RECHERCHE
Changes in structural network topology correlate
with severity of hallucinatory behavior in
Parkinson’s disease
1,2
Julie M. Hall
Joseph R. Phillips
1,2
, Claire O’Callaghan
2,3
2
, Alana J. Muller
, Ahmed A. Moustafa
, Simon J. G. Lewis
1,4
, Kaylena A. Ehgoetz Martens
2
, and James M. Shine
2
,
2
un accès ouvert
journal
1School of Social Sciences and Psychology, Western Sydney University, Milperra, NSW, Australia
2Brain and Mind Centre, University of Sydney, Camperdown, NSW, Australia
Department of Psychiatry and Behavioural and Clinical Neuroscience Institute, University of Cambridge,
Cambridge, United Kingdom
3
4
MARCS Institute, Western Sydney University, Milperra, NSW, Australia
Mots clés: Parkinson’s disease, Visual hallucinations, Diffusion tensor imaging, Network topology,
Graph theory, Connectomics
ABSTRAIT
Inefficient integration between bottom-up visual input and higher order visual processing
regions is implicated in visual hallucinations in Parkinson’s disease (PD). Ici, nous
investigated white matter contributions to this perceptual imbalance hypothesis. Twenty-nine
PD patients were assessed for hallucinatory behavior. Hallucination severity was correlated
to connectivity strength of the network using the network-based statistic approach. Le
results showed that hallucination severity was associated with reduced connectivity within a
subnetwork that included the majority of the diverse club. This network showed overall
greater between-module scores compared with nodes not associated with hallucination
severity. Reduced between-module connectivity in the lateral occipital cortex, insula, et
pars orbitalis and decreased within-module connectivity in the prefrontal, somatosensory,
and primary visual cortices were associated with hallucination severity. Inversement,
hallucination severity was associated with increased between- and within-module
connectivity in the orbitofrontal and temporal cortex, as well as regions comprising the
dorsal attentional and default mode network. These results suggest that hallucination severity
is associated with marked alterations in structural network topology with changes in
participation along the perceptual hierarchy. This may result in the inefficient transfer of
information that gives rise to hallucinations in PD.
RÉSUMÉ DE L'AUTEUR
Inefficient integration of information between external stimuli and internal perceptual
predictions may lead to misperceptions or visual hallucinations in Parkinson’s disease (PD).
Dans cette étude, we show that hallucinatory behavior in PD patients is associated with marked
alterations in structural network topology. Severity of hallucinatory behavior was associated
with decreased connectivity in a large subnetwork that included the majority of the diverse
club, nodes with a high number of between-module connections. En outre, changes
in between-module connectivity were found across brain regions involved in visual
traitement, top-down prediction centers, and endogenous attention, including the occipital,
orbitofrontal, and posterior cingulate cortex. Ensemble, these findings suggest that impaired
Citation: Hall, J.. M., O’Callaghan, C.,
Muller, UN. J., Ehgoetz Martens, K. UN.,
Phillips, J.. R., Moustafa, UN. UN., Lewis,
S. J.. G., & Shine, J.. M.. (2019). Changes
in structural network topology
correlate with severity of hallucinatory
behavior in Parkinson’s disease.
Neurosciences en réseau, 3(2), 521–538.
https://doi.org/10.1162/netn_a_00078
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00078
Informations complémentaires:
https://doi.org/10.1162/netn_a_00078
https://github.com/juliemaehall/
topology
Reçu: 10 Août 2018
Accepté: 21 Décembre 2018
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Julie M. Hall
julie.hall@sydney.edu.au
Éditeur de manipulation:
Danielle Bassett
droits d'auteur: © 2019
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
integration across different sides across different perceptual processing regions may result in
inefficient transfer of information.
INTRODUCTION
Visual hallucinations (VHs) in Parkinson’s disease (PD) exist on a spectrum ranging from simple
misperceptions to complex well-formed images (Barnes & David, 2001). With disease progres-
sion and loss of insight, VHs constitute a major source of distress for the patient (Goetz, 2009;
Schrag, 2004) and comprise a high degree of burden for caregivers (Aarsland et al., 2007).
Risk factors of VHs include older age and disease duration, sleep and mood disturbances, comme
well as cognitive decline (Barnes & David, 2001; Fénelon, Mahieux, Huon, & Ziégler, 2000;
Lenka, Hegde, Arumugham, & Pal, 2017). En outre, previous work has shown that patients
with VHs show disruptions in attentional processing (Hall et al., 2016), reduced performance
on visuoperceptive tasks (Barnes, Boubert, Harris, Lee, & David, 2003; Gallagher et al., 2011;
Ramírez-Ruiz, Junqué, Martí, Valldeoriola, & Tolosa, 2006), and decreased visual contrast sen-
sitivity, color discrimination (Diederich et al., 1998), and acuity (Matsui et al., 2006). Actuel
models of VHs have therefore focused on the interaction of perceptual and attentional dysfunc-
tion (for a review, see Muller, Shine, Halliday, & Lewis, 2014). Specifically, it has been pro-
posed that failure to effectively integrate information from different processing sites across the
perceptual hierarchy is likely to contribute to VHs and misperceptions in PD (Collerton, Perry,
& McKeith, 2005; Diederich, Goetz, & Stebbins, 2005; Muller et al., 2014; Shine, Halliday,
Carlos, Naismith, & Lewis, 2012).
Attention, prior experience, and expectations strongly influence perception. Perceptual pre-
dictions, generated from a myriad of modalities across the brain, guide perceptual processes
to facilitate the interpretation of noisy and ambiguous input (Bar, 2009; Ange, Fries, & Chanteur,
2001; Summerfield et al., 2006). The orbitofrontal cortex (OFC) processes coarse informa-
tion projected from the visual cortex and provides an “initial guess” of an object’s identity
(Summerfield & Koechlin, 2008). Previous work in PD patients with VHs has shown that the
accumulation of sensory evidence is slow and inefficient, which may result in an overreliance
on these top-down predictions (O’Callaghan et al., 2017). Surtout, top-down visual pro-
cessing regions can modulate neural activity in early visual regions, with expected stimuli
leading to reduced activity (Meyer & Olson, 2011). En plus, activity within the default
mode network (DMN), a network involved in mediating endogenous perception, has shown
to be increased during a misperception in this patient population (Shine, Halliday, et coll., 2014).
Donc, VHs may arise when perceptual input is not properly integrated and internally gen-
erated images interfere with the perceptual process (Fletcher & Frith, 2008; Intaite, Noreika,
Soliunas, & Falter, 2013; O’Callaghan et al., 2017; Powers, Kelley, & Corlett, 2016).
While functional neuroimaging studies have made significant contributions to our
understanding (Hepp, Foncke, Olde Dubbelink, et coll., 2017; Ramírez-Ruiz et al., 2008; Shine,
Halliday, et coll., 2014; Shine, Muller, et coll., 2015; Yao et al., 2014), less is known about the
involvement of white matter changes in the manifestation of VHs in PD. Experiments using dif-
fusion tensor imaging (DTI) have reported altered white matter integrity in the optic nerve and
optic radiation (Lee et al., 2016) as well as ascending tracts from the cholinergic nucleus basalis
of Meynert to parietal and occipital cortical regions (Hepp, Foncke, Berendse, et coll., 2017).
Cependant, given the involvement of large-scale brain networks in perception, unique insights
into white matter changes associated with VHs can be gained by investigating whole-brain
network topology. Topological features of the human connectome allow us to describe the
Neurosciences en réseau
522
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
arrangement of connections within and between segregated submodules (Bullmore & Sporns,
2009). Specifically, nodes that integrate these specialist communities are crucial for incor-
porating information streams of different modalities, which is essential for processes such as
perception (Bertolero, Yeo, & D'Esposito, 2015; Muller, O’Callaghan, Walton, Shine, & Lewis,
2017). Donc, investigating network topology can provide novel insights in changes across
different perceptual hierarchies.
The current study aimed to examine whether VHs are associated with changes in structural
network topology. To identify hallucinatory behavior in patients with PD, we assessed perfor-
mance on a computerized task capable of inducing misperceptions (Shine et al., 2012), dans
combination with a validated questionnaire that evaluates hallucinatory behavior in PD pa-
patients (Shine, Mills, et coll., 2015). We aimed to circumvent the sole reliance on self-reported
symptom occurrence while controlling for the possibility of misclassifying hallucinators as non-
hallucinators, when patients did not experience VHs during their clinic visit or when “passage”
hallucinations go unreported. En plus, by creating this composite score, we are able to
assess visual hallucinatory severity, rather than classifying patients into artificial dichotomous
patient groups.
We hypothesized that the severity of hallucinatory behavior would be associated with
ineffective information processing as shown by reduced between-module scores in visual net-
travaux, reflecting reduced visual input to integration centers. En outre, increased between-
module scores across top-down perceptual prediction areas and the DMN could indicate an
overreliance on regions involved in the generation of internal percepts (Shine, O’Callaghan,
Halliday, & Lewis, 2014).
MÉTHODES
Twenty-nine patients with idiopathic PD were included in this study. Demographic information
including age, disease duration, and levodopa equivalence daily dose (LEDD) were obtained
for all participants. All patients were assessed on the Hoehn & Yahr clinical stage (Hoehn &
Yahr, 1998) and the motor aspect of the Movement Disorder Society Unified Parkinson’s Dis-
ease Rating Scale (MDS-UPDRS) part III (Goetz et al., 2008). Global cognition was assessed
using the Mini-Mental State Examination (MMSE; Folstein, Robins, & Helzer, 1983), and set-
shifting performance was assessed using the Trail Making Test Part B minus Part A (TMTB−A;
Tombaugh, 2004). The study was approved by the ethics committee of the University of Sydney
and was in accordance with the principles of the Helsinki Declaration. Written informed con-
sent was obtained from all participants before participation.
