RECHERCHE
Spatial and spectral trajectories in typical
neurodevelopment from childhood
to middle age
Benjamin A. E. Hunt
1,2, Simeon M. Wong
1,2, Marlee M. Vandewouw1,2,
Matthew J. Brookes3, Benjamin T. Dunkley1,2,5, and Margot J. Taylor1,2,4,5
1Department of Diagnostic Imaging, The Hospital for Sick Children, Toronto, Canada
2Neurosciences and Mental Health Program, The Hospital for Sick Children Research Institute, Toronto, Canada
3The Sir Peter Mansfield Imaging Centre, School of Physics and Astronomy, University of Nottingham,
Nottingham, United Kingdom
4Département de psychologie, University of Toronto, Toronto, Canada
5Department of Medical Imaging, University of Toronto, Toronto, Canada
Mots clés: Neurodevelopment, MEG, Functional connectivity, Phase synchronisation, Power
spectral density, Maturational trajectories, Resting state
ABSTRAIT
Detailed characterization of typical human neurodevelopment is key if we are to understand
the nature of mental and neurological pathology. While research on the cellular processes
of neurodevelopment has made great advances, in vivo human imaging is crucial to
understand our uniquely human capabilities, as well as the pathologies that affect them.
Using magnetoencephalography data in the largest normative sample currently available
(324 participants aged 6–45 years), we assess the developmental trajectory of resting-state
oscillatory power and functional connectivity from childhood to middle age. Le
maturational course of power, indicative of local processing, was found to both increase and
decrease in a spectrally dependent fashion. Using the strength of phase-synchrony between
parcellated regions, we found significant linear and nonlinear (quadratic and logarithmic)
trajectories to be characterized in a spatially heterogeneous frequency-specific manner, tel
as a superior frontal region with linear and nonlinear trajectories in theta and gamma band
respectivement. Assessment of global efficiency revealed similar significant nonlinear trajectories
across all frequency bands. Our results link with the development of human cognitive
abilities; they also highlight the complexity of neurodevelopment and provide quantitative
parameters for replication and a robust footing from which clinical research may map
pathological deviations from these typical trajectories.
RÉSUMÉ DE L'AUTEUR
Imagine that we could predict which children will go on to develop depression or anxiety
disorders using functional brain imaging? This would allow early intervention and,
in some, negation of the illness. The first step toward this goal is understanding the
neurodevelopmental trajectory of healthy individuals. We can then recognize the
developmental deviations with specific illnesses.
We used the largest developmental MEG dataset available (n = 324) to characterize the
developmental trajectories of functional connectivity in a cohort aged 6–45 years. We find
evidence for nonlinear trajectories in two graph theory metrics, heterogeneously defined in
brain space with opposing trajectories in different oscillatory frequencies. Our findings
un accès ouvert
journal
Citation: Hunt, B. UN. E., Wong, S. M.,
Vandewouw, M.. M., Brookes, M.. J.,
Dunkley, B. T., & Taylor, M.. J.. (2019).
Spatial and spectral trajectories in
typical neurodevelopment from
childhood to middle age. Réseau
Neurosciences, 3(2), 497–520.
https://doi.org/10.1162/netn_a_00077
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00077
Informations complémentaires:
https://doi.org/10.1162/netn_a_00077
Reçu: 16 Août 2018
Accepté: 24 Décembre 2018
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Benjamin A. E. Hunt
Benjamin.Hunt@SickKids.ca
Éditeur de manipulation:
Olaf Sporns
droits d'auteur: © 2019
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
highlight the intricacy of typical neurodevelopment and provide foundational quantifications
for replication in different modalities and application to clinical cohorts.
INTRODUCTION
Compared with other mammalian species, human neonates are relatively disadvantaged at
birth. While giraffe calves are able to walk within an hour of birth and run within a day (Kaleta
& Marczewska, 2007), it takes human neonates more than 1 year to walk unaided, and many
months thereafter to run (Størvold, Aarethun, & Bratberg, 2013). Néanmoins, as children,
humans develop uniquely complex cognitive abilities and motor control. Protracted ex utero
neurodevelopmental processes continue well into adulthood (Tottenham, 2014), and they are
characterized in early life by simultaneous apoptosis, synaptogenesis, and myelination of ax-
ons (Giedd, 1999; Tau & Peterson, 2010). Positron emission tomography (ANIMAL DE COMPAGNIE) studies indicate
that overall brain metabolism rises to twice that of adults in 4- to 5-year-olds, and remains con-
stant at that level until almost 10 years of age (Chugani, Phelps, & Mazziotta, 1987), which is
thought to indicate the energy demands of these processes. Oligodendrocyte-based develop-
ment continues into adulthood, with cortical myelination continuing to increase until the third
decade of life (Shafee, Buckner, & Fischl, 2015) and white matter volumes peaking in the fifth
decade of life (Paus et al., 2001). Healthy neurodevelopment ex utero is highly complex, avec
different processes following regionally specific, often nonlinear, trajectories ranging in time
span from years to decades. Characterizing typical developmental trajectories is key if we are
to understand why some children develop pathology, such as mental disorders or neurological
conditions, while others remain free from illness. While we have a rapidly increasing compre-
hension of neurodevelopment at a cellular level (par exemple., Tau & Peterson, 2010), a more modest
literature offers descriptions of human neurodevelopment derived from in vivo measurements.
Understanding at this level is crucial, given that the symptomology of many disorders is re-
flected in the impairment of complex and uniquely human cognition and behavior. Here we
characterize typical neurodevelopment using in vivo neurophysiology in a large normative
cohort.
Functional magnetic resonance imaging (IRMf) and electroencephalography (EEG) have
made advances in our conception of human neurodevelopment. Cependant, fMRI depends on
the blood oxygenation level dependent signal, which has a variable lag between neural recruit-
ment and signal change in the order of seconds (Ye¸silyurt, Whittingstall, Uˇgurbil, Logothetis, &
Uludaˇg, 2010). Plus loin, recent improvements in fMRI (pre)processing have called many “clas-
sic” developmental fMRI findings into question, particularly the many studies that failed to
adequately account for head motion–induced artefacts (Grayson & Fair, 2017). EEG offers a
direct measurement of electrophysiology but because of difficulties in modeling the inhomo-
geneous and geometrically complex conductivity profile in the head, the spatial resolution
is limited. Donc, a more promising technique is magnetoencephalography (MEG), lequel
offers a noninvasive and direct measurement of in vivo brain function. Measuring the minute
band-limited magnetic fields generated by synchronous postsynaptic potentials in populations
of pyramidal neurons (Hämäläinen, Hari, Ilmoniemi, Knuutila, & Lounasmaa, 1993), MEG
measurements are on the order of milliseconds and, when combined with appropriately im-
plemented spatial filtering informed by anatomical MRI, offer a millimeter spatial resolution
(Troebinger, López, Lutti, Bestmann, & Barnes, 2014). In combination, these features make
MEG ideally suited to the characterization of neurodevelopment (Lopes da Silva, 2013), les deux
in terms of fundamental electrophysiological features and in terms of functional connectivity
between disparate regions.
498
Functional connectivity:
Nondirectional statistical relations
between signals measured from
disparate brain regions thought to
indicate physiological
communication.
Neurosciences en réseau
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
Neural oscillations:
Rhythmic changes in magnetic field
produced by populations of neurons;
a key mechanism of interregional
communication in the brain.
Power:
The amplitude squared.
Power spectral density:
A measure of a signal’s power
content at specific oscillatory
frequencies.
Resting state:
Measuring changes in brain activity
when the subject is at rest, typically
passively viewing a fixation cross.
Amplitude:
Change of a neural oscillation from
its mean value.
Neural oscillations, as measured by (M.)EEG, are integral to healthy brain function (Fries,
2005). Oscillations are characterized by their oscillatory power and phase, the latter thought
to enable interregional communication (Fries, 2005). Despite being fundamental to the oscilla-
tion, the two measurements are minimally related and provide complementary insights into
underlying neurophysiology. Here we investigated how both features change during typical
neurodevelopment. Despite many decades of research with both EEG and MEG, a limited
literature describes typical neurodevelopment from an electrophysiological perspective. Le
majority of these investigations used EEG with a small number of electrodes, allowing limited
comparisons to MEG. Néanmoins, a consistent finding is that power spectral density (PSD),
linked to local processing, decreases with age in the slower frequency bands (delta, ∼1–4 Hz,
and theta, ∼4–7 Hz; Brookes et al., 2018; Gómez et al., 2017; Miskovic et al., 2015; Perone,
Palanisamy, & Carlson, 2018; Vlahou, Thurm, Kolassa, & Schlee, 2014). There are also some
reports of higher frequency PSD decreasing with age, such as alpha (∼8–13 Hz; Dias et al.,
2015; Gómez et al., 2017) and beta (∼13–30 Hz; Rodriguez-Martinez, Barriga-Paulino, Rojas-
Benjumea, & Gómez, 2015) frequencies. Whitford and colleagues (Whitford et al., 2007)
collected EEG and structural MRI data in 10- to 30-year-old subjects. They found similar curvi-
linear relations between age and PSD and between age and cortical volume in frontal and
parietal lobes. The authors suggested that PSD is related to synaptic pruning, with decreasing
PSD in these lobes indicating decreasing synapse numbers. Cependant, this relationship may not
hold for all frequency bands, with other reports demonstrating an increase in PSD with age in
alpha, beta, and gamma (∼30–70 Hz) bands (Benninger, Matthis, & Scheffner, 1984; Clarke,
Barry, McCarthy, & Selikowitz, 2001; Perone et al., 2018).
While PSD is reported to be intimately linked to neurodevelopment, a more intuitive empiri-
cal focus is developmental changes in how brain regions communicate. Since the first observa-
tion of statistical dependency between isolated regions during rest (Biswal, Yetkin, Haughton,
& Hyde, 1995), the field of functional connectivity (FC) has made great progress. Assessment
of brain function in the absence of a task, often termed the resting state (Fox & Raichle, 2007),
has proven to be a powerful technique for the study of neurodevelopment. Where a task may
be more or less difficult for participants of different ages, the external stimuli during a resting-
state acquisition is identical across the life span. Abnormal MEG-derived resting-state FC has
been found in psychiatric conditions such as schizophrenia (par exemple., Brookes et al., 2016), major
depressive disorder (MDD; par exemple., Nugent, Robinson, Coppola, Furey, & Zarate, 2015), and post-
traumatic stress disorder (PTSD; par exemple., Dunkley et al., 2015). In pediatric populations, reduced
functional connectivity has been found in children diagnosed with autism spectrum disorder
during a working memory task (Urbain et al., 2016) and in children born very preterm (Ye,
AuCoin-Power, Taylor, & Doesburg, 2016). Given this growing evidence base, characteriza-
tion of FC development across the life span in healthy subjects is a highly pertinent research
area.
A limited literature has assessed the electrophysiological basis of neurodevelopment in
terms of whole-brain functional connectivity changes across the life span. Schäfer and col-
leagues (Schäfer, Morgan, Ye, Taylor, & Doesburg, 2014) assessed whole-brain functional con-
nectivity using MEG-derived amplitude envelope correlations in 59 participants aged 6 à
34 années. Using a priori defined canonical resting-state networks, they found global within-
and between-network connectivity to linearly increase with age, in the theta, and most strik-
ingly in the alpha and beta bands. Cependant, a significant amount of evidence indicates cortical
developmental trajectories to take a nonlinear form.
Neurosciences en réseau
499
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
Studies assessing cortical thickness find development to take an inverted U, quadratic trajec-
tory on a near global basis between the ages of 3 et 33 années (Giedd, 1999; Giedd & Rapoport,
2010; Gogtay et al., 2004; Shaw et al., 2008; Vandekar et al., 2015), although some also find
evidence for logarithmic trajectories in a younger sample (1–6 years; Remer et al., 2017). Stud-
ies using fMRI to investigate neurotypical trajectories have also found this to be the case, tel
as Dosenbach et al. (2010) who found asymptotic development curves to be most success-
ful at predicting brain age via machine learning in a sample aged between 7 et 30 années.
Given the intimate relation between hemodynamic and electrophysiological techniques (voir
Hall, Robson, Morris, & Brookes, 2014, for review) and between electrophysiology and corti-
cal structure (Hunt et al., 2016; Tewarie et al., 2016; Tewarie et al., 2014), nonlinear models
are likely to be most successful in capturing electrophysiological maturation.
Whole-brain measures of FC offer great utility (Brookes et al., 2016) but can generate an un-
manageable mass of data for each individual, leading many groups to adopt graph theory met-
rics that collapse across connectivity matrices. Here we used two such measures termed node
strength and global efficiency (GE; Rubinov & Sporns, 2010; Sporns, 2011). Node strength is
derived as the sum of all connections (edges) to a region (node), which measures how con-
nected each region is within the greater network, whereas GE is a global measure assessing the
networks’ ability to transmit information on a global scale and is influenced most by shorter
paths (Rubinov & Sporns, 2010). Previously, GE was measured from resting MEG data, et
was found to relate to functional coupling within networks (de Pasquale, Della Penna, Sporns,
Romani, & Corbetta, 2016). Node strength has been derived consistently and used more widely
than GE across numerous imaging modalities. In MEG, node strength measures are increased
in PTSD (Dunkley et al., 2016) and reduced in mild traumatic brain injury (Pang, Dunkley,
Doesburg, de Costa, & Taylor, 2016), indicating that this metric has sufficient sensitivity to
detect differential functional perturbations in these behaviorally overlapping conditions. Le
developmental trajectory of node strength is somewhat contested within the MRI literature,
with some studies reporting strength decreases between 7 et 22 années (Supekar, Musen, &
Menon, 2009), some reporting age-related increases between 8 et 16 (J.. R.. Sato et al., 2014),
and others characterizing strength using nonlinear analyses, reporting strength to take a nega-
tive quadratic form across a much larger age range (7 à 85 années; Cao et al., 2014). A recent
MEG study found strength to develop nonlinearly between 9 et 25 années, with theta band
strength following a quadratic developmental curve and alpha, beta, and gamma to take a
monotonically increasing or decreasing form (Brookes et al., 2018). Cependant, strength was
averaged across all cortical parcels, precluding regional results and conclusions about topo-
graphic changes in strength.
