RECHERCHE
Functional connectome fingerprinting
across the lifespan
Frédéric St-Onge1,2
, Mohammadali Javanray1,2, Alexa Pichet Binette3,
Cherie Strikwerda-Brown2, Jordana Remz2, R.. Nathan Spreng2,4,5, Golia Shafiei5,
Bratislav Misic5, Étienne Vachon-Presseau6,7,8, and Sylvia Villeneuve1,2,5
1Integrated Program in Neuroscience, Faculty of Medicine, Université McGill, Montréal, Canada
2Research Center of the Douglas Mental Health University Institute, Montréal, Canada
3Clinical Memory Research Unit, Faculty of Medicine, Lund University, Lund, Sweden
4Department of Neurology and Neurosurgery, Institut neurologique de Montréal, Université McGill, Montréal, Canada
5McConnell Brain Imaging Centre, Institut neurologique de Montréal, Université McGill, Montréal, Canada
6Faculty of Dental Medicine and Oral Health Sciences, Université McGill, Montréal, Canada
7Department of Anesthesia, Faculty of Medicine, Université McGill, Montréal, Canada
8Alan Edwards Centre for Research on Pain (AECRP), Université McGill, Montréal, Canada
Mots clés: Functional connectome fingerprinting, IRMf, Lifespan
ABSTRAIT
Systematic changes have been observed in the functional architecture of the human brain with
advancing age. Cependant, functional connectivity (FC) is also a powerful feature to detect
unique “connectome fingerprints,” allowing identification of individuals among their peers.
Although fingerprinting has been robustly observed in samples of young adults, the reliability
of this approach has not been demonstrated across the lifespan. We applied the fingerprinting
framework to the Cambridge Centre for Ageing and Neuroscience cohort (n = 483 âgé 18 à
89 années). We found that individuals are “fingerprintable” (c'est à dire., identifiable) across independent
functional MRI scans throughout the lifespan. We observed a U-shape distribution in
the strength of “self-identifiability” (within-individual correlation across modalities), et
“others-identifiability” (between-individual correlation across modalities), with a decrease
from early adulthood into middle age, before improving in older age. FC edges contributing
to self-identifiability were not restricted to specific brain networks and were different
between individuals across the lifespan sample. Self-identifiability was additionally associated
with regional brain volume. These findings indicate that individual participant-level
identification is preserved across the lifespan despite the fact that its components are
changing nonlinearly.
RÉSUMÉ DE L'AUTEUR
Important interindividual differences are increasingly identified in functional connectivity
études. Cependant, most of the work to date has focused on samples of younger adults, ignoring
older individuals in which brains undergo significant transformation. Ici, we examine
individual-level patterns of functional connectivity—that is, connectome fingerprints—across
the adult lifespan. Our results suggest that connectome fingerprints can reliably predict
individuals from independent brain scans across the lifespan. En plus, régions
contributing to fingerprints differed from person to person, emphasizing the extent of
interindividual differences in the brain. Specific patterns of brain volume, associated with
normative age-related atrophy, may impact the identification of individual-specific functional
un accès ouvert
journal
Citation: St-Onge, F., Javanray, M.,
Pichet Binette, UN., Strikwerda-Brown,
C., Remz, J., Spreng, R.. N., Shafiei, G.,
Misic, B., Vachon-Presseau, É., &
Villeneuve, S. (2023). Functional
connectome fingerprinting across the
lifespan. Neurosciences en réseau, 7(3),
1206–1227. https://doi.org/10.1162/netn
_a_00320
EST CE QUE JE:
https://doi.org/10.1162/netn_a_00320
Informations complémentaires:
https://doi.org/10.1162/netn_a_00320;
https://github.com/villeneuvelab
/projects
Reçu: 1 Août 2022
Accepté: 24 Avril 2023
Intérêts concurrents: Les auteurs ont
a déclaré qu'aucun intérêt concurrent
exister.
Corresponding Authors:
Frédéric St-Onge
frederic.st-onge@mail.mcgill.ca
Sylvia Villeneuve
Sylvia.villeneuve@mcgill.ca
Éditeur de manipulation:
Alex Fornito
droits d'auteur: © 2023
Massachusetts Institute of Technology
Publié sous Creative Commons
Attribution 4.0 International
(CC PAR 4.0) Licence
La presse du MIT
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
un
_
0
0
3
2
0
p
d
/
t
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
connectivity patterns. Our findings highlight the importance of interindividual variability in
functional connectivity, regardless of participant age, when assessing individual differences
in brain function.
INTRODUCTION
The structural and functional organization of the brain is reliably consistent within species
because of strong genetic control of this biological system (Gómez-Robles et al., 2015). Dans
humans, cependant, substantial intraindividual variability has been found at a fine-grained level
(Amico & Goñi, 2018; Finn et al., 2015; Mansour et al., 2021; Mueller et al., 2013). An emerg-
ing body of evidence suggests that interindividual differences in brain connectomes are suffi-
cient to match brain scans and effectively identify individuals among large datasets of brain
images. These “signatures” or “fingerprints” (Finn et al., 2015, 2017) are stable over years
(Guo et al., 2012; Horien et al., 2019; Jalbrzikowski et al., 2020; Ousdal et al., 2020) et
between scan conditions (Finn et al., 2015, 2017; Vanderwal et al., 2017), and they are found
using other brain scanning modalities such as magnetoencephalography (da Silva Castanheira
et coll., 2021). Individual participant identifiability is observable in homogeneous samples of
young adults (Finn et al., 2015; Mueller et al., 2013), yet older adults have been relatively
neglected in the literature.
Cross-sectional studies comparing older and younger adult groups have revealed substantial
differences in functional connectivity (FC) (Chen et al., 2016; Setton et al., 2023; Zonneveld
et coll., 2019), raising questions about the reliability of FC fingerprinting in older adults. Our aims
were to (un) test the stability of fingerprint identification accuracy (c'est à dire., uniqueness of the connec-
tomes, which facilitates individual participant identification; Finn et al., 2015), (b) determine
self-identifiability (c'est à dire., a continuous variable measuring within-individual similarity across
independent observations; Amico & Goñi, 2018), et (c) characterize others-identifiability
(c'est à dire., a continuous variable measuring how similar an individual is relative to others; Amico
& Goñi, 2018). These aims were examined across the lifespan, spanning the full connectome,
as well as within and between large-scale networks, using functional magnetic resonance
imaging (IRMf). We then determined which functional connections between regions (c'est à dire.,
edges) reliably contributed to identification and how these patterns varied across the lifespan.
Enfin, we explored the association between identifiability and brain volume, a significant
predictor of participant age (Gonneaud et al., 2021). We derived fingerprint metrics using a pair
of fMRI conditions (resting state and sensorimotor task) from cognitively healthy adults across
the lifespan in the Cambridge Centre for Ageing and Neuroscience (Cam-CAN) cohort (n =
483; ages 18–89 years) (Shafto et al., 2014; Taylor et al., 2017). Our results indicate that
fingerprint identifiability is a reliable metric across the lifespan. We also show that self- et
others-identifiability measures had nonlinear distributions across the lifespan. Self- et d'autres-
identifiability was high in young adults, decreased into middle age, then increased again into
older adulthood. Elastic net models revealed that the fingerprinting methodology identifies
dominant individual-specific features of FC, reliably demarcating unique patterns for each
healthy adult at each decade of life. Enfin, self-identifiability, but not others-identifiability,
was associated with brain volume in regions known to atrophy in the context of normative
aging. Dans l'ensemble, the results suggest that intraindividual variability in the organization of the
cerveau humain, particularly in older adults, warrants consideration in parallel with normative
trajectories of age-related brain change.
Functional connectome
fingerprinting:
Framework used to test how unique
individuals’ patterns of functional
connectivity are.
Fingerprint identification accuracy:
If an individual’s self-identifiability is
higher than any others-identifiability,
fingerprint identification is accurate.
Self-identifiability:
Correlation (c'est à dire., similarité) entre
the pattern of functional connectivity
of a single individual across fMRI
conditions.
Others-identifiability:
Average of the correlation (c'est à dire.,
similarité) between the pattern of
functional connectivity of an
individual to all other individuals in
the cohort across fMRI conditions.
Edge:
One functional connectivity edge
represents the functional connection
between two nodes.
Elastic net:
Machine learning method used to
predict which combination of
predictors is associated with an
outcome.
Neurosciences en réseau
1207
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
un
_
0
0
3
2
0
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
RÉSULTATS
Analyses were performed on 483 individuals of the Cam-CAN cohort (Shafto et al., 2014;
Taylor et al., 2017) âgé 18 à 89 années. Participants were included if they had at least two
fMRI scans (rest and sensorimotor task modalities) passing quality control. Demographic infor-
mation is presented in Table 1. About half of our sample was composed of females. Most par-
ticipants were right-handed. The final sample comprised at least 50 individuals in each decade
de la vie, except individuals between 80 et 89 years of age, with only 34 included participants.
Fingerprint Identification Accuracy in a Lifespan Cohort
To test the stability of the fingerprint metrics of interest (fingerprint identification accuracy, soi-
identifiability, and others-identifiability; Finn et al., 2015), we correlated the FC pattern of a
given individual to their own FC pattern across Rest and Task conditions (self-identifiability)
and to the FC pattern of all other individuals (others-identifiability; Amico & Goñi, 2018). Si le
self-identifiability correlation coefficient was stronger than any of the others-identifiability
correlation coefficient, then the participant was identified as having a unique signature (finger-
print identification accuracy). This method is illustrated in Figure 1A. Edges used in the iden-
tification paradigm and the rest of the analyses comprised the whole-brain connectome, edges
within a given network (within-network edges), and edges between a given network and all
other network nodes (between-network edges). We used the Schaefer parcellation with 400
nodes (Schaefer et al., 2018) and Yeo’s seven-network solution (Yeo et al., 2011) to derive the
FC and to compute the fingerprint metrics. Results were also replicated using the Power par-
cellation (264 nodes; Supplementary Figure 3A; Power et al., 2011).
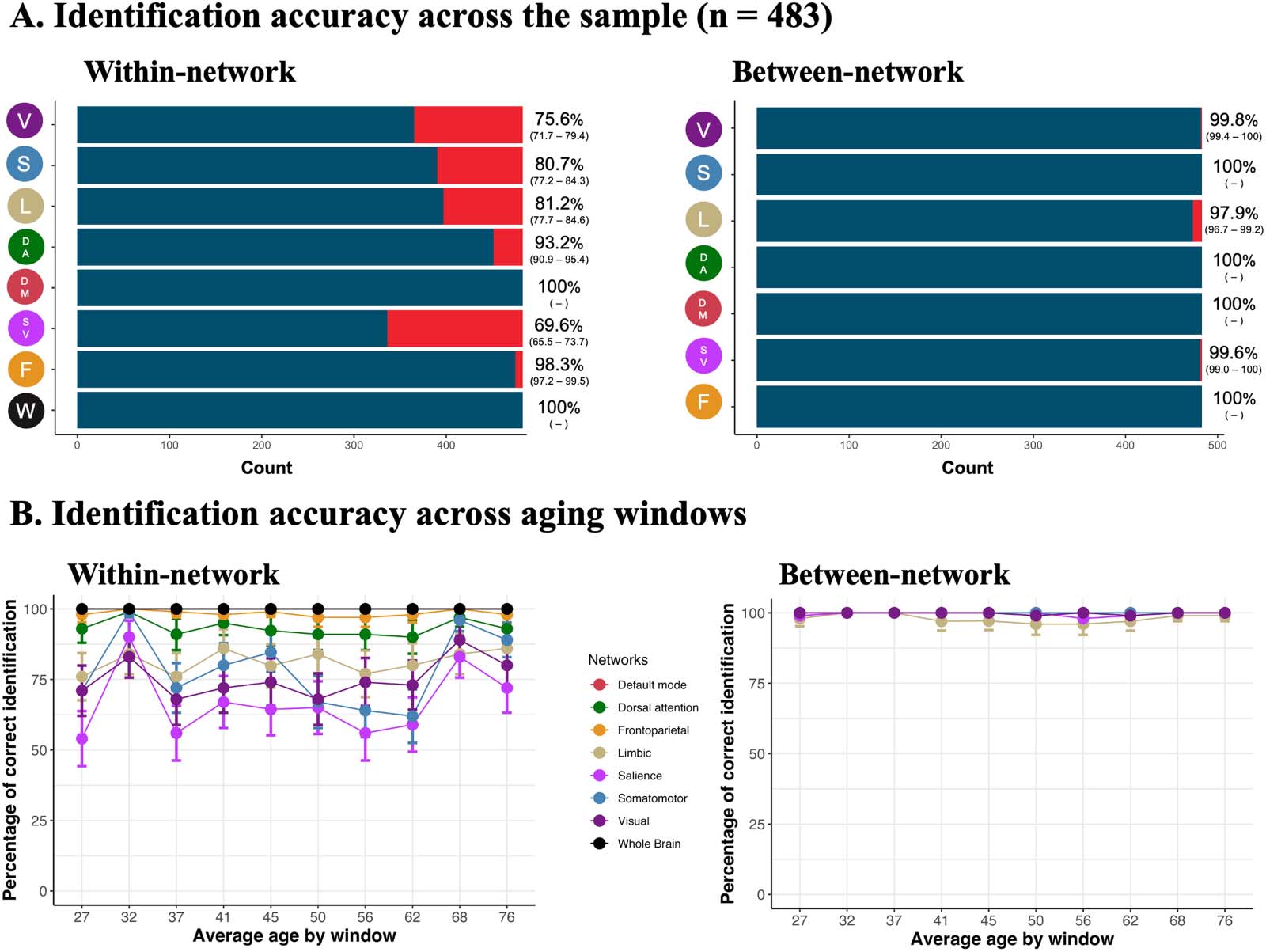
Across the full sample, we found high rates—up to 100% in some networks—of fingerprint
identification (Figure 2A). McNemar tests indicated that identification was increased when
using between-network edges (compared with within-network edges; Supplementary Table 1)
and when using higher associative cortices (default, frontoparietal, and dorsal attention com-
pared with visual, sensorimotor, and limbic; Supplementary Table 2). Using between-network
edges, we were able to achieve 100% fingerprint identification accuracy in the somatomotor,
dorsal attention, default, and frontoparietal networks. Using whole-brain connectome and
within-network edges in the default network, we achieved 100% fingerprint identification
accuracy.
