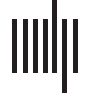RECHERCHE
Resting-state brain network features associated
with short-term skill learning ability in humans
and the influence of N-methyl-D-aspartate
receptor antagonism
1
Zhenxiang Zang
1
, Lena S. Geiger
, Urs Braun
, Hengyi Cao
, Maria Zangl
1
1
1
Carolin Moessnang
Alexander Moscicki
3
, Matthias Ruf
, Janine Reis
1
1
, Emanuel Schwarz
1
, Luanna Dixson
, Janina I. Schweiger
1
, and Heike Tost
, Andreas Meyer-Lindenberg
2
1
1
, Axel Schäfer
1
,
1
,
un accès ouvert
journal
1Department of Psychiatry and Psychotherapy, Research Group System Neuroscience in Psychiatry,
Central Institute of Mental Health, University of Heidelberg, Medical Faculty Mannheim, Mannheim, Allemagne
2Department of Neuroimaging, Central Institute of Mental Health, University of Heidelberg, Medical Faculty Mannheim,
Mannheim, Allemagne
3Department of Neurology and Neurophysiology, Albert-Ludwigs-University, Freiburg, Allemagne
Mots clés: Resting-state fMRI, Short-term motor learning, System neuroscience, Functional brain
réseaux, NMDA receptor-related plasticity
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
un
_
0
0
0
4
5
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
ABSTRAIT
Graph theoretical functional magnetic resonance imaging (IRMf) studies have demonstrated
that brain networks reorganize significantly during motor skill acquisition, yet the
associations between motor learning ability, brain network features, and the underlying
biological mechanisms remain unclear. In the current study, we applied a visually guided
sequential pinch force learning task and graph theoretical analyses to investigate the
associations between short-term motor learning ability and resting-state brain network metrics
dans 60 healthy subjects. We further probed the test-retest reliability (n = 26) and potential
effects of the N-methyl-D-aspartate (NMDA) antagonist ketamine (n = 19) in independent
healthy volunteers. Our results show that the improvement of motor performance after
short-term training was positively correlated with small-worldness ( p = 0.032) and global
efficiency (p = 0.025), whereas negatively correlated with characteristic path length
(p = 0.014) and transitivity (p = 0.025). En outre, using network-based statistics (NBS), nous
identified a learning ability–associated ( p = 0.037) and ketamine-susceptible ( p = 0.027)
cerebellar-cortical network with fair to good reliability (intraclass correlation coefficient
[ICC] > 0.7) and higher functional connectivity in better learners. Our results provide new
evidence for the association of intrinsic brain network features with motor learning and
suggest a role of NMDA-related glutamatergic processes in learning-associated subnetworks.
RÉSUMÉ DE L'AUTEUR
Learning a new motor skill prompts immediate reconfigurations of distributed brain networks
followed by adaptive changes in intrinsic brain circuits related to synaptic plasticity. Ici,
we identify global brain network properties and a cerebellar-cortical functional subnetwork
that are both significantly associated with motor learning ability in a previously trained
visuomotor task in humans. We further show that the associated functional subnetwork
connectivity but not the global brain network properties are susceptible to ketamine. Notre
findings suggest a distinct functional role for learning-related global versus local network
metrics and support the idea of a preferential susceptibility of learning-associated
subnetworks to N-methyl-D-aspartate antagonist and plasticity-related consolidation effects.
Citation: Zang, Z., Geiger, L. S., Brun,
U., Cao, H., Zang, M., Schäfer, UN., . . . &
Tost, H. (2018). Resting-state brain
network features associated with
short-term skill learning ability in
humans and the influence of
N-methyl-D-aspertate receptor
antagonism. Neurosciences en réseau,
2(4), 464–480. https://est ce que je.org/
10.1162/netn_a_00045
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00045
Informations complémentaires:
https://doi.org/10.1162/netn_a_00045
Reçu: 16 Août 2017
Accepté: 11 Janvier 2018
Auteur correspondant:
Heike Tost
heike.tost@zi-mannheim.de
Éditeur de manipulation:
Alex Fornito
droits d'auteur: © 2018
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
Brain functional connectome and skill learning
INTRODUCTION
The acquisition of new motor skills requires the brain to flexibly reconfigure neural circuits
to master a desired performance level (Bassett & Mattar, 2017). Recent studies have demon-
strated that different circuits are involved at distinct stages of learning (Dayan & Cohen, 2011;
Penhune & Steele, 2012). Whereas the initial learning phase engages a widespread network
consisting of primary motor area (M1), supplementary motor area (SMA), basal ganglia (BG),
dorsolateral prefrontal cortex (DLPFC), premotor cortex, and posterior cerebellum, the follow-
ing longer term learning phase relies on a smaller set of brain regions including M1, SMA, BG,
and the lateral cerebellum (Dayan & Cohen, 2011).
En outre, the specific type of motor
learning task determines the preferential involvement of brain regions with sequential learn-
ing challenging cortical areas while more complex sensorimotor tasks with novel kinematic
additionally challenge the BG and cerebellum (Hardwick, Rottschy, Miall, & Eickhoff, 2013).
The interactions between brain regions during motor learning can be studied in the frame-
work of brain networks. By combining network analysis and functional magnetic resonance
imaging (IRMf), recent studies have shown that brain network features including flexibility
(Bassett et al., 2011), connectivity strength, local path length, and nodal efficiency (Heitger
et coll., 2012; Sami & Miall, 2013) change in response to motor learning and can predict its
rate (Bassett et al., 2011). Notably, changes in the brain network architecture cannot only
be assessed during the process of motor learning by using task-based fMRI, but also during
repos. Although there is some evidence that intrinsic network connectivity measures derived
from prior resting-state fMRI (rs-fMRI) predict motor learning abilities (Mawase, Bar-Haim, &
Shmuelof, 2017; Wu, Srinivasan, Kaur, & Cramer, 2014), recent studies also suggest that motor
learning effects can be detected using rs-fMRI after task practice (Albert, Robertson, & Miall,
2009; Sami & Miall, 2013; Sami, Robertson, & Miall, 2014). Cependant, whereas plasticity-
related effects of motor learning likely shape the intrinsic configuration of brain circuits, le
biological mechanisms in humans remain largely unknown.
Plausible molecular mechanisms contributing to motor learning–related network changes
include glutamate-dependent processes (Dayan & Cohen, 2011). Supportive evidence is pro-
vided by animal studies showing that motor training can shift the glutamatergic N-methyl-D-
aspartate (NMDA) receptor subunit composition in BG (Kent, Deng, & McNeill, 2013) et
promote the NMDA-dependent synaptic plasticity in the primary motor cortex of rats (Kida
et coll., 2016), while impaired motor performance was observed in mGluR4 gene knockout mice
(Pekhletski et al., 1996). In humans, evidence for the involvement of glutamate-dependent pro-
cesses during motor learning is less direct. Ici, many studies have focused on the effects of a
common functional polymorphism (Val66Met) in the brain-derived neurotrophic factor (BDNF)
gene (Fritsch et al., 2010; McHughen et al., 2010), a downstream modulator of the molecular
cascade supporting synaptic plasticity linked to motor learning impairments and altered motor
cortical activations in the plasticity-impaired Met allele carriers (Fritsch et al., 2010; McHughen
et coll., 2010; Thomason, Yoo, Glover, & Gotlib, 2009). For the evidence in humans, a study by
Hadj Tahar et al. further showed that the NMDA receptor antagonist amantadine significantly
impairs motor learning in healthy subjects (Hadj Tahar, Blanchet, & Doyon, 2004).
In the current work we aimed to answer two main questions in healthy humans: first,
whether the brain’s resting-state network configuration relates to individual differences in short-
term motor learning; et deuxieme, whether these metrics can be influenced by NMDA receptor
antagonism. We first investigated whether resting-state network features relate to individual
differences in short-term motor learning ability by combining an established sequential vi-
sual isometric pinch force learning task (Reis et al., 2009) with rs-fMRI and graph theoretical
N-methyl-D-aspartate receptor
(NMDA):
NMDA receptor is a glutamate
receptor (ion channel protein)
in the central nervous system
that is very important for synaptic
plasticity and memory function.
Neurosciences en réseau
465
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
un
_
0
0
0
4
5
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain functional connectome and skill learning
Ketamine:
Ketamine is a noncompetitive
NMDA receptor antagonist that
binds specifically to a subsite of
NMDA receptor and blocks ion
channels in the central nervous
système. It is normally used to
induce and maintain general
anesthesia before, pendant,
and after surgery.
Sequential visual isometric
pinch force task:
Subjects are required to pinch
their right thumb and index
finger against a force transducer
to achieve the “target” by
looking at the cursor
displayed on the monitor.
analyses. We hypothesized that both global network diagnostics and functional connectivity
among a circumscribed set of brain visuomotor brain areas would relate to individual mo-
tor learning ability (Doyon & Benali, 2005; Hikosaka, Nakamura, Sakai, & Nakahara, 2002).
Deuxième, we tested whether ketamine influences the functional connectivity of motor learning–
related subnetworks. Ici, we hypothesized that NMDA receptor blockade would decrease
the connectivity of motor learning–related subnetworks.
