RECHERCHE
Optimizing network neuroscience computation of
individual differences in human spontaneous
brain activity for test-retest reliability
Chao Jiang1, Ye He2, Richard F. Betzel3, Yin-Shan Wang4,5, Xiu-Xia Xing6, and Xi-Nian Zuo4,5,7,8
1School of Psychology, Capital Normal University, Beijing, Chine
2School of Artificial Intelligence, Beijing University of Posts and Telecommunications, Beijing, Chine
3Department of Psychological and Brain Sciences, Indiana University, Bloomington, Indiana, Etats-Unis
4State Key Laboratory of Cognitive Neuroscience and Learning, Beijing Normal University, Beijing, Chine
5Developmental Population Neuroscience Research Center, International Data Group/ McGovern
Institute for Brain Research, Beijing Normal University, Beijing, Chine
6Department of Applied Mathematics, College of Mathematics, Faculty of Science,
Beijing University of Technology, Beijing, Chine
7National Basic Science Data Center, Beijing, Chine
8Institute of Psychology, Chinese Academy of Sciences, Beijing, Chine
Mots clés: Individual difference, Reliability, Open science, Spontaneous brain activity, Connectome
ABSTRAIT
A rapidly emerging application of network neuroscience in neuroimaging studies has provided
useful tools to understand individual differences in intrinsic brain function by mapping
spontaneous brain activity, namely intrinsic functional network neuroscience (ifNN). Cependant,
the variability of methodologies applied across the ifNN studies—with respect to node
definition, edge construction, and graph measurements—makes it difficult to directly compare
findings and also challenging for end users to select the optimal strategies for mapping
individual differences in brain networks. Ici, we aim to provide a benchmark for best ifNN
practices by systematically comparing the measurement reliability of individual differences
under different ifNN analytical strategies using the test-retest design of the Human Connectome
Project. The results uncovered four essential principles to guide ifNN studies: (1) use a
whole brain parcellation to define network nodes, including subcortical and cerebellar regions;
(2) construct functional networks using spontaneous brain activity in multiple slow bands;
et (3) optimize topological economy of networks at individual level; et (4) characterize
information flow with specific metrics of integration and segregation. We built an interactive
online resource of reliability assessments for future ifNN (https://ibraindata.com/research/ifNN).
RÉSUMÉ DE L'AUTEUR
It is an essential mission for neuroscience to understand the individual differences in brain
fonction. Graph or network theory offer novel methods of network neuroscience to address
such a challenge. This article documents optimal strategies on the test-retest reliability of
measuring individual differences in intrinsic brain networks of spontaneous activity. Le
analytical pipelines are identified to optimize for highly reliable, individualized network
measurements. These pipelines optimize network metrics for high interindividual variances
and low inner-individual variances by defining network nodes with whole-brain parcellations,
deriving the connectivity with spontaneous high-frequency slow-band oscillations,
constructing brain graphs with topology-based methods for edge filtering, and favoring
un accès ouvert
journal
Citation: Jiang, C., Il, Y., Betzel, R.. F.,
Wang, Y.-S., Xing, X.-X., & Zuo, X.-N.
(2023). Optimizing network neuroscience
computation of individual differences
in human spontaneous brain activity
for test-retest reliability. Réseau
Neurosciences, 7(3), 1080–1108. https://
doi.org/10.1162/netn_a_00315
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00315
Informations complémentaires:
https://doi.org/10.1162/netn_a_00315
Reçu: 14 Août 2022
Accepté: 22 Mars 2023
Corresponding Authors:
Xi-Nian Zuo
xinian.zuo@bnu.edu.cn
Xiu-Xia Xing
xingxx@bjut.edu.cn
Éditeur de manipulation:
Alex Fornito
droits d'auteur: © 2023
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
multilevel or multimodal metrics. These psychometric findings are critical for translating the
functional network neuroscience into clinical or other personalized practices requiring
neuroimaging markers.
INTRODUCTION
Over the past two decades, network neuroscience has helped transform the field of neurosci-
ence (D. Bassett et al., 2020), providing a quantitative methodology framework for modeling
brains as graphs (or networks) composed of nodes (brain regions) and edges (their connec-
tion), namely connectomics (Sporns, 2013un). The organization and topology of macroscale
brain networks can be characterized by a growing suite of connectomic measurements includ-
ing efficiency, centrality, clustering, small-word topology, rich-club, and so forth (Craddock
et coll., 2013). In parallel, resting-state fMRI (rfMRI) has opened up new avenues toward under-
standing the intrinsic human brain function (Biswal et al., 2010). In conjunction with network
neuroscience, rfMRI has led to the emergence of a multidisciplinary field, intrinsic functional
connectomics or network neuroscience (ifNN), in which the brain’s intrinsic, interregional
connectivity is estimated from rfMRI recordings. It has been widely used to investigate the
system-level organization of the human brain function and its relationship with individual dif-
ferences (Dubois & Adolphs, 2016) in developmental (Zuo et al., 2017), sociocultural (Personne,
2018), and clinical conditions (Fornito, Zalesky, & Breakspear, 2015).
Highly reliable measurements are essential for studying individual differences. En général,
reliability characterizes a proportion of measurement variability between different subjects
relative to the overall variability including both between-subject and within-subject (c'est à dire.,
random) components (Xing & Zuo, 2018). It is commonly used to assess the consistency or
agreement between measurements, or the ability to obtain consistent measures over time.
Beyond that, it can also serve as a measure of discriminability (Xing & Zuo, 2018; Zuo, Biswal,
& Poldrack, 2019un; Zuo, Xu, & Milham, 2019b). Par exemple, if a measurement can more
sufficiently capture individual characteristics (c'est à dire., better differentiate a group of individuals),
it will produce higher between-subject variability and thus higher reliability than a measure-
ment underestimating the between-subject variability. Such reliability concept has well-
established statistical theory and applications in fields such as psychology (Elliott, Knodt,
Caspi, Moffitt, & Hariri, 2021un) and medicine (Kraemer, 2014) where it is used in psychometric
theory and diagnosis theory, respectivement. Spécifiquement, in psychology, reliability is important
for assessing the validity of psychological tests, and in medicine, it is important for accurately
diagnosing and treating patients. In the field of human brain mapping, more recent studies
have demonstrated that the measurement reliability is equivalent to the “fingerprint” or
discriminability of the measurement under the Gaussian distribution (Bridgeford et al.,
2021; Milham, Vogelstein, & Xu, 2021). Donc, the optimization of measurement reliability
of the individual differences can help guide ifNN processing and analysis pipelines for
individualized or personalized (par exemple., neurodevelopmental; Herting, Gautam, Chen, Mezher,
& Vetter, 2018) or clinical (Matthieu & Hampshire, 2016) recherche.
Previous studies have demonstrated that many functional network measurements with
rfMRI have limited reliability (Noble, Scheinost, & Constable, 2019; Zuo & Xing, 2014). These
low levels of reliability could be an indication of failure in handling individual variability at
different levels (Elliott, Knodt, & Hariri, 2021b; Hallquist & Hillary, 2019). En particulier, exper-
imental design and processing decisions related to scan duration, determining frequency
Brain topology:
A quantitative description on
structural and functional
relationships of the brain
organization and their links to mind.
Resting state:
A paradigm in neuroimaging
developed to approch the balance
state of brain as a complex system
from a perspective of dynamic
physics, commonly a state of lying-
down quietly without systematic or
specific thoughts.
Measurement reliability:
Proportion of between-subject
variability of a measurement relative
to its overall variability including
both between-subject and within-
subject components.
Neurosciences en réseau
1081
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
t
/
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
Parcellation:
A division into parcels defining
distinct partitions of neural elements
in the brain.
range, and regressing global signal have impacts on rfMRI measurements and thus their
reliability (Noble et al., 2019; Zuo et al., 2013). Although less focused on reliability, existing
network neuroscience studies revealed that their findings are influenced by choices of parcel-
lation templates (Bryce et al., 2021; Wang et al., 2009), edge construction and definition, et
choice of graph metrics (Liang et al., 2012). How these decisions affect the measurement reli-
ability in ifNN deserves further investigation. These analytical choices have been implemented
in different software packages but can vary from one package to another, and thus introduce
more analytic variability (Botvinik-Nezer et al., 2020). Beyond limited examinations on
reliability (Aurich, Filho, da Silva, & Franco, 2015; Braun et al., 2012; Termenon, Jaillard,
Delon-Martin, & Achard, 2016), a systematic investigation into the measurement reliability
is warranted to guide ifNN software use and analyses.
We conducted a systematic ifNN reliability analysis using the test-retest rfMRI data from
the Human Connectome Project (HCP). The HCP has developed its imaging acquisition and
data preprocessing (Glasser et al., 2013) by integrating various strategies optimized for reli-
ability in previous studies (Noble et al., 2019; Noble, Scheinost, & Constable, 2021; Zuo &
Xing, 2014; Zuo et al., 2013). We thus analyzed the minimally preprocessed HCP rfMRI
data and focused our work on four key postanalytic stages: node definition, edge construc-
tion, network measurement, and reliability assessments. À la fin, we propose a set of
principles to guide researchers in performing reliable ifNN, advancing the fieldstandard call
for the best practices in network neuroscience. We released all the codes and reliability data
by building an online platform for sharing the data and computational resources to foster
future ifNN.
THEORY AND METHODS
A typical analysis pipeline in ifNN includes steps for node definition (parcellations) and edge
construction (frequency bands, connectivity estimation and filtering schemes) (Figure 1A). À
determine an optimal pipeline, we combine the most reliable strategies across different parts
of the analysis by comparing the reliability of derived global network metrics. The HCP test-
retest data were employed for reliability evaluation (Figure 1B) using the intraclass correlation
(ICC) statistics on the measurement reliability. Overall reliability assessments associated with
the various analytic strategies as well as their impact on between- and within-subject variabil-
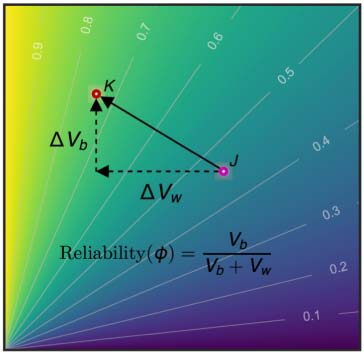
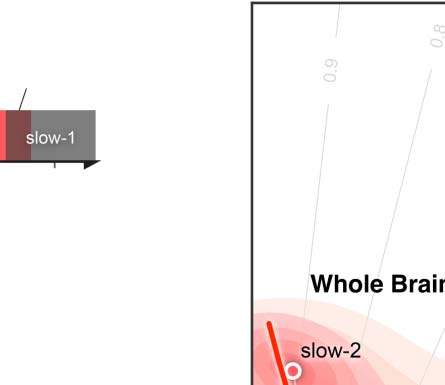
ville (Figure 1C) are investigated. We calculated the between-subject variability (Vb) and within-
subject variability (Vw) and normalized them to values between 0 et 1 by the total sample
variances. The changes in these variability measures, ΔVb and ΔVw, were used to create a
reliability gradient represented by a vector. The length of the arrow reflects the amplitude
of the change in reliability when comparing one choice (pink circle, J.) to another choice
(red circle, K ). The direction of the arrow, JK, indicates the sources of the change in reliability.
Dans ce cas, the reliability increases from a moderate to a substantial level with an increase in
between-subject variability (ΔVb > 0) and a decrease in within-subject variability (ΔVw < 0).
We then determine the optimized pipelines based on the highest reliability measurements,
while documenting the derived both global and local network metrics and both their reli-
ability and variability at an individual level. We first introduce the reliability theory and
assessments.
Reliability Theory and Assessments
Measurement reliability is defined as the extent to which measurements can be replicated
across multiple repeated measuring occasions (Xing & Zuo, 2018; Zuo, Biswal, & Poldrack,
Network Neuroscience
1082
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
Figure 1. Analytical pipelines for reliable ifNN. (A) There are five stages during our analyses: (1) test-retest dataset (white box) downloaded
from HCP website; (2) node definition (green box) defining nodes using a set of brain areas of 24 different partitions of the human brain; (3)
edge construction (yellow box) estimating individual correlation matrices using the six frequency bands (slow 1–6) from Buzsaki’s theoretical
framework on the brain oscillations as well as the widely used empirical frequency band (Slow-emp) and transferring these matrices into
adjacency matrices using 7 × 4 × 12 different strategies on edge construction including band-pass filtering, connectivity estimation and edge
filtering; (4) network analysis (blue box) systematically calculating various brain graph metrics on measurements of information flow; and (5)
reliability assessment (red box) evaluating test-retest reliability with massive linear mixed models. (B) The test-retest data shared multimodal
MRI datasets of 46 subjects in the HCP S1200 release and the HCP Retest release. Each subject underwent the first four test scans on 2 days
(two scans per day: Rest1 and Rest2) and return several months later to finish the four retest scans on another 2 days. (C) Measurement reli-
ability refers to the interindividual or between-subject variability Vb relative to the intraindividual or within-subject variability Vw. Variability of
both between-subject (Vb) and within-subject (Vw) are normalized into between 0 and 1 by the total sample variances. Their changes (ΔVb and
ΔVw) introduce a reliability gradient as represented by the vector (the black arrow). The length of the arrow reflects the amplitude of reliability
changes when the reliability assessment from one choice (pink circle, J ) to another choice (red circle, K ). Further, the arrow’s direction (JK)
indicates the sources of this reliability change. Here the reliability becomes from moderate to substantial level with increases of between-
subject variability (ΔVb > 0) and decreases of within-subject variability (ΔVw < 0).
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
.
t
/
2019a; Zuo, Xu, & Milham, 2019b). Test-retest reliability is the closeness of the agreement
between the results of successive measurements of the same measure and carried out under
the same conditions of measurement.
Linear mixed models. As a group-level statistic, reliability refers to the interindividual or
between-subject variability Vb relative to the intraindividual or within-subject variability Vw.
Both the intra- and inter-individual variances can be estimated using linear mixed model
(LMM). In this study, given a functional graph metric ϕ, we considered a random sample of
P subjects with N repeated measurements of a continuous variable in M visits. ϕijk (for i = 1, …,
N and j = 1, …, M, and k = 1, …, P) denotes the metric from the k th subject’s j th visit and i th
measurement occasions. The three-level LMM models ϕijk as the following equations:
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Graph metric
z}|{
(cid:1)ijk
¼ (cid:3)000|{z}
þ
p0k|{z}
þ
v 0jk|{z}
fixed
intercept
random intercepts
level 3; subjects
random intercepts
level 2; visits
þ eijk|{z}
random
residuals
where γ000 is a fixed parameter (the group mean) and p0k, v0jk, and eijk are independent ran-
dom effects normally distributed with a mean of 0 and variances σp0
2. The term p0k
is the subject effect, v0jk is the visit effect, and eijk is the measurement residual. Age, gender,
and interval (Δt) between two visits are covariants.
2 , and σe
2 , σv0
Network Neuroscience
1083
Optimal network neuroscience computation pipelines
ICC estimation. These variances are used to calculate the test-retest reliability, which is mea-
sured by the dependability coefficient and reflects the absolute agreement of measurements.
The dependability coefficient is a form of ICC commonly, which is the ratio of the variances
due to the object of measurement versus sources of error. To avoid negative ICC values and
obtain more accurate estimation of the sample ICC, the variance components in model are
usually estimated with the restricted maximum likelihood approach with the covariance struc-
ture of an unrestricted symmetrical matrix (Zuo et al., 2013).
