RECHERCHE
Uncovering differential identifiability in
network properties of human brain
functional connectomes
Meenusree Rajapandian1, Enrico Amico1,2, Kausar Abbas1,2,
Mario Ventresca1, and Joaquín Goñi1,2,3
1School of Industrial Engineering, Purdue University, West Lafayette, IN, Etats-Unis
2Purdue Institute of Integrative Neuroscience, West Lafayette, IN, Etats-Unis
3Weldon School of Biomedical Engineering, West Lafayette, IN, Etats-Unis
un accès ouvert
journal
Mots clés: Brain connectomics, Functional connectivity, Fingerprint, Network science, Subject
identifiability
ABSTRAIT
The identifiability framework (If ) has been shown to improve differential identifiability
(reliability across-sessions and -sites, and differentiability across-subjects) of functional
connectomes for a variety of fMRI tasks. But having a robust single session/subject functional
connectome is just the starting point to subsequently assess network properties for
characterizing properties of integration, segregation, and communicability, entre autres.
Naturellement, one wonders whether uncovering identifiability at the connectome level also
uncovers identifiability on the derived network properties. This also raises the question of
where to apply the If framework: on the connectivity data or directly on each network
measurement? Our work answers these questions by exploring the differential identifiability
profiles of network measures when If is applied (un) on the functional connectomes, et (b)
directly on derived network measurements.
Results show that improving across-session reliability of functional connectomes (FCs) aussi
improves reliability of derived network measures. We also find that, for specific network
properties, application of If directly on network properties is more effective. Enfin, nous
discover that applying the framework, either way, increases task sensitivity of network
properties. At a time when the neuroscientific community is focused on subject-level
inferences, this framework is able to uncover FC fingerprints, which propagate to derived
network properties.
RÉSUMÉ DE L'AUTEUR
Functional connectome (FC) fingerprinting recently became a topic of great interest in
network neuroscience. We recently proposed a framework to improve brain fingerprint (c'est à dire.,
identifiability) of FCs, which improves not only test-retest reliability but also the correlation of
FCs with fluid intelligence. Cependant, does this improvement in FC fingerprints propagate to
the derived network measures?
In this work we found that improving the fingerprint (differential identifiability) of the
functional connectome also improves the “fingerprint” of its network properties.
En outre, when using the identifiability framework on the network properties directly,
certain network properties like search information and communicability add to the FC
fingerprint. Enfin, we show that enhancement of the fingerprint in the network measures, dans
a wide range of cognitive tasks, using the identifiability framework also improves task
Citation: Rajapandian, M., Amico, E.,
Abbas, K., Ventresca, M., & Goñi, J..
(2020). Uncovering differential
identifiability in network properties of
human brain functional connectomes.
Neurosciences en réseau, 4(3), 698–713.
https://doi.org/10.1162/netn_a_00140
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00140
Reçu: 21 Novembre 2019
Accepté: 30 Mars 2020
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Joaquín Goñi
jgonicor@purdue.edu
Éditeur de manipulation:
Alex Fornito
droits d'auteur: © 2020
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Uncovering identifiability in properties of functional connectomes
sensitivity in these measures. We show that regardless of whether you are using functional
connectomes or the network properties derived from them, using the If framework on the
functional connectomes would be a beneficial first step.
INTRODUCTION
The analysis of structural and functional human brain connectivity based on network science
has become prevalent for understanding the underlying mechanisms of the human brain. Us-
ing network properties, we are able to understand the topology of brain connectivity patterns
(Fornito, Zalesky, & Bullmore, 2016; Sporns, 2010, 2018), integration and segregation (Cohen
& D'Esposito, 2016; Deco, Tononi, Boly, & Kringelbach, 2015; Fukushima et al., 2018; Sporns,
2013, Sporns & Betzel, 2016), as well as communication dynamics (Avena-Koenigsberger,
Misic, & Sporns, 2018; Costa, Batista, & Ascoli, 2011; Estrada & Hatano, 2008; Petrella, 2011)
and association between human cognition and brain function (Alavash, Hilgetag, Thiel, &
Gießing, 2015; Bola & Sabel, 2015; Davison et al., 2015; Mattar, Betzel, & Bassett, 2016;
Zalesky, Fornito, & Bullmore, 2010). Until recently, many brain connectivity studies used
group-level comparisons, where data from many subjects are collapsed (par exemple., group averag-
ing) into a representative sample of clinical and healthy population (Castellanos, Di Martino,
Craddock, Mehta, & Milham, 2013; Crossley et al., 2014; Fornito, Zalesky, & Breakspear,
2015). Cependant, this comes at a price of potentially ignoring intragroup individual variability
(Seitzman et al., 2019).
Detecting individual differences in functional connectivity profiles thus becomes important,
when associating connectivity profiles with individual behavioral outcomes. Au cours des dernières années,
publicly available functional connectome (FC) datasets (Biswal et al., 2010; Van Essen et al.,
2013) with large sample sizes have enabled the scientific community to account for interindi-
vidual variability in the human functional connectome. A number of promising methods that
can successfully capture these individual differences have been established in recent times
(Gratton et al., 2018; Mars, Passingham, & Jbabdi, 2018; Satterthwaite, Xia, & Bassett, 2018;
Seitzman et al., 2019; Venkatesh, Jaja, & Personne, 2019). Par exemple, work by Finn et al.
(2015) has shown the existence of a recurrent and reproducible fingerprint in functional con-
nectomes estimated from neuroimaging data. This idea has been extended to maximize or
minimize subject-specific and/or task-specific information (Pallarés et al., 2018; Xie et al.,
2018). These subject-specific fingerprints have been used to track fluctuations in attention at
the individual level (Rosenberg et al., 2019).
The “identifiability framework” (Amico and Goñi, 2018b), based on the group-level princi-
pal component analysis of functional connectomes that maximizes differential identifiability,
has been shown to improve functional connectome fingerprints within and across sites, for a
variety of fMRI tasks, over a wide range of scanning length, and with and without global signal
regression (Amico and Goñi, 2018b; Bari, Amico, Vike, Talavage, & Goñi, 2019). En plus,
it has been shown that maximizing differential identifiability on the functional connectomes
provides more robust and reliable associations with cognition (Svaldi, Goñi, Abbas, et coll.,
2019) as well as with disease progression (Svaldi, Goñi, Sanjay, et coll., 2019). The natural next
step is to assess the impact of such a procedure on subsequent network measurements that
characterize topological and communication properties of functional brain networks.
An open question of great relevance for the brain connectomics community is how to mea-
sure and uncover subject fingerprints in network measurements of functional connectivity. Un-
covering reliable connectivity fingerprints is crucial when assessing clinical populations and
when ultimately mapping cognitive characteristics into connectivity (Scheinost et al., 2019;
699
Analyse en composantes principales (APC):
Une réduction de dimensionnalité
technique that uses an orthogonal
transformation to convert a set of
observations of (possibly) correlated
variables into a set of values of
linearly uncorrelated variables called
principal components. Tel
transformation ensures that the first
principal component has the largest
possible variance, and each
subsequent component has the
highest possible variance under the
constraint of being orthogonal to the
preceding components.
Differential identifiability:
A score that quantifies in a test/retest
dataset, on average, how much more
similar are rest/retest functional
connectomes of the same subject
with respect to functional
connectomes of different subjects.
Similarity is measured by Pearsons
correlation coefficient between every
two functional connectivity profiles.
Neurosciences en réseau
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Uncovering identifiability in properties of functional connectomes
Functional magnetic resonance
imaging (IRMf):
A noninvasive technique that
estimates brain activity by detecting
changes associated with blood flow.
The rationale of this technique relies
on the fact that there is a positive
association between cerebral blood
flow and neuronal activation.
Identifiability framework:
A framework based on principal
component analysis to decompose a
test/retest functional connectivity
dataset (or its network-derived
measurements) and subsequently
reconstruct at the optimal number of
components that maximizes the
differential identifiability score.
Functional connectome/connectivity
(FC) matrice:
A network representation of the
functional coupling between brain
régions. Such coupling is usually
measured by quantifying the
statistical dependencies between
timeseries of brain regions (par exemple.,
pairwise Pearson’s correlation,
mutual information) as obtained by
IRMf.
Graph:
An ordered pair formed by a set of
nodes and a set of edges (lequel
represent connections between pairs
of nodes). Nodes are usually
represented by circles, alors que
edges are represented by lines or arcs
connecting pairs of nodes.
Mean first passage time (MFPT):
In a connected graph, MFPT
quantifies the expected number of
steps that it takes for a random
walker to go from a source node to a
target node for the very first time.
The measurement relies on the
transition probability matrix that can
be obtained from a connected graph,
which is indeed an ergodic Markov
chain.
Node strength:
In a weighted graph (c'est à dire., où
edges have assigned numerical
valeurs), it represents the total sum of
the edge weights attached to the
node.
Neurosciences en réseau
Shen et al., 2017; Svaldi, Goñi, Sanjay, et coll., 2019). Our hypothesis is that improvement in
FC fingerprints should also “propagate” to network derived measurements. An organic way of
assessing this would be to track differential identifiability scores of derived network features as
the differential identifiability on the functional connectomes changes. One could also proceed
with the application of the identifiability framework directly on the network-derived features
as opposed to using it on FCs. The above-mentioned approaches rely on different principles
of what a fingerprint in a network-derived measurement. The first one assumes that functional
connectivity data are “holding” the fingerprints and propagating them to any network-derived
measurement. The second one considers functional connectivity data as a proxies to ultimately
estimate a network measurement with a potentially prominent subject fingerprint.
