RECHERCHE
Functional connectivity-based subtypes of
individuals with and without autism
spectrum disorder
Amanda K. Easson1,2, Zainab Fatima3, and Anthony R. McIntosh1,2
1Rotman Research Institute, Baycrest Hospital, Toronto, ON, Canada
2Département de psychologie, University of Toronto, Toronto, ON, Canada
3Département de psychologie, Faculty of Health, Sherman Health Sciences Centre, York University, Toronto, ON, Canada
un accès ouvert
journal
Mots clés: Autism spectrum disorder, Functional connectivity, Clustering, Brain-behavior
relationships, Multivariate statistics, Resting-state networks
ABSTRAIT
Autism spectrum disorder (ASD) is a heterogeneous neurodevelopmental disorder,
characterized by impairments in social communication and restricted, repetitive behaviors.
Neuroimaging studies have shown complex patterns and functional connectivity (FC) in ASD,
with no clear consensus on brain-behavior relationships or shared patterns of FC with
typically developing controls. Ici, we used a dimensional approach to characterize two
distinct clusters of FC patterns across both ASD participants and controls using k-means
clustering. Using multivariate statistical analyses, a categorical approach was taken to
characterize differences in FC between subtypes and between diagnostic groups. Un
subtype was defined by increased FC within resting-state networks and decreased FC across
networks compared with the other subtype. A separate FC pattern distinguished ASD from
controls, particularly within default mode, cingulo-opercular, sensorimotor, and occipital
réseaux. There was no significant interaction between subtypes and diagnostic groups.
Enfin, a dimensional analysis of FC patterns with behavioral measures of IQ, sociale
responsiveness, and ASD severity showed unique brain-behavior relations in each subtype
and a continuum of brain-behavior relations from ASD to controls within one subtype.
These results demonstrate that distinct clusters of FC patterns exist across ASD and controls,
and that FC subtypes can reveal unique information about brain-behavior relationships.
RÉSUMÉ DE L'AUTEUR
Autism spectrum disorder (ASD) is a neurodevelopmental disorder, with high variation in the
types of severity of impairments in social communication and restricted, repetitive behaviors.
Neuroimaging studies have shown complex patterns of communication between brain
régions, or functional connectivity (FC), in ASD. Ici, we defined two distinct FC patterns
and relationships between FC and behavior in a group of participants consisting of individuals
with and without ASD. One subtype was defined by increased FC within distinct networks
of brain regions and decreased FC between networks compared with the other subtype.
A separate FC pattern distinguished ASD from controls. The interaction between subtypes
and diagnostic groups was not significant. Dimensional analyses of FC patterns with
behavioral measures revealed unique information about brain-behavior relations in each
subtype.
Citation: Easson, UN. K., Fatima, Z., &
McIntosh, UN. R.. (2019). Functional
connectivity-based subtypes of
individuals with and without autism
spectrum disorder. Réseau
Neurosciences, 3(2), 344–362.
https://doi.org/10.1162/netn_a_00067
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00067
Informations complémentaires:
https://doi.org/10.1162/netn_a_00067
Reçu: 1 Juin 2018
Accepté: 16 Août 2018
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Auteur correspondant:
Amanda Easson
amanda.easson@mail.utoronto.ca
Éditeur de manipulation:
Alex Fornito
droits d'auteur: © 2018
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
un
_
0
0
0
6
7
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
Functional connectivity:
The statistical association between
the time series of activity of two brain
régions.
INTRODUCTION
Autism spectrum disorder (ASD) is a neurodevelopmental disorder that is characterized by im-
pairments in social cognition as well as restricted and repetitive behaviors (RRBs; Américain
Psychiatric Association, 2013). ASD is highly heterogeneous, with a broad range of the types
and severities of behaviors that can be displayed. Par exemple, verbal and nonverbal IQ are
highly variable in ASD (par exemple., Munson et al., 2008), and RRBs can range from low-level stereo-
typed motor behaviors to higher order behaviors such as insistence on sameness (Américain
Psychiatric Association, 2013). It has been proposed that these complex behavioral features
are associated with atypical patterns of functional connectivity (FC). Such theories include
reduced communication between frontal and posterior brain regions (Just et al., 2012), dans-
creased local FC along with reduced long-range FC (Belmonte et al., 2004; Courchesne &
Pierce, 2005), and an abnormal developmental trajectory of FC compared with typically de-
veloping (TD) individuals (Nomi & Uddin, 2015; Uddin et al., 2013b). Cependant, complexe
patterns of both increased and decreased FC have been found in neuroimaging studies of
ASD, and results are inconsistent across studies (see Hull et al., 2016, Picci et al., 2016, et
Uddin et al., 2013b, for reviews).
It is crucial to consider the heterogeneous nature of ASD, both in terms of behavioral severity
and FC profiles. The importance of this consideration is highlighted by the inconsistent results
regarding relationships between FC and behavioral profiles in individuals with ASD in previous
études (par exemple., Keown et al., 2013; Lee et al., 2016; Monk et al., 2009; Uddin et al., 2013b).
Several recent studies that considered the heterogeneity of neurobiological and behavioral fea-
tures of ASD have reported novel findings regarding brain-behavior relationships. Hahamy,
Behrmann, and Malach (2015) found that idiosyncratic distortions in FC from a “typical” tem-
plate were related to ASD symptom severity. Nunes et al. (2018) reported that incorporation of
vertices along the cortical surface into intrinsic connectivity networks, particularly into default
mode and sensorimotor networks, was more idiosyncratic in ASD and related to symptom
severity.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
un
_
0
0
0
6
7
p
d
t
.
Resting-state network:
A set of brain regions that exhibit
synchronous patterns of activity and
are involved in similar functions.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
FC-based subtypes have the potential to resolve some of the current discrepancies regard-
ing the nature of FC abnormalities in individuals with ASD, and to shed light on the com-
plex relationships between FC and behavior, which may differ between subtypes. Previously,
ASD subtypes have been defined based on clusters of social communication behaviors and
RRBs (Georgiades et al., 2013), structural MRI (Hrdlicka et al., 2005), various neuroanatomical
features (Hong et al., 2017), and FC (Chen et al., 2015). Chen et al. (2015) found two subtypes
that exhibited unique FC patterns in different resting-state networks (RSNs), and differed in
terms of ASD symptom severity. Hong et al. (2017) found that prediction of individual scores
on the Autism Diagnostic Observation Schedule (ADOS) greatly improved when subtypes
were considered, compared with considering all ASD participants as one group. Ainsi, brain-
based subtyping has the potential to elucidate brain-behavior relationships that are unique
to each subtype, as certain behaviors may result from complex interplay between local and
distributed processing in the brain. One limitation of these studies is that they did not include
both ASD and TD participants in the subtyping procedures. Because of the heterogeneity of
ASD symptomatology and inconsistent reports of FC profiles in ASD, it is important to consider
FC patterns that may be shared among those with ASD and controls. Recent work revealed
shared FC patterns between ASD and TD participants, and between TD individuals and other
clinical groups, including ADHD and schizophrenia (Spronk et al., 2018). En outre, taking
a dimensional approach to examining FC can reveal information about brain-behavior rela-
tionships that exist as a continuum across typical development and clinical diagnoses. Rashid
Neurosciences en réseau
345
Functional connectivity subtypes of ASD and controls
K-means clustering:
A machine learning method used to
identify distinct patterns (clusters) dans
a set of data.
et autres. (2018) demonstrated a continuum of the relationship between neurobiological features
and subclinical ASD symptoms in healthy controls. En plus, Muller and Amaral (2017)
highlighted the importance of “studying functional systems dimensionally within the [recherche
domain criteria] framework” and defining biological subtypes of ASD to make progress toward
customized treatments and behavioral interventions.
In the present study, we used a data-driven, dimensional approach to characterize subtypes
based on distinct clusters of FC in all participants, and to relate FC patterns to specific be-
havioral profiles in these subtypes. We used k-means clustering, an unsupervised machine
learning technique, to define subtypes by using functional connections as features. Suivant, nous
implemented multivariate statistical analyses that, when applied to neuroimaging data, reveal
optimal relationships between measures of brain activity and experimental design or group
membership. Using this approach categorically, we characterized connections that were reli-
ably different between subtypes, and between ASD and TD participants. We also character-
ized dimensional relationships between particular FC patterns and a set of behaviors across
participants in both diagnostic groups within each subtype. It was hypothesized that defining
FC-based subtypes in a sample of both ASD and TD participants by using data-driven metrics
would reveal unique information about brain-behavior interactions.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
un
_
0
0
0
6
7
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Bootstrapping:
A resampling method in which
statistics are recalculated on a
random sample of data while
maintaining group membership.
RÉSULTATS
FC-Based Subtypes of ASD and TD Participants
FC-based subtypes were defined using k-means clustering. It was necessary to regress the ef-
fects of age and acquisition site out of the FC matrices prior to performing k-means clustering
(see Materials and Methods). When these effects were not removed, there was a significant
difference in the distribution of scan sites between the two subtypes defined by k-means clus-
tering, X2 (4, N=266) = 78.60, p < 0.001. After the scan sites were regressed from the data,
the resulting subtypes were significantly different in age, t(264) = 2.50, p = 0.01; thus, effects
of both site and age were regressed from the data.