To determine how stable identifiability was throughout the lifespan, we adapted a between-
participant sliding-window approach (Figure 1C; Váša et al., 2018). Briefly, participants were
ordered by age, and slices of overlapping participants were selected to create groups of par-
ticipants across different ages of the lifespan. Using this approach, identifiability was stable
throughout the lifespan across networks (Figure 2B, Supplementary Figure 1; approche
described in Figure 1C). Enfin, employing McNemar tests, identification rates using within-
network edges were similar for partial correlation-derived FC compared with product–moment
correlation-derived FC, but were superior when using between-network edges (Supplementary
Tables 3 et 4).
We also tested whether using a random collection of nodes selected throughout the brain
would yield comparable fingerprint identification accuracy rather than using the nodes of the
defined Yeo networks. We created two random networks by randomly selecting a subset of
nodes from the Schaefer parcellation, de 22 et 91 nodes, respectively matching the size of
the limbic and default networks in our study. Using the 22-node random network yielded
worse accuracy than any other network at 16%, but using the 91-node random network
yielded high accuracy at 94% (Supplementary Figure 2A).
Node:
A parcel representing a single region
from a brain atlas. Multiple nodes
compose a network.
Sliding-window approach:
Sample selection method in which
participants are ordered by a variable
(ici, âge) and iteratively selected to
create overlapping subsamples.
Neurosciences en réseau
1208
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
/
t
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
un
_
0
0
3
2
0
p
d
/
.
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
F
toi
n
c
t
je
o
n
un
je
c
o
n
n
e
c
t
o
m
e
F
je
n
g
e
r
p
r
je
n
t
je
n
g
un
c
r
o
s
s
t
h
e
je
je
F
e
s
p
un
n
Tableau 1. Demographics information. Handedness was measured as a continuous variable from −100 (fully left-handed) à 100 (full right-handed). Motion is reported
as the average frame displacement for each modality. The number of frames is the number of fMRI frames remaining after removing frames with excessive motion.
Counts are presented for categorical data, while average and standard deviation are presented for continuous data.
Decades of
age variables
Sex (F)
Sex (M.)
18–29 (n = 71)
40
30–39 (n = 88)
43
40–49 (n = 95)
50
50–59 (n = 68)
34
60–69 (n = 71)
30
70–79 (n = 56)
27
31
45
45
34
41
29
80–89 (n = 34) Dans l'ensemble (n = 483)
15
19
239
244
Mean (SD)
Age
25.00 (3.50)
34.83 (2.64)
44.54 (3.06)
54.51 (2.89)
64.45 (2.84)
75.16 (2.97)
83.06 (2.39)
50.49 (18.24)
Handedness
75.01 (49.57)
78.08 (48.75)
74.22 (55.35)
75.91 (54.06)
77.15 (52.92)
85.09 (42.77)
86.53 (36.06)
77.82 (50.12)
Motion
(mm)(Rest)
Motion
(mm)(Task)
Number of
frames(Rest)
Number of
frames(Task)
0.175 (0.048)
0.173 (0.047)
0.196 (0.057)
0.203 (0.050)
0.221 (0.041)
0.235 (0.047)
0.238 (0.046)
0.201 (0.054)
0.153 (0.044)
0.147 (0.042)
0.170 (0.051)
0.182 (0.049)
0.198 (0.046)
0.209 (0.047)
0.230 (0.052)
0.178 (0.053)
238.06 (31.14)
246.08 (26.66)
230.31 (34.58)
227.62 (32.12)
223.28 (41.48)
211.82 (38.74)
205.88 (38.79)
229.05 (36.23)
242.18 (29.75)
247.85 (20.68)
237.46 (28.61)
234.29 (26.11)
233.41 (32.38)
215.39 (39.24)
200.44 (43.10)
233.84 (32.90)
t
N
e
w
o
r
k
N
e
toi
r
o
s
c
e
n
c
e
je
1
2
0
9
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
un
_
0
0
3
2
0
p
d
.
/
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
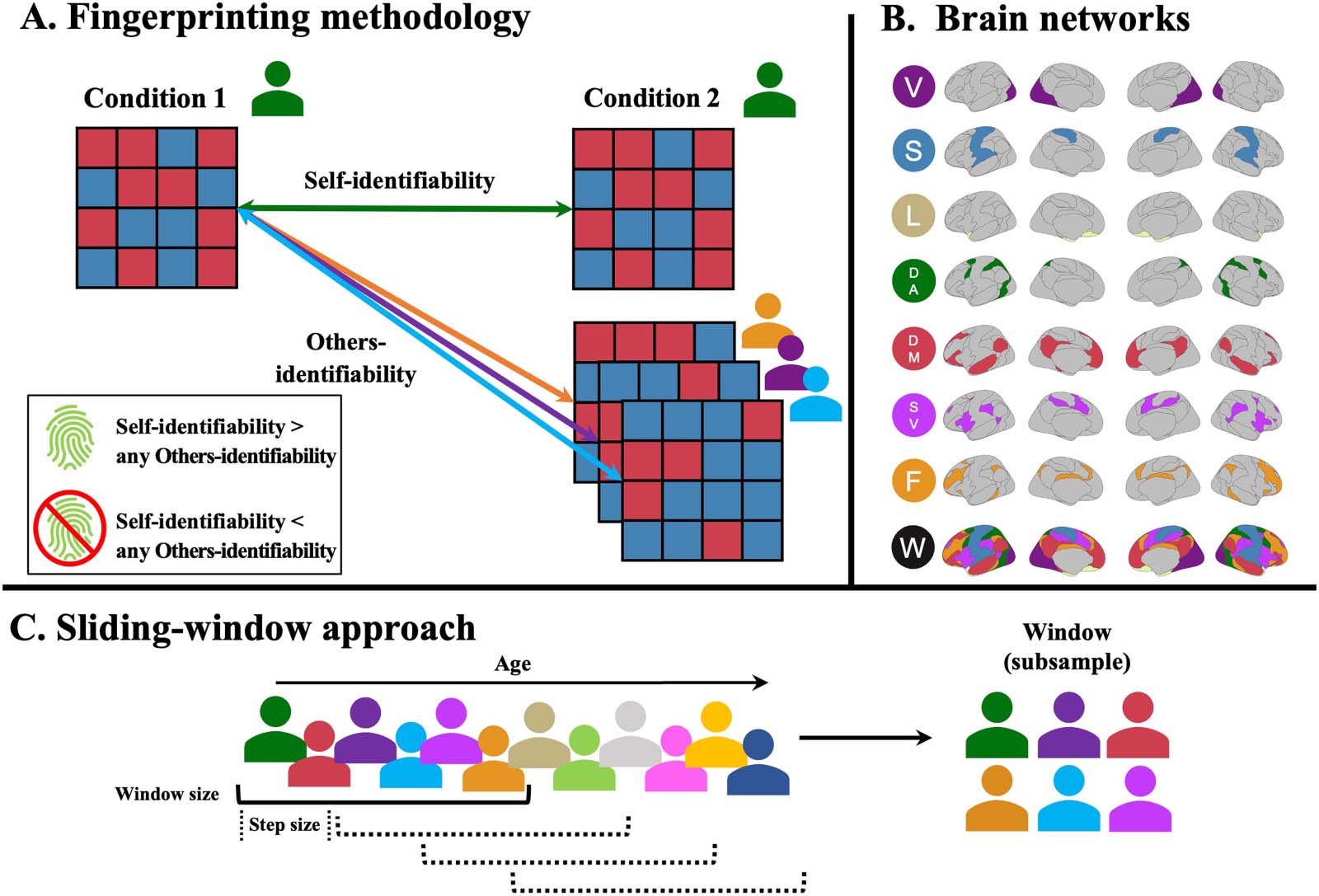
Illustration of the methodology. (UN) Illustration of the fingerprinting framework. Fingerprints are established by the correlation of the
Chiffre 1.
functional connectivity of participants between conditions. The correlation within the same individual constitutes self-identifiability, tandis que le
correlation between individuals constitutes others-identifiability (Amico & Goñi, 2018). If the self-identifiability is higher than any other others-
identifiability for a given participant, they are successfully identified (Finn et al., 2015). (B) Yeo functional networks used in the analyses of the
papier. V = visual, S = somatomotor, L = limbic, DA = dorsal attention, DM = default mode, SV = salience/ventral attention, F = frontoparietal,
W = whole brain. (C) Illustration of the sliding-window approach to select subgroups of participants. Subsamples of participants (window size)
were chosen iteratively by taking the participants from the cohort, ordered by age, and slowly moving along (step size) the lifespan. Ce
method yields subsets of overlapping participants across the lifespan, offering a cross-sectional, semi-continuous overview of changes during
aging. Window size and step size were varied to obtain different combinations of subsamples.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
un
_
0
0
3
2
0
p
d
.