MÉTHODES
The dataset used here is composed of the 100 unrelated subjects of the Human Connectome
Project Release Q3 (Van Essen et al., 2013). Per HCP protocol, all subjects gave written in-
formed consent to the HCP consortium. Each subject consists of two fMRI resting-state runs and
seven fMRI tasks: gambling, relational, sociale, working memory, moteur, langue, and emo-
tion. Data acquisition for each subject and for each task consists of two fMRI sessions, lequel
are tagged here as test and retest. A cortical parcellation into 360 brain regions as proposed by
Glasser et al. (2013) was employed with an additional 14 subcortical regions for completeness
(Amico & Goñi, 2018un, 2018b). The HCP functional preprocessing pipeline was used (Glasser
et coll., 2013; Smith et al., 2013), followed by further processing as described in Amico, Arenas,
and Goñi (2019) and Amico and Goñi (2018b) for both resting-state and task fMRI data. Pour
each subject and fMRI session, a symmetric weighted connectivity matrix (the functional con-
nectome) was obtained by computing Pearson’s correlation coefficients between pairs of nodal
time courses. For a detailed description of all the preprocessing steps, refer to Amico and Goñi
(2018b). Enfin, before finding the below network properties, all negative correlations are set
to a small value of epsilon (MATLAB command eps, equivalent to 2.22 × 10 − 16). Please note
that we used the value of epsilon and not 0 to ensure the following two properties for all FCs
assessed: (un) FCs are connected graphs; (b) The derived Markov Chains (as obtained by the
transition probability matrices) are regular and hence permit mean first passage time (MFPT)
computation (Kemeny & Snell, 1976).
Network Properties
Graph theoretic measures have played a key role in understanding the attributes of brain net-
works in general, and of functional connectomes in particular (Fornito et al., 2016; Rubinov
& Sporns, 2010; Sporns, 2010). Here we select a set of node and node pair properties (c'est à dire.,
properties that are a function of a single node or a pair of nodes, respectivement) to assess their
fingerprinting characteristics. A functional connectome is a symmetric square correlation ma-
trix that may be seen as an undirected weighted graph. Let G = (V, W ) be an undirected
weighted graph with set of nodes V = {v1, v2, . . . , vn} and weights W = [wij], where wij is
the strength of the edge between nodes vi and vj.
1. Degree strength
The degree strength of a node (Ki) in an undirected binary graph is the number of edges
that are connected to the node. Ici, we consider the weighted sum of the edges con-
nected to the node i.
Ki =
n
∑
j=1
wij
700
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Uncovering identifiability in properties of functional connectomes
2. Shortest path length
The shortest path length (SPL) between two nodes of an undirected graph is defined as
the minimum number of edges (and thus steps) that separate the two nodes. For an
undirected weighted graph, it is the path that results in the smallest value of the sum of
the inverse of edge weights that constitute a path between a pair of nodes i and j. Pour
such a path, that consists of the following sequence of nodes, Ωi↔j = {je, X, oui, . . . , z, j}
with corresponding sequence of edge weights πi↔j = {wix, wxy, . . . , wzj}, the shortest
path length is:
SPLij = ∑
wlm∈πi↔j
1
wlm
.
Search information (SI):
An information theoretical
measurement that quantifies in bits
how hidden a shortest path is (from a
source node to a target node), comme
embedded in the graph. Ce
measurement may be applied to
binary and weighted graphs.
Note that Ωi↔j = Ωj↔i for shortest paths in any undirected graph.
3. Search information
The search information (SIij) for two nodes i and j is the information required to follow
the shortest path (Rosvall, Trusina, Minnhagen, & Sneppen, 2005); c'est, the negative
log of the product of probability of taking the correct exit at every node along the shortest
chemin. Autrement dit, it can be considered as the information required to reach node j
starting from node i. For a path between nodes i and j that has a sequence of nodes
Ωi→j = {je, X, oui, . . . , z, j}, with probability of taking the path P(πi→j) = Πl∈Ω∗
1/kl,
the search information for the path is (Goñi et al., 2014)
i→j
SIij = − log2 P(πi→j).
Note that SIij 6= SIji.
4. Mean first passage time
The MFPT is the expected (on average) number of steps a random walker takes to reach
node j (for the the first time) from node i (Kemeny & Snell, 1976). The Mean First Passage
Time (MFPT) for a pair of nodes with source i and target j is
MFPTij =
ζ jj − ζij
ϕj
where ϕ is the left eigenvector associated with eigenvalue 1, Z = [ζij] is the fundamen-
tal matrix computed as Z = (I − P + Φ)−1. Here I is the n × n identity matrix, P is
the transition matrix and Φ is an n × n matrix with each column corresponding to the
probability vector ϕ such that ∀j Φij = ϕi. Please note that MFPTij 6= MFPTji.
5. Driftness
We use a measure of communication called driftness (Costa et al., 2011), which is the
ratio of the mean first passage time and the shortest path of a pair of nodes i and j. Con-
sidering that SPij is the best possible scenario path for a random-walk, this measurement
is modulating the mean first passage times with respect to the fastest routes within the
network to go from node i to j. Ainsi, note that Wij ≥ 1.
Wij =
MFPTij
SPij
6. Communicability
Communicability between two nodes i and j is a measure of network integration com-
puted as a weighted sum of number of all possible walks between them (Estrada &
Hatano, 2008). Ici, we use a normalization method proposed to handle the dis-
proportionate influence of highly connected nodes (also known as hubs) in a graph
Neurosciences en réseau
701
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Uncovering identifiability in properties of functional connectomes
(Crofts & Higham, 2009). Note that this is frequently the case when assessing functional
connectomes.
Cij = [eD−0.5 AD−0.5
]ij
where D = diag(K) and K = [ki] where ki is the degree strength of node i, as defined
au-dessus de.
7. Clustering Coefficient
The clustering coefficient of a node is the tendency of its neighbors to form cliques. C'est
the ratio of the total number of triangles that a node forms with its neighbors to the total
number of possible triangles that can be formed.
CCi =
2ti
ki(ki − 1)
where ti = 1/2 ∑j,h∈V(wijwihwjh)1/3 is the geometric mean of triangles around node i
for weighted networks.
8. Betweenness Centrality
The betweenness centrality of a node is the fraction of all shortest paths in a network that
contain that node.
Bi =
1
(n − 1)(n − 2)
ρhj(je)
ρhj
∑
h,j∈V
h6=j,h6=i,j6=i
where ρhj(je) is the number of shortest paths between h and j that pass through i. It can
be seen as a measurement of to what extent a node “lies” between other pairs of nodes
when accounting specifically for shortest-paths.
Group-Level Principal Component Analysis and Differential Identifiability
Briefly describing the Identifiability Framework (If ) introduced in Amico and Goñi (2018b), le
functional connectomes of each subject (test and retest) are vectorized and added to a matrix,
the columns of which are the runs (test and retest) of each subject, while the rows are the
functional connectivity values of brain region pairs. The m principal components of this matrix
are then ranked by variance explained and included, in an iterative fashion, to reconstruct
the functional connectomes (Amico & Goñi, 2018b). This is done separately for each task
and rest. Following the reconstruction of the functional connectomes, we then compute the
network property of interest for each subject, on each run (test and retest). This is referred to as
NP(If {FC}) in all further sections, where NP is the network property and FC is the functional
connectome.
We also extend the framework by using this decomposition — reconstruction procedure on
the network properties. Dans ce cas, the network properties are computed on the original func-
tional connectomes for each subject and run. Each network property is then vectorized and
added to a matrix. Note that this is similar to how functional connectomes were rearranged
in the NP(If {FC}) and in Amico and Goñi (2018b). Cependant, the rows of this matrix now
consists of the network property values corresponding to a pair of brain regions in case of pair-
wise properties or a brain region when node properties are derived. The principal components
of this matrix are then extracted and iteratively reconstructed using m number of components
with the highest explained variance. Since the network properties are the ones being decom-
posed in this case, the result of the reconstruction is the corresponding network properties of
each individual and each run. This method is subsequently referred to as If {NP(FC)}).
Neurosciences en réseau
702
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Uncovering identifiability in properties of functional connectomes
We use differential identifiability (Amico, & Goñi, 2018b) to assess the individual fingerprint
of each network property. For each method described above, the network properties derived
are used to compute the identifiability matrix. Each position of the identifiability matrix i, j
denotes the correlation between the network property of subject i test and subject j retest.
Alors, along the diagonal elements, we have the correlation of a network property between
the subject test and retest called Iself . The non-diagonal elements are the correlations between
a run of a subject i and subject j where i and j are different (Iothers). The differential identifiability
is then defined as,
Idiff = (Iself − Iothers) × 100
Intraclass correlation (ICC):
An inferential statistic for quantitative
measurements that are organized into
groupes. ICC describes how strongly
units in the same group resemble
l'un l'autre. A typical application
consists of the assessment of
consistency or reproducibility of
quantitative measurements made by
different observers measuring the
same quantity.
Intraclass correlation coefficient (ICC) represents how strongly measures of a group are
in agreement with each other (Bartko, 1966; McGraw & Wong, 1996). The higher the ICC
valeur, the higher the level of agreement. We use ICC (Shrout & Fleiss, 1979) to assess the task
sensitivity of a network measure, for each brain region pair and every subject. Dans ce cas,
the members of the groups are the different runs (test and retest) of a subject; the different
groups represent the different fMRI task conditions (and rest). The mean task sensitivity is then
taken across all subjects and reported. For this assessment, the functional connectome (ou
the network property If {NP(FC)}) was optimally reconstructed, c'est, using the number of
components that gave the highest Idiff score for that task.