R e l
i a b i
l
i
t y ϕð Þ ¼
Vb
Vb þ Vw
¼
σ2
p0
p0 þ σ2
e
σ2
(ICC)
The ICC statistics on the measurement reliability are categorized into five common levels:
0 < ICC ≤ 0.2 (slight); 0.2 < ICC ≤ 0.4 (fair); 0.4 < ICC ≤ 0.6 (moderate); 0.6 < ICC ≤ 0.8
(substantial); and 0.8 < ICC < 1.0 (almost perfect). A metric with moderate to almost perfect
test-retest reliability (ICC ≥ 0.4) is commonly expected in practice. The ICC level should not
be judged only based upon the point statistical estimation of ICC but its confidence intervals
(CI) (Koo & Li, 2016). We employed the nonparametric conditional bootstrap method for
1,000 times to estimate their 95% CIs.
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Statistics evaluation. Our analyses can produce big data of 524,160 ICCs (419,328 for the
global network metrics). These ICCs are grouped into four categories (parcellation, frequency
band, connectivity transformation, and edge filtering scheme), each of which has different
choices. Given each choice of a category, we estimated its density distributions of ICCs and
calculated two descriptive statistics: (1) mean ICC values, which measures the general reli-
ability under the given choice; (2) number of almost perfect (noap) ICC values, which mea-
sures the potential reliability under the given choice.
We further perform Friedman rank sum test to evaluate whether the location parameters of
the distribution of ICCs are the same in each choice. Once the Friedman test is significant, we
employ the pairwise Wilcoxon signed rank test for post hoc evaluations to compare ICCs
between each pair of the distributions under different choices. The statistical significance
levels are corrected with Bonferroni method for controlling the familywise error rate at a level
of 0.05. We develop a method to visualize and evaluate the change of ICCs (i.e., reliability
gradient) between different choices (Figure 1C). Specifically, the reliability can be plotted as a
function of Vb and Vw in its anatomy plane (Xing & Zuo, 2018; Zuo, Xu, & Milham, 2019b). The
gradient of reliability between two choices is modeled by the vector (i.e., the black arrow), and
decomposed into changes of individual variability. The systematic evaluation on the reliability of
the global network metrics determines the optimal network neuroscience by combining the most
reliable pipeline choices, which further generated the nodal metrics’ reliability.
Specifically, using the HCP test-retest dataset, our analytic procedure implemented the four
postanalytic stages: node definition, edge construction, network measurement, and reliability
assessments. The test-retest rfMRI dataset underwent the standardized preprocessing pipeline
developed by the HCP team (Glasser et al., 2013). The second step defines nodes (green box)
using sets of brain areas based on 24 partitions, and then extracts the nodal time series. During
the third step (yellow box), individual correlation matrices are first estimated based upon the
six frequency bands derived from Buzsaki’s theoretical framework on brain oscillations
(Buzsaki & Draguhn, 2004) along with the classical band widely used (0.01–0.08 Hz). These
matrices are then converted into adjacency matrices using 4 × 12 = 48 strategies on edge
filtering. In the fourth step, we performed graph analyses (blue box) by systematically calcu-
lating the brain graph metrics at global, modular, and nodal scales. Finally, test-retest reliability
Network Neuroscience
1084
Optimal network neuroscience computation pipelines
was evaluated (red box) as ICCs with the linear mixed models as described above. We present
details of the other three stages of analyses in the following sections.
Test-Retest Dataset
The WU-Minn Consortium in HCP shared a set of test-retest multimodal MRI datasets of 46
subjects from both the S1200 release and the Retest release. These subjects were retested using
the full HCP 3T multimodal imaging and behavioral protocol. Each subject underwent the four
scans on 2 days (two scans per day: Rest1 versus Rest2) during the first visit and returned
several months later to finish the four scans on another two days during the second visit
(Figure 1B). The test-retest interval ranged from 18 to 328 days (mean: 4.74 months; standard
deviation: 2.12 months). Only 41 subjects (28 females, age range: 26–35 years; 13 males,
age range: 22–33 years) had full length rfMRI data across all the eight scans, 2 visits × 2 days ×
2 (LR and RL encoding directions), and were included in the subsequent analyses. Then we
averaged across the RL and LR encodings for each day, so each subject had four repeated
measurements in the ICC estimation. This sample size is larger than the minimal sample size
(N = 35) for fair reliability with 80% power and significance level of 0.05 based on the above-
mentioned test-retest design (four observations per subject) (Bujang & Baharum, 2017). The
HCP rfMRI protocols for scanning and preprocessing images have been optimized for
reliability.
During the scanning, participants were instructed to keep their eyes open and to let their
mind wander while fixating on a cross-hair projected on a dark background. Data were
collected at the 3T Siemens Connectome Skyra MRI scanner with a 32-channel head coil.
All functional images were acquired using a multiband gradient-echo EPI imaging sequence
(2 mm isotropic voxel, 72 axial slices, TR = 720 ms, TE = 33.1 ms, flip angle = 52°, field of
view = 208 × 180 mm2, matrix size = 104 × 90 and a multiband factor of 8). A total of 1,200
images was acquired for a duration of 14 min and 24 s. Details on the imaging protocols can
be found in Smith et al. (2013).
The protocols of rfMRI image preprocessing and artifact removal procedures are docu-
mented in detail elsewhere and generated the minimally preprocessed HCP rfMRI images.
Artifacts were removed using the ICA-based X-noiseifier (ICA + FIX) procedure, followed by
MS-MAll for intersubject registration. The preprocessed rfMRI data were represented as a time
series of grayordinates (4D), combining both cortical surface vertices and subcortical voxels
(Glasser et al., 2013).
Node Definition
A brain graph defines a node as a brain area, which is generally derived by an element of brain
parcellation (parcel) according to borders or landmarks of brain anatomy, structure, or function
as well as an element of volume (voxel) in imaging signal acquisition or a cluster of voxels
(Sporns, 2013b). Due to the high computational demand of voxel-based brain graph, in this
study, we defined nodes as parcels according to the following brain parcellation strategies
(Figure 2A). A surface-based approach has been demonstrated to outperform other
approaches for fMRI analysis (Coalson, Van Essen, & Glasser, 2018; Zuo et al., 2013), and thus
the nodes are defined in the surface space (total 24 surface parcellation choices). Of note, we
adopted a naming convention for brain parcellations as follows: ParcAbbr-NumberOfParcels’
(e.g., LGP-100 or its whole-brain version wbLGP-458).
HCP multimodal parcellation (MMP). A cortical parcellation generated from multimodal images
of 210 adults from the HCP database, using a semiautomated approach (Glasser et al., 2016).
Brain graph:
A mathematical abstract using graph
thery to define a brain system as a set
of nodes and interconnecting edges.
Network Neuroscience
1085
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
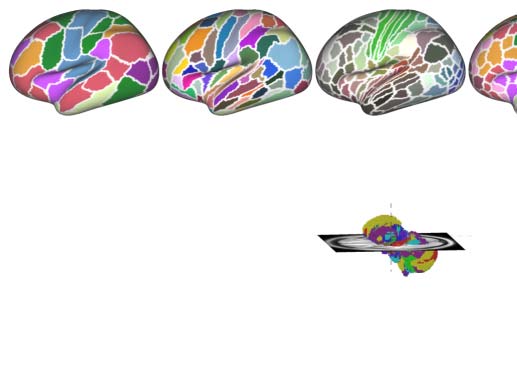
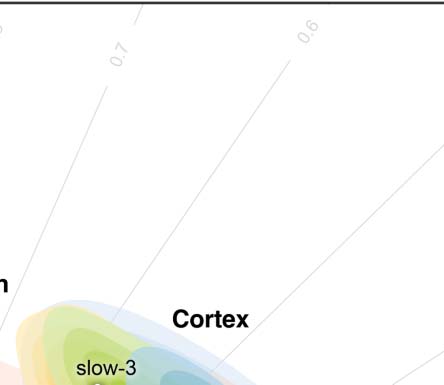
Figure 2. Parcellation choices impact measurement reliability and individual variability. (A) Node definitions are derived from the process of
spatially partitioning the human cortex and whole brain (including both cortical and subcortical nodes) at various resolutions; see more details
of these name abbreviations in Theory and Methods. (B) Density plots are visualized for distributions of the ICCs under the various parcellation
choices on node definition. These density distributions are ranked from top to bottom according to decreases of the mean ICCs while the four
colors depict the four quantiles. (C) Reliability gradient between any one whole-brain parcellation choice and its corresponding cortical par-
cellation choice is decomposed into the axis of changes of the between-subject variability (ΔVb) and the axis of changes of the within-subject
variability (ΔVw). This gradient can be represented as a vector, which is the black arrow from the origin with an angle θ with the x axis, while
the color encodes this angle and the transparency or the length reflects the magnitude of the degree of ICC improvement. According to the
anatomy of reliability, the optimal space is in the second quadrant (quadII) while the first and third quadrant (quadI and quadIII) are suboptimal
for reliability. (D) The improvement in the reliability of the pipeline, which is defined from the cortical parcellations to the corresponding
whole-brain parcellations (including the subcortex), is illustrated by gradient arrows in the plane of individual variability, while controlling
for all other processing steps. Each arrow represents a specific global metric, while controlling for all other processing steps. The position of
the arrows reflects the magnitude of between- and within-subject variability changes (ΔVb, ΔVw), and the size of the arrows indicates the
magnitude of ICC changes.
Cortical regions are delineated with respect to their function, connectivity, cortical architec-
ture, and topography, as well as expert knowledge and meta-analysis results from the literature
(Glasser et al., 2016). The atlas contains 180 parcels for each hemisphere.
Local-global parcellation (LGP). A gradient-weighted Markov random field model integrating
local gradient and global similarity approaches produces the novel parcellations (Schaefer
et al., 2018). The final version of LGP comes with a multiscale cortical atlas including 100,
200, 300, 400, 500, 600, 700, 800, 900, and 1,000 parcels (equal numbers across the two hemi-
spheres). One benefit of using LGP is to have nodes with almost the same size, and these nodes
are also assigned to the 4acommon large-scale functional networks (Thomas Yeo et al., 2011).
Brainnetome parcellation (BNP). Both anatomical landmarks and connectivity-driven informa-
tion are employed to develop this volumetric brain parcellation (Fan et al., 2016). Specifically,
anatomical regions defined as in Desikan et al. (2006) are parcellated into subregions using
functional and structural connectivity fingerprints from HCP datasets. Cortical parcels are
obtained by projecting their volume space to surface space. It is noticed that the original
BNP contains both cortical (105 areas per hemisphere) and subcortical (36 areas) regions
but only the 210 cortical parcels are included for the subsequent analyses.
Network Neuroscience
1086
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
Functional connectivity:
Relationship defined by temporal
dependency between two functional
units (e.g., brain areas).
Natural logarithm linear law:
A special case of power law to divide
frequencies of brain oscillations into
specific bands with relatively
constant relationship to each other
on a natural logarithmic scale and a
constant ratio between any pair of
neighboring bands.
Inclusion of subcortical areas has been shown to contain unignor-
Whole-brain parcellation (wb).
able influences on brain graph analyses (D. Greene et al., 2020; Noble et al., 2019), and we thus
also constructed brain graphs with subcortical structures in volume space as nodes by adding
these nodes to the cortical brain graphs. To get a high-resolution subcortical parcellation, we
adopted the 358 subcortical parcels in Ji et al. (2019). The authors employed data of 337 unre-
lated HCP healthy volunteers and extended the MMP cortical network partition into subcortex.
This results in a set of whole-brain parcellations by combining these subcortical parcels with the
aforementioned cortical parcellations, namely wbMMP, wbLGP, and wbBNP. We noticed that
the wbMMP-718 has been named by Ji et al. (2019) as the Cole-Anticevic Brain-wide Network
Partition, and we thus renamed the wbMMP-718 as wbCABP-718 for consistency.
Edge Construction
After defining the node with each parcellation, in each parcel, regional mean time series were
estimated by averaging the vertex time series at each time point. To construct an edge between
a pair of nodes, their representative time series entered into the following steps in order: band-
pass filtering, internode connectivity transformation, and edge filtering.
Band-pass filtering. Resting-state functional connectivity studies have typically focused on
fluctuations below 0.08 Hz or 0.1 Hz (Biswal, Zerrin Yetkin, Haughton, & Hyde, 1995; Fox
& Raichle, 2007) and assumed that only these frequencies contribute significantly to inter-
regional functional connectivity while other frequencies are artifacts (Cordes et al., 2001).
In contrast, however, other studies have found that specific frequency bands of the rfMRI oscil-
lations make unique and neurobiologically meaningful contributions to resting-state functional
connectivity (Salvador et al., 2005; Zuo & Xing, 2014). More recently, with fast fMRI methods,
some meaningful FC patterns were reported across much higher frequency bands (Boubela
et al., 2013). These observations motivate exploring a range of frequency bands beyond those
typically studied in resting-state functional connectivity studies.
Buzsaki and Draguhn (2004) proposed a hierarchical organization of frequency bands
driven by the natural logarithm linear law. This offers a theoretical template for partitioning
rfMRI frequency content into multiple bands (Figure 3A). The frequencies occupied by these
bands have a relatively constant relationship to each other on a natural logarithmic scale and
have a constant ratio between any given pair of neighboring frequencies (Buzsáki, 2009).
These different oscillations are linked to different neural activities, including cognition,
emotion regulation, and memory (Achard, Salvador, Whitcher, Suckling, & Bullmore, 2006;
Buzsáki, 2009; Fox & Raichle, 2007). Advanced by the fast imaging protocols offered by the
HCP scanner, the short scan interval (TR = 720 ms) allows us to obtain more oscillation classes
that the traditional rfMRI method. We incorporate Buzsaki’s framework (Buzsaki & Draguhn,
2004; Penttonen & Buzsáki, 2003) with the HCP fast-TR datasets by using the DREAM toolbox
(Gong et al., 2021) in the Connectome Computation System (Xing, Xu, Jiang, Wang, & Zuo,
2022; Xu, Yang, Jiang, Xing, & Zuo, 2015). It decomposed the time series into the six slow
bands as illustrated in Figure 3A.
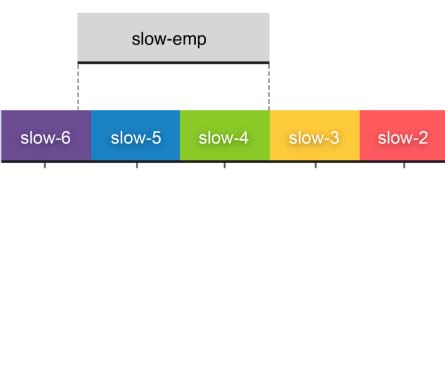
Connectivity transformation. For each scan, individual nodal representative time series were
band-pass filtered with each of the six frequency bands, and another empirical frequency
band, slow-emp (0.01–0.08 Hz). The Pearson’s correlation rij 2 [−1, 1] between the filtered
time series of each pair of nodes i = 1, …, N, j = 1, …, N was calculated (N is the number of
nodes). These correlation values provided an estimation on the edge strengths between the
two nodes and formed a N × N symmetric correlation matrix R = (rij) for each given subject,
scan, parcellation, and frequency band.
Network Neuroscience
1087
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
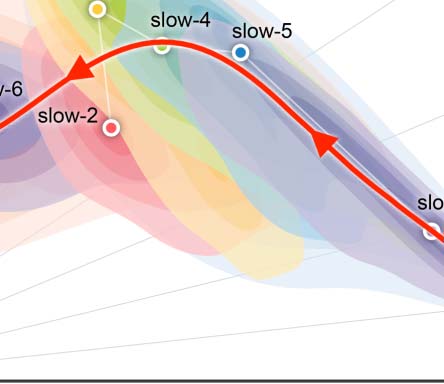
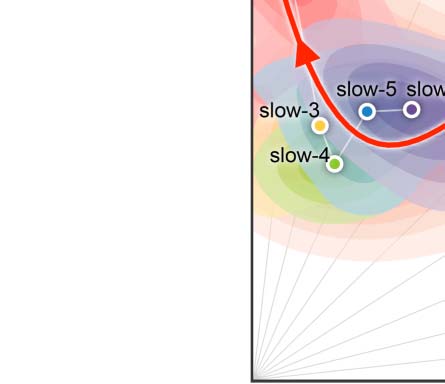
Figure 3. Reliability gradient across the slow bands and changes of related individual variability. (A) Classes of frequency bands for slow
oscillations derived from the natural logarithm linear law. (B) Density plots are visualized for the ICC distributions under the various frequency
bands. These density distributions are ranked from top to bottom according to decreases of the mean ICCs, while the vertical lines depict the
four quartiles. (C) Network measurements are projected onto the reliability anatomy plane coordinated by both between- and within-subject
variability. These dot plots are fitted into the topographic (contour) maps where the local maxima for each band is labeled as a circle. To
highlight the trend of increasing reliability as the frequency band increases, a fourth-order polynomial curve (represented by a red line) is
fitted to the frequency contour plot peak points, tracing the reliability flow along slow-to-fast oscillations in the cortex and whole brain.