MATERIALS AND METHODS
Participants
Sixty healthy right-handed volunteers (mean age 26.6 ± 7.5 années, 33 men) underwent visuo-
motor training followed by a resting-state fMRI scan (mean training duration: 26.9 ± 5.7 min;
mean time interval between motor training and fMRI scan: 45.8 ± 7.5 min). Exclusion criteria
included MRI contraindications, a history of psychiatric and neurological illness, prior head
trauma, and current alcohol or drug abuse. None of the subjects had a first-degree relative with
a psychiatric disorder or received psychopharmacological treatment. All participants provided
written, informed consent for a protocol approved by the Ethics Committee of the University
of Heidelberg.
Visuomotor Learning Task
Behavioral training consisted of a single session with a modified version of an established (Reis
et coll., 2009) sequential visual isometric pinch force task. Subjects were seated 80 cm in front
of a 28-inch monitor depicting a home position and five target gates (G1-G5, Chiffre 1) alors que
holding a force transducer between their right thumb and index finger. The application of
pinch force moved a screen cursor from the home position in a right hand direction toward
the target gates, whereas relaxation resulted in a leftward cursor movement back toward the
home position. The distance of the cursor to the home position increased logarithmically with
increasing pinch force in order to make the task more difficult. Subjects were instructed to
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
un
_
0
0
0
4
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 1. Setup of the sequential visual isometric pinch task (see Materials and Methods for de-
tails). Subjects were asked to move the cursor into the highlighted targets (c'est à dire., G2) as fast and
as accurate as possible. The sequence of targets was 2-5-3-1-4. (Manikin illustration ©Petr Ciz –
Fotolia.com).
Neurosciences en réseau
466
Brain functional connectome and skill learning
Skill measure:
Skill measure is an index to
reflect the performance level
by combining speed and
accuracy with a nonlinear
formula.
modulate their pinch force so that the cursor navigated as quickly and accurately as possible
along the following sequence: home-G2-home-G5-home-G3-home-G1-home-G4. After getting
familiar with the setting, subjects performed four training blocks consisting of 35 trials (com-
pleted sequences) chaque. Movement times per trial were measured from movement onset in
the home position to stopping at the last gate (G4). Error rates were calculated as ratio of gates
per block with over- or undershooting cursor movements (missed gates).
Definition of Skill Learning
Following prior work with this task (Reis et al., 2009), we calculated individual skill measures
for each block using the following formula:
skill measure = ln(
1 − error rate
error rate(ln (duration)5.424)
)
(1)
where duration is the average movement time across the trials of the block, and error rate
is the rate of over- and undershoots across the trials of the block (Reis et al., 2009). Sur
all training blocks, individual differences in skill learning ability were calculated based on
the difference in the skill measure between the last and first training block (skill learning =
skill measureblock4 − skill measureblock1).
MRI Data Acquisition
Neuroimaging was performed on a 3T MRI scanner (Siemens Trio, Erlangen, Allemagne) equipped
with a 32 channel multi-array head-coil. Details on MRI sequences are given in Supporting
Information (Zang, Geiger, Brun, Cao, Zang, Schäfer, . . . & Tost, 2018).
fMRI Data Processing
Image processing was performed using standard routines implemented in the Statistical Para-
metric Mapping software (SPM8, http://www.fil.ion.ucl.ac.uk/spm/software/spm8/) et le
Data Processing Assistant for Resting-State fMRI toolbox [DPARSF, (C. Yan & Zang, 2010)]. All
images were realigned to the first image of the time series, corrected for slice timing, spatially
normalized to the Montreal Neurological Institute (MNI) EPI template, and spatially smoothed
with an 8-mm full-width at half-maximum Gaussian kernel. For each participant, we then ex-
tracted the mean time series from 264 brain regions derived from the Automated Anatomical
Labeling (AAL 116) brain atlas (Tzourio-Mazoyer et al., 2002) by random parcellation (Zalesky,
Fornito, Harding, et coll., 2010). From the node time series, we regressed out the time series of
white matter and cerebrospinal fluid masks (derived from SPM tissue probability maps thresh-
olded at 90% for CSF and 99% for WM; Cao et al., 2014) and the six head motion parameters
from the realignment step. The resulting residual time series were temporally filtered using a
0.01–0.1 Hz band-pass filter.
Quantification of Head Displacements
The functional connectivity estimates and network diagnostics derived from resting-state fMRI
may be impacted by motion artifacts (Power, Barnes, Snyder, Schlaggar, & Petersen, 2012;
Satterthwaite et al., 2013; C. G. Yan et al., 2013). To account for this, we used in-house
software to estimate averaged frame-wise displacement (FD) (Power et al., 2012) and included
average FD as covariates of noninterest in our analyses.
Neurosciences en réseau
467
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
un
_
0
0
0
4
5
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain functional connectome and skill learning
Construction of Connectivity Matrices
For the construction of brain networks, we computed pairwise Pearson correlation coeffi-
cients between the processed time series of each node, which resulted in a 264 × 264 deux-
dimensional matrix for each subject. We then thresholded the matrices in 1% intervals over a
range of 40 densities from 1% à 40% to generate binary graphs (par exemple., dans le 1% thresholded
matrix only the top 1% of the highest positive correlations are represented by assigning a value
de 1 to the internode connections).
Calculation and Analysis of Graph Diagnostics
On the global brain network level, graph features were computed using the Brain Connectivity
Toolbox (Rubinov & Sporns, 2010). Specifically, for each density, we calculated seven reliable
(Cao et al., 2014) global brain network markers that reflect the integration and segregation of
whole-brain network and were shown to be in association with cognitive functions (Alavash
et coll., 2015): Transitivity, characteristic path length, global efficiency, small-worldness, mod-
ularity Q (Newman, 2006), assortativity, and mean connectivity coefficient. The detailed de-
scriptions of the seven markers are given in the Supporting Information (Zang et al., 2018).
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
un
_
0
0
0
4
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Network-based statistics (NBS):
NBS is a method to perform group
level statistics on correlation
matrix by using a general linear
model. An arbitrary initial
threshold is needed to determine
the network cluster, et
permutations will be performed
on the determined network for
addressing multiple comparison
correction.
For association with the degree of skill learning, the network properties were averaged
across densities and introduced as a dependent variable into separate linear regression mod-
els with skill learning as independent variables of interest and age, sex, and averaged FD as
covariates of noninterests. Hochberg’s stepwise p value adjustment method (Hochberg, 1988)
was used to correct raw p values for multiple hypothesis testing.
Network-Based Statistics
We analyzed the connectivity matrices with network-based statistics (NBS) to identify clusters
of node connections associated with skill learning ability. Compared with the mass-univariate
testing of independent links, NBS offers higher statistical power by identifying connected com-
ponents from a set of uncorrected thresholded links that are significantly associated with a vari-
able of interest (Zalesky, Fornito, & Bullmore, 2010), and then uses a randomization approach
to evaluate the null hypothesis on the level of connected subclusters (rather than individually
for each connection). Following prior procedures (Wang et al., 2013), we defined initial linear
regression models for each of the (N (N-1))/2 = 34, 716 (N = 264) possible links in the con-
nectivity matrices. The regression models included skill learning as an independent variable
of interest and age, sex, and the averaged FD as covariates of noninterest. From the resulting
p matrix, we defined a set of suprathreshold connections by isolating all links with t > 3.48
and p < 5 × 10−4
and used M = 5, 000 permutations (Wang, Zuo, & He, 2010) to estimate
the null distribution during permutation testing of the identified cluster association.
Supplemental Analyses
To further probe the quality of the skill learning–related NBS result, we further (a) quantified
the test-retest reliability of the mean connectivity of the identified cerebellar-cortical cluster,
(b) considered the potential role of structural confounds by testing the relationship between
skill learning and gray matter volume of the nodes contributing to the cluster, and (c) explored
the effects of low-dose ketamine as NMDA receptor antagonist on skill learning ability and
the connectivity of the identified NBS cluster. Additionally, we aimed to probe (d) the robust-
ness of our results by using a more conservative head motion correction approach and (e) the
Network Neuroscience
468
Brain functional connectome and skill learning
Global network features:
In the current study, global
network features are defined as
small-worldness, global efficiency,
characteristic path length,
modularity Q, transitivity,
assortativity, and mean
correlation coefficient.
specificity of the association between motor learning ability and global network features by
controlling for the mean functional connectivity as covariate of noninterest. Finally, we exam-
ined the identified cerebellar-cortical network association to skill learning with respect to po-
tential effects of the choice of the initial cluster-forming significance threshold and parcellation
scheme for NBS, respectively, by exploring the outcome of (f) two additional cluster-forming
significance thresholds (p < 0.001, p < 0.0001) and (g) an alternative whole-brain functional
atlas (Rosenberg et al., 2016) containing a comparable number of nodes (268 parcellations)
as our AAL-based atlas.
As previous studies have demonstrated that the reliability of functional
Test-retest reliability.
connectivity estimates is spatially heterogeneous (Mueller et al., 2015), we aimed to establish
the robustness of the connectivity estimates in the identified subnetwork before further explor-
ing it in the context of a pharmacological challenge study. To quantify the test-retest reliability
of the connectivity phenotype, we reanalyzed the resting-state reliability data reported in Cao
et al. (2014). Following the nomenclature of Fleiss (1986), we considered an ICC value below
0.4 as poor, 0.4–0.75 as fair to good, and >0.75 as excellent. Detailed information about
fMRI data is given in the Supporting Information (Zang et al., 2018).