/
t
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
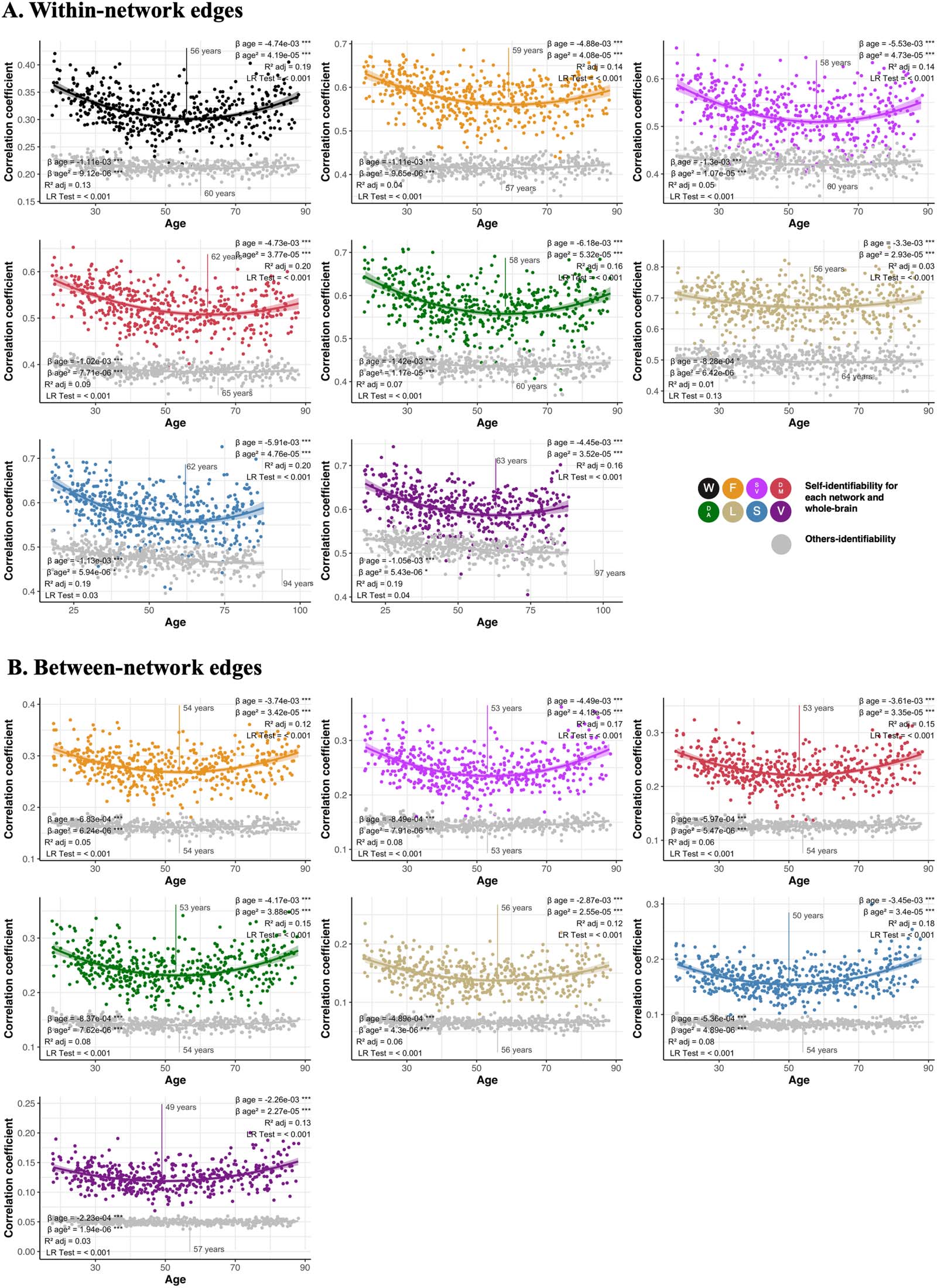
Both Self-Identifiability and Others-Identifiability Change Nonlinearly and in Parallel Across the Lifespan
We found, using quadratic regressions and nested likelihood ratio tests, that both self-
identifiability and others-identifiability differed nonlinearly across the lifespan across all
réseaux (for both within- and between-network edges; Chiffre 3). Using Stimson’s equation
for quadratic models (Stimson et al., 1978), we further found that both self-identifiability and
others-identifiability appeared to decrease until 49–63 years of age, before increasing
(Chiffre 3). Results of all models, except for others-identifiability in the limbic network,
remained significant when bootstrapping and controlling for sex, handedness, motion, et
number of fMRI frames available. Results were also very similar when using the Power atlas
(Supplementary Figure 3B) across all networks for the self-identifiability, and for the whole-
brain, default, frontoparietal, and dorsal attention networks for the others-identifiability. Nous
also found the same results when using our two random networks described in the previous
section (Supplementary Figure 2B).
To ensure that the nonlinearity of our results wasn’t driven by the oldest participants having
higher self- and others-identifiability, we excluded participants above 80 years of age and
repeated the analyses. We found nearly identical results (Supplementary Figure 4). Enfin,
we tested whether results would be similar when functional connectivity was generated using
product–moment correlations of the blood oxygen level–dependent (AUDACIEUX) signal between
nodes instead of using partial correlations. We found few associations between self-
Neurosciences en réseau
1210
Functional connectome fingerprinting across the lifespan
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
t
/
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
un
_
0
0
3
2
0
p
d
t
/
.
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 2. Unique connectomes across the lifespan, réseaux, and tasks. Fingerprint identifiability in the pair of Rest and Task conditions.
Panel (UN) illustrates the fingerprint identification accuracy across the entire sample using within- and between-network edges. The blue color in
the bar graphs and the percentages (with confidence intervals; alpha = 0.05) to the right of the graphs indicate the proportion of individuals
correctly identified. Network acronyms on the y-axes match graphics in Figure 1B and represent the specific functional network used for
identification. In panel (B), we used a between-individual, age-group sliding-window approach to plot how stable the fingerprint identification
accuracy was across the lifespan for each network.
identifiability and age using this method. Cependant, in networks where the association existed,
it exhibited a U-shape (Supplementary Figure 5).
Regions Contributing to Self-Identifiability Across the Lifespan
To determine which FC edges contributed to self-identifiability, we used an elastic net model
paired with the age-group sliding-window approach. Spécifiquement, we aimed to determine
which combination of FC edges across the brain were predictive of self-identifiability in both
a training and a testing set, in each age window. We applied these elastic net models to each
age window across our sliding-window parameters. We report the model performance of the
elastic net models in the left-out test set (c'est à dire., whether edges identified in the training model
also predicted self-identifiability in the left-out test set). We also report the nodal density for
each node in each age window. The nodal density indicates the extent to which edges from a
given node contribute to self-identifiability (Amico & Goñi, 2018). For each node, we summed
the number of edges identified by the elastic net as being important for prediction within each
node and divided this number by the total number of edges per node (400). While we report
the results for only the window size 100 and step size 40, the results were identical across
sliding-window parameters.
Dans l'ensemble, prediction of self-identifiability within each age window did not generalize to
any left-out samples and exhibited poor model performance across all age windows
Nodal density:
Sum of number of edges contributing
to an outcome within a node,
divided by the number of edges in
the node.
Neurosciences en réseau
1211
Functional connectome fingerprinting across the lifespan
je
D
o
w
n
o
un
d
e
d
F
r
o
m
h
t
t
p
:
/
/
d
je
r
e
c
t
.
m
je
t
.
/
t
/
e
d
toi
n
e
n
un
r
t
je
c
e
–
p
d
je
F
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
un
_
0
0
3
2
0
p
d
t
.
/
F
b
oui
g
toi
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Chiffre 3. Differences in self- and others-identifiability across the lifespan. Change in self-identifiability (colors) and others-identifiability (gray)
are represented using either within-network edges (UN) or between-network edges (B). Each graph represents a different network, following
acronyms and color schemes of Figure 1B. The beta coefficient of the age term and its quadratic term are presented at the top of the graph.
We also present the adjusted R2 of the model and the p value of the nested likelihood ratio indicating the nonlinearity of the relationship. Le
p value of predictors surviving inclusion of covariates and execution of the bootstrapping are indicated by asterisks (***p < 0.001, **p < 0.01,
*p < 0.05, °p < 0.10). The age at which the curve changed direction was calculated from Stimson’s equation (Stimson et al., 1978) and is
illustrated on the graphs.
Network Neuroscience
1212
Functional connectome fingerprinting across the lifespan
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
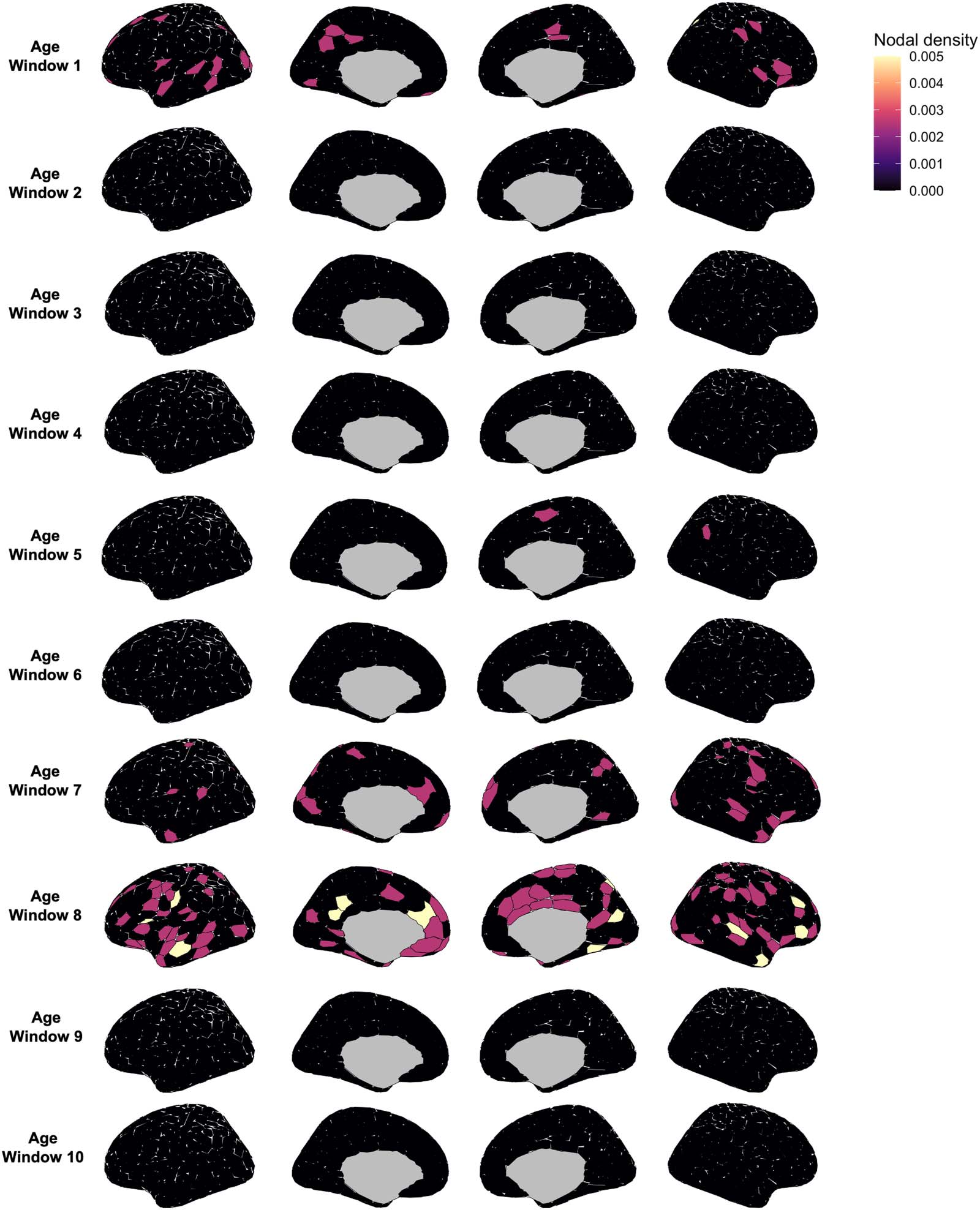
Figure 4. Distribution of nodes predicting self-identifiability across the brain. For each age window (see sliding-window approach,
Figure 1C), we plot the nodal density (sum of number of edges identified by the elastic net as being predictive of self-identifiability divided
by the total number of nodes) using the Schaefer atlas (400 nodes). A higher nodal density indicates that the node had a higher proportion of
edges contributing to self-identifiability. Average age in each window matches averages in Figure 2.
(Supplementary Figure 6). Furthermore, no specific nodes had more predictive edges than
others (Figure 4). In fact, in many windows, the elastic net did not identify any combination
of edges that were predictive of self-identifiability. These results suggest that no combination of
FC brain edges can reliably predict self-identifiability across individuals.
A potential issue with using the elastic net model with our data is the massive number of pre-
dictors in the model (79,800 edges) and the sparse nature of the FC matrices derived from partial
correlations rather than product–moment correlations. This could explain the lack of
Network Neuroscience
1213
Functional connectome fingerprinting across the lifespan
Temporal similarity profile:
Correlation of a wide range of BOLD
signal features between each node,
yielding how similar the features of a
node are to the features of a different
node.
generalizability in the left-out test sets, as well as the lack of nodes identified consistently as
predictive. To ensure that our results were not driven by these limitations, we adapted three
more methods to verify these results: a connectome predictive modeling approach, an intraclass
correlation approach, and a clustering approach. The connectome predictive modeling and
intraclass correlation approaches tested whether FC in individual edges in specific networks
were driving self-identifiability. Alternatively, the clustering approach aimed to confirm whether
patterns of individual FC were shared across the brain in each age window. Detailed discussions
of these methods are found in the Supporting Information. All these additional analyses converged
toward the same conclusion as the elastic net model: Regions contributing to self-identifiability
vary across the lifespan and between individuals (Supplementary Figures 7 to 14).
Variability in FC, but Not Variability in BOLD Features, Differs Over the Lifespan
We additionally tested whether other markers of variability in fMRI signal differed over the
lifespan (Supplementary Figure 15). To this end, we derived two types of measures, variability
in FC and variability in BOLD temporal similarity profile. Within-individual variability in FC
was defined as the variance coefficient of FC values within a given network for a given indi-
vidual. We defined the between-individual variability in FC as the distance (subtraction)
between the variability in FC of an individual to the average variability in FC of the group.