Bistable Percept Paradigm
All patients performed the bistable percept paradigm (BPP; Shine et al., 2012), a behavioral
task capable of inducing misperceptions in susceptible patients. In this task, patients were
presented with either single or bistable percepts (c'est à dire., “hidden” images as shown in Figure 1) pour
a maximum of 30 s in a randomized order. The participant had to decide whether the stimulus
was a single or hidden image by a button press and describe to the examiner what they had
seen. The recorded responses included the following: (un) correct single or correct hidden, (b)
“missed,” recorded when the subject perceived a single image when a bistable percept was
présenté, et (c) “misperceptions,” recorded when a subject incorrectly identified a single
image as a bistable image, c'est, incorrectly reported an image that was not presented on the
screen.
MDS Unified Parkinson’s Disease
Rating Scale:
A commonly used measure of
Parkinson’s disease clinical symptom
severity.
Bistable percept paradigm:
A computer-based task using bistable
percepts to measure visual
misperceptions.
Neurosciences en réseau
523
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
Psychosis and Hallucinations
Questionnaire:
A 20-item self-reported questionnaire
to measure hallucinatory
phenomenology.
Chiffre 1. Example of single and hidden images of the BPP (Shine et al., 2012).
Psychosis and Hallucinations Questionnaire
All patients completed the Psychosis and Hallucinations Questionnaire (PsycH-Q; Shine, Mills,
et coll., 2015). The PsycH-Q (Part A) consists of three subscales including (un) visual mispercep-
tion, which includes questions about the presence of VHs, passage hallucinations, and three
frequently reported contents of VHs including people, animals, and objects; (b) sensory mis-
perceptions, including audition, touch, olfaction, and gustation; et (c) disordered thought
and psychotic behavior. Participants rated the frequency of their symptoms on a 5-point Likert
scale, ranging from 0 (“never experienced”) à 4 (“experienced daily”). The total score was
calculated by summing the responses (Shine, Mills, et coll., 2015; see Supporting Information,
Hall et al., 2019). Part B of the PsycH-Q assesses symptoms related to VHs (c'est à dire., attention and
dormir) and was not included in this study.
Composite Score
The percentage of misperceptions on the BPP (“indirect” measure of VH) and the total score
on the PsycH-QA (“direct” measure of hallucinatory behavior) were standardized and then
summed to create a composite score that reflected the severity of hallucinatory behavior (ici-
after referred to as the hallucination severity score, HSS). The HSS was correlated with the
demographic variables using parametric or nonparametric correlations depending on the dis-
tribution of the variables and was used as a correlate in the imaging analysis.
MRI Acquisition
All participants underwent magnetic resonance imaging (IRM) using a 3-Tesla General Elec-
tric Discovery MR750 scanner (GE Medical Systems) with an 8-channel phased array head
coil. Diffusion-weighted images (DWI) were obtained by using echo-planar imaging sequences
avec 61 different motion-probing gradient directions (TR/TE: 7,025/80 ms, 55 transverse slices,
slice thickness: 2.5 mm, matrice: 256 × 256, FOV: 240 × 240 mm). The effective diffusion
2
weighting was b = 1,000 s/mm
)
were obtained at the beginning of each diffusion sequence. 3-D T1-weighted, anatomical im-
ages were obtained (TR/TE/TI: 7.2/2.7/450 ms, voxel size 1 × 1 × 1 mm, 196 transverse slices,
256 × 256 matrice, FOV: 256 × 256 mm, flip angle 12
). The 3-D T1 images were used for
individual registration between T1-weighted anatomical and the DWI images and cortical
parcellation using FreeSurfer (version 5.3; http://surfer.nmr.mgh.harvard.edu).
, and four volumes with no diffusion weighting (b = 0 s/mm
◦
2
Neurosciences en réseau
524
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
Tractography:
Three-dimensional representation of
the brain’s white matter tracts derived
from diffusion MRI data.
Network-based statistic:
A statistical method to identify
connections and networks
comprising the connectome
associated with behavior.
Diffusion Tensor Imaging Preprocessing and Deterministic Fiber Tracking
DTI preprocessing was performed using the FMRIB Software Library (FSL, http://fsl.fmrib.
ox.ac.uk). The preprocessing steps were as follows: (un) DTI images were corrected for sus-
ceptibility, head motion, and eddy current–induced geometrical distortions using FSL’s tool
eddy; (b) a binary brain mask was created using bet; (c) images were realigned using a rigid
body registration to the b = 0 image; alors (d) a tensor was fitted in each voxel (Chang, Jones, &
Pierpaoli, 2005), suivi de (e) the computation of the fractional anisotropy (FA) level based
on the eigenvalues for each voxel, in order to determine the preferred diffusion direction within
a voxel. FA thus serves as a surrogate measure of white matter integrity, with lower levels of
FA reflecting reduced white matter integrity (Sun et al., 2003; van den Heuvel & Sporns, 2011;
Verstraete et al., 2010). The preferred diffusion direction information was then used to recon-
struct the white matter tracts of the brain using a deterministic tracking approach based on
the fiber assignment by continuous tracking (FACT) algorithme (Mori, Crain, Chacko, & van
Zijl, 1999). Deterministic tractography yields less false positive tracts compared with proba-
bilistic methods (Bastiani, Shah, Goebel, & Roebroeck, 2012). False positives are detrimental
in network modularity as they occur more prevalently between than within modules (Zalesky
et coll., 2016). A streamline was started from eight seeds within each voxel of the brain (gray
and white matter) following the main diffusion direction of the voxel and stopped when (un)
the FA value < 0.1; (b) the traced fiber made a turn > 45◦
; ou (c) the tract left the brain mask.
The images were acquired when reverse phase-encoding direction approaches were not the
standard procedure within acquisition protocols, which could have influenced the registra-
tion of diffusion and anatomical images. Donc, anatomically constrained tractography was
not applied (Forgeron, Tournier, Calamante, & Connelly, 2012). The atlas presented by Cammoun
et autres. (2012) was used, y compris 219 cortical regions and 14 subcortical regions. Le pondéré
brain network was calculated for each participant, and consistency thresholding at 50% était
appliqué (c'est à dire., including the tracts found in 50% of the patients; de Reus & van den Heuvel,
2013). The mean density of the thresholded group matrix was 8.7%. To verify the results were
not skewed by the choice of threshold, we also applied the thresholding method that retained
most consistent edges across subjects but controlling for their distance (c'est à dire., the consistency of
edges within “bins” based on their length to avoid preferential retention of short edges; Misic
et coll., 2015). The mean density of the group matrix using this threshold was 13.2%.
Network-Based Statistic
A network-based statistic (NBS) analysis was applied to investigate whether the HSS was asso-
ciated with altered connectivity strength in an interconnected subnetwork of the brain (Zalesky,
Fornito, & Bullmore, 2010). NBS is a nonparametric method for connectome-wide analysis,
which aims to detect specific pairs of brain regions showing a significant effect of interest, alors que
controlling for family-wise error (FWE) rate. Surtout, no inferences of individual connec-
tions are made; instead the null hypothesis can only be rejected at the subnetwork level.
En tant que tel, NBS is similar to the cluster-based multiple-comparison approaches used in stan-
dard functional MRI analysis. To identify changes in subnetworks associated with the HSS, le
t statistic was set at 1.7, determined using the critical value of the t distribution for our sam-
ple size (Field, 2009). En plus, we verified the robustness of the results by controlling for
disease severity using the MDS-UPDRS III (motor part) as covariate. Connections were deemed
significant at FWE-corrected p value < 0.05 (one-sided) using 5,000 permutations.
To investigate whether the subnetwork involved particular functional networks, we inves-
tigated whether nodes in the subnetwork that correlated with the HSS overlapped with previ-
ously defined resting-state networks. To this end, seven canonical resting-state networks from
Network Neuroscience
525
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
a
_
0
0
0
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
the Yeo et al. (2011) atlas were overlaid with the structural parcellation and the percentage of
nodes from each network included within the structural subnetwork that inversely related to
HSS was calculated for each resting-state network. To analyze whether this overlap occurred
significantly above chance, we randomly permuted the resting-state network identity of each
region (5,000 iterations) and used the overlap between the randomized vector and the original
node assignment to populate a null distribution. To test whether each individual resting-state
network overlapped with the significant subnetwork, their overlap was compared with the null
distributions. A resting-state network was identified as targeted if the true overlap was more
than the 97.5th percentile of null distribution (i.e., the top 2.5%). A network was considered
not to be associated with the HSS if the overlap was less than the 2.5th percentile of the null
distribution.
Graph Theoretical Analysis
The graph organizational measures were computed using the Brain Connectivity Toolbox (http://
www.brain-connectivity-toolbox.net; Rubinov & Sporns, 2010). The thresholded, weighted
brain networks were then partitioned into modules, which are nonoverlapping groups of highly
connected nodes that are only sparsely connected with other modules, using the Louvain algo-
rithm (Rubinov & Sporns, 2010). To account for the stochastic nature of the Louvain algorithm,
a consensus partition was identified by calculating the module assignment for each node 500
times. To define an appropriate value for the resolution parameter (γ), the Louvain algorithm
was iterated 100 times across a range of values (0.5–2.0 in steps of 0.1) of the group mean con-
nectivity matrix and then the similarity of the resultant partitions was estimated using mutual
information. The γ parameter of 1.9 provided the most robust estimates of topology across the
iterations and was used to determine the optimal resolution of the network modularity.
After the nodes were assigned to their modules, their intra- and intermodular connectivity
were calculated. Intramodular connectivity was calculated using the module degree z-score Wi
(see Equation 1), in which a positive score reflects high within-module connections (compared
with the node’s average number of connections), and negative z-scores denote the opposite. In-
termodular connectivity was calculated using the participation coefficient Bi (see Equation 2).