Although GE has been used less frequently than node strength, a recent study by Kahn and
colleagues (Khan et al., 2018) adopted nonlinear graph theory analyses. Using MEG-derived
amplitude envelope connectivity (AEC) from a large sample (n = 162 participants) âgé 7 à
29 années, the authors found significant nonlinear increases in GE in the gamma band only. Us-
ing machine learning based on a composite of graph metrics, the authors found linear increases
in the beta band to best predict subject age, et, using a different graph metric composite,
found the most successful predictors of age in the gamma band to take a quadratic (inverted
U) formulaire. In contrast to the study by Schäfer et al. (2014), the group did not find any significant
effects in the alpha band.
Here we characterized neurodevelopmental connectivity, measured by MEG, in the largest
developmental cohort that currently exists (N = 324, from a sample of 425). Because of the
mixed MRI and MEG literature and our uniquely large sample size, we conducted an initial
Amplitude envelope connectivity:
Functional connectivity measured
by assessing correlations over
time between the amplitudes of
electrophysiological signals from
two brain regions.
Neurosciences en réseau
500
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
Phase synchrony:
A term to describe a shared timing
of the peaks and troughs of
oscillations/brain waves between
two regions.
Beamformer:
A class of spatial filter used to infer
the region of the brain responsible
for magnetic field changes measured
by MEG sensors.
Adjacency matrix:
A way of presenting brain
connectivity data. Each matrix
element corresponds to the
connection strength between two
brain regions.
analysis of node strength, derived from measurements of phase synchrony between regions.
Given the recent findings of Brookes and colleagues, we hypothesized that strength would
be characterized predominantly by negative quadratic curves in theta, nonlinearly increasing
curves in alpha and beta, and nonlinearly decreasing curves in gamma frequency strength.
We also measured GE and hypothesized to replicate the findings of Khan et al. (2018), OMS
identified a significant nonlinear GE trajectory in the gamma band. We then conducted a PSD
analysis to evaluate the relation between whole-brain band-limited PSD and age. Based on the
previous EEG literature, we hypothesized PSD to decrease globally with age most strikingly in
the theta frequency band. As there have been mixed reports of the relation between PSD and
age in higher frequencies, we predicted a significant change with age. Enfin, we assessed for
any differences in signal-to-noise ratio (SNR) across our sample.
RÉSULTATS
Following data quality control, a cohort of 324 participants (from an initial group of 425, âgé
5.9 à 45.5 années) was included in our analyses (mean age [SD]: 18.8 [9.9], 116 females).
Resting-state MEG data were epoched into 10-s “trials” and an automated procedure was used
to remove trials containing significant artefact or excessive head movement (>7 mm). MEG
data were coregistered to individual anatomical MRIs and, pour chaque participant, a single-shell
head model constructed. The center of mass of each automated anatomical labeling (AAL)
atlas parcel (Tzourio-Mazoyer et al., 2002) was nonlinearly unwarped into subject space, et
a beamformer class of spatial filter was used to project MEG data into source space to derive a
single signal time course for each AAL parcel. Regional signals were frequency filtered into four
classical frequency bands (theta: 4–7 Hz, alpha: 8–14 Hz, beta: 15–30 Hz, gamma: 31–80 Hz)
and used to measure PSD and to estimate functional connectivity using the weighted phase
lag index (Vinck, Oostenveld, van Wingerden, Battaglia, & Pennartz, 2011). See the Methods
section for a schematic illustration of our processing pipeline (Chiffre 6).
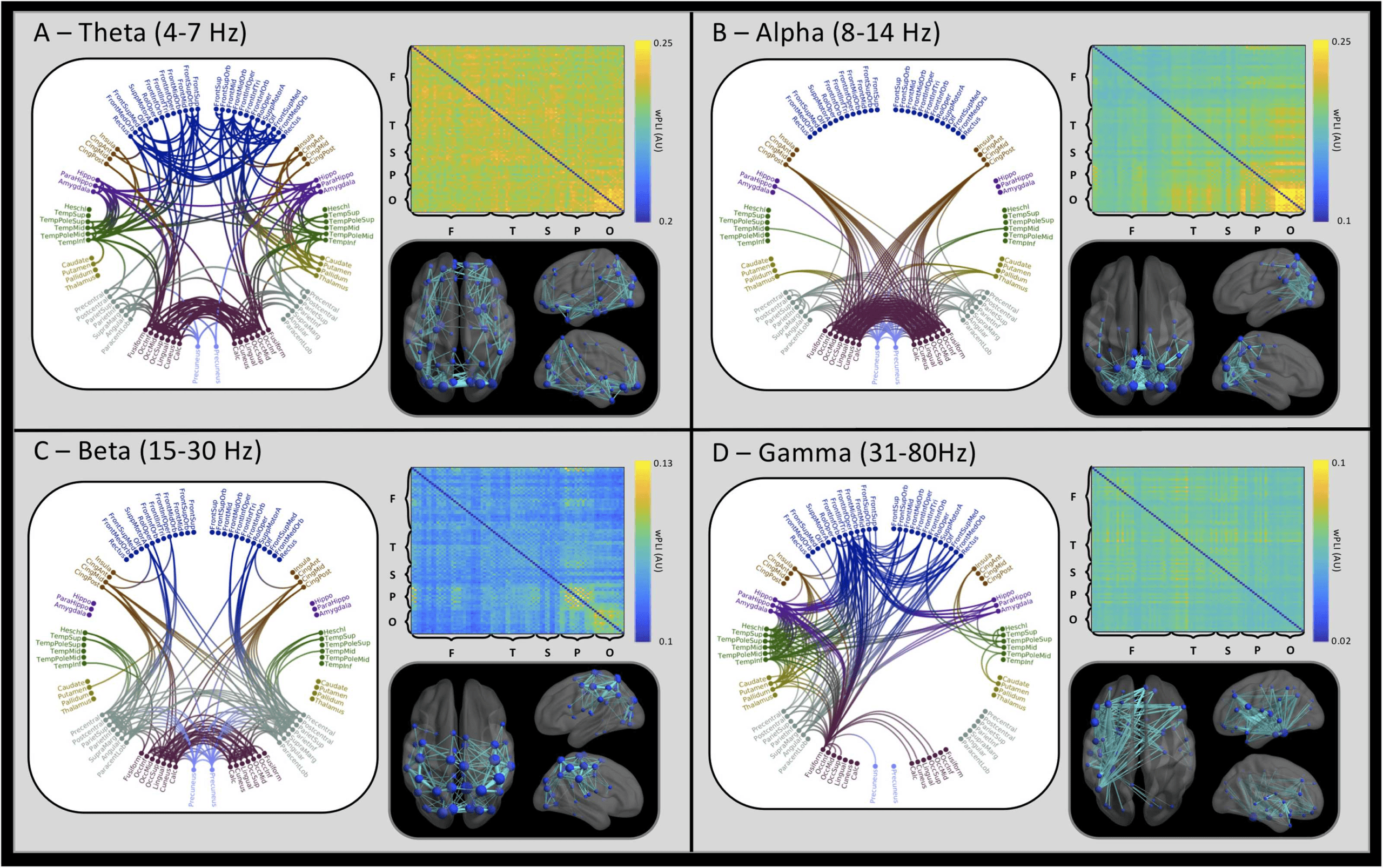
Grand-Average Functional Connectivity Highlights Spectral Segregation
The weighted phase lag index (wPLI) assumes that a functional connection exists between
two regional signals if the phase of one signal consistently leads or lags another over time.
Values of wPLI range from 0 à 1, avec 0 indicating a completely random/symmetric phase
distribution and 1 indicating a highly asymmetric distribution; wPLI was calculated between
all node pairs of the AAL atlas, within each frequency band, enabling a whole-brain measure
of neural synchronization for each participant.
Grand-averaged adjacency matrices are plotted in Figure 1, accompanied by glass brains
and circular connectivity plots showing the top 5% of connections, for each frequency band.
Adjacency matrices plot the wPLI value between two brain regions, with greater values indi-
cating a greater functional connection between regions. Matrices are arranged by lobe, avec
homologous regions plotted in neighboring rows and columns (left then right). The strongest
connections within the theta band are broadly distributed, as shown in the circular connectiv-
ity plot, whereas the alpha band adjacency matrix is dominated by high occipital phase syn-
chrony, indicated by the bright yellow square. This feature is echoed in the glass brain, avec
a high concentration of connections within the occipital lobe and a high degree of occipital
symmetry in the alpha circle plot. While the beta band glass brain (Figure 1C) also includes
many occipital connections, there is a reduced local concentration with many connections
entering the parietal lobe, particularly across the motor cortices. The gamma frequency wPLI
adjacency matrix is characterized by broadly distributed high connection weights. Plotting the
Neurosciences en réseau
501
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 1. Grand-averaged functional connectivity (wPLI). Within each frequency quadrant, connections falling within the 95th percentile are
plotted in the circle plots and glass brains. The circle plots are arranged anatomically such that left hemisphere regions appear on the left of the
plot. Regional connections (edges) within the glass brains are scaled within band, with thicker edges indicating a greater connection strength.
The spheres indicating individual regions (nodes) are scaled by connectivity strength, with larger spheres indicating greater connectivity values
to connected regions, within the 95th percentile of connections. Adjacency matrix labels indicate the lobular segregation of the matrix, dans le
order of frontal (F), temporal (T), subcortical (S), pariétal (P.), and occipital (Ô). Note that adjacency matrix color axes are scaled independently
by band.
Edge weights:
A graph theory term for the
connection strength between
nodes/regions of a graph/network.
highest 5% of edge weights onto the glass brain highlights a hemispheric asymmetry, with a
greater number of connections in the left hemisphere for gamma. The circle plot reveals that
many left hemisphere regions connect bilaterally to the frontal lobes and subcortical structures
such as the hippocampus.
Graph Theoretic Measurements Mature With Nonlinear Trajectories
Measurements of node strength and global efficiency were entered into a cross-validation
regime, enabling the selection of the best fitting curve, based upon a linear, quadratic or loga-
rithmic model. This technique offers robust curve selection that is not influenced by the com-
plexity or number of coefficients of a curve (see Methods). While other experimenters have
chosen model-free techniques to characterize neurodevelopment, such as local regression
(Khan et al., 2018), we adopted this fitting regime as our models can be described com-
pletely by four or fewer coefficients, facilitating replication. Following completion of the cross-
validation, the best fitting curve type was regressed with the full dataset and an empirical
significance level, derived from null distributions, was obtained. The resultant p values were
Neurosciences en réseau
502
Determining typical neurophysiological trajectories
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
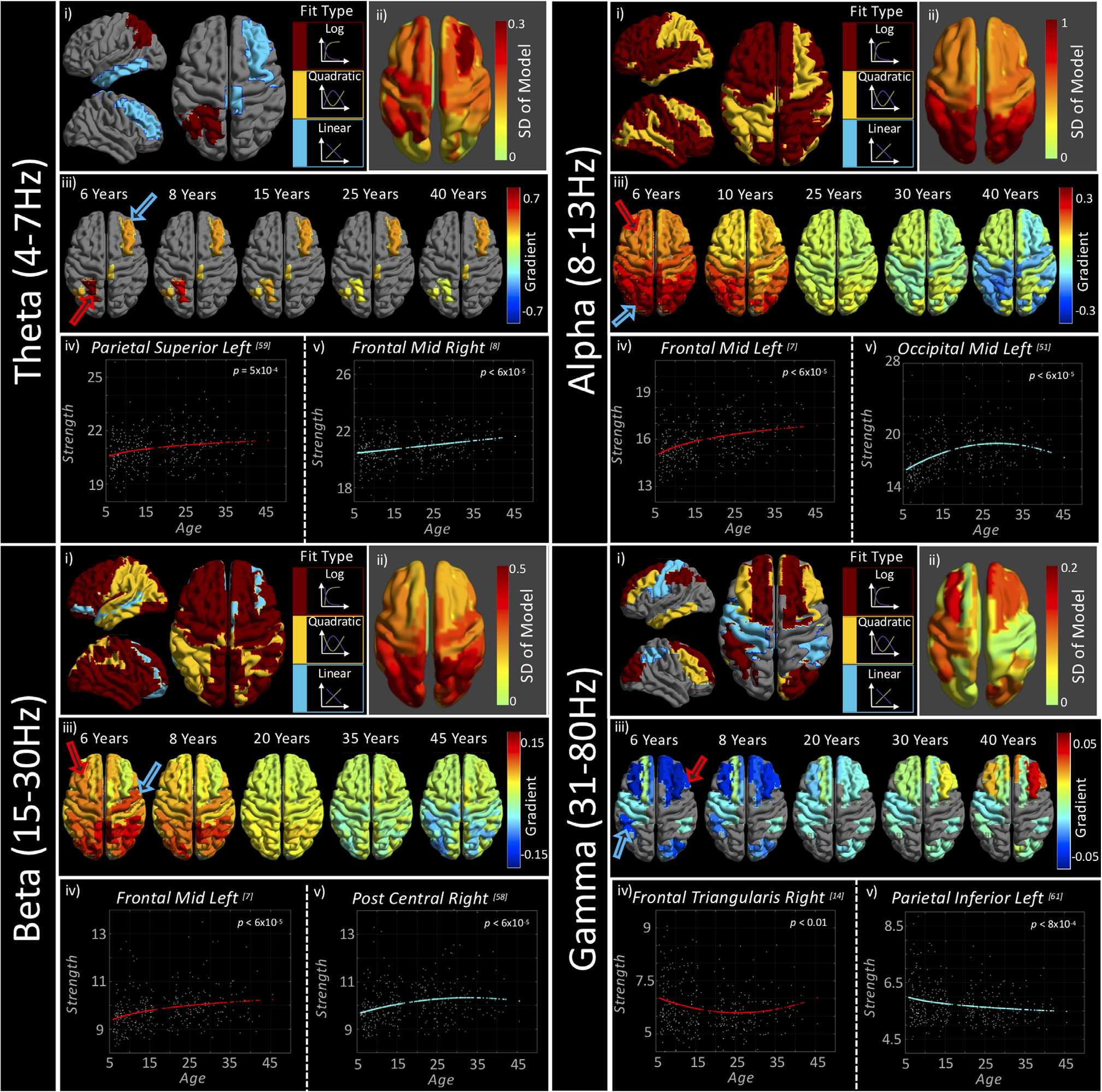
s
Chiffre 2. Typical neurodevelopmental trajectories assessed with wPLI-S. Each quadrant (band) contains the following sections: Section (je)
plots curve types that were found to be statistically significant on a regional basis. Any regions in gray indicate regions with nonsignificant
fits. Section (ii) presents the standard deviation (SD) of the model, indicating to what extent the data change over the developmental course.