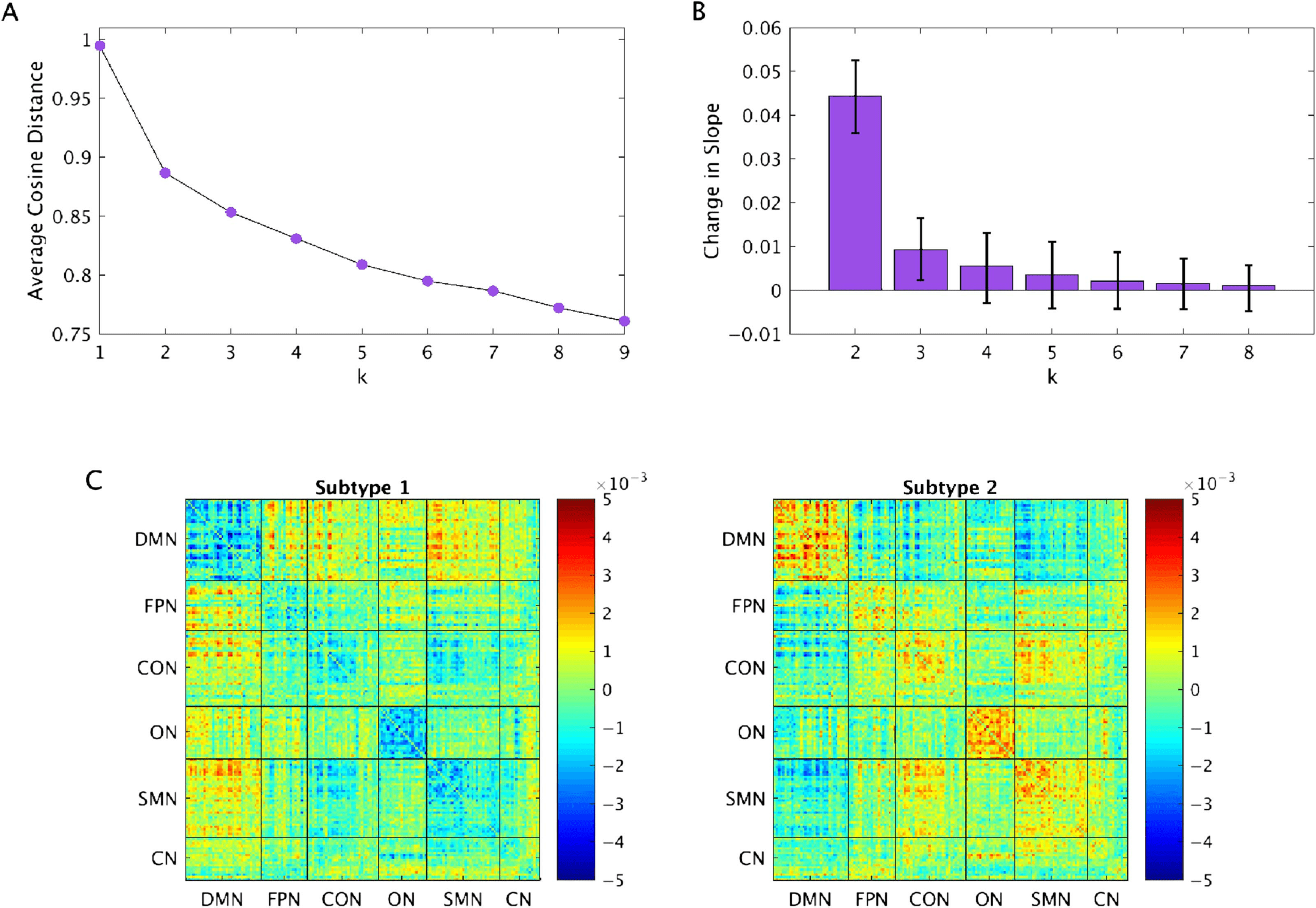
The optimal number of clusters, as determined by the elbow point criterion, was 2 (Fig-
ure 1A). Using a bootstrapping procedure to evaluate the reliability of the optimal number of
clusters, it was found that the optimal number of clusters was 2 in 500/500 bootstrap samples
(Figure 1B). Qualitatively, it can be seen that the change in slope for k = 2 was always much
greater than the change in slope for any other value of k. For example, the mean change in
slope for k = 2 was 4.81 times greater than that for k = 3, and confidence intervals do not
overlap. For values of k = 3 or more, the intervals do overlap.
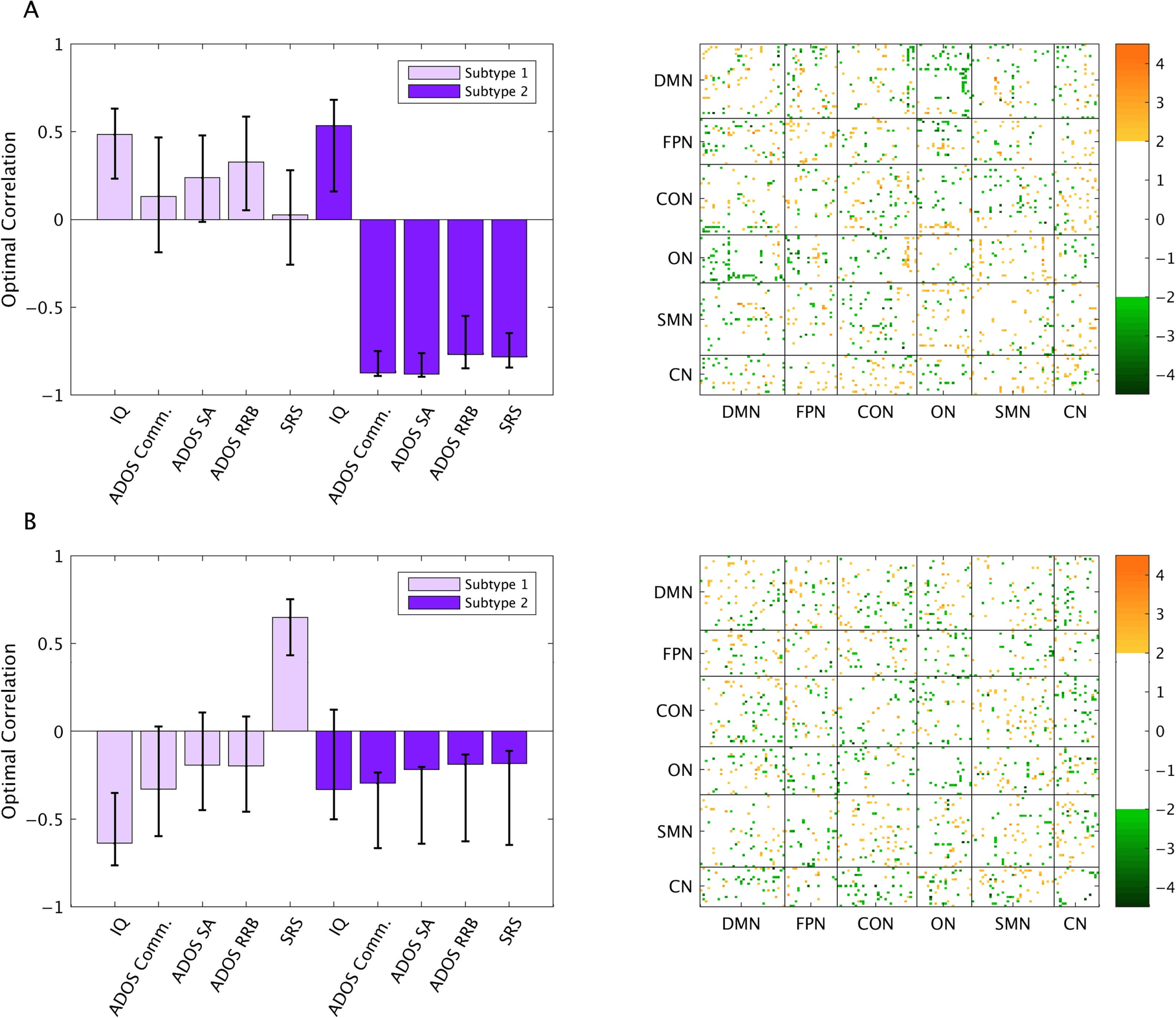
Subtype 1 consisted of 85 ASD participants and 54 TD participants. Subtype 2 consisted of
60 ASD participants and 67 TD participants. Qualitatively, Subtype 1 was defined by stronger
FC between networks, particularly between the default mode network (DMN) and other net-
works, and weaker FC within networks relative to Subtype 2 (Figure 1C).
Importantly, subtypes did not differ in demographics or behavior, including IQ, eye status,
medication use, presence of comorbidities, head motion, or the parameters (scan site and age)
that were regressed out of the FC matrices (Supporting Information Table S3, Easson, Fatima, &
McIntosh, 2019). Although subtypes differed in ADOS communication scores (t(112) = 2.62,
p = 0.01), they were marginally significantly different in ADOS total scores (t(130) = 1.87,
p = 0.06) and differences in the social responsiveness scale (SRS; Constantino & Gruber,
2005) scores approached significance (t(136) − 1.71, p = 0.09), these differences were driven
by the fact that there were more TD participants with these scores in Subtype 2 compared
Network Neuroscience
346
Functional connectivity subtypes of ASD and controls
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 1.
(A) Elbow point plots, indicating that the optimal number of clusters is 2. (B) Bootstrapping procedure to evaluate the reliability
of the optimal number of clusters. Error bars show 95% confidence intervals. (C) Subtype centroids. DMN = default mode network; FPN =
fronto-parietal network; CON = cingulo-opercular network; ON = occipital network; SMN = sensorimotor network; CN = cerebellar network.
with Subtype 1. SRS scores did not differ between ASD participants in Subtypes 1 and 2, and
also did not differ between TD participants in Subtypes 1 and 2. ADOS scores did not differ
between ASD participants in Subtypes 1 and 2, but could not be compared for TD participants
in Subtypes 1 and 2 because ADOS scores were only available for 2 TD participants in Sub-
type 1 and 12 TD participants in Subtype 2.