Variability in temporal similarity profile was defined in the same way but using BOLD signal
feature profiles instead of FC (Shafiei et al., 2020). More information on these measures is
available in the Methods section. For both measures we calculated a within-individual vari-
ance measure and between-individual variance measure. These were calculated at the whole-
brain and network levels, and for each fMRI condition (Rest and Task). In contrast to the
fingerprinting results, within-individual variability in FC decreased linearly across the lifespan
in all networks, and between-individual variability in FC did not change with age in any net-
work. No associations were found between age and variability in temporal profile similarity.
Finally, we tested whether a nonlinear change in amplitude of the BOLD signal with age could
be driving the nonlinear association between self-identifiability and age. Amplitude was calcu-
lated as the average of the amplitude of the BOLD signal for all nodes in a given network, for each
participant. We found that there was no association between amplitude of the BOLD signal and
age in the Rest modality. In the Task modality, amplitude of the BOLD signal was associated
nonlinearly with age (Supplementary Figure 16), but self-identifiability was not associated with
BOLD signal amplitude (Supplementary Figure 17). We conclude that it is very unlikely that the
BOLD signal amplitude drives the nonlinear association between self-identifiability and age.
Self-Identifiability Is Associated With Gray Matter Volume
Our initial results suggest that fingerprint accuracy is reliably achieved in adults over the life-
span. This is likely achieved due to the variability across people in the strongest FC patterns
across the brain. We finally wanted to assess whether this identifiability was associated with
age-sensitive variables, such as brain volume. To do so, we used three morphometric networks
derived from an independent study as our outcome of interest (Pichet Binette et al., 2020): one
frontal network (strongest age-related changes), a limbic network, which includes the hippo-
campus and the medial temporal lobe (moderate associations with both Alzheimer’s disease
and age-related changes), and an occipital network (weakest age-related effect). Gray matter
volume was extracted for all three networks and used in our analyses.
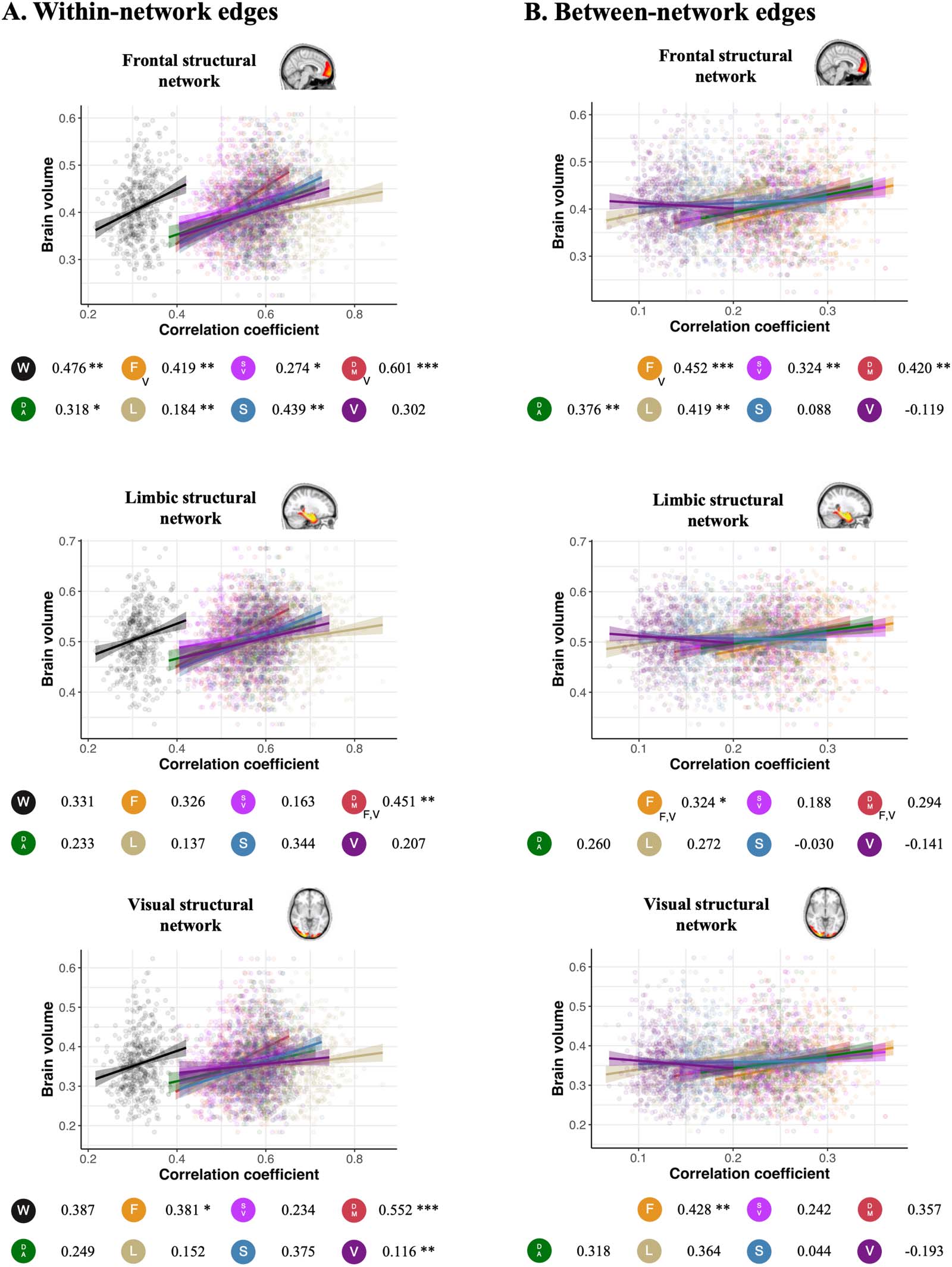
Lower self-identifiability was associated with lower gray matter volume in the frontal structural
network over and above the effect of age and other covariates (Figure 5). This connectome-wide
Network Neuroscience
1214
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
.
t
/
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Figure 5. Association between gray matter volume and self-identifiability. Scatterplots presenting the association between self-identifiability,
derived using (A) within- and (B) between-network edges, and gray matter volume in three gray matter morphometric networks: frontal struc-
tural network (age-sensitive network), limbic structural network (Alzheimer’s/age-related network), and visual structural network (“control”
network). Data points, regression slopes, and bubbles below the graph follow the color scheme of Figure 1B. The beta coefficient of the
relationship between the self-identifiability and the brain volume is indicated beside each network bubble. The p value of each predictor
surviving comparison with covariates and bootstrapping is denoted by asterisks next to the beta coefficient (***p < 0.001, **p < 0.01, *p <
0.05). Models surviving all confounders for all three morphometric networks were compared using Vuong’s test for non-nested models. A letter
at the bottom right of the network acronym indicates that the association was stronger using that specific structural network compared with the
other networks referred to by the letter ( V = visual, F = frontal, L = limbic).
Network Neuroscience
1215
Functional connectome fingerprinting across the lifespan
result was recapitulated in the within- and between-network self-identifiability metrics (except
for self-identifiability in the visual networks for within-network edges and visual and somato-
motor networks for between-network edges).
Others-identifiability was not associated with brain volume (Supplementary Figure 18).
Using our markers of variability in FC and BOLD signal, we found that decreased within-
individual variability in FC was associated with decreased brain volume across networks
(Supplementary Figure 19) but to a lower extent than self-identifiability. Similar to others-
identifiability, there was no association between between-individual variability in FC and
brain volume. We also did not find any association between variability of temporal profile
similarity and brain volume.
DISCUSSION
We found that connectome-based fingerprinting is reliable across the lifespan. Fingerprint
identification accuracy was robust even though self-identifiability and others-identifiability
show a nonlinear cross-sectional distribution across the lifespan. We also found that the
highest weighted edges contributing to self-identifiability varied across individuals. This
interindividual variability was observed within each decade of life. Furthermore, relative to
whole-brain and within-network FC, between-network FC provided more reliable identifica-
tion estimates and the number of nodes (and related edges) were more important than their
localization to identify individuals using brain functional proprieties. Finally, we found that
self-identifiability (but not others-identifiability) was associated with frontal brain volume, a
morphometric feature known to atrophy with advancing age (Pichet Binette et al., 2020).
Our findings indicate that fingerprinting remains robust across the lifespan, despite
observed age-related changes in FC in older adulthood (Geerligs et al., 2015; Zonneveld
et al., 2019). We found high fingerprinting identification accuracy across the lifespan, with
perfect to nearly perfect identifiability accuracy in the whole-brain connectomes and in the
frontoparietal and default mode networks. Higher identification rates for associative cortical
FC compared with unimodal networks have been found previously (Airan et al., 2016; Amico
& Goñi, 2018; Finn et al., 2015; Horien et al., 2018, 2019; Van De Ville et al., 2021; Vanderwal
et al., 2021) and have been suggested to be the result of high interindividual variability in FC
within these regions (Geerligs et al., 2015; Mueller et al., 2013). While this might partly be the
case, networks such as the default mode network include a larger number of nodes than do some
unimodal networks. In supplementary analyses we showed that we could achieve almost perfect
identification using a large random network composed of the same number of nodes as the ones
used in the default mode network. This identification was significantly diminished in a random
network that included the same number of nodes as the ones used in the limbic network. These
results suggest that high identification may largely depend on the amount of information provided
to perform this identification. This is in line with previous studies suggesting that finer grained
parcellations—that is, more nodes—yield higher identification accuracy (Finn et al., 2015).
The amount of information available for each participant might therefore be more important
than the specific cortical topography when identifying individuals.
Next, we explicated fingerprint identification accuracy into its two components: self-
identifiability and others-identifiability (Amico & Goñi, 2018). We found that both components
have a nonlinear trajectory across the lifespan. These U-shaped trajectories comprise high
scores in young adults, which decrease into middle age and increase in older adulthood. This
phenomenon was present across networks, and it impacted metrics of self-identifiability more
than others-identifiability. While U-shape trajectories have previously been reported in
Network Neuroscience
1216
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
functional and structural lifespan studies (DuPre & Spreng, 2017; Kupis et al., 2021; Nadig
et al., 2021), the exact cause of this phenomenon is unknown. This nonlinear trajectory could
be explained by a number of factors. Interindividual heterogeneity in anatomy is most variable
in early and late life, with more homogeneity observed between middle-aged adults (Nadig
et al., 2021). This variance could boost individual identifiability in the youngest and oldest
participants. Additionally, middle-aged women may have lower self-identifiability between
fMRI sessions because of perimenopausal fluctuations in estrogen and progesterone (Pritschet
et al., 2020), which impact brain function on multiple scales. Previous studies in young adults
found no longitudinal change in fingerprinting accuracy over a 2- to 3-year follow-up (Ousdal
et al., 2020), suggesting that a longitudinal lifespan approach is needed to better characterize
age-related change occurring over decades.
We also found that change occurring with age in self-identifiability and others-identifiability
mainly occurs in parallel in all networks, including the random networks, which likely
explains why the high identification accuracy is preserved across the lifespan despite age-
related changes in its two components. As long as the balance between self-identifiability
and others-identifiability is preserved, healthy individuals can be identified among a large
group of individuals with similar accuracy (Horien et al., 2019; Kaufmann et al., 2018).
Findings from developmental cohorts suggest that fingerprint metrics increase rapidly from a
few days after birth and stabilize in young adults (Horien et al., 2019; Hu et al., 2022;
Jalbrzikowski et al., 2020; Kaufmann et al., 2018; Vanderwal et al., 2021). Additionally, neu-
ropsychiatric symptoms lead to a decrease in self-identifiability (Kaufmann et al., 2017, 2018).
From a developmental perspective, fingerprints stabilize in early adulthood, but they may be
disrupted when the brain is affected by an overt neuropsychiatric or neurological disorder, or
subclinical perturbations, resulting in mid-life declines in identifiability. The older adult cohort
could represent either a stabilization of these effects, or a more carefully screened sample of
healthy individuals that persist in the developmental stability of identification.
Our findings are consistent with work suggesting that fMRI connectivity is composed of
distinct, unique individual-specific and shared task-specific components (Gratton et al.,
2018; Mantwill et al., 2022). Using four different approaches (elastic net, connectome predic-
tive modeling, edge-wise intraclass correlation, and clustering), we did not find consistent
edges contributing to self-identifiability in our sample across individuals. This aligns with
our large random network yielding high identification accuracy, suggesting that brain signa-
tures, and their associated brain regions, are unique across individuals. These analytical
approaches reinforce the finding that between-individual variance in functional brain connec-
tomics are critical to the identification of individual functional fingerprints.