Section (iii) plots axial brain images indicating the gradient between age and wPLI-strength. Hotter colors indicate a steeply increasing gradient
(par exemple., the upward portion of a quadratic or log fit) and cooler colors indicate a steeply decreasing gradient; note the change in color axes with
frequency band. Regions in gray are regions where the best fit was found not to be significant. Arrows on the first brain in (iii) correspond,
in color, to the graphs plotted in (iv) et (v), plotting the relationship between age and wPLI-strength, with the colored line indicating the
model that best characterizes that regional relationship. See Figures S7 and S8 (Hunt et al., 2019) for videos depicting gradient change from
6 à 45 years of age. Video S7 presents gradients only for significantly characterized curves, and S8 presents all gradients. Table S10 (Hunt
et coll., 2019) details the best fitting model coefficients for each region and frequency band. We also present a figure focused on subcortical
trajectories in the supplementary material, Figure S1 (Hunt et al., 2019).
Neurosciences en réseau
503
Determining typical neurophysiological trajectories
Gradient:
The rate of change or “steepness” of
a curve.
corrected for multiple comparisons using a false discovery rate correction at a threshold of
q < 0.05, within each frequency band.
Figure 2 presents the results of the curve fitting analysis for node strength measurements. The
top row of each subfigure (i) first presents the best model type on a regional basis, for statistically
significant fits only (q < 0.05). The second plot of the top row (ii) presents the extent to which
the data change across the life span, calculated as the standard deviation (SD) of the best fitting
model. The second row (iii) presents the statistically significant curve gradients for exemplar
ages. These gradients are also presented as both significance thresholded and unthresholded
videos (S7 and S8, respectively; Hunt et al., 2019). The third row presents exemplar graphs
(iv and v) for two regions (indicated by colored arrows in [iii]). Note that the color axis of the
brain plots has a direct relation to change in node strength, with, for example, a value of 0.7
indicating that for every year of development, node strength increases by 0.7. The theta band is
characterized by predominantly linear trajectories, with only left parietal regions fitted with a
more complex logarithmic curve. The regions with the greatest change in strength, as indicated
by hotter colors in the upper right brain plot, are also those to be significantly characterized by
our curves within the theta band. These regions form one half of an asymmetric frontoparietal
network. The left graph (red curve) plots an exemplar logarithmic fit, and the right graph (blue
curve) plots a linear fit from the right frontal region.
In the alpha band, all regional strength measures were found to be statistically significant,
with the logarithmic curve most successfully characterizing the development of node strength.
The few alpha band regions that were characterized by a quadratic curve closely match those
in theta band that were found to contain significant fits, with an asymmetric frontoparietal
distribution. Positive alpha band curve gradients were greatest in the occipital lobes and the
highest negative gradients, toward the older participants in our cohort, occur in parietal and
sensorimotor cortices, indicating that strength in these regions descends more rapidly with age
than those in the occipital regions.
The beta frequency band was significantly characterized by all three curve types, although
the majority of regions were best described by a logarithmic model. As observed in the
alpha band, occipital regions were found to have the steepest positive developmental gra-
dient. However, within the beta frequency band the negative gradients for quadratic fits were
approximately equal across occipito-parietal nodes. The best model for the right somatosen-
sory cortex (postcentral gyrus) is plotted in the right graph (blue); note that the peak of this
quadratic fit occurs at approximately 30 years old.
In the theta, alpha, and beta bands, nonlinear models were all positively characterized;
node strength first increased with age. In the gamma band the opposite was true, with all
linear and nonlinear fits characterized by negative coefficients, indicating reducing strength in
childhood. This relation is most apparent in a quadratic form in the frontal regions, which at
6 years old are steeply negative (a 0.05 decrease in strength per year of age) and at 40 years
are steeply positive (a 0.05 increase in strength per year).
These patterns are echoed in a detailed analysis of subcortical regions, presented in the
Supporting Information (Figure S1; Hunt et al., 2019). The alpha and beta bands were signif-
icantly characterized by nonlinear trajectories in all subcortical regions. The theta band was
significantly characterized by a positive linear model in the left caudate region. The gamma
band was also found to develop linearly, with decreasing strength in the right thalamus and
left parahippocampal region.
Network Neuroscience
504
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
a
_
0
0
0
7
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
a
_
0
0
0
7
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
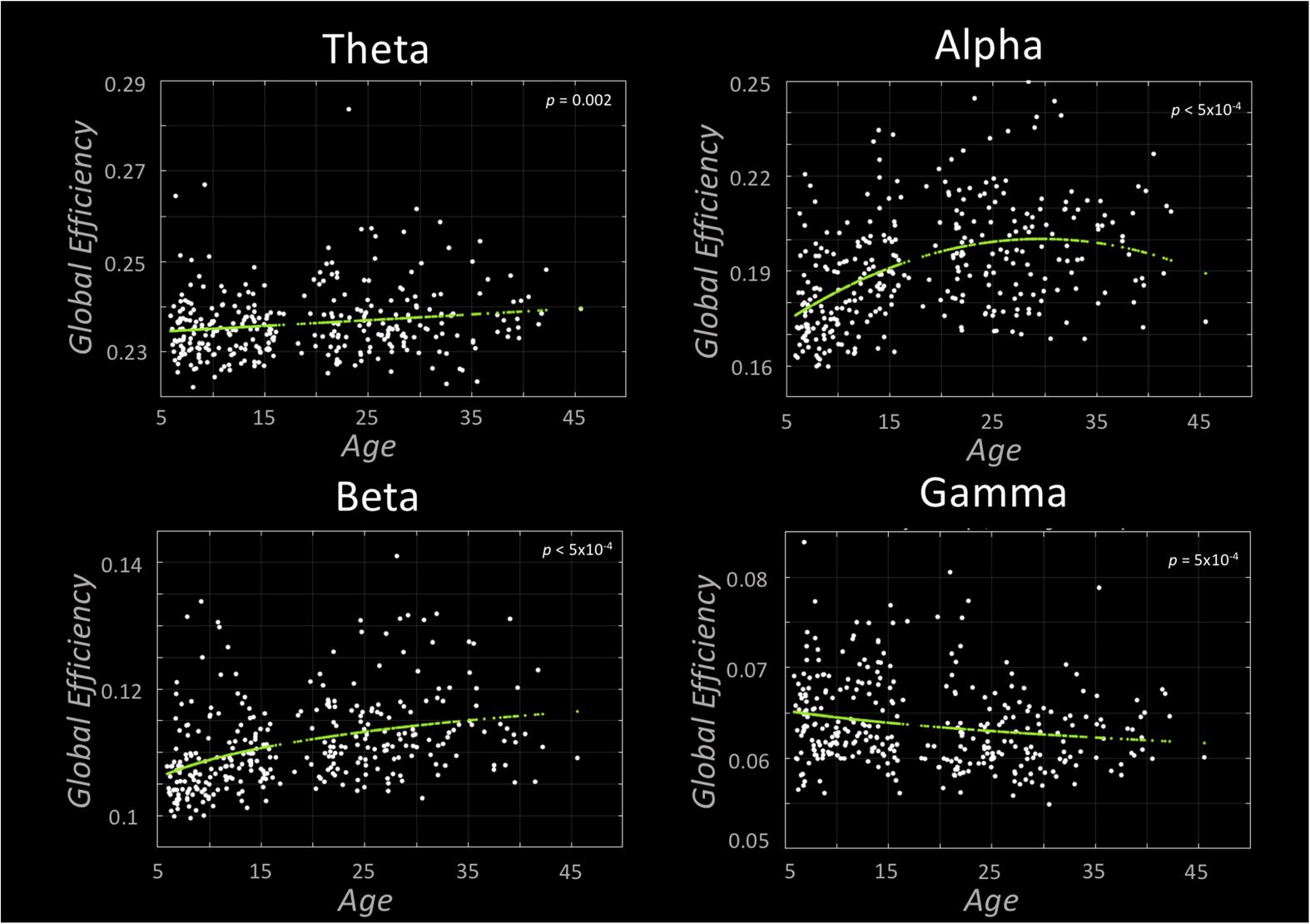
Figure 3. Typical neurodevelopmental trajectories assessed with global efficiency (GE). Each plot-
presents the statistically significant curve fits following our cross-validation analysis. Theta band GE
is significantly characterized by a linearly increasing fit, whereas the alpha and gamma bands were
characterized by a quadratic curve. The beta band was significantly characterized by a logarithmic
curve.
Figure 3 presents the results of our global efficiency (GE) curve fitting results, where a higher
GE score indicates that information is passed more easily throughout the entire network. In all
bands, GE development was significant. Theta band GE was found to increase linearly with
age. Alpha and gamma band GE was significantly characterized by quadratic curves, the former
peaking at 30 years of age. Beta GE was characterized by a logarithmic model.
Maturation of Spectral Power Is Regionally Specific
PSD was calculated for each trial and each subject. Trial-averaged PSD values were then corre-
lated with subject age for each region of the AAL atlas. In keeping with the majority of previous
studies (Benninger et al., 1984; Miskovic et al., 2015; Vlahou et al., 2014), we used linear cor-
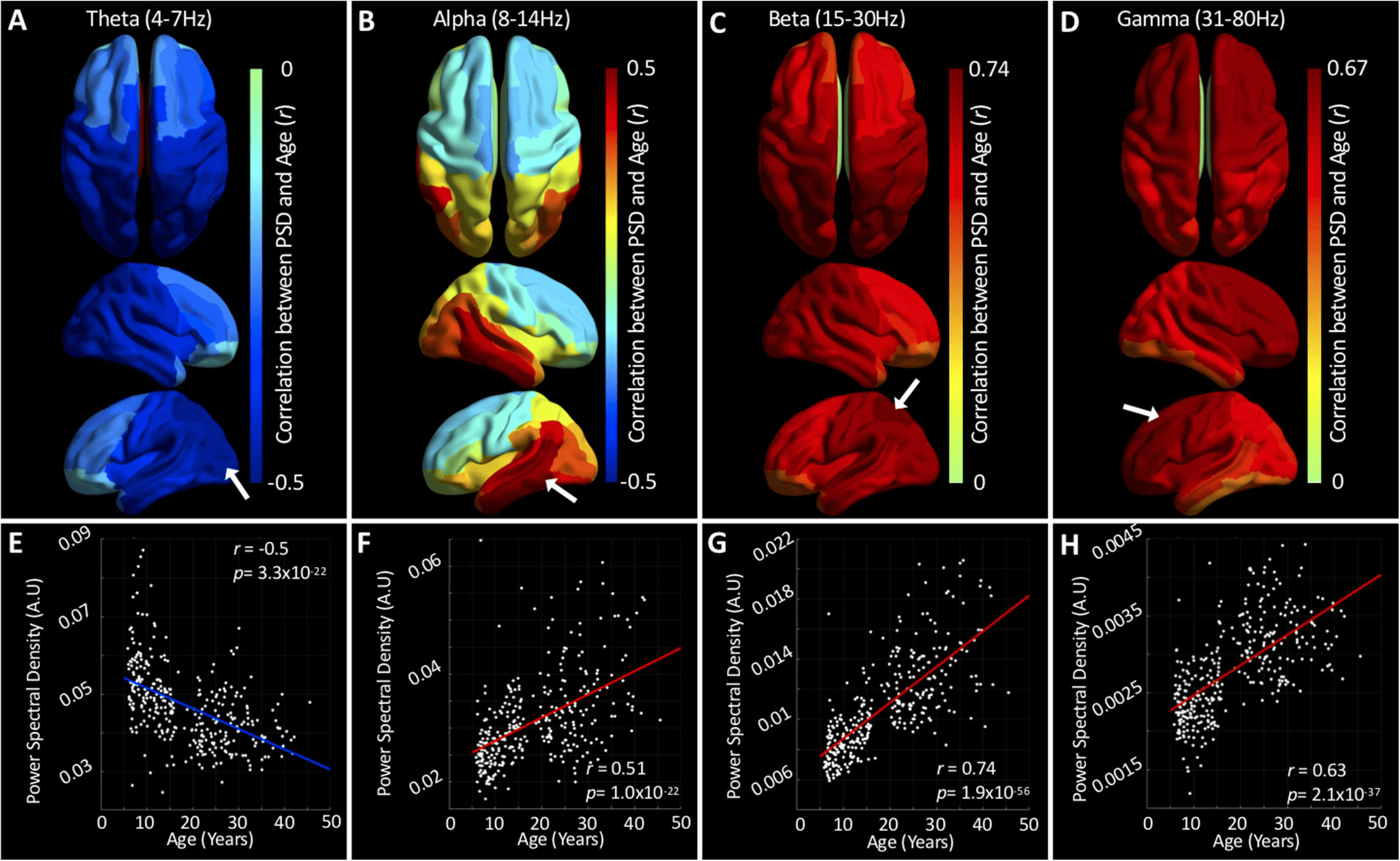
relational analyses to investigate the relation between age and PSD. Figure 4, A–D, presents
regional maps of bivariate Pearson correlation coefficients (r) between PSD and age for the
four frequency bands. Scatterplots for exemplar regions (indicated by white arrows) for each
band are presented in Figure 4, E–H.