Multivariate Analysis of Subtype and Diagnostic Group Differences in FC
We used a multivariate statistical approach to determine differences in FC between subtypes
and between ASD and TD participants. The reliability of these patterns was determined via
bootstrap sampling. A functional connection was considered to be reliable, or stable, if the
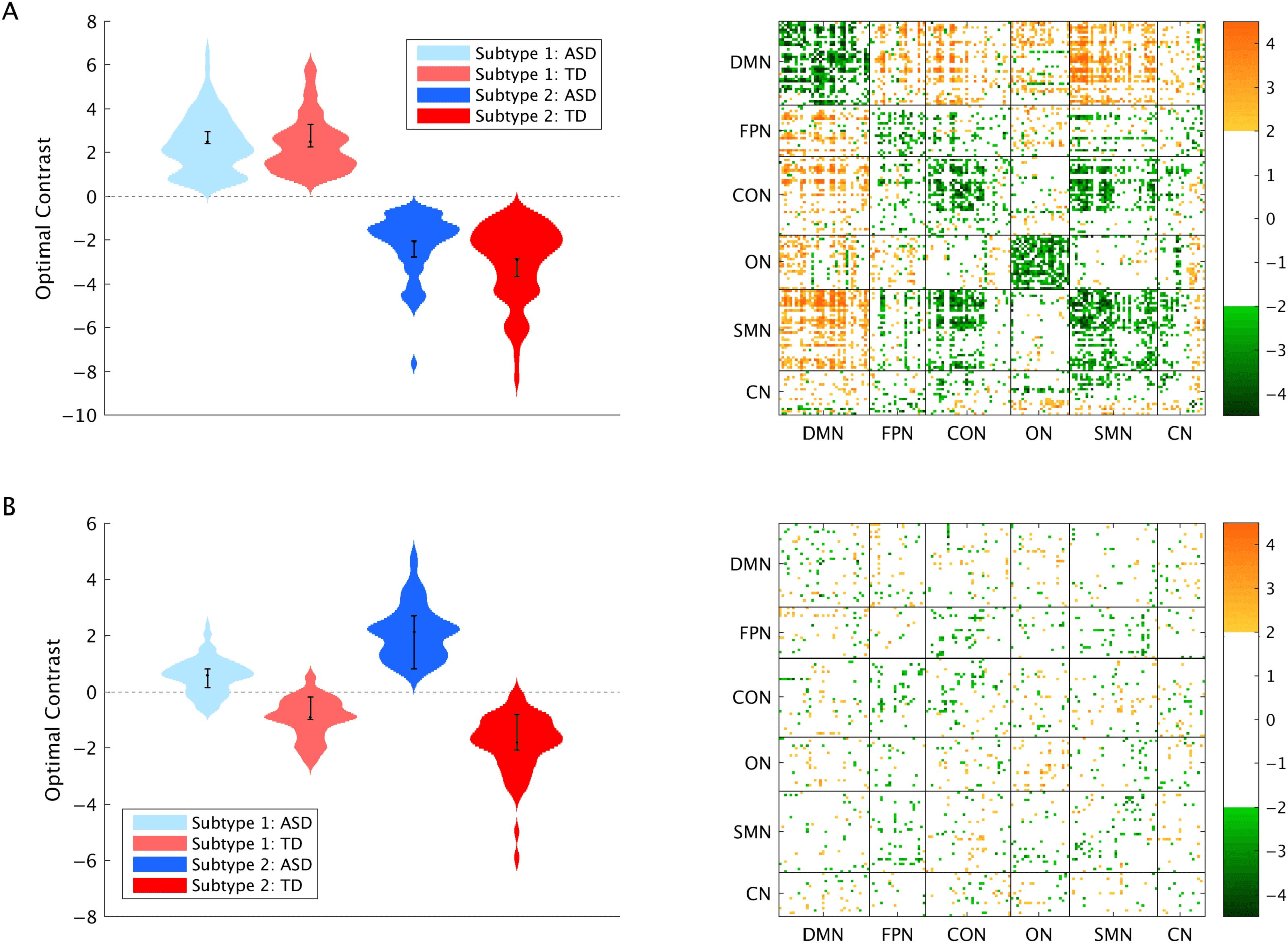
absolute value of its bootstrap ratio (BSR) exceeded 2. This analysis revealed two signifcant
patterns. The first pattern showed stable differences in FC between subtypes ( p < 0.001,
61.07% of variance explained, Figure 2A), whereby Subtype 2 was characterized by stron-
ger FC within RSNs, and weaker FC between RSNs, compared with Subtype 1. The contrast
expression for this FC pattern (Supporting Information Figure S3, Easson et al., 2019) revealed
that functional connections with significant positive BSRs, on average, were positive in
Network Neuroscience
347
Functional connectivity subtypes of ASD and controls
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 2. Results from the multivariate group analysis. (A) First pattern and (B) second pattern, and the associated BSRs for each connection at
a threshold of ±2. Error bars show 95% confidence intervals determined through bootstrap resampling.
Subtype 1 and negative in Subtype 2, and vice versa for negative BSRs. The second pattern
revealed a contrast between diagnostic groups in both subtypes (p = 0.02, 21.74% of variance
explained, Figure 2B), with a diffuse spatial pattern. The contrast expression (Supporting In-
formation Figure S4, Easson et al., 2019) revealed that functional connections with significant
positive BSRs, on average, were negative in the ASD group and positive in the TD group,
and vice versa for negative BSRs. The third pattern, which revealed a subtype by diagnosis
interaction, was not significant, p = 0.92.
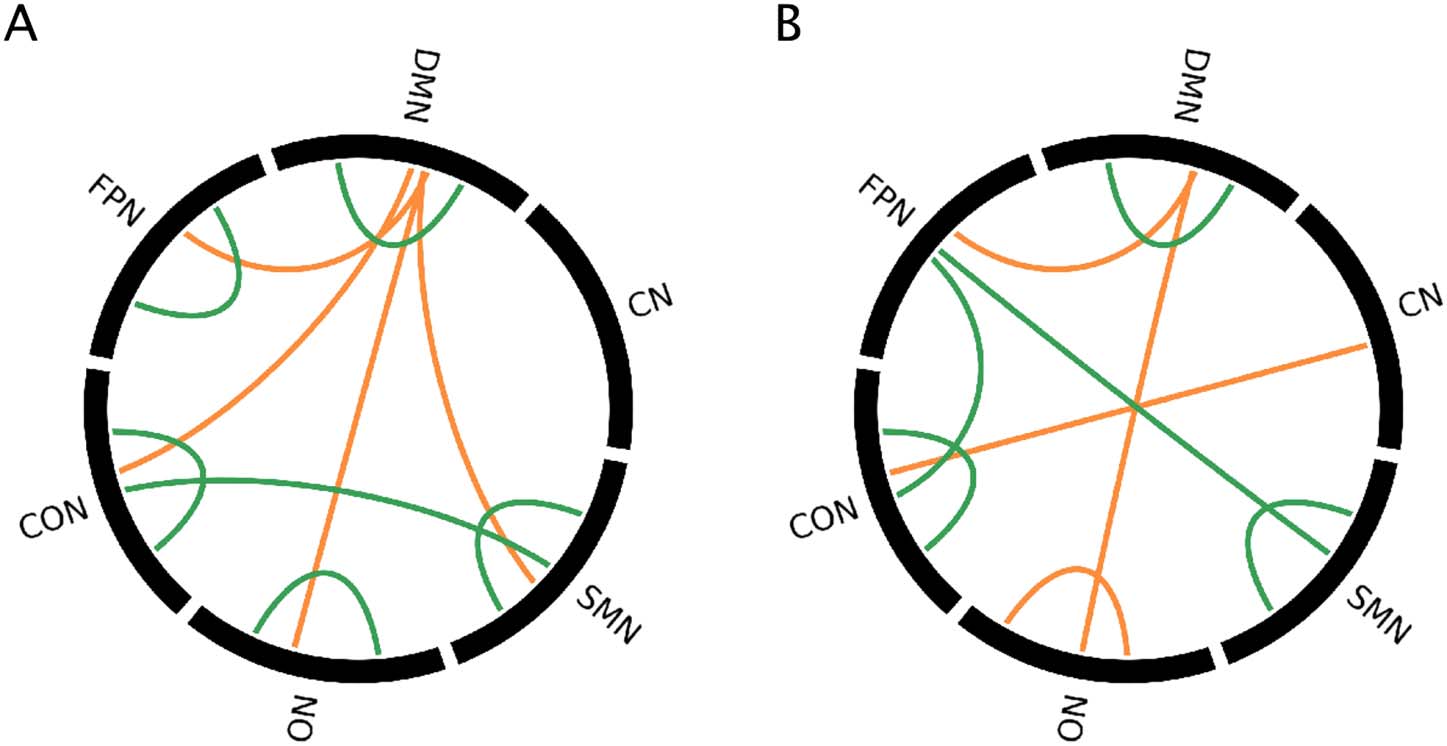
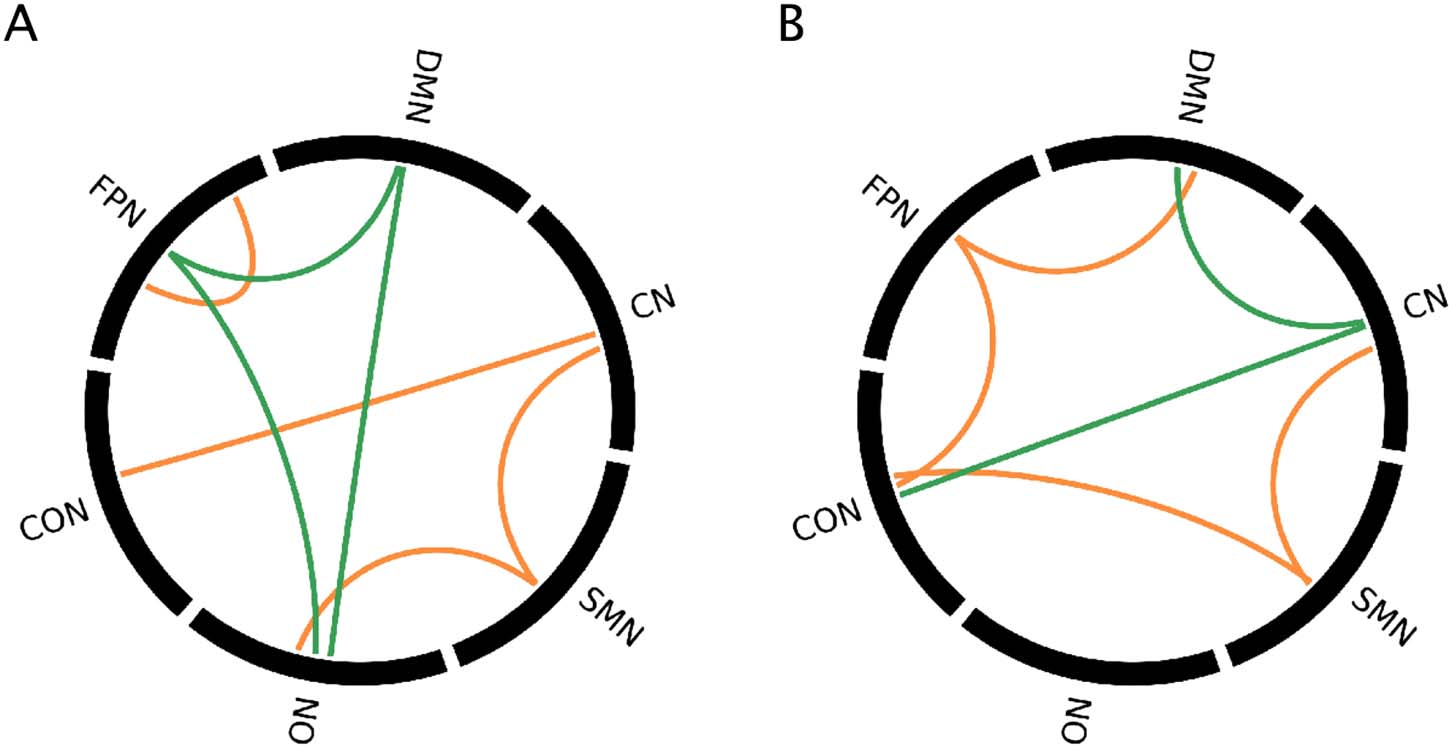
In addition to examining individual connections that differed between subtypes (first spatial
pattern) and diagnostic groups (second spatial pattern), the significance of the average spatial
patterns within and between RSNs was evaluated using permutation tests (see Materials and
Methods) and is shown in Figure 3.