Finally, we aimed to determine whether individual-level FC fingerprint metrics were asso-
ciated with brain volume. We compared brain volume associations with self-identifiability and
others-identifiability. Whole-brain connectomes, within-network FC, and between-network FC
self-identifiability were associated with brain volume in frontal areas known to be particularly
affected by normal age-related atrophy (Pichet Binette et al., 2020). We also found consistent
associations for self-identifiability between default and frontoparietal networks and brain
volume in the hippocampus and medial temporal lobe (limbic structural network; known to
be vulnerable in Alzheimer’s disease). The occipital structural regions were preserved over the
lifespan and did not impact self-identifiability (Pichet Binette et al., 2020). In contrast, there
was no consistent association between others-identifiability and brain volume.
Self-identifiability may be associated with a number of factors that differentially impact
individuals over the lifespan (Kaufmann et al., 2017, 2018). For instance, the default network
Network Neuroscience
1217
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
is affected by a wide range of neurological disorders (de Lange et al., 2019) and aging
(Zonneveld et al., 2019). Consistent with this idea, reduced fingerprint identifiability within
the default network has been associated with mental health disorders in youth (Kaufmann
et al., 2017) and reduced brain volume in older adults (Ousdal et al., 2020). Individual-level
FC measures, rather than group-level measures, may be more appropriate to detect age- or
disease-related functional changes (Finn & Constable, 2016), either by directly accounting
for interindividual differences (Finn & Rosenberg, 2021) or, at least partially, by ignoring
group-level noise in fMRI signal (Amico & Goñi, 2018). Targeting individual-level differences
might be particularly important in aging research, as aging individuals present high diversity
of lifestyle and medical history, leading to diverse age-related outcomes (Daskalopoulou
et al., 2019).
As an additional note, the findings above also expand methodological aspects in the field of
fingerprinting. First, we used partial correlation to generate FC matrices, which were then used
for fingerprinting. Partial correlations resulted in higher identification rates in our study com-
pared with product–moment correlations. Partial correlations produce sparse matrices exclud-
ing, to some extent, noise inherent to fMRI (Marrelec et al., 2006). Other methods aiming at
removing noise in FC matrices have also found success in improving fingerprint identification
accuracy (Amico & Goñi, 2018). As noise affects individuals across the sample in a similar
manner (Amico & Goñi, 2018), removing noise should therefore better isolate individual-
specific features. Second, we computed identification using between-network edges as well
as within-network edges. In most cases, between-network edges provided better identification
rates compared with within-network edges. To our knowledge, all studies on fingerprinting to
date have used within-network edges to compute identification. However, while within-
network edges represent communities of nodes working tightly together (Sporns & Betzel,
2016), between-network connections are necessary to various brain functions. In aging in par-
ticular, the segregation of these networks tend to change (Chan et al., 2014), and as such
between-network edges may better represent overall brain network communications. Our
study therefore also suggests potential methodological aspects to consider in future finger-
printing studies, particularly when considering aging populations.
Strengths and Limitations
The main limitation of this study is its cross-sectional nature. As such, it is difficult to determine
how fingerprint metrics change during the lifespan within individuals, limiting the interpreta-
tion of our conclusions. Future work would benefit from longitudinal studies examining iden-
tifiability across scans separated by many years. Similarly, because of quality control issues,
the eldest participants in our cohort were underrepresented, and as such, the nonlinearity of
our findings need to be interpreted with caution. However, supporting the nonlinearity of the
findings, the nonlinearity between age and self-identifiability was still preserved when remov-
ing the oldest participants. Finally, we included only two fMRI modalities in our analyses. As
other studies have shown that the choice of the task can influence the ability to fingerprint,
future studies should replicate our findings using other combinations of fMRI tasks.
Conclusions
Across our analyses, we found that FC patterns allow for precise fingerprinting between indi-
viduals and that this discrimination is reliable across the lifespan. High identification rates
were observed across the Cam-CAN cohort, accompanied by age-related effects on
individual-level (self-identifiability) and group-level (others-identifiability) FC patterns. Accu-
rate fingerprinting of FC was observed across the lifespan, even though edges contributing to
Network Neuroscience
1218
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
self-identifiability differed between individuals across networks and across the decades of life.
Finally, we show that individual-level self-identifiability (instead of group-level others-
identifiability) is associated with brain volume in regions vulnerable to age-related atrophy.
Together, the present findings illuminate the potential utility of individual-level measures that
demarcate age-related brain change. Group-level differences in FC have revealed reliable
patterns attributed to the aging brain. However, individual differences in FC patterns are likely
to play a key role in predicting brain health and associated functional outcomes.
MATERIALS AND METHODS
Participants
Data used in the preparation of this work were obtained from the Cam-CAN repository (available
at https://www.mrc-cbu.cam.ac.uk/datasets/camcan/). The Cam-CAN cohort is a lifespan cross-
sectional population-based cohort, composed of cognitively healthy participants aged between
18 and 89 years of age residing in the United Kingdom. Full details for the study participants
and recruitment can be found elsewhere (Shafto et al., 2014; Taylor et al., 2017). Participants
underwent several brain imaging procedures at one time point. Special attention was given to
recruiting persons from different decades and to balance both men and women. This study
was approved by the Cambridgeshire 2 Research Ethics Committee (reference: 10/H0308/50).
Magnetic Resonance Imaging
Full details on the imaging data collection and on the fMRI tasks used are available elsewhere
(Taylor et al., 2017). Briefly, MRI data were acquired on a 3T Siemens TIM Trio scanner with a
32-channel head coil for a 1-hr session. T1-weighted MPRAGE sequences were acquired for
structural imaging, and T2*-weighted EPI sequences were acquired for fMRI imaging (261 vol-
umes with 32 axial slices each; slice thickness of 3.7 mm; interslice gap of 0.74 mm; TR =
1,970 ms; TE = 30 ms; flip angle = 78 degrees; FOV = 192 mm × 192 mm; voxel size =
3 mm × 3 mm × 4.44 mm). Participants underwent two different fMRI conditions during
one session: a resting-state condition (Rest), where participants were asked to keep their eyes
closed for 8 min, 40 s; and a sensorimotor task (Task), also of 8 min, 40 s, in which participants
were asked to press a button when audiovisual stimuli were presented.
Functional images from the two modalities were preprocessed using the NeuroImaging
Analysis Kit, version 0.12.4 (NIAK; https://niak.simexp-lab.org/; Bellec et al., 2011; Gonneaud
et al., 2020). The first three volumes of each run were suppressed to allow the magnetization to
reach equilibrium. Images were slice-timing corrected, and rigid-body motion parameters
were estimated for each time frame. For registration, T1-weighted images were linearly and
nonlinearly registered to MNI space. The rigid-body transform, fMRI-to-T1 transform, and
T1-to-stereotaxic transform were all combined, and the functional volumes were resampled
in the MNI space at a 3-mm isotropic resolution. To account for potentially excessive motion,
frame displacement was calculated for each volume, and those with more than 0.5 frame dis-
placement were removed with one prior adjacent frame and two consecutive frames after.
Time series with less than 40% of their original data after removing excessive motion were
discarded from subsequent analyses (Orban et al., 2015). Next, slow time drifts, cerebrospinal
fluid, average white matter signal, and motion artifacts (first principal components of the six
rigid-body motion parameters, and their squares) were removed from the fMRI time series, and
fMRI volumes were smoothed with a 6-mm Gaussian kernel. Finally, fMRI time series for each
region of the Schaefer atlas (n = 400) were extracted using Nilearn 0.6.2 (Abraham et al.,
2014). Partial correlations were used to generate FC between regions, accounting for the signal
Network Neuroscience
1219
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
in all other brain regions. This process generates sparser functional connectivity matrices that
are thought to account for more noise than using simple product–moment correlation and rep-
resent more direct connections between regions (Marrelec et al., 2006; Zalesky et al., 2012).
We generated FC matrices for the task and resting-state fMRI runs.
All structural images were preprocessed using Statistical Parametric Mapping (SPM12,
https://www.fil.ion.ucl.ac.uk/spm/software/spm12/) in MATLAB version 2012a, as part of a pre-
vious study (Pichet Binette et al., 2020). Images were segmented into gray matter, white matter,
and CSF components. Then, a group-specific template was created using the Diffeomorphic
Anatomical Registration through Exponentiated Lie Algebra toolbox (DARTEL; Ashburner,
2007), which was then registered nonlinearly to the MNI-ICBM152 template. Finally, each
individual participant’s gray matter map was registered back to the group template, before
being smoothed with an 8-mm3 isotropic Gaussian kernel.
Sliding-Window Analysis
Across multiple analyses, we used a between-participant sliding-window approach to study
differences across the lifespan in a semi-continuous manner (Váša et al., 2018; Figure 1C).
First, participants were ordered by age (from youngest to oldest). Then, we iteratively selected
subsamples of overlapping participants varying two parameters: window size (i.e., the number
of participants in each subsample) and step size (i.e., the number of participants skipped
before selecting the next window). We used window sizes of 100 or 150 participants and used
a step size of either 25 or 40 participants. As such, participants in adjacent windows over-
lapped by 60% to 80%. Main analyses report results using a window size of 100 and step size
of 40, while results using other parameters are reported in supplementary analyses.
Fingerprinting
Our main interest was functional connectome fingerprinting, which encompasses three mea-
sures of interest: fingerprint identification accuracy (Finn et al., 2015), self-identifiability, and
others-identifiability (Amico & Goñi, 2018). In the fingerprinting framework, the FC matrix of a
given individual obtained from one fMRI condition is correlated to the FC matrix of the same
individual obtained during a different fMRI condition by computing the correlation coefficient
of the vectorized upper triangle z-values between sessions. This results in a within-individual
correlation (i.e., self-identifiability). This process is repeated for all within- and between-
participant FC matrices, permitting the computation of both within- and between-individual
similarities (i.e., others-identifiability). Finally, fingerprint identification accuracy is estimated
from both self- and others-identifiability measures: A fingerprint is considered identifiable
when self-identifiability exceeds the magnitude of others-identifiability (Amico & Goñi,
2018; Finn et al., 2015; see Figure 1A for schematic overview). This process was done for
the whole-brain connectome as well as for within-network and between-network edges for
each network.
The fingerprint framework was adapted from the original methodologies by Finn et al.
(2015) and Amico and Goñi (2018). The FC obtained for the Rest and Task modalities were
first normalized using Fisher’s r-to-z transform. Product–moment correlations were then
used to correlate FC matrices obtained from the Rest and Task conditions, deriving self-
identifiability and others-identifiability. These values where then stored in a similarity matrix.
For each row of the matrix, a fingerprint identification was considered accurate if self-
identifiability (diagonal elements of the matrix) was higher than any other others-identifiability
(off-diagonal elements of the matrix). Self-identifiability and others-identifiability were
Network Neuroscience
1220
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
/
t
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
computed based on the FC between 400 parcels. Additionally, the parcel information corre-
sponding to the Yeo seven networks was leveraged to demarcate within-network FC. Then, we
composed aggregates of between-network FC by taking any edges between each other net-
work. For example, if the visual network was composed of nodes 1 to 61 (rows and columns
1 to 61 in the matrix), within-network edges comprised edges where both rows and columns
were between 1 and 61. Between-network edges comprised edges in rows 1 to 61 but in
columns 62 to 400 (Yeo et al., 2011).
Additionally, we created two random networks to test whether we could find high identi-
fication accuracy using edges belonging to a random assortment of nodes rather than using
predefined networks. We selected two random sets of nodes across the brain. The number
of nodes chosen were 22 and 91, to match the size of the smallest (limbic) and largest (default)
networks (Schaefer et al., 2018; Yeo et al., 2011) included in our study. Fingerprinting
methodology described above was applied to the edges of these two random networks.
Predicting Self-Identifiability Using Combinations of Edges
We adapted an elastic net approach to predict self-identifiability from the FC edges in our
sample. Specifically, we used the sliding-window method to select subsamples of participants
across the lifespan. Within each window, we first removed the diagonal and lower triangle
of the functional connectivity matrices of the participants and flattened the remaining
79,800 edges. The connectivity values were then standardized, and participants were ran-
domly split in a training and testing subset (85% training, 15% testing). Connectivity values
across all 79,800 edges were used to predict self-identifiability. In the training set, we used
a 5 k-fold cross-validation with a grid search to select the optimal L1 ratio for our elastic net
model. Once the optimal L1 ratio was selected, an elastic net model was fitted on the entire
training set. The model was then used to predict self-identifiability in the testing set. Perfor-
mance in the testing set was reported as the variance explained (R2) and the root mean
square error (RMSE).