Theta band PSD was negatively correlated with age in all regions, with maximally negative
correlations occurring in the occipital lobe. Alpha band PSD comprises both positive and
negative correlations with age, with negative correlations confined to the superior frontal and
anterior parietal regions. Beta band PSD was positively correlated with age in all regions, with
maximal correlation occurring on the superior border between occipital and parietal lobes.
Finally, gamma band PSD estimates were also found to be positively correlated with age in all
regions, albeit to a marginally lower extent than those in the beta band.
Network Neuroscience
505
Determining typical neurophysiological trajectories
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
a
_
0
0
0
7
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. The relationship between age and power spectral density. A–D present regional correlations between PSD and age for each
frequency band. Hotter colors indicate a positive correlation and cooler colors indicate the opposite relationship. E–H plot the correlation
between PSD and age for exemplar regions (indicated by white arrows in A–D). See Figure S3 (Hunt et al., 2019) for the results of an analysis
investigating the extent to which these differences arise because of age-related changes in SNR (A–D) or head motion (E–G).
To further investigate our PSD results, we performed supplementary analyses. The first, pre-
sented in Figure S2 (Hunt et al., 2019), investigated the presence of heteroskedasticity of PSD
values across our age range; we found little evidence for systematic effects aside from the
theta band, where the variance in all regions was found to be greater in younger subjects. Our
second analysis demonstrated quantitatively that PSD effects were not driven by SNR changes
across our cohort (Figure S3; Hunt et al., 2019).
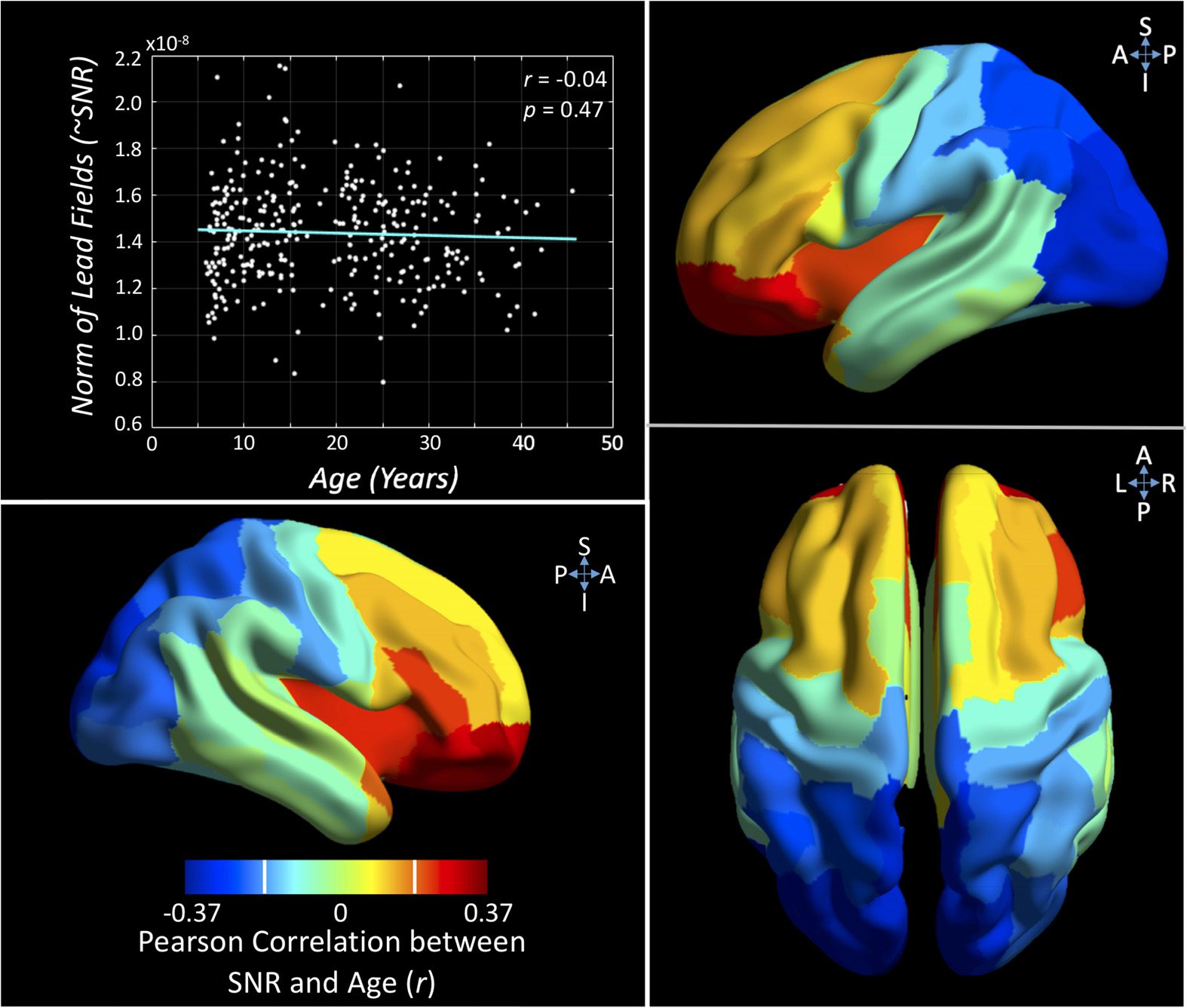
Signal-to-Noise Ratio Is Related to Age
Our sample spans an age range of almost 40 years, containing data from a diverse set of
head/brain sizes. As the MEG signal decreases rapidly with distance, it is important to assess
whether the age of the participant significantly impacts our ability to accurately measure neuro-
magnetic signal. Given the known head size and the location of the sensors with respect to
the brain, we modeled the expected signal-to-noise ratio individually for every subject. We
found no significant effect of age on the average SNR across the head (r = −0.04, p = 0.47).
Figure 5 also plots the regional correlation values between SNR and age; here a negative
correlation implies greater SNR for younger subjects, and a positive correlation the opposite.
Note that the threshold for a significant regional correlation is r ≥ 0.19 (t > 3.5, pcorr < 0.05).
Network Neuroscience
506
Determining typical neurophysiological trajectories
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
a
_
0
0
0
7
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Assessing the influence of age on signal-to-noise ratio. The upper left graph plots the
correlation between a proxy for global signal-to-noise ratio (SNR; Frobenius norm of the lead fields)
and age (nonsignificant, p > 0.05). The brain plots expand on this relation by plotting the Pearson
correlation between SNR and age on a regional basis. The white vertical bars on color scale indicate
threshold for significant correlation (r = ±0.19). See Figure S5A (Hunt et al., 2019) for a greater
exploration of this effect.
This spatial analysis revealed younger subjects to have significantly greater SNR in occipital
regions and reduced SNR in inferior frontal regions. Figure S5A (Hunt et al., 2019) presents a
further exploration of this, plotting SNR against age for a frontal and occipital region.
DISCUSSION
Typical neurodevelopment is a highly complex process with many different cellular changes
occurring over many timescales (Tau & Peterson, 2010). Characterization of these trajectories is
key if we are to understand the developmental origins of complex human behaviors as well as
neurological and cognitive pathology. In the present study, we used magnetoencephalographic
measurements of power spectral density (PSD) and functional connectivity (FC) to characterize
developmental trajectories as measured by this powerful technique. Our sample is the largest
currently available, placing us in a unique position to conduct a well-powered foundational
étude. Our results replicate and extend the previously reported negative relationship between
theta band PSD and age, and add to the weight of evidence that higher frequency bands (beta
and gamma) exhibit globally reversed correlation coefficients compared with slow-wave oscil-
latory power. Our calculation of weighted phase lag index (wPLI) revealed spatially different
topologies for the strongest connections within each frequency band, with alpha wPLI, pour
Neurosciences en réseau
507
Determining typical neurophysiological trajectories
example, characteristically marked by dense occipito-occipito connections. De plus, node
strength, based upon measurement of wPLI, was found to exhibit complex relations with age
in a regional and frequency-specific manner, a pattern mirrored in measurements of global
efficiency.
Power Spectral Density: Replication and Novelty
The finding of a negative relationship between theta band PSD and age has been reported
widely (Miskovic et al., 2015; Rodriguez-Martinez et al., 2015; Smit et al., 2012; Whitford
et coll., 2007). Three studies, to the authors’ knowledge, have reported age-related increases
in PSD in higher frequencies, and our results buttress these findings (Benninger et al., 1984;
Clarke et al., 2001; Perone et al., 2018). We report, for the first time, frontal decreases and
occipital, pariétal, and temporal increases in alpha band PSD with age (Figure 4B). Whitford
et autres. (2007) argued the physiological basis of PSD is linked to cortical volume, as the team
found similar relations between age and PSD and age and cortical volume. While our current
results preclude direct comment on this claim, our findings revealing PSD to be both positively
and negatively correlated with age in the alpha band suggests a more complex relationship.
Whitford et al. used voxel-based morphometry to derive measures of cortical volume. Although
this technique has been used extensively, a more detailed approach is to measure different
properties of the cortex as opposed to volume, such as cortical thickness, local gyrification,
and surface area, and to make inferences on microstructural properties such as myelination
(Geades et al., 2016). These measures have different developmental trajectories (Winkler et al.,
2010) and relate nontrivially to one another. It would therefore be pertinent for future studies
to investigate the relationship between these more detailed measurements of cortical structure
and PSD.
Grand-Average wPLI: Insights from Gamma Band Asymmetry
Chiffre 1 plots the grand-averaged wPLI, collapsing across age groups. Taking the strongest 95%
of connections, striking symmetry is seen in the theta, alpha, and beta bands. This is in contrast
to the profound asymmetry seen in the gamma band. This asymmetry is biased toward the left
hemisphere with many within-hemisphere connections. Gamma frequency oscillations are
produced in superficial cortical layers (Buffalo, Fries, Landman, Buschman, & Désimone, 2011)
and are therefore considered to be primarily associated with local communication. The left
hemisphere has been found to be biased toward within-hemisphere connections (Gotts et al.,
2013), whereas the right hemisphere shows no such bias. This may underlie the asymmetry in
the group-averaged gamma band plots.
wPLI-S and GE: Windows to Cognitive and Sensory Development
Our analysis of wPLI-strength (wPLI-S) revealed highly heterogeneous developmental trajecto-
ries both within and between frequency bands. The major trends across bands were echoed in
GE analysis. It is noteworthy that quadratic and asymptotic/logarithmic models suggest distinct
underlying processes. Quadratic model fits indicate processes that are continually evolving
across the life span (or sample age range). A quadratic form may arise either from a single
physiological phenomenon with a quadratic trajectory, or from two nontemporally coincident
processes, the second of which occurs at the peak or trough of the quadratic form. In contrast,
a logarithmic model suggests an early period of rapid change that gradually plateaus.
Theta band wPLI-S was characterized primarily by linear relations with age, albeit in a small
number of regions. Theta is understood to be a frequency band associated with long-range
Neurosciences en réseau
508
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
cognitive communication in the brain; the linear relations seen may reflect the steady increase
in cognitive abilities across age, even into mid-adulthood. This process would involve increas-
ing the capacity for information transfer in the global network, explaining the steady increase in
GE across our cohort. We also found two regions in the left parietal cortex to be characterized
by logarithmically increasing wPLI-S with age. This brain region is particularly involved in writ-
ing (Menon & Desmond, 2001), and this more rapid early increase may index the acquisition
and refinement of this skill in our younger participants.