Multivariate Analyses of FC-Behavior Relationships
A multivariate brain-behavior analysis was used to assess relationships between FC and a set of
behavioral measures in the two subtypes, including IQ, ADOS scores (communication; COMM),
Network Neuroscience
348
Functional connectivity subtypes of ASD and controls
Significant contributions of RSN pairs to each pattern for positive and negative BSRs, for
Figure 3.
the (A) first pattern and (B) second pattern from the multivariate group analysis. Orange = positive
BSRs, green = negative BSRs.
social affect, and RRB, and scores on the SRS. The full set of behavioral measures was available
for 51 participants (49 ASD, 2 TD) in Subtype 1 and 50 participants (38 ASD, 12 TD) in Subtype
2. ADI-R scores were not included, as only 28 participants in Subtype 1 and 26 participants in
Subtype 2 had the full set of behavioral measures including ADI-R scores. Furthermore, none
of the participants with the full set of scores including ADI-R scores were TD participants.
The analysis revealed three significant patterns. The first pattern (p = 0.03, 32.09% co-
variance explained) revealed stable relationships between FC and IQ and ADOS RRB scores
in Subtype 1, and stable relationships between FC and all behavioral measures in Subtype 2.
This first pattern was a contrast between Subtypes 1 and 2 in terms of relationships with FC and
ADOS RRB scores, such that connections that were reliably positively correlated with ADOS
RRB scores in Subtype 1 were negatively correlated in Subtype 2, and vice versa. The next
significant pattern was the third pattern (p = 0.008, 10.82% covariance explained), which
revealed a different spatial pattern that exhibited stable correlations with IQ and SRS in Sub-
type 1, and with all ADOS scores and SRS in Subtype 2. Additionally, there was a contrast
between Subtypes 1 and 2 in terms of correlations between FC and SRS scores. The seventh
pattern (p = 0.003, 4.45% covariance explained) revealed a contrast between Subtypes 1 and 2
in terms of correlations between FC and ADOS communication scores, as well as stable cor-
relations between FC and ADOS social affect scores in Subtype 1.
Overall, it can be seen that connections that show stable correlations with behavior are
diffuse. Patterns that accounted for more than 10% of the covariance between FC and behavior
(i.e., patterns 1 and 3) are shown in Figure 4, and the corresponding contrast expressions are
shown in Supporting Information Figure S5 and S6 (Easson et al., 2019). The stability of these
FC-behavior relationships within and between RSNs are shown in Figure 5.
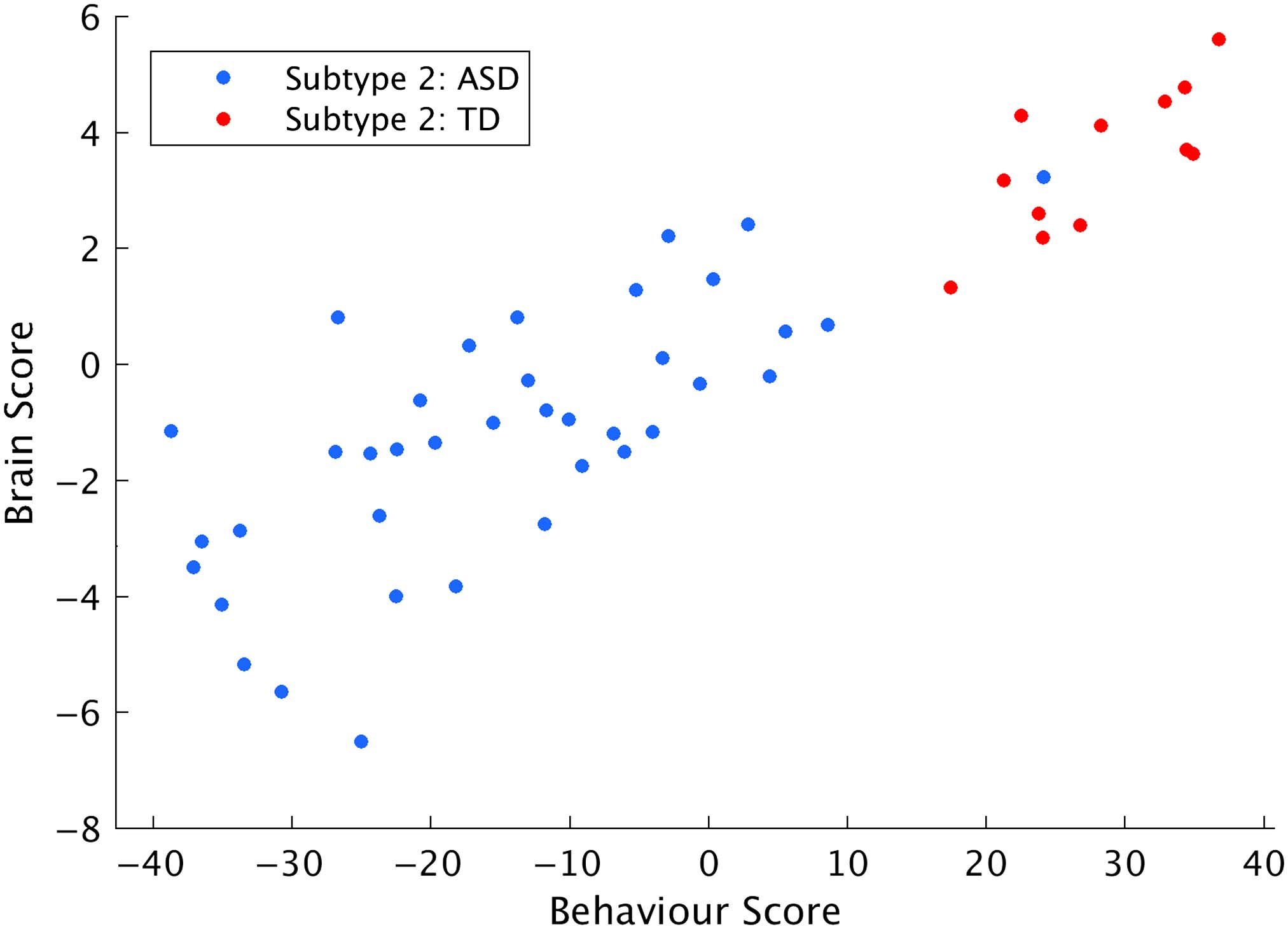
Continuum of FC-Behavior Relationships Across Diagnostic Groups
The relationship between brain and behavior scores for ASD and TD participants in Subtype 2
was evaluated for the first pattern of the multivariate brain-behavior analysis, which explained
the greatest proportion of covariance between FC and the set of behavioral measures. The
Network Neuroscience
349
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 4. Results from the multivariate brain-behavior analysis. (A) First pattern and (B) third pattern, and the associated BSRs for each connec-
tion at a threshold of ±2. Error bars show 95% confidence intervals determined through bootstrap resampling.
continuum of scores for both brain and behavior variables illustrates that there is a pattern of FC
that covaries with the severity of behaviors across the autism spectrum and typical development
(Figure 6). This analysis was only performed in Subtype 2, as there were only 2 TD participants
in Subtype 1 who had the full set of behavior measures.
Relationships Between Multivariate Group and Brain-Behavior Spatial Patterns
We then determined the relationship between the patterns from the multivariate group anal-
ysis and the multivariate brain-behavior analysis by correlating the brain saliences for each
Network Neuroscience
350
Functional connectivity subtypes of ASD and controls
Figure 5.
9A) first pattern and (B) third pattern. Orange = positive BSRs, green = negative BSRs.
Significant contributions of RSN pairs to each pattern for positive and negative BSRs for
Permutation testing:
A resampling method in which
statistics are recalculated on data
with shuffled group labels to obtain a
null distribution to which the original
test statistic is compared.
analysis, and evaluated the significance of these correlations using permutation testing. There
was a significant correlation between the first brain-behavior pattern and the second group
pattern (r = 0.40, p < 0.001), indicating that the continuum of FC-behavior relationships was
associated with the diagnostic pattern from the group analysis. The correlations between the
other patterns were not significant (brain-behavior pattern 1 and group pattern 1: r = −0.06,
p = 0.81; brain-behavior pattern 3 and group pattern 1: r = 0.005, p = 0.45; brain-behavior
pattern 3 and group pattern 2: r = 0.07, p = 0.13.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 6. Brain and behavior scores for Subtype 2, from the first pattern of the multivariate brain-
behavior analysis.
Network Neuroscience
351
Functional connectivity subtypes of ASD and controls
DISCUSSION
Overview
This study reveals distinct clusters of FC patterns across a cohort of both ASD and TD partici-
pants. We characterized differences in FC between subtypes and diagnostic groups, and showed
that individuals within each subtype exhibit different relationships between FC and behavior.
The continuum of brain and behavior scores across ASD and TD participants reveals that FC
phenotypes observed in ASD extend to typical development in relation to behavioral severity.
Comparison of FC Between Subtypes and Diagnostic Groups
Two subtypes were defined for all participants. When all four groups were considered in a
multivariate analysis (i.e., ASD Subtype 1, ASD Subtype 2, TD Subtype 1, and TD Subtype 2),
the strongest pattern, not surprisingly, was a contrast between subtypes. Regardless of diag-
nostic group, Subtype 2 was defined by greater FC within networks and lower FC between
networks, especially between the DMN and other RSNs, compared with Subtype 1. Connec-
tions within networks tended to be positive on average in Subtype 2 and negative in Subtype 1,
indicating reduced interactions among brain regions within these networks in Subtype 1.
Connections between networks that were lower in Subtype 2 tended to be negative, but were
positive on average in Subtype 1 (Supporting Information Figure S3, Easson et al., 2019). As
anticorrelations between RSNs are hypothesized to signify the ability for regions that are
relevant for certain cognitive functions to become activated with concurrent deactivation of
irrelevant regions (Fox et al., 2005; Greicius et al., 2003), these abilities may be affected in
Subtype 1. Using the Power atlas (Power et al., 2011), previous FC-based ASD subtypes also
revealed unique FC profiles between subtypes within and between RSNs (Chen et al., 2015).