Low Bi values indicate few between-module connections, whereas high Bi values indicate
uniformly distributed connections across modules (Hall, 2018). High Wi and high Bi scores
are not mutually exclusive (Guimerà & Nunes Amaral, 2005).
Wi =
κi − κsi
σκsi
(1)
Module degree z-score:
A measure of diversity of
intramodular connections of
individual nodes.
Participation coefficient:
A measure of diversity of
intermodular connections of
individual nodes.
Equation 1: Module degree z-score Wi, where κi is the strength of the connections of region i
to other regions in its module si, κsi is the average of κ over all the regions in si, and σκsi is the
standard deviation of κ in si.
Bi = 1 − ∑nM
s=1
(cid:2)
(cid:3)2
κis
κi
(2)
Equation 2: Participation coefficient Bi, where κ
is is the strength of the positive connections
of region i to regions in module s, and κ
i is the sum of strengths of all positive connections of
region i. The participation coefficient of a region is therefore close to 1 if its connections are
uniformly distributed among all the modules and 0 if all of its links are within its own module.
To test whether nodes within the subnetwork identified using the NBS analysis differed
from nodes not included in the subnetwork, the average Wi and Bi of the subnetwork were
contrasted against the average Wi and Bi of the nodes not included in the subnetwork using
nonparametric permutation testing.
Network Neuroscience
526
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
a
_
0
0
0
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
Diverse club:
A subset of nodes in a network with
high participation coefficients that
exhibit properties consistent with an
integrative network function.
To test whether the HSS correlated with the Wi and Bi nodes across the whole-brain con-
nectome, a Spearman’s rho correlation was performed followed by a nonlinear permutation
test using 5,000 iterations to control for multiple comparisons (Nichols & Holmes, 2002), using
an alpha of 0.05. This approach was repeated using the different threshold (Misic et al., 2015),
and the outcome was correlated to the Wi and Bi using the original threshold. Both the Wi and
the Bi scores calculated using the aforementioned threshold highly correlated with the Wi
and Bi scores calculated with the consensus threshold (r = 0.92 and r = 0.94, respectively),
indicating that the results were not biased by the chosen thresholding method.
Diverse Club Analysis
We identified the “diverse club” of the network, which comprised the top 20% of Bi nodes
(Bertolero, Yeo, & D’Esposito, 2017). These nodes play an important role in network inte-
gration, and changes to these nodes could affect between-module communication (Bertolero
et al., 2017). We normalized the diverse club coefficient in reference to a null model: A random
vector with a preserved modular structure was created by randomizing the mean participation
coefficient of each node for 5,000 iterations. The diverse club was identified as those regions
with a participation coefficient greater than the 95th percentile of the permuted distribution.
We investigated whether the number of diverse club nodes was significantly higher within
the subnetwork associated with the HSS, compared with nodes that were not included in the
subnetwork identified using the NBS analysis.
RESULTS
Demographics
Table 1 presents the descriptive variables of the 29 patients. The mean percentage of misper-
ceptions on the BPP was 18.48 (range: 0–49), and the mean score on the PsycH-QA was 9.48
(range: 0–34, max score = 52), highlighting a diverse range of hallucinatory behavior in the
patient cohort. The PsycH-QA and the BPP score showed a positive, significant correlation
(r = 0.52, p = 0.004). Finally, to verify the severity score to the “gold standard,” we corre-
lated the HSS in a large cohort of patients with PD and Lewy body dementia (n = 75) to the
MDS-UPDRS item 2 and found a correlation of r = 0.53 (p < 0.001). However, given higher
construct validity (Shine, Mills, et al., 2015), we opted to include the PsycH-QA and BP scores
in the composite score (HSS) for the remainder of our analysis.
Table 1. Demographics and clinical variables
Mean (range)
66.8 ± 8 (51 – 84)
5.8 ± 4 (1.2–16)
617.8 ± 392 (125–1548)
Variable
Age (y)
Duration (y)
LEDD
H & Y
MDS-UPDRS III
MMSE
TMTB−A
BPP % misperceptions
PsycH-QA
LEDD = levodopa equivalence daily dose; MDS-UPDRS III = motor part of the Movement Disorder
Society Unified Parkinson’s Disease Rating Scale; H & Y = Hoehn and Yahr; MMSE = Mini-Mental
= Trail Making Test Part B – Part A; BPP = bistable percept paradigm;
State Examination; TMTB−A
PsycH-QA
Correlation with HSS r (p value)
0.18 (0.345)
−0.05 (0.785)
−0.06 (0.767)
0.01 (0.949)
0.29 (0.125)
−0.06 (0.762)
0.33 (0.080)
−
−
1.9 ± 0.5 (1–3)
30.0 ± 14 (7–55)
28.5 ± 2 (25–30)
78.9 ± 65 (−1−123)
18.5 ± 17 (0–49)
9.5 ± 8 (0–34)
= Psychosis and Hallucinations Questionnaire, Part A.
Network Neuroscience
527
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
a
_
0
0
0
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
The HSS showed a positive correlation trending towards significance with the TMTB−A
(r = 0.33, p = 0.08). No significant correlations were observed between the HSS and other
demographic and clinical variables.
The HSS Correlated With Decreased Connectivity in a Large Subnetwork
As illustrated in Figure 2, the NBS analysis revealed a subnetwork comprising 183 edges (8%
of the edges in the thresholded connectivity matrix) and 127 nodes with reduced FA-based
connectivity strength correlated to the HSS (p < 0.05). Using disease severity as a covariate, the
NBS analysis revealed a similar subnetwork comprising 177 edges and 135 nodes that showed
a correlation with the HSS that was trending towards significance (p = 0.059). The effects
presented with a fairly liberal threshold, suggesting the changes related to the HSS are subtle
yet topological extended (Zalesky et al., 2010). The size of networks identified using a range
of t statistics are presented in the Supporting Information (Hall et al., 2019). No significant
subnetwork was identified in the opposite direction (positive correlation between the HSS
and connectivity strength). Furthermore, the group average Bi score within the subnetwork
was 0.506, which was significantly higher (p < 0.05) than nodes outside this network, which
show a group average of 0.310. The group average Wi score of the nodes within the network
was higher than the group average Wi score of nodes outside the network (0.149 and −0.178,
respectively), yet this difference did not remain significant when controlling for disease severity
(p = 0.104).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
a
_
0
0
0
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. NBS analysis reveals a subnetwork, comprising 183 edges and 127 nodes with reduced
connectivity strength correlated to increased HSS (p < 0.05). Figure visualized with BrainNet Viewer
(Xia, Wang, & He, 2013).
Network Neuroscience
528
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
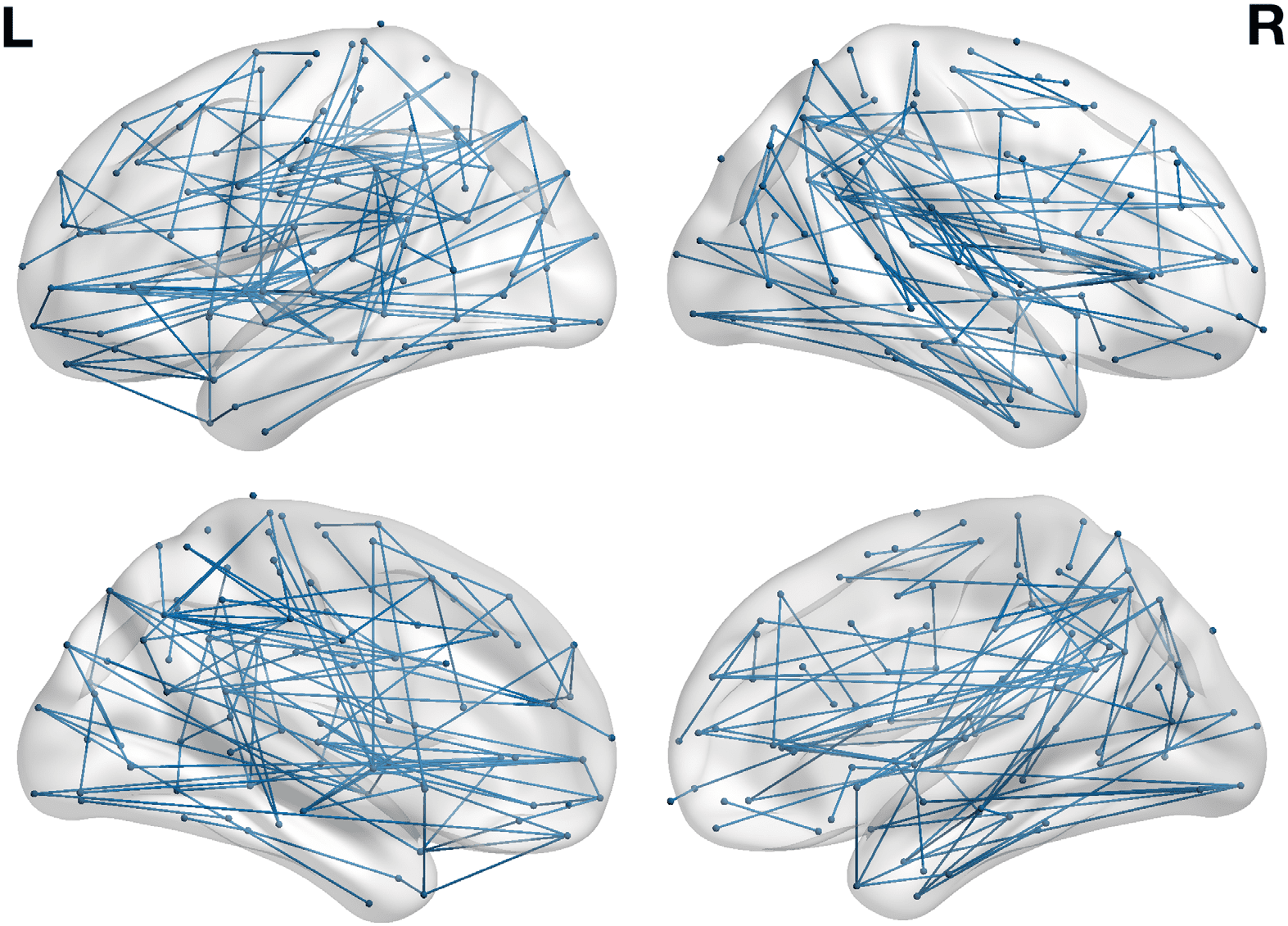
Figure 3. Overlap between the identified structural subnetwork and functional resting-state net-
works. (A) The percentage of nodes included in the subnetwork for each resting-state network.