Many network metrics are not well defined for negatively weighted connections. In order to
ensure that the connection weights are positive only, we applied four types of transformations
to the symmetric correlation matrix: the positive (Equation pos), absolute (Equation abs), expo-
nential (Equation exp) and distance-inverse (Equation div) functions, respectively. This avoids
the negative values in the internode connectivity matrix W = (wij) where zij = tanh−1 (rij) is
Fisher’s z−transformation.
(cid:2)
(cid:2)
(cid:2)
(cid:2)
zij þ zij
2
wij ¼
Þ
2 0; ∞½
(cid:2)
(cid:2)
wij ¼ zij
(cid:2)
(cid:2) 2 0; ∞½
Þ
wij ¼ ezij 2 0; ∞½
Þ
wij ¼
q
2
ffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffiffi
(cid:5)
(cid:4)
2 (cid:2) 1 − rij
2 0; ∞ð
Þ
(pos)
(abs)
(exp)
(div)
The connectivity matrix represents a set of the node parcels and relational quantities
between each pair of the nodes, and will serve as the basis of following edge filtering proce-
dure for generation of the final brain graphs.
Connectome:
A term describing the whole-brain
graph and initiating the field of
connectomics.
In a graph, edges represent a set of relevant interactions of crucial importance to
Edge filtering.
obtain parsimonious descriptions of complex networks. Filtering valid edges can be highly
challenging due to the lack of ‘ground truth’ of the human brain connectome. To provide a
Network Neuroscience
1088
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
reliable way of building candidate edges, we sampled the following 12 schemes on edge
filtering and applied them to the connectivity matrices.
Absolute weight thresholding (ABS). This approach selects those edges that exceed a manually
defined absolute threshold (e.g., correlations higher than 0.5), setting all correlations smaller
than 0.5 to 0 (ABS05). This is a simple approach to reconstruct networks (Hagmann et al.,
2007).
It is a common step in the reconstruction of functional brain
Proportional thresholding (PROP).
networks to ensure equal edge density across subjects (D. Bassett et al., 2009; Rubinov,
Sporns, van Leeuwen, & Breakspear, 2009; van den Heuvel et al., 2017). It keeps the number
of connections fixed across all individuals to rule out the influence of network density on the
computation and comparison of graph metrics across groups. This approach includes the
selection of a fixed percentage of the strongest conncections as edges in each individual net-
work or brain graph. Compared to ABS, PROP has been argued to reliably separate density
from topological effects (Braun et al., 2012; Ginestet, Nichols, Bullmore, & Simmons, 2011)
and to result in more stable network metrics (Garrison, Scheinost, Finn, Shen, & Constable,
2015). This makes it a commonly used approach for network construction and analysis in
disease-related studies. Here, we focused on two thresholds that are commonly reported in
the literature: 10% (PROP10) and 20% (PROP20).
Degree thresholding (DEG). The structure of a graph can be biased by the number of existing
edges. Accordingly, statistical measures derived from the graph should be compared against
graphs that preserve the same average degree, K. A threshold of the degree can be chosen to
produce graphs with a fixed mean degree (e.g., K = 5, DEG5), which is the average nodal
degrees of an individual graph from a single subject’s scan. Many network neuroscience
studies have taken this choice for K = 5 (S. I. Dimitriadis, Laskaris, Del Rio-Portilla, & Koudounis,
2009; Micheloyannis et al., 2006; Milo et al., 2002; Stam, Jones, Nolte, Breakspear, & Scheltens,
2007). We also include the DEG15 for denser graphs of the brain networks.
Global cost efficiency optimization (GCE). Given a network with a cost ρ, its global efficiency is
a function of the cost Eg(ρ), and its GCE is J(ρ) = Eg(ρ) − ρ. Several studies suggested that
brain networks, in particular those with small-world topology, maximize their global cost
efficiency (D. S. Bassett et al., 2008), that is, Jmax = maxρ J(ρ). Computationally, this scheme
is implemented by looping all network costs (e.g., adding edges with weights in order) to
find the Jmax where the corresponding edge weight was determined as the threshold for edge
filtering. In this sense, GCE is an individualized and optimized version of ABS, PROP, and
DEG, while the latter three are commonly employed with a fixed threshold for all
individuals.
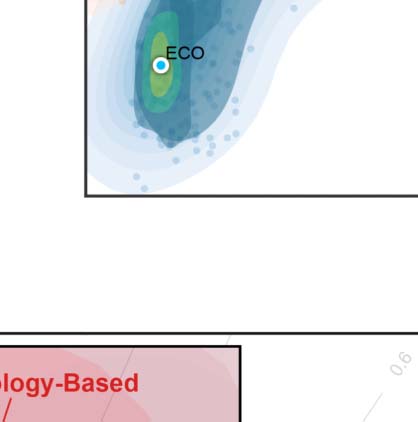
Overall efficiency cost optimization (ECO). Both global and local efficiency are important graph
features to characterize the structure of complex systems in terms of integration and segrega-
tion of information (Latora & Marchiori, 2001). ECO was proposed to determine a network
density threshold for filtering out the weakest links (De Vico Fallani, Latora, & Chavez,
2017). It maximizes an extension of Jmax, the ratio between the overall (both global and local)
efficiency and its wiring cost maxρ Jext(ρ) = (Eg(ρ) + Eloc(ρ))/ρ, where Eloc denotes the network
local efficiency. The study (Latora & Marchiori, 2001) also demonstrated that, to maximize J,
these networks have to be sparse with an average node degree K ≃ 3.
Minimum spanning tree (MST). This is an increasingly popular method for identifying the
smallest and most essential set of connections while ensuring that the network forms a fully
Network Neuroscience
1089
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
connected graph (Guo, Qin, Chen, Xu, & Xiang, 2017; Meier, Tewarie, & Van Mieghem, 2015;
Otte et al., 2015; van Nieuwenhuizen et al., 2018). The tenet of using MST is to summarize infor-
mation and index the structure of the graph, and thus remove edges with redundant information
(Mantegna, 1999). Specifically, an MST filtered graph will contain N nodes connected via N − 1
connections with minimal cost and no loops. This addresses key issues in existing topology
filtering schemes that rely on arbitrary and user-specified absolute thresholds or densities.
Orthogonal minimum spanning tree (OMST). This topological filtering scheme was proposed
recently (S. Dimitriadis, Antonakakis, Simos, Fletcher, & Papanicolaou, 2017) to maximize
the information flow over the network versus the cost by selecting the connections via the
OMSTs. It samples the full-weighted brain network over consecutive rounds of MST that are
orthogonal to each other. Practically, we extracted the first MST, and then we cleared their
connections and we tracked the second MST from the rest of the network connections, and
so forth. Such an iterative procedure (stopped by the Mth MST) can get orthogonal MSTs and
topologically filter brain network by optimizing the GCE under the constrains by the MST,
leading to an integration of both GCE and MST:
max
n2 1;M½
(cid:3)
ð
J ρ nMSTs
ð
ð
ð
Þ ¼ Eg ρ nMSTs
Þ
ð
Þ − ρ nMSTs
Þ
Þ
Planar maximally filtered graph (PMFG). The idea underneath PMFG (Tumminello, Aste, Di Matteo,
& Mantegna, 2005) is to filter a dense matrix of weights by retaining the largest possible
subgraph while imposing global constraints on the derived network topology. Edges with
the strong connection weights are retained while constraining the subgraph to be a (spanning)
tree globally. Similarly, during the PMFG construction, the largest weights are retained while
constraining the subgraph to be a planar graph globally. The PMFG algorithm searches for the
maximum weighted planar subgraph by adding edges one by one. The resulting matrix is
sparse with 3(N − 2) edges. It starts by sorting all the edges of a dense matrix of weights in
nonincreasing order and tries to insert every edge in the PMFG. Edges that violate the planarity
constraint are discarded.
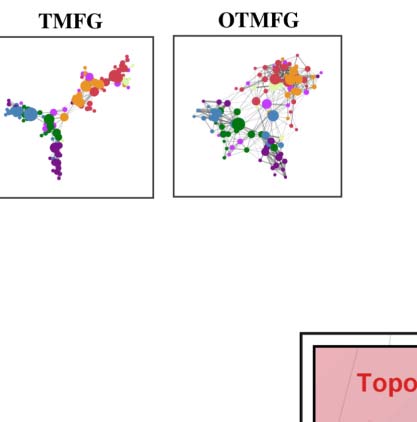
Triangulated maximally filtered graph (TMFG). The algorithm for implementing PMFG is compu-
tationally expensive, and is therefore impractical when applied to large brain networks
(Massara, Di Matteo, & Aste, 2016). A more efficient algorithm, TMFG, was developed that
exhibited greatly reduced computational complexity compared to PMFG. This method cap-
tures the most relevant information between nodes by approximating the network connectivity
matrix with the endorsement association matrix and minimizing spurious associations. The
TMFG-derived network contains three-node (triangle) and four-node (tetrahedron) cliques,
imposing a nested hierarchy and automatically generates a chordal network (Massara et al.,
2016; Song, Di Matteo, & Aste, 2012). Although TMFG is not widely applied in network neuro-
science studies, it has been applied elsewhere and proven to be a suitable choice for modeling
interrelationships between psychological constructs like personality traits (Christensen, Kenett,
Aste, Silvia, & Kwapil, 2018).
Orthogonal TMF graph (OTMFG). To combine both the TMFG’s efficiency and OMST’s accu-
racy, we propose OTMFG to maximize the information flow over the network versus the cost
by selecting the connections of the orthogonal TMFG. It samples the full-weighted brain net-
work over consecutive rounds of TMFG that are orthogonal to each other.
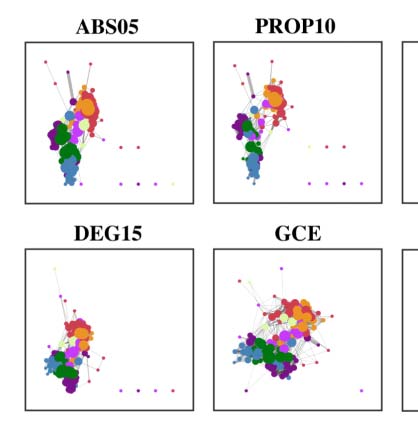
In summary, as illustrated in Figure 4A, the 12 edge filtering schemes transform a fully
weighted matrix into a sparse matrix to represent the corresponding brain network. They can
Network Neuroscience
1090
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
t
/
.
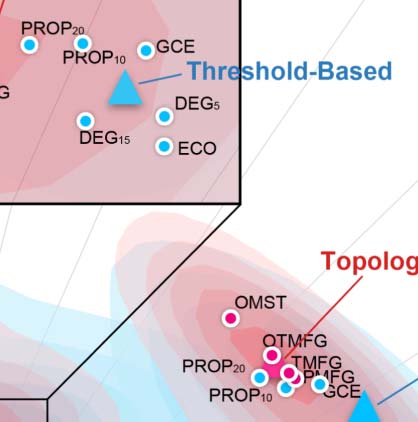
Edge filtering schemes and their networking performance. (A) Twelve schemes of filtering edge are applied to an individual
Figure 4.
connectivity matrix, resulting in the 12 brain networks with their nodes colored as the Yeo2011-7Networks (Thomas Yeo et al., 2011).
(B) Global cost efficiency are plotted against network wiring costs of all the brain networks derived with the 12 edge filtering schemes
from all the individual rfMRI scans. Red dots represent the topology-based, while blue dots are for threshold-based networks. These dot
plots are fitted into the topographic (contour) maps where the local maxima for each filtering choice is labeled as a circle. (C) Density plots
are for ICC distributions under various the 12 edge filtering schemes. These density distributions are ranked from top to bottom according to
decreases of the mean ICCs, while the two colors depict the topology-based and threshold-based schemes. Four quartiles were indicated by
vertical lines. (D) Network measurements are projected onto the reliability anatomy plane coordinated by both between- and within-subject
variability. Red dots represent the topology-based, while blue dots are for threshold-based networks. The topographic (contour) maps fit the
dots and label the local maxima as a circle for each scheme and the global maxima as a triangle for the topology and threshold groups,
respectively.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
be categorized into two classes: threshold-based versus topology-based schemes. ABS05,
PROP10, PROP20, DEG5, DEG15, ECO, and GCE rely on a threshold for filtering and retaining
edges with higher weights than the threshold. These schemes normally ignore the topological
structure of the entire network and can result in isolated nodes. In contrast, the topology-based
methods including MST, OMST, PMFG, TMFG, and OTMFG all consider the global network
topology in determining which edges to retain. As illustrated in Figure 4B, all the schemes are
plotted in the ρ − Jmax plane for their network economics.
Network Neuroscience
1091
Optimal network neuroscience computation pipelines
Table 1.
A list of the employed network metrics derived with graph theory
Measure
Attribute
Global efficiency of the network
Symbol
Eg
Reference
Latora & Marchiori, 2001
Integration
Average shortest path length of the network
Pseudo diameter of the network
Global
Clustering coefficient of the network
Lp
D
Cp
Watts & Strogatz, 1998
Bouttier, Di Francesco, & Guitter, 2003
Watts & Strogatz, 1998
Segregation
Local efficiency of the network
Elocal
Latora & Marchiori, 2001
Modularity of the network
Transitivity of the network
Local characteristic path length of nodes
Efficiency of nodes
Local efficiency of nodes
Clustering coefficient of nodes
Nodal
Centrality
Pagerank centrality of nodes
Degree centrality of nodes
Eigenvector centrality of nodes
Resolvent centrality of nodes
Subgraph centrality of nodes
Betweenness centrality of nodes
Q
Tr
Lpi
Enodal,i
Elocal,i
Cpi
Pci
Dci
Eci
Rci
Sci
Bci
Newman, 2004
Newman, 2003
Watts & Strogatz, 1998
Latora & Marchiori, 2001
Latora & Marchiori, 2001
Watts & Strogatz, 1998
Page, Brin, Motwani, & Winograd, 1999
Pastor-Satorras, Vázquez, &
Vespignani, 2001
Newman, 2008
Estrada & Higham, 2010
Estrada & Rodriguez-Velazquez, 2005
Freeman, 1978
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
.