RÉSULTATS
The dataset used for this study consisted of fMRI scans of the 100 unrelated subjects from
the Human Connectome Project (Van Essen et al., 2013). Pour chaque sujet, we computed
18 whole-brain functional connectivity matrices: 4 corresponding to resting-state (2 sessions,
each with test and retest), et 14 corresponding to each of the seven tasks (each including
two runs; test-retest). The multimodal parcellation used here, as proposed by Glasser et al.
(2016), includes 360 cortical brain regions. For completeness, 14 subcortical regions were
added (Amico & Goñi, 2018un), hence producing functional connectome matrices (square,
symmetric) of size 374 × 374.
In this work, we study the effects of If on the identifiability profiles of network properties in
two different scenarios: (un) when applying differential identifiability on functional connectivity,
NP(If {FC}), et (b) when applying differential identifiability directly on network properties,
If {NP(FC)}.
NP(If {FC}): The functional connectomes (FCs) of each task (including rest) were vector-
ized, organized together, and then decomposed into principal components and subsequently
reconstructed by adding an increasing number of components ordered by their variance ex-
plained. After every such reconstruction, a number of network measurements (see the Methods
section for details) were computed for each FC, and Idiff was found on the derived network
properties. This is compared with the Idiff score estimated directly from the reconstructed func-
tional connectomes – If {FC}. By doing so, we extend the differential identifiability framework
to uncover fingerprints in network properties derived from functional connectomes.
For each task, we observed an optimal point of reconstruction where the differential iden-
tifiability on the FCs was maximized (voir la figure 1). This optimal point was always in the
neighborhood of half the maximum number of components (which is equal to the number of
subjects in the data) and produced Idiff values much higher than fully reconstructed data, que
est, using all the components. These results reaffirm those reported by Amico and Goñi (2018b).
We then assessed Idiff on the following node pair network properties: shortest path length (SPL),
Neurosciences en réseau
703
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Uncovering identifiability in properties of functional connectomes
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
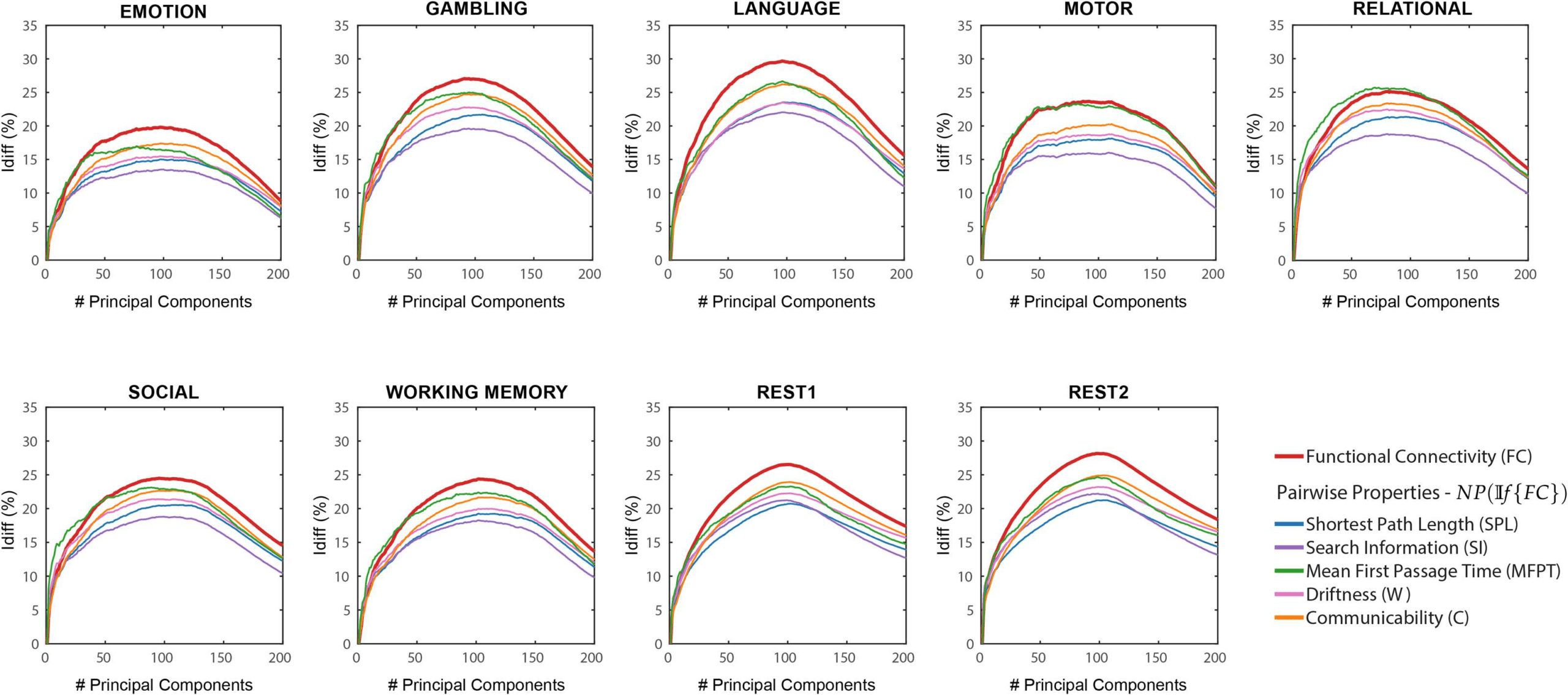
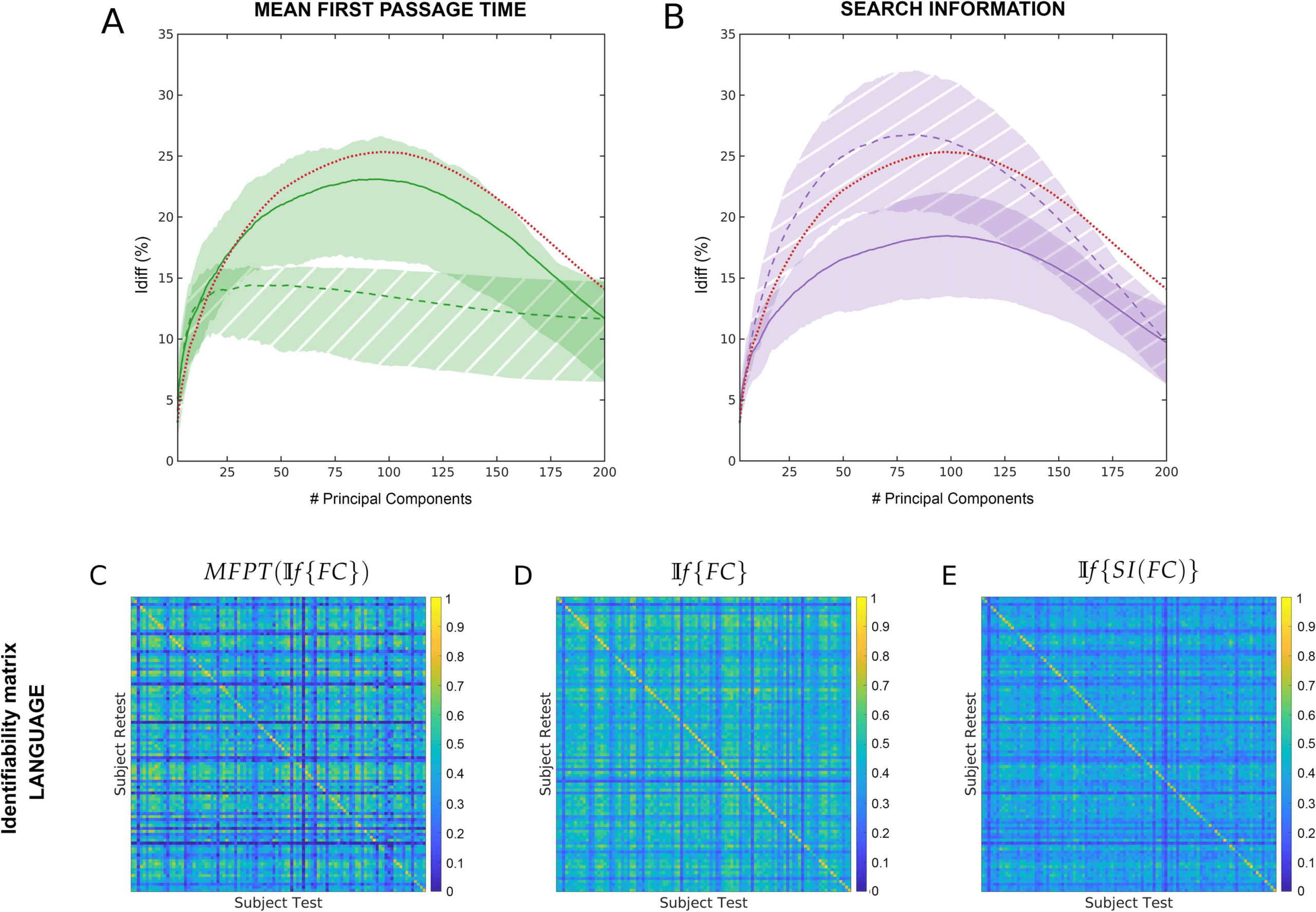
Chiffre 1. NP(If {FC}) Differential identifiability (Idiff ) profiles of pairwise properties for different fMRI tasks as a function of the number of
principal components used for reconstruction. Ici, the identifiability framework was applied on the functional connectomes (If {FC}). Chaque
plot shows, for each fMRI task, the Idiff score associated with functional connectivity (red solid line) and the Idiff scores on network properties
derived from the reconstructed functional connectomes, NP(If {FC}) (see legend) for different numbers of components.
search information (SI), mean first passage time (MFPT), driftness (W), and communicability
(C). In all cases, there was an optimal regime of number of components that maximized Idiff
(voir la figure 1). Dans l'ensemble, the Idiff score on all the network properties and functional connec-
tomes reach the peak at a similar number of principal components, ranging between 80 et
110. We can also see that the Idiff on functional connectomes is generally higher than those
on the network properties for all the tasks and for most of the number of components. Un
exception is MFPT on motor task where the Idiff scores on FC and MFPT produced very similar
results for the entire range of principal components. Another exception is MFPT on relational
task where the peak Idiff of MFPT(If {FC} is greater than that of If {FC} but the margin of
difference is really small (≈ 0.59).