Generators of alpha frequency oscillations have been identified across the cortical laminae
(Bollimunta, Mo, Schroeder, & Ding, 2011; Dougherty, Cox, Ninomiya, Leopold, & Maier,
2017), suggesting that they may modulate both local and long-range connectivity (Uhlhaas,
Haenschel, Nikoli´c, & Chanteur, 2008). Alpha band oscillations have been linked closely with
cortical inhibition (Pfurtscheller & Lopes da Silva, 1999), which is thought to be a key pro-
ponent of healthy brain function (Klimesch, 2012), as well as memory (Freunberger, Fellinger,
Sauseng, Gruber, & Klimesch, 2009; Jensen, Gelfand, Kounios, & Lisman, 2002; J.. Sato et al.,
2018), an essential cognitive skill. Several reports indicate that cognitive processes requiring
cortical inhibition, such as cognitive control (Houghton & Tipper, 1996), continue to develop
in adolescence and adulthood (Adleman et al., 2002; Rubia et al., 2006), as do mnemonic
skills (Ornstein & Light, 2010). Ici, we found alpha band wPLI-S to be characterized pre-
dominantly by logarithmic curves in the frontal lobes, and an approximately even distribution
of quadratic and logarithmic curves in other regions. Given the protracted development of the
complex cognitive activities associated with alpha oscillations, our nonlinear characterization
of alpha band wPLI-S extends the existing literature pertaining to this ubiquitous frequency
band; its widespread effects underscore the importance of cortical inhibitory and memory
functions across all regions in the brain. The rapid frontal increases in childhood are consis-
tent with the emergence of increasing ability in these executive functions, and the quadratic
curves are consistent with some later decline, particularly in memory. This potential decline is
supported by the GE results, which suggest that the peak efficiency of the network is at approx-
imately 30 ans. Plus loin, GE is most sensitive to short-range connections, meaning that
the regions with logarithmic wPLI-S may be preferentially involved in long-range subnetworks.
Similar to the alpha band results, our analysis revealed beta band wPLI-S to be predom-
inantly characterized by nonlinear curves. Prior studies have found beta band graphs to be
closely linked to brain structure (Hunt et al., 2016; Tewarie et al., 2014), which is known to
develop nonlinearly (Paus et al., 2001; Shafee et al., 2015). Invasive electrophysiological stud-
ies have identified beta band oscillations to be generated in deeper cortical layers (Sun & Dan,
2009), principally implicating this frequency in long-range communication (Brookes et al.,
2011), and in motor control (Weinrich et al., 2017). We observe high symmetry between motor
cortices, both being fitted with logarithmically increasing curves with similar curve gradients.
While the majority of motor maturation occurs before the youngest members of our cohort,
motor refinement continues through adolescence and into early adulthood (Smits-Engelsman
& Wilson, 2013), which may explain the logarithmic trajectories of the pre- and postcentral
gyri. We also observed the highest degree of hemispheric symmetry within beta band curve
gradients—potentially implicating a connectivity preference for homologous connections.
In both the alpha and the beta band, the frontal lobes are predominantly characterized by
logarithmic curves, whereas the parietal, temporal, and occipital lobes include a high num-
ber of quadratic fits. Our measurement of wPLI-S is indicative of overall connection strength
to other brain regions. En tant que tel, it is tempting to argue that the rapid increase in frontal lobe
connectivity is temporally coincident with both the selective strengthening of increasingly
Neurosciences en réseau
509
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
important functional connections, and the physical pruning of redundant connections—
generating a net increase in wPLI-S that plateaus post-adolescence (Uhlhaas, 2011). This theory
is supported by the GE results, as the pruning of redundant connections would lead to increased
efficiency in the global network. The regions that tended toward a quadratic trajectory in
alpha and beta band wPLI-S are similar to the network identified by Douaud et al. (2014),
encompassing transmodal regions within the parietal, occipital, and temporal lobes. The group
found this network to be late to mature in youth and early to decline in old age, and also
demonstrated these regions to a show heightened vulnerability to disorders occurring early
(early onset psychosis) and late (Alzheimer’s disease) in the life span. The quadratic curves we
found may be the early harbingers of later declines in function in these brain areas.
We found gamma frequency PSD to increase globally with age in addition to negative
nonlinear trajectories with connectivity strength. We interpret this as a concomitant increase in
local processing (c'est à dire., within the cortical parcel) and decrease in long-range integration. These
results replicate, to some extent, the results recently presented by Brookes et al. (2018), OMS,
in a smaller study, found whole-brain strength (regional strength summed across all parcels) à
decrease nonlinearly in the gamma frequency band. In the current study, regions exhibiting the
greatest developmental increase in gamma power (Figure 4D) were those bilateral superior-
frontal parcels that also exhibit nonlinearly decreasing gamma strength trajectories (Chiffre 2).
This is consistent with the prolonged maturation of the frontal lobes and is associated with
the greatest age-related changes in cognitive functions. These changes are in contrast to fewer
frontal effects seen in the other frequency bands, underscoring the possible linkage between
the frontal regions and the executive functions they support, within the gamma frequency.
Alternativement, the more general effects seen in the lower frequency bands, particularly alpha,
would reinforce the protracted development of widespread networks that are necessary for
human cognition.
We hypothesized GE in the gamma band to take a quadratic (inverted U) formulaire, given the re-
sults of Khan et al. (2018). We did find gamma had a quadratic form, but with an opposite sign
to the findings by Kahn et al. (a U rather than an inverted U). While our approach is very sim-
ilar to their study, we chose to use a phase-based metric of connectivity (wPLI), whereas Kahn
and colleagues used amplitude envelope connectivity. While wPLI and AEC are undoubtedly
related, they measure neuronal processes on different timescales. AEC-based analyses assess
connectivity at a temporal resolution of ∼1 Hz (the resolution of the envelope of amplitude
changes), whereas phase-based techniques remain in the millisecond range. Khan and col-
leagues used a fine-grained parcellation including more than 400 cortical regions, alors que
we used the more commonly selected AAL atlas (Tzourio-Mazoyer et al., 2002) consisting
de 90 parcels, including cortical and subcortical regions. Plus loin, our age range is larger and
more evenly distributed than that used by Kahn et al. (7 à 29 années, with the greatest density
of participants >20 years), which may mean that the early life trajectory was poorly character-
ized in their curve fitting. Donc, we believe this discrepancy may be accounted for by the
differing temporal and spatial scales at which the graph metrics are computed and the different
characteristics of our cohorts.
Future Directions and Limitations
The models used in the current study, and the methods used to create them, are highly gener-
alizable across laboratories and imaging modalities. En tant que tel, we encourage future studies to
incorporate these models into their research and assess how their data converges or deviates
from these trajectory patterns. De la même manière, we encourage those with data from clinical cohorts to
Brain parcellation:
A division of the brain based on
structural or functional features into a
number of discrete regions or parcels.
Neurosciences en réseau
510
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
assess how their findings diverge from this general model of typical development. While we
derived this model from MEG data, it would be interesting to assess the overlap between MEG
and EEG as well as MEG and fMRI.
The cohort used in this study was limited in age span by the fundamental physics of tradi-
tional MEG systems, where participants younger than approximately 5 years of age will have
heads too small, and subsequently would be too far from sensors, for reasonable SNR in MEG.
To perform MEG in younger subjects, several targeted systems exist that are specifically de-
signed to have smaller dewars such that the sensors are closer to the heads of young children
(par exemple., He et al., 2018) or for infants (par exemple., Gaetz et al., 2015). There have also been excit-
ing developments in next-generation MEG sensors, in particular optically pumped magneto-
meters (OPMs; Boto et al., 2016; Boto et al., 2018), and necessary magnetic field nulling
technologie (Holmes et al., 2018). These new sensors can be placed directly onto the scalp,
meaning an increase in SNR and negation of head movement–related artefacts. With con-
tinued development and optimization, these sensors will provide new possibilities for MEG
recordings in neonates and young children.
A further limitation is that our sample was unevenly distributed between the sexes (voir
Figure S9 in the Supporting Information; Hunt et al., 2019), meaning that it is possible our
results are more applicable to males than females. To assess this empirically, we performed
100 additional analyses with subsampling of males to match females, totaling 540 million
permutations (see the Supporting Information, section 1.4; Hunt et al., 2019) and found alpha,
beta, and gamma band wPLI-S curve types to be stable even when sexes were matched. Theta
band wPLI-S was less stable, potentially implicating that sexual dimorphisms mainly localize
to this band. This would also explain why there were few significant fits in the primary analysis
of theta band wPLI-S. Assessment of sex differences in the theta band would be an exciting
direction for future research.
In conclusion, we have performed extensive investigation of typical human neurodevelop-
ment using a large developmental MEG database. We found PSD to closely relate to age in a
frequency- and region-dependent manner. wPLI-S was significantly characterized as linearly,
quadratically, and logarithmically changing across development. Alpha and beta frequency
bands best characterized the relationship between wPLI-S and age, with the greatest number
of significantly fitted regions. Gamma band wPLI-S was found to exhibit opposite relations to
other bands, suggesting a drop in long-range connectivity with increasing age. GE was sig-
nificantly associated with age in all frequency bands, also revealing spectrally distinct linear
and nonlinear trajectories, largely consistent with wPLI-S. We have shown PSD and wPLI-S
to be highly sensitive to the fundamental functional architecture of the developing brain. Notre
foundational study provides a concrete footing from which future studies can detail links with
specific cognitive functions, and clinical research may begin to decisively map divergent patho-
logical trajectories in neurodevelopmental disorders.
MATERIALS AND METHODS
Human Subjects
A retrospective database was assembled consisting of 425 typically developing research par-
ticipants from The Hospital for Sick Children (SickKids), scanned between August 2010 et
Juin 2017. Participants were recruited to a number of different projects led by MJT and BTD. À
be included in the present analysis, participants were required to be free of MEG or MRI con-
traindications, neurological disorders, and any developmental or psychiatric disorders, and to
Neurosciences en réseau
511
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
have completed a 5-min resting-state acquisition with a static fixation cross. Following exclu-
sion based on task completion and preprocessing of data (including excessive head movement
[>7 mm]), a final cohort of 324 participants (mean age: 18.8, standard deviation: 9.9, 116 fe-
males, range: 5.9 à 45.5 années; for sex distribution see Figure S9, Hunt et al., 2019) étaient
included in the main analyses. All studies received ethical approval from The Hospital for Sick
Children Research Ethics Board.
MEG and MRI Data Acquisition
Five minutes of resting-state MEG data were acquired in supine position using a CTF 151-
channel system (CTF-MISL, Coquitlam, Canada) positioned within a magnetically shielded
room (MSR). Participants were instructed to fixate on a centrally positioned gray cross (+) within
a circle on a black background, back projected onto a screen inside the MSR. Data were
acquired at a sampling frequency of 600 Hz operating in third-order synthetic gradiometry
configuration. Prior to acquisition, participants were fitted with three head position indicator
coils, located at the nasion and left and right preauricular points. These coils were tracked
continuously, enabling a moment-to-moment measurement of head motion. The location of
these coils was recorded, and MRI-visible markers positioned at these locations to coregister
between MEG and MRI data.
An anatomical T1-weighted image was also acquired for all participants using a 3T Mag-
netom Tim Trio (Siemens AG, Erlangen, Allemagne) MRI system using a 12-channel head coil
running an MPRAGE pulse sequence (TR = 2,300 ms; LE = 2.9 ms; flip angle = 9
; field of
view = 240 × 256 × 192 mm; slice thickness 1 mm3).
◦
Preprocessing
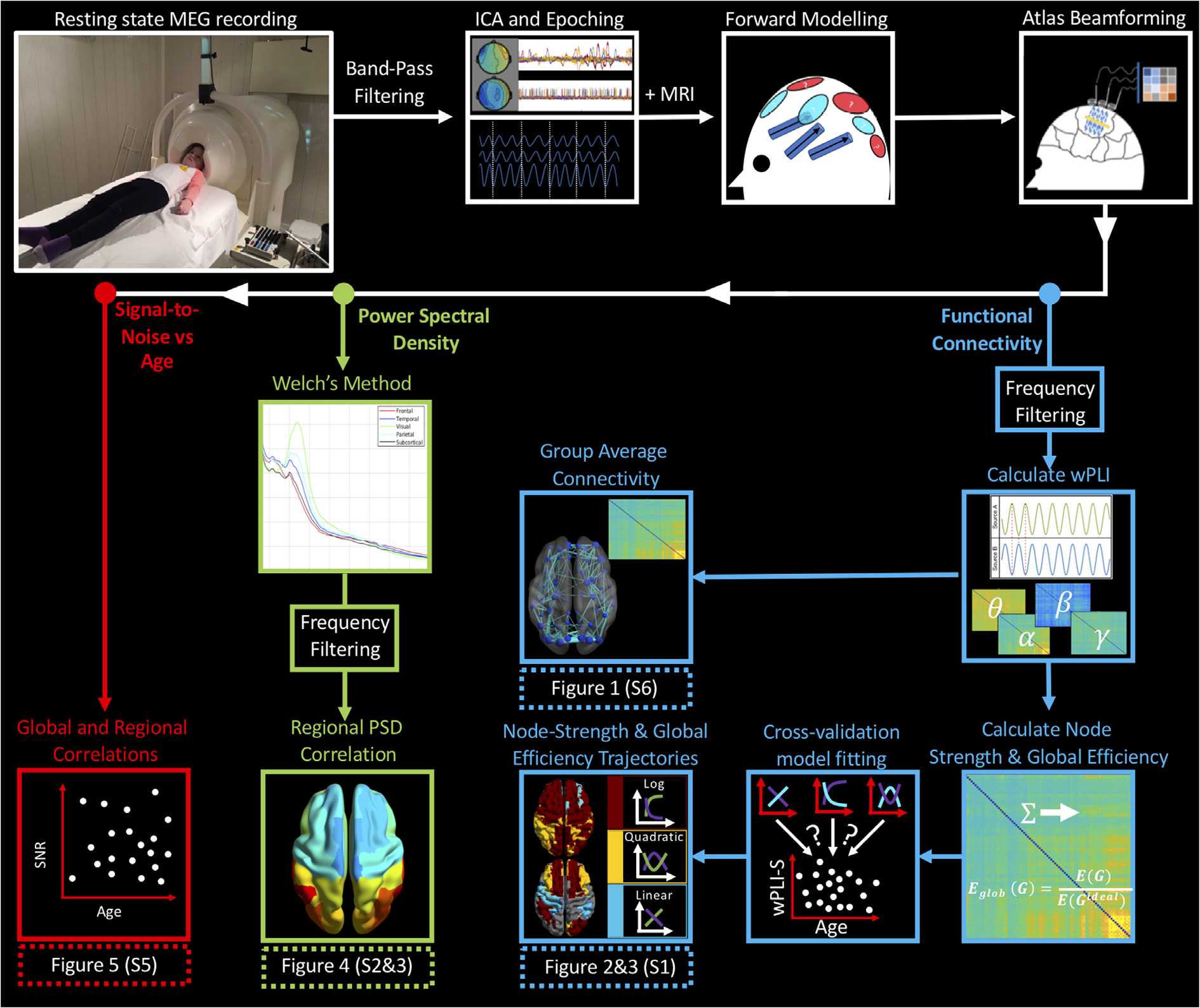
Chiffre 6 presents a schematic of our processing pipeline.