A second pattern revealed diffuse functional connections that differed between diagnostic
groups in both subtypes. ASD participants showed decreased FC within the sensorimotor net-
work (SMN), DMN, and cingulo-opercular network (CON), but greater FC within the occipital
network (ON). Atypical FC of sensorimotor regions has been reported in ASD (Anderson et al.,
2011a; Mostofsky et al., 2009; Turner et al., 2006). Abnormal DMN functioning in ASD has
been related to difficulties with self-referential processing, redirecting attention from external to
internal processing, and theory of mind (e.g., Assaf et al., 2010). Various studies have reported
decreased FC between DMN regions in ASD (Assaf et al., 2010; Kennedy & Courchesne, 2008;
Monk et al., 2009; Weng et al., 2010), although hyperconnectivity has also been reported
(Monk et al., 2009; Uddin et al., 2013a). Decreased FC within the CON is in line with previous
studies that showed difficulties with set maintenance in ASD (Kaland, Smith, & Mortensen,
2008; Miller et al., 2015). Increased FC in the ON is consistent with findings of increased local
connectivity in primary visual regions (Keown et al., 2013) and increased involvement of extra-
striate cortex (Shen et al., 2012) in ASD. Additionally, increased FC was found between the
DMN and FPN, DMN and ON, and CON and CN in ASD participants. These connections were
positive on average in ASD, but negative on average in controls (Supporting Information
Figure S4, Easson et al., 2019). Previous studies have reported reduced negative connectiv-
ity in ASD, which was described as reduced functional segregation of networks (Rudie et al.,
2012, 2013a). However, other between-network connections (FPN-CON and FPN-SMN) ex-
hibited a greater degree of anti-correlation in ASD. The functional significance of decreased
anticorrelations between some RSNs, but increased anticorrelations between others, remains
to be explored.
The third pattern, showing a subtype by diagnosis interaction, was not significant, thus
revealing additive effects of subtype and diagnosis on FC patterns. Therefore, the expression
Network Neuroscience
352
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
of the subtypes does not depend on the diagnosis; the manifestation of the subtypes in ASD is
not different from controls.
Comparison of FC-Behavior Relationships Between Subtypes
Reliable correlations between FC and behavior were observed within and between RSNs
for IQ and ADOS RRB scores for Subtype 1, and all behavioral measures for Subtype 2, show-
ing that similar behavioral profiles can be associated with different functional correlates in
the brain. Previous studies have reported mixed results regarding FC-behavior relationships
in ASD (e.g., Keown et al., 2013; Lee et al., 2016; Monk et al., 2009; Uddin et al., 2013b).
For instance, Lee et al. (2016) reported a negative correlation between RRBs and connec-
tions involving certain DMN regions, whereas Monk et al. (2009) reported a positive correla-
tion. Our results highlight the importance of considering FC-based subtypes when examining
brain-behavior relationships in ASD and controls.
Importantly, individuals in each subtype
did not differ significantly in IQ or SRS scores, and ASD participants in the two subtypes did
not differ significantly in ADOS scores. Thus, there is unique information about FC-based
subtypes that is not accessible by using behavior alone. This finding of unique brain-behavior
relationships in different subtypes is supported by previous work that showed that predic-
tion of ADOS scores improved when subtypes of ASD, based on unique neuroanatomical
profiles, were considered, as opposed to considering all ASD participants as a single group
(Hong et al., 2017). This study, as well as the FC-based subtypes study by Chen et al. (2015),
also utilized data from the Autism Brain Imaging Data Exchange (ABIDE); thus, it is possible
that there is overlap in the participants in our study and these previous studies.
The multivariate brain-behavior analysis supports the idea that instead of being a categorical
diagnosis, ASD should indeed be considered as an extreme of a continuum of both neuro-
biological and behavioral features that can also be observed in TD individuals (Constantino &
Todd, 2003; Rashid et al., 2018). In other words, there is normal variation in FC across both
ASD and TD participants (Figure 6), but too much of this natural variation is associated with a
diagnosis of ASD. This idea is supported by the continuum of brain and behavior scores from
pattern 1 of the brain-behavior analysis for Subtype 2, and the significant correlation between
the spatial pattern for this pattern and the second pattern from the group analysis, that is, the
contrast in FC between diagnostic groups.
This dimensional approach has also been reinforced by ASD studies that accounted for
heterogeneity of the relationships between behavior and various neurobiological features
(Hahamy et al., 2015; Nunes et al., 2018). Recently, it has been noted that different features of
brain function are variable even among TD individuals, and a certain feature cannot be consid-
ered to be an impairment unless it is accompanied by behavioral symptoms (Muller & Amaral,
2017). Our results support this idea by showing that some FC patterns are (a) similar among
subsets of ASD and TD participants and (b) correlated with behavioral severity. The similarity
of FC patterns in ASD and controls has also been demonstrated by Spronk et al. (2018), who
showed that FC patterns between TD participants and several clinical groups, including ASD,
ADHD, and schizophrenia, are highly correlated.
Limitations
One limitation of our study is that we defined subtypes using a single data preprocessing
strategy. However, inconsistent results between FC studies in ASD relate, in part, to different
preprocessing choices (Hull et al., 2016). For instance, Ciric et al. (2017) showed that global
signal regression reduces the relationship between FC and head motion, but can result in
Network Neuroscience
353
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
distance-dependent artifacts in FC unless used in combination with censoring methods. Global
signal regression and low-pass filtering have been shown to affect group differences in FC
between participants with and without ASD (Gotts et al., 2013; Muller et al., 2011). The length
of fMRI scans may also contribute to heterogeneity across studies: increasing scan lengths
improves the reliability of FC estimates (Birn et al., 2013). It is therefore crucial to gain a better
understanding of how preprocessing choices and scanning parameters affect group differences
in FC, and to compare FC-based subtypes across different preprocessing strategies.
Unfortunately, with the current dataset, we did not have a way to investigate age, as when
age was not controlled for in the FC data, the resulting subtypes from k-means clustering dif-
fered significantly in age. Therefore, we cannot rule out the possibility that FC-based sub-
types may manifest differently in various age groups. Uddin et al. (2013b) reported complex
developmental trajectories of FC in ASD, with primarily hyperconnectivity in childhood and
hypoconnectivity in adulthood. Furthermore, Anderson et al. (2011b) found that classification
accuracy of ASD based on FC was higher in younger compared with older cohorts, suggesting
that FC patterns between ASD and controls may be more distinct in childhood. Thus, although
unique FC subtypes exist across a broad age range, there may be differences in FC clusters in
children, adolescents, and adults. In this study, age ranges differed between scan sites, making
it difficult to differentiate between age and site effects on FC. Larger samples within smaller
age ranges from a single site are therefore required to study the manifestation of subtypes in
different age groups.
Furthermore, we examined the continuum of brain and behavior scores across both ASD
and TD participants in Subtype 2; however, ADOS scores were available for only 2 TD partic-
ipants in Subtype 1. To better characterize the continuum of FC-behavior relationships across
participants in different subtypes, future studies should collect measures of subclinical ASD be-
haviors using scales such as the SRS and Autism-Spectrum Quotient (AQ; Baron-Cohen et al.,
2001), which is a brief self-report of ASD traits.
Finally, ABIDE consists of data from high-functioning individuals with ASD, thus limiting
the generalizability of our findings. Including individuals with low-functioning ASD will be
important in future studies to determine if similar subtypes exist among these individuals.
Conclusions
Multivariate analyses of FC-based subtypes highlight the importance of considering the hetero-
geneity of FC patterns and behavior, and reveal the continuum of brain-behavior relationships
in individuals with and without ASD. As subtypes exhibited different relationships between FC
and behavior, it will be important to determine if individuals with ASD in different subtypes
exhibit unique responses to treatments and behavioral therapies.
MATERIALS AND METHODS
Participants
Resting-state fMRI data from 145 males with ASD and 121 TD males were acquired from
the Preprocessed Connectomes Project (Craddock et al., 2013; http://www.preprocessed-
connectomes-project.org/abide). The data had been obtained from ABIDE (Di Martino et al.,
2014; http://www.fcon_1000.projects.nitrc.org/indi/abide) and preprocessed using the Con-
nectome Computation System (CCS) pipeline (Xu et al., 2015). Participants were excluded if
their age was greater than 40, full scale IQ was less than 75, mean framewise displacement
(FD) during the resting-state fMRI scan was greater than 0.20 mm, percentage of data points
Network Neuroscience
354
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
exceeding 0.20 mm was greater than 20%, and/or scans were rated as good by less than two
(out of 3) raters as per the ABIDE quality assessment protocol (http://preprocessed-connectomes-
project.org/abide/quality_assessment.html). Groups were matched for age, IQ, mean FD, and
the percentage of data points exceeding 0.20 mm. ASD diagnoses were confirmed using ADOS
(Lord et al., 2000) and/or the Autism Diagnostic Interview-Revised (ADI-R; Lord et al., 1994).
Participant characteristics are shown in Table 1, along with the number of scores that were
available for ADOS, ADI-R, and SRS if these scores were not available for all participants. Par-
ticipant characteristics for each site are described in Supporting Information Table S1 (Easson
et al., 2019).
fMRI Preprocessing
Data from five sites (New York University Lagone Medical Center, University of Utah School
of Medicine, San Diego State University, Trinity Centre for Health Sciences, and University
of Michigan) using a TR of 2,000 ms were included. The proportion of ASD compared with
TD subjects was not significantly different across sites, X2(4, N = 266) = 5.07, p = 0.28.
Written, informed consent or assent was obtained for all participants in accordance with re-
spective institutional review boards. Additional information about scanner types and param-
eters can be found on the ABIDE website (http://www.fcon_1000.projects.nitrc.org/indi/abide).
The CCS preprocessing steps, which had been carried out as part of the Preprocessed Connec-
tomes Project, were as follows: dropping the first four volumes, removing and interpolating
temporal spikes, slice timing correction, motion correction, brain mask creation, 4D global
mean-based intensity normalization, boundary-based registration of functional to anatomi-
cal images, anatomical segmentation of gray matter, white matter and cerebrospinal fluid,
nuisance parameter regression (including 24 motion parameters, white matter and CSF sig-
nals, linear and quadratic trends, and the global signal), band-pass filtering (0.01 to 0.1 Hz),
and registering functional images to the MNI template. The final preprocessed time series for
each subject were obtained from the Preprocessed Connectomes Project. We chose to use
data that had the global signal regressed out, as this step has been shown to help mitigate
differences across multiple sites (Power et al., 2014). Furthermore, it has been shown re-
cently that global signal regression attenuates artifactual changes in BOLD signal that are
introduced by head motion (Byrge & Kennedy, 2017; Ciric et al., 2017; Power et al., 2017). It
should also be noted that without global signal regression, FC-based subtypes differed in head
motion (both mean FD, t(264) = −4.68, p < 0.001, and percentage of frames above 0.2 mm,
t(264) = −5.02, p < 0.001). We also implemented ICA denoizing by using ICA-AROMA
(Pruim et al., 2015a, 2015b) and found that the resulting subtypes still differed in mean FD,
t(264) = −3.49, p < 0.001, and percentage of frames exceeding 0.2 mm, t(264) = −3.96,
p < 0.001. Furthermore, regional artifacts were evident in the cluster centroids when the data
were preprocessed without global signal regression (Supporting Information Figure S2A, Easson
et al., 2019), and without global signal regression but with ICA denoizing using ICA-AROMA
(Supporting Information Figure S2B, Easson et al., 2019).