The elastic net outputs coefficients indicating which edges significantly contributed to
model performance. To determine their topography in the brain, we adapted a nodal density
approach (Amico & Goñi, 2018). Briefly, edges were resized to a 400 × 400 functional con-
nectivity matrix format and binarized. Then, the sum of the binary coefficients for each node
(i.e., each row of the matrix) was divided by the number of edges for that node (i.e., 400). This
yields a nodal density measure for all 400 nodes of the Schaefer atlas, where more density
indicates that edges in that node are more important. These results were then projected to a
brain map.
Age-Related Outcome: Structural Aging Morphometric Network
We tested the relationship between functional fingerprint metrics (self-identifiability and
others-identifiability) and structural age-related changes. Three morphometric networks
derived in an independent study were used as our outcome of interest (Pichet Binette et al.,
2020). Briefly, a large cohort of cognitively unimpaired younger and older adults and partic-
ipants on the Alzheimer disease spectrum were grouped and independent component analysis
was used to derive statistically independent structural brain networks. Then, volume in each
component was used in a receiver operating characteristic analysis to determine whether brain
volume could accurately classify individuals in their corresponding group. From this study, we
chose one frontal network (ICA01 in the original study, which was the network showing the
strongest age-related changes), a limbic network (ICA10, which showed moderate associations
Network Neuroscience
1221
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
with both Alzheimer’s disease and age-related changes), and an occipital network (ICA15,
which showed the weakest age-related effect). Gray matter volume was then extracted for
all three morphometric networks for each participant. These morphometric networks were
chosen rather than using parcel-level or network-level gray matter measures, as we wanted
age-associated gray matter measures and as their association with fingerprint measures could
be more directly comparable between networks.
Supplementary Analyses on FC Variability
Additionally, we also determined whether other markers of variability in fMRI signal changed
during the aging process. We derived two types of measures, variability in FC and variability in
BOLD temporal similarity profile.
Both types of measures were calculated using custom Python scripts. Variability in FC for
each individual was computed as the variance coefficient of FC coefficients at a whole-brain
level and within each network of interest. To obtain between-individual variability, we first
computed the average variability in FC across the sample. Then, for each individual, we com-
puted the absolute distance between their variability and the mean variability of the group. A
greater distance indicates more between-individual variability.
Variability within and between individuals in BOLD signal features was computed as the
variance in temporal profile similarity of BOLD signal features (Shafiei et al., 2020). To do so,
BOLD signal features were extracted from each brain region using the highly comparative time
series analysis toolbox (Fulcher & Jones, 2017; Fulcher et al., 2013). Time series were first
z-scored and then fed to the toolbox where 7,700 features were extracted from the BOLD
signal. Because the number of fMRI frames varied between individuals after the preprocessing,
some features had missing or constant values across participants and were therefore dropped.
This resulted in a final 6,192 features remaining. Extracted features were then z-scored again to
ensure comparability between features. For each participant, the extracted features time series
for each region were correlated using product–moment correlations to determine how similar
time series were between each brain region, resulting in a 400 × 400 temporal similarity pro-
file similarity matrix. Finally, variability in temporal similarity profile was computed identically
to the variability in FC.
As a final verification, we computed BOLD signal amplitude and related this measure to
age and self-identifiability. In each of the 400 brain regions, for each participant, the amplitude
of the time series were computed by subtracting the minimum from the maximum BOLD
value. Then, the amplitude of the nodes within the network was averaged to obtain a single
amplitude measure for each network for each participant.
Software
The sliding-window, fingerprinting framework, elastic net, edge-wise intraclass correlation
analyses, connectome predictive modeling, clustering, and variability in FC analyses were
adapted and developed using Python 3.8.5 (Python Software Foundation, https://www
.python.org/; NumPy 1.19.1, Harris et al., 2020; pandas 1.1.3, McKinney, 2010; Pandas
Development Team, 2020; scikit-learn 0.24.0, Abraham et al., 2014; SciPy 1.7.1, Virtanen
et al., 2020; pingouin 0.5.2, Vallat, 2018) on Béluga, a high-performance computing resource
hosted by the Digital Research Alliance of Canada, running on CentOS 7.9. The temporal sim-
ilarity profile analysis was done using MATLAB 2021b (highly comparative time series analysis
toolbox; Fulcher & Jones, 2017; Fulcher et al., 2013) and Python 3.8.5. All statistical analyses
and graphs were done in R v4.1.2 (R Core Team, 2020; tidyverse 1.3.0, Wickham et al., 2019;
Network Neuroscience
1222
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
boot 1.3-28, Canty & Ripley, 2021; Davison & Hinkley, 1997; patchwork 1.1.1, Lin Pedersen,
2020; ggnewscale 0.4.8, Campitelli, 2022; ggseg 1.6.5, Mowinckel & Vidal-Piñeiro, 2020)
using R Studio (“Ghost Orchid” Release [077589bc, 2021-09-20]) for macOS Monterey
12.6. All code related to the analyses are available at https://github.com/villeneuvelab
/projects (St-Onge, 2022). The code related to the fingerprinting analysis was also adapted
to the openly available sihnpy Python package (https://sihnpy.readthedocs.io/).
Statistical Analyses
Fingerprint identifiability was computed for all networks for the Rest and Task conditions for
both within- and between-network edges. Percentage of identified individuals with confidence
intervals were calculated. Paired McNemar tests were used to compare identification in a
given network to the identification of the same network using a different fingerprint type
(i.e., using within- vs. between-network edges). We used this approach to compare network
performance in fingerprint identifiability within each modality pair (e.g., comparing proportion
of identification using the default vs. somatomotor networks). A family-wise Bonferroni correc-
tion was applied to each set of comparisons to account for multiple comparisons. Finally, we
used the sliding-window approach where the fingerprint identification accuracy in each age
window was computed and visualized.
To study the relationship between self-identifiability and age, and others-identifiability and
age, we used multiple linear regression models. We used polynomial (quadratic) regressions
when the linearity assumption was violated and, in that case, used Stimson’s equation to
derive the peak’s coordinates (Stimson et al., 1978). Assumption violation was determined
by comparing fitted to residual values. We also used nested likelihood ratio tests to test
whether models using quadratic terms outperformed linear models on model fit to confirm
the nonlinearity of the relationship. Each model included common confounders known to
affect either FC or fingerprints as covariates of no interest: the number of frames post-scrubbing
(Amico & Goñi, 2018; Horien et al., 2018; Xu et al., 2016), the mean frame displacement
(i.e., movement; Amico & Goñi, 2018; Geerligs et al., 2017; Guo et al., 2012; Horien et al.,
2018; Jalbrzikowski et al., 2020), the sex of participants (Finn et al., 2017), and a continuous
handedness measure (Bailey et al., 2020). One model was generated for each network for
both self-identifiability using within- and between-network edges (45 models total). The
same number of models was generated to study the association between others-identifiability
and age. Because of the high number of regressions, we applied a bootstrap resampling
procedure to each model as a way to account for multiple comparisons (Westfall, 2011).
Specifically, we generated bootstrapped bias-corrected and accelerated confidence intervals
for the β-coefficients (where coefficients not overlapping with 0 were considered significant)
of the main exposure. All analyses were repeated using within- and between-individual var-
iability in FC and BOLD signal (i.e., variability in FC and variability in temporal similarity
profiles) in each fMRI task in each network and within the whole brain.
All procedures described above were reapplied to study the relationship between finger-
print metrics and brain volume. Individual variability in FC and temporal similarity profile
were also associated with age and with gray matter volume using the methodology described
above. These metrics were used to determine whether other fMRI measures would also show
interindividual variability across the lifespan. To test whether the association was stronger in
the frontal structural network, the limbic structural network, or the visual structural network,
we used Vuong’s test for non-nested models (Vuong, 1989). Models were repeated including
all covariates with and without age.
Network Neuroscience
1223
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
t
/
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
ACKNOWLEDGMENTS
The authors would like to thank Jonathan Gallego Rudolf, Yara Yakoub, Ting Qiu, and Julien
Menes for useful feedback on the project, and Gabriel St-Onge for technical help in imple-
menting the connectome predictive modeling. We would also like to thank the reviewers
for providing their time to review this paper and for sharing their thoughtful insights into
our results, which significantly strengthened the paper. Data collection and sharing for this
project were provided by the Cambridge Centre for Ageing and Neuroscience (Cam-CAN).
Computations were enabled in part by support from Calcul Québec (calculquebec.ca) and
the Digital Research Alliance of Canada (alliancecan.ca), who provided access to the Béluga
high-performance computing resource. This research was undertaken thanks in part to funding
from the Canada First Research Excellence Fund and Fonds de recherche du Québec, awarded
to the Healthy Brains, Healthy Lives initiative at McGill University. The funding organizations
did not have a role in writing the manuscript.
SUPPORTING INFORMATION
Supporting information for this article is available at https://doi.org/10.1162/netn_a_00320.
AUTHOR CONTRIBUTIONS
Frédéric St-Onge: Conceptualization; Data curation; Formal analysis; Methodology; Software;
Visualization; Writing–original draft; Writing–review & editing. Mohammadali Javanray:
Conceptualization; Formal analysis; Writing–review & editing. Alexa Pichet Binette: Concep-
tualization; Methodology; Writing–review & editing. Cherie Strikwerda-Brown: Conceptuali-
zation; Writing–review & editing. Jordana Remz: Conceptualization; Software; Writing–review
& editing. R. Nathan Spreng: Methodology; Supervision; Writing–review & editing. Golia
Shafiei: Methodology; Writing–review & editing. Bratislav Misic: Methodology; Supervision;
Writing–review & editing. Étienne Vachon-Presseau: Methodology; Supervision; Writing–
review & editing. Sylvia Villeneuve: Conceptualization; Funding acquisition; Methodology;
Resources; Supervision; Visualization; Writing–original draft; Writing–review & editing.
FUNDING INFORMATION
Cam-CAN, UK Biotechnology and Biological Sciences Research Council, Award ID:
BB/H008217/1. Frédéric St-Onge, Healthy Brains, Healthy Lives through the Canada First
Research Excellence Fund and the Fonds de Recherche du Québec, Award ID: NA. Frédéric
St-Onge, Mitacs (https://dx.doi.org/10.13039/501100004489), Award ID: NA. Frédéric
St-Onge, Fonds de Recherche du Québec–Santé (https://dx.doi.org/10.13039/501100000156),
Award ID: 316992.
REFERENCES
Abraham, A., Pedregosa, F., Eickenberg, M., Gervais, P., Mueller, A.,
Kossaifi, J., Gramfort, A., Thirion, B., & Varoquaux, G. (2014).
Machine learning for neuroimaging with scikit-learn. Frontiers
in Neuroinformatics, 8, 14. https://doi.org/10.3389/fninf.2014
.00014, PubMed: 24600388
Airan, R. D., Vogelstein, J. T., Pillai, J. J., Caffo, B., Pekar, J. J., & Sair,
H. I. (2016). Factors affecting characterization and localization of
interindividual differences in functional connectivity using MRI.
Human Brain Mapping, 37(5), 1986–1997. https://doi.org/10
.1002/hbm.23150, PubMed: 27012314
Amico, E., & Goñi, J. (2018). The quest for identifiability in human
functional connectomes. Scientific Reports, 8(1), 8254. https://
doi.org/10.1038/s41598-018-25089-1, PubMed: 29844466
Ashburner, J. (2007). A fast diffeomorphic image registration
algorithm. NeuroImage, 38(1), 95–113. https://doi.org/10.1016/j
.neuroimage.2007.07.007, PubMed: 17761438
Network Neuroscience
1224
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
/
t
.
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
Bailey, L. M., McMillan, L. E., & Newman, A. J. (2020). A sinister
subject: Quantifying handedness-based recruitment biases in
current neuroimaging research. European Journal of Neurosci-
ence, 51(7), 1642–1656. https://doi.org/10.1111/ejn.14542,
PubMed: 31408571
Bellec, P., Carbonell, V., Perlbarg, C., Lepage, O., Lyttelton, V.,
Fonov, A., Janke, J., Tohka, A., & Evans, A. (2011). A neuroim-
aging analysis kit for MATLAB and Octave. Proceedings of the
17th International Conference on Functional Mapping of the
Human Brain.
Campitelli, E. (2022). ggnewscale: Multiple fill and colour scales in
“ggplot2” [R].