/
t
Network Analysis
We performed graph theory–driven network analysis by calculating several common graph-
based metrics for the resulting graphs. These measures, broadly, can be interpreted based
on whether they characterize the extent to which network structure allows for integrated or
segregation information flow. Examples of integrative measures include average shortest path
length (Lp), global efficiency (Eg), and pseudo diameter (D). Segregation measures include clus-
tering coefficient (Cp), local efficiency (Elocal), transitivity (Tr), modularity (Q), and a suite of
nodal centrality measures (Table 1). All the metrics are calculated using the Brain Connectivity
Toolbox (Rubinov & Sporns, 2010). We employed graph-tool (https://graph-tool.skewed.de)
and NetworKit (https://networkit.github.io) to achieve high performance comparable (both
in memory usage and computation time) to that of a pure C/C++ library.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
RESULTS
Whole-brain Networks Are More Reliable Than Cortical Networks
We evaluated reliability based on 24 different parcellation choices (Figure 2A). In the follow-
ing parts of the paper, we name a parcellation as ‘ParcAbbr-NumberOfParcels’ (e.g., LGP-100
or its whole-brain version wbLGP-458). We found significant differences in ICC distributions
across the 24 parcellation choices (Figure 2B, Friedman rank sum test: χ2 = 20379.07, df = 23,
Network Neuroscience
1092
Optimal network neuroscience computation pipelines
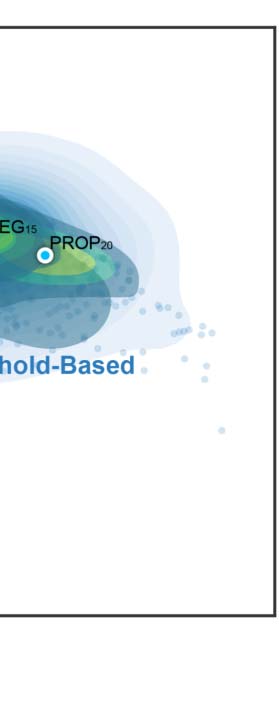
p < 2.2 × 10−16, effect size WKendall = 0.377). The mean ICCs range from slight (LGP-1000) to
substantial (wbLGP-458). Given a particular parcellation and definition of nodes, we illustrate
the density distribution of its ICCs under all other strategies (edge definition and metric deri-
vation). Notably, whole-brain parcellations yield higher measurement reliability than parcella-
tions of cerebral cortex on their own (the effect sizes > 0.65). This improvement in reliability
seems not simply a by-product of having more parcels. We chose the parcellations in which
the number of parcels (400 ≤ n ≤ 1,000) almost overlapped between the cortex and the whole
brain, and found no correlation between the number of parcels and the median ICCs (r =
−0.11, p = 0.7). We report the mean ICC and the number of almost perfect (noap) ICCs
(≥0.8) as the descriptive statistics for the density distributions. The wbLGP-458 (mean ICC:
0.671; noap ICC: 519), wbLGP-558 (mean ICC: 0.671; noap ICC: 540) and The wbBNP-
568 (mean ICC: 0.664; noap ICC: 511) are the three most reliable choices (see more details
of the post hoc Wilcoxon signed rank test in Supporting Information Table S7). Among the
cortical parcellations, the LGP-500 (mean ICC: 0.362; noap ICC: 0), LGP-400 (mean ICC:
0.342; noap ICC: 0), and LGP-600 (mean ICC: 0.340; noap ICC: 0) are the three most reliable
choices (Supporting Information Table S3).
To better understand the effect of introducing 358 subcortical parcels into the cortical par-
cellations, we decomposed the reliability changes into a two-dimensional representation of
changes of individual variability (Figure 2C, and D). This idea was motivated by the analysis
of reliability derived with individual variability (Xing & Zuo, 2018; Zuo, Xu, & Milham, 2019b)
as in Figure 1C. For each ICC under a given parcellation choice, we calculated the related
between-subject variability Vb and within-subject variability Vw. Changes in the individual
variability associated with the reliability improvements from cortical to whole-brain pipelines
were plotted along with ΔVb and ΔVw as arrows. These arrows are distributed across the three
quadrants (quadI: 0.94%; quadII: 59.99%; quadIII: 39.07%). We noticed that most of these
arrows were distributed into the optimal quadrant where the improvements of test-retest
reliability by the whole-brain parcellation choices largely attributing to the increases of
between-subject variability and decreases of within-subject variability. The decreases of both
between-subject and within-subject variability may also strengthen the measurement reli-
ability (the suboptimal quadIII in Figure 2).
Spontaneous Brain Activity Portrays More Reliable Networks in Higher Slow Bands
Brain oscillations are hierarchically organized, and their frequency bands were theoretically
driven by the natural logarithm linear law (Buzsaki & Draguhn, 2004). By analogy, rfMRI oscil-
lations can, similarly, be partitioned into distinct frequency bands. Advanced by the fast imag-
ing protocols (TR = 720 ms), HCP test-retest data allow to obtain more oscillation classes than
traditional rfMRI acquisitions (typical TR = 2 s). We incorporate Buzsaki’s framework with the
HCP dataset using the DREAM toolbox (Gong et al., 2021) in the Connectome Computation
System to decompose the time series into the six slow bands (Figure 3A): slow-6 (0.0069–
0.0116 Hz), slow-5 (0.0116–0.0301 Hz), slow-4 (0.0301–0.0822 Hz), slow-3 (0.0822–
0.2234 Hz), slow-2 (0.2234–0.6065 Hz), and slow-1− (0.6065–0.6944 Hz).
We noticed that, due to the limited sampling rate (TR), this slow-1− only covers a small part
of the full slow-1 band (0.6065–1.6487 Hz) – we indicate this above. We also included the
frequency band, slow-emp (0.01–0.08 Hz) for the sake of comparison, as it is covers a range
commonly used in rfMRI studies. A significant effect on order (χ2 = 9283.536, df = 6, p < 2.2 ×
10−16, WKendall = 0.192) across the frequency bands was revealed based on the density distri-
butions of ICC (Figure 3B): slow-2, slow-1−, slow-3, slow-emp, slow-4, slow-5, and slow-6.
Spontaneous brain acitivity:
Activity not attributable to specific
brain inputs or outputs and
representative neuronal activity
intrinsically generated and organized
in the brain.
Network Neuroscience
1093
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
Post hoc paired tests indicated that any pairs of neighboring bands are significantly different
from one another, with measurement reliability increasing with faster frequency bands. Note,
however, that slow-1− (mean ICC: 0.564) did not fit into this trend, possibly due to its limited
coverage of the full band. But remarkably, slow-1− exhibited the largest number of almost pre-
fect ICCs for potential reliability (noap ICC: 1746). Slow-emp (mean ICC: 0.519; noap ICC:
434) contains overlapping frequencies with both slow-4 (mean ICC: 0.560; noap ICC: 441)
and slow-5 (mean ICC: 0.494; noap ICC: 285), and higher ICCs than the two bands but the
effect sizes are small to moderate (slow-emp vs. slow-4: 0.193; slow-emp vs. slow-5: 0.485).
Slow-6 is the choice with the lowest ICCs (mean ICC: 0.331; noap ICC: 154) compared to
other bands (large effect sizes: r > 0.57).
To visualize reliability variation across frequency bands, we plotted a trajectory tracing reli-
ability flow along the five full (slow-6 to slow-2) bands in the reliability plane, whose axes
correspond to between-versus within-subject variability (Figure 3C). As expected, this nonlin-
ear trajectory contains two stages of almost linear changes of the network measurement reli-
ability from slow to fast oscillations: whole brain versus cortex. In each case, the reliability
improvements attribute to both increases of between-subject variability and decreases of
within-subject variability while the improvements of whole-brain network measurement reli-
ability were largely driven by the increased variability between subjects.
Topological Economics Individualize Highly Reliable Functional Brain Networks
Estimating functional connections can be highly challenging due to the absence of a ‘ground
truth’ human functional connectome. To provide a reliable way of building candidate edges of
the connections, we sampled the 12 schemes on graph edge filtering (Figure 4A), which turn a
fully connected matrix into a sparse graphical representation of the corresponding brain net-
travail. These schemes can be categorized into two classes: threshold-based versus topology-
based schemes. Threshold-based schemes usually use a threshold to preserve those edges
whose strengths are above a cutoff value, such as ABS05, PROP10, PROP20, DEG5, et
DEG15. Threshold-based schemes are widely used in network neuroscience and ignore the
intrinsic topological structure of the entire brain network (par exemple., leading to multiple connected
components or isolated nodes). In contrast, topology-based schemes such as MST, OMST,
PMFG and TMFG come from other scientific disciplines and are optimized based on the entire
network topology (see Materials and Methods). To combine both the TMFG’s efficiency and
OMST’s accuracy, we proposed the OTMFG. All the schemes are plotted in the plane of cost
versus global cost efficiency to better visualize the economical properties of the derived net-
travaux (Figure 4B). These plots are fitted into the topographic (contour) maps where the local
maxima for each filtering choice is labeled as a circle. The human brain networks achieve
higher global efficiency with lower cost by using topology-based schemes compared to
threshold-based schemes, suggesting increasingly optimal economics.
Significant differences in test-retest reliability were detectable across these 12 edge filtering
schemes (χ2 = 9784.317, df = 11, p < 2.2 × 10−16, WKendall = 0.189, see Figure 4C). Among the
topology-based schemes, OMST (mean ICC: 0.608; noap ICC: 765), OTMFG (mean ICC:
0.602; noap ICC: 781), and TMFG (mean ICC: 0.570; noap ICC: 767) were the three most
reliable choices. They showed significantly greater reliability than the three most reliable
threshold-based, respectively: PROP20 (mean ICC: 0.593; noap ICC: 632), PROP10 (mean
ICC: 549; noap ICC: 445), and GCE (mean ICC: 0.533; noap ICC: 352). Mean reliability of
MST are slight to fair (mean ICC: 0.309) but its number of almost perfect reliability (noap
ICC:362) is still higher than all threshold-based schemes except PROP10 and PROP20.
Network Neuroscience
1094
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
Network measurements are labeled based on topology and threshold groups and projected
onto the reliability anatomy plane, whose axes represent between- and within-subject vari-
ability (Figure 4D). The contour maps are reconstructed for each scheme based upon the
individual variability of all the related network measurements. The topology-based methods
(red) showed overall higher ICCs than the threshold-based methods (blue), improvements
that could be attributed to increases in between-subject variability and decreases of
within-subject variability. These observations are consistent between cortex and whole-brain
networks while topology-based whole brain network are almost perfectly reliable (meaning
almost perfect reliability, i.e., ICC ≥ 0.8).
We also explored connection transformation and edge weights, two factors included in
edge filtering, the choices of connectivity transformation, and weighing edges, regarding their
measurement reliability. Positive (Equation pos) (mean ICC: 0.512; noap ICC: 1,031) and expo-
nential (Equation exp) transformation (mean ICC: 0.509; noap ICC: 1,855) were the two most
reliable choices. Comparing to the positive and absolute (Equation abs) (mean ICC: 0.508;
noap ICC: 1,050) transformation, the exponential and distance-inverse (Equation div) (mean
ICC: 0.500; noap ICC: 1,031) transformation show larger number of almost perfect ICCs.
Weighted graphs are also more reliable than the binary graphs, while the normalized weighted
graphs demonstrated the highest ICCs, reflecting both the increased between-subject vari-
ability and decreased within-subject variability.
Network Integration and Segregation Can Serve Reliable Metrics of Information Flow
The previous extensive data analysis suggests that the optimally reliable pipeline should (1)
define network nodes using a whole-brain parcellation, (2) filter the time series with higher
frequency bands, (3) transform the connectivity using positive transformation, and (4) construct
network edges using topology-based methods and normalized weights. Using the optimal
pipelines, we evaluated the reliability levels of various metrics from network neuroscience
and their differences across individuals. Focusing on the optimized pipeline with the highest
ICCs of the various choices (wbLGP-458, slow-2, pos, normalized weights, OMST), we
reported test-retest reliability of the measurements as well as their corresponding individual
variability. In Figure 5A, we found that the global network measurements of information
segregation and integration are at the level of almost perfect reliability except for the modu-
larity Q (ICC = 0.46, 95% CI = [0.252, 0.625]). These high-level ICCs are derived with large
between-subject variability and small within-subject variability (Figure 5B). These findings are
reproducible across the other two parcellation choices (wbCABP-718, wbBNP-458). In
consideration of “Ease-of-Use” for researchers and higher cortical resolution, we mapped
the “Out-of-the-Box” Cole-Anticevic Brain-wide Network Partition (wbCABP-718) for nodal
metrics visualization.
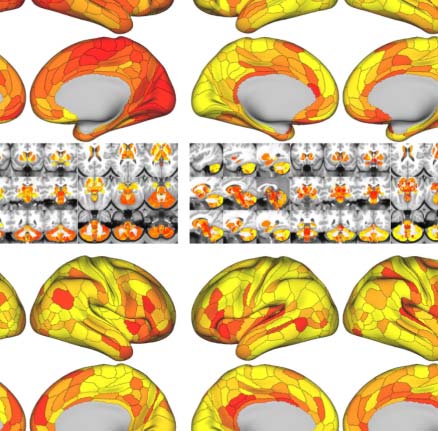
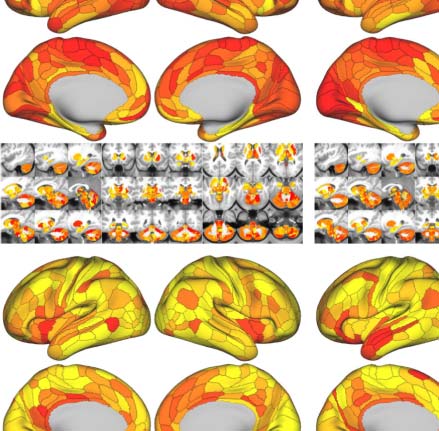
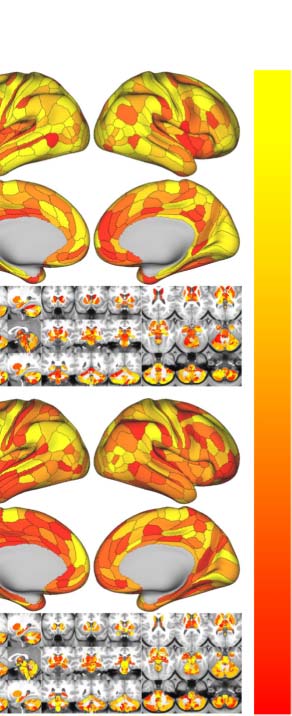
Similar to the global metrics, shortest path length Lp and nodal efficiency Enodal exhibited
the highest ICCs (almost perfect test-retest reliability), while ICCs of other nodal metrics
remained less than 0.6. To visualize node-level network metrics, we reported results derived
from the wbCABP-718 choice. To improve spatial contrasts of reliability, we ranked the parcels
according to their ICCs and visualized the ranks in Figure 5C. Most nodal metrics are more
reliable across the 360 cortical areas than the 358 subcortical areas ( Wilcoxon tests: all
p values less than 0.001, corrected for multiple comparisons). However, Lp, Enodal, and Bc exhib-
ited higher across subcortical areas than cortical areas (corrected p < 0.001). Across the human
cerebral cortex, the right hemispheric areas demonstrated more reliable Cp (corrected p <
0.0036) than the left hemispheric areas. Interesting patterns of the reliability gradient are also
Network Neuroscience
1095
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
a
_
0
0
3
1
5
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Measurement reliability and variability of global/nodal network metrics under the optimized pipeline. (A) Spider plots are visual-
ized for ICCs (test-retest) with the 95% confidence intervals (CIs, shadow bands) for the global metrics of network integration, segregation.
Integrative measures include average shortest path length (Lp), global efficiency (Eg), and pseudo diameter (D). Segregation measures include
clustering coefficient (Cp), local efficiency (Eloc), transitivity (Tr ), and modularity (Q). (B) The reliability anatomy was plotted as a function of
between-subject variability (Vb) and within-subject variability (Vw). (C) Ranks of ICCs across the 360 cortical parcels and the 358 subcortical
parcels in the optimal pipeline (wbCABP-718, slow-2, pos, OMST) are depicted. Ten nodal metrics are assessed including local characteristic
path length of node (Lplocal), nodal efficiency (Enodal), local efficiency of nodes (Elocal), nodal clustering coefficient (Cp), pagerank centrality (Pc),
degree centrality (Dc), eigenvector centrality (Ec), resolvent centrality (Rc), subgraph centrality (Sc), and betweeness centrality (Bc).
observable along large-scale anatomical directions (dorsal > ventral, posterior > anterior)
across the nodal metrics of information segregation and centrality. These spatial configuration
profiles on the reliability reflected their correspondence on interindividual variability of these
metrics, characterising the network information flow through the slow-2 band.