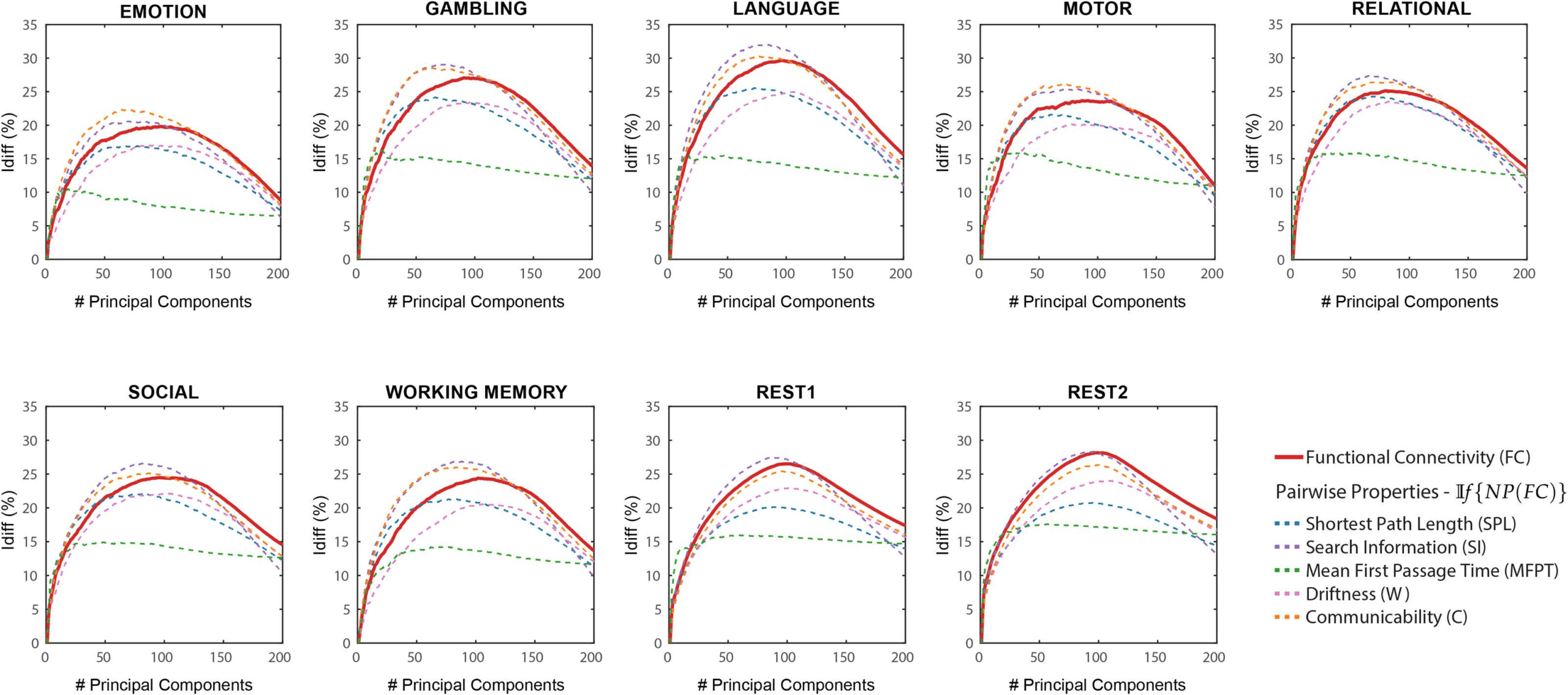
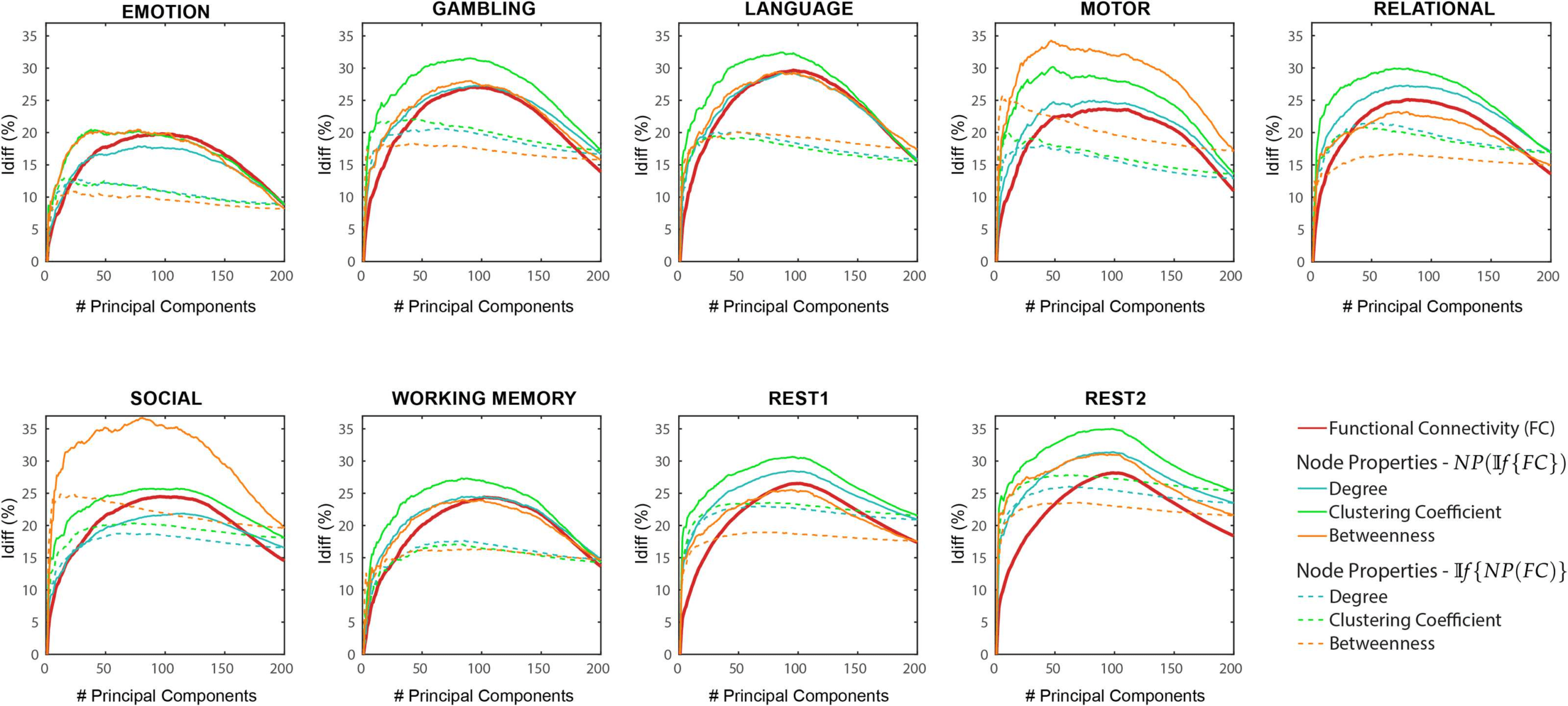
In If {NP(FC)}) the different network properties (refer Methods) were first derived from the
original functional connectomes and subsequently decomposed and reconstructed using the
identifiability framework. Idiff scores were computed on these reconstructed network prop-
erties for a different number of components and compared with those computed from the
reconstructed FCs (voir la figure 2).
As opposed to results shown in Figure 1, which used NP(If {FC}), network properties have
heterogeneous Idiff profiles with respect to number of components. Compared with Idiff from
If {FC}, search information has a higher peak Idiff score for all tasks, while communicability
has a higher peak Idiff score for all tasks except resting state. We also find that MFPT has a
very different Idiff profile compared with other network properties. The Idiff profiles of MFPT
from If {MFPT(FC)} increases as we add the first few component and saturates or decreases
gradually as more components are added (starting at around 20 components for all tasks). Ce
is unlike other network properties and functional connectomes that share similar Idiff profiles
Neurosciences en réseau
704
Uncovering identifiability in properties of functional connectomes
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
If {NP(FC)} Differential identifiability (Idiff ) profiles of pairwise properties for different fMRI tasks as a function of the number of
Chiffre 2.
principal components used for reconstruction. Ici, the identifiability framework was applied directly on the network properties derived from
the original functional connectomes (If {NP(FC)}). Each plot shows, for each fMRI task, the Idiff score associated with functional connectivity
(red solid line) and the Idiff scores on reconstructed network properties derived from the original functional connectomes, If {NP(FC)} (voir
legend) for different numbers of components.