DMN = default mode network; VAN = ventral attentional network; DAN = dorsal attentional net-
work. (B) The functional resting-state networks of the Yeo et al. (2011) atlas.
The Subnetwork Includes All Subcortical Nodes but Did Not Target a Specific Cortical
Resting-State Network
The subnetwork that showed decreased connectivity strength correlated with the HSS included
all 14 subcortical nodes (p < 0.05). As illustrated in Figure 3, the subnetwork further included
nodes across the cortex. However, none of the other the resting-state networks were signifi-
cantly correlated with the HSS (p > 0.05), yet the somatomotor network was relatively spared
(p < 0.05).
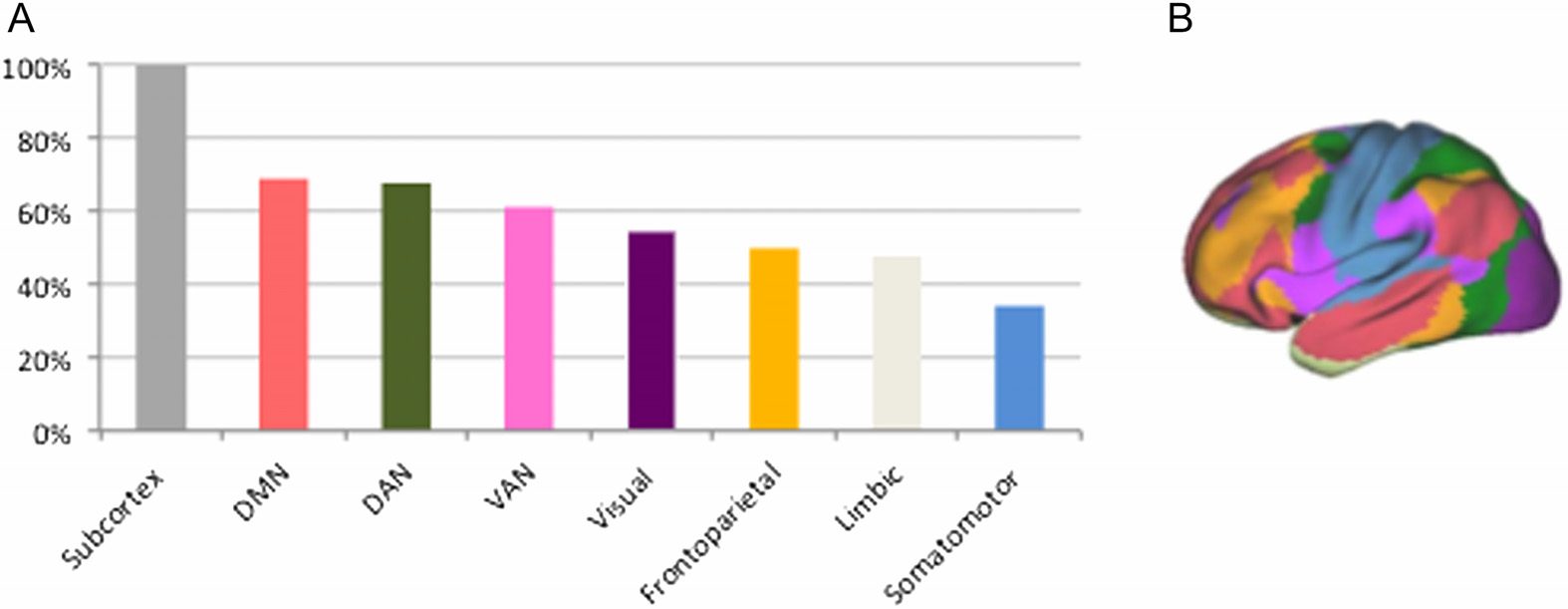
Nodes in the Subnetwork Show High Participation Scores
Eighteen nodes were included in the diverse club (see Supporting Information, Hall et al.,
2019). Seventeen of the eighteen nodes (94%) of the diverse club were included in the afore-
mentioned subnetwork, which was deemed significantly above chance (p < 0.001). As
illustrated in Figure 4, nodes with high participation coefficients were more often part of the
subnetwork.
The HSS Correlated With Wi and Bi Scores
When investigating the whole structural connectome, the HSS positively correlated to regional
Bi (i.e., higher participation scores were associated with higher severity values) for nodes in
the left medial OFC, a node in the right anterior and left posterior cingulate, precuneus, and
the caudal middle frontal gyrus. Furthermore, nodes in the right occipital, pars orbitalis, and
insula showed negative correlations between the HSS and participation coefficient (i.e., lower
participation scores were associated with higher scores on the HSS; see Table 2 and Figure 5).
However, when controlling for disease severity, the insula and medial OFC were only trending
towards significance, while the lateral occipital cortex and anterior posterior cingulate did not
remain significant.
Increased HSS scores were further associated with increased module degree z-scores in the
right thalamus, bilateral lingual, left medial OFC, pars opercularis, supramarginal gyrus, and
superior temporal cortices. The right lateral occipital cortex also showed a positive correlation,
but this did not remain significant after controlling for disease severity. Decreased HSS scores
were associated with increased module degree z-scores in the bilateral precuneus, left parts
triangularis, rostral middle frontal, and superior parietal cortex. The right pericalcarine and
Structural connectome:
Brain connectivity graphs obtained
from diffusion MRI and tractography.
Network Neuroscience
529
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
a
_
0
0
0
7
8
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
a
_
0
0
0
7
8
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Nodes ranked according to the Bi scores. Blue: nodes included in the subnetwork; gray:
nodes not included in the subnetwork correlated to the HSS.
Table 2. Spearman’s rho correlation between the participation coefficient and the HSS (p < 0.05;
permutation test)
X
Y
Z
Rho
Rho*
Subnetwork
Node
Positively correlated
Frontal
ctx-lh-medialorbitofrontal_2
ctx-rh-caudalmiddlefrontal_2
Parietal
ctx-lh-precuneus_2
Cingulate
ctx-lh-posteriorcingulate_2
ctx-rh-superiorfrontal_3
Negatively correlated
Frontal
ctx-rh-parsorbitalis_1
Occipital
ctx-rh-lateraloccipital_5
Insular
ctx-rh-insula_1
* Rho after controlling for disease severity;
right hemisphere.
−5
40
33
15
−20
39
0.37
0.43
#
0.35
0.53
−10
−44
−8
11
−43
40
46
21
40
0.40
0.37
0.57
0.36
0.53
0.31ˆ
43
−10
−0.46
−0.46
−73
2
−0.39
−0.21ˆ
43
47
36
−18
# p < 0.1; ˆ p > 0.1. ctx-lh = left hemisphere; ctx-rh =
−0.40
−0.32
13
(cid:2)
#
(cid:2)
−
(cid:2)
(cid:2)
(cid:2)
(cid:2)
(cid:2)
Neurosciences en réseau
530
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
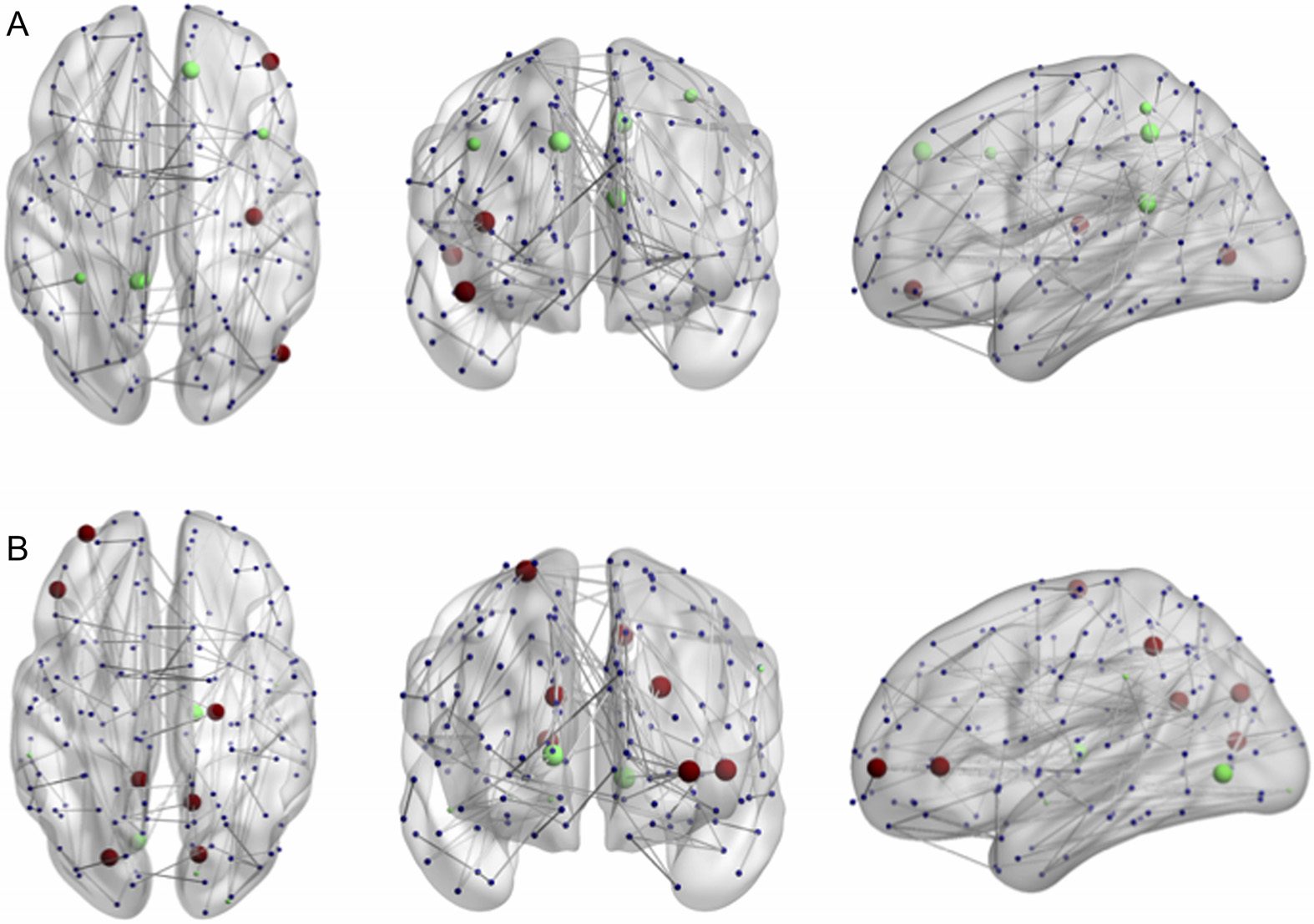
Chiffre 5. Significant changes in between- and within-module scores associated with the HSS.