Structural correlates. We analyzed the high-resolution structural data with the voxel-based
morphometry toolbox (VBM8, http://dbm.neuro.uni-jena.de/vbm8/) by using default parame-
ters. Detailed descriptions of preprocessing the structural data are provided in the Supporting
Information (Zang et al., 2018). We then extracted the GM volume of the nodes contributing
to the identified NBS cluster and entered the sum GM volume as a dependent variable in a
regression model that included skill learning as independent variable of interest and age and
sex as covariates of noninterest (significance level: p < 0.05).
To quantify the effects of the NMDA receptor antagonist ketamine
NMDA receptor challenge.
on the identified cerebellar-cortical subnetwork, we analyzed the ketamine challenge data re-
ported in (Francois et al., 2016; Grimm et al., 2015). In this study, resting-state fMRI data were
acquired in 24 healthy individuals (12 women, mean age 25 years, mean body weight 70 kg)
undergoing three consecutive fMRI sessions over the course of 3 weeks. The pharmacolog-
ical protocol followed a double blind, placebo-controlled, order randomized, three-period
cross-over design with single intravenous doses of either saline (placebo condition), ketamine
(0.5 mg/kg body weight), or scopolamine (4 μg/kg body weight). Drug infusions started 73.8 ±
13.8 minutes prior to the resting-state scan and were 40.02 ± 6.02 min in duration. The vi-
suomotor learning task started 15.6 ± 3.5 min after infusion onset and was completed around
the end of the infusion (at 40.4 ± 8.61 min). To ensure this, we used the same experimen-
tal setup for the training as in the main study except for a slightly shorter training duration
(25 trials for each block, 4 blocks in total). Since we did not test a hypothesis assuming effects
of mAch-blockade (scopolamine condition) on the identified subnetwork phenotype, we only
analyzed the rs-fMRI data from the ketamine and placebo conditions in the current study. For
this, we created a covariate of noninterest coding for the order of ketamine and placebo condi-
tions (ketamine first, placebo first) that was included in the applied repeated-measure ANOVA.
For pharmacokinetic analysis, blood samples for quantification of norketamine plasma lev-
els were drawn immediately before and after the MRI scan (see Francois et al., 2016, and
Grimm et al., 2015, for details). The time interval between ketamine and placebo infusion was
9.6 ± SD 3.5 days. One subject was excluded because of side effects under ketamine
(Francois et al., 2016); four more subjects were excluded because they had already partic-
ipated in either the current (3 subjects) or in other visuomotor learning studies (1 subject).
Network Neuroscience
469
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
a
_
0
0
0
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain functional connectome and skill learning
In total, 19 subjects were included in further data analyses. The processing of the behavioral
data, rs-fMRI data, node definitions, and construction of connectivity matrices followed the
protocol described above. To test for drug effects, we extracted the mean connectivity from
the links of the cerebellar-cortical subnetwork identified in the NBS analysis (see Results sec-
tion) and used a repeated-measures ANOVA with drug as within-subjects factor and age, sex
(as factor), body mass index (BMI), the order (as factor) of drug, and the differences of aver-
aged FD (placebo vs. ketamine condition) as covariates of noninterest. To directly relate the
connectivity indices of the identified subnetwork to the administration of the drug in the ke-
tamine condition, we quantified intravenous norketamine levels by chromatographic analysis
from the blood samples taken immediately prior to the MRI scan. For details on the blood
sample processing, please refer to Francois et al. (2016). We used a linear regression model
in which the norketamine values were introduced as a dependent variable, the mean connec-
tivity estimates from the network links as independent variable of interest and age, sex, BMI,
and averaged FD as nuisance covariates (significance level: p < 0.05). Detailed descriptions
about drug administration are provided in the Supporting Information (Zang et al., 2018).
As global differences in con-
Controlling for mean individual functional connectivity differences.
nectivity strength might directly influence network properties (van den Heuvel et al., 2017),
we aimed to replicate our results by using the individual mean functional connectivity average
over all connections as an additional covariate of noninterest in our analyses.
Since sharp in-scanner motion can introduce system-
Scrubbing to correct for head motion.
atic, artificial connectivity (Power et al., 2012), we aimed to replicate our findings by using
a “scrubbing” approach as described in detail in Power et al. (2012). In short, all frames of
the time series with a FD >0.5 mm were removed. Two subjects were excluded from this
analysis because their number of spikes exceeded 10% of the total time points, leaving a total
de 58 sujets.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
un
_
0
0
0
4
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
RÉSULTATS
Skill Learning Ability
Training improved sequential visual isometric pinch task performance as indicated by a sig-
nificant decrease in the trial durations and error rates and a significant increase in the skill
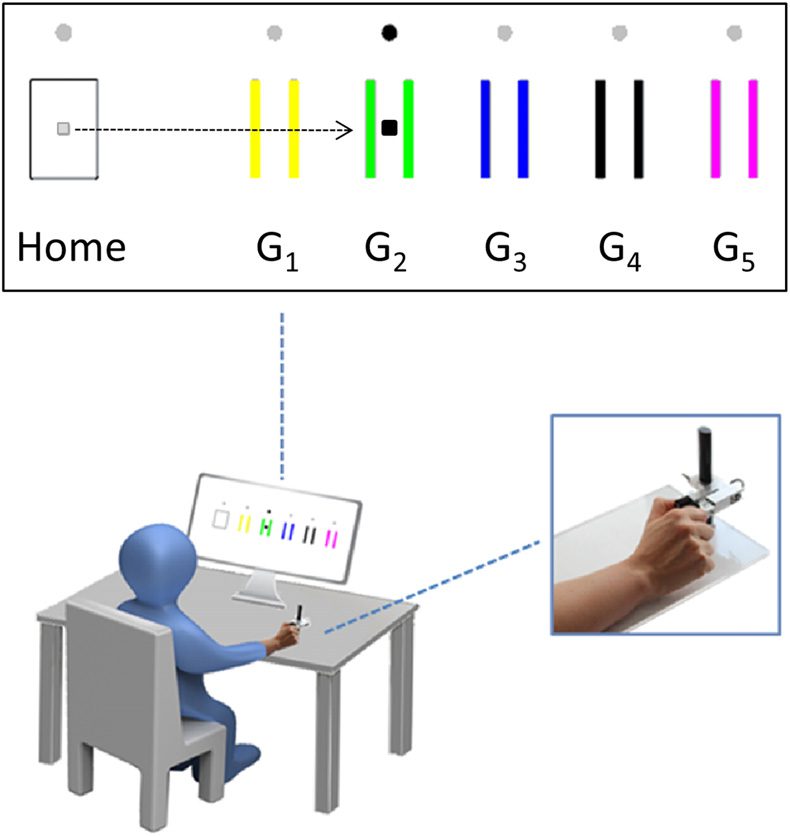
measure (Figure 2A) across blocks (F(3,57) values > 4.27, all p values < 0.009). The analysis of
the skill learning measure confirmed a significant increase in skill performance (skill measure
block 4 to block 1) at the end of the training (one-sample t test, t(59) = 11.43, p = 1.2 × 10−16
).
Relationship to Graph-Based Diagnostics
At the global brain network level, we observed significant associations between the individual
skill learning ability and four of the seven graph diagnostics. Whereas positive correlations
were found for small-worldness (t(55) = 2.73, r = 0.35, praw = 0.008, pcorr = 0.032, Figure 2B)
and global efficiency (t(55) = 2.90, r = 0.36, praw = 0.005, pcorr = 0.025, Figure 2C), we
detected negative associations for characteristic path length (t(55) = −3.33, r = −0.41, praw =
0.002, pcorr = 0.014, Figure 2D) and transitivity (t(55) = −2.92, r = −0.37, praw = 0.005,
pcorr = 0.025, Figure 2E). We observed no significant associations between skill learning ability
and assortativity and modularity Q of the network (all pcorr values > 0.225; Table S2, Zang
et coll., 2018). En outre, there was no significant correlation between skill learning ability
Neurosciences en réseau
470
Brain functional connectome and skill learning
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
un
_
0
0
0
4
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 2.
(UN) Skill increase in the sequential visual isometric pinch task across the training blocks (dots depict the mean values of the skill
measure across blocks). Error bars indicate standard errors. (B–E) Regression plots show significant associations of short-term motor learning
ability (block 4 to block 1) and resting-state fMRI-derived graph diagnostics (adjusted for covariates and constant; see Results section for details)
after controlling for age, sex, and FD.
and whole-brain mean connectivity (r = −0.11, praw = 0.41; Table S2, Zang et al., 2018). All
calculated graphs displayed small-world network properties (σ = γ/λ > 1, range 1.02–2.69).