The time series of 160 4.5-mm spherical regions of interest (ROIs) from the Dosenbach atlas
(Dosenbach et al., 2010) were obtained (see Supporting Information Table S2 and Supporting
Information Figure S1, Easson et al., 2019). Regions in this atlas were selected from meta-
analyses of task-related fMRI studies and categorized into six different RSNs: the DMN, fronto-
parietal network (FPN), CON, ON, SMN, and cerebellar network (CN). Additional details of
the fMRI preprocessing steps can be found on the Preprocessed Connectomes Project website
(http://www.preprocessed-connectomes-project.org/abide).
Network Neuroscience
355
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
Variable
N
Age
IQ
Mean FD
Percent FD > 0.2 mm
Handedness
Eye status
Scan site
Medication use
Comorbidities
ADOS Total
ADOS Communication
ADOS Social
ADOS RRB
ADI-R Social
ADI-R Verbal
ADI-R RRB
SRS
Tableau 1. Participant characteristics.
ASD
Mean ± SD
[range]
145
16.47 ± 6.46
[7.13–39.10]
TD
Mean ± SD
[range]
121
16.03 ± 5.70
[6.47–31.78]
107.57 ± 16.32
[76–148]
110.08 ± 11.61
[80–133]
0.07 ± 0.04
[0.02–0.19]
4.69 ± 5.27
[0–19.33]
120 RH
21 LH
121 open
24 closed
NYU: 59
SDSU: 11
TRINITY: 18
UM: 26
USM: 31
27 yes
86 Non
32 unknown
0.07 ± 0.03
[0.03–0.19]
3.92 ± 1.29
[0–19.33]
109 RH
10 LH
95 open
26 closed
NYU: 52
SDSU: 10
TRINITY: 16
UM: 29
USM: 14
0 yes
106 Non
15 unknown
28 yes
117 no/unknown
0 yes
121 no/unknown
11.69 ± 3.68
[5–22]
(N = 118)
3.89 ± 1.55
[0–8]
(N = 100)
7.89 ± 2.81
[2–14]
(N = 100)
2.04 ± 1.46
[0 – 7]
(N = 98)
19.07 ± 5.44
[7–30]
(N = 108)
15.38 ± 4.36
[2–25]
(N = 109)
5.66 ± 2.60
[0–12]
(N = 109)
1.14 ± 1.17
[0–4]
(N = 14)
0.50 ± 0.65
[0–2]
(N = 14)
0.64 ± 0.84
[0–3]
(N = 14)
0.07 ± 0.27
[0–1]
(N = 14)
N/A
N/A
N/A
Significance
t(264) = 0.58, p = 0.56
t(264) = −1.43, p = 0.15
t(264) = 1.32, p = 0.19
t(264) = 1.29, p = 0.20
X2(1, N = 266) = 0.52, p = 0.13
X2(1, N = 266) = 1.38, p = 0.35
X2(4, N = 266) = 5.07, p = 0.28
N/A
N/A
t(130) = 10.64, p < 0.001
t(112) = 8.06, p < 0.001
t(112) = 9.56, p < 0.001
t(110) = 5.00, p < 0.001
N/A
N/A
N/A
92.56 ± 31.00
[26–164]
(N = 89)
20.59 ± 12.43
[1–56]
(N = 49)
t(136) = 15.56, p < 0.001
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
356
Functional connectivity subtypes of ASD and controls
Functional Connectivity
Each subject’s fMRI time series was truncated to 145 time points, which was the minimum
number of time points across subjects. FC was defined by Fisher z-transformed Pearson cor-
relations for each ROI pair across all time points for each participant. The effects of age and
acquisition site (represented as a Helmert basis) were regressed out of the FC matrices. As it
has been recently shown that despite implementing preprocessing steps that aim to correct for
head motion in resting-state fMRI, residual motion effects can contaminate FC estimates (Ciric
et al., 2017), a multivariate brain-behavior analysis was performed to determine if there were
relationships between FC and head motion metrics (mean FD and percentage of frames ex-
ceeding 0.2 mm). There was not a significant relationship between FC and motion (p = 0.57).
K-Means Clustering
K-means clustering was used to define subtypes of distinct FC patterns. The lower triangle of
each participant’s FC matrix was used, such that the matrix for k means was in the form sub-
jects × FC. The k-means algorithm begins with an initialization of k centroids. Then, in the
assignment step, each participant is assigned to the closest centroid by using the cosine dis-
tance, defined as one minus the cosine of the included angle between each subject’s FC values
and each cluster’s centroids, which are treated as vectors. Next, in the centroid update step,
new centroids are defined as the mean of the data points that are currently assigned to that
centroid. These two steps are repeated iteratively until convergence, when cluster assignments
no longer change.
The “elbow point” criterion was used to determine the optimal number of clusters. To deter-
mine the elbow point, the average cosine distance between a cluster’s centroids and the FC
values of participants assigned to that particular cluster is calculated for each cluster, then
averaged across clusters to obtain a single distance metric for each value of k. These distances
are then plotted as a function of k, and the “elbow” is defined as the value of k where the change
in the rate of decrease in distance is sharpest. Values from k = 2 to k = 8 were tested (but also
included k = 1 in the elbow point plot as a reference point). Furthermore, we evaluated the
reliability of the number of clusters by using bootstrap resampling. Fifty percent of the sample
was selected at random, and these were grouped into subtypes using the k-means algorithm
for values of k from 2 to 8. The elbow criterion was then used to select the ideal value of k for
the bootstrap sample. This process was repeated 500 times to determine the reliability of the
optimal number of clusters.
Partial Least Squares
Partial least squares (PLS) is a multivariate statistical technique that is used to optimally re-
late brain activity to experimental design or group membership in the form of latent variables
(McIntosh et al., 1996; McIntosh & Lobaugh, 2004; Krishnan et al., 2011). PLS software, which
is implemented in Matlab, is available for download from research.baycrest.org/pls-software.
In mean-centering PLS, patterns relating a matrix of brain variables (in the form subjects × brain
variables) and group membership are calculated. For this study, the brain variables were the FC
values in the lower triangle of each subject’s FC matrix (12,720 connections). Mean-centering
PLS was used to examine differences in FC between subtypes and between ASD and TD
participants.
By using singular value decomposition (SVD), orthogonal patterns that express the maximal
covariance between the brain variables and group membership are computed. The resulting
Partial least squares (PLS):
A statistical method for
characterizing relationships between
a set of brain variables and group
membership or a set of behaviors.
Latent variables:
“Hidden” variable within observable
variables that are not measured
directly, but rather are inferred via
statistical procedures.
Network Neuroscience
357
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
patterns are sorted in order of the proportion of covariance between the brain and design/
behavior variables that the pattern accounts for, with the first pattern accounting for the most
covariance. Each pattern consists of saliences (weights) and a singular value. The brain sa-
liences indicate which brain variables (in this case, functional connections) best charac-
terize the relationship between the brain variables and group differences. Design saliences
indicate the group differences profiles that best characterize this relationship. Singular values
indicate the proportion of covariance between the brain and design matrices that each pattern
accounts for. Brain scores, which represent each subject’s contribution to each pattern, are
calculated by multiplying the original matrix of brain variables by the brain saliences.
In behavior PLS, a matrix of behavior variables is also included in the analysis to deter-
mine design-dependent (in this case, group-dependent) relationships between the brain vari-
ables and behavior. For this study, behavioral PLS was used to examine associations between
FC and a set of behavioral variables including IQ, ADOS scores (communication, social affect,
and RRBs), and scores on the SRS in each subtype.
The statistical significance of each pattern was determined using permutation testing. For
this procedure, the rows (participants) of the matrix of brain variables are reshuffled, and new
singular values are obtained using SVD. In this study, this procedure was repeated 1,000 times
to create a distribution of singular values. The p value associated with the original singular
value is defined as the proportion of singular values from the sampling distribution that are
greater than the original singular value, thus representing the probability of obtaining a singu-
lar value larger than the original value under the null hypothesis that there is no association
between the brain variables and group membership.
In addition to determining the statistical significance of each pattern, the reliability of the
brain saliences can also be determined by utilizing a bootstrapping procedure. Bootstrap sam-
ples are generated by randomly sampling subjects with replacement, while ensuring that group
membership is maintained. In this study, 500 bootstrap samples were generated. Creating
bootstrap samples allows one to determine which brain variables are stable, regardless of
which participants are included in the analysis. The BSR, defined as the ratio of the brain sa-
lience to the standard error of the salience (as estimated by the bootstrap procedure), is a mea-
sure of this stability. Reliable connections were defined as those that surpassed a BSR threshold
of __±2.0, which corresponds roughly to a 95% confidence interval.
As FC values can take on positive or negative values, positive BSRs could correspond to
either stronger positive or weaker negative connectivity in one group compared with the other,
and negative BSRs could indicate weaker positive or stronger negative connectivity. Thus, ex-
pressions of FC PLS contrasts were generated for each group. Positive expressions were gen-
erated by averaging connections (Fisher z-transformed Pearson correlation coefficients) that
had BSRs greater than 2 across all participants in each group. A similar procedure was per-
formed for negative expressions, that is, for connections showing BSRs less than −2.
In addition to assessing the contribution of each individual connection to the group differ-
ences, we were interested in determining the extent to which network-level FC, both within and
between RSNs, contributed to the group differences. This was of particular interest because
of hypotheses that ASD may be characterized by atypical FC within and between networks
(e.g., Hull et al., 2016; Rudie & Dapretto, 2013b). To assess the relative contributions of each
RSN to the spatial patterns, the BSR-thresholded spatial maps (i.e., adjacency matrices in the
form connections × connections) were separated into positive BSRs and negative BSRs. These
maps were thresholded such that connections with a BSR less than 2 but greater than −2 were
Network Neuroscience
358
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
set to 0. Positive BSRs greater than 2 were set to 1, and negative BSRs less than −2 were set to
−1. All thresholded BSRs within each pair of networks were then averaged to obtain a 6 × 6
matrix showing the average contribution of each network pair to the spatial pattern, separately
for positive and negative BSRs. To assess the significance of these contributions, the order of
connections in the BSR-thresholded matrices was permuted while keeping the RSN labels the
same, and then the above procedure was repeated to calculate the RSN contributions. This pro-
cess was repeated 1,000 times to obtain a distribution of average contribution values for each
RSN pair. Then, the significance of the original contribution is defined as the proportion of
contribution values from the sampling distribution that are greater than or equal to the original
value.