(UN) Nodes with a significant correlation between participation coefficients and the HSS. (B) Nodes
with a significant correlation between the module degree z-score and HSS. Green indicates a posi-
tive correlation, red indicates a negative correlation. Larger nodes are part of the subnetwork. Chiffre
visualized with BrainNet Viewer (Xia et al., 2013).
precentral gyrus also a showed negative correlation with the HSS, yet this did not remain
significant after controlling for disease severity (see Table 3 and Figure 5).
DISCUSSION
The aim of this study was to determine whether changes in structural network topology were
associated with hallucinatory behavior in PD. We showed that severity of hallucinatory behav-
ior was negatively correlated with connectivity across a bilateral subnetwork. Regions within
this subnetwork showed higher participation compared with regions outside this network. Le
loss of connectivity strength may force the system to adapt and reroute information across less
efficient pathways, impeding the standard sensory integration process. Surtout, 94% of the
nodes in the diverse club were included in this subnetwork. This community of high participa-
tion nodes is thought to control the integration of relatively segregated regions (Bertolero et al.,
2017). En effet, the diverse connectivity pattern of these nodes makes them crucial for the func-
tional coordination of brain regions during tasks, and activity in these nodes predicts changes
in the coupling of other regions (Bertolero et al., 2017). Severity of hallucinatory behavior may
thus be the result of impaired integration and segregation of brain networks or “modules,»
affecting effective information transfer. Enfin, we showed regional changes in participation
associated with hallucination severity (the HSS score), with a positive correlation between par-
ticipation scores in the medial OFC, cingulate, precuneus, and middle frontal gyrus and the
HSS and negative correlation with participation scores in the lateral occipital cortex, pars or-
bitalis, and insula. These findings suggest a reweighting of the regions along the perceptual
hierarchy, which may give rise to hallucinations.
Neurosciences en réseau
531
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
Tableau 3. Spearman’s rho correlation between the module degree z-score and the HSS (p < 0.05; permutation test) X Y Z Rho Rho* Subnetwork Node Positively correlated Subcortical Right-Thalamus-Proper Frontal ctx-lh-medialorbitofrontal_1 ctx-lh-parsopercularis_2 Parietal ctx-lh-supramarginal_3 Temporal ctx-rh-superiortemporal_4 Occipital ctx-rh-lateraloccipital_3 ctx-lh-lingual_2 ctx-rh-lingual_1 Negatively correlated Frontal ctx-lh-parstriangularis_1 ctx-lh-rostralmiddlefrontal_5 ctx-rh-precentral_6 Parietal ctx-lh-precuneus_2 ctx-lh-superiorparietal_6 ctx-rh-precuneus_2 Occipital ctx-rh-pericalcarine_1 13 −17 7 0.49 0.45 −7 −47 52 14 −11 15 0.40 0.41 0.43 0.44 −55 −34 35 0.45 0.53 46 −6 −11 0.57 0.47 −94 −70 −83 33 56 −17 −44 −76 −54 −6 0 −7 2 2 67 46 29 26 0.39 0.42 0.41 −0.50 −0.57 −0.39 −0.46 −0.41 −0.39 # 0.34 0.50 0.37 −0.43 −0.50 −0.35 # −0.43 −0.47 −0.43 27 −10 14 −43 −31 22 −10 −22 12 15 (cid:2) − − − − − (cid:2) − (cid:2) (cid:2) (cid:2) (cid:2) (cid:2) (cid:2) * Rho after controlling for disease severity; right hemisphere. −75 # p < 0.1; ˆ p > 0.1. ctx-lh = left hemisphere; ctx-rh =
−0.24ˆ
−0.38
12
(cid:2)
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Lower participation of the lateral occipital cortex may reflect reduced early visual process-
ing, resulting in ineffective accumulation of visual information from the environment. Previous
work using a Bayesian drift diffusion model has demonstrated that accumulation speed and
quality of perceptual information are reduced in PD patients with VH (O’Callaghan et al.,
2017). En outre, reduced quality or integration of visual stimuli may increase perceptual
uncertainty, a suggestion that aligns with increased participation in the dorsal anterior cingu-
late cortex (Keri, Decety, Roland, & Gulyas, 2004). Perceptual uncertainty may place excessive
emphasis on top-down prediction centers, which subsequently could lead to a reduced activity
in early visual regions (Meyer & Olson, 2011). Cependant, the reduced participation score in
the occipital and anterior cingulate cortex may not be specific to hallucinatory behavior, comme
the negative correlations did not remain significant after controlling for disease severity.
This emphasis on top-down visual processing centers is supported by the increased partici-
pation coefficient and module degree z-score in the medial OFC. The OFC has an integrative
function across brain networks, as evidenced by its high participation coefficient. En plus,
this region is thought to facilitate recognition during visual perception by integrating incom-
ing sensory information with previous experiences and expectations (Panichello, Cheung, &
Bar, 2012). During typical visual perception, the OFC is activated early in response to vi-
sual stimuli, receiving low spatial frequency signals from the visual cortex (Bar et al., 2006).
Notably, only stimuli resembling known objects are shown to activate the OFC, which in turn
Neurosciences en réseau
532
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
generates a semantic association and provides a predictive signal
to the visual system
(Chaumon, Kveraga, Barrett, & Bar, 2014). Inversement, visual stimuli that carry no meaningful
association do not activate the OFC in healthy individuals. Ainsi, it could be speculated that
because of decreased quality of visual input, inappropriate recruitment of the OFC occurs,
which may result in ascribing false associative information to visual stimuli.
The manifestation of VHs has previously been recognized as a dysfunction between the
attentional networks (Shine, Muller, et coll., 2015). Specifically, patients with VHs are shown to
be less able to recruit the dorsal attentional network (DAN), which enables the selection of
appropriate sensory stimuli (Corbetta, Patel, & Shulman, 2008). With reduced control of this
réseau, ambiguous stimuli might instead be interpreted by the ventral attentional network
(VAN) and the DMN, which are less well equipped for this task. Our results showed increased
participation in the posterior cingulate cortex (PCC), a key hub of the DMN. PCC activity has
been implicated in regulating the focus of attention, specifically the shift from the external
world into internal mentation (Weissman, Roberts, Visscher, & Woldorff, 2006). En outre,
the PCC is involved in mind wandering and supports internally directed cognition (Leech &
Sharp, 2013). A failure to suppress PCC activity may lead to the intrusion of internal thoughts
into task performance (Sonuga-Barke & Castellanos, 2007). De plus, a positive correlation
was found between the HSS and the module degree z-score of the left pars opercularis, un
node in the VAN, a network that is activated when expectations in perception are violated
(Corbetta & Shulman, 2002; Shine, Halliday, et coll., 2014). Inversement, a negative correlation
between the HSS scores and module degree score and participation coefficients was found
in other nodes of the VAN, namely the left pars triangularis, the right pars orbitalis, et en-
sula. The left pars triangularis supports resolving competition between simultaneously active
representations (Badre & Wagner, 2007), while the insula plays an important role in dynami-
cally shifting attention between the attentional control networks (Menon & Uddin, 2010). Le
anterior insula has previously been shown to be reduced in volume in PD patients with VH
(Ibarretxe-Bilbao et al., 2010; Shine, Halliday, et coll., 2014). Ensemble, these results suggest that
ineffective communication between attentional networks in the brain may predispose an in-
dividual to hallucinate. Surprisingly, the participation coefficient of a node within the DAN
(“ctx-rh-caudalmiddlefrontal_2”) showed a positive correlation with the HSS. This node was
not part of the subnetwork, and it may be possible that this is a compensatory response to
the loss of connectivity strength in the other DAN regions. Notably, the connectivity matrix
shows between module connections of this region with nodes in the somatomotor and the
frontoparietal network, but not with the DMN or VAN.