Relationship to Subnetwork Functional Connectivity
Although we did not detect a significant association between short-term skill learning ability
and the mean correlation estimates of the whole-brain functional connectome (r = −0.11,
praw = 0.41; Table S2, Zang et al., 2018), significantly associated brain subnetworks likely
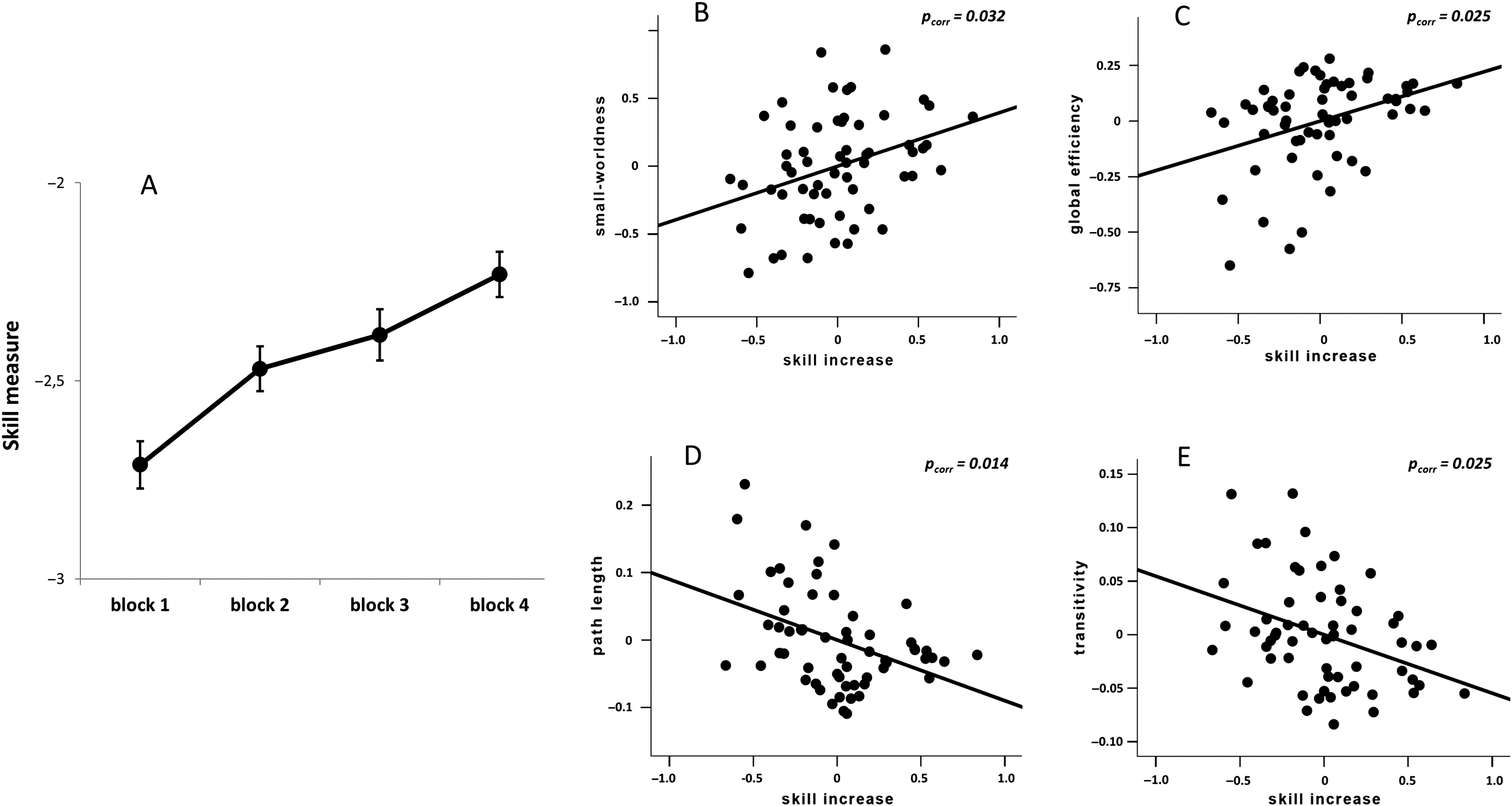
exister. Consistent with this notion, NBS identified a cluster of links with a significant positive
association between skill learning ability and the functional connectivity estimates of the clus-
ter links (uncorrected initial p < 5 × 10−4
, FWE corrected p = 0.037; Figure 3). The cluster
consisted of 69 nodes and 91 links mainly interconnecting the cerebellum, frontal, and pari-
etal lobes. Specifically, most of the links of this cluster connected area 7b and area 8 of the
left cerebellum to the cortex, in particular, to nodes mapping in proximity to M1, primary sen-
sory cortex, SMA, dorsal premotor cortex, intraparietal sulcus, and the motion sensitive visual
processing area V5. A detailed description of all nodes and links of the identified subnetwork
is provided in Table S1 (Zang et al., 2018).
Supplemental Analyses
The test-retest reliability analysis of the connectivity estimates of the
Test-retest reliability.
NBS subnetwork yielded an ICC2,1 of 0.72 and an ICC3,1 of 0.73, respectively. This indicates
Network Neuroscience
471
Brain functional connectome and skill learning
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
Figure 3.
Illustration of the NBS-derived cerebellar-cortical functional network associated with
short-term skill learning. Spheres represent center-of-gravity coordinates of the NBS-derived regions.
Images are visualized using BrainNet Viewer (Xia, Wang, & He, 2013). Detailed information can
be found in Table S1 (Zang et al., 2018).
f
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
a
_
0
0
0
4
5
p
d
t
.
an almost excellent robustness of the connectivity estimates of the cluster identified to relate
to skill learning ability.
The structural analysis did not provide any evidence for an association
Structural analysis.
between the mean gray matter volume of the 69 subnetwork nodes and skill learning ability
(t(56) = −0.33, p = 0.74). Also, we detected no significant correlations between mean gray
matter volume of the 69 subnetwork nodes and the mean functional connectivity estimates of
the 91 links of the NBS subnetwork (t(56) = −0.10, p = 0.92) or any of the four whole-brain
graph features identified to relate to skill learning ability (all |t(56)| < 1.28, p > 0.21). Ce
makes the influence of structural confounds on skill learning ability and its association with
the identified NBS subnetwork unlikely.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Effects of NMDA receptor challenge. We detected no significant behavioral differences for skill
increase (block 4 to block 1, F(1,17) = 0.33, p = 0.86), task duration (F(1,17) = 1.82, p = 0.20)
or error rate (F(1,17) = 0.48, p = 0.50) between the placebo and ketamine conditions (drug
In comparison to placebo, application of
order was included as covariate of noninterest).
ketamine did not result in significant differences in global network measures (all F(1,12) < 0.83,
p > 0.38) or whole-brain mean connectivity (F(1,12) = 2.04, p = 0.18), but significantly
decreased the mean functional connectivity of the learning-associated cerebellar-cortical
Neurosciences en réseau
472
Brain functional connectome and skill learning
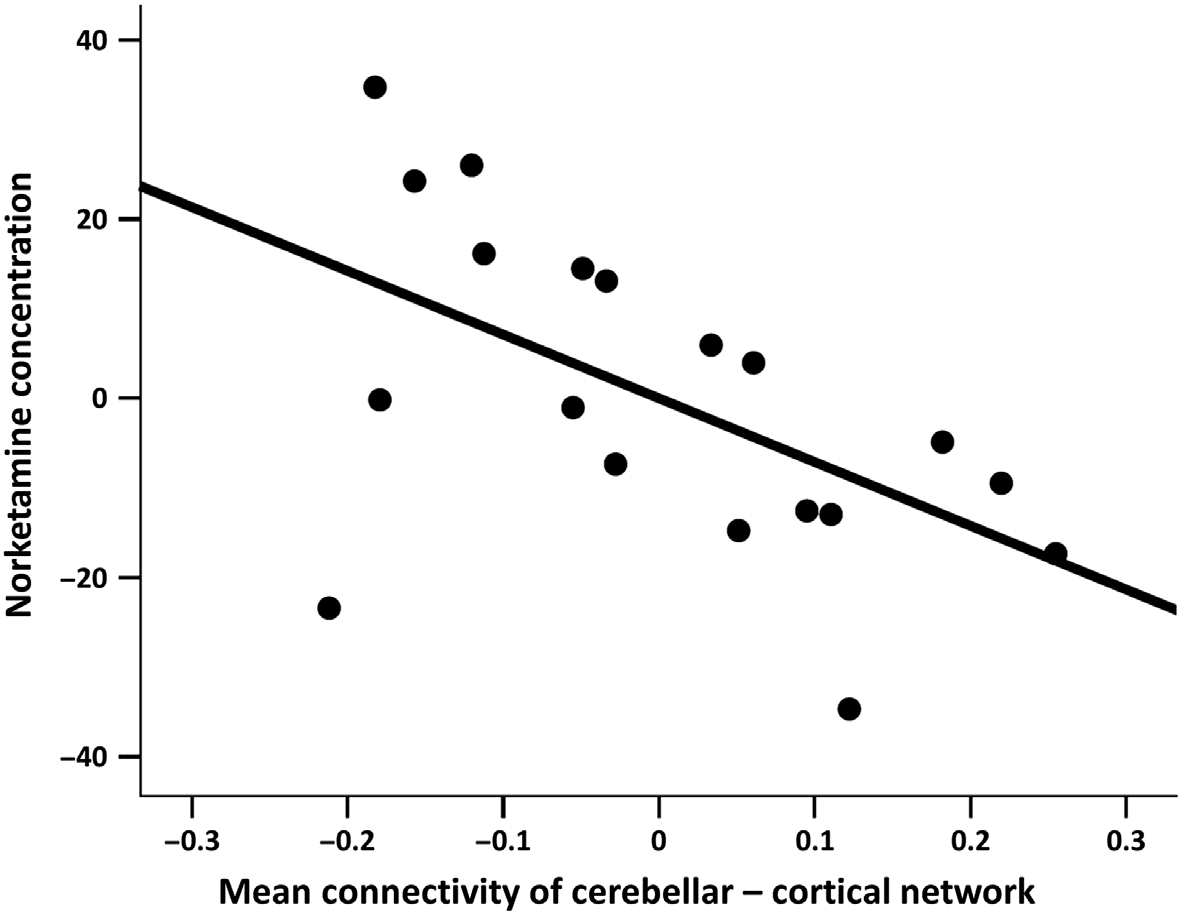
Chiffre 4. Partial correlation plot of negative correlation between blood Norketamine concen-
tration and the mean connectivity of the NBS-derived cerebellar-cortical network in the ketamine
condition (p = 0.032; adjusted for covariates and constant), controlled for age, sex, BMI, and FD.
réseau (F(1,12) = 6.38, p = 0.027).