Building An Open Resource for Reliable Network Neuroscience
The results presented here represent huge costs in terms of computational resources (plus que
1,728,000 core-hours on CNGrid), supported by Chinese Academy of Sciences (https://cscgrid
.cas.cn). Derivations of the ICCs and their linear mixed models were implemented in R and
Python. As is our practice in open science, we have started to provide an online platform
on the reliability assessments (https://ibraindata.com/research/ifNN/reliabilityassessment).
The big reliability data were designed into an online database for providing the community
a resource to search reliable choices and help with final decision-making. The website for this
Neurosciences en réseau
1096
Optimal network neuroscience computation pipelines
online database provided more details of the reliability data use. We shared all the codes,
figures, and other reliability resources via the website (https://ibraindata.com/research/ifNN
/database).
DISCUSSION
This study examined the series of processing and analysis decisions in constructing graphical
representations of brains’ intrinsic spontaneous activity. The focus, ici, was on identifying the
pipeline that generated reliable, individualized networks and network metrics. The results of
our study suggest that to derive reliable global network metrics showing higher interindividual
variances and lower intraindividual variances, one should use whole-brain parcellations to
define network nodes, focus on higher frequencies in the slow band for time series filtering
to derive the connectivity, and use the topology-based methods for edge filtering to construct
sparse brain graphs. Regarding network metrics, multilevel or multimodal metrics appear more
reliable than single-level or single-model metrics. Deriving reliable measurements is critical in
network neuroscience, especially for translating network neuroscience into personalized prac-
tice. Based on these results, we provide four principles toward optimal functional network
neuroscience for reliability of measuring individual differences.
Principle I: Use A Whole Brain Parcellation to Define Network Nodes
The basic unit of a graph is the node. Cependant, variability across brain parcellations can yield
different graphs, distorting network metrics and making it difficult to compare findings across
études (Wang et al., 2009; Zalesky et al., 2010). In many clinical applications (Fornito et al.,
2015; Matthieu & Hampshire, 2016) researchers aim to identify disease-specific connectivity
profiles of the whole brain, including cortical and subcortical structures, as well as cerebellum.
A recent review has raised the concern that many studies have focused on restricted sets of
nodes, Par exemple, cortex only, and has called for a field standard for the best practices in
clinical network neuroscience (Hallquist & Hillary, 2019), which requires almost perfectly reli-
able measurements (Xing & Zuo, 2018). Our meta-reliability assessments revealed such high
reliability of measurements made involving functional brain networks can be achieved,
through the inclusion of high-resolution subcortical nodes. This provides strong evidence that
whole-brain node use should be part of the standard analysis pipeline for network neurosci-
ence applications. These improvements of reliability can be attributed to increases in between-
subject variability coupled with reductions in within-subject variability relative to networks of
cortical regions alone. One possible neuroanatomical explanation is that distant areas of cere-
bral cortex are interconnected by the basal ganglia and thalamus while also communicating
with different regions of the cerebellum via polysynaptic circuits, forming an integrated con-
nectome (Bostan & Strick, 2018). These subcortical structures have been suggested to play a
role in both primary (par exemple., moteur) and higher order function (par exemple., learning and memory), alors que
studies using rfMRI have delineated the resting-state functional connectivity (RSFC) maps
between these subcortical structures and cortical networks of both primary and high-order
les fonctions. Fait intéressant, a recent work revealed that interindividual variance in cerebellar RSFC
networks exceeds that of cortex (Marek et al., 2018). Entre-temps, these RSFC maps are highly
individualized and stable within individuals (D. Greene et al., 2020), indicating that they
possess reliable characteristics. In line with our observations, we argue that inclusion of the
subcortical structures as network nodes can enhance the between-subject variability and
stabilize the within-subject variability by providing more comprehensive measurements on
the entirety of the brain connectivity.
Neurosciences en réseau
1097
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
Principle II: Generate Functional Networks using Spontaneous Brain Activity in Multiple Slow Bands
It has been a common practice in rfMRI research to estimate the RSFC profile based on BOLD
time series of the intrinsic spontaneous brain activity from the low frequency (0.01–0.1 Hz or
0.01–0.08 Hz). Cependant, the test-retest reliability of RSFC measurements derived from this
frequency band has been limited, with ICCs less than 0.4 (Noble et al., 2019; Zuo & Xing,
2014). Existing studies, cependant, have advocated adopting a multifrequency perspective to
examine the amplitude of brain activity at rest (Zuo et al., 2010) and its network properties
(Achard et al., 2006). This approach has been spurred along by recent advances in multi-
banded acquisitions and fast imaging protocols, offering rfMRI studies a way to examine
spontaneous brain activity at much higher frequencies that may contain neurobiologically
meaningful signals (Gong et al., 2021). Our study provides strong evidence of highly reliable
signals across higher slow-frequency bands, which are derived with the hierarchical frequency
band theory of neuronal oscillation system (Buzsaki & Draguhn, 2004). Spécifiquement, a spec-
trum of reliability increases was evident from slow bands to fast bands. This reflects greater
variability of the network measurements between subjects and less measurement variability
within subject between the higher and lower bands of the slow frequencies. En théorie, chaque
frequency band has an independent role in supporting brain function. Lower frequency bands
are thought to support more general or global computation with long-distance connections to
integrate specific or local computation, which are driven by higher slow bands based on
short-distance connections (Buzsáki, 2009). Our findings of high reliability (interindividual
differences) are perfectly consistent with this theory from a perspective of individual variability.
Previous findings have found that high-order associative (par exemple., default mode and cognitive con-
trol) networks are more reliable than the primary (par exemple., somatomotor and visual) réseaux
(Noble et al., 2019; Zuo & Xing, 2014; Zuo, Xu, & Milham, 2019b). A novel frequency-based
perspective on these network-level individual differences can be inspired directly by our
observations on the multiple bands.
Principle III: Optimize Topological Economy to Construct Network Connections at Individual Level
There is no gold standard on for human functional connectomes, leading to a plurality of
approaches for inferring and constructing brain network connections. Threshold-based
methods focus on the absolute strength of connectivity, retaining connections that are above
some user-defined threshold and oftentimes involve applying the same threshold to all
sujets. Although this approach mitigates potential biases in network metrics associated with
differences in network density, it may inadvertently also lead to decreased variability between
sujets. This is supported by our results showing that threshold-based methods yield low reli-
ability of network measurements. On the other hand, the human brain is a complex network
that is also near optimal in terms of connectional economy, balancing trade-offs of cost with
functionality (Bullmore & Sporns, 2012). In line with this view, certain classes of topology-
based methods for connection definition may hold promise for individualized network con-
struction. Spécifiquement, each individual brain optimizes its economic wiring in terms of cost and
efficiency, reaching a trade-off between minimizing costs and allowing the emergence of
adaptive topology. Our results demonstrate that such highly individualized functional connec-
tomes generated by the topology-based methods are more reliable than those generated by the
threshold-methods. This reflects the increase of individual differences in functional connec-
tomes attributing to the optimal wiring economics at individual level. The topological optimi-
zation also brings other benefits such as ensuring that a graph forms a single connected
component and preserving weak connections. En effet, there is increasing evidence supporting
the hypothesis that weak connections are neurobiologically meaningful and explain individual
Neurosciences en réseau
1098
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
differences in mind, behavior, and demographics as well as disorders (Santarnecchi, Galli,
Polizzotto, Rossi, & Rossi, 2014). Weak connections in a graph may be consistent across data-
sets and reproducible within the same individual over multiple scanning sessions and therefore
be reliable. Weak connections might also play nontrivial roles in transformed versions of
the original brain network, Par exemple, so-called “edge-based functional connectivity”
(Faskowitz, Esfahlani, Jo, Sporns, & Betzel, 2020). Among these topology-based methods,
MST is the simplest and most promising filtering method if computational efficiency is the pri-
ority. MST can obtain a graph with the same number of nodes and edges, and it is not sensitive
to scaling effects, because its structure only depends on the order rather than the absolute
values of the edges. Although MST loses some local network measurements due to the limited
number of edges, it has some other unique metrics that can be calculated (par exemple., leaf fraction,
tree hierarchy). A better alternative might be TMFG, which is computationally very efficient
and statistically robust, while the OMST and OTMFG are the most reliable choices by prior-
itizing significant individual differences.
Principle IV: Characterize Information Flow with Both Network Integration and Segregation Metrics
Intrinsic functional networks reflect the outcomes of communication processes and informa-
tion flows between pairs of brain regions. How the information and other signals propagate
between pairs of brain regions can be assayed using network neuroscientific metrics and is
essential to understanding normative connectome function and its variation in clinical settings.
While the ground truth of functional connectome remains unknown (and may not exist), net-
work models can help validate the imaging-based reconstructions of human functional net-
travaux (D. Bassett et al., 2020). From a perspective of individual differences, reliable network
measures are the basis of achieving valid ifNN measurements (Zuo, Xu, & Milham, 2019b).
Our findings indicated that both the brain network segregation and integration could be reli-
ably measured with functional connectomics using rfMRI by the optimized pipelines. At the
global level, measures of information integration, Par exemple, characteristic path length and
efficiency, were more reliable than those of information segregation, Par exemple, modularity
and clustering coefficient. Our results also revealed that measures of integration were more
stable across different scan sessions (c'est à dire., the test-retest) for an individual subject than the seg-
regation measurements, while interindividual variability are measured at the similar level for
both integration and segregation metrics. At the nodal level, mapping reliability of the network
measurements revealed interesting spatial patterns. Spécifiquement, we found that cortical areas
were generally associated with more reliable local measurements compared to subcortical
domaines. This may reflect different functional roles for human cortex and subcortex. Par exemple,
the differences in reliability of path-based metrics might reflect the fact that there are more
cortical within-community paths, while between-community paths are more common in sub-
cortex. Beyond this cortical-subcortical gradient, reliability of the nodal information flow also
fit the left-right asymmetry and dorsal-ventral as well as posterior-anterior gradient, implying
the potential validity of individual differences in information flow attributing to evolutionary,
genetic, and anatomical factors (Chen et al., 2013; Rakić, 2009). To facilitate the utility of reli-
able network integration and segregation metrics in ifNN, we integrated all the reliability
resources into an online platform for reliability queries on specific metrics of information flow
(https://ibraindata.com/research/ifNN).
Reproducibility and Generalizability
Both reproducibility and generalizability are cornerstones of modern sciences, and remain
challenging as a scientific research frontier (Munafo et al., 2017; Yarkoni, 2022). Dans ce
Neurosciences en réseau
1099
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
.
/
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
recherche, we adopt a big data approach by deeply sampling the parameters (more than 524k
parametric settings) of various steps in the network construction and analysis pipeline to sys-
tematically explore the reliability of functional brain network measurements. This provided
robust experimental evidence supporting four key principles that will foster optimal ifNN
research and application. These principles can serve as the basis for building guidelines on
the use of ifNN to map individual differences. Standard guidelines are essential for improve-
ments of reproducibility and generalizability in research practice, and our work provides basic
ressources, initiating such standardization in future network neuroscience. We note, cependant,
that while our approach was extensive, it was not exhaustive (likely impossible)—the analyt-
ical sampling procedure could miss many other existing choices. The processing decisions that
yield reliable connectomic measurements may yield the most reliable network statistics, mais
there may be another way to process data that yields overall a higher level of reliability in
network measures. Regarding the statistical benefits of our sampling analytics in the parametric
space of the ifNN pipelines, we discuss below the implications of the present research for
reproducible and generalizable network neuroscience.
Reliability and validity. Pipelines of generating highly reliable measurements are central to the
experimental design of studying individual differences (Matheson, 2019; Zuo, Xu, & Milham,
2019b). Given a statistical power, for a fixed sample size, experiments designed with more
reliable pipelines can detect bigger effects of interests. On the other side, to detect a fixed
effect size, experiments designed with more reliable pipelines would be more powerful or
logistically economical (par exemple., needless samples). This has very important implications on the
recent arguments about “big data versus small data” (Marek et al., 2022; Rosenberg & Finn,
2022; Tibon, Geerligs, & Campbell, 2022), which must take the reliability into account when
first designing an experiment (Gratton, Nelson, & Gordon, 2022), and has been increasingly
appreciated by the field of network neuroscience (Helwegen, Libedinsky, & van den Heuvel,
2023). From a perspective of experimental design, reliability is more straightforward to repro-
ducibility but validity is related to generalizability. Donc, we clarify that the measurement
reliability is not the final goal but the validity (Finn & Rosenberg, 2021; Noble et al., 2021),
which is not easily ready for a direct examination as a reliability assessment (Zuo, Xu, &
Milham, 2019b). The reliable pipeline we proposed produced biologically plausible findings
according to the four principles we discussed, likely reflecting its potential validity of measur-
ing individual differences in intrinsic brain functional organization. Validation on the use of
our proposed principles represents a promising arena for fostering future network neurosci-
ence studies such as personality (Hilger & Markett, 2021) or brain developmental charts
(Bethlehem et al., 2022), with potential novel fMRI paradigms (Elliott, Knodt, & Hariri,
2021b; Finn, Glerean, Hasson, & Vanderwal, 2022) or more precise neuroimaging technology
(Toi et al., 2022).
Reliability does not necessarily equate to but indeed provides an up bound of validity. Dans
some cases, increasing reliability may cause a decrease in validity, particularly if the sources of
reliability are not related to the underlying construct of interests. Par exemple, physiological
noise and head motion can be highly reliable as biological traits, but may not be involved in
the investigated cognitive processes. Previous studies have shown that head motion can intro-
duce artifacts into the fMRI data, which can affect the reliability of functional connectivity
measures (Power et al., 2014). En particulier, head motion may have a nonuniform impact
on different edge filtering methods and network metrics. Certain methods that rely on the
strength of functional connections, such as threshold-based approaches, may be more sensi-
tive to head motion artifacts than topology-based methods that focus on the overall structure of
the network. Measures that are highly reliable due to the inclusion of these contaminants may
Neurosciences en réseau
1100
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
.
/
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
not be valid indicators of the underlying construct. Cependant, head motion may not always be
purely noise and may contain some neurobiologically meaningful signals (Zeng et al., 2014;
Zhou et al., 2016). Donc, it is important to carefully consider the potential impact of head
motion when choosing an edge filtering method and interpreting the resulting functional
connectivity measures, as well as the trade-off between controlling for motion artifacts and
preserving potentially meaningful signals in the data. In the context of graph theory, bruit
can affect both the reliability and validity of graph metrics. Par exemple, noise in the data
can result in higher reliability of certain graph metrics, but this may not necessarily reflect
the true underlying network structure. This is because noise may lead to inflated
correlations between certain regions, resulting in overestimations of network connectivity
and thus higher reliability. Cependant, these measurements may not be valid indicators of the
true network structure and may not accurately reflect the underlying cognitive processes
being studied. On the other hand, the removal of noise may lead to decreased reliability,
but may improve the validity of the measurement by reducing the influence of unrelated
sources of variance. The optimal choices for maximizing reliability in our study may also
have implications for interpretability and generalizability. Par exemple, the inclusion of
subcortical structures in the parcellation scheme may increase the interpretability of the
résultats, as these structures play a key role in the functional organization of the brain. Sur
the other hand, the choice of connectivity transformation and edge weighting may have
implications for the generalizability of the results, as different methods may produce
different results depending on the specific characteristics of the data. Further research is
warranted to fully understand the consequences of these choices on interpretability,
generalizability, and other aspects of the measurement process.