Enfin, all nodes that showed negative correlations with the HSS were included in the
subnetwork. Decreased within-module scores were found across the prefrontal and the
somatosensory association cortex, as well as in the primary visual cortex, while there was
a positive correlation between the HSS and the bilateral secondary visual cortex, perhaps as a
result of the decreased visual input from V1. En plus, the supramarginal gyrus, a node that
has been shown to be functionally active during spatial perception but also during visual im-
agery (Ganis, Thompson, & Kosslyn, 2004), showed an increased module degree z-score with
increasing severity of VHs. En outre, a positive correlation with the HSS and the module
degree score in the superior temporal cortex, a region involved in auditory processing, était
trouvé. It could be speculated that increased visual uncertainty may stimulate other sensory
processing areas. Inversement, previous work in schizophrenia has shown increased activation
in the superior temporal cortex during a mismatch between expected and perceived auditory
feedback (Fu et al., 2008). The HSS also included hallucinations of other sensory modali-
liens, and it could be speculated that a greater emphasis is being placed on predictions of
Neurosciences en réseau
533
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
auditory stimuli, as reflected as increased within-module connectivity in the superior temporal
cortex.
This study has several limitations worth noting. D'abord, the DWI data were acquired without
EPI distortion correction. This may have affected the accuracy of registration between DWI and
T1 images in the frontal and temporal cortices. Because of relatively low diffusion weighting
used in the current MRI protocol, it was chosen to employ DTI rather than more sophisticated
methods such as constrained spherical deconvolution, an algorithm that more adequately deals
with multiple fiber directions within one voxel than DTI. En outre, after controlling for dis-
ease severity, the identified subnetwork was only trending towards significance. The correlation
between the HSS and the participation coefficients of the lateral occipital and anterior cingulate
did not remain significant, while the OFC and insula were only trending towards significance,
and similar patterns were found for the module degree z-score: correlation with the HSS and
nodes in the occipital and precentral regions did not remain significant. Cependant, this may
be expected given the close relationship between overall symptom severity and phenotypic
symptom severity. Surtout, the correlation coefficients showed notable consistency across
the analyses, suggesting the HSS was able to distinguish interindividual variability. While the
current study included PD patients with no self-reported hallucinations (n = 6), no data from a
healthy control group were acquired, which could have aided in disassociating the significant
network measures and disease burden. The LEDD score was not associated with the HSS in
the current study, and the relation between hallucinations and a LEDD-related side effect has
been dismissed in previous work (Fénelon et al., 2000; Graham, Grünewald, & Sagar, 1997;
Sanchez-Ramos, Ortollm, & Paulson, 1996; Shergill, Walker, & Le Katona, 1998). En outre,
hallucinations may occur in the absence of dopaminergic treatment in PD or dementia with
Lewy bodies (Ala, Lequel, Sung, & Frey, 1997). Donc, the current study chose not to include
LEDD score as a covariate. En outre, the investigation was conducted in a relatively small
group of PD patients, and future studies should replicate our findings in a larger sample size.
Enfin, this study chose to include the PsycH-Q to assess hallucinations. This is an in-depth
questionnaire with high construct validity; cependant, patients have to report their hallucinations
themselves. Ideally, a trained neurologist or qualified researcher, who can probe the patients
when in doubt, should assist with this process.
CONCLUSIONS
We cautiously conclude that hallucinatory behavior in PD patients is associated with marked
alterations in structural network topology. Severity of hallucinatory behavior was associated
with decreased connectivity in a large subnetwork that included the majority of the diverse
club. These changes may result in an inefficient rerouting of information across less efficient
pathways, which may lead to impaired visual integration processes. En outre, nodes within
the orbitofrontal cortex and temporal lobes showed increased participation scores, tandis que le
visual association cortex, insula, and middle frontal gyrus showed decreased scores associ-
ated with the HSS score. These findings suggest that impaired integration across different re-
gions along the perceptual hierarchy may result in inefficient transfer of information. A failure
to effectively switch between attentional networks and the intrusion of internal percepts could
give rise to perceptual glitches, such as misperceptions and hallucinations.
REMERCIEMENTS
We thank the patients and their families who contribute to our research at the Parkinson’s Dis-
ease Research Clinic. We thank Dr. Váša for sharing his thresholding code (https://github.com/
Neurosciences en réseau
534
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
frantisekvasa/matlab_general). The DWI data were processed during the 10kin1day initiative at
the Dutch Connectome Lab. This research was supported by Sydney Informatics Hub, funded
by the University of Sydney.
CONTRIBUTIONS DES AUTEURS
Julie M. Hall: Conceptualisation; Conservation des données; Analyse formelle; Enquête; Methodol-
ogy; Gestion de projet; Visualisation; Rédaction – ébauche originale. Claire O’Callaghan: Données
curation; Enquête; Méthodologie; Rédaction – révision & édition. Alana Justine Muller: Données
curation; Rédaction – révision & édition. Kaylena A. Ehgoetz Martens: Rédaction – révision & edit-
ing. Joseph R. Phillips: Rédaction – révision & édition. Ahmed A. Moustafa: Rédaction – révision &
édition. Simon J. G. Lewis: Acquisition de financement; Enquête; Surveillance; Rédaction – révision
& édition. James M.. Shine: Conceptualisation; Conservation des données; Analyse formelle; Enquête;
Méthodologie; Surveillance; Rédaction – révision & édition.
INFORMATIONS SUR LE FINANCEMENT
Claire O’Callaghan, National Health and Medical Research Council Neil Hamilton Fairley
Fellowship, Australia, Award ID: 1091310. Simon J. G. Lewis, National Health and Medical
Research Council of Australia, Award ID: 1037746. Simon J. G. Lewis, National Health and
Medical Research Council of Australia, Award ID: 1095127. Simon J. G. Lewis, National
Health and Medical Research Council of Australia and the Australian Research Council, De-
mentia Fellowship, AWARD ID: 1110414. James M.. Shine, National Health and Medical Re-
search Council Project Grant, Australia, Award ID: 1156536. James M.. Shine, The University
of Sydney Robinson Fellowship.
RÉFÉRENCES
Aarsland, D., Bronnick, K., Ehrt, U., De Deyn, P.. P., Tekin, S., Emre,
M., & Cummings, J.. L. (2007). Neuropsychiatric symptoms in pa-
tients with Parkinson’s disease and dementia: Frequency, profile
and associated care giver stress. Journal of Neurology, Neuro-
surgery, and Psychiatry, 78(1), 36–42. https://doi.org/10.1136/
jnnp.2005.083113
Ala, T. UN., Lequel, K. H., Sung, J.. H., & Frey, W. H. (1997). Hallucina-
tions and signs of parkinsonism help distinguish patients with de-
mentia and cortical Lewy bodies from patients with Alzheimer’s
Journal de
disease at presentation: A clinicopathological study.
Neurologie, Neurochirurgie, and Psychiatry, 62(1), 16–21.
Badre, D., & Wagner, UN. D.
(2007). Left ventrolateral prefrontal
cortex and the cognitive control of memory. Neuropsychologie,
45(13), 2883–2901. https://doi.org/10.1016/j.neuropsychologia.
2007.06.015
Bar, M., Kassam, K. S., Ghuman, UN. S., Boshyan,
Bar, M.. (2009). The proactive braerin: Memory for predictions. Philo-
sophical Transactions of the Royal Society B: Sciences biologiques,
364(1521), 1235–1243. https://doi.org/10.1098/rstb.2008.0310
J., Schmid,
UN. M., Dale, UN. M., . . . Halgren, E. (2006). Top-down fa-
cilitation of visual recognition. Actes de la Nationale
Académie des Sciences, 103(2), 449–454. https://doi.org/10.1073/
pnas.0507062103
Barnes, J., Boubert, L., Harris, J., Lee, UN., & David, UN. S. (2003). Re-
ality monitoring and visual hallucinations in Parkinson’s disease.
Neuropsychologie, 41(5), 565–574.
Barnes, J., & David, UN. (2001). Visual hallucinations in Parkinson’s
maladie: A review and phenomenological survey. Journal of Neu-
rology, Neurochirurgie, and Psychiatry, 70(6), 727–733.
Bastiani, M., Shah, N. J., Goebel, R., & Roebroeck, UN. (2012). Hu-
man cortical connectome reconstruction from diffusion weighted
IRM: The effect of tractography algorithm. NeuroImage, 62(3),
1732–1749. https://doi.org/10.1016/j.neuroimage.2012.06.002
Bertolero, M.. UN., Yeo, B. T. T., & D'Esposito, M.. (2015). The modu-
lar and integrative functional architecture of the human brain.
Actes de l'Académie nationale des sciences, 112(49),
E6798–E6807. https://doi.org/10.1073/pnas.1510619112
Bertolero, M.. UN., Yeo, B. T. T., & D'Esposito, M.. (2017). The diverse
club. Nat Commun, 8(1), 1277. https://doi.org/10.1038/s41467-
017-01189-w
Bullmore, E., & Sporns, Ô. (2009). Complex brain networks: Graph
theoretical analysis of structural and functional systems. Nature
Reviews Neuroscience, 10(3), 186.