En outre, the mean connectivity of the cerebellar-
cortical network was significantly negatively correlated with individual Norketamine concen-
trations (46.1 ± 21.6 ng/ml, t(13) = −2.40, r = −0.55, p = 0.032, Chiffre 4) in the ketamine
condition (âge, sex, FD, and BMI were controlled as covariates of noninterest). Average FD
(p = 0.424) and the time interval between drug infusion and resting-state scan (p = 0.219)
were not significantly different between the ketamine and placebo conditions. A trend toward
a main effect of drug order was found (p = 0.06). The results indicate that the functional con-
nectivity of the NBS-derived cerebellar-cortical network is affected by ketamine and negatively
associated to the concentration of the major active metabolite (c'est à dire., Norketamine).
Controlling for individual mean functional connectivity differences. Using the mean functional
connectivity as an additional covariate of noninterests, we replicated the findings that skill
increase is significantly positively associated with small-worldness (p = 0.008) and global ef-
ficiency (p = 0.005) and negatively associated with characteristic path length (p = 0.002) et
transitivity (p = 0.006). De la même manière, we replicated our NBS results by using the mean correla-
tion as an additional covariate of noninterest. Specifically, we found both statistically equiva-
lent (same uncorrected initial threshold, different number of links in network; FWE corrected
p < 0.001, 220 links, ICC > 0.75) and density-equivalent (similar number of links in network,
but more strict initial threshold p < 0.0001; FWE corrected p < 0.001, 87 links, ICC > 0.72)
cerebellar-cortical subnetworks that were significantly associated with short-term motor learn-
ing ability and were modulated by ketamine (F(1,11) = 6.97, p = 0.023 for the subnetwork with
220 links; F(1,11) = 4.93, p = 0.048 for the subnetwork with 87 links). Age, sex, order, BMI,
mean FD, and mean connectivity were controlled for as covariates of noninterests. De plus,
a marginal significant negative correlation was found between the mean connection of the sta-
tistically equivalent subnetwork and the Norketamine concentration (r = −0.51, p = 0.060),
whereas the mean connection of the density-equivalent subnetwork was significantly corre-
lated with the Norketamine concentration (r = −0.56, p = 0.037).
Neurosciences en réseau
473
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
un
_
0
0
0
4
5
p
d
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain functional connectome and skill learning
Scrubbing. Using a more stringent motion correction approach, we could replicate our pre-
vious findings that motor learning is positively correlated with small-worldness (r = 0.39,
p = 0.003) and global efficiency (r = 0.41, p = 0.002) and negatively correlated with transi-
tivité (r = −0.39, p = 0.003; Table S2, Zang et al., 2018). Cependant, motor learning was no
longer correlated with characteristic path length (r = −0.24, p = 0.074; Table S2, Zang et al.,
2018).
En outre, we could replicate our finding of a cerebellar-cortical subnetwork that was
correlated with learning rate. In detail, 81 connections linking bilateral cerebellum to visual,
sensorimotor, pariétal, and frontal areas were positively correlated with skill increase (uncor-
rected initial p < 5 × 10−4
, FWE corrected p = 0.036) while controlling age and sex as
covariates of noninterest (FD was no longer controlled as covariate of noninterest since we
used FD for scrubbing). This subnetwork also had a fair to good reliability (ICC > 0.73). Nous
then extracted the mean connectivity of the 81 connections and found a significant main ef-
fect of drug (F(1,13) = 5.10, p = 0.042) controlling for age, sex, order, and BMI as covariates
of noninterest. The mean connectivity of the 81 connections was also significantly negatively
correlated with Norketamine concentrations (r = −0.55, p = 0.03) when age, sex, and BMI
were controlled as covariates of noninterest.
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
un
_
0
0
0
4
5
p
d
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
To further explore the robustness of our NBS finding,
Influence of initial threshold definition.
we repeated our skill learning association analysis by using two different initial thresholds for
cluster definition, c'est, p < 0.001 (less strict) and p < 0.0001 (more strict). Notably, a less
strict initial p value should result in a larger but more unspecific network, whereas a stricter
initial p value should provide a more specific, but smaller network. As expected, using an
initial threshold of p < 0.0001, we found a similar but smaller network of 38 links including
the left cerebellum and cortical areas that were significantly correlated with skill learning (FWE
pcorr = 0.012). Moreover, the p < 0.001 initial threshold resulted in a larger, cerebellar-
cortical network consisting of 134 links that were significantly correlated with skill learning
(FWE pcorr = 0.048). We conclude from these observations that the reported association of the
cerebellar-cortical network with skill learning is observed across a range of initial t threshold
definitions for NBS.
To further probe our AAL-based findings for potential effects
Influence of parcellation choice.
of parcellation choice, we repeated our analysis with a recently published functional parcella-
tion atlas including 268 nodes (Rosenberg et al., 2016). Notably, the choice of this particular
functional atlas was motivated by the fact that it contains a comparable number of node defi-
nitions and covers the cerebellum in adequate detail, which is an important prerequisite given
that we employed a motor learning paradigm challenging subcortical and cerebellar structures.
All other data processing and analysis procedures were kept identical to our initial AAL-based
analysis. Similar to our AAL-based analysis, we detected a significant positive correlation of
skill increase with small-worldness (r = 0.28, p = 0.031) and global efficiency (r = 0.36,
p = 0.005) and a significant negative correlation of skill increase with transitivity (r = −0.38,
p = 0.002).
In addition, the NBS analysis with the Rosenberg atlas resulted in a very sim-
ilar but interestingly less reliable (ICC2,1 = 0.63, ICC3,1 = 0.64) cerebellar-cortical network
with 69 links that showed a significant positive association with skill learning (FWE corrected
p = 0.044, Figure S1, Zang et al., 2018). Moreover, comparable to our AAL-based findings, the
mean connectivity of this network was significantly negatively correlated with Norketamine
concentrations (r = −0.62, p = 0.014). We conclude from these observations that (a) our
AAL-based NBS findings do not relate to the choice of this particular parcellation scheme, and
Network Neuroscience
474
Brain functional connectome and skill learning
(b) that the choice of a functional parcellation atlas does not necessarily improve the reliability
of the examined connectivity estimates.
DISCUSSION
In the current resting-state fMRI study, we found several global brain network features to be
significantly associated with individual motor learning ability. Furthermore, using a well-
established motor learning task (Reis et al., 2009), we identified a cerebellar-cortical functional
subnetwork that was (a) significantly associated with short-term learning ability and (b) signif-
icantly modulated by NMDA receptor antagonism. We discuss our findings in more detail in
the following paragraphs.
First, we demonstrate that short-term motor learning ability is associated with several global
network features that characterize a network’s capability to process information efficiently
(Bullmore & Sporns, 2009; Rubinov & Sporns, 2010). Specifically, global efficiency and small-
worldness were positively correlated with motor learning ability, whereas transitivity and char-
acteristic path length were negatively correlated. Although these learning-associated global
network features are highly correlated with each other, they converge on the idea that higher
network integration may favor better short-term motor learning ability. This notion is in line
with previous studies demonstrating that higher network integration is beneficial for a range of
brain functions, including intelligence (van den Heuvel, Stam, Kahn, & Hulshoff Pol, 2009) and
working memory (Alavash, Doebler, Holling, Thiel, & Giessing, 2015). Importantly, whereas
previous studies have suggested that motor learning changes resting-state connectivity patterns
in terms of local network measures (Sami et al., 2014; Zhang et al., 2012), global resting-state
network characteristics of the brain have been shown to be relatively stable (Braun et al., 2012;
Cao et al., 2014) and untouched by the effects of motor learning (Heitger et al., 2012; Sami
& Miall, 2013). Taken together, this may indicate that those global features of brain networks
rather reflect the brain’s general capability to master a task independent of training-induced
alterations.
Second, we identified a highly plausible cerebellum-centered network with links between
cerebellar, visuospatial, sensorimotor, frontal, and temporal regions that were positively asso-
ciated with an individual’s learning ability. We further provided evidence suggesting that the
associated subnetwork is relatively reliable and robust against a variety of potential influencing
factors including local gray matter volume, age, sex, head motion, individual mean functional
connectivity differences, and the choice of the initial cluster-forming significance threshold
and parcellation scheme, respectively.