D'abord, data were bandpass filtered between 1 et 150 Hz, and notch filters were applied at
60 et 120 Hz. Deuxième, an independent component analysis (ICA) was performed on MEG
data (Muthukumaraswamy, 2013). A single experimenter examined components and those
deemed arising from eye blinks or cardiac muscles were removed from the data. Suivant, un
automated data rejection algorithm was used to detect and mark segments of data containing
excessive head movement (>7 mm) or artefact. The remaining clean data were epoched into
as many 10-s “trials” as available. Note that these trials are arbitrary divisions of a continuous
resting-state recording and are epoched for computational reasons. Participants with greater
than five trials remaining following this procedure were taken forward for primary analyses.
The mean number of trials per participant was 24.5.
Forward Model and Beamforming
A linearly constrained minimum variance (LCMV; Van Veen, van Drongelen, Yuchtman, &
Suzuki, 1997) spatial filter, implemented in Field Trip (Oostenveld, Fries, Maris, & Schoffelen,
2011), was used to enter source space. The forward model was based upon a dipole approx-
imation (Sarvas, 1987) and a realistically shaped single-shell approximation (Nolte, 2003).
Dipole orientation was determined using the maximum eigenvector and fixed across trials.
Covariance was calculated within a 1–150 Hz covariance window spanning the entire experi-
ment and regularized using the Tikhonov method with a regularization parameter equal to 5%
of the maximum eigenvalue of the unregularized matrix. The LCMV output was normalized
by estimated noise, the result being termed the neural activity index (NAI), to avoid biasing
measurements toward the center of the head. Subject-level data were transformed onto the
Neurosciences en réseau
512
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
t
.
Chiffre 6. Processing pipeline schematic. MEG data were first bandpass filtered, and independent component analysis (ICA) avait l'habitude de
de-noise the data. Following this, data were epoched into 10-s “trials” (for processing efficiency) and beamformed to parcels of the AAL atlas.
Functional connectivity (blue): Data were frequency filtered into bands and functional connectivity calculated using the weighted phase lag
index (wPLI). wPLI-S and GE were entered into a cross-validation curve/model fitting algorithm, which fit either linear, quadratic, or logarithmic
curves to data, quantifying changing graph properties with age. Power spectral density (vert): Using atlas-beamformed data, we used Welch’s
method to calculate PSD. These values were binned into frequency bands and Pearson’s correlation was used to infer relations between PSD
and age. Signal-to-noise (red): Using the lead fields generated to solve the forward problem, we investigated whether any relation between
SNR and age existed, using both global and regional SNR/lead field values. Note that figures in brackets appear in the Supporting Information
section (Hunt et al., 2019).
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
90-parcel AAL atlas (Tzourio-Mazoyer et al., 2002) and a beamformer spatial filter was used
to derive a single signal from each region at the region’s center of mass.
Assessing for Age-Related Confounds
Given that dipolar magnetic field, the source of the MEG signal, falls with a 1
r2 relation, head
size may represent a significant confound in MEG studies of development, as smaller heads
may be accompanied by reduced signal-to-noise ratio. To ensure that our results were not
biased by head size, an assessment of SNR was performed using each subject’s lead fields. Le
forward field (forward model) models the field strength at the sensor level arising from a dipole
of unit strength at a given voxel (Robinson & Rose, 1993). Derivation of forward fields takes
Neurosciences en réseau
513
Determining typical neurophysiological trajectories
account of the subject’s head position relative to MEG sensors. Measurement of the average
forward fields for each subject provides a measure of SNR per subject. We calculated forward
fields from the center of mass of each AAL region for each subject. We then took the root
mean square over orientations and, for the overall relation, the Frobenius norm over channels
and regions, and the mean over channels for individual region correlations. These results were
plotted against age, and bivariate Pearson’s correlation was used to assess for a statistically
significant relationship.
We also assessed for age-related differences in the number of trials per subject. As would
be expected, data from younger subjects were prone to greater head movement and number of
artefacts, therefore more trials from younger subjects were removed during the artefact rejec-
tion procedure. To counteract this, we calculated the average number of trials for participants
less than 11 ans (21 trials) and randomly removed trials from the data of older subjects
with a greater number of trials than this threshold.
Our Supporting Information (Hunt et al., 2019) presents further analyses assessing for age-
related confounds. Figure S5A presents an expanded analysis of the SNR plot from Figure 5.
Here we plot the SNR from exemplar regions, in the frontal and occipital lobes, against partic-
ipant age. Figure S5B assesses the influence of the trial balancing procedure described above.
Plotting the number of trials against age both before and after the balancing reveals the removal
of significant relations between age and the amount of data entered into analyses.
Power Spectral Density (PSD) Calculation
Trial-wise regional time courses, generated by the beamformer spatial filter, were mean-centered.
PSD was then estimated using Welch’s method (‘pwelch’ function in Matlab [MathWorks,
Natick, Etats-Unis]). The resultant values were trial-averaged and portioned into four frequency
bands of interest (theta: 4−7 Hz, alpha: 8−14 Hz, beta: 15−30 Hz, gamma: 31−80 Hz) et
for each, the mean in the frequency domain derived. To characterize the relationship between
age and PSD, bivariate Pearson correlations were computed between participant ages and PSD
estimates for each frequency band and region. Regional PSD correlations were plotted using
Brain Net Viewer (Xia, Wang, & Il, 2013).
Calculating Weighted Phase Lag Index Strength and Global Efficiency
The weighted phase lag index (wPLI) estimates the asymmetry, around zero, of the phase dif-
ference between two signals weighted by the magnitude of the imaginary cross-spectrum. C'est
assumed that a functional connection exists between two regional signals if one signal con-
sistently leads or lags another over time. wPLI values range from 0 à 1, avec 0 indicating a
completely random/symmetric phase distribution to 1, indicating a highly asymmetric distri-
bution. This technique is advantageous as compared with the original phase lag index (Stam,
Nolte, & Daffertshofer, 2007) as it has been shown to be less sensitive to volume conduc-
tion and noise sources more generally (Vinck et al., 2011) and has been used successfully
in many EEG and MEG studies. Trial-wise, frequency filtered, regional time courses were en-
tered into an in-house-coded wPLI routine, exactly following the procedure described in the
original publication (Vinck et al., 2011). Time courses were mean-centered before using the
Hilbert transform to extract the instantaneous phase of the signal for each region-pair, trial,
frequency band, and subject. Trial-wise wPLI values were mean-averaged to generate a single
phase synchronization value per region, frequency band, and subject. This process generated
an adjacency matrix for each subject and frequency band, characterizing whole-brain phase
synchronization. To generate the grand-average adjacency matrices in Figure 1, a mean-average
Neurosciences en réseau
514
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
across all participants was derived. Glass brains were plotted using Brain Net Viewer software
(Xia et al., 2013). To calculate wPLI-strength (wPLI-S), the weights for each regional AAL node
were summed in each frequency band, generating an [AAL nodes (90) × frequency bands (4)]
array of values for each subject; this is equivalent to summing across a single dimension of
the adjacency matrix. To calculate global efficiency (GE), we used the corresponding function
(‘charpath’) in Brain Connectivity Toolbox (Rubinov & Sporns, 2010). This function requires
a distance matrix, which was created by taking the inverse of the individual wPLI adjacency
matrices and using the ‘distance_wei’ function implemented in Brain Connectivity Toolbox.
Curve Fitting Algorithm
D'abord, we constructed models for the linear (1), quadratic (2), and logarithmic (3) curves as
follows:
y = a
X + un
1
x2 + b
2,
X + b
2
X + c
3,
3) + c
(c
2
y = b
1
1log10
y = c
4,
(1)
(2)
(3)
1−2, b
1−3, and c
where x represents subject age, un
1−4 represent coefficients, and y represents
our wPLI-S or GE measurements for all participants. Alors, for each frequency, and for wPLI-S
region, we performed a cross-validation routine. D'abord, we randomly split subject-level data into
two equal groups, termed Fit and Test. Deuxième, all three models were fitted to the Fit group.
Troisième, these same models were then fitted to the Test group and the residual difference, derived
as the absolute difference between the model and the data, was recorded. This process was
repeated 1,000 times, avec 1,000 different random group allocations. Following completion
of the iterations, the best fitting model was chosen as that with the lowest median residual. Par
taking the median residual across iterations, we protect against overfitting and ensure that the
model selection is not biased toward more complex model forms. Note that the coefficients
for all best fitting models are detailed in Table S10 (Hunt et al., 2019).
Determining Statistical Significance
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
un
_
0
0
0
7
7
p
d
t
.
To determine whether the curve characterizing the relationship between wPLI-S/GE and age
was significantly better than chance, we derived an empirical null distribution. This was based
on the reasoning that if age had no effect on the graph metric, then randomly shuffling the
ages would have no effect on the estimated curve F statistic. D'abord, the best fitting model was
fit to the entire dataset for a region and frequency band and the F statistic derived. Suivant,
the best fitting model was fit to data with the ages randomly shuffled and a null F statistic
calculated. This process was repeated 15,000 times to generate a distribution of null F statistics.
A significance value was then derived as the proportion of null F statistics less than that for the
nonshuffled data. To control for multiple comparisons across regions, strength measurements
were thresholded using a false discovery rate, implemented using the Benjamini-Hochberg
procedure, at q < 0.05.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Calculating Curve Gradients
To visualize the trajectories more clearly, we calculated the gradients (∇) of each curve type
for each year of age (xn),
Network Neuroscience
∇linear = a
1,
xn) + b
1
c
2
xn) ln (10) + c
1
2,
.
3
∇quadratic = 2(b
c
∇
log
=
(c
2
(4)
(5)
(6)
515
Determining typical neurophysiological trajectories
We used the calculated gradients to visualize maturation, both on static brains and as videos
(S7 and S8; Hunt et al., 2019), using Brain Net Viewer (Xia et al., 2013), where each frame
(1/s) presents the gradient values for each region in that year of age.
ACKNOWLEDGMENTS
We wish to acknowledge the following individuals who acquired the data used in this publi-
cation: Rachel Leung, Vanessa Vogan, Sarah Mossad, Julie Sato, Veronica Yuk, MyLoi Huynh,
and Julie Lu. We also wish to thank the participants who kindly donated their time to multi-
ple studies at The Hospital for Sick Children. Finally, we wish to acknowledge Professor Krish
Singh of Cardiff University Brain Research Imaging Centre (CUBRIC), who kindly shared his
code for plotting the circular connectivity graphs.
AUTHOR CONTRIBUTIONS
Benjamin Alexander Edward Hunt: Conceptualization; Data curation; Formal analysis; Investi-
gation; Methodology; Project administration; Software; Visualization; Writing – original draft;
Writing – review & editing. Simeon M. Wong: Data curation; Formal analysis; Methodology;
Software; Writing – review & editing. Marlee M. Vandewouw: Methodology; Software; Visu-
alization; Writing – review & editing. Matthew J. Brookes: Conceptualization; Methodology;
Supervision; Writing – review & editing. Benjamin T. Dunkley: Conceptualization; Method-
ology; Supervision; Writing – review & editing. Margot J. Taylor: Conceptualization; Funding
acquisition; Methodology; Supervision; Writing – original draft; Writing – review & editing.
FUNDING INFORMATION
Margot
J. Taylor, Canadian Institutes of Health Research (http://dx.doi.org/10.13039/
501100000024), Award ID: MOP-106582. Margot J. Taylor, Canadian Institutes of Health Re-
search (http://dx.doi.org/10.13039/501100000024), Award ID: MOP-119541. Margot J. Taylor,
Canadian Institutes of Health Research (http://dx.doi.org/10.13039/501100000024), Award
ID: MOP-137115. Margot J. Taylor, Canadian Institutes of Health Research, Award ID: MOP-
142379. Benjamin Alexander Edward Hunt, Hospital for Sick Children (http://dx.doi.org/10.
13039/501100006126), Award ID: Restracomp. Benjamin Alexander Edward Hunt, Hospital
for Sick Children (http://dx.doi.org/10.13039/501100006126), Award ID: The Centre for Brain
and Mental Health Abe Bresver Fellowship.
REFERENCES
Adleman, N. E., Menon, V., Blasey, C. M., White, C. D., Warsofsky,
I. S., Glover, G. H., & Reiss, A. L. (2002). A developmental fMRI
study of the Stroop color-word task. NeuroImage, 16(1), 61–75.
https://doi.org/10.1006/nimg.2001.1046
Benninger, C., Matthis, P., & Scheffner, D. (1984). EEG develop-
ment of healthy boys and girls. Results of a longitudinal study.
