics of ‘small-
world’ networks. Nature, 393(6684), 440–442. https://est ce que je.org/10
.1038/30918, PubMed: 9623998
Westerberg, C. E., Florczak, S. M., Weintraub, S., Mesulam, M.. M.,
Maréchal, L., Zee, P.. C., & Paller, K. UN. (2015). Mémoire
improvement via slow-oscillatory stimulation during sleep in older
adultes. Neurobiology of Aging, 36(9), 2577–2586. https://doi.org
/10.1016/j.neurobiolaging.2015.05.014, PubMed: 26116933
Wilson, M.. UN., & McNaughton, B. L. (1994). Reactivation of hippo-
campal ensemble memories during sleep. Science, 265(5172),
676–679. https://doi.org/10.1126/science.8036517, PubMed:
8036517
Ye, Q., Zou, F., Dayan, M., Lau, H., Hu, Y., & Kwok, S. C. (2019).
Individual susceptibility to TMS affirms the precuneal role in
meta-memory upon recollection. Structure et fonction du cerveau,
224(7), 2407–2419. https://doi.org/10.1007/s00429-019-01909-6,
PubMed: 31254060
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
5
3
7
3
4
1
9
6
0
4
9
7
n
e
n
_
un
_
0
0
2
0
1
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neurosciences en réseau
756