Notably, the identified subnetwork is highly plausible since it connects several key areas
involved in the early phase of visuomotor learning, including M1, SMA, premotor cortex, V5,
parietal cortex, and cerebellum (Bassett et al., 2011; Doyon et al., 2002; Hikosaka et al., 2002;
Zhang et al., 2012). Among these regions, M1, SMA, premotor, and visual cortex in particular
have been related to the computational integration of spatial motor demands (Hikosaka et al.,
2002) and the handling of on-line visual feedback (Dong et al., 2012) during the acquisition of
complex motor skills. Both functions are crucially important in the early learning phase of our
complex motor learning paradigm which requires constant visually guided feedback control
and real-time adjustments of executed motor programs. In addition, several parietal regions
participated in the cerebellum-centered network, an observation that is in line with the sug-
gested role of these regions in motor imagery learning (Zhang et al., 2012), a key element for
planning the upcoming movements’ kinetic parameters (Kuang, Morel, & Gail, 2016). More-
over, the involvement of bilateral DLPFC is consistent with previous motor learning studies
Network Neuroscience
475
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
a
_
0
0
0
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain functional connectome and skill learning
(Bassett, Yang, Wymbs, & Grafton, 2015; Heitger et al., 2012) and may plausibly relate to the
high level of visual attention demands (Barbey et al., 2013) and complex sequential memory
input in motor learning tasks (Toni and Passingham, 1999), especially during the early learning
phase (Bassett et al., 2015). Furthermore, the central role of the cerebellum in our identified
subcircuit is in good agreement with prior PET and fMRI studies. These studies demonstrated a
crucial role of the cerebellum as an error detector and parameter modifier of motor reference
plans in early learning phases (Doyon et al., 2002; Penhune & Steele, 2012). This has been
evidenced, for example, by severe impairments in certain aspects of motor learning (e.g., reac-
tion time) due to lack of behavioral adjustment in the face of errors in patients with cerebellar
lesions (Laforce & Doyon, 2001; Smith & Shadmehr, 2005). Although the observed association
between connectivity of the cerebellum-centered subnetwork and motor learning ability could
be interpreted as a stronger intrinsic capability of the network architecture in superior learn-
ers, it could also be argued that the association is a consequence of learning-induced motor
memory consolidation (Albert et al., 2009; Sami et al., 2014) since the resting-state scan was
acquired posttraining.
Third, consistent with prior system-level ketamine studies in humans (Kraguljac et al., 2016;
Niesters et al., 2012), we found that the cerebellum-centered network was significantly
modulated by NMDA receptor antagonism and its connectivity was negatively correlated
with blood-level Norketamine concentrations. Interestingly, the motor learning performance
before the scan itself was not affected (Francois et al., 2016; van Loon et al., 2016) by low-
dose ketamine infusion. Similar observations were made in object-recognition and reward-
anticipation fMRI studies, in which authors showed significantly altered BOLD responses but
no main effect of drug under low-dose (e.g., ≤0.5 mg/kg) ketamine administration during task
performance. This might indicate that the administered drug dose was sufficient to alter neural
functional interactions in the identified cerebellum-centered subnetwork, but below the dose
In addition,
level at which overt interruptions of motor learning behavior become evident.
the absence of behavioral differences between the drug conditions suggests that the observed
connectivity differences are unlikely to be the consequence of drug-induced changes in mo-
tor performance. The detected changes in cerebellar-cortical network connectivity suggest a
role for NMDA receptor-dependent glutamatergic neurotransmission that may relate to con-
solidation processes. This interpretation is consistent with previous reports of a strong depen-
dence of memory consolidation processes (Volianskis et al., 2015), BDNF genotype (Gosselin
et al., 2016), and plasticity-related protein synthesis in the motor cortex (Luft, Buitrago, Ringer,
Dichgans, & Schulz, 2004). Notably, the fact that we found no modulation of global network
measures by ketamine further supports our earlier interpretation of these whole-brain efficiency
markers as traitlike reflections of the brain’s capability to perform a range of different tasks.
Our study has several limitations worth mentioning. Most importantly, although our finding
of learning-related subnetwork connectivity indices is in line with the hypothesis that motor
training leads to temporary changes in the functional brain network architecture, the direc-
tionality of such an effect cannot be claimed with our cross-sectional data. Even though we
acquired resting-state data after off-line motor learning, the interpretation of a predisposed suit-
ability of intrinsic brain networks for the challenged motor performance is equally plausible
(Mary et al., 2016). Second, although the connectivity within the motor learning–associated
subnetwork was significantly decreased under NMDA receptor blockade, the interpretation
of impaired motor memory consolidation would ultimately require an affected motor perfor-
mance in a preceding motor task. However, as we did not reassess the motor performance af-
ter scanning, we must defer such a proof to future studies. Third, previous resting-state studies
(Albert, Robertson, & Miall, 2009; Barnes, Bullmore, & Suckling, 2009) provided evidence for
Network Neuroscience
476
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
a
_
0
0
0
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain functional connectome and skill learning
an impact of motor and cognitive tasks on the functional configuration of resting-state networks
in subsequent MRI scans. This implies that in the case of drug-dependent differences in task
engagement prior to the scan, variant carry-over effects (instead of or in addition to an NMDA
receptor-related neural plasticity mechanism) may have influenced our drug challenge results.
Notably, we did not detect significant main effects of drug condition on behavioral markers
of training performance, which argues against such an interpretation. We nonetheless cannot
fully exclude that other drug-induced differences in task engagement may have existed and
have been carried over to the following resting-state scan. Fourth, although ketamine mod-
ulated our specific cerebellum-centered subnetwork, ketamine as a noncompetitive NMDA
receptor antagonist may also plausibly influence other brain subnetworks.
In conclusion, we demonstrate that global brain network characteristics and specific subnet-
work connectivity patterns during resting state are associated with motor learning before scan-
ning. We further show that the identified learning-related subnetwork connectivity estimates
are unrelated to the gray matter volume of the nodes, reliable, and susceptible to glutamate
challenge. We posit that the observed differential modulation of the examined whole-brain
graph theoretical versus cerebellar-cortical network features by ketamine may reflect distinct
qualities of learning-related brain function, for example, individual predisposition for learning
new motor skills (global brain network measures) versus glutamate-dependent processes re-
lated to active motor memory consolidation (cerebellar-cortical network connectivity). Taken
together, this investigation may offer valuable information on the neural processes related to
short-term motor learning in humans and provide a starting point for future studies in a still
under-researched area of human neuroscience.
ACKNOWLEDGMENTS
We thank Ilka Alexi, Carolin Dennewill, Tobias Gradinger, Jascha Thiem, Canan Koc, Oliver
Grimm, Leila Haddad, and Mathias Kienow for research assistance.
FUNDING INFORMATION
This study was supported by the German Federal Ministry of Education and Research
(BMBF, grant 01GQ1102 to H.T.). A.M.-L. acknowledges grant support by the European
Community’s Seventh Framework Programme under the grant agreement No. 115008 (Project
EU-NEWMEDS).
AUTHOR CONTRIBUTIONS
Heike Tost, Andreas Meyer-Lindenberg, and Janine Reis: Design of the study. Zhenxiang
Zang, Heike Tost, Lena Geiger, and Urs Braun: Writing. Zhenxiang Zang, Urs Braun, Axel
Schäfer, and Emanuel Schwarz: Data analysis. Zhenxiang Zang, Lena Geiger, Maria Zangl, and
Janina I. Schweiger: Data collection. Axel Schäfer, Matthias Ruf, and Janine Reis: Provided of
experiment materials. Hengyi Cao, Carolin Moessang, Luanna Dixson, Alexander Moscicki,
and Andreas Meyer-Lindenberg: Manuscript revision.
CONFLICT OF INTEREST
A.M.-L. has received consultant fees and travel expenses from Alexza Pharmaceuticals,
AstraZeneca, Bristol-Myers Squibb, Defined Health, Decision Resources, Desitin Arzneimittel,
Elsevier, F. Hoffmann-La Roche, Gerson Lehrman Group, Grupo Ferrer, Les Laboratoires
Servier, Lilly Deutschland, Lundbeck Foundation, Outcome Sciences, Outcome Europe,
Network Neuroscience
477
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
a
_
0
0
0
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain functional connectome and skill learning
PriceSpective; and Roche Pharma and has received speaker’s fees from Abbott, AstraZeneca,
BASF, Bristol-Myers Squibb, GlaxoSmithKline, Janssen-Cilag, Lundbeck, Pfizer Pharma, and
Servier Deutschland. The other authors report no biomedical financial interests or other
potential conflicts of interest.
REFERENCES
Alavash, M., Doebler, P., Holling, H., Thiel, C. M., & Giessing, C.
(2015). Is functional integration of resting state brain networks
an unspecific biomarker for working memory performance?
NeuroImage, 108, 182–193. https://doi.org/10.1016/j.neuroimage.
2014.12.046
Albert, N. B., Robertson, E. M., & Miall, R. C. (2009). The rest-
ing human brain and motor learning. Current Biology, 19(12),
1023–1027. https://doi.org/10.1016/j.cub.2009.04.028
Barbey, A. K., Koenigs, M., & Grafman, J. (2013). Dorsolateral pre-
frontal contributions to human working memory. Cortex, 49(5),
1195–1205. https://doi.org/10.1016/j.cortex.2012.05.022
Barnes, A., Bullmore, E. T., & Suckling, J. (2009). Endogenous hu-
man brain dynamics recover slowly following cognitive effort.