Electroencephalography and Clinical Neurophysiology, 57(1),
1–12.
Biswal, B., Yetkin, F. Z., Haughton, V. M., & Hyde, J. S. (1995).
Functional connectivity in the motor cortex of resting human
brain using echo-planar MRI. Magnetic Resonance in Medicine,
34(4), 537–541. https://doi.org/10.1002/mrm.1910340409
Bollimunta, A., Mo, J., Schroeder, C. E., & Ding, M. (2011). Neu-
ronal mechanisms and attentional modulation of corticothalamic
alpha oscillations. Journal of Neuroscience, 31(13), 4935–4943.
https://doi.org/10.1523/JNEUROSCI.5580-10.2011
Boto, E., Bowtell, R., Krüger, P., Fromhold, T. M., Morris, P. G.,
Meyer, S. S., . . . Brookes, M. J. (2016). On the potential of a new
generation of magnetometers for MEG: A beamformer simula-
tion study. PLoS ONE, 11(8), e0157655. https://doi.org/10.1371/
journal.pone.0157655
Boto, E., Holmes, N., Leggett, J., Roberts, G., Shah, V., Meyer, S. S.,
. . . Brookes, M. J.
(2018). Moving magnetoencephalography
towards real-world applications with a wearable system. Nature,
555(7698), 657–661. https://doi.org/10.1038/nature26147
Brookes, M. J., Groom, M. J., Liuzzi, L., Hill, R. M., Smith, H. J. F.,
Briley, P. M., . . . Liddle, E. B. (2018). Altered temporal stability
in dynamic neural networks underlies connectivity changes in
neurodevelopment. NeuroImage, 174, 563–575. https://doi.org/
10.1016/j.neuroimage.2018.03.008
Brookes, M. J., Tewarie, P. K., Hunt, B. A. E., Robson, S. E.,
Gascoyne, L. E., Liddle, E. B., . . . Morris, P. G. (2016). A multi-layer
Network Neuroscience
516
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
a
_
0
0
0
7
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
network approach to MEG connectivity analysis. NeuroImage,
132, 425–438.https://doi.org/10.1016/j.neuroimage.2016.02.045
Brookes, M. J., Woolrich, M., Luckhoo, H., Price, D., Hale, J. R.,
Stephenson, M. C., . . . Morris, P. G. (2011). Investigating the
electrophysiological basis of resting state networks using mag-
netoencephalography. Proceedings of the National Academy of
Sciences, 108(40), 16783–16788. https://doi.org/10.1073/pnas.
1112685108
Buffalo, E. A., Fries, P., Landman, R., Buschman, T. J., & Desimone,
R. (2011). Laminar differences in gamma and alpha coherence
in the ventral stream. Proceedings of the National Academy of
Sciences, 108(27), 11262–11267. https://doi.org/10.1073/pnas.
1011284108
Cao, M., Wang,
J.-H., Dai, Z.-J., Cao, X.-Y., Jiang, L.-L., Fan,
F.-M., . . . He, Y.
(2014). Topological organization of the hu-
man brain functional connectome across the lifespan. Develop-
mental Cognitive Neuroscience, 7, 76–93. https://doi.org/10.1016/
j.dcn.2013.11.004
Chugani, H. T., Phelps, M. E., & Mazziotta, J. C. (1987). Positron
emission tomography study of human brain functional devel-
opment. Annals of Neurology, 22(4), 487–497. https://doi.org/
10.1002/ana.410220408
Clarke, A. R., Barry, R. J., McCarthy, R., & Selikowitz, M. (2001).
Age and sex effects in the EEG: Development of the normal
child. Clinical Neurophysiology, 112(5), 806–814. https://doi.org/
10.1016/S1388-2457(01)00488-6
de Pasquale, F., Della Penna, S., Sporns, O., Romani, G. L., &
Corbetta, M.
(2016). A dynamic core network and global
efficiency in the resting human brain. Cerebral Cortex, 26(10),
4015–4033. https://doi.org/10.1093/cercor/bhv185
Dias, N. S., Ferreira, D., Reis, J., Jacinto, L. R., Fernandes, L., Pinho,
F., . . . Sousa, N. (2015). Age effects on EEG correlates of the
Wisconsin Card Sorting Test. Physiological Reports, 3(7), e12390.
https://doi.org/10.14814/phy2.12390
Dosenbach, N. U. F., Nardos, B., Cohen, A. L., Fair, D. A., Power,
J. D., Church, J. A., . . . Schlaggar, B. L. (2010). Prediction
of individual brain maturity using fMRI. Science, 329(5997),
1358–1361. https://doi.org/10.1126/science.1194144
Douaud, G., Groves, A. R., Tamnes, C. K., Westlye, L. T., Duff,
E. P., Engvig, A., . . . Johansen-Berg, H. (2014). A common brain
network links development, aging, and vulnerability to disease.
Proceedings of the National Academy of Sciences, 111(49),
17648–17653. https://doi.org/10.1073/pnas.1410378111
Dougherty, K., Cox, M. A., Ninomiya, T., Leopold, D. A., & Maier,
A. (2017). Ongoing alpha activity in V1 regulates visually driven
spiking responses. Cerebral Cortex, 27(2), 1113–1124. https://
doi.org/10.1093/cercor/bhv304
Dunkley, B. T., Doesburg, S. M., Jetly, R., Sedge, P. A., Pang, E. W., &
Taylor, M. J. (2015). Characterising intra- and inter-intrinsic net-
work synchrony in combat-related post-traumatic stress disorder.
Psychiatry Research, 234(2), 172–181. https://doi.org/10.1016/
j.pscychresns.2015.09.002
Dunkley, B. T., Pang, E. W., Sedge, P. A., Jetly, R., Doesburg, S. M.,
& Taylor, M. J. (2016). Threatening faces induce fear circuitry
hypersynchrony in soldiers with post-traumatic stress disorder.
Heliyon, 2(1), e00063. https://doi.org/10.1016/j.heliyon.2015.
e00063
Fox, M. D., & Raichle, M. E. (2007). Spontaneous fluctuations
in brain activity observed with functional magnetic resonance
imaging. Nature Reviews Neuroscience, 8, 700–711. https://doi.
org/10.1038/nrn2201
Freunberger, R., Fellinger, R., Sauseng, P., Gruber, W., & Klimesch,
W. (2009). Dissociation between phase-locked and nonphase-
locked alpha oscillations in a working memory task. Human Brain
Mapping, 30(10),3417–3425. https://doi.org/10.1002/hbm.20766
Fries, P. (2005). A mechanism for cognitive dynamics: Neuronal
communication through neuronal coherence. Trends in Cog-
nitive Sciences, 9(10), 474–480. https://doi.org/10.1016/j.tics.
2005.08.011
Fries, P. (2015). Rhythms for cognition: Communication through
coherence. Neuron, 88( 1), 220–235. https://doi.org/10.1016/
j.neuron.2015.09.034
Gaetz, W., Gordon, R. S., Papadelis, C., Fujiwara, H., Rose, D. F.,
Edgar, J. C., . . . Roberts, T. P. L. (2015). Magnetoencephalogra-
phy for clinical pediatrics: Recent advances in hardware, meth-
ods, and clinical applications. Journal of Pediatric Epilepsy, 4(4),
139–155. https://doi.org/10.1055/s-0035-1563726
Geades, N., Hunt, B. A. E., Shah, S. M., Peters, A., Mougin, O. E., &
Gowland, P. A. (2016). Quantitative analysis of the z-spectrum
using a numerically simulated look-up table: Application to the
healthy human brain at 7T. Magnetic Resonance in Medicine,
78(2), 645–655. https://doi.org/10.1002/mrm.26459
Giedd, J. N. (1999). Brain development, IX: Human brain growth.
American Journal of Psychiatry, 156(1), 4. https://doi.org/10.1176/
ajp.156.1.4
Giedd, J. N., & Rapoport, J. L. (2010). Structural MRI of pediatric
brain development: What have we learned and where are we go-
ing? Neuron, 67(5), 728–734. https://doi.org/10.1016/j.neuron.
2010.08.040
Gogtay, N., Giedd, J. N., Lusk, L., Hayashi, K. M., Greenstein,
D., Vaituzis, A. C., . . . Thompson, P. M. (2004). Dynamic
mapping of human cortical development during childhood
through early adulthood. Proceedings of the National Academy
of Sciences, 101(21), 8174–8179. https://doi.org/10.1073/pnas.
0402680101
Gómez, C. M., Rodríguez-Martínez, E. I., Fernández, A., Maestú,
J., & Gómez, C. (2017). Absolute power spectral
F., Poza,
density changes in the magnetoencephalographic activity during
the transition from childhood to adulthood. Brain Topography,
30(1), 87–97. https://doi.org/10.1007/s10548-016-0532-0
Gotts, S. J., Jo, H. J., Wallace, G. L., Saad, Z. S., Cox, R. W., &
Martin, A. (2013). Two distinct forms of functional lateralization
in the human brain. Proceedings of the National Academy of
Sciences, 110(36), E3435–E3444. https://doi.org/10.1073/pnas.
1302581110
Grayson, D. S., & Fair, D. A. (2017). Development of large-scale
functional networks from birth to adulthood: A guide to the
neuroimaging literature. NeuroImage, 160, 15–31. https://doi.
org/10.1016/j.neuroimage.2017.01.079
Hall, E. L., Robson, S. E., Morris, P. G., & Brookes, M. J. (2014).
The relationship between MEG and fMRI. NeuroImage, 102(Pt. 1),
80–91. https://doi.org/10.1016/j.neuroimage.2013.11.005
Hämäläinen, M., Hari, R.,
J., &
Lounasmaa, O. V. (1993). Magnetoencephalography: Theory,
Ilmoniemi, R.
J., Knuutila,
Network Neuroscience
517
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
a
_
0
0
0
7
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
instrumentation, and applications to noninvasive studies of
the working human brain. Reviews of Modern Physics, 65(2),
413–497. https://doi.org/10.1103/RevModPhys.65.413
He, W., Sowman, P. F., Brock, J., Etchell, A. C., Stam, C. J., &
Hillebrand, A. (2018). Topological segregation of functional net-
works increases in developing brains. bioRxiv:378562. https://
doi.org/10.1101/378562
Holmes, N., Leggett,
J., Boto, E., Roberts, G., Hill, R. M.,
Tierney, T. M., . . . Bowtell, R. (2018). A bi-planar coil system
for nulling background magnetic fields in scalp mounted magne-
toencephalography. NeuroImage, 181, 760–774. https://doi.org/
10.1016/j.neuroimage.2018.07.028
Houghton, G., & Tipper, S. P. (1996). Inhibitory mechanisms of neu-
ral and cognitive control: Applications to selective attention and
sequential action. Brain and Cognition, 30(1), 20–43. https://doi.
org/10.1006/brcg.1996.0003
Hunt, B. A. E., Tewarie, P. K., Mougin, O. E., Geades, N.,
Jones, D. K., Singh, K. D., . . . Brookes, M. J. (2016). Rela-
tionships between cortical myeloarchitecture and electrophys-
iological networks. Proceedings of the National Academy of
Sciences, 113(47), 13510–13515. https://doi.org/10.1073/pnas.
1608587113
Hunt, B. A. E., Wong, S. M., Vandewouw, M. M., Brookes, M. J.,
Dunkley, B. T., & Taylor, M. J. (2019). Supporting information
for “Spatial and spectral trajectories in typical neurodevelop-
ment from childhood to middle age.” Network Neuroscience,
3(2), 497–520. https://doi.org/10.1162/netn_a_00077
Jensen, O., Gelfand, J., Kounios, J., & Lisman, J. E. (2002). Oscil-
lations in the alpha band (9–12 Hz) increase with memory load
during retention in a short-term memory task. Cerebral Cortex,
12(8), 877–882. https://doi.org/10.1093/cercor/12.8.877
Kaleta, T., & Marczewska, S. (2007). The observations concern-
ing reproduction and young development
in captive giraffe
(Giraffa camelopardalis) at Warsaw ZOO. Annals of Warsaw
University of Life Sciences - SGGW. Animal Science, (44). Re-
trieved from https://www.infona.pl//resource/bwmeta1.element.
agro-ea9ab271-54c8-4344-81a8-e7e98f1b90be
Khan, S., Hashmi, J. A., Mamashli, F., Michmizos, K., Kitzbichler,
M. G., Bharadwaj, H., . . . Kenet, T. (2018). Maturation trajec-
tories of cortical resting-state networks depend on the me-
diating frequency band. NeuroImage, 174, 57–68. https://doi.org/
10.1016/j.neuroimage.2018.02.018
Klimesch, W. (2012). Alpha-band oscillations, attention, and con-
trolled access to stored information. Trends in Cognitive Sciences,
16(12), 606–617. https://doi.org/10.1016/j.tics.2012.10.007
Lopes da Silva, F. (2013). EEG and MEG: Relevance to neuro-
science. Neuron, 80( 5), 1112–1128. https://doi.org/10.1016/
j.neuron.2013.10.017
Menon, V., & Desmond, J. E. (2001). Left superior parietal cortex in-
volvement in writing: Integrating fMRI with lesion evidence. Cog-
nitive Brain Research, 12(2), 337–340. https://doi.org/10.1016/
S0926-6410(01)00063-5
Miskovic, V., Ma, X., Chou, C.-A., Fan, M., Owens, M., Sayama, H.,
& Gibb, B. E. (2015). Developmental changes in spontaneous
electrocortical activity and network organization from early to
late childhood. NeuroImage, 118, 237–247. https://doi.org/10.