The rfMRI datasets minimally preprocessed by the HCP pipeline are employed for our
study while many different pipelines are available for rfMRI data preprocessing (see a list of
pipelines in Xu et al., 2015). These different pipelines vary across parametric settings and
orders of various steps of preprocessing, and thus can have different impacts on the reliability
of measuring spontaneous brain activity (Li et al., 2022). Donc, it is very important to
validate whether the present findings are reproducible under another preprocessing pipeline.
Accordingly, we repeated our analyses by leveraging another widely accepted preprocessing
pipeline, fMRIPrep (Esteban et al., 2019). As documented in the Supporting Information, le
major findings supporting the principal guidelines are reproducible while the measurement
reliability derived with fMRIPrep are generally lower than those with the HCP pipeline.
Various within-pipeline parametric settings also as exist other choices not sampled by our
experimental design but remain potentials for further investigation. Par exemple, edge filtering
methods are commonly used for identification and retain only the most important edges in a
graph, based on criteria such as statistical significance or functional relevance. Cependant, ce
approach has the potential to introduce bias and subjectivity in the selection process, et
may not fully capture the higher order structure of a network system. Algebraic topology,
as demonstrated by Giusti, Ghrist, and Bassett (2016) and other recent studies, offers a prom-
ising alternative for high-order edge filtering. By representing relationships between objects as
higher dimensional simplices instead of edges, simplicial complexes can characterize polya-
dic interactions and capture more nuanced aspects of complex network organization. With
the increasing availability of computational tools for the application of algebraic topology to
real data, this framework has the potential to surpass graph theory in understanding the com-
plexities of neural systems.
Other fMRI paradigms. The guidelines we proposed for rfMRI-based network neuroscience
may also provide insights for network neuroscience computation by leveraging task-fMRI or
Neurosciences en réseau
1101
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
.
t
/
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
movie-fMRI. These two paradigms have gained increasing attention in recent years as a means
of measuring functional connectomes (Cole, Bassett, Power, Plus courageux, & Petersen, 2014; Cole
et coll., 2013; Finn et al., 2022). The reliability and predictive power of these measures have
been the subject of a number of studies. Par exemple, a study by Gao et al. (2020) found that
the reliability of movie-fMRI connectivity was influenced by the complexity and duration of
the movie stimulus, with more complex and longer stimuli resulting in higher test-retest reli-
ability. A recent work demonstrated that these improvements of prediction with task-based
functional connectivity were due largely to specific task design, highlighting the importance
of task design in probing valid brain–behavior associations (Zhao et al., 2023). Such task-
based fMRI connectivity methods can detect developmentally related individual differences
in brain and behavior (Kardan et al., 2022). These findings support the notion that task-fMRI
and movie-fMRI can produce more reliable connectivity measures with greater predictive
power for individual differences in cognitive and mental health measures compared to rfMRI,
particularly for tasks and stimuli that elicit strong and sustained activation. According to the
relationships among rest, task, and movie as well as other naturalistic states of the human brain
as a systems entity (Cole, Ito, Bassett, & Schultz, 2016; Finn, 2021; McCormick, Arnemann,
Ito, Hanson, & Cole, 2022), we speculate that the four principles are generalizable to func-
tional network neuroscience based on these nonrest brain states. Cependant, we note that more
research is warranted to fully understand the underlying mechanisms and generalizability of
these findings to different task and movie paradigms as well as their translational applications
(Eickhoff, Milham, & Vanderwal, 2020; Finn & Rosenberg, 2021).
Psychometric and other limitations. When studying the individual differences in brain and
behavior, one must consider the specific research aims and the underlying assumptions of
the study for addressing the reliability trade-off between maximizing between-individual var-
iability and minimizing within-individual variability. Par exemple, if the goal of the study is to
identify population-level age-related trajectories, it may be more important to prioritize max-
imizing between-individual variability in order to capture the full range of individual differ-
ences for a specific population. On the other hand, if the focus of the study is on assessing
within-subject changes over time (par exemple., the individual-level growth), it may be more important
to minimize within-individual variance in order to accurately capture developmental changes
in brain function. It is also important to consider the potential impacts of these trade-off deci-
sions on the validity of measurements of interest. Par exemple, if motion scrubbing or outlier
detection leads to the exclusion of a large number of subjects or time points, this may intro-
duce bias and reduce the generalizability of the results regarding the limited population diver-
sity or temporal stability. Careful consideration of these trade-offs is therefore essential in order
to ensure that the chosen approach is appropriate for the specific research aims and assump-
tions of the study. The reliability theory we proposed leads to a quantitative framework on the
measurement reliability, which delineates the anatomy of reliability changes as a function of
variability changes for both between and within individuals. Our demonstration of the utility of
this framework as a compass of variability (see Figure 2C) can guide the decision-making in
the experimental design as mentioned above.
The design of a psychometric study is normally recommended to recruit a group of partic-
ipants who are stable across the duration of investigation. The stability makes the interpretation
of within-subject variability straightforward as the subject-independent random noise, et le
reliability assessment are more precise; this is also why most psychometric studies were done
in adults although some in children (but with very short duration). Population diversity plays a
critical factor of assuring the generalizability in studying individual differences (UN. S. Vert
et coll., 2022; Ricard et al., 2023). This has been responded by the emerging population
Neurosciences en réseau
1102
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
neuroscience as a new stage of cognitive neuroscience to increase generalizability across pop-
ulations. (Falk et al., 2013; Paus, 2010). Psychometric studies are particularly required for pop-
ulation neuroscience due to the core aim of measuring individual differences in brain and
mind developmental during the life-span (Zuo et al., 2017, 2018). Future psychometric studies
on network neuroscience including those further validating the four principles developed by
the present work should be done for more diverse populations.
Summary and Conclusion
In this study we developed a novel framework to model reliability changes as a function of
changes of between-subject and within-subject variability. This framework was leveraged to
optimize network neuroscience computation of individual differences in human spontaneous
brain activity for test-retest reliability. We showed that reliable network neuroscience compu-
tation should define network nodes with whole-brain parcellations, derive the network edge
weights with spontaneous high-frequency slow-band oscillations, and construct the brain
networks using topology-based methods for multilevel and multimodal characteristics. Notre
findings highlight the importance of psychometric studies to understand the individual differ-
ences in spontaneous brain activity at systems level, and provide the resources and principles
of experimental design to guide future network neuroscience studies.
ACKNOWLEGDMENTS
We thank the Chinese Data-Sharing Warehouse for In-vivo Imaging Brain at National Basic
Science Data Center for informatics resources, the Research Program on Discipline Direction
Prediction and Technology Roadmap of China Association for Science and Technology, et
Apprentissage: Brains, Machines and Children from the Indiana University Office of the Vice Pres-
ident for Research Emerging Area of Research Initiative. The neuroimaging data were provided
by the HCP WU-Minn Consortium, which is funded by the 16 NIH institutes and centers that
support the NIH Blueprint for Neuroscience Research 1U54MH091657 (PIs: David Van Essen
and Kamil Ugurbil), the McDonnell Center for Systems Neuroscience at Washington Univer-
ville. The institutional Research Ethics Boards approved their HCP WU-Minn Consortium
études. Written informed consent was obtained from all participants.
INFORMATIONS À L'APPUI
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00315.
CONTRIBUTIONS DES AUTEURS
Chao Jiang: Conceptualisation; Conservation des données; Analyse formelle; Méthodologie; Ressources; Soft-
ware; Visualisation; Writing—Original draft; Rédaction—Révision & édition. Ye He: Conceptual-
ization; Rédaction—Révision & édition. Richard F. Betzel: Acquisition de financement; Écriture—Originale
brouillon; Rédaction—Révision & édition. Yin-Shan Wang: Conceptualisation; Rédaction—Révision &
édition. Xiu-Xia Xing: Conceptualisation; Ressources; Logiciel; Surveillance; Rédaction—Révision
& édition. Xi-Nian Zuo: Conceptualisation; Conservation des données; Analyse formelle; Funding acquisi-
tion; Enquête; Méthodologie; Gestion de projet; Ressources; Logiciel; Surveillance;
Validation; Visualisation; Writing—Original draft; Rédaction—Révision & édition.
INFORMATIONS SUR LE FINANCEMENT
Xi-Nian Zuo, The STI 2030 – Major Projects, Award ID: 2021ZD0200500.
Neurosciences en réseau
1103
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
t
.
/
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
RÉFÉRENCES
Achard, S., Salvador, R., Whitcher, B., Suckling, J., & Bullmore, E.
(2006). A resilient, low-frequency, small-world human brain
functional network with highly connected association cortical
hubs. Journal des neurosciences, 26(1), 63–72. https://est ce que je.org/10
.1523/JNEUROSCI.3874-05.2006, PubMed: 16399673
Aurich, N., Filho, J., da Silva, UN., & Franco, UN. (2015). Evaluating
the reliability of different preprocessing steps to estimate graph
theoretical measures in resting state fMRI data. Frontiers in Neu-
roscience, 9, 48. https://doi.org/10.3389/fnins.2015.00048,
PubMed: 25745384
Bassett, D., Bullmore, E., Meyer-Lindenberg, UN., Apud, J., Weinberger,
D., & Coppola, R.. (2009). Cognitive fitness of cost-efficient brain
functional networks. Proceedings of the National Academy of
les sciences, 106(28), 11747–11752. https://doi.org/10.1073/pnas
.0903641106, PubMed: 19564605
Bassett, D., Cullen, K., Eickhoff, S., Farah, M., Goda, Y., Haggard,
P., … Ueda, H. (2020). Reflections on the past two decades of
neuroscience. Nature Revues Neurosciences, 21(10), 524–534.
https://doi.org/10.1038/s41583-020-0363-6, PubMed: 32879507
Bassett, D. S., Bullmore, E., Verchinski, B. UN., Mattay, V. S.,
Weinberger, D. R., & Meyer-Lindenberg, UN. (2008). Hierarchical
organization of human cortical networks in health and schizo-
phrenia. Journal des neurosciences, 28(37), 9239–9248. https://
doi.org/10.1523/ JNEUROSCI.1929-08.2008, PubMed:
18784304
Bethlehem, R., Seidlitz, J., Blanc, S., Vogel, J., Anderson, K.,
Adamson, C., … Alexander-Bloch, UN. F. (2022). Brain charts for
the human lifespan. Nature, 604(7906), 525–533. https://doi.org
/10.1038/s41586-022-04554-y, PubMed: 35388223
Biswal, B., Mennes, M., Zuo, X.-N., Gohel, S., Kelly, C., Forgeron, S.,
… Milham, M.. (2010). Toward discovery science of human brain
fonction. Actes de l'Académie nationale des sciences,
107(10), 4734–4739. https://doi.org/10.1073/pnas.0911855107,
PubMed: 20176931
Biswal, B., Zerrin Yetkin, F., Haughton, V., & Hyde, J.. (1995). Func-
tional connectivity in the motor cortex of resting human brain
using echo-planar MRI. Magnetic Resonance in Medicine,
34(4), 537–541. https://doi.org/10.1002/mrm.1910340409,
PubMed: 8524021
Bostan, UN., & Strick, P.. (2018). The basal ganglia and the cerebellum:
Nodes in an integrated network. Nature Revues Neurosciences,
19(6), 338–350. https://doi.org/10.1038/s41583-018-0002-7,
PubMed: 29643480
Botvinik-Nezer, R., Holzmeister, F., Camerer, C., Dreber, UN., Huber,
J., Johannesson, M., … Schonberg, T. (2020). Variability in the
analysis of a single neuroimaging dataset by many teams. Nature,
582(7810), 84–88. https://doi.org/10.1038/s41586-020-2314-9,
PubMed: 32483374
Boubela, R., Kalcher, K., Huf, W., Kronnerwetter, C., Filzmoser, P.,
& Moser, E. (2013). Beyond noise: Using temporal ICA to extract
meaningful information from high-frequency fMRI signal fluctu-
ations during rest. Frontiers in Human Neuroscience, 7, 168.
https://doi.org/10.3389/fnhum.2013.00168, PubMed: 23641208
Bouttier, J., Di Francesco, P., & Guitter, E. (2003). Geodesic dis-
tance in planar graphs. Nuclear Physics B, 663(3), 535–567.
https://doi.org/10.1016/S0550-3213(03)00355-9
Brun, U., Plichta, M., Esslinger, C., Sauer, C., Haddad, L., Grimm,
O., … Meyer-Lindenberg, UN. (2012). Test-retest reliability of
resting-state connectivity network characteristics using fMRI and
graph theoretical measures. NeuroImage, 59(2), 1404–1412.
https://doi.org/10.1016/j.neuroimage.2011.08.044, PubMed:
21888983
Bridgeford, E., Wang, S., Wang, Z., Xu, T., Craddock, C., Dey, J., …
Vogelstein, J.. (2021). Eliminating accidental deviations to
minimize generalization error and maximize replicability: Appli-
cations in connectomics and genomics. PLoS Computational
Biology, 17(9), e1009279. https://doi.org/10.1371/journal.pcbi
.1009279, PubMed: 34529652
Bryce, N., Flournoy, J., Guassi Moreira, J., Rosen, M., Sambook, K.,
Mair, P., & McLaughlin, K. (2021). Brain parcellation selection: Un
overlooked decision point with meaningful effects on individual
differences in resting-state functional connectivity. NeuroImage,
243, 118487. https://doi.org/10.1016/j.neuroimage.2021.118487,
PubMed: 34419594
Bujang, M., & Baharum, N. (2017). A simplified guide to determi-
nation of sample size requirements for estimating the value of
intraclass correlation coefficient: A review. Archives of Orofacial
les sciences, 12(1), 1–11.
Bullmore, E., & Sporns, Ô. (2012). The economy of brain network
organization. Nature Revues Neurosciences, 13(5), 336–349.
https://doi.org/10.1038/nrn3214, PubMed: 22498897
Buzsáki, G., & Draguhn, UN. (2004). Neuronal oscillations in cortical
réseaux. Science, 304(5679), 1926–1929. https://est ce que je.org/10
.1126/science.1099745, PubMed: 15218136
Buzsáki, G. (2009). Rhythms of the brain. Presse universitaire d'Oxford.
Chen, C.-H., Fiecas, M., Gutiérrez, E., Panizzon, M., Eyler, L.,
Vuoksimaa, E., … Kremen, W. (2013). Genetic topography of
brain morphology. Proceedings of the National Academy of
les sciences, 110(42), 17089–17094. https://doi.org/10.1073/pnas
.1308091110, PubMed: 24082094
Christensen, UN., Kenett, Y., Aste, T., Silvia, P., & Kwapil, T. (2018).
Network structure of the wisconsin schizotypy scales-short
forms: Examining psychometric network filtering approaches.