Cammoun, L., Gigandet, X., Meskaldji, D., Thiran, J.. P., Sporns,
O., Do, K. Q., . . . Hagmann, P.. (2012). Mapping the human
connectome at multiple scales with diffusion spectrum MRI. Jour-
nal of Neuroscience Methods, 203(2), 386–397. https://est ce que je.org/
10.1016/j.jneumeth.2011.09.031
Chang, L. C., Jones, D. K., & Pierpaoli, C. (2005). RESTORE: Ro-
bust estimation of tensors by outlier rejection. Magnetic Reso-
nance in Medicine, 53(5), 1088–1095. https://doi.org/10.1002/
mrm.20426
Neurosciences en réseau
535
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
Chaumon, M., Kveraga, K., Barrett, L. F., & Bar, M.. (2014). Vi-
sual predictions in the orbitofrontal cortex rely on associa-
tive content. Cortex cérébral, 24(11), 2899–2907. https://est ce que je.org/
10.1093/cercor/bht146
Collerton, D., Perry, E., & McKeith, je. (2005). Why people see things
that are not there: A novel Perception and Attention Deficit model
for recurrent complex visual hallucinations. Behavioral and Brain
les sciences, 28(6), 737—757; discussion 757–794. https://est ce que je.org/
10.1017/S0140525X05000130
Corbetta, M., Patel, G., & Shulman, G. L. (2008). The reorienting sys-
tem of the human brain: From environvment to theory of mind. Neu-
ron, 58(3), 306–324.https://doi.org/10.1016/j.neuron.2008.04.017
Corbetta, M., & Shulman, G. L. (2002). Control of goal-directed
and stimulus-driven attention in the brain. Nature Reviews
Neurosciences, 3(3), 201–215. https://doi.org/10.1038/nrn755
de Reus, M.. UN., & van den Heuvel, M.. P..
(2013). Estimating
false positives and negatives in brain networks. NeuroImage, 70,
402–409. https://doi.org/10.1016/j.neuroimage.2012.12.066
Diederich, N. J., Goetz, C. G., Raman, R., Pappert, E. J., Leurgans,
S., & Piery, V. (1998). Poor visual discrimination and visual hal-
lucinations in Parkinson’s disease. Clinical Neuropharmacology,
21(5), 289–295.
Diederich, N. J., Goetz, C. G., & Stebbins, G. T. (2005). Repeated
visual hallucinations in Parkinson’s disease as disturbed exter-
nal/internal perceptions: Focused review and a new integrative
model. Movement Disorders, 20(2), 130–140. https://est ce que je.org/10.
1002/mds.20308
Ange, UN. K., Fries, P., & Chanteur, W. (2001). Dynamic predictions:
Oscillations and synchrony in top-down processing. Nature Re-
views Neuroscience, 2, 704. https://doi.org/10.1038/35094565
Fénelon, G., Mahieux, F., Huon, R., & Ziégler, M.. (2000). Halluci-
nations in Parkinson’s disease: Prevalence, phenomenology and
risk factors. Cerveau, 123(4), 733–745.
Field, UN.
(2009). Discovering statistics using SPSS. Londres, ROYAUME-UNI:
SAGE Publications.
Fletcher, P.. C., & Frith, C. D. (2008). Perceiving is believing:
A Bayesian approach to explaining the positive symptoms of
schizophrenia. Nature Revues Neurosciences, 10, 48. https://
doi.org/10.1038/nrn2536
Folstein, M.. F., Robins, L. N., & Helzer, J.. E. (1983). The mini-mental
state examination. Archives of General Psychiatry, 40(7), 812.
Fu, C. H. Y., Brammer, M.. J., Yágüez, L., Allen, P., Matsumoto,
K., Johns, L., . . . van Haren, N. (2008). Increased superior
temporal activation associated with external misattributions of
self-generated speech in schizophrenia. Schizophrenia Research,
100(1), 361–363. https://doi.org/10.1016/j.schres.2007.10.023
Gallagher, D. UN., Parkkinen, L., O’Sullivan, S. S., Spratt, UN., Shah,
UN., Davey, C. C., . . . Lees, UN. J.. (2011). Testing an aetiologi-
cal model of visual hallucinations in Parkinson’s disease. Cerveau,
134(11), 3299–3309.
Ganis, G., Thompson, W. L., & Kosslyn, S. M.. (2004). Brain areas
underlying visual mental imagery and visual perception: An fMRI
étude. Cognitive Brain Research, 20(2), 226–241. https://est ce que je.org/
10.1016/j.cogbrainres.2004.02.012
Goetz, C. G. (2009). Scales to evaluate psychosis in Parkinson’s
maladie. Parkinsonism and Related Disorders, 15(Supplément. 3),
S38–41. https://doi.org/10.1016/S1353-8020(09)70777-1
Goetz, C. G., Tilley, B. C., Shaftman, S. R., Stebbins, G. T., Fahn, S.,
Martinez-Martin, P., . . . Dodel, R.. (2008). Movement Disorder
Society–sponsored revision of the Unified Parkinson’s Disease
Rating Scale (MDS-UPDRS): Scale presentation and clinimetric
testing results. Movement Disorders, 23(15), 2129–2170.
Graham, J.. M., Grünewald, R.. UN., & Sagar, H. J.. (1997). Hallucinosis
in idiopathic Parkinson’s disease. Journal of Neurology, Neuro-
surgery, and Psychiatry, 63(4), 434–440.
Guimerà, R., & Nunes Amaral, L. UN. (2005). Functional cartogra-
phy of complex metabolic networks. Nature, 433, 895. https://
doi.org/10.1038/nature03288
Hall, J.. M.. (2018). Topology: Structural network topology code for
“Alterations in structural network topology contribute to freez-
ing of gait in Parkinson’s disease,” GitHub. https://github.com/
juliemaehall/topology
Hall, J.. M., O’Callaghan, C., Muller, UN. J., Ehgoetz Martens, K. UN.,
(2019). Sup-
Phillips, J.. R., Moustafa, UN. UN., . . . Shine, J.. M..
porting information for “Changes in structural network topology
correlate with severity of hallucinatory behavior in Parkinson’s
disease.” Network Neuroscience, 3(2), 521–538. https://est ce que je.org/
10.1162/netn_a_00078
Hall, J.. M., O’Callaghan, C., Shine, J.. M., Muller, UN. J., Phillips, J.. R.,
Walton, C. C., . . . Moustafa, UN. UN. (2016). Dysfunction in atten-
tional processing in patients with Parkinson’s disease and visual
hallucinations. Journal of Neural Transmission (Vienna), 123(5),
503–507. https://doi.org/10.1007/s00702-016-1528-3
Hepp, D. H., Foncke, E. M.. J., Berendse, H. W., Wassenaar, T. M.,
Olde Dubbelink, K. T. E., Groenewegen, H. J., . . . Schoonheim,
M.. M..
(2017). Damaged fiber tracts of the nucleus basalis of
Meynert in Parkinson’s disease patients with visual hallucina-
tion. Scientific Reports, 7(1), 10112.
Hepp, D. H., Foncke, E. M.. J., Olde Dubbelink, K. T. E., van de Berg,
W. D. J., Berendse, H. W., & Schoonheim, M.. M.. (2017). Loss of
functional connectivity in patients with Parkinson disease and vi-
sual hallucinations. Radiology, 285(3), 896–903. https://est ce que je.org/
10.1148/radiol.2017170438
Hoehn, M.. M., & Yahr, M.. D. (1998). Parkinsonism: Onset, progres-
sion, and mortality. Neurologie, 50(2), 318–318.
Ibarretxe-Bilbao, N., Ramírez-Ruiz, B., Junqué, C., Martí, M.. J.,
Valldeoriola, F., Bargallo, N., . . . Tolosa, E. (2010). Differen-
tial progression of brain atrophy in Parkinson’s disease with and
Journal of Neurology, Neuro-
without visual hallucinations.
surgery, and Psychiatry, 81(6), 650–657. https://doi.org/10.1136/
jnnp.2009.179655
Intaite, M., Noreika, V., Soliunas, UN., & Falter, C. M.. (2013). Interac-
tion of bottom-up and top-down processes in the perception of
ambiguous figures. Recherche sur la vision, 89, 24–31. https://est ce que je.org/
10.1016/j.visres.2013.06.011
Keri, S., Decety, J., Roland, P.. E., & Gulyas, B. (2004). Feature uncer-
tainty activates anterior cingulate cortex. Human Brain Mapping,
21(1), 26–33. https://doi.org/10.1002/hbm.10150
Lee, J.-Y., Yoon, E. J., Lee, W. W., Kim, Oui. K., Lee, J.-Y., & Jeon, B.
(2016). Lateral geniculate atrophy in Parkinson’s with visual hal-
lucination: A trans-synaptic degeneration? Movement Disorders,
31(4), 547–554. https://doi.org/10.1002/mds.26533
Leech, R., & Sharp, D. J.. (2013). The role of the posterior cingulate
cortex in cognition and disease. Cerveau, 137(1), 12–32.
Neurosciences en réseau
536
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
Lenka, UN., Hegde, S., Arumugham, S. S., & Pal, P.. K. (2017). Pattern
of cognitive impairment in patients with Parkinson’s disease and
psychosis: A critical review. Parkinsonism and Related Disorders,
37, 11–18. https://doi.org/10.1016/j.parkreldis.2016.12.025
Matsui, H., Udaka, F., Tamura, UN., Oda, M., Kubori, T., Nishinaka,
K., & Kameyama, M..
Impaired visual acuity as a risk
factor for visual hallucinations in Parkinson’s disease. Journal de
Geriatric Psychiatry and Neurology, 19(1), 36–40. https://est ce que je.
org/10.1177/0891988705284739
(2006).
Menon, V., & Uddin, L. Q.
(2010). Saliency, switching, attention
and control: A network model of insula function. Brain Struc-
ture and Function, 214(5–6), 655–667. https://doi.org/10.1007/
s00429-010-0262-0
Meyer, T., & Olson, C. R.. (2011). Statistical learning of visual
transitions in monkey inferotemporal cortex. Actes du
Académie nationale des sciences, 108(48), 19401–19406. https://
doi.org/10.1073/pnas.1112895108
Misic, B., Betzel, R.. F., Nematzadeh, UN., Goni,
J., Griffa, UN.,
Hagmann, P., . . . Sporns, Ô. (2015). Cooperative and competitive
spreading dynamics on the human connectome. Neurone, 86(6),
1518–1529. https://doi.org/10.1016/j.neuron.2015.05.035
Mori, S., Crain, B. J., Chacko, V. P., & van Zijl, P.. C. (1999). Three-
dimensional tracking of axonal projections in the brain by mag-
netic resonance imaging. Annales de neurologie, 45(2), 265–269.