(voir la figure 2). A summary of maximum Idiff , corresponding number of components used and
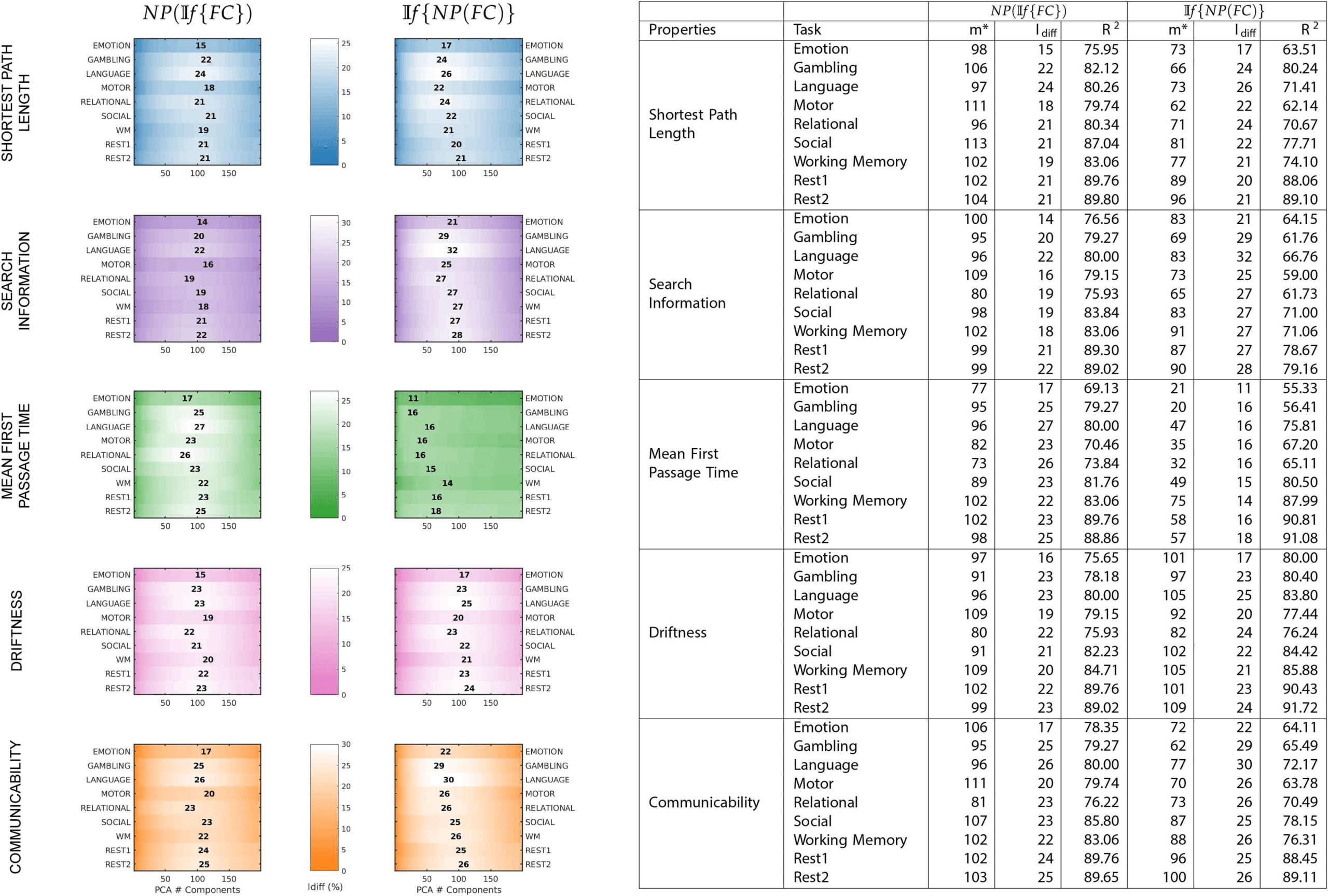
variance retained for NP(If {FC}), and If {NP(FC)} can be seen in Figure 3.
The network property with the most different Idiff profiles was between MFPT(If {FC})
and If {MFPT(FC)}. Search information was the only network property that reached higher
Idiff values for all fMRI tasks for If {SI(FC)}. The difference between search information
and mean first passage time are assessed in detail in Figure 4. shaded area highlights the
variability of Idiff scores across different tasks for NP(If {FC}) (solid area) and If {NP(FC)}
(hatched area). Across all tasks, Idiff on If {SI(FC)} is higher than SI(If {FC}.
Cependant, for Mean First Passage time, Idiff on MFPT(If {(FC)} is higher than (If {MFPT
(FC)}. When SI(If {FC}) is derived and optimally reconstructed, Idiff on search information
is highest across all tasks. Cependant, under full reconstruction m = 200 (which is equivalent to
using the original functional connectomes), Idiff scores are highest for the functional connec-
tome for all fMRI tasks.
We then assessed how differential identifiability varies based on node properties: degree,
betweeness centrality, and clustering coefficient (Chiffre 5). We find that the Idiff profiles of
NP(If {FC} are similar to that of If {FC}. These also give a significantly higher optimal Idiff
score for gambling, langue, moteur, and working memory tasks for all node properties. Es-
pecially in the case of language and motor tasks, betweeness centrality gives a significantly
higher Idiff of 37 et 35 respectively at optimal reconstruction. For If {NP(FC)}, results show
lower and flatter Idiff profiles for all tasks and a wide range of number of components. Idiff pro-
files using NP(If {FC}) of these node properties are in agreement with all pairwise properties
explored so far. In contrast, the Idiff profiles using If {NP(FC)} on these node properties are
similar to If {MFPT(FC)} only.
Neurosciences en réseau
705
Uncovering identifiability in properties of functional connectomes
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
.
t
Chiffre 3. A summary of maximum Idiff values, corresponding to number of components and explained variance retained for each fMRI
task and network property for both NP(If {FC}) and If {NP(FC)}. On the left, each plot shows, for each property and each method—
NP(If {FC}) or If {NP(FC)}—the Idiff score for all tasks. The number mentioned gives the maximum Idiff score for the corresponding task
(y-axis) and the position denotes the number of components (x-axis). On the right is the same information summarized as a table. For each
method and network property, the table gives the number of components used for optimal reconstruction m∗, corresponding maximum Idiff
valeur, and the variance explained at that reconstruction R2.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Intraclass correlation coefficient was used to assess the task sensitivity of each pairwise
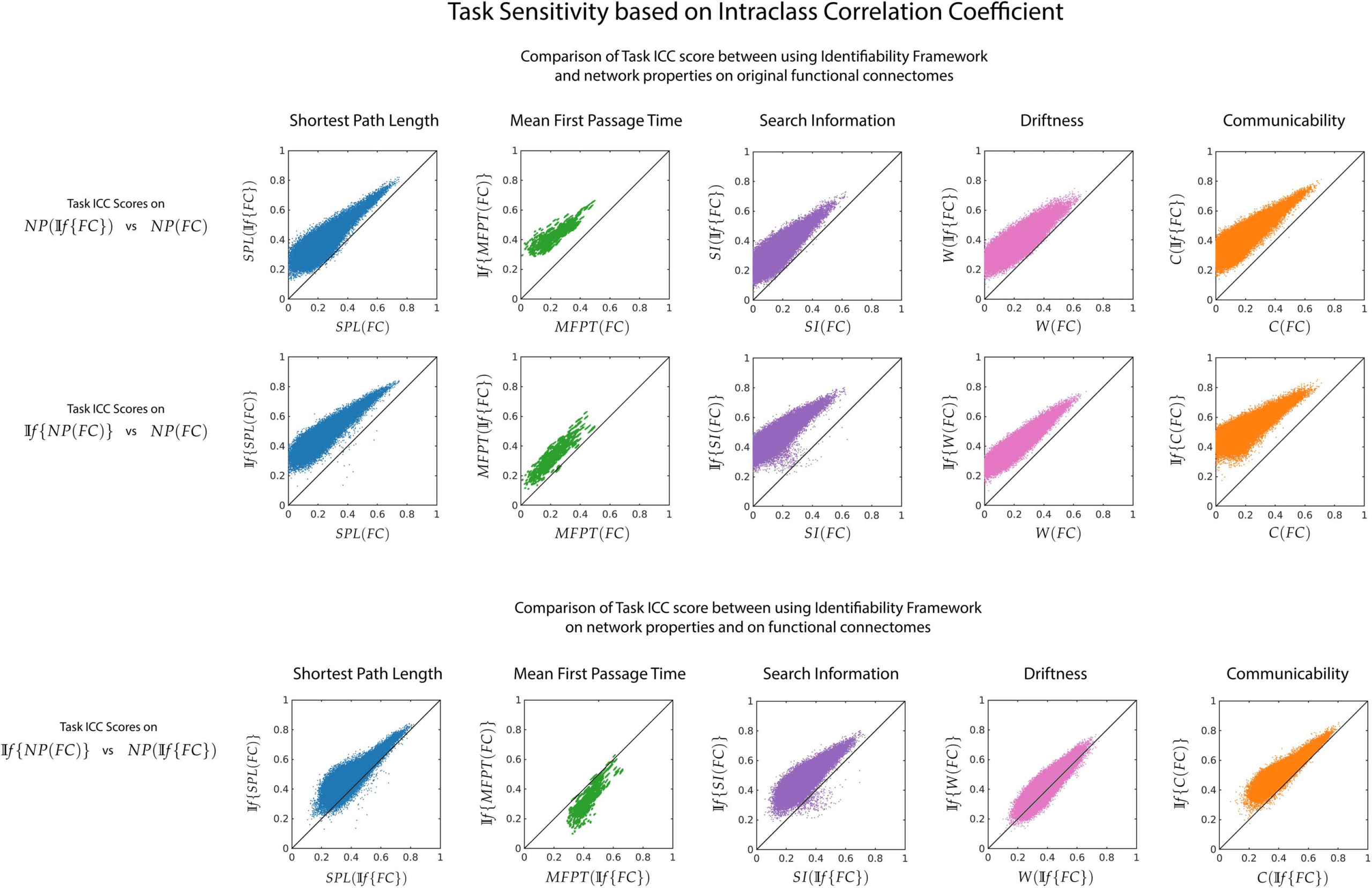
network property for three possible cases: NP(If {FC}) vs NP(FC) (Chiffre 6, top row), If {NP
(FC)} vs NP(FC) (Chiffre 6, middle row) and NP(If {FC}) vs If {NP(FC)} (Chiffre 6, bottom
row). We find that the task sensitivity is higher for all network properties when the identifiability
framework was used (for both NP(If {FC}) and If {NP(FC)}). Between NP(If {FC}) et
If {NP(FC)}, there is no one method that improves task sensitivity for all network properties.