Data Visualization
Connectivity circle plots were created using the plot_connectivity_circle function from the
open-source MNE software package implemented in Python (Gramfort et al., 2013; 2014). All
other figures were created using Matlab (MATLAB 8.6.0 [R2015b], MathWorks, Natick, MA).
Violin plots were created using the distributionPlot.m function (Jonas, 2017).
ACKNOWLEDGMENTS
The authors thank Bratislav Misic and Sam Doesburg for helpful discussions, and the contrib-
utors to the Autism Brain Imaging Exchange and Preprocessed Connectomes Project.
AUTHOR CONTRIBUTIONS
Amanda K. Easson: Conceptualization; Formal analysis; Methodology; Visualization; Writing
– original draft; Writing – review & editing. Zainab Fatima: Conceptualization; Methodology;
Writing – review & editing. Anthony R. McIntosh: Conceptualization; Methodology; Super-
vision; Writing – review & editing.
FUNDING INFORMATION
Anthony R. McIntosh, Natural Sciences and Engineering Research Council of Canada (http://
dx.doi.org/10.13039/501100000038), Award ID: RGPIN-2018-04457. Amanda K. Easson,
Ontario Graduate Scholarship. Amanda K. Easson, Mynne & Harold Soupcoff Fellowship.
Amanda K. Easson, Finkler Graduate Student Fellowship.
REFERENCES
American Psychiatric Association. (2013). Diagnostic and Statistical
Manual of Mental Disorders (5th ed.). Washington, DC: Ameri-
can Psychiatric Association.
Anderson, J. S., Druzgal, T. J., Froehlich, A., DuBray, M. B., Lange,
N., Alexander, A. L., . . . Lainhart, J. E. (2011a). Decreased inter-
hemispheric functional connectivity in autism. Cerebral Cortex,
21(5), 1134–1146. https://doi.org/10.1093/cercor/bhq190
Anderson, J. S., Nielsen, J. A., Froehlich, A. L., DuBray, M. B.,
Druzgal, T. J., Cariello, A. N., . . . Lainhart, J. E. (2011b). Func-
tional connectivity magnetic resonance imaging classification of
autism. Brain, 134(Pt 12), 3742–3754. https://doi.org/10.1093/
brain/awr263
Assaf, M., Jagannathan, K., Calhoun, V. D., Miller, L., Stevens,
M. C., Sahl, R., . . . Pearlson, G. D. (2010). Abnormal functional
connectivity of default mode sub-networks in autism spectrum
disorder patients. NeuroImage, 53(1), 247–256. https://doi.org/
10.1016/j.neuroimage.2010.05.067
Baron-Cohen, S., Wheelwright, S., Skinner, R., Martin, J., & Clubley,
E. (2001). The autism-spectrum quotient (AQ): Evidence from
Asperger syndrome/high-functioning autism, males and females,
Journal of Autism and Develop-
scientists and mathematicians.
mental Disorders, 31(1), 5–17.
Belmonte, M. K., Allen, G., Beckel-Mitchener, A., Boulanger,
L. M., Carper, R. A., & Webb, S. J. (2004). Autism and abnormal
Network Neuroscience
359
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
development of brain connectivity. Journal of Neuroscience,
24(42), 9228–9231. https://doi.org/10.1523/JNEUROSCI.3340-
04.2004
Birn, R. M., Molloy, E. K., Patriat, R., Parker, T., Meier, T. B., Kirk,
G. R., . . . Prabhakaran, V. (2013). The effect of scan length on
the reliability of resting-state fMRI connectivity estimates. Neuro-
Image,83,550–558. https://doi.org/10.1016/j.neuroimage.2013.
05.099
Byrge, L., & Kennedy, D. P. (2017). Identifying and characterizing
systematic temporally-lagged BOLD artifacts. NeuroImage, 171,
376–392. https://doi.org/10.1016/j.neuroimage.2017.12.082
Chen, C., Bailey, B., & Muller, R. A. (2015). Towards autism sub-
types? Unsupervised machine learning using fcMRI features.
Oral Presentation at the Organization for Human Brain Mapping
Annual Meeting, Honolulu, HI, USA.
Ciric, R., Wolf, D. H., Power, J. D., Roalf, D. R., Baum, G. L.,
Ruparel, K., . . . Satterthwaite, T. D. (2017). Benchmarking of
participant-level confound regression strategies for the control
of motion artifact in studies of functional connectivity. Neuro-
Image, 154, 1 7 4–1 8 7. https://doi.org/10.1016/j.neuroimage.
2017.03.020
Constantino, J. N., & Gruber, C. P. (2005). Social Responsiveness
Scale (SRS). Los Angeles, CA: Western Psychological Services.
Constantino, J. N., & Todd, R. D. (2003). Autistic traits in the general
population: A twin study. Archives of General Psychiatry, 60(5),
524–530. https://doi.org/10.1001/archpsyc.60.5.524
Courchesne, E., & Pierce, K. (2005). Why the frontal cortex in
autism might be talking only to itself: Local over-connectivity but
long-distance disconnection. Current Opinion in Neurobiology,
15(2), 225–230. https://doi.org/10.1016/j.conb.2005.03.001
Craddock, C., Benhajali, Y., Chu, C., Chouinard, F., Evans, A.,
Jakab, A., . . . Bellec, P. (2013). The Neuro Bureau Preprocessing
Initiative: Open sharing of preprocessed neuroimaging data and
derivatives. In Neuroinformatics 2013, Stockholm, Sweden.
Di Martino, A., Yan, C. G., Li, Q., Denio, E., Castellanos, F. X.,
Alaerts, K., . . . Milham, M. P. (2014). The autism brain imag-
ing data exchange: Towards a large-scale evaluation of the in-
trinsic brain architecture in autism. Molecular Psychiatry, 19(6),
659–667. https://doi.org/10.1038/mp.2013.78
Dosenbach, N. U., Nardos, B., Cohen, A. L., Fair, D. A., Power,
J. D., Church, J. A., . . . Schlagger, B. L. (2010). Prediction
of individual brain maturity using fMRI. Science, 329(5997),
1358–1361. https://doi.org/10.1126/science.1194144
Easson, A. K., Fatima, Z., & McIntosh, A. R.
(2019). Supporting
Information for “Functional connectivity-based subtypes of indi-
viduals with and without autism spectrum disorder.” Network
Neuroscience, 3(2), 344–362. https://doi.org/10.1162/netn_a_
00067
Fox, M. D., Snyder, A. Z., Vincent, J. L., Corbetta, M., Van Essen,
D. C., & Raichle, M. E. (2005). The human brain is intrinsically
organized into dynamic, anticorrelated functional networks. Pro-
ceedings of the National Academy of Sciences of the United
States of America, 102(27), 9673–9678. https://doi.org/10.1073/
pnas.0504136102
Georgiades, S., Szatmari, P., Boyle, M., Hanna, S., Duku, E.,
Zwaigenbaum, L., . . . Thompson, A. (2013). Investigating pheno-
typic heterogeneity in children with autism spectrum disorder:
Journal of Child Psy-
A factor mixture modeling approach.
chology and Psychiatry, 54(2), 206–215. https://doi.org/10.1111/
j.1469-7610.2012.02588.x
Gotts, S. J., Saad, Z. S., Jo, H. J., Wallace, G. L., Cox, R. W., &
Martin, A. (2013). The perils of global signal regression for
group comparisons: A case study of Autism Spectrum Dis-
orders. Frontiers in Human Neuroscience, 7, 356. https://doi.org/
10.3389/fnhum.2013.00356
Gramfort, A., Luessi, M., Larson, E., Engemann, D. A., Strohmeier,
D., Brodbeck, C., . . . Hamalainen, M. S. (2014). MNE software
for processing MEG and EEG data. NeuroImage, 86, 446–460.
https://doi.org/10.1016/j.neuroimage.2013.10.027
Gramfort, A., Luessi, M., Larson, E., Engemann, D. A., Strohmeier,
D., Brodbeck, C., . . . Hämäläinen, M. S.
(2013). MEG and
EEG data analysis with MNE-Python. Frontiers in Neuroscience,
7, 267. https://doi.org/10.3389/fnins.2013.00267
Greicius, M. D., Krasnow, B., Reiss, A. L., & Menon, V. (2003).
Functional connectivity in the resting brain: A network analysis
of the default mode hypothesis. Proceedings of the National
Academy of Sciences of the United States of America, 100(1),
253–258. https://doi.org/10.1073/pnas.0135058100
Hahamy, A., Behrmann, M., & Malach, R. (2015). The idiosyn-
cratic brain: Distortion of spontaneous connectivity patterns in
autism spectrum disorder. Nature Neuroscience, 18(2), 302–309.
https://doi.org/10.1038/nn.3919
Hong, S. J., Valk, S. L., Di Martino, A., Milham, M. P., & Bernhardt,
B. C. (2017). Multidimensional neuroanatomical subtyping of
Autism spectrum disorder. Cerebral Cortex, 1–11. https://doi.org/
10.1093/cercor/bhx229
.
.
. Urbanek, T.
Hrdlicka, M., Dudova, I., Beranova, I., Lisy, J., Belsan, T., Neuwirth,
(2005). Subtypes of autism by cluster
J.,
analysis based on structural MRI data. European Child &
Adolescent Psychiatry, 14(3), 138–144. https://doi.org/10.1007/
s00787-005-0453-z
Hull, J. V., Jacokes, Z. J., Torgerson, C. M., Irimia, A., & Van Horn,
J. D. (2016). Resting-state functional connectivity in Autism spec-
trum disorders: A review. Frontiers in Psychiatry, 7, 205. https://
doi.org/10.3389/fpsyt.2016.00205
Jonas (2017). Violin Plots for plotting multiple distributions (dis-
tributionPlot.m). MATLAB Central File Exchange. Retrieved from
https://www.mathworks.com/matlabcentral/fileexchange/23661-
violin-plots-for-plotting-multiple-distributions–distributionplot-m.
Just, M. A., Keller, T. A., Malave, V. L., Kana, R. K., & Varma, S.
(2012). Autism as a neural systems disorder: A theory of frontal-
posterior underconnectivity. Neuroscience & Biobehavioral Re-
views, 36(4), 1292–1313. https://doi.org/10.1016/j.neubiorev.