Muller, UN. J., O’Callaghan, C., Walton, C. C., Shine, J.. M., & Lewis,
S. J.. (2017). Retrospective neuropsychological profile of patients
with Parkinson disease prior to developing visual hallucinations.
Journal of Geriatric Psychiatry and Neurology, 30(2), 90–95.
https://doi.org/10.1177/0891988716686830
Muller, UN. J., Shine, J.. M., Halliday, G. M., & Lewis, S. J.. (2014).
Visual hallucinations in Parkinson’s disease: Theoretical models.
Movement Disorders, 29(13), 1591–1598.
Nichols, T. E., & Holmes, UN. P..
(2002). Nonparametric permuta-
tion tests for functional neuroimaging: A primer with examples.
Human Brain Mapping, 15(1), 1–25.
O’Callaghan, C., Hall, J.. M., Tomassini, UN., Muller, UN. J., Walpola,
je. C., Moustafa, UN. UN., . . . Lewis, S. J.. G. (2017). Visual hallu-
cinations are characterized by impaired sensory evidence ac-
cumulation: Insights from hierarchical drift diffusion modeling
in Parkinson’s disease. Biological Psychiatry: Cognitive Neuro-
science and Neuroimaging, 2(8), 680–688. https://est ce que je.org/10.
1016/j.bpsc.2017.04.007
Panichello, M.. F., Cheung, Ô. S., & Bar, M.. (2012). Predictive feed-
back and conscious visual experience. Frontiers in Psychology,
3, 620. https://doi.org/10.3389/fpsyg.2012.00620
Powers, UN. R., Kelley, M., & Corlett, P.. R.. (2016). Hallucinations as
top-down effects on perception. Biological Psychiatry: Cognitive
Neuroscience and Neuroimaging, 1(5), 393–400. https://est ce que je.org/
10.1016/j.bpsc.2016.04.003
Ramírez-Ruiz, B., Junqué, C., Martí, M.. J., Valldeoriola, F., & Tolosa,
E. (2006). Neuropsychological deficits in Parkinson’s disease pa-
tients with visual hallucinations. Movement Disorders, 21(9),
1483–1487. https://doi.org/10.1002/mds.20965
Ramírez-Ruiz, B., Martí, M.. J., Tolosa, E., Falcón, C., Bargalló, N.,
Valldeoriola, F., & Junqué, C. (2008). Brain response to complex
visual stimuli in Parkinson’s patients with hallucinations: A func-
tional magnetic resonance imaging study. Movement Disorders,
23(16), 2335–2343.
Rubinov, M., & Sporns, Ô. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003
Sanchez-Ramos, J.. R., Ortollm, R., & Paulson, G. W. (1996). Vi-
sual hallucinations associated with Parkinson disease. Archives
of Neurology, 53(23), 1265–1268.
Schrag, UN.
(2004). Psychiatric aspects of Parkinson’s disease: Un
update. Journal of Neurology, 251(7), 795–804. https://est ce que je.org/
10.1007/s00415-004-0483-3
Shergill, S. S., Walker, Z., & Le Katona, C. (1998). A preliminary in-
vestigation of laterality in Parkinson’s disease and susceptibility
to psychosis. Journal of Neurology, Neurochirurgie, and Psychia-
try, 65(4), 610–611.
Shine, J.. M., Halliday, G. H., Carlos, M., Naismith, S. L., & Lewis,
S. J.. (2012). Investigating visual misperceptions in Parkinson’s dis-
ease: A novel behavioral paradigm. Movement Disorders, 27(4),
500–505.
Shine, J.. M., Halliday, G. M., Gilat, M., Matar, E., Bolitho, S. J.,
Carlos, M., . . . Lewis, S. J.. (2014). The role of dysfunctional at-
tentional control networks in visual misperceptions in Parkinson’s
maladie. Human Brain Mapping, 35(5), 2206–2219. https://est ce que je.
org/10.1002/hbm.22321
Shine, J.. M., Mills, J.. M.. Z., Qiu, J., O’Callaghan, C., Terpening,
Z., Halliday, G. M., . . . Lewis, S. J.. G. (2015). Validation
de
the psychosis and hallucinations questionnaire in non-
demented patients with Parkinson’s disease. Movement Disor-
ders Clinical Practice, 2(2), 175–181. https://doi.org/10.1002/
mdc3.12139
Shine,
J.. M., Muller, UN.
J., O’Callaghan, C., Hornberger, M.,
Halliday, G. M., & Lewis, S. J.. (2015). Abnormal connectivity
between the default mode and the visual system underlies the
manifestation of visual hallucinations in Parkinson’s disease: UN
task-based fMRI study. NPJ Parkinson’s Disease, 1, 15003. https://
doi.org/10.1038/npjparkd.2015.3
Shine, J.. M., O’Callaghan, C., Halliday, G. M., & Lewis, S. J.. (2014).
Tricks of the mind: Visual hallucinations as disorders of attention.
Progrès en neurobiologie, 116, 58–65. https://est ce que je.org/10.1016/
j.pneurobio.2014.01.004
Forgeron, R.. E., Tournier, J.. D., Calamante, F., & Connelly, UN. (2012).
Anatomically constrained tractography:
Improved diffusion
MRI streamlines tractography through effective use of anatom-
ical information. NeuroImage, 62(3), 1924–1938. https://est ce que je.
org/10.1016/j.neuroimage.2012.06.005
Sonuga-Barke, E. J.. S., & Castellanos, F. X. (2007). Spontaneous
attentional fluctuations in impaired states and pathological
conditions: A neurobiological hypothesis. Neuroscience and
Biobehavioral Reviews, 31(7), 977–986. https://est ce que je.org/10.1016/
j.neubiorev.2007.02.005
Summerfield, C., Egner, T., Vert, M., Koechlin, E., manque, J., &
Hirsch, J.. (2006). Predictive codes for forthcoming perception in
the frontal cortex. Science, 314(5803), 1311–1314. https://est ce que je.
org/10.1126/science.1132028
Summerfield, C., & Koechlin, E. (2008). A neural representation
of prior information during perceptual inference. Neurone, 59(2),
336–347. https://doi.org/10.1016/j.neuron.2008.05.021
Neurosciences en réseau
537
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Changes in structural network topology are associated with visual hallucinations in Parkinson’s disease
Sun, Z., Wang, F., Cui, L., Breeze, J., Du, X., Wang, X., . . .
Zhang, D.
(2003). Abnormal anterior cingulum in patients
with schizophrenia: A diffusion tensor imaging study. Neuro-
Report, 14(14), 1833–1836. https://doi.org/10.1097/01.wnr.
0000094529.75712.48
Tombaugh, T. N. (2004). Trail Making Test A and B: Normative data
stratified by age and education. Archives of Clinical Neuropsy-
cologie, 19(2), 203–214.
Tourneur, T. H., Cookson, J.. C., Wass, J.. UN., Drury, P.. L., Prix, P.. UN.,
& Besser, G. M.. (1984). Psychotic reactions during treatment of
pituitary tumours with dopamine agonists. British Medical Jour-
nal (Clinical Research Ed.), 289(6452), 1101–1103.
van den Heuvel, M.. P., & Sporns, Ô. (2011). Rich-club organ-
ization of the human connectome. Journal des neurosciences,
31(44), 15775–15786.https://doi.org/10.1523/JNEUROSCI.3539-
11.2011
Verstraete, E., van den Heuvel, M.. P., Veldink, J.. H., Blanken, N.,
Mandl, R.. C., Hulshoff Pol, H. E., & van den Berg, L. H. (2010).
Motor network degeneration in amyotrophic lateral sclerosis: UN
structural and functional connectivity study. PLoS ONE, 5(10),
e13664. https://doi.org/10.1371/journal.pone.0013664
Weissman, D. H., Roberts, K., Visscher, K., & Woldorff, M.. (2006).
The neural bases of momentary lapses in attention. Nature
Neurosciences, 9(7), 971.
Xia, M., Wang, J., & Il, Oui. (2013). BrainNet Viewer: A network
visualization tool for human brain connectomics. PLoS One,
8(7), e68910. https://doi.org/10.1371/journal.pone.0068910
Yao, N., Shek-Kwan Chang, R., Cheung, C., Pang, S., Lau, K. K.,
Suckling, J., . . . Chua, S. E. (2014). The default mode network
is disrupted in Parkinson’s disease with visual hallucinations.
Human Brain Mapping, 35(11), 5658–5666.
Yeo, B. T., Krienen, F. M., Sepulcre, J., Sabuncu, M.. R., Lashkari,
D., Hollinshead, M., . . . Buckner, R.. L. (2011). The organization
of the human cerebral cortex estimated by intrinsic functional
Journal de neurophysiologie, 106(3), 1125–1165.
connectivité.
https://est ce que je.org/10.1152/jn.00338.2011
Zalesky, UN., Fornito, UN., & Bullmore, E. T. (2010). Network-based
statistique: Identifying differences in brain networks. NeuroImage,
53(4), 1197–1207. https://doi.org/10.1016/j.neuroimage.2010.
06.041
Zalesky, UN., Fornito, UN., Cocchi, L., Gollo, L. L., van den Heuvel,
M.. P., & Breakspear, M.. (2016). Connectome sensitivity or speci-
ficity: Which is more important? NeuroImage, 142, 407–420.
https://doi.org/10.1016/j.neuroimage.2016.06.035
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
5
2
1
1
0
9
2
7
0
4
n
e
n
_
un
_
0
0
0
7
8
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Neurosciences en réseau
538