PLoS One, 4(8), e6626. https://doi.org/10.1371/journal.pone.
0006626
Bassett, D. S., & Mattar, M. G. (2017). A Network Neuroscience
of Human Learning: Potential to Inform Quantitative Theories
of Brain and Behavior. Trends in Cognitive Sciences, 21(4),
250–264. https://doi.org/10.1016/j.tics.2017.01.010
Bassett, D. S., Wymbs, N. F., Porter, M. A., Mucha, P. J., Carlson,
J. M., & Grafton, S. T. (2011). Dynamic reconfiguration of hu-
man brain networks during learning. Proceedings of the National
Academy of Sciences of the United States of America, 108(18),
7641–7646. https://doi.org/10.1073/pnas.1018985108
Bassett, D. S., Yang, M., Wymbs, N. F., & Grafton, S. T. (2015).
Learning-induced autonomy of sensorimotor systems. Nature
Neuroscience, 18(5), 744–751. https://doi.org/10.1038/nn.3993
Braun, U., Plichta, M. M., Esslinger, C., Sauer, C., Haddad, L.,
Grimm, O., . . . Meyer-Lindenberg, A. (2012). Test-retest reli-
ability of
resting-state connectivity network characteristics
using fMRI and graph theoretical measures. NeuroImage, 59(2),
1404–1412. https://doi.org/10.1016/j.neuroimage.2011.08.044
(2009). Complex brain networks:
graph theoretical analysis of structural and functional systems.
Nature Reviews Neuroscience, 10(3), 186–198. https://doi.org/
10.1038/nrn2575
Bullmore, E., & Sporns, O.
Cao, H., Plichta, M. M., Schafer, A., Haddad, L., Grimm, O.,
Schneider, M., . . . Tost, H. (2014). Test-retest reliability of fMRI-
based graph theoretical properties during working memory, emo-
tion processing, and resting state. NeuroImage, 84, 888–900.
https://doi.org/10.1016/j.neuroimage.2013.09.013
Dayan, E., & Cohen, L. G. (2011). Neuroplasticity subserving motor
skill learning. Neuron, 72(3), 443–454. https://doi.org/10.1016/
j.neuron.2011.10.008
Dong, Z. Y., Liu, D. Q., Wang, J., Qing, Z., Zang, Z. X., Yan, C. G.,
& Zang, Y. F. (2012). Low-frequency fluctuation in continuous
real-time feedback of finger force: a new paradigm for sustained
attention. Neuroscience Bulletin, 28(4), 456–467. https://doi.org/
10.1007/s12264-012-1254-2
Doyon, J., & Benali, H. (2005). Reorganization and plasticity in the
adult brain during learning of motor skills. Current Opinion in
Neurology, 15(2), 161–167. https://doi.org/10.1016/j.conb.2005.
03.004
Doyon, J., Song, A. W., Karni, A., Lalonde, F., Adams, M. M., &
Ungerleider, L. G. (2002). Experience-dependent changes in cere-
bellar contributions to motor sequence learning. Proceedings of the
National Academy of Sciences of the United States of America,
99(2), 1017–1022. https://doi.org/10.1073/pnas.022615199
Fleiss, J. L. (1986). The Design and Analysis of Clinical Experiments.
New York: Wiley.
Francois,
.
.
. Meyer-Lindenberg, A.
J., Grimm, O., Schwarz, A. J., Schweiger,
J., Haller,
L., Risterucci, C.,
(2016). Keta-
mine Suppresses the Ventral Striatal Response to Reward An-
ticipation: A Cross-Species Translational Neuroimaging Study.
Neuropsychopharmacology, 41(5), 1386–1394. https://doi.org/
10.1038/npp.2015.291
Fritsch, B., Reis,
J., Martinowich, K., Schambra, H. M.,
Ji, Y.,
Cohen, L. G., & Lu, B. (2010). Direct current stimulation pro-
motes BDNF-dependent synaptic plasticity: potential implica-
tions for motor learning. Neuron, 66(2), 198–204. https://doi.org/
10.1016/j.neuron.2010.03.035
Gosselin, N., De Beaumont, L., Gagnon, K., Baril, A. A., Mongrain,
V., Blais, H., . . . Carrier, J. (2016). BDNF Val66Met Polymorphism
Interacts with Sleep Consolidation to Predict Ability to Create
New Declarative Memories. Journal of Neuroscience, 36(32),
8390–8398. https://doi.org/10.1523/jneurosci.4432-15.2016
Grimm, O., Gass, N., Weber-Fahr, W., Sartorius, A., Schenker, E.,
Spedding, M., . . . Meyer-Lindenberg, A. (2015). Acute ketamine
challenge increases resting state prefrontal-hippocampal connec-
tivity in both humans and rats. Psychopharmacology (Berl), 232(21-22),
4231–4241. https://doi.org/10.1007/s00213-015-4022-y
Hadj, T. A., Blanchet, P. J., & Doyon, J. (2004). Motor-learning
impairment by amantadine in healthy volunteers. Neuropsycho-
pharmacology, 29(1), 187–194. https://doi.org/10.1038/sj.npp.
1300317
Hardwick, R. M., Rottschy, C., Miall, R. C., & Eickhoff, S. B. (2013).
A quantitative meta-analysis and review of motor learning in
the human brain. NeuroImage, 67, 283–297. https://doi.org/10.
1016/j.neuroimage.2012.11.020
Heitger, M. H., Ronsse, R., Dhollander, T., Dupont, P.,
Caeyenberghs, K., & Swinnen, S. P. (2012). Motor learning-
induced changes in functional brain connectivity as revealed by
means of graph-theoretical network analysis. NeuroImage, 61(3),
633–650. https://doi.org/10.1016/j.neuroimage.2012.03.067
Hikosaka, O., Nakamura, K., Sakai, K., & Nakahara, H. (2002).
Central mechanisms of motor skill learning. Current Opinion in
Neurology, 12(2), 217–222.
Network Neuroscience
478
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
a
_
0
0
0
4
5
p
d
t
.
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain functional connectome and skill learning
Hochberg, Y. (1988). A sharper Bonferroni procedure for multiple
tests of significance. Biometrika, 75, 800–802.
Kent, K., Deng, Q., & McNeill, T. H. (2013). Unilateral skill acquisi-
tion induces bilateral NMDA receptor subunit composition shifts
in the rat sensorimotor striatum. Brain Research, 1517, 77–86.
https://doi.org/10.1016/j.brainres.2013.04.021
Kida, H., Tsuda, Y., Ito, N., Yamamoto, Y., Owada, Y., Kamiya, Y., &
Mitsushima, D. (2016). Motor Training Promotes Both Synaptic
and Intrinsic Plasticity of Layer II/III Pyramidal Neurons in the
Primary Motor Cortex. Cerebral Cortex, 26(8), 3494–3507.
https://doi.org/10.1093/cercor/bhw134
Kraguljac, N. V., Frolich, M. A., Tran, S., White, D. M., Nichols, N.,
(2016). Ketamine modu-
Barton-McArdle, A., . . . Lahti, A. C.
lates hippocampal neurochemistry and functional connectivity:
a combined magnetic resonance spectroscopy and resting-state
fMRI study in healthy volunteers. Molecular Psychiatry. https://
doi.org/10.1038/mp.2016.122
Kuang, S., Morel, P., & Gail, A. (2016). Planning Movements in
Visual and Physical Space in Monkey Posterior Parietal Cortex.
Cerebral Cortex, 26(2), 731–747. https://doi.org/10.1093/cercor/
bhu312
Laforce, R., Jr., & Doyon, J. (2001). Distinct contribution of the
striatum and cerebellum to motor learning. Brain and Cognition,
45(2), 189–211. https://doi.org/10.1006/brcg.2000.1237
Luft, A. R., Buitrago, M. M., Ringer, T., Dichgans, J., & Schulz,
J. B. (2004). Motor skill learning depends on protein synthe-
Journal of Neuroscience,
sis in motor cortex after training.
24(29), 6515–6520. https://doi.org/10.1523/JNEUROSCI.1034-
04.2004
Mary, A., Wens, V., Op de Beeck, M., Leproult, R., De Tiege, X., &
Peigneux, P. (2016). Resting-state Functional Connectivity is an
Age-dependent Predictor of Motor Learning Abilities. Cerebral
Cortex. https://doi.org/10.1093/cercor/bhw286
Mawase, F., Bar-Haim, S., & Shmuelof, L.
(2017). Formation
of Long-Term Locomotor Memories Is Associated with Func-
tional Connectivity Changes in the Cerebellar–Thalamic–Cortical
Network. Journal of Neuroscience, 37(2), 349–361.
McHughen, S. A., Rodriguez, P. F., Kleim, J. A., Kleim, E. D.,
Marchal Crespo, L., Procaccio, V., & Cramer, S. C. (2010). BDNF
val66met polymorphism influences motor system function in the
human brain. Cerebral Cortex, 20(5), 1254–1262. https://doi.org/
10.1093/cercor/bhp189
Mueller, S., Wang, D., Fox, M. D., Pan, R., Lu, J., Li, K., . . .
Liu, H. (2015). Reliability correction for functional connectivity:
Theory and implementation. Human Brain Mapping, 36(11),
4664–4680. https://doi.org/10.1002/hbm.22947
Newman, M. E.