DISCUSSION
Brain connectivity fingerprinting has taken center stage in the neuroscientific community (Byrge
& Kennedy, 2019; Finn et al., 2015; Gratton et al., 2018; Mars et al., 2018; Miranda-Dominguez
et coll., 2014; Satterthwaite et al., 2018; Seitzman et al., 2019; Venkatesh et al., 2019). Comme nous
move in this direction, there is a need to improve the reliability and robustness of individual
Neurosciences en réseau
706
Uncovering identifiability in properties of functional connectomes
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
.
t
Chiffre 4. Assessment of the two most divergent network measurement Idiff profiles. (UN) Across tasks and rest differential identifiability (Idiff )
for mean first passage time as a function of the number of principal components used for reconstruction. Solid line and solid shaded area
represent the results for MFPT(If {FC}). Dashed line and hatched area show results for If {MFPT(FC)} (B) Across tasks and rest differential
identifiability (Idiff ) for search information as a function of the number of principal components used for reconstruction. Solid line and solid
shaded area represent the results for SI(If {FC}). Dashed line and hatched area show results for If {SI(FC)} The differential identifiability
matrice (as defined in the Methods section) is shown at optimal reconstruction for language task for (C) MFPT(If {FC}), (D) If {FC} et (E)
If {SI(FC)}. The diagonal elements in each matrix represent Iself and the non-diagonal elements represent Iothers.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
fingerprint in functional connectomes and on common network measures extracted from func-
tional connectomes. The identifiability framework (If ) has shown the capacity to uncover subject
fingerprint as measured by the Idiff score in human functional connectomes, regardless of the
Improving differential identifiability using the If frame-
fMRI task (Amico & Goñi, 2018b).
work on functional connectomes (FCs) has been shown to improve the test-retest reliability
of FCs and correlation with fluid intelligence (Amico & Goñi, 2018b). Ici, we extend this
framework to show that by maximizing individual fingerprints in the functional connectomes,
we also maximize individual fingerprint in network properties derived from the connectomes.
En outre, we found that uncovering individual fingerprinting on network measurements
also improves task signature.
En outre, we show that in certain network properties, nous
can uncover an even stronger fingerprint if we apply the framework directly on the network
property instead of functional connectomes.
Neurosciences en réseau
707
Uncovering identifiability in properties of functional connectomes
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 5. NP(If {FC}) and If {NP(FC)} Differential identifiability (Idiff ) of node properties for different fMRI tasks as a function of the
number of principal components used for reconstruction. Each plot shows, for each task, the Idiff score associated with functional connectivity
(red solid line), the Idiff scores on the network properties derived from the reconstructed functional connectomes NP(If {FC}) (solid lines,
colors – see legend) and the Idiff scores on the reconstructed network properties derived from the original functional connectomes If {NP(FC)}
(dotted lines, colors – see legend) for different numbers of components.
Numerous work has been done to assess the effect of a change in parameters of the acqui-
sition process and the preprocessing pipelines on test-retest (TRT) reliability of fMRI data (Birn
et coll., 2013; Noble, Scheinost, & Constable, 2019; Noble et al., 2017; Shah, Cramer, Ferguson,
Birn, & Anderson, 2016). The impact of different correlation metrics, inclusion or exclusion
of edges on functional connectomes, as well as the use of global signal regression, have been
explored extensively (Byrge & Kennedy, 2019; Cao et al., 2014; Fiecas et al., 2013; Liang et al.,
2012; Schwarz & McGonigle, 2011; Wang et al., 2011). En plus, TRT reliability is also
seen to be affected by band pass filtering, scan length, sampling rate, network definition of
the weights, and size of voxels for node definition (Braun et al., 2012; Liang et al., 2012;
Liao et al., 2013). Given that the TRT reliability of the fMRI data and the subsequent estima-
tion of functional connectomes are affected by such diverse factors, it is important to explore
the reliability of the derived network properties. Even though TRT reliability is not the only
parameter to take into account when choosing the optimal strategy for brain network analyses,
it surely has to be considered an important factor to help in such an important choice.
Essentially, If works as a group-level data-driven (denoising) procedure where the com-
ponents not contributing towards test-retest reliability of FCs are identified and removed. If
doesn’t just improve the overall TRT reliability of a functional connectome but also improves
it locally on an edge-level (Amico & Goñi, 2018b) which should ensure that both global and
local network properties computed using these denoised functional connectomes are more
reliable and robust. As shown in Figure 1, If not only maximizes subject fingerprint at the FC
level, but also at the network property level, which validated our premise. En outre, this con-
vergent behavior is not present just at the optimal point; the identifiability profile of network
properties follows the identifiability profile of the functional connectomes. En substance, we have
Neurosciences en réseau
708
Uncovering identifiability in properties of functional connectomes
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 6. Effect of If on task sensitivity of network measures. For each pairwise network property, task sensitivity is measured using ICC
between NP(If {FC}) vs NP(FC) (top row), If {NP(FC)} versus NP(FC) (middle) and NP(If {FC}) vs If {NP(FC)} (bottom row). Le
first two rows highlight the fact that the If framework uncovers the inherently distinct signature of different tasks through derived network
properties. The last row shows that certain network properties would benefit more from application of the If framework on the functional
connectomes, while others from application directly on the network properties.
shown that regardless of whether you are using functional connectomes or the network
properties derived from them, using If framework on the functional connectomes would be a
beneficial first step.
A natural next question was to find whether If should be applied on functional connectomes
and then derive the network properties (NP(If {FC})), or to use it directly on the network prop-
erties derived from original functional connectomes (If {NP(FC)}). The two approaches are
an attempt to understand different principles of what a fingerprint is in a network derived mea-
surement. If {NP(FC)} assumes that functional connectomes are “holding” the individual fin-
gerprints and then propagating them to the network measurements. The fact that maximizing
fingerprint of functional connectomes also maximizes the fingerprint in derived network mea-
sures, suggests that functional connectomes do indeed hold a subject fingerprint that is then
transmitted to the derived network properties. On the other hand, we also see that for some
network measures (par exemple., search information), we can uncover a better fingerprint if we apply
the framework directly on the network measure. This suggests that specific network measures
have a subject fingerprint of their own which gets added on to the functional connectome
Neurosciences en réseau
709
Uncovering identifiability in properties of functional connectomes
fingerprint. Ainsi, if under some circumstances, the goal is to maximize the reliability and
the individual variability of a specific network property, one can benefit from applying the If
framework on the network property itself, rather than on FCs.
Notably, in the If {SI(FC)} scénario, the most different Idiff profiles were found between
MFPT and search information (Chiffre 4). Search information consistently provides a better
fingerprint across all tasks than does functional connectome. MFPT, cependant, can neither im-
prove nor match the fingerprint of functional connectomes. Aussi, it can not retain the finger-
print that is otherwise present is the functional connectomes and is then propgated to MFPT
using If {MFPT(FC)}. Ainsi, while some properties (c'est à dire., search information) can derive
higher identifiability than functional connectomes, properties like MFPT need to be computed
on optimally reconstructed functional connectomes to uncover subject identifiability on it.
These findings show that brain fingerprinting can be improved by deriving network mea-
surements that extract multivariate information from bivariate measurements such as pairwise
correlations used to estimate FCs. Specifically, individual fingerprint peaks on network mea-
surements (par exemple., search information) that are more multivariate and requires more information
on the global topology of the functional network. Cependant, if the information is heavily driven
by degree properties (par exemple., MFPT), then there is no improvement on the individual fingerprint
(Chiffre 4). This is strongly corroborated by the Idiff profiles of several node properties under
the If {NP(FC)} scénario. These profiles are very similar to that of MFPT, a network prop-
erty which has a strong negative correlation with the degree of the target node. Although
If {NP(FC)} of these node properties have Idiff profiles similar to If {MFPT(FC)}, the max-
imum Idiff on these node properties are, for some tasks, significantly higher than If {FC}.
Betweeness centrality, Par exemple, has a higher subject identifiability for social and motor
tasks.
It was interesting to observe that under the If {NP(FC)} scénario, betweenness centrality
maximizes differential identifiability using just the first two components for social and mo-
tor tasks and that it was higher than the identifiability of the functional connectomes for any
number of components. Since betweenness centrality can be used to identify integrative com-
munication hubs in FCs (Sporns, 2013), it can be argued that social and motor tasks display a
“hub functional fingerprint”, which can be captured by the first two principal components.
A complementary assessment to the identification of subject fingerprints is to assess the
ability to identify the different tasks used in this study. To do so, we used intraclass correlation
coefficient on the derived network properties. The If framework improved task sensitivity
on the network properties (voir la figure 6). Regardless of using the framework on the original
functional connectomes or on the network properties themselves, a higher task sensitivity is
obtained using one of the process depending on the network property.