Behavior Research Methods, 50(6), 2531–2550. https://doi.org
/10.3758/s13428-018-1032-9, PubMed: 29520631
Coalson, T., Van Essen, D., & Glasser, M.. (2018). The impact of
traditional neuroimaging methods on the spatial localization of
cortical areas. Actes de l'Académie nationale des sciences,
11 5 ( 2 7 ) , E 6 3 5 6 – E 6 3 6 5 . h t t p s : / / d o i . o rg / 1 0 . 1 0 7 3 / p n a s
.1801582115, PubMed: 29925602
Cole, M.. W., Bassett, D. S., Power, J.. D., Plus courageux, T. S., & Petersen,
S. E. (2014). Intrinsic and task-evoked network architectures of
the human brain. Neurone, 83(1), 238–251. https://est ce que je.org/10
.1016/j.neuron.2014.05.014, PubMed: 24991964
Cole, M.. W., Ito, T., Bassett, D. S., & Schultz, D. H. (2016). Activity
flow over resting-state networks shapes cognitive task activa-
tion. Neurosciences naturelles, 19(12), 1718–1726. https://doi.org
/10.1038/nn.4406, PubMed: 27723746
Cole, M.. W., Reynolds, J.. R., Power, J.. D., Repovs, G., Anticevic, UN.,
& Plus courageux, T. S. (2013). Multi-task connectivity reveals flexible hubs
for adaptive task control. Neurosciences naturelles, 16(9), 1348–1355.
https://doi.org/10.1038/nn.3470, PubMed: 23892552
Neurosciences en réseau
1104
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
Cordes, D., Haughton, V., Arfanakis, K., Carew, J., Turski, P., Moritz,
C., … Meyerand, M.. (2001). Frequencies contributing to functional
connectivity in the cerebral cortex in “resting-state” data. Américain
Journal de neuroradiologie, 22(7), 1326–1333. PubMed: 11498421
Craddock, R., Jbabdi, S., Yan, C.-G., Vogelstein, J., Castellanos, F.,
Di Martino, UN., … Milham, M.. (2013). Imaging human connec-
tomes at the macroscale. Nature Methods, 10(6), 524–539.
https://doi.org/10.1038/nmeth.2482, PubMed: 23722212
Desikan, R., Ségonne, F., Fischl, B., Quinn, B., Dickerson, B.,
Blacker, D., … Killiany, R.. (2006). An automated labeling system
for subdividing the human cerebral cortex on MRI scans into
gyral based regions of interest. NeuroImage, 31(3), 968–980.
https://doi.org/10.1016/j.neuroimage.2006.01.021, PubMed:
16530430
De Vico Fallani, F., Latora, V., & Chavez, M.. (2017). A topological
criterion for filtering information in complex brain networks.
Biologie computationnelle PLoS, 13(1), e1005305. https://doi.org
/10.1371/journal.pcbi.1005305, PubMed: 28076353
Dimitriadis, S., Antonakakis, M., Simos, P., Fletcher, J., & Papanicolaou,
UN. (2017). Data-driven topological filtering based on orthogonal
minimal spanning trees: Application to multigroup magnetoen-
cephalography resting-state connectivity. Brain Connectivity,
7(10), 661–670. https://doi.org/10.1089/ brain.2017.0512,
PubMed: 28891322
Dimitriadis, S. JE., Laskaris, N. UN., Del Rio-Portilla, Y., & Koudounis,
G. C. (2009). Characterizing dynamic functional connectivity
across sleep stages from EEG. Brain Topography, 22(2), 119–133.
https://doi.org/10.1007/s10548-008-0071-4, PubMed: 19005749
Dubois, J., & Adolphs, R.. (2016). Building a science of individual
differences from fMRI. Tendances des sciences cognitives, 20(6),
425–443. https://doi.org/10.1016/j.tics.2016.03.014, PubMed:
27138646
Eickhoff, S. B., Milham, M., & Vanderwal, T. (2020). Towards clinical
applications of movie fMRI. NeuroImage, 217, 116860. https://est ce que je
.org/10.1016/j.neuroimage.2020.116860, PubMed: 32376301
Elliott, M., Knodt, UN., Caspi, UN., Moffitt, T., & Hariri, UN. (2021un).
Need for psychometric theory in neuroscience research and
entraînement: Reply to Kragel et al. (2021). Sciences psychologiques,
32(4), 627–629. https://doi.org/10.1177/0956797621996665,
PubMed: 33685291
Elliott, M., Knodt, UN., & Hariri, UN. (2021b). Striving toward transla-
tion: Strategies for reliable fMRI measurement. Trends in Cogni-
tive Sciences, 25(9), 776–787. https://doi.org/10.1016/j.tics.2021
.05.008, PubMed: 34134933
Esteban, O., Markiewicz, C., Blair, R., Moodie, C., Isik, UN.,
Erramuzpe, UN., … Gorgolewski, K. (2019). fMRIPrep: A robust
preprocessing pipeline for functional MRI. Nature Methods,
16(1), 111–116. https://doi.org/10.1038/s41592-018-0235-4,
PubMed: 30532080, PubMed: 16089598
Estrada, E., & Higham, D. J.. (2010). Network properties revealed
through matrix functions. SIAM Review, 52(4), 696–714. https://
doi.org/10.1137/090761070
Estrada, E., & Rodriguez-Velazquez, J.. UN. (2005). Subgraph centrality
in complex networks. Physical Review E, 71(5), 056103. https://est ce que je
.org/10.1103/PhysRevE.71.056103, PubMed: 16089598
Falk, E. B., Hyde, L. W., Mitchell, C., Faul, J., González, R., Heitzeg,
M.. M., … Schulenberg, J.. (2013). What is a representative brain?
Neuroscience meets population science. Actes du
Académie nationale des sciences, 110(44), 17615–17622. https://
doi.org/10.1073/pnas.1310134110, PubMed: 24151336
Fan, L., Li, H., Zhuo, J., Zhang, Y., Wang, J., Chen, L., … Jiang, T.
(2016). The Human Brainnetome Atlas: A new brain atlas based
on connectional architecture. Cortex cérébral, 26(8), 3508–3526.
https://doi.org/10.1093/cercor/bhw157, PubMed: 27230218
Faskowitz, J., Esfahlani, F., Jo, Y., Sporns, O., & Betzel, R.. (2020).
Edge-centric functional network representations of human cere-
bral cortex reveal overlapping system-level architecture. Nature
Neurosciences, 23(12), 1644–1654. https://est ce que je.org/10.1038
/s41593-020-00719-y, PubMed: 33077948
Finn, E. (2021). Is it time to put rest to rest? Tendances cognitives
les sciences, 25(12), 1021–1032. https://doi.org/10.1016/j.tics
.2021.09.005, PubMed: 34625348
Finn, E., Glerean, E., Hasson, U., & Vanderwal, T. (2022). Natural-
istic imaging: The use of ecologically valid conditions to study
brain function. NeuroImage, 247, 118776. https://est ce que je.org/10
.1016/j.neuroimage.2021.118776, PubMed: 34864153
Finn, E., & Rosenberg, M.. (2021). Beyond fingerprinting: Choosing
predictive connectomes over reliable connectomes. Neuro-
Image, 239, 118254. https://doi.org/10.1016/j.neuroimage.2021
.118254, PubMed: 34118397
Fornito, UN., Zalesky, UN., & Breakspear, M.. (2015). The connec-
tomics of brain disorders. Nature Revues Neurosciences, 16(3),
159–172. https://doi.org/10.1038/nrn3901, PubMed: 25697159
Fox, M., & Raichle, M.. (2007). Spontaneous fluctuations in brain
activity observed with functional magnetic resonance imaging.
Nature Revues Neurosciences, 8(9), 700–711. https://est ce que je.org/10
.1038/nrn2201, PubMed: 17704812
Freeman, L. C. (1978). Centrality in social networks conceptual
clarification. Social Networks, 1(3), 215–239. https://est ce que je.org/10
.1016/0378-8733(78)90021-7
Gao, J., Chen, G., Wu, J., Wang, Y., Hu, Y., Xu, T., … Yang, Z.
(2020). Reliability map of individual differences reflected in
inter-subject correlation in naturalistic imaging. NeuroImage,
223, 117277. https://doi.org/10.1016/j.neuroimage.2020.117277,
PubMed: 32818614
Garrison, K. UN., Scheinost, D., Finn, E. S., Shen, X., & Constable,
R.. T. (2015). Le (dans)stability of functional brain network mea-
sures across thresholds. NeuroImage, 118, 651–661. https://est ce que je
.org/10.1016/j.neuroimage.2015.05.046, PubMed: 26021218
Ginestet, C. E., Nichols, T. E., Bullmore, E. T., & Simmons, UN.
(2011). Brain network analysis: Separating cost from topology
using cost-integration. PLoS One, 6(7), e21570. https://doi.org
/10.1371/journal.pone.0021570, PubMed: 21829437
Giusti, C., Ghrist, R., & Bassett, D. S. (2016). Two’s company, three
(ou plus) is a simplex. Journal of Computational Neuroscience,
41(1), 1–14. https://doi.org/10.1007/s10827-016-0608-6,
PubMed: 27287487
Glasser, M., Coalson, T., Robinson, E., Hacker, C., Harwell, J.,
Yacoub, E., … Van Essen, D. (2016). A multi-modal parcellation
of human cerebral cortex. Nature, 536(7615), 171–178. https://
doi.org/10.1038/nature18933, PubMed: 27437579
Glasser, M., Sotiropoulos, S., Wilson, J., Coalson, T., Fischl, B.,
Andersson, J., … Jenkinson, M.. (2013). The minimal preprocessing
pipelines for the Human Connectome Project. NeuroImage, 80,
105–124. https://doi.org/10.1016/j.neuroimage.2013.04.127,
PubMed: 23668970
Neurosciences en réseau
1105
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
Gong, Z.-Q., Gao, P., Jiang, C., Xing, X.-X., Dong, H.-M., Blanc, T.,
… Zuo, X.-N. (2021). DREAM: A toolbox to decode rhythms of
the brain system. Neuroinformatics, 19(3), 529–545. https://est ce que je
.org/10.1007/s12021-020-09500-9, PubMed: 33409718
Gratton, C., Nelson, S., & Gordon, E. (2022). Brain-behavior
correlations: Two paths toward reliability. Neurone, 110(9),
1446–1449. https://doi.org/10.1016/j.neuron.2022.04.018,
PubMed: 35512638
Vert, UN. S., Shen, X., Noble, S., Horien, C., Hahn, C. UN., Arora,
J., … Constable, R.. T. (2022). Brain–phenotype models fail for
individuals who defy sample stereotypes. Nature, 609(7925),
109–118. https://doi.org/10.1038/s41586-022-05118-w,
PubMed: 36002572
Vert, D., Marek, S., Gordon, E., Siegel, J., Gratton, C., Laumann,
T., … Dosenbach, N. (2020). Integrative and network-specific
connectivity of the basal ganglia and thalamus defined in indi-
viduals. Neurone, 105(4), 742–758. https://est ce que je.org/10.1016/j
.neuron.2019.11.012, PubMed: 31836321
Guo, H., Qin, M., Chen, J., Xu, Y., & Xiang, J.. (2017). Machine-
learning classifier for patients with major depressive disorder:
Multifeature approach based on a high-order minimum spanning
tree functional brain network. Computational and Mathematical
Methods in Medicine, 2017, 4820935. https://doi.org/10.1155
/2017/4820935, PubMed: 29387141
Hagmann, P., Kurant, M., Gigandet, X., Thiran, P., Wedeen, V.,
Meuli, R., & Thiran, J.-P. (2007). Mapping human whole-brain
structural networks with diffusion MRI. PLoS One, 2(7), e597.
https://doi.org/10.1371/journal.pone.0000597, PubMed:
17611629
Hallquist, M.. N., & Hillary, F. G. (2019). Graph theory approaches
to functional network organization in brain disorders: A critique
for a brave new small-world. Neurosciences en réseau, 3(1), 1–26.
https://doi.org/10.1162/netn_a_00054, PubMed: 30793071
Helwegen, K., Libedinsky, JE., & van den Heuvel, M.. P.. (2023).
Statistical power in network neuroscience. Tendances cognitives
les sciences, 27(3), 282–301 https://doi.org/10.1016/j.tics.2022.12
.011, PubMed: 36725422
Herting, M., Gautam, P., Chen, Z., Mezher, UN., & Vetter, N. (2018).
Test-retest reliability of longitudinal task-based fMRI: Implica-
tions for developmental studies. Developmental Cognitive Neu-
roscience, 33, 17–26. https://doi.org/10.1016/j.dcn.2017.07.001,
PubMed: 29158072
Hilger, K., & Markett, S. (2021). Personality network neuroscience:
Promises and challenges on the way toward a unifying frame-
work of individual variability. Neurosciences en réseau, 5(3),
631–645. https://doi.org/10.1162/netn_a_00198, PubMed:
34746620
Ji, J., Spronk, M., Kulkarni, K., Repovš, G., Anticevic, UN., & Cole, M..
(2019). Mapping the human brain’s cortical-subcortical func-
tional network organization. NeuroImage, 185, 35–57. https://
doi.org/10.1016/j.neuroimage.2018.10.006 , PubMed:
30291974
Kardan, O., Stier, UN. J., Cardenas-Iniguez, C., Schertz, K. E., Pruin,
J.. C., Deng, Y., … Rosenberg, M.. D. (2022). Differences in the
functional brain architecture of sustained attention and working
memory in youth and adults. PLoS Biology, 20(12), e3001938.
https://doi.org/10.1371/journal.pbio.3001938, PubMed:
36542658
Koo, T., & Li, M.. (2016). A guideline of selecting and reporting
intraclass correlation coefficients for reliability research. Journal
of Chiropractic Medicine, 15(2), 155–163. https://est ce que je.org/10
.1016/j.jcm.2016.02.012, PubMed: 27330520
Kraemer, H. (2014). The reliability of clinical diagnoses: State of the art.
Annual Review of Clinical Psychology, 10, 111–130. https://doi.org
/10.1146/annurev-clinpsy-032813-153739, PubMed: 24387235
Latora, V., & Marchiori, M.. (2001). Efficient behavior of small-world
réseaux. Physical Review Letters, 87(19), 198701. https://est ce que je
.org/10.1103/PhysRevLett.87.198701, PubMed: 11690461
Li, X., Ai, L., Giavasis, S., Jin, H., Feczko, E., Xu, T., … Milham, M.. P..
(2022). Moving beyond processing and analysis-related variation
in neuroscience. bioRxiv. https://doi.org/10.1101/2021.12.01
.470790
Liang, X., Wang, J., Yan, C., Shu, N., Xu, K., Gong, G., & Il, Oui.
(2012). Effects of different correlation metrics and preprocessing
factors on small-world brain functional networks: A resting-state
functional MRI study. PLoS One, 7(3), e32766. https://est ce que je.org/10
.1371/journal.pone.0032766, PubMed: 22412922
Mantegna, R.. (1999). Hierarchical structure in financial markets.
European Physical Journal B, 11(1), 193–197. https://est ce que je.org/10
.1007/s100510050929
Marek, S., Siegel, J., Gordon, E., Raut, R., Gratton, C., Newbold, D., …
Dosenbach, N. (2018). Spatial and temporal organization of the
individual human cerebellum. Neurone, 100(4), 977–993. https://
doi.org/10.1016/j.neuron.2018.10.010, PubMed: 30473014
Marek, S., Tervo-Clemmens, B., Calabro, F., Montez, D., Kay, B.,
H a t o u m , UN . , … Dose nb ach , N.
( 20 22 ). Re pro du cib le
brain-wide association studies require thousands of individuals.
Nature, 603(7902), 654–660. https://doi.org/10.1038/s41586
-022-04492-9, PubMed: 35296861
Massara, G., Di Matteo, T., & Aste, T. (2016). Network filtering for
big data: Triangulated maximally filtered graph. Journal de
Complex Networks, 5(2), 161–178. https://est ce que je.org/10.1093
/comnet/cnw015
Matheson, G. (2019). We need to talk about reliability: Making
better use of test-retest studies for study design and interpretation.