2012.02.007
Kaland, N., Smith, L., & Mortensen, E. L. (2008). Brief report:
Cognitive flexibility and focused attention in children and
adolescents with Asperger syndrome or high-functioning autism
as measured on the computerized version of the Wisconsin
Card Sorting Test. Journal of Autism and Developmental Dis-
orders, 38(6), 1161–1165. https://doi.org/10.1007/s10803-007-
0474-1
Kennedy, D. P., & Courchesne, E. (2008). The intrinsic functional
organization of the brain is altered in autism. NeuroImage, 39(4),
1877–1885. https://doi.org/10.1016/j.neuroimage.2007.10.052
Network Neuroscience
360
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
Keown, C. L., Shih, P., Nair, A., Peterson, N., Mulvey, M. E., &
Muller, R. A. (2013). Local functional overconnectivity in poste-
rior brain regions is associated with symptom severity in autism
spectrum disorders. Cell Reports, 5(3), 567–572. https://doi.org/
10.1016/j.celrep.2013.10.003
Krishnan, A., Williams, L. J., McIntosh, A. R., & Abdi, H. (2011).
Partial least squares (PLS) methods for neuroimaging: A tutorial
and review. NeuroImage, 56(2), 455–475. https://doi.org/10.
1016/j.neuroimage.2010.07.034
Lee, J. M., Kyeong, S., Kim, E., & Cheon, K. A. (2016). Abnormalities
of inter- and intra-hemispheric functional connectivity in Autism
spectrum disorders: A study using the Autism Brain Imaging Data
Exchange Database. Frontiers in Neuroscience, 10, 191. https://
doi.org/10.3389/fnins.2016.00191
Lord, C., Risi, S., Lambrecht, L., Cook, E. H., Jr., Leventhal, B. L.,
DiLavore, P. C., . . . Rutter, M. (2000). The Autism Diagnostic
Observation Schedule-Generic: A standard measure of social
and communication deficits associated with the spectrum of
Journal of Autism and Developmental Disorders, 30(3),
autism.
205–223.
Lord, C., Rutter, M., & Le Couteur, A. (1994). Autism Diagnostic
Interview-Revised: A revised version of a diagnostic interview
for caregivers of individuals with possible pervasive developmen-
tal disorders. Journal of Autism and Developmental Disorders,
24(5), 659–685.
McIntosh, A. R., Bookstein, F. L., Haxby, J. V., & Grady, C. L. (1996).
Spatial pattern analysis of functional brain images using partial
least squares. NeuroImage, 3(3 Pt 1), 143–157. https://doi.org/
10.1006/nimg.1996.0016
McIntosh, A. R., & Lobaugh, N. J. (2004). Partial least squares
analysis of neuroimaging data: Applications and advances.
NeuroImage, 23 Suppl 1, S250–263. https://doi.org/10.1016/
j.neuroimage.2004.07.020
Miller, H. L., Ragozzino, M. E., Cook, E. H., Sweeney, J. A., &
Mosconi, M. W. (2015). Cognitive set shifting deficits and their
relationship to repetitive behaviors in autism spectrum disorder.
Journal of Autism and Developmental Disorders, 45(3), 805–815.
https://doi.org/10.1007/s10803-014-2244-1
Monk, C. S., Peltier, S. J., Wiggins, J. L., Weng, S. J., Carrasco, M.,
Risi, S., & Lord, C. (2009). Abnormalities of intrinsic functional
connectivity in autism spectrum disorders. NeuroImage, 47(2),
764–772. https://doi.org/10.1016/j.neuroimage.2009.04.069
Mostofsky, S. H., Powell, S. K., Simmonds, D. J., Goldberg, M. C.,
Caffo, B., & Pekar, J. J. (2009). Decreased connectivity and cere-
bellar activity in autism during motor task performance. Brain,
132(Pt 9), 2413–2425. https://doi.org/10.1093/brain/awp088
Muller, R. A., & Amaral, D. G.
(2017). Editorial: Time to give up
on Autism Spectrum Disorder? Autism Research, 10(1), 10–14.
https://doi.org/10.1002/aur.1746
Muller, R. A., Shih, P., Keehn, B., Deyoe, J. R., Leyden, K. M., &
Shukla, D. K. (2011). Underconnected, but how? A survey of
functional connectivity MRI studies in autism spectrum disorders.
Cerebral Cortex, 21(10), 2233–2243. https://doi.org/10.1093/
cercor/bhq296
Munson, J., Dawson, G., Sterling, L., Beauchaine, T., Zhou, A.,
Elizabeth, K., . . . Abbott, R. (2008). Evidence for latent classes of
IQ in young children with autism spectrum disorder. American
Journal on Mental Retardation, 113(6), 439–452. https://doi.org/
10.1352/2008.113:439-452
Nunes, A. S., Peatfield, N., Vakorin, V., & Doesburg, S. M.
in
(2018).
autism spectrum disorder. NeuroImage. https://doi.org/10.1016/
j.neuroimage.2018.01.022
Idiosyncratic organization of cortical networks
Nomi, J. S., & Uddin, L. Q. (2015). Developmental changes in
large-scale network connectivity in autism. NeuroImage Clin, 7,
732–741. https://doi.org/10.1016/j.nicl.2015.02.024
Picci, G., Gotts, S. J., & Scherf, K. S. (2016). A theoretical rut:
Revisiting and critically evaluating the generalized under/over-
connectivity hypothesis of autism. Developmental Science, 19(4),
524–549. https://doi.org/10.1111/desc.12467
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes, K. A.,
Church, J. A., . . . Petersen, S. E.
(2011). Functional network
organization of the human brain. Neuron, 72(4), 665–678. https://
doi.org/10.1016/j.neuron.2011.09.006
Power, J. D., Mitra, A., Laumann, T. O., Snyder, A. Z., Schlaggar,
B. L., & Petersen, S. E. (2014). Methods to detect, character-
ize, and remove motion artifact in resting state fMRI. Neuro-
Image, 84, 320–341. https://doi.org/10.1016/j.neuroimage.2013.
08.048
Power, J. D., Plitt, M., Laumann, T. O., & Martin, A.
(2017).
Sources and implications of whole-brain fMRI signals in humans.
NeuroImage, 146, 609–625. https://doi.org/10.1016/j.neuroimage.
2016.09.038
Pruim, R. H. R., Mennes, M., Buitelaar, J. K., & Beckmann, C. F.
(2015a). Evaluation of ICA-AROMA and alternative strategies
for motion artifact removal in resting state fMRI. NeuroImage,
112, 278–287. https://doi.org/10.1016/j.neuroimage.2015.02.
063
Pruim, R. H. R., Mennes, M., van Rooij, D., Llera, A., Buitelaar, J. K.,
& Beckmann, C. F.
ICA-AROMA: A robust ICA-based
strategy for removing motion artifacts from fMRI data. Neuro-
Image, 112, 267–277. https://doi.org/10.1016/j.neuroimage.2015.
02.064
(2015b).
Rashid, B., Blanken, L. M. E., Muetzel, R. L., Miller, R., Damaraju,
E., Arbabshirani, M. R., . . . Calhoun, V.
(2018). Connectivity
dynamics in typical development and its relationship to autis-
tic traits and Autism spectrum disorder. Human Brain Mapping,
39(8), 3127–3142. https://doi.org/10.1002/hbm.24064
Rudie, J. D., Brown, J. A., Beck-Pancer, D., Hernandez, L. M.,
Dennis, E. L., Thompson, P. M., . . . Dapretto, M. (2013a).
Altered functional and structural brain network organization in
autism. NeuroImage Clinical, 2, 79–94. https://doi.org/10.1016/
j.nicl.2012.11.006
Rudie, J. D., & Dapretto, M. (2013b). Convergent evidence of brain
overconnectivity in children with autism? Cell Reports, 5(3),
565–566. https://doi.org/10.1016/j.celrep.2013.10.043
Rudie,
J. D., Shehzad, Z., Hernandez, L. M., Colich, N. L.,
Bookheimer, S. Y., Iacoboni, M., & Dapretto, M. (2012). Reduced
functional integration and segregation of distributed neural sys-
tems underlying social and emotional information processing in
autism spectrum disorders. Cerebral Cortex, 22(5), 1025–1037.
https://doi.org/10.1093/cercor/bhr171
Shen, M. D., Shih, P., Ottl, B., Keehn, B., Leyden, K. M., Gaffrey,
M. S., & Muller, R. A. (2012). Atypical lexicosemantic function
Network Neuroscience
361
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectivity subtypes of ASD and controls
of extrastriate cortex in autism spectrum disorder: Evidence
from functional and effective connectivity. NeuroImage, 62(3),
1780–1791. https://doi.org/10.1016/j.neuroimage.2012.06.008
Spronk, M., Kulkarni, K., Ji, J. L., Keane, B., Anticevic, A., & Cole,
M. W. (2018). A whole-brain and cross-diagnostic perspective on
functional brain network dysfunction. bioRxiv 326728; https://
doi.org/10.1101/326728
Turner, K. C., Frost, L., Linsenbardt, D., McIlroy, J. R., & Muller, R. A.
(2006). Atypically diffuse functional connectivity between cau-
date nuclei and cerebral cortex in autism. Behavioral and Brain
Functions, 2, 34. https://doi.org/10.1186/1744-9081-2-34
Uddin, L. Q., Supekar, K., Lynch, C. J., Khouzam, A., Phillips, J.,
(2013a). Salience network-based
Feinstein, C., . . . Menon, V.
classification and prediction of symptom severity in children
with autism. JAMA Psychiatry, 70(8), 869–879. https://doi.org/
10.1001/jamapsychiatry.2013.104
Uddin, L. Q., Supekar, K., & Menon, V. (2013b). Reconceptualiz-
ing functional brain connectivity in autism from a developmental
perspective. Frontiers in Human Neuroscience, 7, 458. https://
doi.org/10.3389/fnhum.2013.00458
Weng, S. J., Wiggins, J. L., Peltier, S. J., Carrasco, M., Risi, S., Lord,
C., & Monk, C. S. (2010). Alterations of resting state functional
connectivity in the default network in adolescents with autism
spectrum disorders. Brain Research, 1313, 202–214. https://doi.
org/10.1016/j.brainres.2009.11.057
Xu, T., Yang, Z., Jiang, L., Xing, X., & Zuo, X. (2015). A connectome
computation system for discovery of brain science. Science Bul-
letin, 60(1), 86–95. https://doi.org/10.1007/s11434-014-0698-3
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
3
2
3
4
4
1
0
9
2
5
6
1
n
e
n
_
a
_
0
0
0
6
7
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
362