In both cases, le
task reliability of the network properties has improved. The different tasks in the HCP dataset
aim to assess different cognitive processes. Ainsi, the corresponding connectomes and the
network properties derived from them should, at least to some extent, be task specific. We have
shown that using the If framework uncovers task-related fingerprints where unique cognitive
processes result in differential network properties.
To summarize, differential identifiability was found to be always higher on functional con-
nectomes than on any network properties when the identifiability framework (If ) is not used.
When If improved identifiability on functional connectomes, the identifiability on the network
properties also increased. The framework also improved the subject fingerprints of the network
properties. Not only do they improve at the optimal point, but the differential identifiability
follows the same profile on network properties as it does on functional connectomes. We also
find that applying the identifiability framework on the network properties instead of functional
connectomes gives higher differential identifiability for some network properties. At optimal
Neurosciences en réseau
710
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Uncovering identifiability in properties of functional connectomes
reconstruction, we find that search information has higher differential identifiability than func-
tional connectomes across all tasks when the identifiability framework is applied on search
information. This shows that there are network properties that can uncover better identifiabil-
ity with the framework than the functional connectomes themselves. Enfin, we found that
using the identifiability framework (either on functional connectomes or network property)
improves task sensitivity in all network properties.
Our study has some limitations. Only the unrelated subjects of the Human Connectome
project and the cortical parcellation proposed by Glasser et al. (2013) are used in this work.
Other explorations with other atlases, parcellations and/or other estimators of functional cou-
pling (other than Pearson’s correlation coefficient) would expand on the implications of our
travail. We have also limited our study to commonly used five pairwise and three node net-
work properties. Delving into other network properties can strengthen this framework further
and provide additional insights in understanding the associations between brain fingerprints,
functional connectivity, and network derived properties. It could also be possible that relevant
combinations of network measurements (driftness is an example of it) might uncover additional
brain fingerprints and reach even higher differential identifiability levels.
This study can be extended to clinical applications to understand diseases that target specific
functions of the human brain. Par exemple, for assessing pathologies whose signature cannot
be mapped on the functional connectomes themselves but can be assessed using different
network properties derived from them (Bassett & Bullmore, 2009; Fornito & Bullmore, 2015;
Fornito et al., 2015) Dans ce cas, to retain individual differences and to be able to differentiate
healthy population from clinical ones, we need this study to understand the advantages of
using the identifiability framework on the functional connectome or network property. Enfin,
studying the effect of the framework on the structural connectome is another natural extension
of this work.
REMERCIEMENTS
Data were provided (in part) by the Human Connectome Project, WU-Minn Consortium (prin-
cipal investigators: David Van Essen and Kamil Ugurbil; 1U54MH091657), funded by the 16
NIH Institutes and Centers that support the NIH Blueprint for Neuroscience Research; and by
the McDonnell Center for Systems Neuroscience at Washington University. The authors thank
Docteur. Gorka Zamora-Lopez and Dr. Matthieu Gilson for useful comments.
CONTRIBUTIONS DES AUTEURS
Meenusree Rajapandian: Conceptualisation; Conservation des données; Analyse formelle; Visualisation;
En écrivant – Original Draft. Enrico Amico: Conservation des données; Méthodologie; En écrivant – Original Draft.
Kausar Abbas: Analyse formelle; Méthodologie; En écrivant – Original Draft. Mario Ventresca:
Surveillance; En écrivant – Original Draft. Joaquín Goñi: Conceptualisation; Conservation des données; Formal
analyse; Acquisition de financement; Méthodologie; Ressources; Surveillance; En écrivant – Original Draft.
INFORMATIONS SUR LE FINANCEMENT
Joaquín Goñi, National Institutes of Health (U. S.), Award ID: R01EB022574. Joaquín Goñi,
National Institutes of Health, Award ID: R01MH108467. Joaquín Goñi, National Institutes of
Health, Indiana Alcohol Research Center, Award ID: P60AA07611. Joaquín Goñi and Mario
Ventresca, Purdue Discovery Park Data Science Award “Fingerprints of the Human Brain: UN
Data Science Perspective.”
Neurosciences en réseau
711
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Uncovering identifiability in properties of functional connectomes
RÉFÉRENCES
Alavash, M., Hilgetag, C. C., Thiel, C. M., & Gießing, C. (2015). Per-
sistency and flexibility of complex brain networks underlie dual-
task interference. Human brain mapping, 36(9), 3542–3562.
Amico, E., Arenas, UN., & Goñi, J.. (2019). Centralized and distributed
cognitive task processing in the human connectome. Réseau
Neurosciences, 3(2), 455–474.
Amico, E., & Goñi, J.. (2018un). Mapping hybrid functional-structural
connectivity traits in the human connectome. Network Neuro-
science, 2(3), 306–322.
Amico, E., & Goñi, J.. (2018b). The quest for identifiability in human
functional connectomes. Scientific Reports, 8(1), 8254.
Avena-Koenigsberger, UN., Misic, B., & Sporns, Ô. (2018). Commu-
nication dynamics in complex brain networks. Nature Reviews
Neurosciences, 19(1), 17.
Bari, S., Amico, E., Vike, N., Talavage, T. M., & Goñi, J..
(2019).
Uncovering multi-site identifiability based on resting-state func-
tional connectomes. NeuroImage, 202, 115967.
Bartko, J.. J.. (1966). The intraclass correlation coefficient as a mea-
sure of reliability. Psychological Reports, 19(1), 3–11.
Bassett, D. S., & Bullmore, E. T. (2009). Human brain networks in
health and disease. Opinion actuelle en neurologie, 22(4), 340.
Birn, R.. M., Molloy, E. K., Patriat, R., Parker, T., Meier, T. B., Kirk,
G. R., . . . Prabhakaran, V. (2013). The effect of scan length on
the reliability of resting-state fMRI connectivity estimates. Neu-
roImage, 83, 550–558.
Biswal, B. B., Mennes, M., Zuo, X.-N., Gohel, S., Kelly, C., Forgeron,
S. M., . . . Milham, M.. P..
(2010). Toward discovery science of
human brain function. Proceedings of the National Academy of
les sciences, 107(10), 4734–4739.
Bola, M., & Sabel, B. UN. (2015). Dynamic reorganization of brain func-
tional networks during cognition. NeuroImage, 114, 398–413.
Brun, U., Plichta, M.. M., Esslinger, C., Sauer, C., Haddad, L.,
(2012). Test–retest re-
Grimm, O., . . . Meyer-Lindenberg, UN.
liability of resting-state connectivity network characteristics us-
ing fMRI and graph theoretical measures. NeuroImage, 59(2),
1404–1412.
Byrge, L., & Kennedy, D. P.. (2019). High-accuracy individual identi-
fication using a thin slice of the functional connectome. Réseau
Neurosciences, 3(2), 363–383.
Cao, H., Plichta, M.. M., Schäfer, UN., Haddad, L., Grimm, O.,
Schneider, M., . . . Tost, H. (2014). Test–retest reliability of fmri-
based graph theoretical properties during working memory, emo-
tion processing, and resting state. NeuroImage, 84, 888–900.
Castellanos, F. X., Di Martino, UN., Craddock, R.. C., Mehta, UN. D.,
& Milham, M.. P.. (2013). Clinical applications of the functional
connectome. NeuroImage, 80, 527–540.
Cohen, J.. R., & D'Esposito, M.. (2016). The segregation and integra-
tion of distinct brain networks and their relationship to cognition.
Journal des neurosciences, 36(48), 12083–12094.
Costa, L. d. F., Batista, J.. L., & Ascoli, G. UN.
(2011). Communi-
cation structure of cortical networks. Frontiers in Computational
Neurosciences, 5, 6.
Crofts, J.. J., & Higham, D. J.. (2009). A weighted communicability
measure applied to complex brain networks. Journal of the Royal
Society Interface, 6(33), 411–414.
Crossley, N. UN., Mechelli, UN., Scott, J., Carletti, F., Fox, P.. T.,
McGuire, P., & Bullmore, E. T. (2014). The hubs of the human
connectome are generally implicated in the anatomy of brain dis-
ordres. Cerveau, 137(8), 2382–2395.
Davison, E. N., Schlesinger, K. J., Bassett, D. S., Lynall, M.-E.,
Miller, M.. B., Grafton, S. T., & Carlson, J.. M.. (2015). Brain net-
work adaptability across task states. Biologie computationnelle PLoS,
11(1), e1004029.
Deco, G., Tononi, G., Boly, M., & Kringelbach, M.. L.
(2015).
Rethinking segregation and integration: Contributions of
whole-brain modelling. Nature Revues Neurosciences, 16(7),
430–439.
Estrada, E., & Hatano, N.
(2008). Communicability in complex
réseaux. Physical Review E, 77(3), 036111.
Fiecas, M., Ombao, H., Van Lunen, D., Baumgartner, R., Coimbra,
UN., & Feng, D. (2013). Quantifying temporal correlations: A test–
retest evaluation of functional connectivity in resting-state fmri.
NeuroImage, 65, 231–241.
Finn, E. S., Shen, X., Scheinost, D., Rosenberg, M.. D., Huang, J.,
(2015). Functional con-
Chun, M.. M., . . . Constable, R.. T.
nectome fingerprinting: Identifying individuals using patterns of
brain connectivity. Neurosciences naturelles, 18(11), 1664.
Fornito, UN., & Bullmore, E. T.
(2015). Connectomics: A new
paradigm for understanding brain disease. European Neuropsy-
chopharmacology, 25(5), 733–748.
Fornito, UN., Zalesky, UN., & Breakspear, M..