1016/j.neuroimage.2015.06.013
Muthukumaraswamy, S. D. (2013). High-frequency brain activity
and muscle artifacts in MEG/EEG: A review and recommenda-
tions. Frontiers in Human Neuroscience, 7. https://doi.org/10.
3389/fnhum.2013.00138
Nolte, G. (2003). The magnetic lead field theorem in the quasi-static
approximation and its use for magnetoencephalography forward
calculation in realistic volume conductors. Physics in Medicine
and Biology, 48(22), 3637.
Nugent, A. C., Robinson, S. E., Coppola, R., Furey, M. L., & Zarate,
C. A. (2015). Group differences in MEG-ICA derived resting state
networks: Application to major depressive disorder. NeuroImage,
118, 1–12. https://doi.org/10.1016/j.neuroimage.2015.05.051
Oostenveld, R., Fries, P., Maris, E., & Schoffelen, J.-M. (2011). Field-
Trip: Open source software for advanced analysis of MEG, EEG,
and invasive electrophysiological data [Research article]. https://
doi.org/10.1155/2011/156869
Ornstein, P. A., & Light, L. L. (2010). Memory development across
the life span. In W. Overton (Ed.), The handbook of life-span
development. Hoboken, NJ: John Wiley & Sons. https://doi.org/
10.1002/9780470880166.hlsd001009
Pang, E. W., Dunkley, B. T., Doesburg, S. M., da Costa, L., & Taylor,
M. J. (2016). Reduced brain connectivity and mental flexibility in
mild traumatic brain injury. Annals of Clinical and Translational
Neurology, 3(2), 124–131. https://doi.org/10.1002/acn3.280
Paus, T., Collins, D. L., Evans, A. C., Leonard, G., Pike, B., &
Zijdenbos, A. (2001). Maturation of white matter in the human
brain: A review of magnetic resonance studies. Brain Research
Bulletin, 54(3), 255–266.
Perone, S., Palanisamy, J., & Carlson, S. M. (2018). Age-related
change in brain rhythms from early to middle childhood: Links
to executive function. Developmental Science, e12691. https://
doi.org/10.1111/desc.12691
Pfurtscheller, G., & Lopes da Silva, F. H. (1999). Event-related
EEG/MEG synchronization and desynchronization: Basic princi-
ples. Clinical Neurophysiology, 110(11), 1842–1857. https://doi.
org/10.1016/S1388-2457(99)00141-8
Remer, J., Croteau-Chonka, E., Dean, D. C., D’Arpino, S., Dirks,
H., Whiley, D., & Deoni, S. C. L. (2017). Quantifying cortical de-
velopment in typically developing toddlers and young children,
1–6 years of age. NeuroImage, 153, 246–261. https://doi.org/
10.1016/j.neuroimage.2017.04.010
Robinson, S. E., & Rose, D. F. (1993). Current source image esti-
mation by spatially filtered MEG. In M. Hoke, S. N. Erné, Y. C.
Okada, & G. L. Romani (Eds.), Biomagnetism: Clinical aspects
(pp. 761–765). Amsterdam, Netherlands: Elsevier Science Publishers.
Rodriguez-Martinez, E. I., Barriga-Paulino, C. I., Rojas-Benjumea,
M. A., & Gómez, C. M. (2015). Co-maturation of theta and
low-beta rhythms during child development. Brain Topography,
28(2), 250–260. https://doi.org/10.1007/s10548-014-0369-3
J., Nosarti, C., Heyman,
I.,
Taylor, E., & Brammer, M. (2006). Progressive increase of fron-
tostriatal brain activation from childhood to adulthood during
event-related tasks of cognitive control. Human Brain Mapping,
27(12), 973–993. https://doi.org/10.1002/hbm.20237
Rubia, K., Smith, A. B., Woolley,
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003
Network Neuroscience
518
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
a
_
0
0
0
7
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
Sarvas, J. (1987). Basic mathematical and electromagnetic concepts
of the biomagnetic inverse problem. Physics in Medicine and Bi-
ology, 32(1), 11. https://doi.org/10.1088/0031-9155/32/1/004
Sato, J., Mossad, S. I., Wong, S. M., Hunt, B. A. E., Dunkley,
B. T., Smith, M. L., . . . Taylor, M. J. (2018). Alpha keeps it
together: Alpha oscillatory synchrony underlies working memory
maintenance in young children. Developmental Cognitive
Neuroscience, 34, 114–123. https://doi.org/10.1016/j.dcn.2018.
09.001
Sato, J. R., Salum, G. A., Gadelha, A., Picon, F. A., Pan, P. M.,
Vieira, G., . . . Jackowski, A. P. (2014). Age effects on the de-
fault mode and control networks in typically developing chil-
dren. Journal of Psychiatric Research, 58, 89–95. https://doi.org/
10.1016/j.jpsychires.2014.07.004
Schäfer, C. B., Morgan, B. R., Ye, A. X., Taylor, M.
J., &
Doesburg, S. M. (2014). Oscillations, networks, and their devel-
opment: MEG connectivity changes with age. Human Brain Map-
ping, 35(10), 5249–5261. https://doi.org/10.1002/hbm.22547
Shafee, R., Buckner, R. L., & Fischl, B. (2015). Gray matter myeli-
nation of 1555 human brains using partial volume corrected
MRI images. NeuroImage, 105, 473–485. https://doi.org/10.1016/
j.neuroimage.2014.10.054
Shaw, P., Kabani, N. J., Lerch, J. P., Eckstrand, K., Lenroot, R.,
Gogtay, N., . . . Wise, S. P. (2008). Neurodevelopmental tra-
jectories of the human cerebral cortex. Journal of Neuroscience,
28(14), 3586–3594. https://doi.org/10.1523/JNEUROSCI.5309-
07.2008
Smit, D. J. A., Boersma, M., Schnack, H. G., Micheloyannis, S.,
Boomsma, D. I., Hulshoff Pol, H. E., . . . de Geus, E. J. C. (2012).
The brain matures with stronger functional connectivity and de-
creased randomness of its network. PLoS ONE, 7(5), e36896.
https://doi.org/10.1371/journal.pone.0036896
Smits-Engelsman, B. C. M., & Wilson, P. H. (2013). Age-related
changes in motor imagery from early childhood to adulthood:
Probing the internal representation of speed-accuracy trade-offs.
Human Movement Science, 32(5), 1151–1162. https://doi.org/
10.1016/j.humov.2012.06.006
Sporns, O. (2011). The human connectome: A complex network.
Annals of the New York Academy of Sciences, 1224, 109–125.
https://doi.org/10.1111/j.1749-6632.2010.05888.x
Stam, C. J., Nolte, G., & Daffertshofer, A. (2007). Phase lag in-
dex: Assessment of functional connectivity from multi channel
EEG and MEG with diminished bias from common sources.
Human Brain Mapping, 28(11), 1178–1193. https://doi.org/10.
1002/hbm.20346
Størvold, G. V., Aarethun, K., & Bratberg, G. H. (2013). Age for
onset of walking and prewalking strategies. Early Human Devel-
opment, 89(9), 655–659. https://doi.org/10.1016/j.earlhumdev.
2013.04.010
Sun, W., & Dan, Y. (2009). Layer-specific network oscillation and
spatiotemporal receptive field in the visual cortex. Proceedings
of the National Academy of Sciences, 106(42), 17986–17991.
https://doi.org/10.1073/pnas.0903962106
Supekar, K., Musen, M., & Menon, V. (2009). Development of large-
scale functional brain networks in children. PLoS Biology, 7(7),
e1000157. https://doi.org/10.1371/journal.pbio.1000157
Tau, G. Z., & Peterson, B. S. (2010). Normal development of brain
circuits. Neuropsychopharmacology, 35(1), 147–168. https://
doi.org/10.1038/npp.2009.115
Tewarie, P. K., Bright, M. G., Hillebrand, A., Robson, S. E.,
Gascoyne, L. E., Morris, P. G., . . . Brookes, M. J. (2016). Predict-
ing haemodynamic networks using electrophysiology: The role of
non-linear and cross-frequency interactions. NeuroImage, 130,
273–292. https://doi.org/10.1016/j.neuroimage.2016.01.053
Tewarie, P. K., Hillebrand, A., van Dellen, E., Schoonheim, M. M.,
Barkhof, F., Polman, C. H., . . . Stam, C. J.
(2014). Structural
degree predicts functional network connectivity: A multimodal
resting-state fMRI and MEG study. NeuroImage, 97, 296–307.
https://doi.org/10.1016/j.neuroimage.2014.04.038
Tottenham, N. (2014). The importance of early experiences for neuro-
affective development. Current Topics in Behavioral Neuro-
sciences, 16, 109–129. https://doi.org/10.1007/7854_2013_254
Troebinger, L., López, J. D., Lutti, A., Bestmann, S., & Barnes, G.
(2014). Discrimination of cortical laminae using MEG. Neuro-
Image, 102(2), 885–893. https://doi.org/10.1016/j.neuroimage.
2014.07.015
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., . . . Joliot, M. (2002). Automated
anatomical
labeling of activations in SPM using a macro-
scopic anatomical parscellation of the MNI MRI single-subject
brain. NeuroImage, 15(1), 273–289. https://doi.org/10.1006/
nimg.2001.0978
Uhlhaas, P. J.
(2011). The adolescent brain: Implications for the
understanding, pathophysiology, and treatment of schizophrenia.
Schizophrenia Bulletin, 37(3), 480–483. https://doi.org/10.1093/
schbul/sbr025
Uhlhaas, P. J., Haenschel, C., Nikoli´c, D., & Singer, W. (2008).
The role of oscillations and synchrony in cortical networks and
their putative relevance for the pathophysiology of schizophre-
nia. Schizophrenia Bulletin, 34(5), 927–943. https://doi.org/10.
1093/schbul/sbn062
Urbain, C., Vogan, V. M., Ye, A. X., Pang, E. W., Doesburg, S. M.,
& Taylor, M. J. (2016). Desynchronization of fronto-temporal
networks during working memory processing in autism. Human
Brain Mapping, 37(1), 153–164. https://doi.org/10.1002/hbm.
23021
Vandekar, S. M., Shinohara, R. T., Raznahan, A., Roalf, D. R., Ross,
M., Nicholas, D. L., . . . Satterthwaite, T. D. (2015). Topologically
dissociable patterns of development of the human cerebral cor-
tex. Journal of Neuroscience, 35(2), 599–609. https://doi.org/10.
1523/JNEUROSCI.3628-14.2015
Van Veen, B. D., van Drongelen, W., Yuchtman, M., & Suzuki,
A. (1997). Localization of brain electrical activity via linearly
constrained minimum variance spatial filtering. IEEE Transactions
on Bio-Medical Engineering, 44(9), 867–880. https://doi.org/10.
1109/10.623056
Vinck, M., Oostenveld, R., van Wingerden, M., Battaglia, F.,
& Pennartz, C. M. A. (2011). An improved index of phase-
synchronization for electrophysiological data in the presence of
volume-conduction, noise and sample-size bias. NeuroImage,
55(4), 1548–1565. https://doi.org/10.1016/j.neuroimage.2011.
01.055
Network Neuroscience
519
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
a
_
0
0
0
7
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Determining typical neurophysiological trajectories
Vlahou, E. L., Thurm, F., Kolassa, I.-T., & Schlee, W. (2014). Resting-
state slow wave power, healthy aging and cognitive performance.
Scientific Reports, 4. https://doi.org/10.1038/srep05101
Weinrich, C. A., Brittain, J.-S., Nowak, M., Salimi-Khorshidi, R.,
Brown, P., & Stagg, C. J. (2017). Modulation of long-range con-
nectivity patterns via frequency-specific stimulation of human
cortex. Current Biology, 27(19), 3061-3068.e3. https://doi.org/
10.1016/j.cub.2017.08.075
Whitford, T. J., Rennie, C. J., Grieve, S. M., Clark, C. R., Gordon,
E., & Williams, L. M. (2007). Brain maturation in adolescence:
Concurrent changes in neuroanatomy and neurophysiology.
Human Brain Mapping, 28(3), 228–237. https://doi.org/10.1002/
hbm.20273
Winkler, A. M., Kochunov, P., Blangero, J., Almasy, L., Zilles, K., Fox,
P. T., . . . Glahn, D. C. (2010). Cortical thickness or grey matter
volume? the importance of selecting the phenotype for imaging
genetics studies. NeuroImage, 53(3), 1135–1146. https://doi.org/
10.1016/j.neuroimage.2009.12.028
Xia, M., Wang, J., & He, Y. (2013). BrainNet Viewer: A network vi-
sualization tool for human brain connectomics. PLoS ONE, 8(7),
e68910. https://doi.org/10.1371/journal.pone.0068910
Ye, A. X., AuCoin-Power, M., Taylor, M. J., & Doesburg, S. M.
(2016). Disconnected neuromagnetic networks in children born
very preterm: Disconnected MEG networks in preterm chil-
dren. NeuroImage. Clinical, 11, 376–384. https://doi.org/10.1016/
j.nicl.2015.08.016
Ye¸silyurt, B., Whittingstall, K., Uˇgurbil, K., Logothetis, N. K., &
Uludaˇg, K.
(2010). Relationship of the BOLD signal with VEP
for ultrashort duration visual stimuli (0.1 to 5 ms) in humans.
Journal of Cerebral Blood Flow and Metabolism, 30(2), 449–458.
https://doi.org/10.1038/jcbfm.2009.224
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
4
9
7
1
0
9
2
6
7
1
n
e
n
_
a
_
0
0
0
7
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
520