Canty, A., & Ripley, B. (2021). boot: Bootstrap R (S-plus) functions
(1.3-28).
Chan, M. Y., Park, D. C., Savalia, N. K., Petersen, S. E., & Wig, G. S.
(2014). Decreased segregation of brain systems across the
healthy adult lifespan. Proceedings of the National Academy of
Sciences, 111(46), E4997–E5006. https://doi.org/10.1073/pnas
.1415122111, PubMed: 25368199
Chen, P. Y., Chiou, J. M., Yang, Y. F., Chen, Y. T., Hsieh, H. L.,
Chang, Y. L., & Tseng, W. Y. I. (2016). Heterogeneous aging
effects on functional connectivity in different cortical regions: A
resting-state functional MRI study using functional data analysis.
PLOS ONE, 11(9), e0162028. https://doi.org/10.1371/journal
.pone.0162028, PubMed: 27658309
da Silva Castanheira, J., Orozco Perez, H. D., Misic, B., & Baillet, S.
(2021). Brief segments of neurophysiological activity enable
individual differentiation. Nature Communications, 12(1), 5713.
https://doi.org/10.1038/s41467-021-25895-8, PubMed: 34588439
Daskalopoulou, C., Koukounari, A., Wu, Y.-T., Terrera, G. M.,
Caballero, F. F., de la Fuente, J., Tyrovolas, S., Panagiotakos,
D. B., Prince, M., & Prina, M. (2019). Healthy ageing trajectories
and lifestyle behaviour: The Mexican Health and Aging Study.
Scientific Reports, 9(1), 11041. https://doi.org/10.1038/s41598
-019-47238-w, PubMed: 31363117
Davison, A. C., & Hinkley, D. V. (1997). Bootstrap methods and
their applications. Cambridge University Press. https://doi.org
/10.1017/CBO9780511802843
de Lange, S. C., Scholtens, L. H., van den Berg, L. H., Boks, M. P.,
Bozzali, M., Cahn, W., Dannlowski, U., Durston, S., Geuze, E.,
van Haren, N. E. M., Hillegers, M. H. J., Koch, K., Jurado, M. Á.,
Mancini, M., Marqués-Iturria, I., Meinert, S., Ophoff, R. A., Reess,
T. J., Repple, J., … van den Heuvel, M. P. (2019). Shared vulner-
ability for connectome alterations across psychiatric and neuro-
logical brain disorders. Nature Human Behaviour, 3(9), 988–998.
https://doi.org/10.1038/s41562-019-0659-6 , PubMed:
31384023
DuPre, E., & Spreng, R. N. (2017). Structural covariance networks
across the life span, from 6 to 94 years of age. Network Neuro-
science, 1(3), 302–323. https://doi.org/10.1162/NETN_a_00016,
PubMed: 29855624
Finn, E. S., & Constable, R. T. (2016). Individual variation in func-
tional brain connectivity: Implications for personalized
approaches to psychiatric disease. Dialogues in Clinical Neuro-
science, 18(3), 277–287. https://doi.org/10.31887/DCNS.2016
.18.3/efinn, PubMed: 27757062
Finn, E. S., & Rosenberg, M. D. (2021). Beyond fingerprinting:
Choosing predictive connectomes over reliable connectomes.
NeuroImage, 239, 118254. https://doi.org/10.1016/j.neuroimage
.2021.118254, PubMed: 34118397
Finn, E. S., Scheinost, D., Finn, D. M., Shen, X., Papademetris, X., &
Constable, R. T. (2017). Can brain state be manipulated to
emphasize individual differences in functional connectivity? Neu-
roImage, 160, 140–151. https://doi.org/10.1016/j.neuroimage
.2017.03.064, PubMed: 28373122
Finn, E. S., Shen, X., Scheinost, D., Rosenberg, M. D., Huang, J.,
Chun, M. M., Papademetris, X., & Constable, R. T. (2015). Func-
tional connectome fingerprinting: Identifying individuals using
patterns of brain connectivity. Nature Neuroscience, 18(11),
1664–1671. https://doi.org/10.1038/nn.4135, PubMed:
26457551
Fulcher, B. D., & Jones, N. S. (2017). hctsa: A computational frame-
work for automated time-series phenotyping using massive fea-
ture extraction. Cell Systems, 5(5), 527–531. https://doi.org/10
.1016/j.cels.2017.10.001, PubMed: 29102608
Fulcher, B. D., Little, M. A., & Jones, N. S. (2013). Highly compar-
ative time-series analysis: The empirical structure of time series
and their methods. Journal of the Royal Society Interface,
10(83), 20130048. https://doi.org/10.1098/rsif.2013.0048,
PubMed: 23554344
Geerligs, L., Rubinov, M., Tyler, L. K., Brayne, C., Bullmore, E. T.,
Calder, A. C., Cusack, R., Dalgleish, T., Duncan, J., Henson,
R. N., Matthews, F. E., Marslen-Wilson, W. D., Rowe, J. B.,
Shafto, M. A., Campbell, K., Cheung, T., Davis, S., Geerligs, L.,
Kievit, R., … Henson, R. N. (2015). State and trait components of
functional connectivity: Individual differences vary with mental
state. Journal of Neuroscience, 35(41), 13949–13961. https://
doi.org/10.1523/ JNEUROSCI.1324-15.2015, PubMed:
26468196
Geerligs, L., Tsvetanov, K. A., Cam-CAN, & Henson, R. N. (2017).
Challenges in measuring individual differences in functional con-
nectivity using fMRI: The case of healthy aging. Human Brain
Mapping, 38(8), 4125–4156. https://doi.org/10.1002/ hbm
.23653, PubMed: 28544076
Gómez-Robles, A., Hopkins, W. D., Schapiro, S. J., & Sherwood,
C. C. (2015). Relaxed genetic control of cortical organization
in human brains compared with chimpanzees. Proceedings of
the National Academy of Sciences, 112(48), 14799–14804.
https://doi.org/10.1073/pnas.1512646112, PubMed: 26627234
Gonneaud, J., Baria, A. T., Pichet Binette, A., Gordon, B. A., Chhatwal,
J. P., Cruchaga, C., Jucker, M., Levin, J., Salloway, S., Farlow, M.,
Gauthier, S., Benzinger, T. L. S., Morris, J. C., Bateman, R. J.,
Breitner, J. C. S., Poirier, J., Vachon-Presseau, E., & Villeneuve,
S. (2020). Functional brain age prediction suggests accelerated
aging in preclinical familial Alzheimer’s disease, irrespective of
fibrillar amyloid-beta pathology. bioRxiv. https://doi.org/10.1101
/2020.05.06.076745
Gonneaud, J., Baria, A. T., Pichet Binette, A., Gordon, B. A., Chhatwal,
J. P., Cruchaga, C., Jucker, M., Levin, J., Salloway, S., Farlow, M.,
Gauthier, S., Benzinger, T. L. S., Morris, J. C., Bateman, R. J.,
Breitner, J. C. S., Poirier, J., Vachon-Presseau, E., Villeneuve, S.,
Alzheimer’s Disease Neuroimaging Initiative (ADNI), … Vachon-
Presseau, E. (2021). Accelerated functional brain aging in
pre-clinical familial Alzheimer’s disease. Nature Communica-
tions, 12(1), 5346. https://doi.org/10.1038/s41467-021-25492-9,
PubMed: 34504080
Network Neuroscience
1225
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
.
/
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
Gratton, C., Laumann, T. O., Nielsen, A. N., Greene, D. J., Gordon,
E. M., Gilmore, A. W., Nelson, S. M., Coalson, R. S., Snyder,
A. Z., Schlaggar, B. L., Dosenbach, N. U. F., & Petersen, S. E.
(2018). Functional brain networks are dominated by stable group
and individual factors, not cognitive or daily variation. Neuron,
98(2), 439–452. https://doi.org/10.1016/j.neuron.2018.03.035,
PubMed: 29673485
Guo, C. C., Kurth, F., Zhou, J., Mayer, E. A., Eickhoff, S. B., Kramer,
J. H., & Seeley, W. W. (2012). One-year test-retest reliability of
intrinsic connectivity network fMRI in older adults. NeuroImage,
61(4), 1471–1483. https://doi.org/10.1016/j.neuroimage.2012
.03.027, PubMed: 22446491
Harris, C. R., Millman, K. J., van der Walt, S. J., Gommers, R.,
Virtanen, P., Cournapeau, D., Wieser, E., Taylor, J., Berg, S.,
Smith, N. J., Kern, R., Picus, M., Hoyer, S., van Kerkwijk,
M. H., Brett, M., Haldane, A., del Río, J. F., Wiebe, M., Peterson,
P., … Oliphant, T. E. (2020). Array programming with NumPy.
Nature, 585(7825), 357–362. https://doi.org/10.1038/s41586
-020-2649-2, PubMed: 32939066
Horien, C., Noble, S., Finn, E. S., Shen, X., Scheinost, D., &
Constable, R. T. (2018). Considering factors affecting the
connectome-based identification process: Comment on Waller
et al. NeuroImage, 1(203), 172–175. https://doi.org/10.1016/j
.neuroimage.2017.12.045, PubMed: 29253655
Horien, C., Shen, X., Scheinost, D., & Constable, R. T. (2019). The
individual functional connectome is unique and stable over
months to years. NeuroImage, 189, 676–687. https://doi.org/10
.1016/j.neuroimage.2019.02.002, PubMed: 30721751
Hu, D., Wang, F., Zhang, H., Wu, Z., Zhou, Z., Li, G. G., Wang, L.,
Lin, W., & Li, G. G. (2022). Existence of functional connectome
fingerprint during infancy and its stability over months. Journal of
Neuroscience, 42(3), 377–389. https://doi.org/10.1523
/JNEUROSCI.0480-21.2021, PubMed: 34789554
Jalbrzikowski, M., Liu, F., Foran, W., Klei, L., Calabro, F. J., Roeder,
K., Devlin, B., & Luna, B. (2020). Functional connectome finger-
printing accuracy in youths and adults is similar when examined
on the same day and 1.5-years apart. Human Brain Mapping,
41(15), 4187–4199. https://doi.org/10.1002/ hbm.25118,
PubMed: 32652852
Kaufmann, T., Alnæs, D., Brandt, C. L., Bettella, F., Djurovic, S.,
Andreassen, O. A., & Westlye, L. T. (2018). Stability of the brain
functional connectome fingerprint in individuals with schizo-
phrenia. JAMA Psychiatry, 75(7), 749–751. https://doi.org/10
.1001/jamapsychiatry.2018.0844, PubMed: 29799905
Kaufmann, T., Alnæs, D., Doan, N. T., Brandt, C. L., Andreassen,
O. A., & Westlye, L. T. (2017). Delayed stabilization and individ-
ualization in connectome development are related to psychiatric
disorders. Nature Neuroscience, 20(4), 513–515. https://doi.org
/10.1038/nn.4511, PubMed: 28218917
Kupis, L., Goodman, Z. T., Kornfeld, S., Hoang, S., Romero, C.,
Dirks, B., Dehoney, J., Chang, C., Spreng, R. N., Nomi, J. S., &
Uddin, L. Q. (2021). Brain dynamics underlying cognitive flexi-
bility across the lifespan. Cerebral Cortex, 31(11), 5263–5274.
https://doi.org/10.1093/cercor/bhab156, PubMed: 34145442
Lin Pedersen, T. (2020). patchwork: The composer of plots (1.1.1).
Mansour, L. S., Tian, Y., Yeo, B. T. T., Cropley, V., & Zalesky, A.