(2015). The connec-
tomics of brain disorders. Nature Revues Neurosciences, 16(3),
159–172.
Fornito, UN., Zalesky, UN., & Bullmore, E.
(2016). Fundamentals
of brain network analysis. Amsterdam, Netherlands: Academic
Presse.
Fukushima, M., Betzel, R.. F., Il, Y., van den Heuvel, M.. P., Zuo,
(2018). Structure–function relationships
X.-N., & Sporns, Ô.
during segregated and integrated network states of human brain
functional connectivity. Structure et fonction du cerveau, 223(3),
1091–1106.
Glasser, M.. F., Coalson, T. S., Robinson, E. C., Hacker, C. D., Harwell,
J., Yacoub, E., . . . Van Essen, D. C. (2016). A multi-modal par-
cellation of human cerebral cortex. Nature, 536(7615), 171.
Glasser, M.. F., Sotiropoulos, S. N., Wilson, J.. UN., Coalson, T. S.,
Fischl, B., Andersson, J.. L., . . . WU-Minn HCP Consortium.
(2013). The minimal preprocessing pipelines for the human con-
nectome project. NeuroImage, 80, 105–124.
Goñi,
J., van den Heuvel, M.. P., Avena-Koenigsberger, UN.,
de Mendizabal, N. V., Betzel, R.. F., Griffa, UN., . . . Sporns, Ô.
(2014). Resting-brain functional connectivity predicted by ana-
lytic measures of network communication. Actes du
Académie nationale des sciences, 111(2), 833–838.
Gratton, C., Laumann, T. O., Nielsen, UN. N., Vert, D. J., Gordon,
E. M., Gilmore, UN. W., . . . Petersen, S. E. (2018). Functional brain
networks are dominated by stable group and individual factors,
not cognitive or daily variation. Neurone, 98(2), 439–452.
Kemeny, J.. G., & Snell, J.. L. (1976). Markov chains. New York, New York:
Springer-Verlag.
Neurosciences en réseau
712
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Uncovering identifiability in properties of functional connectomes
Liang, X., Wang, J., Yan, C., Shu, N., Xu, K., Gong, G., & Il, Oui.
(2012). Effects of different correlation metrics and preprocessing
factors on small-world brain functional networks: A resting-state
functional MRI study. PLoS ONE, 7(3), e32766.
Liao, X.-H., Xia, M.-R., Xu, T., Dai, Z.-J., Cao, X.-Y., Niu, H.-J., . . .
Il, Oui. (2013). Functional brain hubs and their test–retest reliabil-
ville: A multiband resting-state functional mri study. NeuroImage,
83, 969–982.
Mars, R.. B., Passingham, R.. E., & Jbabdi, S.
(2018). Connectivity
fingerprints: From areal descriptions to abstract spaces. Trends
in Cognitive Sciences, 22(11), 1026–1037.
Mattar, M.. G., Betzel, R.. F., & Bassett, D. S.
(2016). The flexible
brain. Cerveau, 139(8), 2110–2112.
McGraw, K. O., & Wong, S. P..
(1996). Forming inferences about
some intraclass correlation coefficients. Psychological Methods,
1(1), 30.
Miranda-Dominguez, O., Mills, B. D., Carpenter, S. D., Grant, K. UN.,
Kroenke, C. D., Nigg, J.. T., & Fair, D. UN. (2014). Connectotyping:
Model based fingerprinting of the functional connectome. PLoS
ONE, 9(11), e111048.
Noble, S., Scheinost, D., & Constable, R.. T. (2019). A decade of test-
retest reliability of functional connectivity: A systematic review
and meta-analysis. NeuroImage, 203, 116157.
Noble, S., Spann, M.. N., Tokoglu, F., Shen, X., Constable, R.. T., &
Scheinost, D. (2017). Influences on the test–retest reliability of
functional connectivity MRI and its relationship with behavioral
utility. Cortex cérébral, 27(11), 5415–5429.
Pallarés, V., Insabato, UN., Sanjuán, UN., Kühn, S., Mantini, D., Deco,
G., & Gilson, M..
(2018). Extracting orthogonal subject-and
condition-specific signatures from fMRI data using whole-brain
effective connectivity. NeuroImage, 178, 238–254.
Petrella, J.. R..
(2011). Use of graph theory to evaluate brain net-
travaux: A clinical tool for a small world? Radiological Society of
North America.
Rosenberg, M.. D., Scheinost, D., Vert, UN. S., Avery, E. W., Kwon,
Oui. H., Finn, E. S., . . . Chun, M.. M.. (2019). Functional connectiv-
ity predicts changes in attention over minutes, jours, and months.
bioRxiv:700476.
Rosvall, M., Trusina, UN., Minnhagen, P., & Sneppen, K. (2005). Net-
works and cities: An information perspective. Physical Review
Letters, 94(2), 028701.
Rubinov, M., & Sporns, Ô. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069.
Satterthwaite, T. D., Xia, C. H., & Bassett, D. S. (2018). Personal-
ized neuroscience: Common and individual-specific features in
functional brain networks. Neurone, 98(2), 243–245.
Scheinost, D., Noble, S., Horien, C., Vert, UN. S., Lake, E. M.,
(2019). Ten simple rules for
Salehi, M., . . . Constable, R.. T.
predictive modeling of individual differences in neuroimaging.
NeuroImage, 193, 35–45.
Schwarz, UN. J., & McGonigle, J..
(2011). Negative edges and soft
thresholding in complex network analysis of resting state func-
tional connectivity data. NeuroImage, 55(3), 1132–1146.
Seitzman, B. UN., Gratton, C., Laumann, T. O., Gordon, E. M.,
Adeyemo, B., Dworetsky, UN., . . . Petersen, S. E. (2019). Trait-like
Sporns, Ô.
Presse.
Sporns, Ô.
variants in human functional brain networks. Actes du
Académie nationale des sciences, 116(45), 22851–22861.
Shah, L. M., Cramer, J.. UN., Ferguson, M.. UN., Birn, R.. M., & Anderson,
(2016). Reliability and reproducibility of individual differ-
J.. S.
ences in functional connectivity acquired during task and resting
state. Brain and Behavior, 6(5), e00456.
Shen, X., Finn, E. S., Scheinost, D., Rosenberg, M.. D., Chun, M.. M.,
Papademetris, X., & Constable, R.. T. (2017). Using connectome-
based predictive modeling to predict individual behavior from
brain connectivity. Nature Protocols, 12(3), 506.
Shrout, P.. E., & Fleiss, J.. L. (1979). Intraclass correlations: Uses in
assessing rater reliability. Psychological Bulletin, 86(2), 420.
Forgeron, S. M., Beckmann, C. F., Andersson, J., Auerbach, E. J.,
Bijsterbosch, J., Douaud, G., . . . WU-Minn HCP Consortium.
(2013). Resting-state fMRI in the Human Connectome Project.
NeuroImage, 80, 144–168.
(2010). Networks of the brain. Cambridge, MA: AVEC
(2013). Network attributes for segregation and inte-
gration in the human brain. Opinion actuelle en neurobiologie,
23(2), 162–171.
Sporns, Ô.
(2018). Graph theory methods: applications in brain
réseaux. Dialogues in Clinical Neuroscience, 20(2), 111.
Sporns, O., & Betzel, R.. F. (2016). Modular brain networks. Annual
Review of Psychology, 67, 613–640.
Svaldi, D. O., Goñi,
J., Abbas, K., Amico, E., Clark, D. G.,
(2019). Optimisation
identifiability improves connectome predictive
arXiv
Muralidharan, C., . . . Apostolova, L. G.
differential
modeling of cognitive deficits in alzheimer´s disease.
preprint arXiv:1908.06197.
Svaldi, D. O., Goñi, J., Sanjay, UN. B., Amico, E., Risacher, S. L., West,
J.. D., . . . Apostolova, L. (2019). Towards subject and diagnostic
identifiability in the Alzheimer’s disease spectrum based on func-
tional connectomes. In D. Stoyanov, Z. Taylor, E. Ferrante, UN. V.
Dalca, et autres. (Éd.), Graphs in biomedical image analysis and inte-
grating medical imaging and non-imaging modalities (pp. 74–82).
Cham, Suisse: Springer.
Van Essen, D. C., Forgeron, S. M., Barch, D. M., Behrens, T. E., Yacoub,
E., Voiture d'Ugur, K., & WU-Minn HCP Consortium. (2013). The wu-
minn Human Connectome Project: An overview. NeuroImage,
80, 62–79.
Venkatesh, M., Jaja, J., & Personne, L. (2019). Comparing functional
connectivity matrices: A geometry-aware approach applied to
participant identification. bioRxiv:687830.
Wang, J.-H., Zuo, X.-N., Gohel, S., Milham, M.. P., Biswal, B. B., &
Il, Oui. (2011). Graph theoretical analysis of functional brain net-
travaux: Test-retest evaluation on short-and long-term resting-state
functional mri data. PLoS ONE, 6(7), e21976.
Xie, H., Calhoun, V. D., Gonzalez-Castillo, J., Damaraju, E., Miller,
R., Bandettini, P.. UN., & Mitra, S.
(2018). Whole-brain connec-
tivity dynamics reflect both task-specific and individual-specific
modulation: A multitask study. NeuroImage, 180, 495–504.
Zalesky, UN., Fornito, UN., & Bullmore, E. T. (2010). Network-based
statistique: Identifying differences in brain networks. NeuroImage,
53(4), 1197–1207.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
4
3
6
9
8
1
8
6
7
3
6
5
n
e
n
_
un
_
0
0
1
4
0
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Neurosciences en réseau
713