PeerJ, 2019(5), e6918. https://doi.org/10.7717/peerj.6918,
PubMed: 31179173
Matthieu, P., & Hampshire, UN. (2016). Clinical concepts emerging
from fMRI functional connectomics. Neurone, 91(3), 511–528.
https://doi.org/10.1016/j.neuron.2016.07.031, PubMed:
27497220
McCormick, E. M., Arnemann, K. L., Ito, T., Hanson, S. J., & Cole,
M.. W. (2022). Latent functional connectivity underlying multiple
brain states. Neurosciences en réseau, 6(2), 570–590. https://est ce que je
.org/10.1162/netn_a_00234, PubMed: 35733420
Meier, J., Tewarie, P., & Van Mieghem, P.. (2015). The union of
shortest path trees of functional brain networks. Brain Connec-
tivité, 5(9), 575–581. https://doi.org/10.1089/brain.2014.0330,
PubMed: 26027712
Micheloyannis, S., Pachou, E., Stam, C. J., Breakspear, M., Bitsios,
P., Vourkas, M., … Zervakis, M.. (2006). Small-world networks
and disturbed functional connectivity in schizophrenia. Schizo-
phrenia Research, 87(1–3), 60–66. https://est ce que je.org/10.1016/j
.schres.2006.06.028, PubMed: 16875801
Milham, M., Vogelstein, J., & Xu, T. (2021). Removing the reliability
bottleneck in functional magnetic resonance imaging research to
Neurosciences en réseau
1106
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
achieve clinical utility. JAMA Psychiatry, 78(6), 587–588. https://
doi.org/10.1001/jamapsychiatry.2020.4272, PubMed: 33439234
Milo, R., Shen-Orr, S., Itzkovitz, S., Kashtan, N., Chklovskii, D., &
Alon, U. (2002). Network motifs: Simple building blocks of com-
plex networks. Science, 298(5594), 824–827. https://est ce que je.org/10
.1126/science.298.5594.824, PubMed: 12399590
Munafo, M., Nosek, B., Bishop, D., Button, K., Chambers, C.,
Percie Du Sert, N., … Ioannidis, J.. (2017). A manifesto for repro-
ducible science. Nature Human Behaviour, 1(1), 0021. https://
doi.org/10.1038/s41562-016-0021, PubMed: 33954258
Newman, M.. E. (2003). The structure and function of complex
réseaux. SIAM Review, 45(2), 167–256. https://est ce que je.org/10
.1137/S003614450342480
Newman, M.. E. (2004). Fast algorithm for detecting community
structure in networks. Physical Review E, 69(6), 066133. https://
doi.org/10.1103/PhysRevE.69.066133, PubMed: 15244693
Newman, M.. E. (2008). The mathematics of networks. The New
Palgrave Encyclopedia of Economics, 2(2008), 1–12. https://est ce que je
.org/10.1057/978-1-349-95121-5_2565-1
Noble, S., Scheinost, D., & Todd Constable, R.. (2019). A decade of
test-retest reliability of functional connectivity: A systematic
review and meta-analysis. NeuroImage, 203, 116157. https://est ce que je
.org/10.1016/j.neuroimage.2019.116157, PubMed: 31494250
Noble, S., Scheinost, D., & Todd Constable, R.. (2021). A guide to
the measurement and interpretation of fmri test-retest reliability.
Current Opinion in Behavioral Sciences, 40, 27–32. https://est ce que je
.org/10.1016/j.cobeha.2020.12.012, PubMed: 33585666
Otte, W., van Diessen, E., Paul, S., Ramaswamy, R., Subramanyam
Rallabandi, V., Stam, C., & Roy, P.. (2015). Aging alterations in
whole-brain networks during adulthood mapped with the min-
imum spanning tree indices: The interplay of density, connec-
tivity cost and life-time trajectory. NeuroImage, 109, 171–189.
https://doi.org/10.1016/j.neuroimage.2015.01.011, PubMed:
25585021
Page, L., Brin, S., Motwani, R., & Winograd, T. (1999). The PageRank
citation ranking: Bringing order to the web (Technologie. représentant). Stanford,
California: Stanford InfoLab.
Pastor-Satorras, R., Vázquez, UN., & Vespignani, UN. (2001). Dynam-
ical and correlation properties of the internet. Physical Review
Letters, 87(25), 258701. https://doi.org/10.1103/PhysRevLett.87
.258701, PubMed: 11736611
Paus, T. (2010). Population neuroscience: Why and how. Human
Brain Mapping, 31(6), 891–903. https://doi.org/10.1002/ hbm
.21069, PubMed: 20496380
Penttonen, M., & Buzsáki, G. (2003). Natural logarithmic relation-
ship between brain oscillators. Thalamus and Related Systems,
2(2), 145–152. https://doi.org/10.1016/S1472-9288(03)00007-4
Personne, L. (2018). Understanding emotion with brain networks.
Current Opinion in Behavioral Sciences, 19, 19–25. https://est ce que je
.org/10.1016/j.cobeha.2017.09.005, PubMed: 29915794
Power, J.. D., Mitra, UN., Laumann, T. O., Snyder, UN. Z., Schlaggar,
B. L., & Petersen, S. E. (2014). Methods to detect, characterize,
and remove motion artifact in resting state fMRI. NeuroImage,
84(Supplément. C), 320–341. https://doi.org/10.1016/j.neuroimage
.2013.08.048, PubMed: 23994314
Rakić, P.. (2009). Evolution of the neocortex: A perspective from
developmental biology. Nature Revues Neurosciences, 10(10),
724–735. https://doi.org/10.1038/nrn2719, PubMed: 19763105
Ricard, J., Parker, T., Dhamala, E., Kwasa, J., Allsop, UN., & Holmes,
UN. (2023). Confronting racially exclusionary practices in the
acquisition and analyses of neuroimaging data. Nature Neurosci-
ence, 26(1), 4–11. https://doi.org/10.1038/s41593-022-01218-y,
PubMed: 36564545
Rosenberg, M.. D., & Finn, E. S. (2022). How to establish robust
brain-behavior relationships without thousands of individuals.
Neurosciences naturelles, 25(7), 835–837. https://est ce que je.org/10.1038
/s41593-022-01110-9, PubMed: 35710985
Rubinov, M., & Sporns, Ô. (2010). Complex network measures of
brain connectivity: Uses and interpretations. NeuroImage, 52(3),
1059–1069. https://doi.org/10.1016/j.neuroimage.2009.10.003,
PubMed: 19819337
Rubinov, M., Sporns, O., van Leeuwen, C., & Breakspear, M..
(2009). Symbiotic relationship between brain structure and
dynamics. BMC Neuroscience, 10, 55. https://doi.org/10.1186
/1471-2202-10-55, PubMed: 19486538
Salvador, R., Suckling, J., Coleman, M., Pickard, J., Menon, D., &
Bullmore, E. (2005). Neurophysiological architecture of func-
tional magnetic resonance images of human brain. Cérébral
Cortex, 15(9), 1332–2342. https://doi.org/10.1093/cercor/bhi016,
PubMed: 15635061
Santarnecchi, E., Galli, G., Polizzotto, N., Rossi, UN., & Rossi, S.
(2014). Efficiency of weak brain connections support general
cognitive functioning. Human Brain Mapping, 35(9), 4566–4582.
https://doi.org/10.1002/hbm.22495, PubMed: 24585433
Schaefer, UN., Kong, R., Gordon, E., Laumann, T., Zuo, X.-N.,
Holmes, UN., … Yeo, B. (2018). Local-global parcellation of the
human cerebral cortex from intrinsic functional connectivity
IRM. Cortex cérébral, 28(9), 3095–3114. https://est ce que je.org/10
.1093/cercor/bhx179, PubMed: 28981612
Forgeron, S., Beckmann, C., Andersson, J., Auerbach, E., Bijsterbosch, J.,
Douaud, G., … Glasser, M.. (2013). Resting-state fMRI in the Human
Connectome Project. NeuroImage, 80, 144–168. https://est ce que je.org/10
.1016/j.neuroimage.2013.05.039, PubMed: 23702415
Song, W.-M., Di Matteo, T., & Aste, T. (2012). Hierarchical informa-
tion clustering by means of topologically embedded graphs.
PLoS One, 7(3), e31929. https://doi.org/10.1371/journal.pone
.0031929, PubMed: 22427814
Sporns, Ô. (2013un). Making sense of brain network data. Nature
Methods, 10(6), 491–493. https://doi.org/10.1038/nmeth.2485,
PubMed: 23722207
Sporns, Ô. (2013b). Network attributes for segregation and integra-
tion in the human brain. Opinion actuelle en neurobiologie, 23(2),
162–171. https://doi.org/10.1016/j.conb.2012.11.015, PubMed:
23294553
Stam, C., Jones, B., Nolte, G., Breakspear, M., & Scheltens, P..
(2007). Small-world networks and functional connectivity in
Alzheimer’s disease. Cortex cérébral, 17(1), 92–99. https://est ce que je
.org/10.1093/cercor/bhj127, PubMed: 16452642
Termenon, M., Jaillard, UN., Delon-Martin, C., & Achard, S. (2016).
Reliability of graph analysis of resting state fMRI using test-retest
dataset from the Human Connectome Project. NeuroImage, 142,
172–187. https://doi.org/10.1016/j.neuroimage.2016.05.062,
PubMed: 27282475
Thomas Yeo, B., Krienen, F., Sepulcre, J., Sabuncu, M., Lashkari, D.,
Hollinshead, M., … Buckner, R.. (2011). The organization of the
human cerebral cortex estimated by intrinsic functional
Neurosciences en réseau
1107
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3
Optimal network neuroscience computation pipelines
connectivité. Journal de neurophysiologie, 106(3), 1125–1165.
https://est ce que je.org/10.1152/jn.00338.2011, PubMed: 21653723
Tibon, R., Geerligs, L., & Campbell, K. (2022). Bridging the big
(data) gap: Levels of control in small- and large-scale cognitive
neuroscience research. Trends in Neurosciences, 45(7), 507–
516. https://doi.org/10.1016/j.tins.2022.03.011, PubMed:
35469691
Toi, P., Jang, H., Min, K., Kim, S.-P., Lee, S.-K., Lee, J., … Park, J.-Y.
(2022). In vivo direct imaging of neuronal activity at high tempo-
rospatial resolution. Science, 378(6616), 160–168. https://doi.org
/10.1126/science.abh4340, PubMed: 36227975
Tumminello, M., Aste, T., Di Matteo, T., & Mantegna, R..
for filtering information in complex systems.
(2005). A tool
Proceedings of
the National Academy of Sciences, 102(30),
10421–10426. https://doi.org/10.1073/pnas.0500298102,
PubMed: 16027373
van den Heuvel, M.. P., de Lange, S. C., Zalesky, UN., Seguin, C., Yeo,
B. T., & Schmidt, R.. (2017). Proportional thresholding in
resting-state fMRI functional connectivity networks and conse-
quences for patient-control connectome studies: Issues and rec-
ommendations. NeuroImage, 152, 437–449. https://est ce que je.org/10
.1016/j.neuroimage.2017.02.005, PubMed: 28167349
van Nieuwenhuizen, D., Douw, L., Klein, M., Peerdeman, S.,
Heimans, J., Reijneveld, J., … Hillebrand, UN. (2018). Cognitive
functioning and functional brain networks in postoperative
WHO grade I meningioma patients. Journal of Neuro-Oncology,
140(3), 605–613. https://doi.org/10.1007/s11060-018-2987-1,
PubMed: 30219943
Wang, J., Wang, L., Zang, Y., Lequel, H., Tang, H., Gong, Q., … He, Oui.
(2009). Parcellation-dependent small-world brain functional networks:
A resting-state fMRI study. Human Brain Mapping, 30(5), 1511–1523.
https://doi.org/10.1002/hbm.20623, PubMed: 18649353
Watts, D. J., & Strogatz, S. H. (1998). Collective dynamics of
‘small-world’ networks. Nature, 393(6684), 440–442. https://est ce que je
.org/10.1038/30918, PubMed: 9623998
Xing, X.-X., Xu, T., Jiang, C., Wang, Y., & Zuo, X.-N. (2022).
Connectome Computation System: 2015–2021 updates. Science
Bulletin, 67(5), 448–451. https://doi.org/10.1016/j.scib.2021.11
.021, PubMed: 36546160
Xing, X.-X., & Zuo, X.-N. (2018). The anatomy of reliability: A must
read for future human brain mapping. Science Bulletin, 63(24),
1606–1607. https://doi.org/10.1016/j.scib.2018.12.010,
PubMed: 36658850
Xu, T., Lequel, Z., Jiang, L., Xing, X.-X., & Zuo, X.-N. (2015). UN
connectome computation system for discovery science of brain.
Science Bulletin, 60(1), 86–95. https://doi.org/10.1007/s11434
-014-0698-3
Yarkoni, T. (2022). The generalizability crisis. Behavioral and Brain
les sciences, 45, e1. https://doi.org/10.1017/S0140525X20001685,
PubMed: 33342451
Zalesky, UN., Fornito, UN., Harding, JE., Cocchi, L., Yücel, M., Pantelis, C.,
& Bullmore, E. (2010). Whole-brain anatomical networks: Does the
choice of nodes matter? NeuroImage, 50(3), 970–983. https://est ce que je
.org/10.1016/j.neuroimage.2009.12.027, PubMed: 20035887
Zeng, L.-L., Wang, D., Fox, M., Sabuncu, M., Hu, D., Ge, M., …
Liu, H. (2014). Neurobiological basis of head motion in brain
imaging. Actes de l'Académie nationale des sciences,
111(16), 6058–6062. https://doi.org/10.1073/pnas.1317424111,
PubMed: 24711399
Zhao, W., Makowski, C., Hagler, D. J., Garavan, H. P., Thompson,
W. K., Vert, D. J., … Dale, UN. M.. (2023). Task fMRI paradigms
may capture more behaviorally relevant information than
resting-state functional connectivity. NeuroImage, 270, 119946.
https://doi.org/10.1016/j.neuroimage.2023.119946, PubMed:
36801369
Zhou, Y., Chen, J., Luo, Oui. L., Zheng, D., Rao, L.-L., Li, X., … Zuo,
X.-N. (2016). Genetic overlap between in-scanner head motion
and the default network connectivity. bioRxiv. https://est ce que je.org/10
.1101/087023
Zuo, X.-N., Biswal, B., & Poldrack, R.. (2019un). Editorial: Reliability
and reproducibility in functional connectomics. Frontiers in Neu-
roscience, 13, 117. https://doi.org/10.3389/fnins.2019.00117,
PubMed: 30842722
Zuo, X. N., Di Martino, UN., Kelly, C., Shehzad, Z. E., Gee, D. G.,
Klein, D. F., … Milham, M.. P.. (2010). The oscillating brain:
Complex and reliable. NeuroImage, 49(2), 1432–1445. https://
doi.org/10.1016/j.neuroimage.2009.09.037, PubMed:
19782143
Zuo, X.-N., Il, Y., Betzel, R., Colcombe, S., Sporns, O., & Milham,
M.. (2017). Human connectomics across the life span. Trends in
Cognitive Sciences, 21(1), 32–45. https://doi.org/10.1016/j.tics
.2016.10.005, PubMed: 27865786
Zuo, X.-N., Il, Y., Su, X., Hou, X.-H., Weng, X., & Li, Q. (2018).
Developmental population neuroscience: Emerging from
ICHBD. Science Bulletin, 63(6), 331–332. https://est ce que je.org/10
.1016/j.scib.2018.01.008, PubMed: 36658866
Zuo, X.-N., & Xing, X.-X. (2014). Test-retest reliabilities of
resting-state FMRI measurements in human brain functional con-
nectomics: A systems neuroscience perspective. Neurosciences
and Biobehavioral Reviews, 45, 100–118. https://est ce que je.org/10
.1016/j.neubiorev.2014.05.009, PubMed: 24875392
Zuo, X.-N., Xu, T., Jiang, L., Lequel, Z., Cao, X.-Y., Il, Y., … Milham,
M.. (2013). Toward reliable characterization of functional
homogeneity in the human brain: Preprocessing, scan duration,
imaging resolution and computational space. NeuroImage, 65,
374–386. https://doi.org/10.1016/j.neuroimage.2012.10.017,
PubMed: 23085497
Zuo, X.-N., Xu, T., & Milham, M.. (2019b). Harnessing reliability for
neuroscience research. Nature Human Behaviour, 3(8), 768–771.
https://doi.org/10.1038/s41562-019-0655-x, PubMed: 31253883
Neurosciences en réseau
1108
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
0
8
0
2
1
5
4
8
4
5
n
e
n
_
un
_
0
0
3
1
5
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
7
S
e
p
e
m
b
e
r
2
0
2
3