(2021). High-resolution connectomic fingerprints: Mapping
identity and behavior. NeuroImage, 229, 117695.
neural
https://doi.org/10.1016/j.neuroimage.2020.117695, PubMed:
33422711
Mantwill, M., Gell, M., Krohn, S., & Finke, C. (2022). Brain connec-
tivity fingerprinting and behavioural prediction rest on distinct
functional systems of the human connectome. Communications
Biology, 5(1), 261. https://doi.org/10.1038/s42003-022-03185-3,
PubMed: 35332230
Marrelec, G., Krainik, A., Duffau, H., Pélégrini-Issac, M., Lehéricy,
S., Doyon, J., & Benali, H. (2006). Partial correlation for func-
tional brain interactivity investigation in functional MRI. Neuro-
Image, 32(1), 228–237. https://doi.org/10.1016/j.neuroimage
.2005.12.057, PubMed: 16777436
McKinney, W. (2010). Data structures for statistical computing in
Python. In S. J. van der Walt & K. J. Millman (Eds.), Proceedings
of the 9th Python in Science Conference (pp. 56–61). https://doi
.org/10.25080/Majora-92bf1922-00a
Mowinckel, A. M., & Vidal-Piñeiro, D. (2020). Visualization of
brain statistics with R packages ggseg and ggseg3d. Advances in
Methods and Practices in Psychological Science, 3(4), 466–483.
https://doi.org/10.1177/2515245920928009
Mueller, S., Wang, D., Fox, M. D., Yeo, B. T. T., Sepulcre, J.,
Sabuncu, M. R., Shafee, R., Lu, J., & Liu, H. (2013). Individual
variability in functional connectivity architecture of the human
brain. Neuron, 77(3), 586–595. https://doi.org/10.1016/j.neuron
.2012.12.028, PubMed: 23395382
Nadig, A., Seidlitz, J., McDermott, C. L., Liu, S., Bethlehem, R.,
Moore, T. M., Mallard, T. T., Clasen, L. S., Blumenthal, J. D.,
Lalonde, F., Gur, R. C., Gur, R. E., Bullmore, E. T., Satterthwaite,
T. D., & Raznahan, A. (2021). Morphological integration of the
human brain across adolescence and adulthood. Proceedings of
the National Academy of Sciences, 118(14), e2023860118.
https://doi.org/10.1073/pnas.2023860118, PubMed: 33811142
Orban, P., Madjar, C., Savard, M., Dansereau, C., Tam, A., Das, S.,
Evans, A. C., Rosa-Neto, P., Breitner, J. C. S., Bellec, P., Aisen, P.,
Pascoal, T. A., Anthal, E., Appleby, M., Barkun, A., Beaudry, T.,
Benbouhoud, F., Bohbot, V., Brandt, J., … Wang, S. (2015). Test-
retest resting-state fMRI in healthy elderly persons with a family
history of Alzheimer’s disease. Scientific Data, 2, 150043. https://
doi.org/10.1038/sdata.2015.43, PubMed: 26504522
Ousdal, O. T., Kaufmann, T., Kolskår, K., Vik, A., Wehling, E.,
Lundervold, A. J., Lundervold, A., & Westlye, L. T. (2020).
Longitudinal stability of the brain functional connectome is asso-
ciated with episodic memory performance in aging. Human
Brain Mapping, 41(3), 697–709. https://doi.org/10.1002/ hbm
.24833, PubMed: 31652017
Pandas Development Team. (2020). pandas-dev/pandas: Pandas
(1.1.3). Zenodo. https://doi.org/10.5281/zenodo.3509134
Pichet Binette, A., Gonneaud, J., Vogel, J. W., La Joie, R., Rosa-
Neto, P., Louis Collins, D., Poirier, J., Breitner, J. C. S., Villeneuve,
S., Vachon-Presseau, E., Tremblay-Mercier, J., & Madjar, C.
(2020). Morphometric network differences in ageing versus
Alzheimer’s disease dementia. Brain, 143(2), 635–649. https://
doi.org/10.1093/brain/awz414, PubMed: 32040564
Power, J. D., Cohen, A. L., Nelson, S. M., Wig, G. S., Barnes, K. A.,
Church, J. A., Vogel, A. C., Laumann, T. O., Miezin, F. M.,
Schlaggar, B. L., & Petersen, S. E. (2011). Functional network orga-
nization of the human brain. Neuron, 72(4), 665–678. https://doi
.org/10.1016/j.neuron.2011.09.006, PubMed: 22099467
Network Neuroscience
1226
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
t
/
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
t
.
/
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3
Functional connectome fingerprinting across the lifespan
Pritschet, L., Santander, T., Taylor, C. M., Layher, E., Yu, S., Miller,
M. B., Grafton, S. T., & Jacobs, E. G. (2020). Functional reorga-
nization of brain networks across the human menstrual cycle.
N e u ro I m a g e , 2 2 0 , 11 7 0 9 1 . h t t p s : / / d o i . o rg / 1 0 . 1 0 1 6 / j
.neuroimage.2020.117091, PubMed: 32621974
R Core Team. (2020). R: A language and environment for statistical
computing. R Foundation for Statistical Computing.
Schaefer, A., Kong, R., Gordon, E. M., Laumann, T. O., Zuo, X.-N.,
Holmes, A. J., Eickhoff, S. B., & Yeo, B. T. T. (2018). Local-global
parcellation of the human cerebral cortex from intrinsic func-
tional connectivity MRI. Cerebral Cortex, 28(9), 3095–3114.
https://doi.org/10.1093/cercor/bhx179, PubMed: 28981612
Setton, R., Mwilambwe-Tshilobo, L., Girn, M., Lockrow, A. W.,
Baracchini, G., Hughes, C., Lowe, A. J., Cassidy, B. N., Li, J.,
Luh, W.-M., Bzdok, D., Leahy, R. M., Ge, T., Margulies, D. S.,
Misic, B., Bernhardt, B. C., Stevens, W. D., De Brigard, F., Kundu,
P., … Spreng, R. N. (2023). Age differences in the functional
architecture of the human brain. Cerebral Cortex, 33(1), 114–134.
https://doi.org/10.1093/cercor/bhac056, PubMed: 35231927
Shafiei, G., Markello, R. D., De Wael, R. V., Bernhardt, B. C.,
Fulcher, B. D., & Misic, B. (2020). Topographic gradients of
intrinsic dynamics across neocortex. eLife, 9, e62116. https://
doi.org/10.7554/eLife.62116, PubMed: 33331819
Shafto, M. A., Tyler, L. K., Dixon, M., Taylor, J. R., Rowe, J. B.,
Cusack, R., Calder, A. J., Marslen-Wilson, W. D., Duncan, J.,
Dalgleish, T., Henson, R. N., Brayne, C., Bullmore, E., Campbell,
K., Cheung, T., Davis, S., Geerligs, L., Kievit, R., McCarrey, A., …
Matthews, F. E. (2014). The Cambridge Centre for Ageing and
Neuroscience (Cam-CAN) study protocol: A cross-sectional, life-
span, multidisciplinary examination of healthy cognitive ageing.
BMC Neurology, 14(1), 204. https://doi.org/10.1186/s12883-014
-0204-1, PubMed: 25412575
Sporns, O., & Betzel, R. F. (2016). Modular brain networks. Annual
Review of Psychology, 67, 613–640. https://doi.org/10.1146
/annurev-psych-122414-033634, PubMed: 26393868
Stimson, J. A., Carmines, E. G., & Zeller, R. A. (1978). Interpreting
polynomial regression. Sociological Methods and Research, 6(4),
515–524. https://doi.org/10.1177/004912417800600405
St-Onge, F. (2022). Functional connectome fingerprinting through
the lifespan, GitHub. https://github.com/villeneuvelab/projects
Taylor, J. R., Williams, N., Cusack, R., Auer, T., Shafto, M. A.,
Dixon, M., Tyler, L. K., Cam-CAN, & Henson, R. N. (2017). The
Cambridge Centre for Ageing and Neuroscience (Cam-CAN) data
repository: Structural and functional MRI, MEG, and cognitive
data from a cross-sectional adult lifespan sample. NeuroImage,
144, 262–269. https://doi.org/10.1016/j.neuroimage.2015.09
.018, PubMed: 26375206
Vallat, R. (2018). Pingouin: Statistics in Python. Journal of Open
Source Software, 3(31), 1026. https://doi.org/10.21105/joss.01026
Vanderwal, T., Eilbott, J., Finn, E. S., Craddock, R. C., Turnbull, A.,
& Castellanos, F. X. (2017). Individual differences in functional
connectivity during naturalistic viewing conditions. NeuroImage,
157, 521–530. https://doi.org/10.1016/j.neuroimage.2017.06
.027, PubMed: 28625875
Vanderwal, T., Eilbott, J., Kelly, C., Frew, S. R., Woodward, T. S.,
Milham, M. P., & Castellanos, F. X. (2021). Stability and similarity
of the pediatric connectome as developmental measures. Neuro-
Image, 226, 117537. https://doi.org/10.1016/j.neuroimage.2020
.117537, PubMed: 33186720
Van De Ville, D., Farouj, Y., Preti, M. G., Liégeois, R., & Amico, E.
(2021). When makes you unique: Temporality of the human
brain fingerprint. Science Advances, 7(42), eabj0751. https://doi
.org/10.1126/sciadv.abj0751, PubMed: 34652937
Váša, F., Seidlitz, J., Romero-Garcia, R., Whitaker, K. J., Rosenthal,
G., Vértes, P. E., Shinn, M., Alexander-Bloch, A., Fonagy, P.,
Dolan, R. J., Jones, P. B., Goodyer, I. M., Sporns, O., & Bullmore,
E. T. (2018). Adolescent tuning of association cortex in human
structural brain networks. Cerebral Cortex, 28(1), 281–294.
https://doi.org/10.1093/cercor/bhx249, PubMed: 29088339
Virtanen, P., Gommers, R., Oliphant, T. E., Haberland, M., Reddy,
T., Cournapeau, D., Burovski, E., Peterson, P., Weckesser, W.,
Bright, J., van der Walt, S. J., Brett, M., Wilson, J., Millman,
K. J., Mayorov, N., Nelson, A. R. J., Jones, E., Kern, R., Larson,
E., … Vázquez-Baeza, Y. (2020). SciPy 1.0: Fundamental algo-
rithms for scientific computing in Python. Nature Methods,
17(3), 261–272. https://doi.org/10.1038/s41592-019-0686-2,
PubMed: 32015543
Vuong, Q. H. (1989). Likelihood ratio tests for model selection and
non-nested hypotheses. Econometrica, 57(2), 307–333. https://
doi.org/10.2307/1912557
Westfall, P. H. (2011). On using the bootstrap for multiple compar-
isons. Journal of Biopharmaceutical Statistics, 21(6), 1187–1205.
https://doi.org/10.1080/10543406.2011.607751, PubMed:
22023686
Wickham, H., Averick, M., Bryan, J., Chang, W., D’Agostino
McGowan, L., François, R., Grolemund, G., Hayes, A., Henry,
L., Hester, J., Kuhn, M., Lin Pedersen, T., Miller, E., Milton Bache,
S., Muller, K., Ooms, J., Robinson, D., Paige Seidel, D., Spinu,
V., … Yutani, H. (2019). Welcome to the tidyverse. Journal of
Open Source Software, 4(43), 1686. https://doi.org/10.21105
/joss.01686
Xu, T., Opitz, A., Craddock, R. C., Wright, M. J., Zuo, X. N., & Milham,
M. P. (2016). Assessing variations in areal organization for the
intrinsic brain: From fingerprints to reliability. Cerebral Cortex,
26(11), 4192–4211. https://doi.org/10.1093/cercor/ bhw241,
PubMed: 27600846
Yeo, B. T. T., Krienen, F. M., Sepulcre, J., Sabuncu, M. R., Lashkari, D.,
Hollinshead, M., Roffman, J. L., Smoller, J. W., Zöllei, L., Polimeni,
J. R., Fisch, B., Liu, H., & Buckner, R. L. (2011). The organization of
the human cerebral cortex estimated by intrinsic functional con-
nectivity. Journal of Neurophysiology, 106(3), 1125–1165. https://
doi.org/10.1152/jn.00338.2011, PubMed: 21653723
Zalesky, A., Fornito, A., & Bullmore, E. (2012). On the use of cor-
relation as a measure of network connectivity. NeuroImage,
60(4), 2096–2106. https://doi.org/10.1016/j.neuroimage.2012
.02.001, PubMed: 22343126
Zonneveld, H. I., Pruim, R. H., Bos, D., Vrooman, H. A., Muetzel,
R. L., Hofman, A., Rombouts, S. A., van der Lugt, A., Niessen,
W. J., Ikram, M. A., & Vernooij, M. W. (2019). Patterns of func-
tional connectivity in an aging population: The Rotterdam Study.
NeuroImage, 189, 432–444. https://doi.org/10.1016/j
.neuroimage.2019.01.041, PubMed: 30659958
Network Neuroscience
1227
l
D
o
w
n
o
a
d
e
d
f
r
o
m
h
t
t
p
:
/
/
d
i
r
e
c
t
.
m
i
t
.
/
t
/
e
d
u
n
e
n
a
r
t
i
c
e
-
p
d
l
f
/
/
/
/
7
3
1
2
0
6
2
1
5
4
8
7
3
n
e
n
_
a
_
0
0
3
2
0
p
d
/
.
t
f
b
y
g
u
e
s
t
t
o
n
0
9
S
e
p
e
m
b
e
r
2
0
2
3