(2006). Modularity and community structure in
networks. Proceedings of the National Academy of Sciences of
the United States of America, 103(23), 8577–8582. https://doi.
org/10.1073/pnas.0601602103
Niesters, M., Khalili-Mahani, N., Martini, C., Aarts, L., van
Gerven, J., van Buchem, M. A., . . . Rombouts, S. (2012). Effect of
subanesthetic ketamine on intrinsic functional brain connectiv-
ity: a placebo-controlled functional magnetic resonance imag-
ing study in healthy male volunteers. Anesthesiology, 117(4),
868–877. https://doi.org/10.1097/ALN.0b013e31826a0db3
Pekhletski, R., Gerlai, R., Overstreet, L. S., Huang, X. P., Agopyan,
N., Slater, N. T., . . . Hampson, D. R. (1996). Impaired cerebellar
synaptic plasticity and motor performance in mice lacking the
mGluR4 subtype of metabotropic glutamate receptor. Journal of
Neuroscience, 16(20), 6364–6373.
Penhune, V. B., & Steele, C. J. (2012). Parallel contributions of cere-
bellar, striatal and M1 mechanisms to motor sequence learning.
Behav Brain Research, 226(2), 579–591. https://doi.org10.1016/
j.bbr.2011.09.044
Power, J. D., Barnes, K. A., Snyder, A. Z., Schlaggar, B. L., &
Petersen, S. E.
(2012). Spurious but systematic correlations
in functional connectivity MRI networks arise from subject mo-
tion. NeuroImage, 59(3), 2142–2154. https://doi.org/10.1016/
j.neuroimage.2011.10.018
Reis, J., Schambra, H. M., Cohen, L. G., Buch, E. R., Fritsch, B.,
Zarahn, E., . . . Krakauer, J. W. (2009). Noninvasive cortical
stimulation enhances motor skill acquisition over multiple days
through an effect on consolidation. Proceedings of the National
Academy of Sciences of the United States of America, 106(5),
1590–1595. https://doi.org/10.1073/pnas.0805413106
Rosenberg, M. D., Finn, E. S., Scheinost, D., Papademetris, X.,
Shen, X., Constable, R. T., & Chun, M. M. (2016). A neuro-
marker of sustained attention from whole-brain functional
connectivity. Nature Neuroscience, 19(1), 165–171. https://doi.
org/10.1038/nn.4179
Rubinov, M., & Sporns, O. (2010). Complex network measures of
brain connectivity: uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003
Sami, S., & Miall, R. C. (2013). Graph network analysis of immedi-
ate motor-learning induced changes in resting state BOLD. Fron-
tiers in Human Neuroscience, 7, 166. https://doi.org/10.3389/
fnhum.2013.00166
Sami, S., Robertson, E. M., & Miall, R. C. (2014). The time course
of task-specific memory consolidation effects in resting state net-
works. Journal of Neuroscience, 34(11), 3982–3992. https://doi.
org/10.1523/JNEUROSCI.4341-13.2014
J., Calkins, M. E.,
Satterthwaite, T. D., Elliott, M. A., Gerraty, R. T., Ruparel, K.,
Loughead,
. . . Wolf, D. H. (2013). An
improved framework for confound regression and filtering for
control of motion artifact in the preprocessing of resting-state
functional connectivity data. NeuroImage, 64, 240–256. https://
doi.org/10.1016/j.neuroimage.2012.08.052
Smith, M. A., & Shadmehr, R. (2005). Intact ability to learn internal
models of arm dynamics in Huntington’s disease but not cerebel-
lar degeneration. Journal of Neurophysiology, 93(5), 2809–2821.
https://doi.org/10.1152/jn.00943.2004
Thomason, M. E., Yoo, D. J., Glover, G. H., & Gotlib, I. H. (2009).
BDNF genotype modulates resting functional connectivity in
children. Frontiers in Human Neuroscience, 3, 55. https://doi.org/
10.3389/neuro.09.055.2009
Toni, I., & Passingham, R. E. (1999). Prefrontal-basal ganglia path-
ways are involved in the learning of arbitrary visuomotor associa-
tions: A PET study. Experimental Brain Research, 127(1), 19–32.
Tzourio-Mazoyer, N., Landeau, B., Papathanassiou, D., Crivello,
F., Etard, O., Delcroix, N., . . . Joliot, M. (2002). Automated
anatomical labeling of activations in SPM using a macroscopic
anatomical parcellation of the MNI MRI single-subject brain.
Network Neuroscience
479
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
a
_
0
0
0
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Brain functional connectome and skill learning
NeuroImage, 15(1), 273–289. https://doi.org/10.1006/nimg.2001.
0978
van den Heuvel, M. P., de Lange, S. C., Zalesky, A., Seguin, C.,
Yeo, B. T. T., & Schmidt, R. (2017). Proportional thresholding
in resting-state fMRI functional connectivity networks and con-
sequences for patient-control connectome studies:
Issues and
recommendations. NeuroImage, 152, 437–449. https://doi.org/
10.1016/j.neuroimage.2017.02.005
van den Heuvel, M. P., Stam, C. J., Kahn, R. S., & Hulshoff Pol, H. E.
(2009). Efficiency of functional brain networks and intellectual
performance. Journal of Neuroscience, 29(23), 7619–7624. https://
doi.org/10.1523/jneurosci.1443-09.2009
van Loon, A. M., Fahrenfort, J. J., van der Velde, B., Lirk, P. B.,
Vulink, N. C., Hollmann, M. W., . . . Lamme, V. A. (2016). NMDA
Receptor Antagonist Ketamine Distorts Object Recognition by
Reducing Feedback to Early Visual Cortex. Cerebral Cortex,
26(5), 1986–1996. https://doi.org/10.1093/cercor/bhv018
Volianskis, A., France, G., Jensen, M. S., Bortolotto, Z. A., Jane,
D. E., & Collingridge, G. L.
(2015). Long-term potentiation
and the role of N-methyl-D-aspartate receptors. Brain Research,
1621, 5–16. https://doi.org/10.1016/j.brainres.2015.01.016
Wang, J., Zuo, X., Dai, Z., Xia, M., Zhao, Z., Zhao, X., . . . He,
Y.
(2013). Disrupted functional brain connectome in individ-
uals at risk for Alzheimer’s disease. Biological Psychiatry, 73(5),
472–481. https://doi.org/10.1016/j.biopsych.2012.03.026
Wang, J., Zuo, X., & He, Y. (2010). Graph-based network analysis of
resting-state functional MRI. Frontiers in Systems Neuroscience,
4, 16. https://doi.org/10.3389/fnsys.2010.00016
Wu, J., Srinivasan, R., Kaur, A., & Cramer, S. C. (2014). Resting-state
cortical connectivity predicts motor skill acquisition. NeuroImage,
91, 84–90. https://doi.org/10.1016/j.neuroimage.2014.01.026
Xia, M., Wang, J., & He, Y. (2013). BrainNet Viewer: a network vi-
sualization tool for human brain connectomics. PLoS One, 8(7),
e68910. https://doi.org/10.1371/journal.pone.0068910
Yan, C., & Zang, Y. (2010). DPARSF: A MATLAB Toolbox for
“Pipeline” Data Analysis of Resting-State fMRI. Frontiers in
Systems Neuroscience, 4, 13. https://doi.org/10.3389/fnsys.2010.
00013
Yan, C. G., Cheung, B., Kelly, C., Colcombe, S., Craddock, R. C.,
Di Martino, A., . . . Milham, M. P. (2013). A comprehensive as-
sessment of regional variation in the impact of head micromove-
ments on functional connectomics. NeuroImage, 76, 183–201.
https://doi.org/10.1016/j.neuroimage.2013.03.004
Zalesky, A., Fornito, A., & Bullmore, E. T. (2010). Network-based
statistic: identifying differences in brain networks. NeuroImage,
53(4), 1197–1207. https://doi.org/10.1016/j.neuroimage.2010.06.
041
Zalesky, A., Fornito, A., Harding,
I. H., Cocchi, L., Yucel,
M., Pantelis, C., & Bullmore, E. T. (2010). Whole-brain anatom-
ical networks: does the choice of nodes matter? NeuroImage,
50(3), 970–983. https://doi.org/10.1016/j.neuroimage.2009.12.
027
Zang, Z., Geiger, L. S., Braun, U., Cao, H., Zangl, M., Schäfer, A.,
. . . & Tost, H. (2018). Supporting Information for “Resting-state
brain network features associated with short-term skill learn-
ing ability in humans and the influence of N-mythyl-D-aspertate
receptor antagonism.” Network Neuroscience, 2(4), 464–480.
https://doi.org/10.1162/netn_a_00045
Zhang, H., Xu, L., Zhang, R., Hui, M., Long, Z., Zhao, X., & Yao, L.
(2012). Parallel alterations of functional connectivity during exe-
cution and imagination after motor imagery learning. PLoS One,
7(5), e36052. https://doi.org/10.1371/journal.pone.0036052
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
/
2
4
4
6
4
1
0
9
2
2
3
1
n
e
n
_
a
_
0
0
0
4
5
p
d
.
t
f
b
y
g
u
e
s
t
t
o
n
0
8
S
e
p
e
m
b
e
r
2
0
2
3
Network Neuroscience
480